
The effects of physiologically important nonmetallic ligands in the reactivity of metallothionein...
6
The effects of physiologically important nonmetallic ligands in the reactivity of metallothionein towards 5,5 0 -dithiobis(2-nitrobenzoic acid) A new method for the determination of ligand interactions with metallothionein Liina Kangur 1,2 and Peep Palumaa 2 1 Institute of Molecular and Cell Biology, University of Tartu, Estonia; 2 Institute of Basic and Applied Chemistry, Tallinn Technical University, Estonia The reaction of Cd 5 Zn 2 -metallothionein (MT) with 5,5 0 - dithiobis(2-nitrobenzoic acid) (Nbs 2 ) has been studied at different reagent stoichiometries, pH and temperature con- ditions and in the presence of several ligands. At stoichio- metries of Nbs 2 to MT from 0.5 to 5, the reaction followed first order kinetics. The first order rate constants obtained were independent from the concentration of Nbs 2 but were linearly dependent on the concentration of MT. At higher Nbs 2 /MT stoichiometries, the reaction deviates from first order kinetics and the observed rate constant increases. The reactivity of MT towards Nbs 2 has been probed at 4 mM concentration of both reagents where the reaction is mono- phasic and is characterized by a linear Arrhenius plot (E a ¼ 45.8 ^ 2.7 kJ : mol 21 ). It has been demonstrated that metal release at low pH or subtraction from MT by EDTA substantially increases the reactivity of MT towards Nbs 2 . At the same time, a number of nonmetallic ligands moder- ately accelerate the reaction of MT with Nbs 2 and hyperbolic dose –response curves were obtained. The data have been interpreted with the binding of ligands to MT and following MT. Ligand binding constants were calculated as follows: ATP, K ¼ 0.31 ^ 0.06 mM; ADP, K ¼ 0.26 ^ 0.07 mM. Several compounds such as AMP, S-methylglu- tathione, and phosphate had no effect on the reaction, but Zn 21 ions showed an inhibitory effect at micromolar concentrations. Keywords: metallothionein; Ellman’s reagent; 5,5 0 -dithio- bis(2-nitrobenzoic acid); ATP; reactivity of metal thiolate clusters. Metallothioneins (MTs) represent a class of small cysteine- rich proteins adapted for the binding of various ‘soft’ transition metal ions into metal (thiolate clusters) [1]. Mammalian MTs contain 60–68 amino acids with an absolutely conserved pattern of 20 cysteines [2]. The three- dimensional structure of mammalian MT has been deter- mined for MT metalloforms with seven divalent metal ions (Zn 21 and/or Cd 21 ) by NMR [3 – 5] and by X-ray diffraction [6]. The NMR and the crystal structures are similar and they expose two protein domains, both folded into metal-thiolate clusters [7]. The N-terminal b domain (amino acids 1–31) contains a cluster of three metals co-ordinated with nine thiolates, and the C-terminal a domain contains a cluster of four metals co-ordinated with 11 thiolates [7,8]. MT is a ubiquitous protein and in cells it is induced by a variety of stimuli, including metals, hormones, chemical and physical stress factors [1,9]. However, the primary physiological role of MT is unclear and is still a matter for discussion [10,11]. At the same time, it is generally accepted that MTs are multifunctional proteins participating in the metabolism of zinc and copper, in detoxification of toxic metals such as cadmium and mercury, and in the protection of cells against reactive oxygen species and other electro- philes [9,12]. Some tissue-specific MT isoforms may also have additional physiological functions, such as inhibition of neuronal growth in the case of brain-specific MT-3 [13,14]. Metal transfer and release play an important role in functioning of MT and the appropriate mechanisms have been intensively studied. Generally, MT-bound metals are kinetically labile [15] and could be transferred to other proteins or released into the environment. It has been demonstrated that MT can participate in metal transfer and/or exchange with different metalloproteins including enzymes [16 – 18], receptors [19] and transcription factors [20]. The mechanisms of metal release from MT in vivo is not completely understood, but according to some experimental evidence, metals may be released from MT clusters through partial oxidation of MT thiolates [21 – 24]. It has been demonstrated that the neutrophil oxidants can mobilize Zn 21 from MT, which may have injurious or beneficial effects at sites of inflammation [22]. It has also been demonstrated that oxidative metal release from MT by intracellular disulfides (oxidized glutathione) [23] or nitric oxide [25], could operate as important metalloregulatory processes in vivo. In addition, it has been established that oxidative metal release from MT could be regulated by important physiological ligands such as ATP and GTP, Correspondence to P. Palumaa, Institute of Basic and Applied Chemistry, Tallinn Technical University, Ehitajate tee 5, 19086 Tallinn, Estonia, Fax: 1 372 6202020 Tel.: 1 372 6202804, E-mail: [email protected] (Received 15 May 2001, revised 30 July 2001, accepted 31 July 2001) Abbreviations: MT, metallothionein; Nbs 2 , 5,5 0 -dithiobis(2-nitrobenzoic acid); Nbs – , 5-thio(2-nitrobenzoic acid); GSH, glutathione; S-MeGSH, S-methylglutathione Eur. J. Biochem. 268, 4979–4984 (2001) q FEBS 2001
Transcript of The effects of physiologically important nonmetallic ligands in the reactivity of metallothionein...

The effects of physiologically important nonmetallic ligands in thereactivity of metallothionein towards 5,50-dithiobis(2-nitrobenzoicacid)A new method for the determination of ligand interactions with metallothionein
Liina Kangur1,2 and Peep Palumaa2
1Institute of Molecular and Cell Biology, University of Tartu, Estonia; 2Institute of Basic and Applied Chemistry, Tallinn Technical
University, Estonia
The reaction of Cd5Zn2-metallothionein (MT) with 5,50-
dithiobis(2-nitrobenzoic acid) (Nbs2) has been studied at
different reagent stoichiometries, pH and temperature con-
ditions and in the presence of several ligands. At stoichio-
metries of Nbs2 to MT from 0.5 to 5, the reaction followed
first order kinetics. The first order rate constants obtained
were independent from the concentration of Nbs2 but were
linearly dependent on the concentration of MT. At higher
Nbs2/MT stoichiometries, the reaction deviates from first
order kinetics and the observed rate constant increases. The
reactivity of MT towards Nbs2 has been probed at 4 mM
concentration of both reagents where the reaction is mono-
phasic and is characterized by a linear Arrhenius plot
(Ea ¼ 45.8 ^ 2.7 kJ:mol21). It has been demonstrated that
metal release at low pH or subtraction from MT by EDTA
substantially increases the reactivity of MT towards Nbs2.
