
164 CHARACTERIZATION OF THE COMMON ELAND ( TAUROTRAGUS ORYX ) ESTROUS CYCLE

Physiology & Behavior 95 (2008) 484–491
Contents lists available at ScienceDirect
Physiology & Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /phb
The effects of metergoline and 8-OH-DPAT injections into arcuate nucleus and lateralhypothalamic area on feeding in female rats during the estrous cycle
Sérgio Murilo Steffens, Isabel Cristina da Cunha, Danielle Beckman, Ana Paula Fraga Lopes,Moacir Serralvo Faria, José Marino-Neto, Marta Aparecida Paschoalini ⁎Department of Physiological Sciences, Centre of Biological Sciences, Federal University of Santa Catarina (UFSC), Florianópolis, SC, Brazil
Abbreviations: 3V, 3rd ventricle; ARC, arcuate hypopeduncle; f, fornix; ic, internal capsule; mt, mammillothpart of lateral hypoyhalamus; PeFLH, perifornical partpremammillary nucleus, ventral part; VMH, ventromezona incerta.⁎ Corresponding author. Department of Physiolog
University of Santa Catarina, 88040-900 Florianópolis9352; fax: +55 48 3721 9672.
E-mail address: [email protected] (M.A. Paschoalini
0031-9384/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.physbeh.2008.07.019
a b s t r a c t
a r t i c l e i n f oArticle history:
The present study examine Received 11 April 2008Received in revised form 10 July 2008Accepted 17 July 2008Keywords:Metergoline8-OH-DPATFood intakeEstrous cycleHypothalamus
d the effects of local injections of metergoline (MET, an antagonist of 5-HT1/2receptors, 2 and 20 nmol) and 8-hydroxy-2-(di-n-propylamino)-tetralin (8-OH-DPAT, selective 5-HT1Areceptor agonist, 0.6 and 6 nmol) into the arcuate nucleus (ARC) and the lateral hypothalamus (LH), oningestive and non-ingestive behaviors of female rats. These effects were examined during the diurnal periodsof diestrus and estrus in rats adapted to eat a wet mash diet (enriched with 10% sucrose) during 1h for 3consecutive days at the recording chamber. The results showed that 8-OH-DPAT injected into the LHsignificantly reduced food intake at all doses and both cycle stages, while in the ARC these treatments evokedhypophagia only at the highest 8-OH-DPAT dose and only at the estrous phase. MET administered into theARC (at all doses) failed to affect food intake during both estrous stages. On the other hand, food intakedecreased after injection of both doses of MET into the LH of rats during estrous and diestrus phases. In estrusstage, injections of the higher dose of 8-OH-DPAT into the ARC and into the LH decreased the duration offeeding. Latency to start feeding, drinking, and non-ingestive behaviors were not affected by 8-OH-DPAT orMET treatments in the ARC or the LH in both cycle phases. These results indicated that 5-HT1A receptorsparticipate in the serotonergic control of feeding-related mechanisms located at the ARC and the LH. Thesefeeding-related serotonergic circuits in both areas are possibly affected by ovarian hormones that couldincrease sensitivity of ARC neurons to the hypophagic effects of 8-OH-DPAT or increase the efficacy of satietysignals that terminate feeding. In addition, the present data indicated that serotonergic inputs do not exert atonic inhibitory activity on the ARC and the LH feeding-related circuits.
© 2008 Elsevier Inc. All rights reserved.
1. Introduction
Serotonin (5-hydroxytryptamine; 5-HT) plays an important role inthe regulation of food intake [6,15,79]. Activation of 5-HT2C or 5-HT1Breceptors decreases feeding behavior [14,41,45,46,53,76], while activa-tion of 5-HT1A receptors can cause hyperphagia [9,11,17,18,21,35,71] orreduction in food intake [1,19,20,22,23,62]. These ingestive effectsseem to bemediated primarily by 5-HT1A, 5-HT1B and 5-HT2C receptorspresent in the raphe nuclei, amygdaloid complex and hypothalamus[9,11,14,44,45,48,57,71]. The hyperphagic effects of systemic orintracerebroventricular (ICV) injections of metergoline (MET; nonselective antagonist 5-HT1/2) in free-feeding or fully satiated rats
thalamic nucleus; cp, cerebralalamic tract; PLH, peduncular
of lateral hypothalamus; PMV,dial hypothalamic nucleus; ZI,
ical Sciences – CCB, Federal, SC, Brazil. Tel.: +55 48 3721
).
l rights reserved.
[10,12,28,58,80] have been interpreted as suggestive of the presence ofa tonic inhibitory 5-HTaction on central feeding circuits. The posteriorbasolateral amygdala is a neural site where 5-HT terminals may exertsuch tonic inhibitory influence, since electrolytic lesions in poster-odorsal amygdaloid area attenuated feeding evoked by intrarapheinjections of 8-OH-DPAT, and lesions restricted to the posteriorbasolateral amygdala completely block hyperphagic responsesinduced by systemic injection of 8-OH-DPAT [9,71]. In addition, METinjections into the posterior basolateral amygdala elicit hyperphagicresponses [69,70].
The paraventricular nucleus (PVN) of the hypothalamus alsoseems to be involved in the hypophagic effects of 5-HT. Direct in-fusion of 5-HT, of various 5-HT receptor agonists and of 5-HT reuptakeinhibitors into the PVN, suppresses food intake evoked by a variety ofsituations [12,29,37,59,62,64], while intra-PVN injections of METantagonize the hypophagia after 5-HT injections into the PVN [12].However, intra-PVN injections of MET alone failed to affect foodintake in free-feeding rats [10,58], suggesting that 5-HT inputs to thePVN may exert only phasic (but not tonic) effects on feeding behavior.A feeding-related regulatory activity by 5-HT has also been observedin other hypothalamic nuclei implicated in the regulation of energy

485S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
balance such as the arcuate nucleus (ARC) and the lateral hypotha-lamus (LH), that express orexigenic and anorexigenic peptides[44,45,47,84]. It has been demonstrated that 5-HT2C receptor acti-vation stimulates the ARC neurons expressing the anorectic peptidemelanocortin (α-MSH) precursor (the proopiomelanocortin, POMC),while activation of ARC 5-HT1B receptors both inhibits the neuronalactivity of cells expressing the orexigenic peptide agouti-relatedprotein (AgRP) and decreases inhibitory postsynaptic currents ontoPOMC neurons. This control of melanocortin systemmay represent animportant mechanism through which 5-HT reduces food intake[44,45]. The extent to which the 5-HT influences on the hypothalamicregions other than the PVN are related to phasic phenomena or to atonic activity remains to be determined.
