
The Certainty of Whuplak'n and the (un)Certainty of Extinction: Sinixt in British Columbia

Journal ofNeurology, Neurosurgery, and Psychiatry 1984;47: 1020-1028
The effects of movement velocity, mass displaced, andtask certainty on associated postural adjustmentsmade by normal and hemiplegic individualsFB HORAK, P ESSELMAN, ME ANDERSON, MK LYNCH
From the Departments ofRehabilitation Medicine and Physiology and Biophysics, University ofWashingtonSchool ofMedicine, Seattle, Washington, USA
SUMMARY The activation times for trunk and leg muscles were examined in normal and lefthemiplegic subjects who raised their right arms at different velocities in self-paced or reactiontime conditions. Activity in these postural muscles preceded arm displacement, and they were
activated in a similar sequence during all types of rapid movements. The presence and sequencingof associated postural adjustments were more variable during slow movements.
Movement of an extremity, such as the arm, causesdynamic forces to be applied to the trunk, and thesewill act on the multisegmented kinematic chain bet-ween the shoulder and the base of support. Theforces acting between a moving arm and the trunkare complex, but they must include linear centrifugalforces, inertial reactive forces, and torsional move-ments. In addition to the inertial reactions, themovement may cause a static displacement of bodymass with respect to the support base, such that apostural adjustment will be necessary to maintainstability by keeping the center of mass over the ver-tical projection of the supporting base. If stability isto be maintained during voluntary movement, it islikely that neurally controlled postural adjustmentor stabilisation must be produced by muscular activ-ity, as a supplement to reactive forces provided bystructures such as the ligaments. The required mus-cular activity could be controlled in two ways: (1) itcould be centrally programmed, as part of the totalmotor act, or (2) it could be initiated reflexly by theconsequences of the muscular contractions and/orthe movement.
Belen'kii et al' recorded the electromyographic(EMG) activity of the anterior deltoid and severalmuscles in the trunk and lower extremities in stand-ing normal individuals who rapidly raised their arm
Address for reprint requests: Dr Fay B Horak, Neurological Sci-ences Institute, Good Samaritan Hospital and Medical Center,1015 NW 22nd Avenue, Portland, Oregon 97210, USA
Received 16 May 1983 and in final revised form 7 March 1984.Accepted 10 March 1984
to a horizontal position following an auditory signal.Of the muscles examined, the earliest changes inEMG activity occurred in the biceps femoris and inthe lumbosacral muscles of the back. These changesin EMG activity of back and hip muscles precededactivation of anterior deltoid, the prime mover, byas much as 70-80 ms and occurred more rapidlythan voluntary reaction time activation of the sameback and hip muscles. Thus, it would appear thatthis axial muscle activity must be "preprogrammed"as part of the centrally initiated motor commandleading to arm movement and could not result fromsensory input initiated by the arm movement ormuscle activity in the prime movers at the shoulder.
If the preparatory postural muscle activity of sub-jects with neurological dysfunction is to be com-pared to the patterns seen in normal subjects, it isnecessary to know more about the normal variabilityof these patterns and the changes that different fac-tors might produce in preparatory muscle activity. Inthe present paper, we describe the effect of threefactors (movement velocity, mass displaced, andprior knowledge of the movement to be made) onthe timing of EMG activity in muscles acting at theshoulder, lower back, and hip in normal individualsof different ages. Because it also has been suggestedthat deficits in central programming of posturaladjustment and stabilisation may underly the post-ural instability present in many patients with CNSdefects resulting in hemiplegia or Parkinsonism,2-4we also report the patterns of EMG activity in hip,trunk, and shoulder muscles of individuals withleft-sided hemiplegia who were asked to raise theirright arms.
1020
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

The effects ofmovement velocity, mass displaced, and task certainty on associated postural adjustments 1021Preliminary results have been reported in abstract
form.5
MethodsSubjectsNormal individuals. Data were collected from 30 normalvolunteers, 15 men and 15 women, ranging in age from 20to 82 years (mean 47 5 years). There were five individualsin each of the five youngest decades, and the remaining fivewere between the ages of 70 and 82. All were active, many(even those in the 80th decade) engaged regularly in athle-tic activities, and all initially reported no neurological ormusculoskeletal defects. Upon careful questioning, how-ever, several admitted to previous fractures, sprains, ordislocations. Data from these subjects were comparedcarefully with those from the rest of the sample, and nonestood out as remarkable with the exception of those fromone woman who had experienced a right shoulder disloca-tion seven years prior to testing. This woman, however,also was one of the oldest subjects (age 75), and her datawere included because, although they fell on the extremeof the distribution, they were not distinctly separate fromdata from the rest of the subjects.Hemiplegic individuals. Data also were collected from
thirteen left-sided hemiplegic volunteers ranging in agefrom 25 to 79 years (mean 58-5 years). The elapsed timesince their strokes ranged from 2 months to 24 years. Fivewalked without braces and ambulation aids and wereactive enough to play golf or volleyball one-handed orwork part time. An additional two were fairly active anddid not use a wheelchair, although they did have braces,and two used canes as ambulation aids. The remaining sixused a wheelchair some or all of the time. All could standunsupported for the testing session, with rest breaks bet-ween each sequence.
