
Response of bacterioplankton to iron fertilization in the Southern Ocean
Upload
independentCategory
view
2download
0
Importance of Spatial Population Characteristics onthe Fertilization Rates of Sea Urchins
JEAN-SEBASTIEN LAUZON-GUAY1,* AND ROBERT E. SCHEIBLING2
1Biology Department, University of New Brunswick, Bag Service 45111, Fredericton, New BrunswickE3B 6E1, Canada; and 2Department of Biology, Dalhousie University, Halifax,
Nova Scotia B3H 4J1, Canada
Abstract. We show that inclusion of population charac-teristics in coupled advection-diffusion and fertilization-kinetics models results in higher fertilization rates thanthose previously reported in theoretical studies. We incor-porate parameters related to both individuals and popula-tions by running simulations over a large spatial scale andincorporating sperm contribution from multiple males. Wecompare predictions for three subpopulations of the seaurchin Strongylocentrotus droebachiensis (those occupyingkelp beds, barrens, and grazing fronts) to observations fromsmall-scale experiments, and estimate effects of populationsize and current velocity in each subpopulation. Modeloutputs suggest that fertilization rates are low in kelp beds,intermediate in barrens, and high in grazing fronts. In allpopulations, increasing current velocity has a negative ef-fect on the relationship between fertilization rate and down-stream distance of gametes after release, but no effect on therelationship between fertilization rate and elapsed timesince gamete release. Our model output was most sensitiveto changes in the number of spawning males and the spermrelease rate, suggesting that spawning synchrony and highgonadic index could greatly increase the fertilization suc-cess in sea urchins.
Introduction
Fertilization success of sea urchins and other free-spawn-ing benthic invertebrates has been the focus of many em-pirical and theoretical studies over the last 25 years. Inparticular, a seminal paper by Denny and Shibata (1989)
provided a mathematical framework for predicting fertiliza-tion rates under field conditions by combining the steady-state solution of the turbulent advection-diffusion equations(Csanady, 1973) with the fertilization kinetics model devel-oped by Vogel et al. (1982). That model, and variations ofit, enabled researchers to examine various hypotheses aboutdeterminants of fertilization success, including the size andreproductive output of spawning individuals (Babcock etal., 1994; Levitan and Young, 1995; Claereboudt, 1999),the distances between them (Denny and Shibata, 1989;Levitan and Young, 1995; Claereboudt, 1999), and thehydrodynamic environment during spawning (Denny andShibata, 1989; Young et al., 1992; Levitan and Young,1995). The original formulation of the model has beenprogressively modified to approach field conditions, mostcommonly by incorporating terms for reflection of gametesat the seabed (Claereboudt, 1999) and water surface (Bab-cock et al., 1994), and the blocking effect of polyspermy(Styan, 1998; Millar and Anderson, 2003). Attempts atvalidating the Denny and Shibata model (and its deriva-tives) by means of small-scale manipulative field experi-ments with sea urchins and sea stars have generally metwith limited success (Denny and Shibata, 1989; Levitan andYoung, 1995; Metaxas et al., 2002).
Early applications of that model indicated that spermconcentration is likely a major limiting factor, resulting inlow fertilization rates except at very high population den-sity. Experimental work with sea urchins subsequently con-firmed that even at densities of 144 urchins per squaremeter, we could expect fertilization rates below 70%(Wahle and Peckham, 1999). Up to this point, most theo-retical (Denny, 1988; Denny and Shibata, 1989; Young etal., 1992; Metaxas et al., 2002; but see Levitan and Young,1995) and empirical (Pennington, 1985; Levitan, 1991;
Received 27 October 2006; accepted 13 February 2007.* To whom correspondence should be addressed, at Department of
Biology, Dalhousie University, Halifax, Nova Scotia B3H 4J1, Canada.E-mail: [email protected]
Reference: Biol. Bull. 212: 195–205. (June 2007)© 2007 Marine Biological Laboratory
195

Wahle and Peckham, 1999; but see Levitan et al., 1992)studies had been based on a single or a very few spawningmales that contributed to sperm plumes likely to fertilizeeggs. This reinforced the notion of sperm limitation innatural populations without necessarily being representativeof such situations. Ignoring population size in experimentalstudies also has been somewhat misleading. For example,Wahle and Peckham (1999) did not vary the number ofmales in their various “density” trials, but changed thedistance between the males and egg batches, falsely assum-ing that only the closest male could contribute to fertiliza-tion. Further, because eggs were enclosed in baskets, fertili-zation was limited to males spawning upstream from thebasket, as these eggs could not encounter downstreammales. The results of that study were nonetheless assumed tobe representative and have been used to predict fertilizationsuccess in wild populations (Meidel and Scheibling, 2001).
Modeling work also led to another assumption: that mostfertilization occurs very rapidly once eggs have been re-leased (Denny and Shibata, 1989; Levitan and Young,1995). Again, early modeling work focused on the fertili-zation of eggs emanating from a female directly down-stream of a single spawning male (Denny and Shibata,1989; Meidel, 1999). Given that the advection-diffusionmodel predicts a rapid decrease in sperm concentration assperm are transported downstream, eggs that are not fertil-ized rapidly are soon within much diluted sperm and thelikelihood of fertilization drops. Therefore, because themodel assumes that sperm come from upstream males,fertilization rates are obviously greater in the immediatevicinity of the female (as close to the spawning male aspossible), before eggs are advected from the source. Giventhe longevity of eggs (Meidel and Yund, 2001), however,they could eventually encounter a spawning male as theyare transported downstream, where high fertilization ratesalso could occur. Although informative, single-male modelsdo not provide much insight into fertilization as it wouldproceed in nature. This weakness highlights the importanceof including parameters that are relevant to the populationas a whole rather than to the individuals.
Population parameters were included in a model by Levi-tan and Young (1995) to predict fertilization rate in theechinoid Clypeaster rosaceus. They simulated sperm cloudsresulting from multiple males randomly distributed on two-dimensional grids of varying size and examined the fertili-zation rates along the downstream axis from a spawningfemale. They demonstrated that population density is im-portant, but that a large population size could compensatefor a low density. Claereboudt (1999) used a similar model,but in three dimensions, to examine the effect of density andaggregation on fertilization rate of sea urchins. His modelaccounts for diffusion of eggs, but not for the fact that acertain proportion of these eggs will already have beenfertilized before they reach a given point downstream of the
female. Instead, he assumed that the proportion of eggsremaining unfertilized in any cell of his array was relatedonly to fertilization in that particular cell. He obtainedresults similar to those of Levitan and Young (1995): den-sity is important—but for a given population size, fertiliza-tion rate will increase with the degree of aggregation.