Serotonergic influences on feeding-related hypothalamic neuronsmay be also mediated by 5-HT1A receptors. High densities of thesereceptors were revealed in neurons of the PVN, the supraopticnucleus, the ventromedial nucleus, as well as in the ARC and the LHarea, by the use of an antiserum to the 5-HT1A receptor [10]. In thatstudy, Collin and colleagues also showed that 5-HT1A receptorimmunoreactivity was especially intense in neuropeptide Y (NPY)-,Agrp-, POMC- and cocaine- and amphetamine-regulated transcript-containing (CART) neurons of the ARC. In the LH, 5-HT1A receptorimmunoreactivity was observed in melanin-concentrating hormone-and orexin-containing neurons, suggesting that an important action ofserotonergic input to these peptidergic neurons in the ARC and LHmay be mediated by 5-HT1A receptors. As the 5-HT1A immunoreactiv-ity was never seen in nerve fibers or nerve endings [8], the control ofneuropeptide released by 5-HT may be mediated by possiblypostsynaptic 5-HT1A receptors. To the best of our knowledge, thereare no functional studies in the literature on the possible participationof these receptors on feeding mechanisms present in the ARC or in theLH.
Feeding control by serotonergic circuits seems to be influenced bythe fluctuation in concentration of ovarian steroids during the estrouscycle. Rats in the diestrus phase have been reported to be moresensitive to hyperphagic effects of systemic 8-OH-DPAT injectionsthan rats in estrus phase [83]. Furthermore, MET injections in theposterior basolateral amygdala differentially affect feeding of pala-table diets over the estrous cycle: the hyperphagic effects of theseinjections are more evident in diestrus than in estrus female rats [70].In the same line, it has been shown that the anorectic effect offenfluramine is more intense in estrous as compared to diestrous stageof estrous cycle females [27]. Still, the eating-inhibitory effect offenfluramine was increased by estrogen treatment in ovariectomizedrats [74]. Additionally, estrogen treatment reduced 8-OH-DPAT-induced hyperphagia in ovariectomized, hormone-primed rats [75].In the present work, we examined the behavioral and ingestive effectsof local microinjections of METand of 8-OH-DPAT into the ARC and theLH. We also investigated whether or not the different estrous cyclephases influence these effects.
2. Materials and methods
2.1. Animals and surgery
All the experimental procedures described below were conductedin strict adherence to the recommendations found in the “Principles ofanimal care” (NIH, 1985) and were approved by the local Committeefor Ethics in Animal Research (CEUA-UFSC, protocols no. 372/CEUAand 23080.007968/2003-71). Adults Wistar rats (female, 198 to 228 gbody weight), maintained in individual cages at a temperature of 22–24 °C, on a 12:12 light–dark cycle (lights on at 06:00 h) and with freeaccess to food andwater were used throughout the experiments. After1 week of adaptation to the housing conditions, rats were anesthe-tized with ketamine hydrochloride (100 mg/kg, i.p.) and xylazine(20 mg/kg, i.p.), and were stereotaxically implanted with a unilateral
stainless steel guide cannula (30 G) aimed 2 mm dorsal to the ARC orthe LH, according to coordinates described by Paxinos and Watson[72]. The cannula, anchored to the skull with jeweler screws and fixedwith dental cement, was maintained patent between experiments byan inner removable stylet.
2.2. Drugs and injections
The 5-HT1A receptor agonist 8-Hydroxy-2-(di-n-propylamino)-tetra-lin (8-OH-DPAT, 0.6 and 6 nmol; Sigma Chemical Co., St. Louis,MO, USA)and the 5-HT1/2 receptor antagonist, metergoline (MET 2 and 20 nmol;Tocris Cookson Inc., EllisvilleMO,USA)were freshly dissolved in ascorbicacid 5% in distilled water, which was used as vehicle in controlexperiments. These doses were derived from previous studies on thebehavioral effects of local injections of 8-OH-DPAT [9,17,18,35,40] andMET [10,12,13,28,80] as effective in selectively influencing feedingbehavior in rats and pigeons. Injections were made through an innercannula (33 G) that extended 2mmbeyond the tip of the guide cannulaconnected by polyethylene tubing to a Hamilton micro syringe (1 µl)fitted to an injection pump. The injected volumes (0.2 µl) wereadministered over a period of 60 s and a further 60 s was allowed forthe solution to diffuse from the cannula.
2.3. Estrous cycle phase determination
Vaginal mucosal samples were obtained daily between 10:00–11:00 h starting in the third day after surgery. Vaginal secretion wascollected with a plastic pipette filled with 10 µl of normal saline (NaCl0.9%) by inserting the tip into the rat vagina. For each animal, vaginalfluid was placed on glass slides. The slides were observed unstainedunder a light microscope, with 10 and 40×objectives. The phase ofthe estrous cycle was assigned using standard, vaginal smears-basedand traditional criteria [32,61]. Vaginal mucosal samples wereperformed and examined independently by two observers tocharacterize the stage of ovarian cycle. As diestrus is preceded bylow plasma levels of estradiol and estrus is preceded by highsecretion of estradiol [32], we have chosen these two stages ofovarian cycle to carry out the present study. Another reason forhaving chosen these phases is that female rats eat less during estrusthan in diestrus [2,24–26]. At the experiment onset, all rats displayedregular estrous cycles of 4–5 days.
2.4. Experimental procedures