Testing procedureSubjects stood on the floor in their stocking feet or flatshoes, with approximately 6 inch ( 15 cm) separation of theheels. Right arm position (shoulder joint angle) was meas-ured by an electrogoniometer arrangement. A long rigidaluminum tube was strapped to the arm with velcro bandsjust above the wrist and the elbow. This tube kept theelbow angle fixed in an extended position and accepted athin rod that slid longitudinally within the tube. The rodwas attached via a parallelogram with hinged corners to arotary potentiometer centered on the shoulder joint.6 Thisarrangement allowed sagittal rotational changes in shoul-der joint angle to be measured without the need for exactmatching of the mechanical coupling apparatus to transla-tional or adduction-abduction movements of the arm.Beckman miniature silver-silver chloride disc electrodes
were applied to the skin with adhesive collars positionedapproximately 2 cm apart near the midpoint of the rightanterior deltoid, right and left biceps femoris, and right andleft paraspinal (primarily erector spinae) muscles at thelevel of the iliac crest. Electrode placement was checked byrecording electromyographic (EMG) activity produced asthe subject flexed the shoulder (anterior deltoid), stoodupright from a flexed position at the waist (paraspinals),and flexed the knee to lift the foot, hyperextended the kneeand hip with the foot planted, and externally rotated the
leg at the hip (biceps femoris). Differentially-recordedEMG signals were preamplified with unity gain inputstages situated near the subject, and signals were transmit-ted via a photocouple to variable gain amplifiers for furtheramplification. Amplified EMG signals, potentiometer out-put, and DC voltage steps indicating signal light activation(see below) were recorded on 7 channel analog FM tape.Each normal subject was tested in four experimental
paradigms. These will be designated (1) fast self-paced, (2)slow self-paced, (3) reaction time, and (4) weighted self-paced. The instructions and subject behavior in each wereas follows:
(1) Fast self-paced. The subject stood with his arms at hisside and was asked to elevate his right arm in the parasagit-tal plane as quickly as possible to approximately horizon-tal, hold it there for a brief period of time, rapidly return tothe starting position, and repeat this sequence until askedto stop. He was to determine his own repetition rate, butduring the performance, subjects often were encouraged tosustain the start and elevated positions for longer periodsof time so that steady baseline position values could beobtained and swinging movements were eliminated. Sub-jects also were encouraged repeatedly throughout the trialsto make the movements as rapidly as possible.
(2) Slow self-paced. Subjects were asked to make armdisplacements similar to those described above, but atslower and more variable movement speeds. If subjectsmade only very rapid or very slow movements during thesetrials, they were asked to slow down or speed up theirmovements to insure a variety of movement velocities.
(3) Reaction time. Subjects faced two lights mounted atabout eye level. When the right light was illuminated, theywere to raise their right arm as rapidly as possible and keepit elevated until the light was extinguished. When the leftlight was illuminated, they were to raise their left arm(without an attached recording device) in a similar manner.The experimenter used low acoustic noise switches toilluminate the lights in random order and at variable inter-vals.
(4) Weighted self-paced. A 0 9 kg sandbag weight wasstrapped to the right wrist, and the subject was asked torepeat the rapid movements described in the fastself-paced paradigm.
All requested movements were demonstrated beforeeach test run, the subject made several practice move-ments, and then data were gathered for 20-30 trials duringeach paradigm.
Hemiplegic subjects were asked to do only the fast andweighted self-paced movements with their right, non-involved arm, each for 15-20 trials.
Data analysisTaped data were photographed from an oscilloscope traceonto moving kymograph film, and measurements of theduration and amplitude of the movement and time ofEMG onset for each muscle relative to movement onsetwere measured from projected film images. Movementonset was defined as the first detectable change in restingarm position measured from the potentiometer trace(fig 1). For normal subject data, the filmed image and timeand amplitude calibrations were projected onto an opaquesurface. The arm of an electronic digitiser (Numonics)
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from
Horak, Esselman, Anderson, LynchX1hJ!,dl Id hi IL
L Parospinci \ ,,A| - .-,r"... - -, FT L9 rwwv. v:hI.
Arm positinlOOms
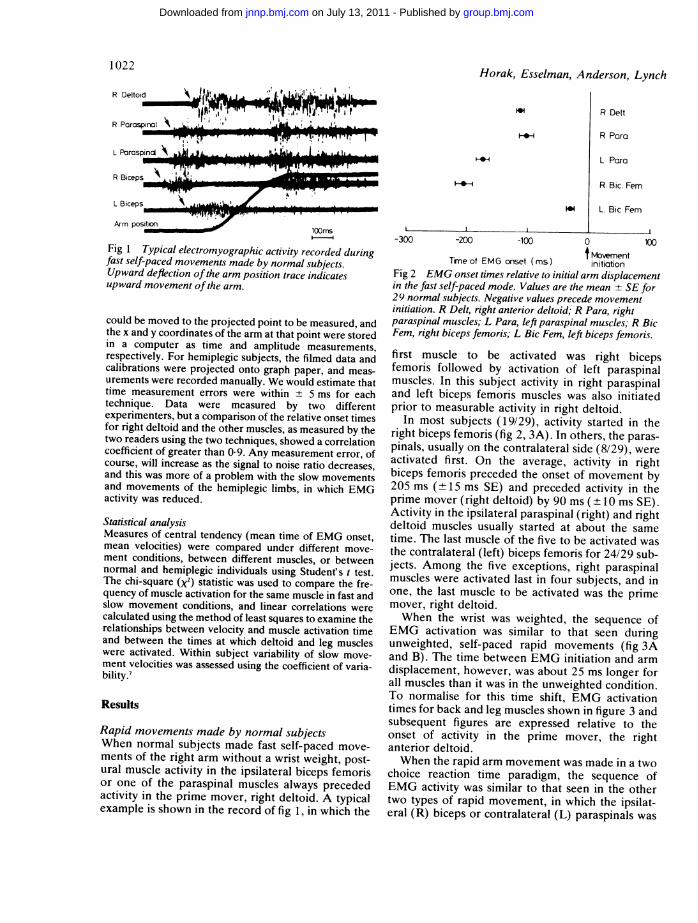
Fig 1 Typical electromyographic activity recorded duringfast self-paced movements made by normal subjects.Upward deflection ofthe arm position trace indicatesupward movement ofthe arm.