Early field experiments did not yield high fertilizationrates, which reinforced the conclusion of low fertilizationsuccess indicated by most models. Meidel and Yund (2001)criticized the short duration of field experiments, indicatingthat fertilization in the laboratory can occur for several hoursafter spawning, whereas most field studies are on the order ofminutes. Recent studies have shown that time-integrated (2–4days) fertilization rates in the field can reach as high as 100%(Wahle and Gilbert, 2002; Gaudette, 2004). These findings castdoubt on the reliability of short-term experiments as indicatorsof fertilization success in natural populations.
Our study attempts to reconcile empirical evidence forhigh fertilization rates observed in the field with the tradi-tional Denny and Shibata (1989) model of advection-diffu-sion and fertilization kinetics. To do so, we incorporateparameters related to individuals as well as to the populationas a whole by running simulations over a large spatial scale,and by incorporating sperm contribution from multiplemales. We compare predictions of our model for threesubpopulations of the green sea urchin Strongylocentrotusdroebachiensis Muller 1776—namely those occurring in(1) kelp beds (low urchin density and high biomass of erectmacroalgae), (2) barrens (intermediate urchin density andabsence of erect macroalgae), and (3) grazing fronts (high-density aggregations of urchins at the kelp bed-barrensinterface)—to previous observations in small-scale experi-ments. We then estimate how fertilization rates are affectedby population size and current velocity in each subpopula-tion, and evaluate parameters most likely to influence thefertilization success.
Materials and Methods
Fertilization model
To predict the concentration of sperm from a single malein a three-dimensional simulation array, we start with thesteady-state solution to the turbulent diffusion-advectionequation (Csanady, 1973; Denny, 1988):
S� x,y,z� �Qsu�
2��y�zu *2x 2 �exp � � y2u�2
2�y2u*
2x2 ��z � s�2u�2
2�z2u*
2x2 �� exp � � y2u�2
2�y2u*
2x2 ��z � s�2u�2
2�z2u*
2x2 �� (1)
where S is the concentration of sperm (m�3) from one maleat distance x, y, and z from the male, Qs is the sperm releaserate per urchin (s�1), u� is the mean current velocity (m �
196 J-S. LAUZON-GUAY AND R. E. SCHEIBLING

s�1), u* is the frictional velocity (m � s�1), s is the height atwhich sperm is released (m), and �y and �z are respectivelythe horizontal and the vertical coefficients relating frictionvelocity to directional diffusivity. This equation assumesthat gametes are reflected at the bottom (z � 0) and lostabove some value of z. We expanded this equation toincorporate a reflective boundary at a height D (D � 12 m,the average water depth at our field sites) by adding a mirrorsource at the air-water boundary to prevent loss of eggs andsperm at this interface (Babcock et al., 1994). To simulatea large population size in the cross-current direction, weincorporated reflective boundaries in the y-dimension byadding mirror sources on both sides of our simulation grid:
S� x,y,z� �Qsu�
2��y�zu*2x2�exp � � y2u�2
2�y2u*
2x2� � exp
� ��2Y1 � y�2u�2
2�y2u*
2x2 � � exp � ��2Y2 � y�2u�2
2�y2u*
2x2 ���exp � ��z � s�2u�2
2�z2u*
2x2 � � exp � ��z � s�2u�2
2�z2u*
2x2 � � exp
� ��2D � z � s�2u�2
2�z2u*
2x2 �� (2)
where D is the depth of the water column (m), and Y1 andY2 represent the distances (m) between an urchin and theedges (in the y-dimension) of our simulation array (seebelow). Estimates of parameters relating to turbulent diffu-sion are difficult to evaluate. For simplicity we assumed that
�y � 2.25 and �z � 1.25 (Denny and Shibata, 1989), whileu* was set at 10% of the mean current velocity (u�) as isgenerally assumed (Levitan and Young, 1995). Diffusion inthe x-dimension is negligible compared to advection, andtherefore excluded from the model.
To account for multiple males spawning synchronously,we randomly assigned x- and y-coordinates to a number ofmale urchins (n) in our simulation array (the z-coordinate ofeach male was set to 0 since adult urchins are on the seabed). Density of males was chosen to be representative ofeach of our three subpopulations (Table 1). Our simulationarray measured 200 m by 20 m by 12 m (x: parallel to flow,y: horizontal and perpendicular to flow, z: vertical) dividedinto 1,920,000 cells measuring 0.1 m by 0.5 m by 0.5 m(2000 by 40 by 24 cells). These dimensions for the cells ofthe arrays provided a good compromise between computa-tion time and loss of accuracy; any further decrease in thecell size resulted in less than 1% change in the modeloutput. We can sum the concentration of sperm obtainedfrom Equation 2 over the total number of males (n) to obtainthe total sperm concentration (ST):
ST� x,y,z� � �i�1
n
Si� x � xi,y � yi,z� (3)
where xi and yi are the x- and y-coordinates of male i in thearray.
