
The effects of recreational Paracentrotus lividus fishing on distribution patterns of sea urchins at...
8
Fisheries Research 81 (2006) 37–44 The effects of recreational Paracentrotus lividus fishing on distribution patterns of sea urchins at Ustica Island MPA (Western Mediterranean, Italy) Paola Gianguzza a,∗ , Mariachiara Chiantore b , Chiara Bonaviri a , Riccardo Cattaneo-Vietti b , Ilaria Vielmini b , Silvano Riggio a a Dipartimento di Biologia Animale, Universit` a degli Studi di Palermo, via Archirafi 18, 90123 (Pa), Italy b DIPTERIS, Universit` a di Genova, Corso Europa 26, 16132 Genova, Italy Received 13 January 2006; received in revised form 31 May 2006; accepted 1 June 2006 Abstract This study evaluated the effects of recreational Paracentrotus lividus fishing on average density and size of this edible sea urchin, and its indirect effects on Arbacia lixula on barren substrates of Ustica Island MPA (SW Italy, Mediterranean Sea). Size, single and pooled species densities, density of large individuals (>40 mm long) and small individuals (<20 mm long) of the two species were estimated by scuba diving in autumn 2003, spring and summer 2004 at two sites impacted by P. lividus fishing (Punta Cavazzi and Pagliaro, take zone C) and one control (Cala Sidoti, no take zone). Two samplings were performed in each season. We found that reduced densities of P. lividus and A. lixula occurred at the fished sites compared with the unfished sites. P. lividus was always larger and more abundant at the protected site than the fished ones, whereas A. lixula had a larger density at the protected site and greater size at the fished sites. The significantly lower density of A. lixula at the sites subjected to fishing of P. lividus could suggest that A. lixula did not benefit from this kind of fishing and thus the two sea urchins might partially share the same habitat and resources in the barren habitats of “Ustica Island” MPA. These outcomes emphasize the role of regulated fishing in maintaining urchin populations at such low densities that interspecific competition is not induced. © 2006 Elsevier B.V. All rights reserved. Keywords: Sea urchins; Fishery; Paracentrotus lividus; Arbacia lixula; MPA; Mediterranean Sea 1. Introduction Marine biologists have widely demonstrated that human exploitation of inshore resources alters the organization and structure of entire marine communities (Steneck, 1998; Watling and Norse, 1998; Turner et al., 1999; Dayton et al., 2002). Intensive fishing leads to dramatic reductions in the abundance of target stocks (direct effect) and, accordingly, this may induce indirect effects on non-target species through food web interactions, such as cascade reactions (Pinnegar and Polunin, 2004). Sea urchins are keystone species of shal- low benthic communities in a wide range of marine habitats and, at the same time, some urchin species are exploited ∗ Corresponding author. Tel.: +39 0916230106; fax: +39 0916230144. E-mail address: [email protected] (P. Gianguzza). worldwide for the delicacy of their gonads (Keesing and Hall, 1998; Lawrence, 2001). Many studies quantified the conse- quent rarefaction and reduction of size and biomass of the target species (Ros and Niell, 1981; R´ egis, 1988; Carter and Van Blaricom, 2002) but little is known about the possible alterations that this fishing may provoke in subtidal com- munities (Rogers-Bennett and Pearse, 2001) and on other co-occurring non-edible sea urchin species (Emlet, 2002; Guidetti et al., 2004). The sea urchins Paracentrotus lividus (Lamarck, 1816) and Arbacia lixula (Linneo, 1758) (Echinodermata: Echi- noidea) are the most ubiquitous echinoids in the Mediter- ranean sublittoral zone and it has been suggested that their grazing may cause important changes in the distribution pat- terns of benthic communities, exerting a paramount role in the transition from macroalgal beds to coralline barrens, 0165-7836/$ – see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.fishres.2006.06.002
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The effects of recreational Paracentrotus lividus fishing on distribution patterns of sea urchins at...

A
idi(awspfi©
K
1
eaW2atfala
0d
Fisheries Research 81 (2006) 37–44
The effects of recreational Paracentrotus lividus fishingon distribution patterns of sea urchins at Ustica Island
MPA (Western Mediterranean, Italy)
Paola Gianguzza a,∗, Mariachiara Chiantore b, Chiara Bonaviri a,Riccardo Cattaneo-Vietti b, Ilaria Vielmini b, Silvano Riggio a
a Dipartimento di Biologia Animale, Universita degli Studi di Palermo, via Archirafi 18, 90123 (Pa), Italyb DIPTERIS, Universita di Genova, Corso Europa 26, 16132 Genova, Italy
Received 13 January 2006; received in revised form 31 May 2006; accepted 1 June 2006
bstract
This study evaluated the effects of recreational Paracentrotus lividus fishing on average density and size of this edible sea urchin, and itsndirect effects on Arbacia lixula on barren substrates of Ustica Island MPA (SW Italy, Mediterranean Sea). Size, single and pooled speciesensities, density of large individuals (>40 mm long) and small individuals (<20 mm long) of the two species were estimated by scuba divingn autumn 2003, spring and summer 2004 at two sites impacted by P. lividus fishing (Punta Cavazzi and Pagliaro, take zone C) and one controlCala Sidoti, no take zone). Two samplings were performed in each season. We found that reduced densities of P. lividus and A. lixula occurredt the fished sites compared with the unfished sites. P. lividus was always larger and more abundant at the protected site than the fished ones,hereas A. lixula had a larger density at the protected site and greater size at the fished sites. The significantly lower density of A. lixula at the
ites subjected to fishing of P. lividus could suggest that A. lixula did not benefit from this kind of fishing and thus the two sea urchins mightartially share the same habitat and resources in the barren habitats of “Ustica Island” MPA. These outcomes emphasize the role of regulatedshing in maintaining urchin populations at such low densities that interspecific competition is not induced.2006 Elsevier B.V. All rights reserved.