could be moved to the projected point to be measured, andthe x and y coordinates of the arm at that point were storedin a computer as time and amplitude measurements,respectively. For hemiplegic subjects, the filmed data andcalibrations were projected onto graph paper, and meas-urements were recorded manually. We would estimate thattime measurement errors were within + 5 ms for eachtechnique. Data were measured by two differentexperimenters, but a comparison of the relative onset timesfor right deltoid and the other muscles, as measured by thetwo readers using the two techniques, showed a correlationcoefficient of greater than 09. Any measurement error, ofcourse, will increase as the signal to noise ratio decreases,and this was more of a problem with the slow movementsand movements of the hemiplegic limbs, in which EMGactivity was reduced.
Statistical analysisMeasures of central tendency (mean time of EMG onset,mean velocities) were compared under different move-ment conditions, between different muscles, or betweennormal and hemiplegic individuals using Student's t test.The chi-square (x2) statistic was used to compare the fre-quency of muscle activation for the same muscle in fast andslow movement conditions, and linear correlations werecalculated using the method of least squares to examine therelationships between velocity and muscle activation timeand between the times at which deltoid and leg muscleswere activated. Within subject variability of slow move-ment velocities was assessed using the coefficient of varia-bility.'
Results
Rapid movements made by normal subjectsWhen normal subjects made fast self-paced move-ments of the right arm without a wrist weight, post-ural muscle activity in the ipsilateral biceps femorisor one of the paraspinal muscles always precededactivity in the prime mover, right deltoid. A typicalexample is shown in the record of fig 1, in which the
-300 -200 -100 0 100
tMovemnentTineof EMG onset (ms) initiationFig 2 EMG onset times relative to initial arm displacementin the fast self-paced mode. Values are the mean + SE for29 normal subjects. Negative values precede movementinitiation. R Delt, right anterior deltoid; R Para, rightparaspinal muscles; L Para, left paraspinal muscles; R BicFem, right biceps femoris; L Bic Fem, left biceps femoris.first muscle to be activated was right bicepsfemoris followed by activation of left paraspinalmuscles. In this subject activity in right paraspinaland left biceps femoris muscles was also initiatedprior to measurable activity in right deltoid.
In most subjects (19/29), activity started in theright biceps femoris (fig 2, 3A). In others, the paras-pinals, usually on the contralateral side (8/29), wereactivated first. On the average, activity in rightbiceps femoris preceded the onset of movement by205 ms (± 15 ms SE) and preceded activity in theprime mover (right deltoid) by 90 ms ( 10 ms SE).Activity in the ipsilateral paraspinal (right) and rightdeltoid muscles usually started at about the sametime. The last muscle of the five to be activated wasthe contralateral (left) biceps femoris for 24/29 sub-jects. Among the five exceptions, right paraspinalmuscles were activated last in four subjects, and inone, the last muscle to be activated was the primemover, right deltoid.When the wrist was weighted, the sequence of
EMG activation was similar to that seen duringunweighted, self-paced rapid movements (fig 3Aand B). The time between EMG initiation and armdisplacement, however, was about 25 ms longer forall muscles than it was in the unweighted condition.To normalise for this time shift, EMG activationtimes for back and leg muscles shown in figure 3 andsubsequent figures are expressed relative to theonset of activity in the prime mover, the rightanterior deltoid.When the rapid arm movement was made in a two
choice reaction time paradigm, the sequence ofEMG activity was similar to that seen in the othertwo types of rapid movement, in which the ipsilat-eral (R) biceps or contralateral (L) paraspinals was
1022
R Deltoid
.'
I.:
R Delt
R Para
L Para
R. Bic. FernL. Bic Fern
mmommommaim I p
I 6mwft..-
--. - Am I .
L B iceps 'k u 1.6. Ilm%. .ML - "NOM4
'I I !P.TW I rl" -v- Ifrllfdr"it"IR PcirctspirK31 . .0 I_,,&.L . - -1-i - 6
UPWI-11,'T IL i ..