At the same time, a number of nonmetallic ligands moder-
ately accelerate the reaction of MT with Nbs2 and
hyperbolic dose–response curves were obtained. The data
have been interpreted with the binding of ligands to MT and
following MT. Ligand binding constants were calculated as
follows: ATP, K ¼ 0.31 ^ 0.06 mM; ADP, K ¼ 0.26 ^
0.07 mM. Several compounds such as AMP, S-methylglu-
tathione, and phosphate had no effect on the reaction, but
Zn21 ions showed an inhibitory effect at micromolar
concentrations.
Keywords: metallothionein; Ellman’s reagent; 5,50-dithio-
bis(2-nitrobenzoic acid); ATP; reactivity of metal thiolate
clusters.
Metallothioneins (MTs) represent a class of small cysteine-rich proteins adapted for the binding of various ‘soft’transition metal ions into metal (thiolate clusters) [1].Mammalian MTs contain 60–68 amino acids with anabsolutely conserved pattern of 20 cysteines [2]. The three-dimensional structure of mammalian MT has been deter-mined for MT metalloforms with seven divalent metal ions(Zn21 and/or Cd21) by NMR [3–5] and by X-ray diffraction[6]. The NMR and the crystal structures are similar and theyexpose two protein domains, both folded into metal-thiolateclusters [7]. The N-terminal b domain (amino acids 1–31)contains a cluster of three metals co-ordinated with ninethiolates, and the C-terminal a domain contains a cluster offour metals co-ordinated with 11 thiolates [7,8].
MT is a ubiquitous protein and in cells it is induced by avariety of stimuli, including metals, hormones, chemicaland physical stress factors [1,9]. However, the primaryphysiological role of MT is unclear and is still a matter fordiscussion [10,11]. At the same time, it is generally accepted
that MTs are multifunctional proteins participating in themetabolism of zinc and copper, in detoxification of toxicmetals such as cadmium and mercury, and in the protectionof cells against reactive oxygen species and other electro-philes [9,12]. Some tissue-specific MT isoforms may alsohave additional physiological functions, such as inhibitionof neuronal growth in the case of brain-specific MT-3[13,14].
Metal transfer and release play an important role infunctioning of MT and the appropriate mechanisms havebeen intensively studied. Generally, MT-bound metals arekinetically labile [15] and could be transferred to otherproteins or released into the environment. It has beendemonstrated that MT can participate in metal transferand/or exchange with different metalloproteins includingenzymes [16–18], receptors [19] and transcription factors[20]. The mechanisms of metal release from MT in vivois not completely understood, but according to someexperimental evidence, metals may be released from MTclusters through partial oxidation of MT thiolates [21–24].It has been demonstrated that the neutrophil oxidants canmobilize Zn21 from MT, which may have injurious orbeneficial effects at sites of inflammation [22]. It has alsobeen demonstrated that oxidative metal release from MT byintracellular disulfides (oxidized glutathione) [23] or nitricoxide [25], could operate as important metalloregulatoryprocesses in vivo. In addition, it has been established thatoxidative metal release from MT could be regulated byimportant physiological ligands such as ATP and GTP,
Correspondence to P. Palumaa, Institute of Basic and Applied
Chemistry, Tallinn Technical University, Ehitajate tee 5, 19086 Tallinn,
Estonia, Fax: 1 372 6202020 Tel.: 1 372 6202804,
E-mail: [email protected]
(Received 15 May 2001, revised 30 July 2001, accepted 31 July 2001)
Abbreviations: MT, metallothionein; Nbs2,
5,50-dithiobis(2-nitrobenzoic acid); Nbs–, 5-thio(2-nitrobenzoic acid);
GSH, glutathione; S-MeGSH, S-methylglutathione
Eur. J. Biochem. 268, 4979–4984 (2001) q FEBS 2001
through their binding to MT [26]. The binding of ATP toMT has been re-evaluated by another lab and in contrastto previous studies, no evidence of the interaction of MTwith ATP was detected [27]. One possible way to shedlight into these conflicting results of the interaction of MTwith ATP, and with other physiologically importantnonmetallic ligands, is by the elaboration and use ofnew methods for the detection of ligand interactions withMT.
In the present study, ligand binding to MT was exposed bymeans of the ligand-induced changes in the reactivity of MTthiolates towards 5,50-dithiobis(2-nitrobenzoic acid) (Nbs2).The application of the method for the determination ofbinding constants for a series of nonthiolate ligandsincluding ATP are demonstrated and possible uses of thenew method for MT studies are discussed.
M A T E R I A L S A N D M E T H O D S
Rabbit liver MT was isolated from rabbits exposed tocadmium salt according to a general procedure [28]. MT-1and MT-2 isoforms were separated by ion-exchangechromatography in a DEAE-cellulose matrix (Whatman),and the isolated Cd5Zn2-MT-2 form was used in allexperiments. The protein concentration was determinedfrom the absorption of the metal-free protein at 220 nm in0.1 M HCl (1220 ¼ 47 300 M
21:cm21) [29]. The metalcontent was determined by atomic absorption spectroscopyon a PerkinElmer 372 instrument.
Kinetic studies were performed in 10-mm thermostatedspectrophotometer cuvettes, and the reaction was started bythe addition of MT to the buffer solution containing Nbs2
and other substances. The reaction was followed at 412 nmon a UV-1601PC instrument (Shimadzu). Kinetic data wereanalysed using the GRAPHPAD PRISM program.