Seven days after surgery, the rats bearing LH- or ARC-directedcannuli were habituated to a wet mash enriched 10% sucrose [28],presented in clear glass Petri dishes (5 cm diameter) during 1 h for 3consecutive days at the recording chamber. Pelleted regular food wasoffered during the rest of the time. Water was provided in glassbottles, which were closed at the top, with a spout that was projectedthrough the cage wires. After this habituation period, enough for therats to reach a maximal and stable intake of a scheduled test meal ofsweet mash food, the animals were injected with vehicle (ascorbicacid 5%); 8-OH-DPAT (0.6 or 6 nmol) or MET (2 or 20 nmol). Eachanimal received only one treatment (drug dose or vehicle) and wasexamined in only one estrous cycle stage. Immediately after theinjection, the rats were placed in the recording chamber, and wereallowed for a 1 h period of access to a wet mash diet and water.During this period, the latency and duration of six behavioralcategories (eating, drinking, grooming, locomotion, rearing andresting) defined and used in a previous study [42] were continuouslyrecorded by a video camera perpendicularly located 60 cm above thecage floor for subsequent detailed behavioral analysis throughEtholog 2.2 [68]. The back and lateral walls as well as the floor cagewere coated with a black adhesive plastic paper. In order to facilitatebehavioral recording, the front wall of the test cage had a mirror with
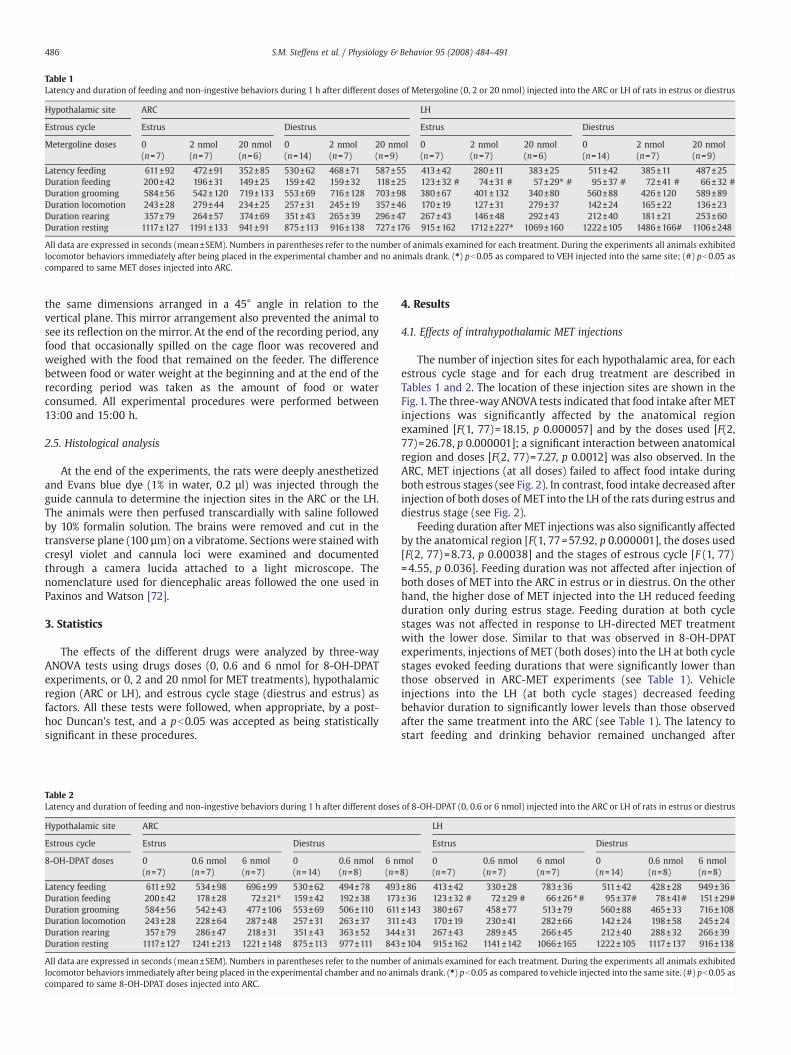
Table 1Latency and duration of feeding and non-ingestive behaviors during 1 h after different doses of Metergoline (0, 2 or 20 nmol) injected into the ARC or LH of rats in estrus or diestrus
Hypothalamic site ARC LH
Estrous cycle Estrus Diestrus Estrus Diestrus
Metergoline doses 0(n=7)
2 nmol(n=7)
20 nmol(n=6)
0(n=14)
2 nmol(n=7)
20 nmol(n=9)
0(n=7)
2 nmol(n=7)
20 nmol(n=6)
0(n=14)
2 nmol(n=7)
20 nmol(n=9)
Latency feeding 611±92 472±91 352±85 530±62 468±71 587±55 413±42 280±11 383±25 511±42 385±11 487±25Duration feeding 200±42 196±31 149±25 159±42 159±32 118±25 123±32 # 74±31 # 57±29⁎ # 95±37 # 72±41 # 66±32 #Duration grooming 584±56 542±120 719±133 553±69 716±128 703±98 380±67 401±132 340±80 560±88 426±120 589±89Duration locomotion 243±28 279±44 234±25 257±31 245±19 357±46 170±19 127±31 279±37 142±24 165±22 136±23Duration rearing 357±79 264±57 374±69 351±43 265±39 296±47 267±43 146±48 292±43 212±40 181±21 253±60Duration resting 1117±127 1191±133 941±91 875±113 916±138 727±176 915±162 1712±227⁎ 1069±160 1222±105 1486±166# 1106±248
All data are expressed in seconds (mean±SEM). Numbers in parentheses refer to the number of animals examined for each treatment. During the experiments all animals exhibitedlocomotor behaviors immediately after being placed in the experimental chamber and no animals drank. (⁎) pb0.05 as compared to VEH injected into the same site; (#) pb0.05 ascompared to same MET doses injected into ARC.
486 S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
the same dimensions arranged in a 45° angle in relation to thevertical plane. This mirror arrangement also prevented the animal tosee its reflection on the mirror. At the end of the recording period, anyfood that occasionally spilled on the cage floor was recovered andweighed with the food that remained on the feeder. The differencebetween food or water weight at the beginning and at the end of therecording period was taken as the amount of food or waterconsumed. All experimental procedures were performed between13:00 and 15:00 h.
2.5. Histological analysis
At the end of the experiments, the rats were deeply anesthetizedand Evans blue dye (1% in water, 0.2 µl) was injected through theguide cannula to determine the injection sites in the ARC or the LH.The animals were then perfused transcardially with saline followedby 10% formalin solution. The brains were removed and cut in thetransverse plane (100 µm) on a vibratome. Sections were stained withcresyl violet and cannula loci were examined and documentedthrough a camera lucida attached to a light microscope. Thenomenclature used for diencephalic areas followed the one used inPaxinos and Watson [72].
3. Statistics
The effects of the different drugs were analyzed by three-wayANOVA tests using drugs doses (0, 0.6 and 6 nmol for 8-OH-DPATexperiments, or 0, 2 and 20 nmol for MET treatments), hypothalamicregion (ARC or LH), and estrous cycle stage (diestrus and estrus) asfactors. All these tests were followed, when appropriate, by a post-hoc Duncan's test, and a pb0.05 was accepted as being statisticallysignificant in these procedures.
Table 2Latency and duration of feeding and non-ingestive behaviors during 1 h after different doses
Hypothalamic site ARC
Estrous cycle Estrus Diestrus
8-OH-DPAT doses 0(n=7)
0.6 nmol(n=7)
6 nmol(n=7)
0(n=14)
0.6 nmol(n=8)
6 n(n=
Latency feeding 611±92 534±98 696±99 530±62 494±78 493Duration feeding 200±42 178±28 72±21⁎ 159±42 192±38 173Duration grooming 584±56 542±43 477±106 553±69 506±110 611Duration locomotion 243±28 228±64 287±48 257±31 263±37 311Duration rearing 357±79 286±47 218±31 351±43 363±52 344Duration resting 1117±127 1241±213 1221±148 875±113 977±111 843
All data are expressed in seconds (mean±SEM). Numbers in parentheses refer to the numberlocomotor behaviors immediately after being placed in the experimental chamber and no ancompared to same 8-OH-DPAT doses injected into ARC.
4. Results
4.1. Effects of intrahypothalamic MET injections
The number of injection sites for each hypothalamic area, for eachestrous cycle stage and for each drug treatment are described inTables 1 and 2. The location of these injection sites are shown in theFig. 1. The three-way ANOVA tests indicated that food intake after METinjections was significantly affected by the anatomical regionexamined [F(1, 77)=18.15, p 0.000057] and by the doses used [F(2,77)=26.78, p 0.000001]; a significant interaction between anatomicalregion and doses [F(2, 77)=7.27, p 0.0012] was also observed. In theARC, MET injections (at all doses) failed to affect food intake duringboth estrous stages (see Fig. 2). In contrast, food intake decreased afterinjection of both doses of MET into the LH of the rats during estrus anddiestrus stage (see Fig. 2).