Fertilization kinetics has traditionally been described bythe model developed by Vogel et al. (1982):
Table 1
Description and values of variables used for our three subpopulations (KB: Kelp bed, PB: Permanent barrens, GF: Grazing front) of urchins(Strongylocentrotus droebachiensis)
Variable name Symbol Unit All Value KB PB GF
Horizontal diffusion coefficient1 �y — 2.25 — — —Vertical diffusion coefficient1 �z — 1.25 — — —Sperm release rate2 Qs s�1 — 1.9 � 107 2.7 � 107 2.9 � 107
Current velocity u� m � s�1 0.05, 0.10, or 0.20 — — —Frictional velocity1 u* m � s�1 0.1 u� — — —Population density4 N m�2 — 14 71 136Density of spawning male4 — m�2 — 0.67 3.87 18.40Number of spawning males in array n 4000 m�2 — 2680 15480 73600Height of gamete release s m 0.05 — — —Fertilizable surface area of an egg3 � m2 1.65 � 10�10 — — —Depth of water column D m 12 — — —Length of cell in flow direction c m 0.5 — — —Egg concentration emanating from a female E m�3 — — — —Sperm concentration emanating from one male S m�3 — — — —Total sperm concentration from all males in array ST m�3 — — — —Percentage of eggs fertilized F % — — — —Percentage of eggs fertilized in plane j Zj % — — — —Sperm-egg contact time — s — — — —
1 Denny and Shibata, 1989; 2 Meidel, 1999; 3 Levitan, 1993; 4 Meidel and Scheibling, 2001.
197FERTILIZATION IN SEA URCHIN

F� x,y,z� � 1 � exp� � ��u*ST�x,y,z�� (4)
where F is the percentage of eggs fertilized (%), � is thefertilizable surface area of an egg (m2), and � is the sperm-egg contact time (s) (i.e., the amount of time spent in aspecific cell). The fertilizable area of an egg is 1% of the eggcross-sectional area (Vogel et al., 1982) calculated from theegg diameter (1.45 � 10�4 m for Strongylocentrotus droe-bachiensis; Levitan, 1993). The egg-sperm contact time (�)is equal to the time spent by sperm and eggs in each cell,which is a function of the current velocity (m � s�1) and thelength (m) of the cell in the flow direction (c):
� � c/u� (5)
Because the fertilization rate does not depend on egg con-centration, we can calculate the potential fertilization ratefor each cell of our simulation area. First, we calculated theproportion of eggs that could be fertilized (P) in any verticalplane j along x, if all eggs emanating from a female reacheda plane j in an unfertilized state:
Pj �
�y�0
40 �z�0
24
F� j,y,z� E� j,y,z�
�y�0
40 �z�0
24
E� j,y,z�
(6)
where E(j,y,z) is the number of eggs (fertilized and virgin)in each cell of the plane j emanating from a single femalelocated at a given position on the x-axis. The number ofeggs from a female in each cell is obtained using Equation2, and by replacing S by E and Qs by 1. From Equation 4,the fertilization success is independent from the concentra-tion of eggs, and therefore we scaled the egg release rate to1. From Equation 6, we can calculate the proportion ofunfertilized eggs (1 � Pj) in a plane that can be transportedto the next plane (j � 1). By assuming that mixing occursbetween each time step, we can account for a reducednumber of unfertilized eggs reaching planes farther awayfrom the female, because of fertilization, by calculating theproportion of eggs not already fertilized in a previous planethat reach plane j, and are fertilized there, as:
Zj � �k�l:
j
�1 � Pk� (7)
Where � is the arithmetic product from k equals 1 to j. Toobtain the total proportion of eggs fertilized from release toany distance d from the female, we can sum Zj from 0 to d.
Fertilization success in different subpopulations
We compared fertilization rates of eggs spawned by asingle female (located at x � 0, y � 10, and z � 0 in our
array) among three subpopulations of urchins (kelp bed,barrens, and grazing front) at low current velocity (0.05 m �s�1). Urchin density is generally the greatest in grazingfronts and lowest in kelp beds, while the barrens haveintermediate densities (Meidel and Scheibling 2001). Ur-chins at a grazing front also are generally larger than thosein the barrens or the kelp beds. We used results from aliterature survey of urchins in these subpopulations (Meideland Scheibling, 2001) to parameterize each simulation (Ta-ble 1). The only variable not available from this study is thesperm release rate per urchin in each subpopulation. For thatvariable we used values from Meidel (1999), who calculatedsperm release rate (Qs) as:
Qs �G
�(8)
where G is the dry weight of sperm release (g), is thespawning time (s) and � is the dry weight per sperm (3.66 �10�12 for S. droebachiensis; Thompson, 1979). Mass ofsperm released per urchin and sperm release time werecalculated from results of field and laboratory experiments(Meidel, 1999). Sperm release rate per urchin is higher ingrazing fronts than in barrens or kelp beds (Table 1).
Effect of environmental factors on fertilization success
We used parameters from the three subpopulations (Table1) to evaluate the effect of current velocity (0.05, 0.10, and0.20 m � s�1) on the fertilization rate of eggs spawned by asingle female located at x � 0 and y � 10 (on the upstreamboundary and 10 m from each along-stream boundary).Although we recognize that kelp beds affect current velocityand modify flow characteristics (Wahle and Peckham,1999), we do not incorporate a decrease in current velocityin the kelp bed. This decrease would change above the frondcanopy and Equation 2 would not be valid. Although ourmodel limits a direct comparison of fertilization rate withinand outside of kelps beds under a given ambient flowregime, it does provide estimates of fertilization success fora kelp bed subpopulation over the same range of flow ratesas that applied to the other subpopulations. For the sake ofsimplicity, we assume that current is unidirectional (flowingfrom x � 0 to x � 200 in our array) and horizontallyuniform in our simulation for all subpopulations.
We also evaluated the effect of the position of the femalewithin the population (x � 0, 25, and 50 m from theupstream boundary) on the fertilization success in eachsubpopulation, using a current velocity of 0.05 m � s�1.Clear boundaries in natural populations are not to be ex-pected (except for a sharp discontinuity at the kelp bed-barren interface), but this allows us to estimate the effect ofpopulation size on fertilization rates. A female located atx � 0 and y � 10 would have no male upstream and eggscould be fertilized only by downstream males, whereas eggs
198 J-S. LAUZON-GUAY AND R. E. SCHEIBLING
released at x � 25 and 50 m could be fertilized by malesspawning both upstream and downstream from the point ofegg release.
Elasticity analysis
Elasticity analysis is a type of sensitivity analysis thatuses small proportional perturbations to measure the re-sponse of the model to changes in parameter values (Cas-well, 2001). The elasticity () of the model prediction tosmall (10%) increases in parameter (p) values was calcu-lated as:
p � 100Xp � X0
X0(9)
where X0 is the model output given the original value ofparameter p, while Xp is the model output given modifiedparameter p (Barbeau and Caswell, 1999). We used distanceto reach 90% fertilization (the highest rate reached in allsubpopulations after 200 m) as the model output for thisanalysis. Sperm release rate (Qs), current velocity (u�), fric-tional velocity (u*), the coefficients �y and �z, the fertiliz-able area of an egg (�), and the number of spawning males(n) were parameters that were used to examine the model’selasticity.