Analytical gel filtration was performed on a Superdex 75column (1 � 30 cm) (Pharmacia), attached to the chroma-tography system combined with a FPLC injection valve,P-500 pump (Pharmacia) and UV detector Uvicord S (LKB,Sweden). The column was eluted with a 20-mM Tris/HClbuffer (pH 8.0) containing 0.15 M KCl.
Reagents were obtained from the following sources: Tris
base, ADP (free acid), EDTA:4Na, S-methylglutathione(S-MeGSH) were from Sigma; ATP:2Na was from SERVA;AMP:Na was from Boehringer Mannheim; 5,50-dithiobis(2-nitrobenzoic acid) and KCl were from Fluka.
R E S U L T S
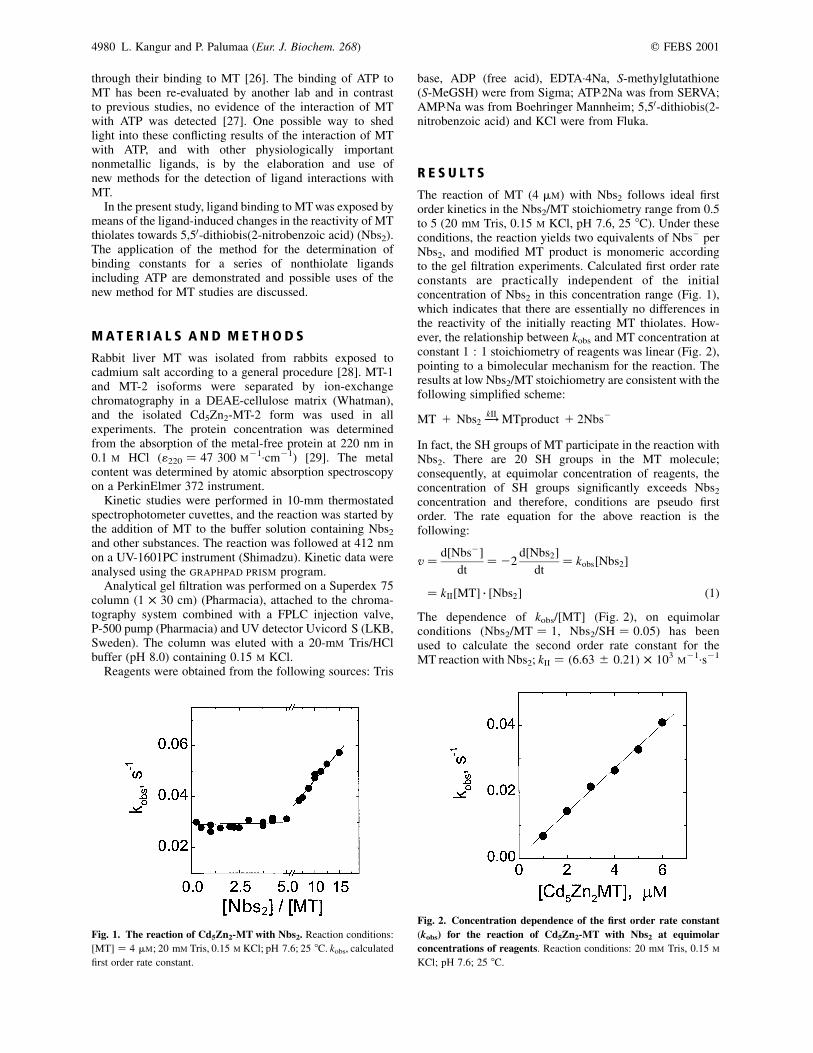
The reaction of MT (4 mM) with Nbs2 follows ideal firstorder kinetics in the Nbs2/MT stoichiometry range from 0.5to 5 (20 mM Tris, 0.15 M KCl, pH 7.6, 25 8C). Under theseconditions, the reaction yields two equivalents of Nbs– perNbs2, and modified MT product is monomeric accordingto the gel filtration experiments. Calculated first order rateconstants are practically independent of the initialconcentration of Nbs2 in this concentration range (Fig. 1),which indicates that there are essentially no differences inthe reactivity of the initially reacting MT thiolates. How-ever, the relationship between kobs and MT concentration atconstant 1 : 1 stoichiometry of reagents was linear (Fig. 2),pointing to a bimolecular mechanism for the reaction. Theresults at low Nbs2/MT stoichiometry are consistent with thefollowing simplified scheme:
MT 1 Nbs2kIIÿ!MTproduct 1 2Nbs2
In fact, the SH groups of MT participate in the reaction withNbs2. There are 20 SH groups in the MT molecule;consequently, at equimolar concentration of reagents, theconcentration of SH groups significantly exceeds Nbs2
concentration and therefore, conditions are pseudo firstorder. The rate equation for the above reaction is thefollowing:
v ¼d½Nbs2�
dt¼ 22
d½Nbs2�
dt¼ kobs½Nbs2�
¼ kII½MT��½Nbs2� ð1Þ
The dependence of kobs/[MT] (Fig. 2), on equimolarconditions (Nbs2/MT ¼ 1, Nbs2/SH ¼ 0.05) has beenused to calculate the second order rate constant for theMT reaction with Nbs2; kII ¼ (6.63 ^ 0.21) � 103
M21:s21
Fig. 1. The reaction of Cd5Zn2-MT with Nbs2. Reaction conditions:
[MT] ¼ 4 mM; 20 mM Tris, 0.15 M KCl; pH 7.6; 25 8C. kobs, calculated
first order rate constant.
Fig. 2. Concentration dependence of the first order rate constant
(kobs) for the reaction of Cd5Zn2-MT with Nbs2 at equimolar
concentrations of reagents. Reaction conditions: 20 mM Tris, 0.15 M
KCl; pH 7.6; 25 8C.