Feeding duration after MET injections was also significantly affectedby the anatomical region [F(1, 77=57.92, p 0.000001], the doses used[F(2, 77)=8.73, p 0.00038] and the stages of estrous cycle [F (1, 77)=4.55, p 0.036]. Feeding duration was not affected after injection ofboth doses of MET into the ARC in estrus or in diestrus. On the otherhand, the higher dose of MET injected into the LH reduced feedingduration only during estrus stage. Feeding duration at both cyclestages was not affected in response to LH-directed MET treatmentwith the lower dose. Similar to that was observed in 8-OH-DPATexperiments, injections of MET (both doses) into the LH at both cyclestages evoked feeding durations that were significantly lower thanthose observed in ARC-MET experiments (see Table 1). Vehicleinjections into the LH (at both cycle stages) decreased feedingbehavior duration to significantly lower levels than those observedafter the same treatment into the ARC (see Table 1). The latency tostart feeding and drinking behavior remained unchanged after
of 8-OH-DPAT (0, 0.6 or 6 nmol) injected into the ARC or LH of rats in estrus or diestrus
LH
Estrus Diestrus
mol8)
0(n=7)
0.6 nmol(n=7)
6 nmol(n=7)
0(n=14)
0.6 nmol(n=8)
6 nmol(n=8)
±86 413±42 330±28 783±36 511±42 428±28 949±36±36 123±32 # 72±29 # 66±26⁎# 95±37# 78±41# 151±29#±143 380±67 458±77 513±79 560±88 465±33 716±108±43 170±19 230±41 282±66 142±24 198±58 245±24±31 267±43 289±45 266±45 212±40 288±32 266±39±104 915±162 1141±142 1066±165 1222±105 1117±137 916±138
of animals examined for each treatment. During the experiments all animals exhibitedimals drank. (⁎) pb0.05 as compared to vehicle injected into the same site. (#) pb0.05 as
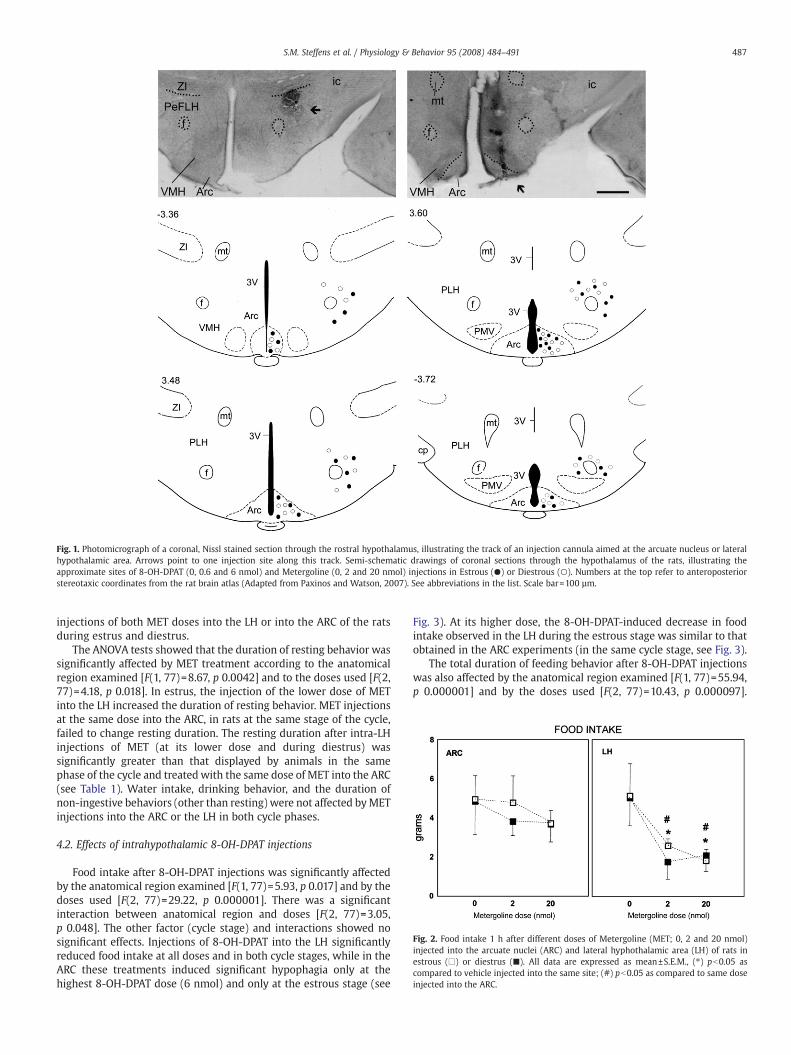
Fig. 1. Photomicrograph of a coronal, Nissl stained section through the rostral hypothalamus, illustrating the track of an injection cannula aimed at the arcuate nucleus or lateralhypothalamic area. Arrows point to one injection site along this track. Semi-schematic drawings of coronal sections through the hypothalamus of the rats, illustrating theapproximate sites of 8-OH-DPAT (0, 0.6 and 6 nmol) and Metergoline (0, 2 and 20 nmol) injections in Estrous (●) or Diestrous (○). Numbers at the top refer to anteroposteriorstereotaxic coordinates from the rat brain atlas (Adapted from Paxinos and Watson, 2007). See abbreviations in the list. Scale bar=100 µm.
Fig. 2. Food intake 1 h after different doses of Metergoline (MET; 0, 2 and 20 nmol)injected into the arcuate nuclei (ARC) and lateral hyphothalamic area (LH) of rats inestrous (□) or diestrus (■). All data are expressed as mean±S.E.M., (⁎) pb0.05 ascompared to vehicle injected into the same site; (#) pb0.05 as compared to same doseinjected into the ARC.
487S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
injections of both MET doses into the LH or into the ARC of the ratsduring estrus and diestrus.
The ANOVA tests showed that the duration of resting behavior wassignificantly affected by MET treatment according to the anatomicalregion examined [F(1, 77)=8.67, p 0.0042] and to the doses used [F(2,77)=4.18, p 0.018]. In estrus, the injection of the lower dose of METinto the LH increased the duration of resting behavior. MET injectionsat the same dose into the ARC, in rats at the same stage of the cycle,failed to change resting duration. The resting duration after intra-LHinjections of MET (at its lower dose and during diestrus) wassignificantly greater than that displayed by animals in the samephase of the cycle and treated with the same dose of MET into the ARC(see Table 1). Water intake, drinking behavior, and the duration ofnon-ingestive behaviors (other than resting) were not affected byMETinjections into the ARC or the LH in both cycle phases.
4.2. Effects of intrahypothalamic 8-OH-DPAT injections
Food intake after 8-OH-DPAT injections was significantly affectedby the anatomical region examined [F(1, 77)=5.93, p 0.017] and by thedoses used [F(2, 77)=29.22, p 0.000001]. There was a significantinteraction between anatomical region and doses [F(2, 77)=3.05,p 0.048]. The other factor (cycle stage) and interactions showed nosignificant effects. Injections of 8-OH-DPAT into the LH significantlyreduced food intake at all doses and in both cycle stages, while in theARC these treatments induced significant hypophagia only at thehighest 8-OH-DPAT dose (6 nmol) and only at the estrous stage (see
Fig. 3). At its higher dose, the 8-OH-DPAT-induced decrease in foodintake observed in the LH during the estrous stage was similar to thatobtained in the ARC experiments (in the same cycle stage, see Fig. 3).
The total duration of feeding behavior after 8-OH-DPAT injectionswas also affected by the anatomical region examined [F(1, 77)=55.94,p 0.000001] and by the doses used [F(2, 77)=10.43, p 0.000097].