DDtiA^ \ ';^r% D ceFb
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from
The effects ofmovement velocity, mass displaced, and task certainty on associated postural adjustments 1023
0
0
0
Fast self-pacedR Parm
@-* L Para0 RBF
LBF
Weighted
*
Reaction time
400
U,E
80
Et 300,9
1Da
>au
-i
-100 -50 0 50 100t R Delt
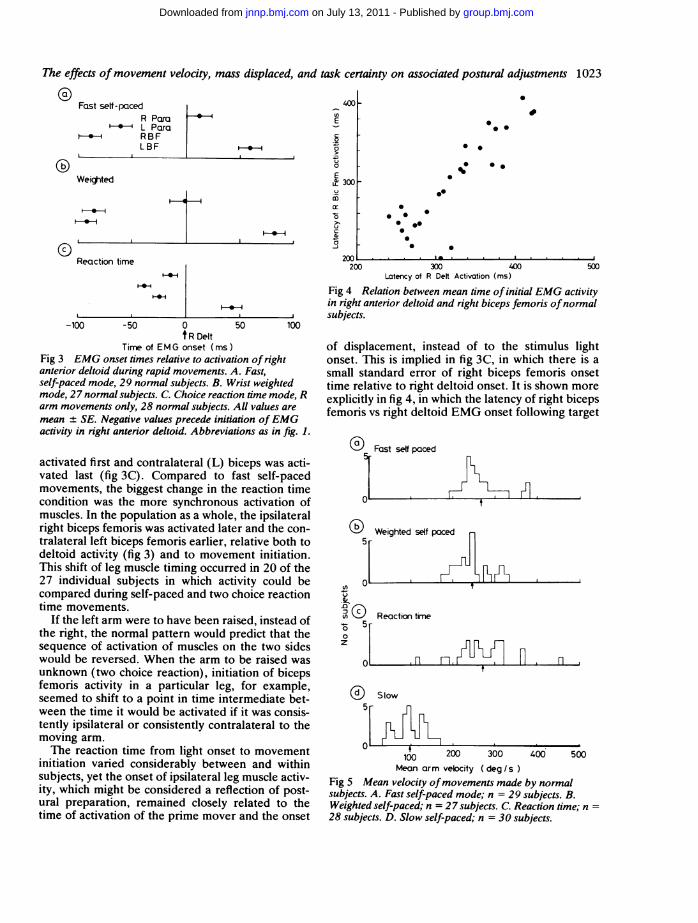
Time of EMG onset (ms)Fig 3 EMG onset times relative to activation of rightanterior deltoid during rapid movements. A. Fast,self-paced mode, 29 normal subjects. B. Wrist weightedmode, 27 normal subjects. C. Choice reaction time mode, Rarm movements only, 28 normal subjects. All values aremean + SE. Negative values precede initiation ofEMGactivity in right anterior deltoid. Abbreviations as in fig. 1.
activated first and contralateral (L) biceps was acti-vated last (fig 3C). Compared to fast self-pacedmovements, the biggest change in the reaction timecondition was the more synchronous activation ofmuscles. In the population as a whole, the ipsilateralright biceps femoris was activated later and the con-tralateral left biceps femoris earlier, relative both todeltoid activity (fig 3) and to movement initiation.This shift of leg muscle timing occurred in 20 of the27 individual subjects in which activity could becompared during self-paced and two choice reactiontime movements.
If the left arm were to have been raised, instead ofthe right, the normal pattern would predict that thesequence of activation of muscles on the two sideswould be reversed. When the arm to be raised wasunknown (two choice reaction), initiation of bicepsfemoris activity in a particular leg, for example,seemed to shift to a point in time intermediate bet-ween the time it would be activated if it was consis-tently ipsilateral or consistently contralateral to themoving arm.The reaction time from light onset to movement
initiation varied considerably between and withinsubjects, yet the onset of ipsilateral leg muscle activ-ity, which might be considered a reflection of post-ural preparation, remained closely related to thetime of activation of the prime mover and the onset
.
0* 0
; *.
*0
0*a* 0S00
.
0
200 300 400 500Latency of R Deft Activation (ms)
Fig 4 Relation between mean time of initial EMG activityin right anterior deltoid and right biceps femoris ofnormalsubjects.
of displacement, instead of to the stimulus lightonset. This is implied in fig 3C, in which there is asmall standard error of right biceps femoris onsettime relative to right deltoid onset. It is shown moreexplicitly in fig 4, in which the latency of right bicepsfemoris vs right deltoid EMG onset following target
'-' Fast self paced
il-
( Weighted self paced5
ni L- A Lu
.5-3 O Reaction time
z
Fn. l n
OD Slow
5p
200 300 400 500Mean arm velocity ( deg / s)
Fig 5 Mean velocity ofmovements made by normalsubjects. A. Fast self-paced mode; n = 29 subjects. B.Weighted self-paced; n = 27 subjects. C. Reaction time; n28 subjects. D. Slow self-paced; n = 30 subjects.
. n
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

1024
light onset is plotted for each subjitime of EMG initiation in both musc
siderably, consistent with a wide rang4tion times for the various subjects, btimes in the two muscles were positiwith a correlation coefficient of 0-92Mean velocities for movements ma
ject in the fast self-paced, weightedtime conditions are shown in fig 5, Avelocities for the sample, as indicatecin the histograms, were 267, 244, anmfor fast, weighted, and reaction tilrespectively. Mean velocities of w
ments were significantly slower thatself-paced or reaction time movemei0-001). There was no significant diever, between movement velocities ipaced and reaction time modes, alth(bility of velocities of reaction time n
greater.