A; Med
w1qtVamcG
eywords: Sea urchins; Fishery; Paracentrotus lividus; Arbacia lixula; MP
. Introduction
Marine biologists have widely demonstrated that humanxploitation of inshore resources alters the organizationnd structure of entire marine communities (Steneck, 1998;atling and Norse, 1998; Turner et al., 1999; Dayton et al.,
002). Intensive fishing leads to dramatic reductions in thebundance of target stocks (direct effect) and, accordingly,his may induce indirect effects on non-target species throughood web interactions, such as cascade reactions (Pinnegar
nd Polunin, 2004). Sea urchins are keystone species of shal-ow benthic communities in a wide range of marine habitatsnd, at the same time, some urchin species are exploited∗ Corresponding author. Tel.: +39 0916230106; fax: +39 0916230144.E-mail address: [email protected] (P. Gianguzza).
anrgtt
165-7836/$ – see front matter © 2006 Elsevier B.V. All rights reserved.oi:10.1016/j.fishres.2006.06.002
iterranean Sea
orldwide for the delicacy of their gonads (Keesing and Hall,998; Lawrence, 2001). Many studies quantified the conse-uent rarefaction and reduction of size and biomass of thearget species (Ros and Niell, 1981; Regis, 1988; Carter andan Blaricom, 2002) but little is known about the possiblelterations that this fishing may provoke in subtidal com-unities (Rogers-Bennett and Pearse, 2001) and on other
o-occurring non-edible sea urchin species (Emlet, 2002;uidetti et al., 2004).The sea urchins Paracentrotus lividus (Lamarck, 1816)
nd Arbacia lixula (Linneo, 1758) (Echinodermata: Echi-oidea) are the most ubiquitous echinoids in the Mediter-
anean sublittoral zone and it has been suggested that theirrazing may cause important changes in the distribution pat-erns of benthic communities, exerting a paramount role inhe transition from macroalgal beds to coralline barrens,
3 eries Re
c1HtdeieaNwipeao
tcmafighaalrgepoli
ot(m1wsrigaeetcii(str
usas1o(fiosotltrP(fis
2
2
s(pwAwC) equally share the remaining area.
According to the institutive decree (D.I. 12 November1986) only research activities are authorized in the no takezone, whereas recreational activities, such as scuba div-
8 P. Gianguzza et al. / Fish
haracterised by lower complexity and diversity (Verlaque,987; Sala et al., 1998; Boudouresque and Verlaque, 2001;ereu et al., 2004). Kempf (1962) investigated extensively
he ecology of these co-occurring species and firstly reportedifferent patterns in their slope distributions (P. lividus is gen-rally more abundant on horizontal surfaces while A. lixulas more abundant on vertical substrata) and feeding prefer-nces. P. lividus is reported to feed mainly on fleshy algaend suspended organic particles (Kempf, 1962; Verlaque andedelec, 1983; Frantzis et al., 1988; Bulleri et al., 1999),hile A. lixula prefers encrusting corallines. This background
nformation could indicate that the two species do not com-ete, but this issue is still a matter of debate (see Bullerit al., 1999). However, to our knowledge no information isvailable on barren grounds characterized by a serious lossf sublittoral biomass and biodiversity.
Besides its ecological role, P. lividus is known for its dis-inctive aroma and the desirable taste of its gonads that,ooked or raw, are considered a gastronomic delicacy inany Mediterranean areas (Turon et al., 1995; Palacın et
l., 1998; Lawrence, 2001). The ecological effects of theshery of P. lividus on the species itself are poorly investi-ated (Guidetti et al., 2004). There is evidence that intensiveuman predation is a key process in determining P. lividusbundance in Mediterranean hard bottom communities: inrecent paper Guidetti et al. (2004) show that intensive P.
ividus fishing affects not only its size, but may also have indi-ect consequences on non-target species, such as significantlyreater average density and biomass of the co-occurring, non-dible, black sea urchin, A. lixula. The authors suggest aossible compensative mechanism between the abundancef the two sea urchins species: when the dominant species, P.ividus, is predated, the abundance of Arbacia lixula tends toncrease.
There are, however, no data about a possible impactf P. lividus fishing on its distribution patterns and onhe co-occurring A. lixula in a marine protected areaMPA). Before the creation in 1986 of the Ustica Islandarine protected area (MPA) (western Mediterranean Sea,
0◦43′43′′E–38◦42′20′′N), Ustica rocky sublittoral bottomsere affected by an intensive fishing activity on the edible
ea urchin P. lividus. After the instauration of the protectionegime, the lack of human fishing pressure caused a sharpncrease of P. lividus density (about 20 sea urchins/m2 Rig-io, pers. commun.), with consequent transformation of algalssemblages into barren areas, dominated by a few species ofncrusting algae, as already described in other areas (Frantzist al., 1988; Sala, 1997; Bulleri et al., 1999). In order to stophe growth of this sea urchin species, the Ustica Town Coun-il (the MPA management body at the time) allowed, startingn 1994, recreational (i.e. non-commercial) sea urchin fish-ng in the take zone (zone C). This regulamentated activity
50 P. lividus animals per person per day, during the summereason) compensated local people for the adoption of a sus-ainable policy and gave us the chance to study the effects ofecreational P. lividus fishing within a MPA.F(
search 81 (2006) 37–44
The present study has been conducted at a place where searchin predation is most probably limited to human fishingince fish predators such as Diplodus sargus (Linneo, 1758)nd Diplodus vulgaris (E. Geoffroy St. Hilaire, 1817) arecarce in the MPA of Ustica Island (La Mesa and Vacchi,999), and subsequently offers an opportunity to improveur knowledge of the effects of human fishing on echinoidsEmlet, 2002). We examined the direct effects of P. lividusshing on the species itself and the possible indirect effectsn the co-occurring, but not fished, A. lixula on barren rockyubstrates of Ustica Island MPA. According to the designf asymmetrical analyses of variance (ACI), we comparedhe average density and size structure of P. lividus and A.ixula between one protected and two fished areas. This study,herefore, aims to test the following hypotheses, on barrenocky substrates of Ustica Island MPA: (1) the fishery of. lividus affects its average density and size structure and2) the co-occurring A. lixula increases in numbers at theshed sites, due to the reduced competition with the exploitedpecies.