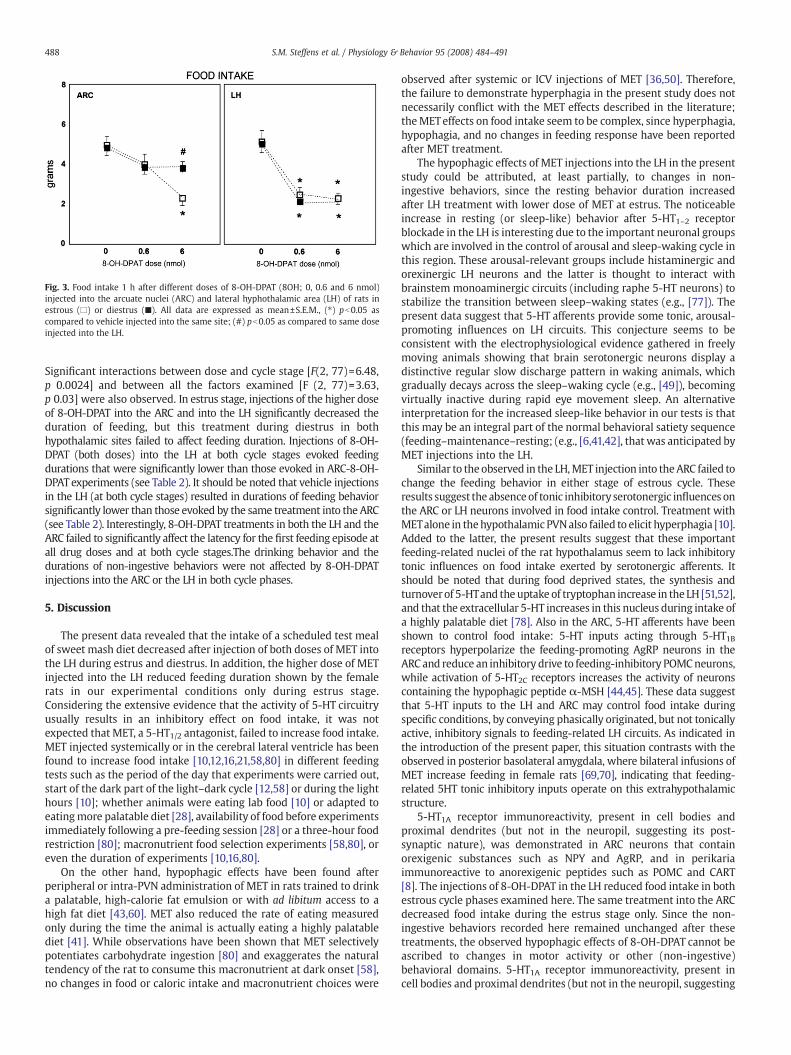
Fig. 3. Food intake 1 h after different doses of 8-OH-DPAT (8OH; 0, 0.6 and 6 nmol)injected into the arcuate nuclei (ARC) and lateral hyphothalamic area (LH) of rats inestrous (□) or diestrus (■). All data are expressed as mean±S.E.M., (⁎) pb0.05 ascompared to vehicle injected into the same site; (#) pb0.05 as compared to same doseinjected into the LH.
488 S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
Significant interactions between dose and cycle stage [F(2, 77)=6.48,p 0.0024] and between all the factors examined [F (2, 77)=3.63,p 0.03] were also observed. In estrus stage, injections of the higher doseof 8-OH-DPAT into the ARC and into the LH significantly decreased theduration of feeding, but this treatment during diestrus in bothhypothalamic sites failed to affect feeding duration. Injections of 8-OH-DPAT (both doses) into the LH at both cycle stages evoked feedingdurations that were significantly lower than those evoked in ARC-8-OH-DPAT experiments (see Table 2). It should be noted that vehicle injectionsin the LH (at both cycle stages) resulted in durations of feeding behaviorsignificantly lower than those evoked by the same treatment into the ARC(see Table 2). Interestingly, 8-OH-DPAT treatments in both the LH and theARC failed to significantly affect the latency for the first feeding episode atall drug doses and at both cycle stages.The drinking behavior and thedurations of non-ingestive behaviors were not affected by 8-OH-DPATinjections into the ARC or the LH in both cycle phases.
5. Discussion
The present data revealed that the intake of a scheduled test mealof sweet mash diet decreased after injection of both doses of MET intothe LH during estrus and diestrus. In addition, the higher dose of METinjected into the LH reduced feeding duration shown by the femalerats in our experimental conditions only during estrus stage.Considering the extensive evidence that the activity of 5-HT circuitryusually results in an inhibitory effect on food intake, it was notexpected that MET, a 5-HT1/2 antagonist, failed to increase food intake.MET injected systemically or in the cerebral lateral ventricle has beenfound to increase food intake [10,12,16,21,58,80] in different feedingtests such as the period of the day that experiments were carried out,start of the dark part of the light–dark cycle [12,58] or during the lighthours [10]; whether animals were eating lab food [10] or adapted toeatingmore palatable diet [28], availability of food before experimentsimmediately following a pre-feeding session [28] or a three-hour foodrestriction [80]; macronutrient food selection experiments [58,80], oreven the duration of experiments [10,16,80].
On the other hand, hypophagic effects have been found afterperipheral or intra-PVN administration of MET in rats trained to drinka palatable, high-calorie fat emulsion or with ad libitum access to ahigh fat diet [43,60]. MET also reduced the rate of eating measuredonly during the time the animal is actually eating a highly palatablediet [41]. While observations have been shown that MET selectivelypotentiates carbohydrate ingestion [80] and exaggerates the naturaltendency of the rat to consume this macronutrient at dark onset [58],no changes in food or caloric intake and macronutrient choices were
observed after systemic or ICV injections of MET [36,50]. Therefore,the failure to demonstrate hyperphagia in the present study does notnecessarily conflict with the MET effects described in the literature;theMETeffects on food intake seem to be complex, since hyperphagia,hypophagia, and no changes in feeding response have been reportedafter MET treatment.
The hypophagic effects of MET injections into the LH in the presentstudy could be attributed, at least partially, to changes in non-ingestive behaviors, since the resting behavior duration increasedafter LH treatment with lower dose of MET at estrus. The noticeableincrease in resting (or sleep-like) behavior after 5-HT1–2 receptorblockade in the LH is interesting due to the important neuronal groupswhich are involved in the control of arousal and sleep-waking cycle inthis region. These arousal-relevant groups include histaminergic andorexinergic LH neurons and the latter is thought to interact withbrainstem monoaminergic circuits (including raphe 5-HT neurons) tostabilize the transition between sleep–waking states (e.g., [77]). Thepresent data suggest that 5-HT afferents provide some tonic, arousal-promoting influences on LH circuits. This conjecture seems to beconsistent with the electrophysiological evidence gathered in freelymoving animals showing that brain serotonergic neurons display adistinctive regular slow discharge pattern in waking animals, whichgradually decays across the sleep–waking cycle (e.g., [49]), becomingvirtually inactive during rapid eye movement sleep. An alternativeinterpretation for the increased sleep-like behavior in our tests is thatthis may be an integral part of the normal behavioral satiety sequence(feeding–maintenance–resting; (e.g., [6,41,42], that was anticipated byMET injections into the LH.