Slow, self-paced movements made bjectsWhen subjects were asked to raise tislowly, the mean velocity of movemesubjects (97.9 deg/s +6-5 deg/s SE)half the velocity of the fast, self-paci(267 deg/s, fig 5D). Under these conding, and often the sequencing, of actious muscles changed from those seemovements.Although the initial measurable
during slow movements most often w;
R Para
L Para A
R. Bic Fem i-
L Bic Fem
-200
F-
I-
-100 0
R DeltTime of EMG onset (ims
Fig 6 EMG onset times relative to activatanterior deltoid during slow movements misubjects. Mean + SE of onset times are shcmovements(-), for movements with meanS100 degls.(-), and for movements with>100 degls (A). Negative values precede iEMG activity in right anterior deltoid. Abfig
Horak, Esselman, Anderson, Lynch
ect. The mean (right) biceps femoris (11/30 subjects) or one of theles varied con- paraspinal muscles (ipsilateral in 2/30 subjects, con-e of mean reac- tralateral in 8/30 subjects), in nine of the 30 sub-ut EMG onset jects, deltoid showed the earliest measurable activ-vely correlated ity. When expressed relative to right deltoid activa-!(p < 0.0001). tion, activity in right biceps femoris and the paraspi-de by each sub- nal muscles on both sides was initiated later during1, and reaction slow movements than during rapid self-pacedi-C. The mean movements (compare fig 6, circles, with fig 3A).I by the arrows There also was considerably more variation in thed 270 degrees/s time at which individual muscles were activated, andme conditions, as a result, the mean onset times for the ipsilateraleighted move- (right) biceps femoris and contralateral (left) paras-n those of fast pinals for the entire sample were later than that ofnts (t test, p < right deltoid (fig 6), even though one of the formerifference, how- two muscle groups was activated before deltoid inin the fast self- most individual subjects. The right biceps femoris,ough the varia- especially, showed considerable variation in time ofnovements was activation and actually was the last muscle to be
activated in eight of the 30 subjects. This is in con-trast to the muscle activation sequence for fast self-
y normal sub- paced movements, in which this muscle never hadthe latest mean onset time in any of the subjects
heir right arms tested.nts made by all During slow self-paced movements, some backwas less than and leg muscles never showed measurable activity,ed movements or the activity appeared in fewer trials than it didlitions, the tim- during rapid movement trials. For example, thevity in the vari- mean probability with which EMG activity wasn during rapid recorded in right biceps femoris for all subjects
dropped from 0-93 during fast self-paced move-EMG activity ments to 0-67 during slow movements. For the leftas in ipsilateral biceps femoris EMG, the drop was from 0-98 (fast)
to 0-70 (slow). This change was significant at the-*-------I 0-05 level for both leg muscles (x2). For paraspinal
muscles, there was only a small, insignificant trendtoward a reduced incidence of measurable activity
-I during slow movements, compared to rapid ones.If subjects are divided into those with mean slow
movement velocities less than 100 deg/s and thosegreater or equal to 100 deg/s (fig 6, squares andtriangles, respectively), then the mean EMG onsettimes (relative to deltoid activity) were consistently
A* , | earliest in those suibjects with higher mean velocities
(triangles).100 200 The trial-to-trial variability in velocity of slow
movements made by individual subjects also wasgreater than it was for fast self-paced movements.The mean coefficient of variability was 0-11 for theaion ofright velocities of fast, self-paced and 0-33 for the vel-ade bynormal ocities of slow self-paced movements. The variabil-
velocities ity in the velocity of movements made by each sub-I mean velocities ject allowed us to examine the correlation betweeninitiation of movement velocity on individual trials and the onset5reviations as in times of the muscles studied. In many individual
subjects, there also was a statistically significant
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

The effects ofmovement velocity, mass displaced, and task certainty on associated postural adjustments 1025
inverse correlation between the time of muscle acvation prior to movement onset and the velocitythe associated movement. This tendency for muscto become activated earlier with higher velocmovements was most often significant for the priimover, right deltoid (16 of 30 subjects; p < 0-05=-0 430 to -0.874). For the other muscles, whmeasurable activity was present in more than fivethe slow movement trials, the correlation betwemovement velocity and the lead time at whichmeasurable increase in EMG activity coulddetected was significant (p < 0-05) for right parasnals in eight of 25 subjects, for left paraspinals inof 22 subjects, for right biceps femoris in eight ofsubjects, and for left biceps femoris in 10 of 20 stjects.
Relationships between movement parameters and aTo allow future comparison of movement variabbetween normal subjects and patient populaticwith restricted age distributions, we examined trelationship between age and the movement amuscle timing variables measured.
Figure 7 shows mean movement velocity plottas a function of age for each of the normal subjestudied in the fast self-paced, reaction time, weigted, and slow self-paced conditions. There wastendency for the velocity to be inversely correlatwith age in the fast and weighted self-paced contions, but this was statistically significant only iweighted movements (r = 0-49, p < 0-01). The laof a significant correlation in the fast self-pacmode is, however, largely the result of the rajmovements made by the 82 yr-old-subject, and
(i) Fast self-paced
4001
300[tn
' 200
100 __ .j __
(b Weighted0> 400-E
c 300...
2000
100 ,'.
0 Reaction time
* *- c * : :.S.-:0 ::
*.0
(i) Slow
20 40 60 80 20 40 60 8CAge (years)
Fig 7 Relation between mean arm velocity and age fornormal subjects studied during fast self-paced (A), wristweighted (B), reaction time (C), and slow self-pacedmovements (D).
cti-of
-lescityvme5, renof'enh abe,pi-1119.19ub-
zgelesons
one compares mean velocities for subjects underand over 50 years of age, the velocities aresignificantly slower for the older subjects duringboth the fast self-paced and the weighted self-pacedmovements (t test, p < 0-001). Movement velocitiesfor subjects under and over 50 years of age were notsignificantly different, however, for movementsmade in the reaction time or slow paradigms.