. Materials and methods
.1. Study area
The study was carried out at Ustica Island, located in theouthern Tyrrhenian Sea, 60 km north of the Sicilian coastFig. 1). The MPA of Ustica Island, created in 1986, encom-asses a total area of 16,000 ha and it contains three zonesith different degrees of protection. The no take zone (zone) extends for 65 ha along the western part of the island,hereas the general reserve (zone B) and the take zone (zone
ig. 1. The zonation of the Ustica Island MPA. The study sites Punta CavazziF1), Punta Parrino (F2) and Cala Sidoti (P) are indicated by the black arrows.
eries Research 81 (2006) 37–44 39
ipir1C
eCFztIiigu
2
emoTwawhted(l15b
U2a(tvdsrhivcrabpo
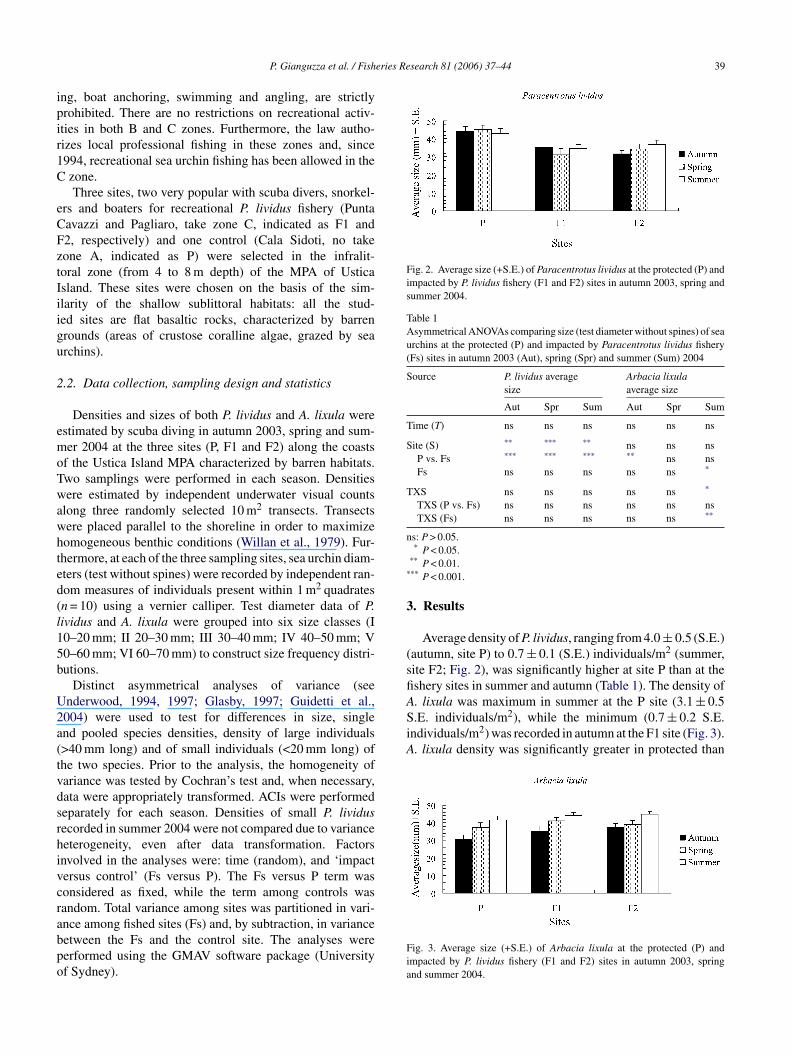
Fig. 2. Average size (+S.E.) of Paracentrotus lividus at the protected (P) andimpacted by P. lividus fishery (F1 and F2) sites in autumn 2003, spring andsummer 2004.
Table 1Asymmetrical ANOVAs comparing size (test diameter without spines) of seaurchins at the protected (P) and impacted by Paracentrotus lividus fishery(Fs) sites in autumn 2003 (Aut), spring (Spr) and summer (Sum) 2004
Source P. lividus averagesize
Arbacia lixulaaverage size
Aut Spr Sum Aut Spr Sum
Time (T) ns ns ns ns ns ns
Site (S) ** *** ** ns ns nsP vs. Fs *** *** *** ** ns nsFs ns ns ns ns ns *
TXS ns ns ns ns ns *
TXS (P vs. Fs) ns ns ns ns ns nsTXS (Fs) ns ns ns ns ns **
ns: P > 0.05.
*
3
(sfiA. lixula was maximum in summer at the P site (3.1 ± 0.5S.E. individuals/m2), while the minimum (0.7 ± 0.2 S.E.individuals/m2) was recorded in autumn at the F1 site (Fig. 3).A. lixula density was significantly greater in protected than
P. Gianguzza et al. / Fish
ng, boat anchoring, swimming and angling, are strictlyrohibited. There are no restrictions on recreational activ-ties in both B and C zones. Furthermore, the law autho-izes local professional fishing in these zones and, since994, recreational sea urchin fishing has been allowed in thezone.Three sites, two very popular with scuba divers, snorkel-
rs and boaters for recreational P. lividus fishery (Puntaavazzi and Pagliaro, take zone C, indicated as F1 and2, respectively) and one control (Cala Sidoti, no takeone A, indicated as P) were selected in the infralit-oral zone (from 4 to 8 m depth) of the MPA of Usticasland. These sites were chosen on the basis of the sim-larity of the shallow sublittoral habitats: all the stud-ed sites are flat basaltic rocks, characterized by barrenrounds (areas of crustose coralline algae, grazed by searchins).