4980 L. Kangur and P. Palumaa (Eur. J. Biochem. 268) q FEBS 2001
(20 mM Tris, 0.15 M KCl, pH 7.6, 25 8C). The bimolecularrate constant calculated this way characterizes the reactionof natively metallated MT with Nbs2; it has a differentabsolute value and meaning from the bimolecular rateconstants presented in the literature. Bimolecular rateconstants in the literature are commonly determined in alarge excess of Nbs2 over MT [30–35]. At high Nbs2/MTstoichiometry the reaction is not monophasic and the kineticdata are usually analyzed by using the following four-termrate equation:
rate ¼ k1S½MT�1 ðk2S½Nbs2�½MT�Þ1 ðk1f½MT�Þ
1 ðk2f½Nbs2�½MT�Þ ð2Þ
where indexes s and f refer to the slow and fast steps of thereaction, and k1 and k2 are the first and second order rateconstants, respectively [31,32,35,36]. Under these con-ditions, the dependence of kobs on [Nbs2] at a constantconcentration of MT was used to calculate the bimolecularrate constants [31,32,35,36]. Our data confirm that at higherNbs2/MT stoichiometries ([Nbs2]/[MT] . 5) the reactiondeviates from first order kinetics and the calculated apparentfirst order rate constant increases (Fig. 1). However, we didnot analyze the kinetic data at higher Nbs2/MT stoichio-metries as it was not in line with the aims of present study.
Based on the obtained kinetic data, the following standardconditions were selected for probing the MT reactivitytowards Nbs2: an [Nbs2]/[MT] stoichiometry of 1 : 1;concentration of reagents were 4 mM. All followingexperiments were conducted using these standard conditions.
pH dependence
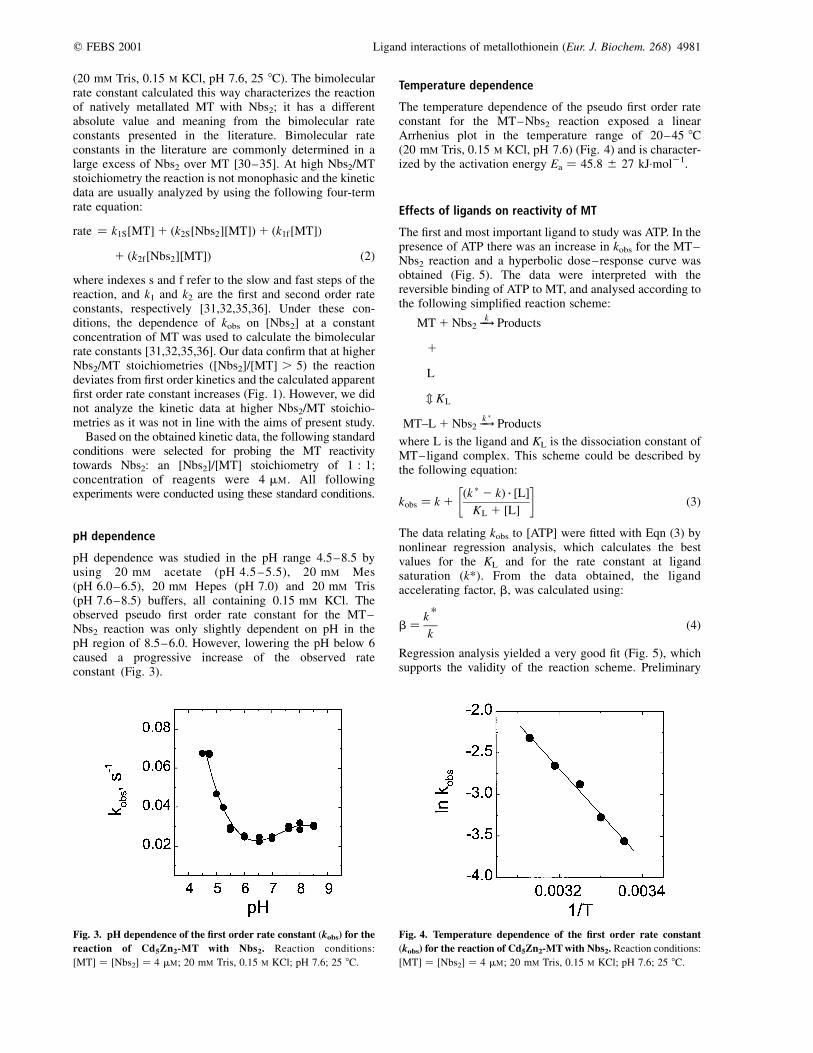
pH dependence was studied in the pH range 4.5–8.5 byusing 20 mM acetate (pH 4.5 – 5.5), 20 mM Mes(pH 6.0–6.5), 20 mM Hepes (pH 7.0) and 20 mM Tris(pH 7.6–8.5) buffers, all containing 0.15 mM KCl. Theobserved pseudo first order rate constant for the MT–Nbs2 reaction was only slightly dependent on pH in thepH region of 8.5–6.0. However, lowering the pH below 6caused a progressive increase of the observed rateconstant (Fig. 3).
Temperature dependence
The temperature dependence of the pseudo first order rateconstant for the MT–Nbs2 reaction exposed a linearArrhenius plot in the temperature range of 20–45 8C(20 mM Tris, 0.15 M KCl, pH 7.6) (Fig. 4) and is character-ized by the activation energy Ea ¼ 45.8 ^ 27 kJ:mol21.