Older subjects did tend to initiate movementsafter a longer delay in the reaction time paradigm, asshown by a statistically significant correlation bet-ween age and reaction time (r = +0 33, p < 0-05).
In spite of the significant interaction between ageand movement velocity and the weak interactionbetween age and reaction time, there were nosignificant correlations between age and the timerelative to deltoid activity at which other individualmuscles were activated in any of the movement con-ditions studied.
the Rapid movements made by hemiplegic subjectsLnd Many movements made with the right arm by left
hemiplegic subjects, even in the fast self-paced orted weighted paradigms, were much slower than thosects made by normal subjects. For example, the dura-gh- tions of fast, self-paced movements ranged from 198s a to 622 ms for normal subjects, whereas they rangedted from 355 to 1256 ms for hemiplegic individuals. Todi- allow comparisons between EMG patterns associ-for ated with rapid movements made by the two groupsack of subjects, only those fast self-paced and weightedwed movements with durations less than 800 ms werepid analyzed quantitatively for hemiplegic subjects.if The mean times of initial EMG activation, ex-
pressed relative to right deltoid onset time, arerepresented by the circles in fig 8 for the four backand leg muscles. The sequences of muscle activationin both the fast self-paced and weighted self-pacedmodes were similar to those seen in normal subjects,although the latency of postural muscle activity onthe hemiplegic side relative to deltoid activity waslater than in normal subjects (triangles in fig 8).Right biceps femoris was the first muscle activated inall hemiplegic subjects studied, both during fastself-paced and weighted movements. In these 13
* hemiplegic subjects, activation of right bicepsfemoris preceded right deltoid activity by 106-2 ms(+ 110 ms SE) in the fast self-paced mode and by74-0 ms (+15-6 ms SE) in the weighted mode.These values were not significantly different from
0 those measured for normal subjects.The paraspinal muscles on the left (hemiplegic)
side were activated next after right biceps femoris innine of the 13 hemiplegic subjects, at an average of47-7 ms (±+ 12-2 ms SE) prior to right deltoid activa-tion during fast self-paced movements and 13-2 ms
00
0* * 0
%* O. 0
"la 0 0O* .0 0
00 4
0 0
00 0
0
* e 0 : I
000
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

Horak, Esselman, Anderson, LynchC) Fast self-pacedR ParaL Para aRBF F-t&LBF
(c) Weighted
A
(fig 2). If the function of the left biceps femorisactivity was to provide hip extensor activity neces-sary for support when the trunk weight was shifteddiagonally away from the raised right arm, then itmight be predicted that the time of left bicepsfemoris activation would have a closer relationshipto the time at which arm elevation was completedthan it would to the time at which it was initiated.This, in fact, was the case. On the average, leftbiceps femoris activity was initiated 443 ms prior totermination of the fast self-paced right arm move-ments made by hemiplegic subjects and 426 ms
-200 -100 0 m 20
f R DeltTime of EMG onset (ims
Fig 8 EMG onset times relative to activation of rightanterior deltoid for left hemiplegic subjects (circles)compared to normal individuals (triangles) taken fromfig 3A and B. A. Fast self-paced mode; n = 13. B. Weightedmode; n = 8. All values for hemiplegic subjects are meansSE. Negative values precede initiation ofEMG activity inright anterior deltoid.
(+ 13-7 ms SE) before right deltoid activation dur-ing weighted movements. These contralateral para-spinal muscles were activated well in advance ofright deltoid in both normal and hemiplegic sub-jects, in the fast self-paced condition, but there wasno significant difference in the activation time be-tween the two groups of subjects. For weightedmovements, however, left paraspinal activity wasmeasured later in hemiplegic than in normalsubjects (p < 0-001).As was the case for normal subjects, the ipsilateral
(right) paraspinal muscles were activated at aboutthe same time as the right deltoid in the fast self-paced condition. The slightly earlier onset in hemi-plegics, compared to normals, during fast self-pacedmovements (compare fig 8A and 3A) was notsignificant at the 0-05 level, and there also was nosignificant difference between the time of right para-spinal activation during weighted movements.As in most normal subjects, the left (hemiplegic)
biceps femoris was the last muscle to show measur-able activity in 11 of 13 hemiplegic subjects studiedduring fast self-paced movements and in all of theeight subjects studied during weighted movements.In both conditions, the onset of measurable activityoccurred later in hemiplegic than in normal indi-viduals (compare circles and triangles in fig 8A andB), p < 0-001). Measurable left biceps femoris activ-ity during fast, self-paced movements of the rightarm was initiated at a mean of 104 ms after move-ment initiation in left hemiplegic individuals,whereas in normal individuals, left biceps femorisactivity was initiated 37 ms prior to movement onset
prior to termination of fast self-'paced movementsmade by normal subjects.Discussion
The results of the present study indicate that whenfreestanding individuals raise one arm as rapidly aspossible, they use essentially the same sequence ofhip, back, and shoulder muscle activation, whetherthe movement is self-initiated or visually triggered,and regardless of the load, their advance knowledgeof which arm is to be raised, or the presence ofcontralateral hemiplegia consequent to a cerebralvascular accident. When they raise the arm moreslowly, the sequence of muscle activation becomesmore variable, and some muscles with early activityprior to rapid movement show no measurable activ-ity at all.Other investigators have examined the activity of
leg and, in some cases, back muscles during simplereaction time tasks requiring unilateral elevation ofthe same arm following either an auditory' or a vis-ual stimulus.8 9 All of these groups found essentiallythe same sequence of activity in back and hip exten-sor muscles as was seen during any of the rapidmovements in the present study. The lead time ofipsilateral hip muscle activation relative to activa-tion of the prime mover varied, however, duringrapid movements made under different conditions.In the simple reaction time condition, in which thesame arm was raised on each trial and, as a consequ-ence, there was temporal, but no spatial uncertaintyabout the movement to be made, ipsilateral hipextensor activity led deltoid activity by about 40-55ms (biceps femoris, Belen'kii et al;' semitendinosus,Bouisset and Zattara8) although in the report byLee,9 this timing difference was not statisticallysignificant. These values during non-choice reactiontime trials were intermediate between the bicepsfemoris lead time (93 ms) measured in the fast self-paced condition of the present study, in which therewas neither spatial nor temporal uncertainty, andthe lead time (23 ms) of the same muscle in the twochoice reaction time condition, in which both tem-