.2. Data collection, sampling design and statistics
Densities and sizes of both P. lividus and A. lixula werestimated by scuba diving in autumn 2003, spring and sum-er 2004 at the three sites (P, F1 and F2) along the coasts
f the Ustica Island MPA characterized by barren habitats.wo samplings were performed in each season. Densitiesere estimated by independent underwater visual counts
long three randomly selected 10 m2 transects. Transectsere placed parallel to the shoreline in order to maximizeomogeneous benthic conditions (Willan et al., 1979). Fur-hermore, at each of the three sampling sites, sea urchin diam-ters (test without spines) were recorded by independent ran-om measures of individuals present within 1 m2 quadratesn = 10) using a vernier calliper. Test diameter data of P.ividus and A. lixula were grouped into six size classes (I0–20 mm; II 20–30 mm; III 30–40 mm; IV 40–50 mm; V0–60 mm; VI 60–70 mm) to construct size frequency distri-utions.
Distinct asymmetrical analyses of variance (seenderwood, 1994, 1997; Glasby, 1997; Guidetti et al.,004) were used to test for differences in size, singlend pooled species densities, density of large individuals>40 mm long) and of small individuals (<20 mm long) ofhe two species. Prior to the analysis, the homogeneity ofariance was tested by Cochran’s test and, when necessary,ata were appropriately transformed. ACIs were performedeparately for each season. Densities of small P. lividusecorded in summer 2004 were not compared due to varianceeterogeneity, even after data transformation. Factorsnvolved in the analyses were: time (random), and ‘impactersus control’ (Fs versus P). The Fs versus P term wasonsidered as fixed, while the term among controls wasandom. Total variance among sites was partitioned in vari-
nce among fished sites (Fs) and, by subtraction, in varianceetween the Fs and the control site. The analyses wereerformed using the GMAV software package (Universityf Sydney).Fia
* P < 0.05.** P < 0.01.** P < 0.001.
. Results
Average density of P. lividus, ranging from 4.0 ± 0.5 (S.E.)autumn, site P) to 0.7 ± 0.1 (S.E.) individuals/m2 (summer,ite F2; Fig. 2), was significantly higher at site P than at theshery sites in summer and autumn (Table 1). The density of
ig. 3. Average size (+S.E.) of Arbacia lixula at the protected (P) andmpacted by P. lividus fishery (F1 and F2) sites in autumn 2003, springnd summer 2004.
40 P. Gianguzza et al. / Fisheries Research 81 (2006) 37–44
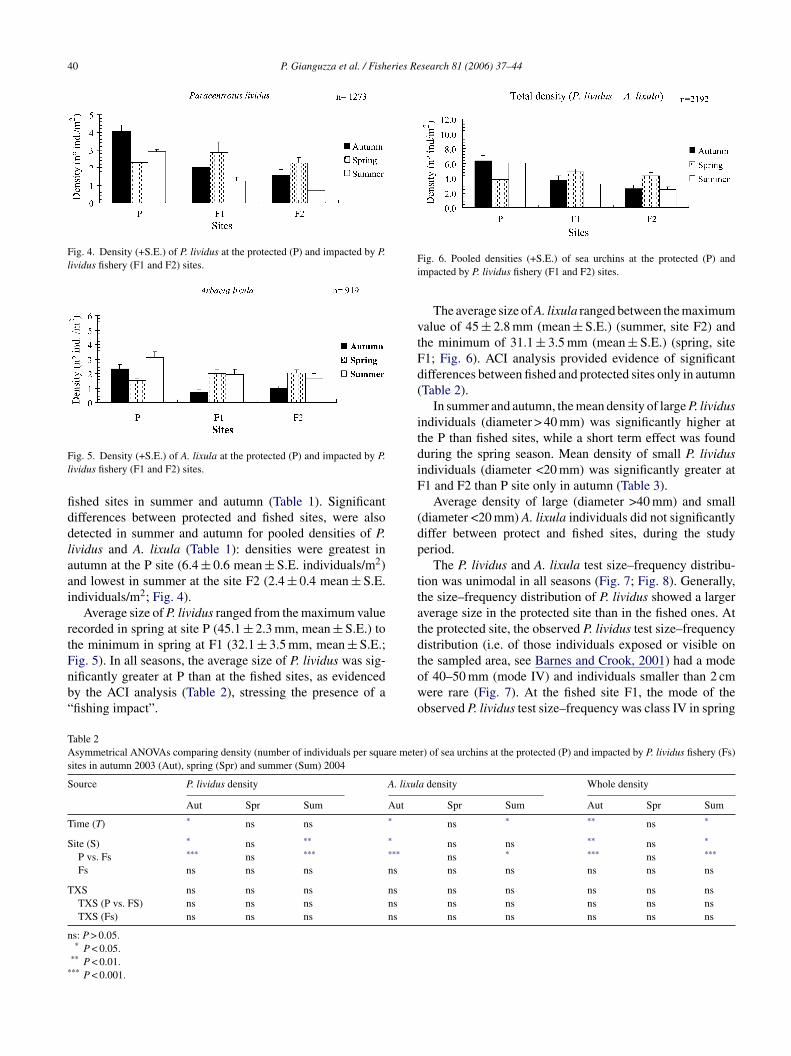
Fig. 4. Density (+S.E.) of P. lividus at the protected (P) and impacted by P.lividus fishery (F1 and F2) sites.
Fl
fiddlaai
rtFnb“
Fi
vtFd(
itdiF
(dp
ttatd
TAs
S
T
S
T
n
*
ig. 5. Density (+S.E.) of A. lixula at the protected (P) and impacted by P.ividus fishery (F1 and F2) sites.
shed sites in summer and autumn (Table 1). Significantifferences between protected and fished sites, were alsoetected in summer and autumn for pooled densities of P.ividus and A. lixula (Table 1): densities were greatest inutumn at the P site (6.4 ± 0.6 mean ± S.E. individuals/m2)nd lowest in summer at the site F2 (2.4 ± 0.4 mean ± S.E.ndividuals/m2; Fig. 4).