Results
Fertilization success in different subpopulations
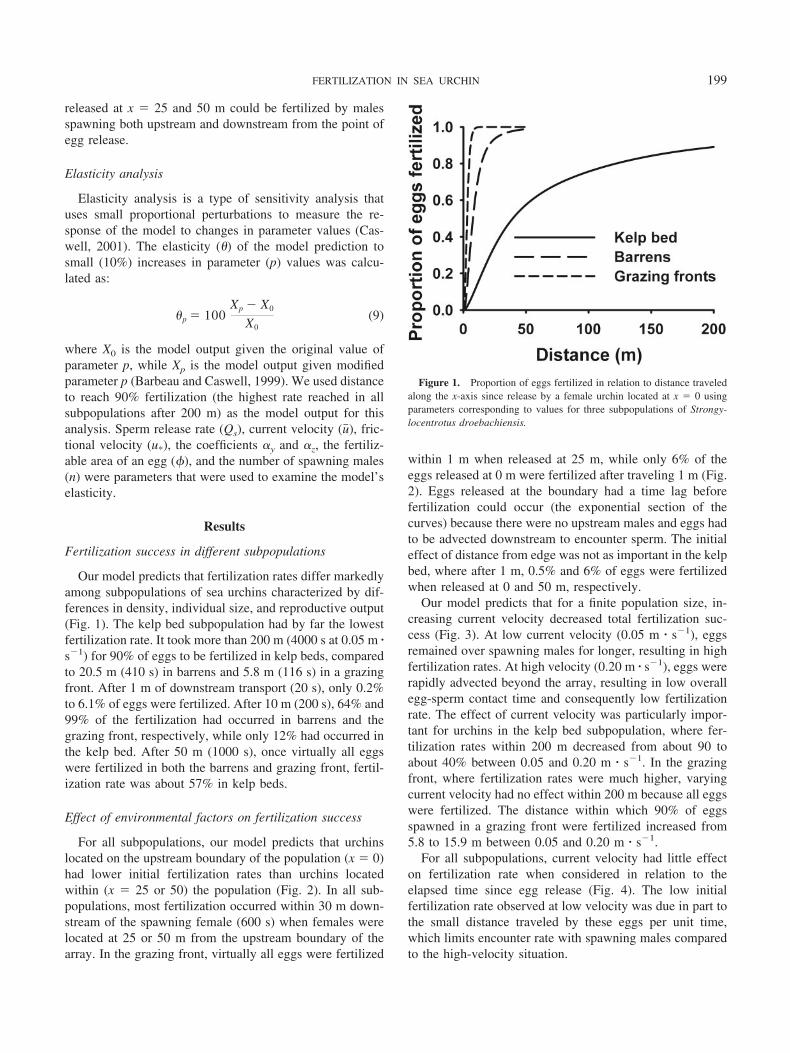
Our model predicts that fertilization rates differ markedlyamong subpopulations of sea urchins characterized by dif-ferences in density, individual size, and reproductive output(Fig. 1). The kelp bed subpopulation had by far the lowestfertilization rate. It took more than 200 m (4000 s at 0.05 m �s�1) for 90% of eggs to be fertilized in kelp beds, comparedto 20.5 m (410 s) in barrens and 5.8 m (116 s) in a grazingfront. After 1 m of downstream transport (20 s), only 0.2%to 6.1% of eggs were fertilized. After 10 m (200 s), 64% and99% of the fertilization had occurred in barrens and thegrazing front, respectively, while only 12% had occurred inthe kelp bed. After 50 m (1000 s), once virtually all eggswere fertilized in both the barrens and grazing front, fertil-ization rate was about 57% in kelp beds.
Effect of environmental factors on fertilization success
For all subpopulations, our model predicts that urchinslocated on the upstream boundary of the population (x � 0)had lower initial fertilization rates than urchins locatedwithin (x � 25 or 50) the population (Fig. 2). In all sub-populations, most fertilization occurred within 30 m down-stream of the spawning female (600 s) when females werelocated at 25 or 50 m from the upstream boundary of thearray. In the grazing front, virtually all eggs were fertilized
within 1 m when released at 25 m, while only 6% of theeggs released at 0 m were fertilized after traveling 1 m (Fig.2). Eggs released at the boundary had a time lag beforefertilization could occur (the exponential section of thecurves) because there were no upstream males and eggs hadto be advected downstream to encounter sperm. The initialeffect of distance from edge was not as important in the kelpbed, where after 1 m, 0.5% and 6% of eggs were fertilizedwhen released at 0 and 50 m, respectively.
Our model predicts that for a finite population size, in-creasing current velocity decreased total fertilization suc-cess (Fig. 3). At low current velocity (0.05 m � s�1), eggsremained over spawning males for longer, resulting in highfertilization rates. At high velocity (0.20 m � s�1), eggs wererapidly advected beyond the array, resulting in low overallegg-sperm contact time and consequently low fertilizationrate. The effect of current velocity was particularly impor-tant for urchins in the kelp bed subpopulation, where fer-tilization rates within 200 m decreased from about 90 toabout 40% between 0.05 and 0.20 m � s�1. In the grazingfront, where fertilization rates were much higher, varyingcurrent velocity had no effect within 200 m because all eggswere fertilized. The distance within which 90% of eggsspawned in a grazing front were fertilized increased from5.8 to 15.9 m between 0.05 and 0.20 m � s�1.
For all subpopulations, current velocity had little effecton fertilization rate when considered in relation to theelapsed time since egg release (Fig. 4). The low initialfertilization rate observed at low velocity was due in part tothe small distance traveled by these eggs per unit time,which limits encounter rate with spawning males comparedto the high-velocity situation.
Figure 1. Proportion of eggs fertilized in relation to distance traveledalong the x-axis since release by a female urchin located at x � 0 usingparameters corresponding to values for three subpopulations of Strongy-locentrotus droebachiensis.