1026
"_4
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

The effects ofmovement velocity, mass displaced, and task certainty on associated postural adjustments 1027poral and spatial uncertainty existed. Thus, althoughspatial and temporal uncertainty did not change thesequential pattern of muscle activation, they didchange the timing of postural muscle activity.There was a strong correlation between the reac-
tion time onset of EMG activity in the prime mover(right deltoid) and in the muscle (right bicepsfemoris) with the earliest associated posturaladjustment activity, indicating that the early hipmuscle activity was more closely related to themovement initiation than to the stimulus light onset.Cordo and Nashner'° reported only a non-significanttrend for this relationship, but this may have beendue to their rather small sample size.The relative timing of EMG activity was much
more sensitive to the mean velocity of the move-ment than it was to the mass displaced. This isperhaps not surprising, since if the preparatoryEMG activity serves to help compensate for themaximum disturbing forces due to inertial and tor-sional moments, compensation must be maximumbetween the maximum acceleration and maximumvelocity phases of the movement, irrespective of themass displaced. If we had been able to quantify theamplitude of EMG activity or the force it produced,it is probable that these would have varied morewith mass displaced than did the timing of EMGonset.The major measured variable that influenced both
the timing of measurable EMG activity and theprobability of its occurrence in back and leg muscleswas the velocity of the ensuing movement. It is truethat the lower magnitude of EMG activity duringslow movements decreased the signal to noise ratioand would have delayed the time at which EMGactivity was detectable and, hence, increased thetiming measurement error. The amplitude of EMGactivity in anterior deltoid also was decreased, how-ever, and since timing was expressed relative to rightdeltoid activity, it is unlikely that measurementerrors could completely explain the results. It ismore probable that, since less stabilisation force isnecessary during low acceleration and low velocitymovements, this aspect of preparatory muscle activ-ity simply is not programmed as rigidly in associa-tion with slow movements. The relation betweenmovement velocity and EMG onset times is impor-tant to consider in studies of associated posturaladjustments made by aged or neurologicallyimpaired subjects, some of whom may make move-ments at slower velocities. Changes in associatedpostural adjustments may be related to the velocityof the movements, and not to the age or pathology,per se.
In addition to the influence of acceleration andvelocity on early preparatory-EMG activity, the sub-
ject' s initial postural stability also affects this pre-paratory activity. Cordo and Nashner'° found thatthe early changes in EMG activity recorded in legmuscles prior to rapid, self-initiated displacement orsound-triggered push or pull movements of the armwere markedly decreased in amplitude if the subjectwas supported at the shoulder or even touched aguard rail with a single finger.
Because these early associated postural adjust-ments precede any displacement, or even any con-traction of the prime movers, it is hypothesized thatthey are centrally generated and are not reflexive innature.' 49I0 Since damage to CNS structures couldinterrupt the ability to generate this normal patternof associated postural adjustments, it is of interest toexamine the activity of muscles on a hemiparetic orhemiplegic side of the body when the contralateral(normal) arm is activated. In our left hemiplegicsubjects, the sequence of activity in the five musclesstudied was the same as that seen in normal subjects,both in the weighted and unweighted conditions.Measurable activity on the contralateral hemipareticside, both at the waist and the hip, occurred at alater time than it did in normal individuals, espe-cially during weighted movements. This occurreddespite the fact that the patients displayed varyingamounts of spasticity on the hemiplegic side. Therelative latencies of muscle activity on the non-hemiplegic side (ipsilateral to the raised arm) werenormal. Pal'tsev and El'ner3 reported that if subjectswith frontal lobe damage raised their paretic arm,activity in the ipsilateral (hemiparetic) bicepsfemoris also occurred later than it did in normals,but they did not present enough data to determinewhether its activation was delayed when it was con-tralateral to the raised normal arm.The delay in preparatory postural activity on the
hemiplegic side during self paced movements wasmore striking in the left biceps femoris than in theleft paraspinal muscles (fig 8). Although greaterbilateral innervation of trunk muscles than leg mus-cles may account for this difference, any possiblesparing of innervation was inadequate to preventstatistically significant delays in left paraspinal onsetunder conditions of increased load.Although our hemiplegic subjects raised their
arms more slowly than normal subjects when askedto move as rapidly as possible, the pattern of pre-paratory postural activity in hemiplegic subjectsdoes not resemble the patterns seen in normal sub-jects moving their arms at slower velocities (com-pare figs 6 & 8A). When normal subjects raisedtheir arms slowly, right paraspinal and right bicepsfemoris activity was usually initiated after rightdeltoid. In contrast, the hemiplegic subjects demon-strated a sequence in the activity of leg and back
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

1028
muscles on the non-hemiplegic (right) side that wassimilar to that seen in the faster moving normal sub-jects. In fact, we may speculate that preparatorypostural muscle activity on the non-hemiplegic sidewas utilised by hemiplegic subjects at slower armvelocities in order to compensate for their more pre-carious stance.