Average size of P. lividus ranged from the maximum valueecorded in spring at site P (45.1 ± 2.3 mm, mean ± S.E.) tohe minimum in spring at F1 (32.1 ± 3.5 mm, mean ± S.E.;
ig. 5). In all seasons, the average size of P. lividus was sig-ificantly greater at P than at the fished sites, as evidencedy the ACI analysis (Table 2), stressing the presence of afishing impact”.towo
able 2symmetrical ANOVAs comparing density (number of individuals per square mete
ites in autumn 2003 (Aut), spring (Spr) and summer (Sum) 2004
ource P. lividus density A. lixul
Aut Spr Sum Aut
ime (T) * ns ns *
ite (S) * ns ** *
P vs. Fs *** ns *** ***
Fs ns ns ns ns
XS ns ns ns nsTXS (P vs. FS) ns ns ns nsTXS (Fs) ns ns ns ns
s: P > 0.05.* P < 0.05.
** P < 0.01.** P < 0.001.
ig. 6. Pooled densities (+S.E.) of sea urchins at the protected (P) andmpacted by P. lividus fishery (F1 and F2) sites.
The average size of A. lixula ranged between the maximumalue of 45 ± 2.8 mm (mean ± S.E.) (summer, site F2) andhe minimum of 31.1 ± 3.5 mm (mean ± S.E.) (spring, site1; Fig. 6). ACI analysis provided evidence of significantifferences between fished and protected sites only in autumnTable 2).
In summer and autumn, the mean density of large P. lividusndividuals (diameter > 40 mm) was significantly higher athe P than fished sites, while a short term effect was founduring the spring season. Mean density of small P. lividusndividuals (diameter <20 mm) was significantly greater at1 and F2 than P site only in autumn (Table 3).
Average density of large (diameter >40 mm) and smalldiameter <20 mm) A. lixula individuals did not significantlyiffer between protect and fished sites, during the studyeriod.
The P. lividus and A. lixula test size–frequency distribu-ion was unimodal in all seasons (Fig. 7; Fig. 8). Generally,he size–frequency distribution of P. lividus showed a largerverage size in the protected site than in the fished ones. Athe protected site, the observed P. lividus test size–frequencyistribution (i.e. of those individuals exposed or visible onhe sampled area, see Barnes and Crook, 2001) had a mode
f 40–50 mm (mode IV) and individuals smaller than 2 cmere rare (Fig. 7). At the fished site F1, the mode of thebserved P. lividus test size–frequency was class IV in springr) of sea urchins at the protected (P) and impacted by P. lividus fishery (Fs)
a density Whole density
Spr Sum Aut Spr Sum
ns * ** ns *
ns ns ** ns *
ns * *** ns ***
ns ns ns ns ns
ns ns ns ns nsns ns ns ns nsns ns ns ns ns

P. Gianguzza et al. / Fisheries Research 81 (2006) 37–44 41
Table 3Asymmetrical ANOVAs comparing density (number of individuals per square meter) of the sea urchins larger than 40 mm and shorter than 20 mm at theprotected (P) and impacted by P. lividus fishery (Fs) sites in autumn 2003 (Aut), spring (Spr) and summer (Sum) 2004
Source P. lividus > 40 mm A. lixula > 40 mm P. lividus < 20 mm P. lividus < 20 mm
Aut Spr Sum Aut Spr Sum Aut Spr Sum Aut Spr Sum
Time (T) ns ns * ns ns ns ns ns ns ns ns np
Site (S) * ns ** ns ns ns * ns ns ns ns npP vs. Fs *** ns *** ns ns ns *** ns ns ns ns npFs ns * ns ns ns ns ns ns ns ns ns np
TXS ns ns ns ns ns ns ns ns ns ns ns npTXS (P vs. FS) ns * ns ns ns ns ns ns ns ns ns npTXS (Fs) ns ns ns ns ns ns ns ns ns ns ns np
ns: P > 0.05. np: not performed* P < 0.05
** P < 0.01*** P < 0.001
Fig. 7. Size–frequency distribution of P. lividus at the protected (P) andimpacted by P. lividus fishery (F1 and F2) sites in autumn 2003, spring andsummer 2004. Size classes (I 10–20 mm; II 20–30 mm; III 30–40 mm; IV40–50 mm; V 50–60 mm; VI 60–70 mm).
Fig. 8. Size–frequency distribution of A. lixula at the protected (P) andimpacted by P. lividus fishery (F1 and F2) sites in autumn 2003, springand summer 2004. Size classes (I 10–20 mm; II 20–30 mm; III 30–40 mm;IV 40–50 mm; V 50–60 mm; VI 60–70 mm).

4 eries Re
au(ttIw
sIst(taa
4
aeEtB(hHutgjpojaa1
acaotFgbmte
irbG
nhsaTftScsp(wtt2
PoIirpmePbtsaawDttiPftobfiedr2
ppfta
2 P. Gianguzza et al. / Fish
nd autumn and III springing summer (Fig. 7) and individ-als larger than 40 mm were rare in all sites in all seasonsFig. 7). The observed P. lividus test size–frequency distribu-ion recorded in the fished site F2 was not affected by seasons:he modal class was formed by sea urchins belonging to modeII (Fig. 7) and also in this case sea urchins larger than 40 mmere rare.At the protected site the observed A. lixula test
ize–frequency was class II and III in autumn, class III andV in spring and class IV in summer (Fig. 8). At the fishedite F1, the observed A. lixula test size–frequency distribu-ion did not change seasonally and the mode was class IVFig. 8). The apparent A. lixula test size–frequency distribu-ion recorded in the fishery site F2 was affected by the season:modal class III was recorded in autumn, whereas in springnd summer the modal class was IV (Fig. 8).
. Discussion
Sea urchins are fished commercially worldwide (Keesingnd Hall, 1998), but still little is known about the indirectffects of urchin fishing on subtidal species and communities.xperimental fishing of adult red sea urchins Strongylocen-
rotus franciscanus (Agassiz, 1863) in California (Rogers-ennett and Pearse, 2001) and local urchins in South Africa
Tarr et al., 1996) showed that the fishery of adult urchins mayave a negative effect on the abundance of juvenile abalonesaliotis rufescens (Swainson, 1822) and also on a number ofnfished species. Rogers-Bennett and Pearse (2001) stressedhat adult sea urchins and their spines are an example of bio-enic structures that turns out an essential fish habitats foruvenile abalones and other benthic organism from severalhyla (see also De Waal, 2005). Furthermore the spine canopyf adult sea urchins is most likely also an important habitat foruvenile sea urchins (Tegner and Dayton, 1977), especially inreas where alternative cryptic habitats (e.g., crevices, holesnd undersides of boulders) are rare or absent (Breen et al.,985).