199FERTILIZATION IN SEA URCHIN
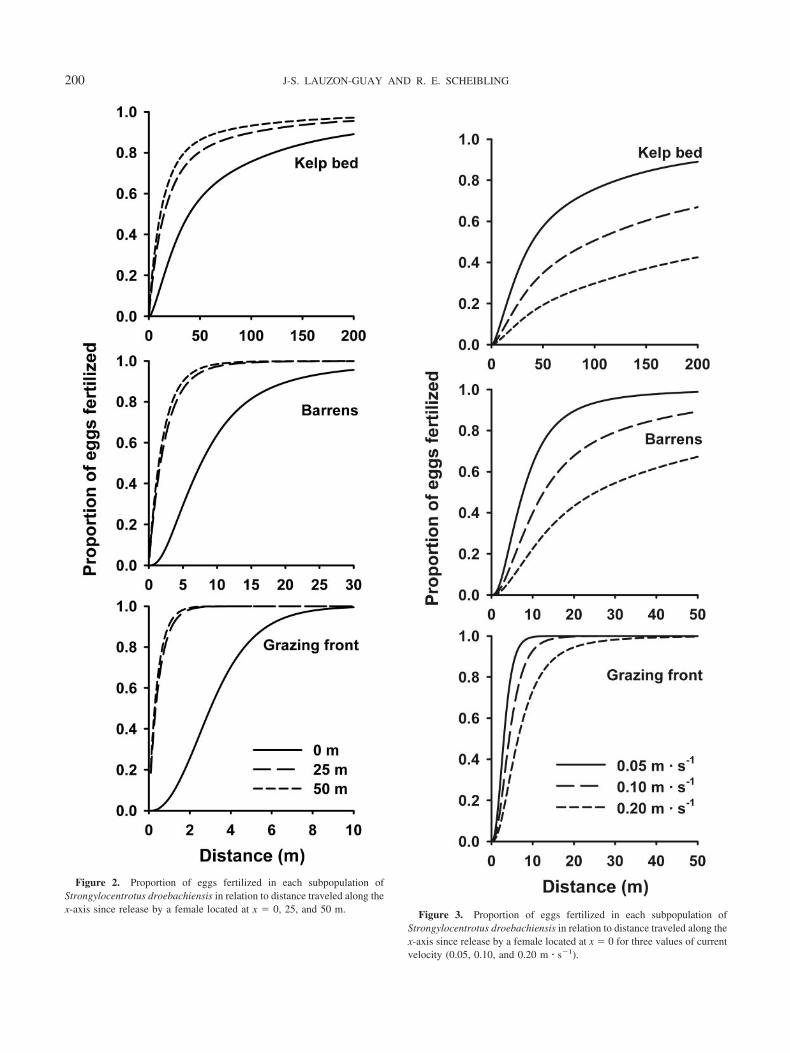
Figure 2. Proportion of eggs fertilized in each subpopulation ofStrongylocentrotus droebachiensis in relation to distance traveled along thex-axis since release by a female located at x � 0, 25, and 50 m. Figure 3. Proportion of eggs fertilized in each subpopulation of
Strongylocentrotus droebachiensis in relation to distance traveled along thex-axis since release by a female located at x � 0 for three values of currentvelocity (0.05, 0.10, and 0.20 m � s�1).
200 J-S. LAUZON-GUAY AND R. E. SCHEIBLING

Elasticity analysis
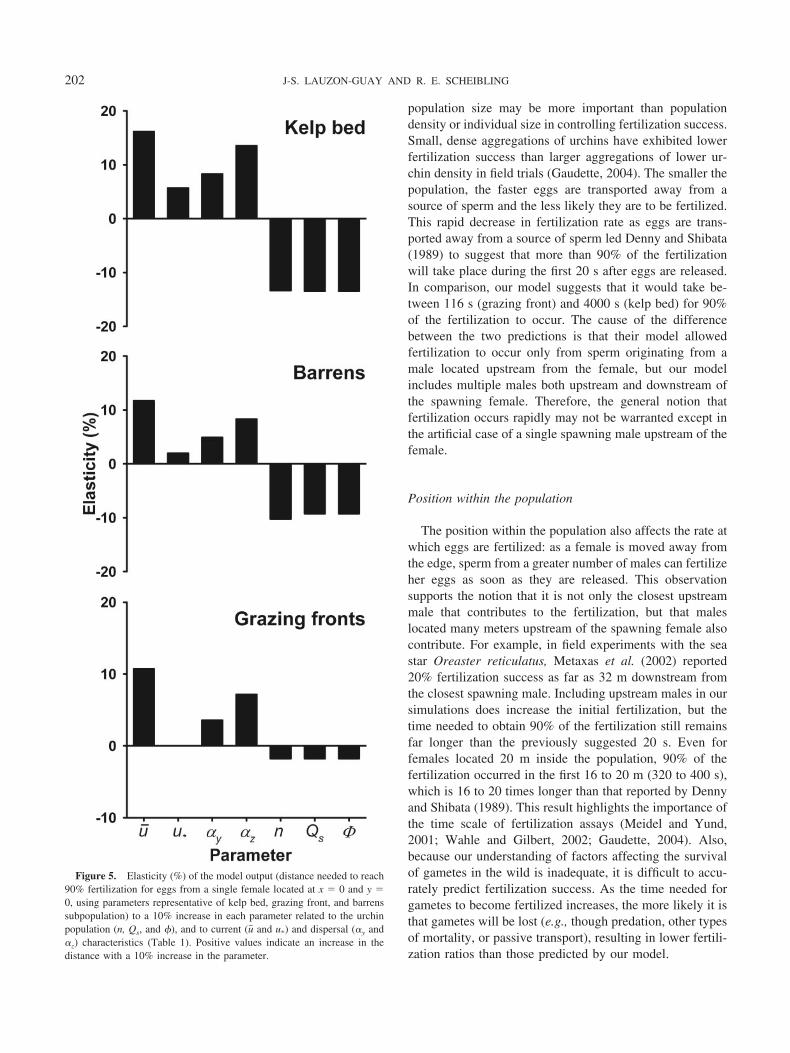
The distance needed to reach 90% fertilization (the modeloutput we used) was most sensitive to changes in parametervalues in the kelp beds, intermediate in the barrens, andfairly robust in the grazing front (Fig. 5). Increasing thedensity of spawning males, the sperm release rate, and thefertilizable area of an egg by 10% all decreased the distance(and thus increase fertilization rate) by 1.8% to 13.5%,whereas increasing hydrodynamic parameters that affectgamete dispersal (u� , u*, �y and �z) increased the distance(i.e., decrease fertilization rate) by as much as 16.2%. Themodel output was most sensitive in all subpopulations to anincrease in current velocity, which increased the distanceneeded to reach 90% fertilization success by 10.7% to16.2%.