Thus, any preprogrammed descending signal thatwould determine the bilateral sequence of muscleactivation seemed to be able to reach the cord.Whether such signals descend on separate sides ofthe brainstem or whether they descend on one sideand distribute bilaterally at the lower brainstem orcord is not known, although associated posturalresponses will occur bilaterally in cats after motorcortex lesions on one side."l Either might have beenthe case in our hemiplegic subjects, since the lateractivation of muscles on the hemiparetic side couldhave resulted either because information descendingon the undamaged side reached a less excitable con-tralateral motoneuronal pool or because the numberof undamaged descending fibres on the side of theinjury simply was insufficient to activate motor-neurons on the hemiplegic side quite as early as wasthe case for normal subjects. Irrespective of whetherthe delay in timing is determined at a high or a lowlevel, it is possible that hemiplegic patients were notcapable of making movements of the "uninvolved"arm as rapidly as normal because of delays in anti-cipatory activity of contralateral hemiplegic musclesimportant in postural stabilisation.
Early associated postural adjustments presumablyprovide stability for the ensuing movement. Of thevariables examined in this study, the velocity of themovement to be made appeared to be mostsignificant in determining the presence and timing ofactivity in the postural muscles examined.
It is a pleasure to acknowledge the technical assis-tance of Mark Guthrie and Beverly Bedell.
Horak, Esselman, Anderson, LynchThis study was supported in part by Research Grant#G008200020 from the National Institute of Hand-icapped Research, Department of Education,Washington, DC 20202.
References
'Belen'kii VY, Gurfinkel VS, Pal'tsev YI. Elements ofcontrol of voluntary movements. Biophysics1967; 12:135-41.
2 Martin JP. The. Basal Ganglia and Posture. Philadelphia:Lippincott 1967.
3Pal'tsev YI, El'ner AM. Preparatory and compensatoryperiod during voluntary movement in patients withinvolvement of the brain of different localization.Biophysics, 1967; 12:161-8.
4Traub MM, Rothwell JC, Marsden CD. Anticipatorypostural reflexes in Parkinson's disease and otherakinetic-rigid syndromes and in cerebellar ataxia.Brain 1980; 103:393-412.
Horak FB, Anderson ME. Preparatory postural activityassociated with movement. Physical Therapy1980;60:580.
6 Lamorux LW. Kinematic measurements in the study ofhuman walking. Bulletin of Prosthetic Research1971; 10: 3-84.
Snedecor GW, Cochran WG. Statistical Methods. Ames:The Iowa State Press 1967.
8 Bouisset S, Zattara M. A sequence of postural move-ments precedes voluntary movement. NeuroscienceLetters 1967; 12:135-41.
9 Lee WA. Anticipatory control of postural and task mus-cles during rapid arm flexion. Journal of MotorBehavior 1980; 12:185-96.
'° Cordo PJ, Nashner LM. Properties of postural adjust-ment associated with rapid arm movements. JNeurophysiol 1982;47:287-302.
"Massion J. Role of motor cortex in postural adjustmentsassociated with movement. In: Asanuma H and Wil-son VJ, eds. Integration in the nervous system.Tokyo-New York: Igaku-Shoin, 1979.
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from

doi: 10.1136/jnnp.47.9.1020 1984 47: 1020-1028J Neurol Neurosurg Psychiatry
F B Horak, P Esselman, M E Anderson, et al. hemiplegic individuals.adjustments made by normal andon associated postural
certaintymass displaced, and task The effects of movement velocity,
http://jnnp.bmj.com/content/47/9/1020Updated information and services can be found at:
These include:
References http://jnnp.bmj.com/content/47/9/1020#related-urls
Article cited in:
serviceEmail alerting
online article.article. Sign up in the box at the top right corner of the Receive free email alerts when new articles cite this
Notes
http://group.bmj.com/group/rights-licensing/permissionsTo request permissions go to:
http://journals.bmj.com/cgi/reprintformTo order reprints go to:
http://group.bmj.com/subscribe/To subscribe to BMJ go to:
group.bmj.com on July 13, 2011 - Published by jnnp.bmj.comDownloaded from
Copyright © 2022 FDOKUMEN














![Meaningful Adjustments: Musical Performance and Ritual Action in a South Indian Temple [AAWM 4-1]](https://static.fdokumen.com/doc/165x107/63333237b94d623842021472/meaningful-adjustments-musical-performance-and-ritual-action-in-a-south-indian.jpg)