Effects of ligands on reactivity of MT
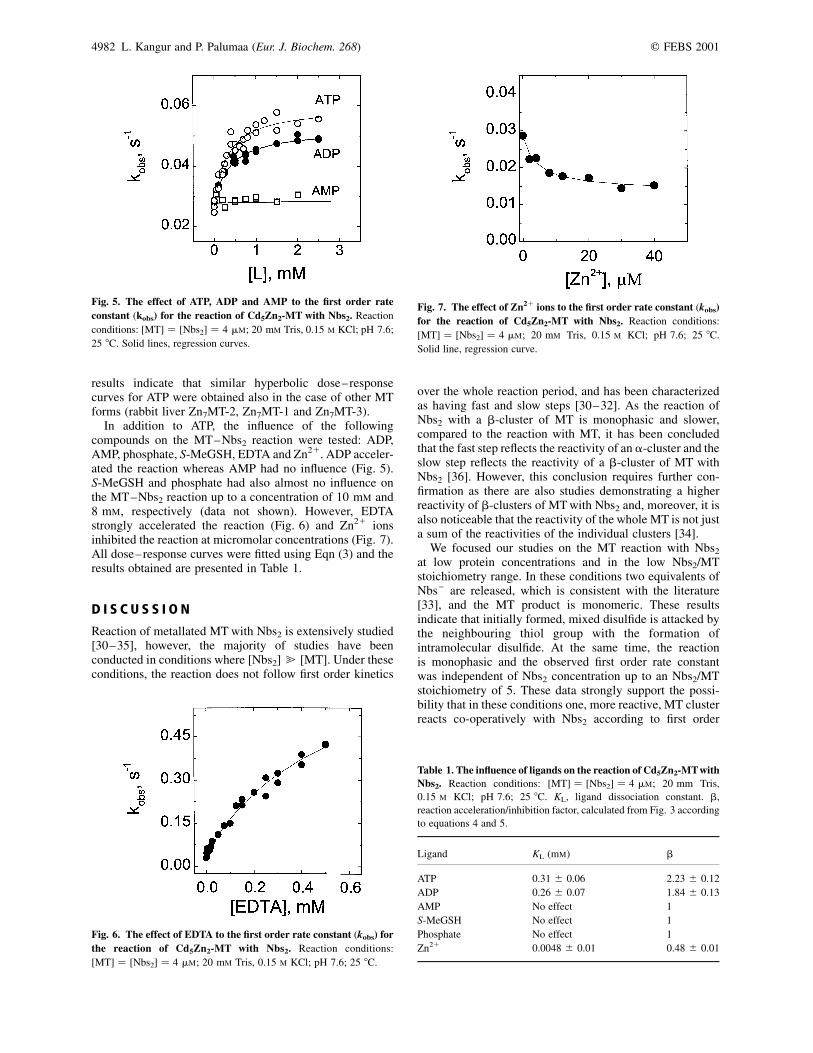
The first and most important ligand to study was ATP. In thepresence of ATP there was an increase in kobs for the MT–Nbs2 reaction and a hyperbolic dose–response curve wasobtained (Fig. 5). The data were interpreted with thereversible binding of ATP to MT, and analysed according tothe following simplified reaction scheme:
MT 1 Nbs2kÿ! Products
1
L
m KL
MT–L 1 Nbs2k *
ÿ! Products
where L is the ligand and KL is the dissociation constant ofMT–ligand complex. This scheme could be described bythe following equation:
kobs ¼ k 1ðk * 2 kÞ�½L� �
KL 1 ½L� ��
� �ð3Þ
The data relating kobs to [ATP] were fitted with Eqn (3) bynonlinear regression analysis, which calculates the bestvalues for the KL and for the rate constant at ligandsaturation (k*). From the data obtained, the ligandaccelerating factor, b, was calculated using:
b ¼k *
kð4Þ
Regression analysis yielded a very good fit (Fig. 5), whichsupports the validity of the reaction scheme. Preliminary
Fig. 3. pH dependence of the first order rate constant (kobs) for the
reaction of Cd5Zn2-MT with Nbs2. Reaction conditions:
[MT] ¼ [Nbs2] ¼ 4 mM; 20 mM Tris, 0.15 M KCl; pH 7.6; 25 8C.
Fig. 4. Temperature dependence of the first order rate constant
(kobs) for the reaction of Cd5Zn2-MT with Nbs2. Reaction conditions:
[MT] ¼ [Nbs2] ¼ 4 mM; 20 mM Tris, 0.15 M KCl; pH 7.6; 25 8C.
q FEBS 2001 Ligand interactions of metallothionein (Eur. J. Biochem. 268) 4981
results indicate that similar hyperbolic dose–responsecurves for ATP were obtained also in the case of other MTforms (rabbit liver Zn7MT-2, Zn7MT-1 and Zn7MT-3).
In addition to ATP, the influence of the followingcompounds on the MT–Nbs2 reaction were tested: ADP,AMP, phosphate, S-MeGSH, EDTA and Zn21. ADP acceler-ated the reaction whereas AMP had no influence (Fig. 5).S-MeGSH and phosphate had also almost no influence onthe MT–Nbs2 reaction up to a concentration of 10 mM and8 mM, respectively (data not shown). However, EDTAstrongly accelerated the reaction (Fig. 6) and Zn21 ionsinhibited the reaction at micromolar concentrations (Fig. 7).All dose–response curves were fitted using Eqn (3) and theresults obtained are presented in Table 1.
D I S C U S S I O N
Reaction of metallated MT with Nbs2 is extensively studied[30–35], however, the majority of studies have beenconducted in conditions where [Nbs2] @ [MT]. Under theseconditions, the reaction does not follow first order kinetics
over the whole reaction period, and has been characterizedas having fast and slow steps [30–32]. As the reaction ofNbs2 with a b-cluster of MT is monophasic and slower,compared to the reaction with MT, it has been concludedthat the fast step reflects the reactivity of an a-cluster and theslow step reflects the reactivity of a b-cluster of MT withNbs2 [36]. However, this conclusion requires further con-firmation as there are also studies demonstrating a higherreactivity of b-clusters of MT with Nbs2 and, moreover, it isalso noticeable that the reactivity of the whole MT is not justa sum of the reactivities of the individual clusters [34].
We focused our studies on the MT reaction with Nbs2
at low protein concentrations and in the low Nbs2/MTstoichiometry range. In these conditions two equivalents ofNbs– are released, which is consistent with the literature[33], and the MT product is monomeric. These resultsindicate that initially formed, mixed disulfide is attacked bythe neighbouring thiol group with the formation ofintramolecular disulfide. At the same time, the reactionis monophasic and the observed first order rate constantwas independent of Nbs2 concentration up to an Nbs2/MTstoichiometry of 5. These data strongly support the possi-bility that in these conditions one, more reactive, MT clusterreacts co-operatively with Nbs2 according to first order
Fig. 5. The effect of ATP, ADP and AMP to the first order rate
constant (kobs) for the reaction of Cd5Zn2-MT with Nbs2. Reaction
conditions: [MT] ¼ [Nbs2] ¼ 4 mM; 20 mM Tris, 0.15 M KCl; pH 7.6;
25 8C. Solid lines, regression curves.