Similar to the observed in the LH,MET injection into theARC failed tochange the feeding behavior in either stage of estrous cycle. Theseresults suggest the absenceof tonic inhibitory serotonergic influences onthe ARC or LH neurons involved in food intake control. Treatment withMETalone in thehypothalamic PVNalso failed to elicit hyperphagia [10].Added to the latter, the present results suggest that these importantfeeding-related nuclei of the rat hypothalamus seem to lack inhibitorytonic influences on food intake exerted by serotonergic afferents. Itshould be noted that during food deprived states, the synthesis andturnoverof 5-HTand the uptake of tryptophan increase in the LH [51,52],and that the extracellular 5-HT increases in this nucleus during intake ofa highly palatable diet [78]. Also in the ARC, 5-HT afferents have beenshown to control food intake: 5-HT inputs acting through 5-HT1Breceptors hyperpolarize the feeding-promoting AgRP neurons in theARC and reduce an inhibitory drive to feeding-inhibitory POMCneurons,while activation of 5-HT2C receptors increases the activity of neuronscontaining the hypophagic peptide α-MSH [44,45]. These data suggestthat 5-HT inputs to the LH and ARC may control food intake duringspecific conditions, by conveying phasically originated, but not tonicallyactive, inhibitory signals to feeding-related LH circuits. As indicated inthe introduction of the present paper, this situation contrasts with theobserved in posterior basolateral amygdala, where bilateral infusions ofMET increase feeding in female rats [69,70], indicating that feeding-related 5HT tonic inhibitory inputs operate on this extrahypothalamicstructure.
5-HT1A receptor immunoreactivity, present in cell bodies andproximal dendrites (but not in the neuropil, suggesting its post-synaptic nature), was demonstrated in ARC neurons that containorexigenic substances such as NPY and AgRP, and in perikariaimmunoreactive to anorexigenic peptides such as POMC and CART[8]. The injections of 8-OH-DPAT in the LH reduced food intake in bothestrous cycle phases examined here. The same treatment into the ARCdecreased food intake during the estrus stage only. Since the non-ingestive behaviors recorded here remained unchanged after thesetreatments, the observed hypophagic effects of 8-OH-DPAT cannot beascribed to changes in motor activity or other (non-ingestive)behavioral domains. 5-HT1A receptor immunoreactivity, present incell bodies and proximal dendrites (but not in the neuropil, suggesting

489S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
its postsynaptic nature), was demonstrated in ARC neurons thatcontain orexigenic substances such as NPY and AgRP, and in perikariaimmunoreactive to anorexigenic peptides such as POMC and CART [8].In addition, 5-HTmay be released at ARC sites where 5-HT1A receptorsare present, since serotonergic fibers were found in close associationwith cells immunoreactive to the 5-HT1A receptor, possibly acting onfeeding-related peptidergic mechanisms in this region [8,44,45].
Unlike the observed in the ARC, both doses of 8-OH-DPAT evokedhypophagic effects when injected in the LH. It has been shown thatextracellular 5-HT release decreased in the LH after peripheral injectionof 8-OH-DPAT in freely feeding rats without access to food after druginjection (during 120 min of experimental period). When food wasavailable to satiated rats during the experimental session, the feedingresponse elicited by 8-OH-DPAT induced an increase in the extracellular5-HT, compared to the group without access to food after 8-OH-DAPTinjection. The extracellular 5-HT was not affected by 8-OH-DPATtreatment in food-deprived animals with or without access to foodduring experimental period. In food-deprived animals with access tofood, 8-OH-DPATdecreased food intake and theunchangedextracellular5-HT released in this condition could be ascribed to low foodconsumption observed in this experiment [84]. These results suggestthat 5-HT inputs to the LH are active only when the intake actuallyoccurs. While these results confirm the presence of hypophagic 5-HTinputs to LH neurons, they also suggest that these inputs are related toepisodic signals originated by gastrointestinal processing of food.Interestingly, it has been reported that activation of 5-HT1B or 5-HT2Creceptors in LH failed to affect food intake [47], suggesting that thehypophagic effects of 5-HT inputs to LHmay bemediated by other 5-HTreceptor type. The present data suggest that 5-HT1A receptors couldmediate the 5-HT hypophagic effects in the LH. In the LH, 5-HT1Areceptor immunoreactivity was observed in cell bodies containing twoorexigenic peptides (MCH and orexin A), and 5-HT transporter-containing nerve fibers were demonstrated in association with 5-HT1Areceptor immunoreactive cell bodies [8]. The hypophagic responsesinduced by 8-OH-DPAT in the LH could be then mediated by thedecrease in the release of MCH and/or orexin.
Food intake evoked by 8-OH-DPAT treatment differs depending onnutritional state. Hypophagia in fasted- and hyperphagia in non fooddeprived-animals have been seen after treatmentwith 8-OH-DPAT [18–21,23]. The hyperphagia evoked by 8-OH-DPAT was attributed toactivation of 5-HT1A somatodentritic autoreceptor found in the medianand dorsal raphe, leading to an inhibition of 5-HT neuronal activity andreduced synthesis and release of 5-HT in forebrain terminal areas[9,12,29]. This proposed mechanism suggests the presence of tonicinhibitory serotonergic inputs from raphe neurons to the forebrainstructures preventing food intake. Since the treatmentwithMET did notelicit any hyperphagic responses, such a tonic inhibitory mechanism onfood intake is apparently absent of the ARC and LH.
Doses of 5-HT1A receptor agonists, which normally increasefeeding in non-deprived rats presented to their normal rat chow,can decrease food intake when animals are presented with palatablefood [22,82]. Corroborating these results, the treatment with 5-HTin the PVN decreased carbohydrate intake, but did not change bothprotein and fat intake. This 5-HT inhibitory effect on carbohydrateconsumption was blocked by 5-HT1A and 5-HT1B receptors antago-nists [63]. In addition, the injection of 8-OH-DPAT into the PVNdecreased carbohydrate intake [62]. Collectively, these data suggestthat activation of 5-HT1A receptors seems to reduce the carbohy-drate intake and that these receptors could be found on the PVN[62,64], ARC and LH (present results). In general, the effects ofserotonergic agents on food intake have been shown to varydepending on numerous factors, including palatability [17,41,80],macronutrient content [36,50,58,80], feeding schedule and period ofthe light/dark cycle [10,12,21,23,28]. Such a situation implies thatthe changes in feeding behavior observed in the present paper maybe not significant to all of the feeding-relevant contexts, being valid
for situations when a scheduled feeding regimen of a high palatablediet is present.