Discussion
Fertilization success in different subpopulations
Our model predicts higher fertilization rates than thosereported by Meidel and Scheibling (2001) using the samepopulation parameters. On the basis of results of a fieldexperiment by Wahle and Peckham (1999), Meidel andScheibling (2001) predicted fertilization rates of 14.6%,43.3%, and 62.0% in kelp bed, barrens, and grazing frontsubpopulations, respectively. Our model predicts these val-ues after eggs are advected for only 3.6, 6.7, and 11.5 mrespectively. These distances are negligible given the spatialextent of populations of Strongylocentrotus droebachiensis,which can span hundreds of kilometers of coastline andextend hundreds of meters to kilometers offshore (e.g.,Wharton and Mann; 1981, Moore and Miller, 1983; Bradyand Scheibling, 2005). These results suggest that the Meideland Scheibling model underestimates fertilization successby using Wahle and Peckham’s (1999) data from small-scale field experiments. However, our model assumes thatmixing occurs between each time step (equation 7), inflatingthe probability of an egg being fertilized. Nonetheless,given the rapid and high level of fertilization obtained in oursimulations, this assumption would likely not affect thegeneral behavior of our results.
Our model uses a specific spatial area occupied by asubpopulation (200 m by 20 m). The actual area occupiedby each subpopulation in nature likely varies between sub-populations and would be difficult to estimate. The size ofthe grazing front (2 m wide; Lauzon-Guay and Scheibling,2007) would always be smaller than our simulation array;nevertheless, because our model predicts greater than 90%fertilization success in the first few meters after spawning,the spatial extent of our array will have little effect on themodel prediction for the grazing front. The spatial extent ofa barren or kelp bed subpopulation will have a greater effecton the fertilization success of eggs spawned. In these cases,
Figure 4. Proportion of eggs fertilized in each subpopulation ofStrongylocentrotus droebachiensis in relation to time spent in water col-umn since release by a female located at x � 0 for three values of currentvelocity (0.05, 0.10, and 0.20 m � s�1).
201FERTILIZATION IN SEA URCHIN
population size may be more important than populationdensity or individual size in controlling fertilization success.Small, dense aggregations of urchins have exhibited lowerfertilization success than larger aggregations of lower ur-chin density in field trials (Gaudette, 2004). The smaller thepopulation, the faster eggs are transported away from asource of sperm and the less likely they are to be fertilized.This rapid decrease in fertilization rate as eggs are trans-ported away from a source of sperm led Denny and Shibata(1989) to suggest that more than 90% of the fertilizationwill take place during the first 20 s after eggs are released.In comparison, our model suggests that it would take be-tween 116 s (grazing front) and 4000 s (kelp bed) for 90%of the fertilization to occur. The cause of the differencebetween the two predictions is that their model allowedfertilization to occur only from sperm originating from amale located upstream from the female, but our modelincludes multiple males both upstream and downstream ofthe spawning female. Therefore, the general notion thatfertilization occurs rapidly may not be warranted except inthe artificial case of a single spawning male upstream of thefemale.
Position within the population
The position within the population also affects the rate atwhich eggs are fertilized: as a female is moved away fromthe edge, sperm from a greater number of males can fertilizeher eggs as soon as they are released. This observationsupports the notion that it is not only the closest upstreammale that contributes to the fertilization, but that maleslocated many meters upstream of the spawning female alsocontribute. For example, in field experiments with the seastar Oreaster reticulatus, Metaxas et al. (2002) reported20% fertilization success as far as 32 m downstream fromthe closest spawning male. Including upstream males in oursimulations does increase the initial fertilization, but thetime needed to obtain 90% of the fertilization still remainsfar longer than the previously suggested 20 s. Even forfemales located 20 m inside the population, 90% of thefertilization occurred in the first 16 to 20 m (320 to 400 s),which is 16 to 20 times longer than that reported by Dennyand Shibata (1989). This result highlights the importance ofthe time scale of fertilization assays (Meidel and Yund,2001; Wahle and Gilbert, 2002; Gaudette, 2004). Also,because our understanding of factors affecting the survivalof gametes in the wild is inadequate, it is difficult to accu-rately predict fertilization success. As the time needed forgametes to become fertilized increases, the more likely it isthat gametes will be lost (e.g., though predation, other typesof mortality, or passive transport), resulting in lower fertili-zation ratios than those predicted by our model.
Figure 5. Elasticity (%) of the model output (distance needed to reach90% fertilization for eggs from a single female located at x � 0 and y �0, using parameters representative of kelp bed, grazing front, and barrenssubpopulation) to a 10% increase in each parameter related to the urchinpopulation (n, Qs, and �), and to current (u� and u*) and dispersal (�y and�z) characteristics (Table 1). Positive values indicate an increase in thedistance with a 10% increase in the parameter.
202 J-S. LAUZON-GUAY AND R. E. SCHEIBLING

Current velocity
Previous models (Denny and Shibata, 1989; Levitan andYoung, 1995) predicted that current velocity has a negativeeffect on fertilization rate, especially when that rate is lowfor other reasons. Similarly, although he did not test this inhis simulations, Claereboudt (1999) suggested that currentvelocity would have an effect opposite to that of spermrelease rate in his model because sperm concentration isproportional to sperm release rate but inversely proportionalto current velocity (Equation 1). However, according to ourfertilization kinetic model (Equation 4), fertilization rateshould be independent of current velocity for an infinitelylarge population. Any decrease in sperm concentration dueto increased current velocity is compensated by a propor-tional increase in frictional velocity, such that the exponen-tial term of the fertilization equation remains unchanged(i.e., S � u��1, u* � u� , and F � 1 � exp (u*S)). While theconcentration of sperm decreases with increasing currentvelocity, the proportion of eggs fertilized for a constantsperm concentration also decreases. However, because egg-sperm contact time (�), which is inversely proportional tothe current velocity (Equation 4), decreases as fast as fric-tional velocity (u*) increases with current velocity, the fer-tilization rate in each cell will decrease with current veloc-ity. This results in a lower fertilization rate after a givendistance, because eggs in high current velocity will travelthat distance more rapidly. However, if we look at theproportion of eggs fertilized as a function of time since theywere spawned, we obtain very similar results across a rangeof current velocities (Fig. 4). This differs from the model ofLevitan and Young (1995) because our fertilization rate is afunction of frictional velocity whereas theirs is a function ofsperm swimming speed. Sperm swimming speed (0.088mm � s�1; Levitan 1993) is orders of magnitude lower thanfrictional velocity (0.1 u�), which is more generally used tocalculate the probability of a sperm-egg encounter.