Fig. 6. The effect of EDTA to the first order rate constant (kobs) for
the reaction of Cd5Zn2-MT with Nbs2. Reaction conditions:
[MT] ¼ [Nbs2] ¼ 4 mM; 20 mM Tris, 0.15 M KCl; pH 7.6; 25 8C.
Fig. 7. The effect of Zn21 ions to the first order rate constant (kobs)
for the reaction of Cd5Zn2-MT with Nbs2. Reaction conditions:
[MT] ¼ [Nbs2] ¼ 4 mM; 20 mM Tris, 0.15 M KCl; pH 7.6; 25 8C.
Solid line, regression curve.
Table 1. The influence of ligands on the reaction of Cd5Zn2-MTwith
Nbs2. Reaction conditions: [MT] ¼ [Nbs2] ¼ 4 mM; 20 mm Tris,
0.15 M KCl; pH 7.6; 25 8C. KL, ligand dissociation constant. b,
reaction acceleration/inhibition factor, calculated from Fig. 3 according
to equations 4 and 5.
Ligand KL (mM) b
ATP 0.31 ^ 0.06 2.23 ^ 0.12
ADP 0.26 ^ 0.07 1.84 ^ 0.13
AMP No effect 1
S-MeGSH No effect 1
Phosphate No effect 1
Zn21 0.0048 ^ 0.01 0.48 ^ 0.01
4982 L. Kangur and P. Palumaa (Eur. J. Biochem. 268) q FEBS 2001

kinetics. From our kinetic data it could not be concludedwhich cluster of MT participates in the reaction. However, at1 : 1 stoichiometry, only native MT reacts with Nbs2, whichwas the main reason why these conditions have beenselected to probe MT reactivity towards Nbs2.
The results of pH dependence provide the followinginformation about the MT–Nbs2 reaction. First, it suggeststhat in a neutral pH range, Nbs2 does not react with free SHgroups but probably with metal co-ordinated thiolates,which have substantially lower pKa values compared to freeSH groups. Consequently, the dissociation of metals is notnecessary for the reaction to occur at neutral pH. Secondly, asubstantial increase in the reaction rate at lower pH values isprobably caused by metal release in acidic conditions, andshows that even under acidic conditions, free SH groupshave a much higher reactivity towards Nbs2, comparedwith metal-bound thiolates. At higher pH values (pH ¼ 7.6)EDTA, which is known as a potent metal extracting ligand[37], caused a substantial increase in the observed rateconstant and there was only a slight tendency for saturationobserved at 0.5 mM concentration of EDTA (Fig. 6).Together, these results indicate that partly metal-depletedMT exposes sufficiently higher rate constants towards Nbs2,compared with completely metallated MT. At the sametime, free Zn21 ions inhibit the reaction (Fig. 7) indicatingthat Zn21 competes with Nbs2 for interaction with solventexposed thiolates, which probably serve as additional low-affinity binding sites for Zn21 ions in the structure ofCd5Zn2-MT. Additional binding sites are known for Cd21
ions in the structure of Cd7MT [38–40], and it is reasonablethat similar possibilities, but with different chemicalaffinities, should also be present for Zn21 ions. In summary,the obtained results indicate that at neutral pH, Nbs2
probably reacts with solution-exposed metal co-ordinatedMT thiolates. It is realistic that the Nbs2 attack of these MTthiolates is sensitive to steric factors, and it is expected thatslight changes in the structure of clusters induced by ligandbinding might change the reactivity of MT towards Nbs2.
Our results confirm the influence of ATP on the reactivityof MT towards Nbs2 [26], and demonstrate further that ATP,as well as ADP, can moderately change the reactivity of MTtowards Nbs2 and hyperbolic dose–response curves areobserved. Kinetic data fitted well to a simple bindingisotherm allowing the determination of ligand dissociationconstants and acceleration factors. Acceleration factors, inthe case of ligands studied, range from 1.8 to 2.2, which aremuch lower than in the case of metal release. These resultsare consistent with the binding of ATP and ADP to MT andare inconsistent with the possibility that these ligandsaccelerate the reaction of MTwith Nbs2 through the extractionof metals from MT. It is not clear why this equilibrium, if itexists, could not be detected by various physico-chemicalmethods applied in recent studies [27]. However, ligandbinding studies of MT as peculiar small SH- and metal-richproteins are prone to experimental difficulties.
The ligands studied so far are adenosine phosphates,inorganic phosphate and glutathione derivative. Our dataconfirm that ATP in submillimolar concentrations can bindto Cd5Zn2-MT. The binding constant that was determined inthis study (KL ¼ 0.31 ^ 0.06 mM) is in the same range asthe binding constant determined by equilibrium gel filtration(0.18 ^ 0.03 mM) [26], and the slight difference could arisefrom differences in the MT sample and methodology used.
According to our data, ADP had a similar influence onreactivity of MT, but AMP and inorganic phosphate had noeffect to the reactivity of MT. Inorganic phosphate has beenselected as a ligand for two reasons. First, it has beendemonstrated that inorganic phosphate is co-crystallizedwith Cd5Zn2-MT and in crystals it interacts with MT [6].Secondly, it has been demonstrated that inorganic phosphatebinds to dimeric and globular forms of mammalian Cd-MT[38,39]. Absence of an effect in the case of AMP andiorganic phosphate can not be interpreted only by the lack ofbinding, as there exists the possibility that these ligands bindto MT, but have no influence on the reactivity of MTclusters.