Other studies proved that low density of sea urchin hadpositive effect on algal recovery: see the case of S. fran-
iscanus on kelp forest communities in California (Dean etl., 1984; Tegner and Dayton, 1999) and that of P. lividusn non-crustose algae in Mediterranean and Atlantic habi-ats (Palacın et al., 1998; Verlaque, 1987; Sala et al., 1998).urthermore, experimental sea urchin removal studies sug-est that sea urchin decrease triggers an increase of density,iomass and species richness of a Kenyan coral-reef fish com-unity, because herbivorous fish species positively respond
o an increase in available algal food resources (McClanahant al., 1996).
Regarding the Mediterranean Sea, there is evidence that
ntensive human fishing of P. lividus is a key process ineducing abundance and size of the species itself on hardottom communities (Gras, 1987; Regis, 1988). Recently,uidetti et al. (2004) showed that intensive P. lividus removalafb1
search 81 (2006) 37–44
ot only affects the size of the species itself but may alsoave positive indirect consequences on non-target species,uch as triggering significant increase in average densitynd biomass of the co-occurring black sea urchin, A. lixula.herefore, the authors suggest that A. lixula could benefit
rom the fishing of the dominant species (P. lividus), andhat a compensative mechanism (see also McClanahan andhafir, 1990) would be invoked on the abundance of the twoompetitor sea urchins species in the shallow rocky reefs ofouthern Apulia (SE Italy, Mediterranean Sea). This com-ensative hypothesis was also tested by Tegner and Dayton1977) and by recent field observations along the north-estern coast of America: S. franciscanus fishing enhances
he abundance of the smaller and not fished Strongylocen-rotus purpuratus (Stimpson, 1857) (see, Guidetti et al.,004).
The present study provides evidence that recreational. lividus fishing could alter distribution patterns of co-ccurring sea urchins on barren rocky substrates of Usticasland MPA. We can reasonably state that this kind of fish-ng decreases density of both target and non-target species:educed densities always occurred at the fished sites com-ared with the protected sites. These results are not in agree-ent with literature data (Tegner and Dayton, 1977; Guidetti
t al., 2004). In addition, this research shows that recreational. lividus fishing affects the sea urchin size–frequency distri-utions in opposing ways. P. lividus was always larger athe protected site than at the fished ones, whereas A. lixulahowed greater size in the fished ones. The fact that recre-tional fishing, as well as commercial, reduces the mean sizend density of the fished sea urchins has been proved world-ide and is predictable (Carter and Van Blaricom, 2002).ifferently, for A. lixula we observed larger densities in
he protected site and greater size in the fished ones, andhis finding is worthy of consideration. In fact, the signif-cantly lower density of A. lixula at the sites subjected to. lividus harvesting suggests that A. lixula does not benefitrom P. lividus fishing and is probably negatively affected byhe fishery itself (general disturbance and erroneous fishingf the non-edible species). However, the larger size coulde due to reduced intra-specific competition. Thus, thesendings could confirm preliminary observations providingvidence that the two Mediterranean sea urchins, havingifferent diets, do not compete for resources in the barrenocky substrates of “Ustica Island” MPA (Vielmini et al.,005).
Yet, before drawing any conclusion about sea urchin com-etition, more detailed field experimental studies should beerformed. In fact, in addition to competition, other differentactors are considered to affect the sea urchin populations inemperate coastal areas (for reviews, see Sala et al. (1998)nd Pinnegar et al. (2000)) and these are: predation (both fish
nd human), recruitment, early mortality, epidemic diseases,ood availability, complexity of the substrate, nature of thearren ground and availability of refuges (Tegner and Dayton,977; Jangoux, 1987; McClanahan and Shafir, 1990; Sala and
eries R
ZT
ostieuobaarpes
5
tmiisossgstratti
A
cfimtrsimsepdU
R
B
B
B
B
C
D
D
D
E
F
G
G
G
G
H
J
K
K
P. Gianguzza et al. / Fish
abala, 1996; Lopez et al., 1998; Shears and Babcock, 2003;uya et al., 2005).
The fact that the population structure of sea urchinsbserved in the MPA of Ustica Island is unimodal, couldupport outcomes of La Mesa and Vacchi (1999) who foundhat the presence of D. sargus and sparids in general, scarcen the Ustica Island MPA and thus fish predation does notxert control on the abundances of P. lividus and A. lix-la. Scheibling (1996) showed that usually, in the presencef intense fish predation, the P. lividus size–distribution isimodal and this is because juveniles (∼20 mm long) areble to shelter in crevices or holes and adults (∼40 mm long)ctively escape from predation (Guidetti and Mori, 2005 andeferences therein). According to our results, only humanredation negatively affects the population structure of thedible sea urchin, truncating its adult cohort at the fisheryites (Fig. 7).