The current study also showed that ovarian hormones may affectthe emergence of the hypophagic effect elicited by 8-OH-DPATinjected into the ARC. In contrast, the 8-OH-DPAT-evoked hypophagiain the LH was not affected by ovarian steroids, since the magnitude ofthe reduction in the food intake in estrus was similar to that observedin diestrus. This result indicated that control of feeding by 5 HT1Areceptors of ARC could be influenced by gonadal steroid hormones.The estradiol seems to be the ovarian hormone involved in thechanges of responsiveness of neurons of ARC to 8-OH-DPAT, sincehyperphagia evoked by systemic injection of 8-OH-DPAT was affectedby estrous cycle stage, while treatment with estradiol in ovariecto-mized rats attenuated the hyperphagia induced by a 5-HT1A agonist[31,75,83]. In female monkeys, postsynaptic 5-HT1A receptors in thehypothalamus are downregulated by treatment with estrogen andestrogen combined with progesterone, but not by progesterone alone[63]. In addition, a cyclic regimen of estrogen treatment in ovariecto-mized rats increased the anorectic effect of fenfluramine [74]. Thehyperphagic response observed after ovariectomy is attributed to thedecline in circulating levels of estradiol, not progesterone, as estradiolreplacement alone returns to normal the food intake in the ovariecto-mized rats [33].
The responsiveness to serotonergic inhibitory influences on the foodintake seems tobemoreevidentduring theestrus than in theotherphasesof ovarian cycle. The infusion of MET into the posterior basolateralamygdala (pBLA) inducedamore intense increase in food intake in estrousrats, relative to diestrous rats. Consequently, the inhibitory 5-HT inputs onfood intake could bemore intense in the estrous phase of cyclewithin thepBLA [70]. In addition, it has been found that fenfluramine-inducedanorexiawas greater during estrus, relative to diestrus [27,74]. In contrast,thehyperphagia inducedby 5-HT1A agonistwasmore evident in diestrousfemales than in proestrous and estrous rats. [83]. As the hyperphagicresponse ismediated bypresynaptic 5-HT1A autoreceptor andhypophagicresponse is mediated by postsynaptic receptors, we could suggest thatestrogen affects differently pre- or postsynaptic 5-HT receptors. In thedorsal raphenucleus, the results presented in the literature are similar andsuggest that the estrogen reduces the function of the 5-HT1A somatoden-dritic receptor [55,63,65,73].
In the hypothalamus, postsynaptic 5-HT1A receptor function changesin response to estrogen in ovariectomized [3,4,29,30,37,61] or due to thefluctuations in 5HT levels during the regular cycle of female rats [38,39],thus altering the responsiveness of hypothalamic neurons to 8-OH-DPAT. It is possible that changes in 5-HT1A receptors binding sites in theARC [5,30], without affecting the 5-HT release in the synapse cleft, couldbe sufficient to allow for a 8-OH-DPAT-evoked hypophagic effect in theestrus stage. In agreementwith this suggestion, it has been reported thatestrous females showed relatively constant extracellular 5-HT in themediobasal hypothalamus between the light and the dark portions ofthe light/dark cycle [65]. Furthermore, normal density of serotonintransporter (SERT) sites in ARC did not change in both ovariectomizedand ovariectomized estrogen-treated female rats [66,81]. Also inovariectomized rats, the treatment with a combination of estradioland progesterone did not influence 5HT synthesis in the medianeminence or mediobasal hypothalamus [54]. Our data suggest thatovarian steroids hormones could increase the efficacy of satietyinfluences mediated by ARC 5-HT1A receptors activation, since thereduction in the feeding duration was observed only during the estrusstage. In line with this hypothesis, it has been reported that estradioldecreased meal size by increasing the potency of anorexigeniccompounds (e.g., cholecystokinin and serotonin) that signal mealtermination and by decreasing the strength of orexigenic compounds(e.g., melanin-concentrating hormone and ghrelin) that sustain a meal[7,26,34,67].
Although it has been reported that food intake decreased duringestrus relative to diestrus in normally cycling rats [2,24–26], this effect

490 S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
was not observed in the current study. Food intake exhibited byvehicle-treated rats in estrus was similar to that exhibited by vehicle-treated rats in diestrus. This discrepancy may be explained by thetraining of the animals to eat a sucrose rich diet in the middle of theday. It has been reported that animals tested during middle lightphase showed similar food consumption in estrus and diestrus [70].The typical low food intake during estrus has been observed duringdark hours of the light-dark cycle [2,24,25,27], when most of dailyfood is eaten [56]. The intense food intake observed in the light phaseof day in the present experiments could be due to the high palatabilityof diet. It must be noted that “estrus”, in the present paper, refers tothe phase of the cycle defined by the vaginal smears. The phase of thecycle was detected in the morning after the ovulation night, and theexperiments were conduced also in the light period, 13 to 15 h afterovulation. This period may be not fully representative of thebehavioral changes present at the initial moments of the behavioralestrous phase [3].
In conclusion, the data presented here revealed that tonicinhibitory serotonergic inputs do not operate on the ARC or LHneurons. Additionally, the current study demonstrated that 5-HT1Areceptors located in the ARC as well as in the LH are components of theserotonergic circuit controlling food intake. Both serotonergic circuitswere affected by ovarian steroids levels, possibly by fluctuations in theblood estrogen concentration, which could directly increase sensitiv-ity of ARC neurons to the hypophagic effects of 8-OH-DPAT or couldincrease the efficacy of satiety influences ARC or LH evoked by 5-HT1Areceptors activation.
Acknowledgements
The present work was supported by CNPq research grants to J.M.N.(proc. 471888/03-6) and to M.A.P. (proc. 474762/01-7).
References
[1] Arkle M, Ebenezer IS. Ipsapirone suppresses food intake in food-deprived rats byan action at 5-HT(1A) receptors. Eur J Pharmacol 2000;408:273–6.
[2] Asarian L, Geary N. Modulation of appetite by gonadal steroid hormones. PhilosTrans R Soc Lond B Biol Sci 2006;361:1251–63.
[3] Becker JB, Arnold AP, Berkley KJ, Blaustein JD, Eckel LA, Hampson E, et al. Strategiesand methods for research on sex differences in brain and behavior. Endocrinology2005;146:1650–73.
[4] Bethea CL, Lu NZ, Gundlah C, Streicher JM. Diverse Actions of Ovarian Steroids inthe Serotonin Neural System. Front Neuroendocrinol 2002;23:41–100.
[5] Biegon A, Fischette CT, Rainbow TC, McEwen BS. Serotonin receptor modulation byestrogen in discrete brain nuclei. Neuroendocrinology 1982;35:287–91.
[6] Blundell JE. Serotonin manipulations and the structure of feeding behavior.Appetite 1986;7(Suppl):39–56.
[7] Clegg DJ, Brown LM, Zigman JM, Kemp CJ, Strader AD, Benoit SC, et al. Estradiol-dependent decrease in the orexigenic potency of ghrelin in female rats. Diabetes2007;56:1051–8.
[8] Collin M, Bäckberg M, Önnestam K, Meister B. 5-HT1A receptor immunoreactivityin hypothalamic neurons involved in body weight control. Neuroreport2002;13:945–51.
[9] Coscina DV, Currie PJ, Bishop C, Parker GC, Rollins BL, King BM. Posterodorsalamygdala lesions reduce feeding stimulated by 8-OH-DPAT. Brain Res2000;883:243–9.
[10] Coscina DV, Feifel D, Nobrega JN, Currie PJ. Intraventricular but not intraparaven-tricular nucleus metergoline elicits feeding in satiated rats. Am J Physiol1994;266:1562–7.