Glutathione (GSH) is so far the only low molecular massorganic ligand whose binding to the mammalian MT hasbeen definitely demonstrated [41]. Moreover, based onmolecular graphics analysis it has been suggested that GSHis bound into the cleft on the N-terminal three-metal clusterof MT and the thiol group of GSH could be bound to Zn21
ion [41]. The method described in the present study isrestricted to studies of nonthiolate ligands, and therefore it isnot applicable for GSH. Instead of GSH, we studied theeffect of S-methylGSH on the reactivity of MT; onlyminimal accelerating effects, if any, were observed. Theabsence of an effect may indicate that S-MeGSH, asopposed to GSH, does not interact with Cd5Zn2-MT at theconcentrations studied. However, it is also possible thatunder the conditions studied, Nbs2 and S-MeGSH interactwith different MT clusters and do not interfere with eachother.
The presented data demonstrate the application of Nbs2 inprobing the reactivity of metal thiolate clusters of MT andtheir sensitivity towards different environmental stimuli,including metallic and organic ligands. As the reactivity ofMT reflects the capacity of MT to release and transfermetals, a new approach could be useful for studies of MTaction. The method has been successfully applied in order todetermine the binding constants of important physiologicalligands such as ATP and ADP. The physiological importanceof ATP binding to MT is not clear from present in vitrostudies. However, the results agree with an earlier conclu-sion that under in vivo conditions, MT is essentially com-plexed with ATP [26]. However, it is also evident that theenergetic status of the cell (ATP/ADP ratio) can not regulatethe metal transfer potential of MT as ADP and ATP hadquite similar effects to the reactivity of MT. Therefore, it cannot be excluded that binding of ATP reflects anotherunknown function of MT. Elaborated technology opens thepossibility for screening the broader set of physiologicalligands to the reactivity of MT, which may be of crucialimportance for functional and mechanistic studies of MT.
A C K N O W L E D G E M E N T S
This work was supported by the Estonian Science Foundation Grant no.
4287. The authors would like to thank Mrs Hilja Lopp and Mrs Maie
Loorits for their kind help in the preparation of metallothionein.
R E F E R E N C E S
1. Kagi, J.H.R. & Schaffer, A. (1988) Biochemistry of metallothio-
nein. Biochemistry 27, 8509–8515.
2. Binz, B.A. & Kagi, J.H.R. (1999) Metallothionein: molecular
q FEBS 2001 Ligand interactions of metallothionein (Eur. J. Biochem. 268) 4983

evolution and classification. In Metallothionein IV (Klaassen, J.,
ed.), pp. 7–13. Birkhauser, Basel, Switzerland.
3. Frey, M.H., Wagner, G., Vasak, M., Sorensen, O.W., Neuhaus, D.,
Worgotter, E., Kagi, J.H.R., Ernst, R.E. & Wuthrich, K. (1985)
Polypeptide metal cluster connectivities in metallothionein-2 by
novel 1H-113Cd heteronuclear two-dimensional NMR experiments.
J. Am. Chem. Soc. 107, 6847–6851.
4. Arseniev, A., Schultze, P., Worgotter, E., Braun, W., Wagner, G.,
Vasak, M.J.H.R.K. & Wuthrich, K. (1988) 3-Dimensional structure
of rabbit liver [Cd7]metallothionein-2a in aqueous-solution
determined by nuclear magnetic-resonance. J. Mol. Biol. 201,
637–657.
5. Messerle, B., Schaffer, A., Vasak, M., Kagi, J.H.R. & Wuthrich, K.
(1990) 3-Dimensional structure of human [113Cd7]-
metallothionein-2 in solution determined by nuclear-magnetic-
resonance spectroscopy. J. Mol. Biol. 214, 765–779.
6. Robbins, A.H., McRee, D.E., Williamson, M., Collett, S.A.,
Xuong, N.H., Furey, W.F., Wang, B.C. & Stout, C.D. (1991)
Refined crystal-structure of Cd, Zn metallothionein at 2.0 A
resolution. J. Mol. Biol. 221, 1269–1993.
7. Braun, W., Vasak, M., Robbins, A.H., Stout, C.D., Wagner, G.,
Kagi, J.H.R. & Wuthrich, K. (1992) Comparison of the NMR
solution structure and the X-ray crystal-structure of rat metal-
lothionein-2. Proc. Natl Acad. Sci. USA 89, 10124–10128.
8. Vasak, M. & Kagi, J.H.R. (1994) Metallothioneins. In Encyclo-
pedia of Inorganic Chemistry (King, R.B., ed.), pp. 2229–2241.
J Wiley & Sons Ltd, New York, USA.
9. Kagi, J.H.R. (1991) Overview of metallothionein. Methods
Enzymol. 205, 613–626.
10. Palmiter, R.D. (1998) The elusive function of metallothioneins.
Proc. Natl Acad. Sci. USA 95, 8428–8430.
11. Vasak, M. & Hasler, D.W. (2000) Metallothioneins: new functional
and structural insights. Curr. Opin. Chem. Biol. 4, 177–183.
12. Viarengo, A., Burlando, B., Ceratto, N. & Panfoli, I. (2000)
Antioxidant role of metallothioneins: a comparative overview. Cell.
Mol. Biol. 46, 407–417.
13. Uchida, Y., Takio, K., Titani, K., Ihara, Y. & Tomonaga, M. (1991)
The growth inhibitory factor that is deficient in the Alzheimers-
disease brain is a 68-amino acid metallothionein-like protein.
Neuron 7, 337–347.
14. Yu, W.H., Lukiw, W.J., Bergeron, C., Niznik, H.B. & Fraser, P.E.
(2001) Metallothionein III is reduced in Alzheimer’s disease. Brain
Res. 894, 37–45.
15. Maret, W., Larsen, K.S. & Vallee, B.L. (1997) Coordination
dynamics of biological zinc ’clusters’ in metallothioneins and in
the DNA-binding domain of the transcription factor Gal4. Proc.
Natl Acad. Sci. USA 94, 2233–2237.
16. Churchich, J.E., Scholz, G. & Kwok, F. (1989) Activation of
pyridoxal kinase by metallothionein. Biochim. Biophys. Acta 996,
181–186.
17. Jiang, L.J., Maret. W. & Vallee. B.L. (1998) The glutathione redox
couple modulates zinc transfer from metallothionein to zinc-depleted
sorbitol dehydrogenase. Proc. Natl Acad. Sci. USA 95, 3483–3488.