. Conclusions
To summarise, in the present study the first hypothesisested (that recreational P. lividus fishing affects abundance,
ean size and size–frequency distribution of the speciestself) was confirmed, while the second one (a compensativencrease of the co-occurring, non-edible species in the fishedites) has to be rejected. In fact, we observed that the removalf the edible species does not trigger an increase of the den-ity of A. lixula. In the fished sites, also A. lixula densityhows a decrease during the fishing season, while its sizeets larger in autumn. This reinforces the hypothesis that thepecies do not compete, but are both affected by fishing dis-urbance, and that intraspecific competition may have someole in affecting the size of the species (reduced numbersllow larger size to be attained). These outcomes emphasizehe role of regulated fishing in maintaining urchin popula-ions at such low densities that interspecific competition is notnduced.
cknowledgements
The authors are indebted to Davide Agnetta, Davide Perri-one and Claudia Scianna for their indispensable help duringeld work. Paolo Guidetti, Marco Milazzo and Fabio Badala-enti for their precious comments on an earlier draft of
his manuscript. Laura Roger-Bennet and one anonymouseviewer made constructive and challenging comments thatubstentially improved the paper. Julie-Anne Buck for revis-ng the English text. The Harbour Office of Palermo, the MPA
anagement body of the Ustica Island, for assistance. Thistudy was funded by the MATT (Ministero dell’Ambiente
Difesa del Territorio) research project Monitoraggio delleopolazioni di Paracentrotus lividus e Arbacia lixula ai finiella tutela della diversita biologica dell’ Amp “Isola distica”.L
L
esearch 81 (2006) 37–44 43
eferences
arnes, D.K.A., Crook, A.C., 2001. Quantifying behavioural determinantsof the coastal European sea urchin Paracentrotus lividus. Mar. Biol.138, 1205–1212.
oudouresque, C.F., Verlaque, M., 2001. Ecology of Paracentrotuslividus. In: Lawrence, J.M. (Ed.), Edible Sea Urchins. Elsevier, Ams-terdam, pp. 177–216.
reen, P.A., Carolsfeld, W., Yamanaka, K.L., 1985. Social behaviour ofjuvenile red sea urchins Strongylocentrotus franciscanus (Agassiz). J.Exp. Mar. Biol. Ecol. 92, 45–61.
ulleri, F., Benedetti-Cecchi, L., Cinelli, F., 1999. Grazing by the seaurchins Arbacia lixula L. and Paracentrotus lividus Lam. in the North-west Mediterranean. J. Exp. Mar. Biol. Ecol. 241, 81–95.
arter, S.K., Van Blaricom, G.R., 2002. Effect of experimental harvest onred sea urchins (Strongylocentrotus franciscanus) in northern Wash-ington. Fish. Bull. 100, 662–673.
ayton, P.K., Thrush, S.F., Coleman, F.C., 2002. Ecological Effects ofFishing in Marine Ecosystems of the United States. Virginia, PewOcean Commission, Arlington, p. 45.
ean, T.A., Schroeter, S.C., Dixon, J.D., 1984. Effects of grazing by twospecies of sea urchins (Strongylocentrotus franciscanus and Lytech-inus anamesus) on recruitment and survival of two species of kelp(Macrocystis pyrifera and Pterygophora californica). Mar. Biol. 78,301–313.
e Waal, S.W.P., 2005. Boulders or urchins? Selecting seeding sites forjuvenile Haliotis midae along the Namaqualand coast of South Africa.Afr. J. Mar. Sci. 27, 501–504.
mlet, R.B., 2002. Ecology of adult sea urchins. In: Yokota, Y., Matranga,V., Smolenicka, Z. (Eds.), Sea Urch: From Basic Biology to Aqua-culture. Balkema Publisher, Elsevier, Rotterdam, Amsterdam, pp.111–114.
rantzis, A., Berthon, J.F., Maggiore, F., 1988. Relation trophique entreles oursins Arbacia lixula et Paracentrotus lividus (Echinoidea regu-laria) et le phytobenthos infralittoral superficiel de la baie de Port-Cros (Var France). Sci. Rep. Port-Cros Natl. Park 14, 81–140.
lasby, T.M., 1997. Analysing data from post-impact studies using asym-metrical analyses of variance: a case study of epibiota on marinas.Aust. J. Ecol. 22, 448–459.
ras, G., 1987. Evolution des stocks de l’oursin comestible Paracentrotuslividus dans le quartier maritime de Marseille (France), soumis a unepˆeche intensive, entre les campagnes 1984–1985 et 1986–1987. In:Boudouresque, C.F. (Ed.), Colloque International sur Paracentrotuslividus et les Oursins Comestibles. GIS Posidonie Publisher, Marseille,pp. 363–370.
uidetti, P., Mori, M., 2005. Morpho-functional defences of Mediter-ranean sea urchins, Paracentrotus lividus and Arbacia lixula, againstfish predators. Mar. Biol. 147, 797–802.
uidetti, P., Terlizzi, A., Boero, F., 2004. Effects of the edible sea urchin,Paracentrotus lividus, fishery along the Apulian rocky coast (SE ItalyMediterranean Sea). Fish. Res. 66, 287–297.
ereu, M., Cabala, M., Linares, C., Sala, E., 2004. Temporal and spatialvariability in settlement of the sea urchin Paracentrotus lividus in theNW Mediterranean. Mar. Biol. 144, 1011–1018.
angoux, M., 1987. Diseases of Echinodermata. I. Agents microorganismsand protistans. Dis. Aquat. Org. 2, 147–162.
eesing, J.K., Hall, K.C., 1998. Review of harvests and status of wordssea urchin fisheries points to opportunities for aquaculture. J. Shell.Res. 17, 1597–1604.
empf, M., 1962. Recherches d’ecologie compare sur Paracentrotuslividus (Lmk.) et Arbacia lixula (L. ) (I). Rec. Trav. St. Mar. End.25, 47–116.
a Mesa, G., Vacchi, M., 1999. An analysis of the coastal fish assem-blages of the Ustica Island Marine Reserve (Mediterranean Sea). Mar.Ecol. 20, 147–165.
awrence, J.M., 2001. Edible Sea Urchins: Biology and Ecology. Elsevier,Amsterdam.