As previously mentioned, fertilization success is largely aconsequence of whether or not eggs are carried away fromthe spawning population before being fertilized, which inturn is determined by the size of the population and thecurrent velocity. Even when individuals are small and at lowurchin density, and the current is strong, eggs will have ahigh probability of eventually being fertilized if the popu-lation is large. This conclusion is supported by recent evi-dence extending the period of egg viability in S. droe-bachiensis to days after spawning (Meidel and Yund, 2001),compared to previous estimates that ranged in hours (Epel etal., 1998; Wahle and Peckham, 1999). The possibility thateggs are initially retained on the aboral surface of the femaleurchin because of their high viscosity (Thomas, 1994: Yundand Meidel, 2003) also may increase fertilization success(Bishop, 1998) by subjecting eggs to high sperm concen-trations for a longer period before they are advected from
the population. Information on retention time of eggs undervarying flow conditions, and the extent of sperm penetrationof the egg matrix, is needed to evaluate how retentionaffects fertilization success, but it would most likely de-crease the importance of population size.
We intentionally modeled gamete dispersion using a sim-plified version of the flow dynamics observed in nature. Thechange in flow characteristics within a kelp bed was notincluded in our model, although it likely has an importanteffect on fertilization success (Wahle and Peckham, 1999).A reduction in flow velocity below the canopy could resultin fertilization occurring over a shorter distance; however,the time required for fertilization to occur may not beaffected. We focused our model on the fertilization in uni-directional flow although many shallow populations in ex-posed areas are subject to oscillatory flow conditions. Sucha flow regime would have dramatically different effects onthe dispersion and mixing of gametes. Also, we ignored theeffect of a boundary layer that could retain gametes nearbottom and reduce their rate of advection, especially eggsthat are negatively buoyant (Thomas, 1994). An increasedconcentration of gametes in the boundary layer couldgreatly increase fertilization rates. These are important as-pects of flow that warrant further investigation.
Elasticity analysis
Changing the number of spawning males (n) or the spermrelease rate (Qs) has the same effect on the total number ofsperm present in the system (Equations 2 and 3). However,these factors will have different effects on the fertilizationrates observed. Increasing the number of males increases theprobability of having a male very close to the spawningfemale and increases the probability of having eggs fertil-ized rapidly (i.e., before eggs are transported to an area oflower sperm concentration). This effect, combined with theincreased number of sperm present in the system, makes themodel output slightly more sensitive to the number of malesthan to the sperm release rate. The number of males spawn-ing can also be seen as spawning synchrony. Our resultssuggest that spawning synchrony would have a large effecton fertilization success, especially in barrens and kelp bedsubpopulations. Mass spawning of S. droebachiensis hasbeen observed during a multi-species mass spawning (C.P.Dumont, Universite Laval, pers. comm.). Like an increasein the number of spawning males, aggregative behaviorassociated with spawning (Young et al., 1992) could have apositive effect on fertilization success, except for urchins ina grazing front that are continuously at high density. Theelasticity of the model output to changes in current velocitysuggests that a low-density subpopulation (e.g., in a kelpbed) would be particularly influenced by strong currentvelocity. Population size would also be a critical factor.Eggs spawned by a female in a small population would be
203FERTILIZATION IN SEA URCHIN

rapidly transported outside of the population, which woulddecrease their fertilization success. As discussed above, thiseffect would become less pronounced as the size of thepopulation increased.
Our current understanding of the long-term behavior andsurvival of gametes renders any prediction of zygote pro-duction difficult and site-dependent. Even if we assume ahigh survival rate for gametes, uncertainty about populationsize within a given area compromises the accuracy of ourprediction of fertilization success. However, given that theareal extent of urchin populations is typically on the order of1000 m2, we could expect fertilization success on the orderof 90% to be common for S. droebachiensis in the field.This success rate is much higher than that previously pro-posed for this species on the basis of theoretical approaches.This result emphasizes the need to incorporate ecologicallyrelevant parameters other than the “single spawning male,upstream from the female” model often used in the past. Italso suggests that more focus should be put on factors likelyto control survival, development, and settlement of larvae,because these might be more important than fertilizationsuccess in controlling recruitment.
We have demonstrated that by including parameters atthe population level, traditional models of advection-diffu-sion and fertilization kinetics can predict fertilization ratesin accordance with those suggested by recent field observa-tions. These findings underscore the importance of using theappropriate scale in modeling ecological phenomena inorder to obtain relevant theoretical predictions.
Acknowledgments
We thank James Watmough and Myriam Barbeau forhelpful discussions, and two anonymous reviewers for com-ments on a previous version of this manuscript. This re-search was funded by a Discovery Grant from the NaturalSciences and Engineering Research Council of Canada(NSERC) to RES. JSLG was supported by a CanadianGraduate Scholarship from NSERC, a Vaughan GraduateFellowship, and a Magee Postgraduate Merit Award.
Literature Cited
Babcock, R. C., C. N. Mundy, and D. Whitehead. 1994. Sperm diffu-sion models and in situ confirmation of long-distance fertilization in thefree-spawning asteroid Acanthaster planci. Biol. Bull. 186: 17–28.
Barbeau, M. A., and H. Caswell. 1999. A matrix model for short-termdynamics of seeded populations of sea scallops. Ecol. Appl. 9: 266–287.
Bishop, J. D. D. 1998. Fertilization in the sea: are the hazards ofbroadcast spawning avoided when free-spawned sperm fertilize re-tained eggs? Proc. R. Soc. Lond. B 265: 725–731.