18. Maret, W., Jacob, C., Vallee, B.L. & Fischer, E.H. (1999) Inhibitory
sites in enzymes: zinc removal and reactivation by thionein. Proc.
Natl Acad. Sci. USA 96, 1936–1940.
19. CanoGauci, D.F. & Sarkar, B. (1996) Reversible zinc exchange
between metallothionein and the estrogen receptor zinc finger.
FEBS Lett. 386, 1–4.
20. Zeng, J., Vallee, B.I. & Kagi, J.H.R. (1991) Zinc transfer from
transcription factor-IIIa fingers to thionein clusters. Proc. Natl
Acad. Sci. USA 88, 9984–9988.
21. Fliss, H. & Menard, M. (1991) Hypochlorous acid-induced mobiliz-
ation of zinc from metalloproteins. Arch. Biochem. Biophys. 287,
175–179.
22. Fliss, H. & Menard, M. (1992) Oxidant-induced mobilization of
zinc from metallothionein. Arch. Biochem. Biophys. 293, 195–199.
23. Maret, W. (1994) Oxidative metal release from metallothionein via
zinc thiol-disulfide interchange. Proc. Natl Acad. Sci. USA 91,
237–241.
24. Maret, W. & Vallee, B.L. (1998) Thiolate ligands in metallothio-
nein confer redox activity on zinc clusters. Proc. Natl Acad. Sci.
USA 95, 3478–3482.
25. Pearce, L.L., Wasserloos, K., St Croix, C.M., Gandley, R., Levitan,
E.S. & Pitt, B.R. (2000) Metallothionein, nitric oxide and zinc
homeostasis in vascular endothelial cells. J. Nutr. 130,
1467S–1470S.
26. Jiang, L.J., Maret, W. & Vallee, B.L. (1998) The ATP-metallo-
thionein complex. Proc. Natl Acad. Sci. USA 95, 9146–9149.
27. Zangger, K., Oz, G. & Armitage, I.M. (2000) Re-evaluation of
the binding of ATP to metallothionein. J. Biol. Chem. 275,
7534–7538.
28. Vasak, M. (1991) Standard isolation procedure for metallothionein.
Methods Enzymol. 205, 41–44.
29. Buhler, R.H.O. & Kagi, J.H.R. (1979) In Metallothionein (Kagi,
J.H.R.N. & Nordberg, M., eds), pp. 211–220. Birkhauser Verlag,
Basel, Switzerland.
30. Li, T.Y., Minkel, D.T., Shaw, C.F. & Petering, D.H. (1981) On the
reactivity of metallothioneins with 5,50-dithiobis-(2-nitrobenzoic
acid). Biochem. J. 193, 441–446.
31. Savas, M.M., Petering, D.H. & Shaw, C.F. (1991) On the rapid,
monophasic reaction of the rabbit liver metallothionein alpha-
domain with 5,50-dithiobis-(2-nitrobenzoic acid) (DTNB). Inorg.
Chem. 30, 581–583.
32. Zhu, Z.W., Goodrich, M., Isab, A.A. & Shaw, C.F. (1992) Biphasic
reactions of DTNB with lobster Cd6-metallothionein-2 and Cd5Cu-
metallothionein-2 which have 2 type-B (M3S9) Clusters. Inorg.
Chem. 31, 1662–1667.
33. Savas, M.M., Shaw, C.F. & Petering, D.H. (1993) The oxidation of
rabbit liver metallothionein-II by 5,50-dithiobis-(2-nitrobenzoic
acid) and glutathione disulfide. J. Inorg. Biochem. 52, 235–249.
34. Jiang, L.J., Vasak, M., Vallee, B.L. & Maret, W. (2000) Zinc
transfer potentials of the alpha- and beta-clusters of metallothionein
are affected by domain interactions in the whole molecule. Proc.
Natl Acad. Sci. USA 97, 2503–2508.
35. Xing, B.G., Shi, Y.B. & Tang, W.X. (2000) Kinetic studies of the
reactions of some metal reconstituted metallothioneins with the
electrophilic disulfide 5,50-dithiobis-(2-nitrobenzoic acid) (DTNB).
Biometals 13, 295–300.
36. Munoz, A., Petering, D.H. & Shaw, C.F. (1999) Reactions of
electrophilic reagents that target the thiolate groups of metallothio-
nein clusters: preferential reaction of the alpha-domain with 5,50-
dithio-bis(2-nitrobenzoate) (DTNB) and aurothiomalate (AuSTm).
Inorg. Chem. 38, 5655–5659.
37. Gan, T., Munoz, A., Shaw, C.F. & Petering, D.H. (1995) Reaction
of Cd111 (7)-metallothionein with EDTA 2 a reappraisal. J. Biol.
Chem. 270, 5339–5345.
38. Palumaa, P. & Vasak, M. (1992) Binding of inorganic phosphate to
the cadmium-induced dimeric form of metallothionein from rabbit
liver. Eur. J. Biochem. 205, 1131–1135.
39. Palumaa, P. & Vasak, M. (1993) Formation and spectroscopic
characterization of a novel monomeric cadmium- and phosphate-
containing form of metallothionein. Biochemistry 32,
2874–2879.
40. Cols, N., Romero-Isart, N., Capdevila, M., Oliva, B., Gonzalez-
Duarte, P., Gonzalez-Duarte, R. & Atrian, S. (1997) Binding of
excess cadmium (II) to Cd-7-metallothionein from recombinant
mouse Zn-7-metallothionein 1. UV-VIS absorption and circular
dichroism studies and theoretical location approach by surface
accessibility analysis. J. Inorg. Biochem. 68, 157–166.
41. Brouwer, M., Hoexum Brouwer, T.C. & Cashon, R.E. (1993) A
putative glutathione-binding site in Cd,Zn-metallothionein identified
by equilibrium binding and molecular-modeling studies. Biochem.
J. 294, 219–225.
4984 L. Kangur and P. Palumaa (Eur. J. Biochem. 268) q FEBS 2001




