4 eries R
L
M
M
P
P
P
R
R
R
S
S
S
S
S
S
T
T
T
T
T
T
U
U
V
V
V
4 P. Gianguzza et al. / Fish
opez, S., Turon, X., Montero, E., Palacin, C., Duarte, C.M., Tar-juelo, I., 1998. Larval abundance, recruitment and early mortal-ity in Paracentrotus lividus (Echinoidea) Interannual variability andplankton–benthos coupling. Mar. Ecol. Prog. Ser. 172, 239–251.
cClanahan, T.R., Shafir, S.H., 1990. Causes and consequences of seaurchin abundance and diversity in Kenyan coral reef lagoons. Oecolo-gia 83, 362–370.
cClanahan, T.R., Kamukuru, A.T., Muthiga, N.A., Gilagabher Yebio,M., Obura, D., 1996. Effect of sea urchin reductions on algae, coral,and fish populations. Cons. Biol. 10, 136–154.
alacın, C., Giribet, G., Carner, S., Dantart, L., Turon, X., 1998. Lowdensities of sea urchins influence the structure of algal assemblagesin the western Mediterranean. J. Sea. Res. 39, 281–290.
innegar, J.K., Polunin, N.V.C., 2004. predicting indirect effects of fishingin Mediterranean rocky littoral communities using a dynamic simula-tion model. Ecol. Model. 172, 249–267.
innegar, J.K., Polunin, N.V.C., Francour, P., Badalamenti, F., Chemello,R., Harmelin-Vivien, M.L., Hereu, B., Milazzo, M., Zabala, M.,D’Anna, G., Pipitone, C., 2000. Trophic cascades in benthic marineecosystems: lessons for fisheries and protected-area management. Env-iron. Conserv. 27, 179–200.
egis, M.B., 1988. Impact et resultats de la Campagne “Sauvon l’oursincomestible: Paracentrotus lividus” (Echinodermata, Echinoida). In:Regis, M.B. (Ed.), Actes du VI Seminaire International sur les Echin-odermes. Ile des Embiez, 11–22. Var-France, pp. 150–157.
ogers-Bennett, L., Pearse, J.S., 2001. Indirect benefits of marine pro-tected areas for juvenile abalone. Cons. Biol. 15, 642–647.
os, J., Niell, F.X., 1981. Algunos datos biometricos de Paracentrotuslividus (Lmk.) Arbacia lixula (L. ) y Sphaerechinus granularis (Lmk.).Oecol. Aquat. 5, 227–231.
ala, E., 1997. Fish predators and scavengers of the sea urchin Paracen-trotus lividus in protected areas of the north-west Mediterranean Sea.Mar. Biol. 129, 531–539.
ala, E., Zabala, M., 1996. Fish predation and the structure of the seaurchin Paracentrotus lividus population in the NW Mediterranean.Mar. Ecol. Prog. Ser. 140, 71–81.
ala, E., Boudouresque, C.F., Harmelin-Vivien, M., 1998. Fishing, trophiccascades, and the structure of algal assemblages: evaluation of an oldbut untested paradigm. Oikos 82, 425–439.
cheibling, R.E., 1996. The role of predation in regulating sea urchinpopulations in eastern Canada. Oceanol. Acta 19, 421–430.
hears, N.T., Babcock, R.C., 2003. Continuing trophic cascade effectsafter 25 years of no take marine reserve protection. Mar. Ecol. Prog.Ser. 246, 1–16.
W
W
esearch 81 (2006) 37–44
teneck, R.S., 1998. Human influences on coastal ecosystem: doesoverfishing create trophic cascades? Trends Ecol. Evol. 13, 429–430.
arr, R.J.Q., Williams, P.V.G., McKenzie, A.J., 1996. Abalone, sea urchinsand rock lobster: a possible ecological shift that may affect traditionalfisheries. S. Afr. J. Mar. Sci. 17, 319–323.
egner, M.J., Dayton, P.K., 1977. Sea urchin recruitment patterns andimplications of commercial fishing. Science 196, 324–326.
egner, M.J., Dayton, P.K., 1999. Ecosystem effects of fishing. TrendsEcol. Evol. 14, 261–262.
urner, S.J., Thrush, S.F., Hewitt, J.E., Cummings, V.J., Funnell, G., 1999.Fishing impacts and the degradation or loss of habitat structure. Fish.Manage. Ecol. 6, 40–420.
uron, X., Giribert, G., Lopez, S., Palacın, C., 1995. Growth andpopulation structure of Paracentrotus lividus (Echinodermata: Echi-noidea) in two contrasting habitats. Mar. Ecol. Prog. Ser. 122, 193–204.
uya, F., Jerez, P.S., Haroun, R.J., 2005. Influence of fishing and func-tional group of algae on sea urchin control of algal communities inthe eastern Atlantic. Mar. Ecol. Prog. Ser. 287, 255–260.
nderwood, A.J., 1994. On beyond BACI: sampling designs that mightreliably detect environmental disturbances. Ecol. Appl. 4, 3–15.
nderwood, A.J., 1997. Experiments in Ecology. Their Logic Designand Interpretation Using Analysis of Variance Cambridge UniversityPress, Cambridge.
erlaque, M., 1987. Relations entre Paracentrotus lividus (Lamarck)et le phytobenthos de Mediterranee occidentale. In: Boudoresque,C.F. (Ed.), Colloque International sur Paracentrotus lividus et lesOursins Comestibles. GIS Posidonie Publications, Marseille, pp. 5–36.
erlaque, M., Nedelec, H., 1983. Biologie de Paracentrotus lividus(Lamarck) sur substratum rocheux en Corse (Mediterranee France):alimentation des adultes. Vie Milieu 33, 191–202.
ielmini, I., Chiantore, M., Gianguzza, P., Bonaviri, C., Cattaneo-ViettiR., Riggio, S., 2005. Protection effects on feeding and reproduc-tion of Paracentrotus lividus and Arbacia lixula on barren groundsat Ustica Island MPA (Western Mediterranean, Italy). Abstract15 Congresso Nazionale S.It.E. Torino, 12–14 Settembre 2005,p. 145.
atling, L., Norse, E.A., 1998. Effects of mobile fishing gear on marinebenthos. Cons. Biol. 12, 1178–1179.
illan, R.C., Dollimore, J.M., Nicholson, J., 1979. A survey of fish pop-ulations at Karikari Peninsula, Northland, by scuba diving. NZ J. Mar.Fresh. Res. 13, 447–458.
