Brady, S. M., and R. E. Scheibling. 2005. Repopulation of the shallowsubtidal zone by green sea urchins (Strongylocentrotus droebachiensis)following mass mortality in Nova Scotia, Canada. J. Mar. Biol. Assoc.UK 85: 1511–1517.
Caswell, H. 2001. Matrix Population Models. Sinauer, Sunderland, MA.Claereboudt, C. 1999. Fertilization success in spatially distributed pop-
ulations of benthic free-spawners: a simulation model. Ecol. Model.121: 221–233.
Csanady, G. T. 1973. Turbulent Diffusion in the Environment. D.Reidel, Dordrecht, Netherlands.
Denny, M. W. 1988. Biology and the Mechanics of the Wave-sweptEnvironment. Princeton University Press, Princeton.
Denny, M. W., and M. F. Shibata. 1989. Consequences of surf-zoneturbulence for settlement and external fertilization. Am. Nat. 134:859–889.
Epel, D., M. Kaufman, L. Xiao, H. Kibak, and C. Patton. 1998.Enhancing use of sea urchin eggs and embryos for cell and develop-mental studies: method for storing spawned eggs for extended periods.Mol. Biol. Cell 9: 182a.
Gaudette, J. 2004. Disponibilite du sperme au cours de la saison de laponte de l’oursin vert, Strongylocentrotus droebachiensis, dans despopulations de differentes tailles. M.Sc. dissertation, Universite Laval,Quebec City, Canada.
Lauzon-Guay, J.-S., and R. E. Scheibling. 2007. Behaviour of seaurchin (Strongylocentrotus droebachiensis) grazing fronts: food-medi-ated aggregation and density-dependent facilitation. Mar. Ecol. Prog.Ser. 329: 191–204.
Levitan, D. R. 1991. Influence of body size and population density onfertilization success and reproductive output in a free-spawning inver-tebrate. Biol. Bull. 181: 261–268.
Levitan, D. R. 1993. The importance of sperm limitation to the evolutionof egg size in marine invertebrates. Am. Nat. 141: 517–536.
Levitan, D. R., and C. M. Young. 1995. Reproductive success in largepopulations: empirical measures and theoretical predictions of fertili-zation in the sea biscuit Clypeaster rosaceus. J. Exp. Mar. Biol. Ecol.190: 221–241.
Levitan, D. R., M. A. Sewell, and F. S. Chia. 1992. How distributionand abundance influence fertilization success in the sea urchin Strongy-locentrotus franciscanus. Ecology 73: 248–254.
Meidel, S. K. 1999. Reproductive ecology of the sea urchin Strongylo-centrotus droebachiensis. Ph.D. dissertation, Dalhousie University,Halifax, Canada.
Meidel, S. K., and R. E. Scheibling. 2001. Variation in egg spawningamong subpopulations of sea urchins Strongylocentrotus droebachien-sis: a theoretical approach. Mar. Ecol. Prog. Ser. 213: 97–110.
Meidel, S. K., and P. O. Yund. 2001. Egg longevity and time-integratedfertilization in a temperate sea urchin (Strongylocentrotus droebachien-sis). Biol. Bull. 201: 84–94.
Metaxas, A., R. E. Scheibling, and C. M. Young. 2002. Estimatingfertilization success in marine benthic invertebrates: a case study withthe tropical sea star Oreaster reticulatus. Mar. Ecol. Prog. Ser. 226:87–101.
Millar, R. B., and M. J. Anderson. 2003. The kinetics of monospermicand polyspermic fertilization in free-spawning marine invertebrates. J.Theor. Biol. 224: 79–85.
Moore, D. S., and R. J. Miller. 1983. Recovery of macroalgae followingwidespread sea urchin mortality with a description of the nearshorehard-bottom habitat on the Atlantic coast of Nova Scotia. Can. Tech.Rep. Fish. Aquat. Sci. 1230: vii � 94 p.
Pennington, J. T. 1985. The ecology and fertilization of echinoid eggs:the consequence of sperm dilution, adult aggregation, and synchronousspawning. Biol. Bull. 169: 417–430.
Styan, C. A. 1998. Polyspermy, egg size, and the fertilization kinetics offree-spawning marine invertebrates. Am. Nat. 152: 290–297.
Thomas, F. I. M. 1994. Physical properties of gametes in three seaurchin species. J. Exp. Biol. 194: 263–284.
Thompson, R. J. 1979. Fecundity and reproductive effort in the bluemussel (Mytilus edulis), the sea urchin (Strongylocentrotus droe-
204 J-S. LAUZON-GUAY AND R. E. SCHEIBLING

bachiensis), and the snow crab (Chionoecetes opilio) from populationsin Nova Scotia and Newfoundland. J. Fish. Res. Board Can. 36:955–964.
Vogel, H., G. Czihak, P. Chang, and W. Wolf. 1982. Fertilizationkinetics of sea urchin eggs. Math. Biosci. 58: 189–216.
Wahle, R. A., and A. E. Gilbert. 2002. Detecting and quantifying malesea urchin spawning with time-integrated fertilization assays. Mar.Biol. 140: 375–382.
Wahle, R. A., and S. H. Peckham. 1999. Density-related reproductivetrade-offs in the green sea urchin, Strongylocentrotus droebachiensis.Mar. Biol. 134: 127–137.
Wharton, W. G., and K. H. Mann. 1981. Relationship betweendestructive grazing by the sea urchin, Strongylocentrotus droebachien-sis, and the abundance of American lobster, Homarus americanus, onthe Atlantic coast of Nova Scotia. Can. J. Fish. Aquat. Sci. 38:1339–1349.
Young, C. M., P. A. Tyler, J. L. Cameron, and S. G. Rumrill. 1992.Seasonal breeding aggregations in low-density populations of thebathyal echinoid Stylocidaris lineata. Mar. Biol. 113: 603–612.
Yund, P. O., and S. K. Meidel. 2003. Sea urchin spawning in benthicboundary layers: Are eggs fertilized before advecting away from fe-males? Limnol. Oceanogr. 48: 795–801.
205FERTILIZATION IN SEA URCHIN
Copyright © 2022 FDOKUMEN




