[11] Currie PJ, Coscina DV. Diurnal variations in the feeding response to 8-OH-DPATinjected into the dorsal or median raphe. Neuroreport 1993;4:1105–7.
[12] Currie PJ, Coscina DV. Metergoline potentiates natural feeding and antagonizes theanorectic action of medial hypothalamic 5-hydroxytryptamine. PharmacolBiochem Behav 1996;53:1023–8.
[13] Da Silva RA, Da Silva AS, Poffo MJ, Ribas DC, Faria MS, Marino-Neto J, et al. Feedingbehavior aftermetergoline or GR-46611 injections into the paraventricular nucleusof the hypothalamus in the pigeon. Behav Brain Res 2007;179:248–57.
[14] De Vry J, Schreiber R. Effects of selected serotonin 5-HT(1) and 5-HT(2) receptoragonists on feeding behavior: possible mechanisms of action. Neurosci BiobehavRev 2000;24:341–53.
[15] Dourish CT. Multiple serotonin receptors: opportunities for new treatments forobesity? Obes Res 1995(Suppl 4):449–62.
[16] Dourish CT, Clark ML, Fletcher A, Iversen SD. Evidence that blockade of post-synaptic5-HT1 receptors elicits feeding in satiated rats. Psychopharmacology 1989;97:54–8.
[17] Dourish CT, Cooper SJ, Gilbert F, Coughlan J, Iversen SD. The 5-HT1A agonist 8-OH-DPAT increases consumption of palatable wet mash and liquid diets in the rat.Psychopharmacology 1988;94:58–63.
[18] Dourish CT, Hutson PH, Curzon G. Characteristics of feeding induced by theserotonin agonist 8-hydroxy-2-(di-n-propylamino) tetralin (8-OH-DPAT). BrainRes Bull 1985;15:377–84.
[19] Ebenezer IS, Arkle MJ, Tite RM. 8-Hydroxy-2-(di-n-propylamino)-tetralin inhibitsfood intake in fasted rats by an action at 5-HT1A receptors. Methods Find Exp ClinPharmacol 2007;29:269–72.
[20] Ebenezer IS, Parrott RF, Vellucci SV. Effects of the 5-HT1A receptor agonist 8-OH-DPAT on operant food intake in food-deprived pigs. Physiol Behav 1999;67:213–7.
[21] Ebenezer IS, Surujbally A. The effects of 8-hydroxy-2-(di-n-propylamino)-tetralin(8-OH-DPAT) on food intake in non-deprived C57BL6 mice. Eur J Pharmacol2007;559:184–8.
[22] Ebenezer IS, Tite RM. Differential effects of the 5-HT1A agonist gepirone on foodintake in rats may depend on their motivation to eat. Methods Find Exp ClinPharmacol 2003;25:727–31.
[23] Ebenezer IS, Vellucci SV, Parrott RF. The differential effects of intravenouslyadministered 8-OH-DPAT on operant food intake in satiated and food-deprivedpigs are mediated by central 5-HT(1A) receptors. Physiol Behav 2001;73:223–7.
[24] Eckel LA, Geary N. Endogenous cholecystokinin's satiating action increases duringestrus in female rats. Peptides 1999;20:451–6.
[25] Eckel LA, Houpt TA, Geary N. Spontaneous meal patterns in female rats with andwithout access to running wheels. Physiol Behav 2000;70:397–405.
[26] Eckel LA, Moore SR. Diet-induced hyperphagia in the rat is influenced by sex andexercise. Am J Physiol Regul Integr Comp Physiol 2004;287:1080–5.
[27] Eckel LA, Rivera HM, Atchley DP. The anorectic effect of fenfluramine is influencedby sex and stage of the estrous cycle in rats. Am J Physiol Regul Integr Comp Physiol2005;288:1486–91.
[28] Fletcher PJ. Increased food intake in satiated rats induced by the 5-HT antagonistsmethysergide, metergoline and ritanserin. Psychopharmacology 1988;96:237–42.
[29] Fletcher PJ, Coscina DV. Injecting 5-HT into the PVN does not prevent feedinginduced by injecting 8-OH-DPAT into the raphe. Pharmacol Biochem Behav1993;46:487–91.
[30] Flügge G, Pfender D, Rudolph S, Jarry H, Fuchs E. 5HT1A-receptor binding in thebrain of cyclic and ovariectomized female rats. J Neuroendocrinol 1999;11:243–9.
[31] Frankfurt M, McKittrick CR, Mendelson SD, McEwen BS. Effect of 5,7- dihydrox-ytryptamine, ovariectomy and gonadal steroids on serotonin receptor binding inrat brain. Neuroendocrinology 1994;59:245–50.
[32] FreemanME. The neuroendocrine control of the ovarian cycle of the rat. In: Knobil,E, Neil, JD, editors. The physiology of reproduction, 3th ed, vol. 2. New York:Academic Press & Elsevier; 2006. p. 2327–88.
[33] Geary N, Asarian L. Cyclic estradiol treatment normalizes body weight and testmeal size in ovariectomized rats. Physiol Behav 1999;67:141–7.
[34] Geary N, Trace D, McEwen B, Smith GP. Cyclic estradiol replacement increases thesatiety effect of CCK-8 in ovariectomized rats. Physiol Behav 1994;56:281–9.
[35] Gilbert F, Dourish CT. Effects of the novel anxiolytics gepirone, buspirone andipsapirone on free feeding and on feeding induced by 8-OH-DPAT. Psychophar-macology 1987;93:349–52.
[36] Grignaschi G, Fanelli E, Scagnol I, Samanin R. Studies on the role of serotoninreceptor subtypes in the effect of sibutramine in various feeding paradigms in rats.Br J Pharmacol 1999;127:1190–4.
[37] Grignaschi G, Sironi F, Samanin R. The 5-HT1B receptor mediates the effect of D-fenfluramine on eating caused by intra-hypothalamic injection of neuropeptide Y.Eur J Pharmacol 1995;274:221–2.
[38] Gundlah C, Pecins-Thompson M, Schutzer WE, Bethea CL. Ovarian steroid effectson serotonin 1A, 2A and 2C receptor mRNA in macaque hypothalamus. Mol BrainRes 1999;63:325–39.
[39] Gundlah C, Simon LD, Auerbach SB. Differences in hypothalamic serotoninbetween estrous phases and gender: an in vivo microdialysis study. Brain Res1998;785:91–6.
[40] Häckl LP, de Oliveira Richter G, Faria MS, Paschoalini MA, Marino-Neto J.Behavioral effects of 8-OH-DPAT injections into pontine and mesencephalicareas containing 5-HT-immunoreactive perikarya in the pigeon. Brain Res2005;1035:154–67.
[41] Halford JC, Blundell JE. The 5-HT1B receptor agonist CP-94,253 reduces food intakeand preserves the behavioural satiety sequence. Physiol Behav 1996;60:933–9.
[42] Haldford JC,Wanninayake SCD, Blundell JE. Behavioral Satiety Sequence (BSS) for theDiagnosis ofDrugAction onFood Intake. Pharmacol BiochemBehav 1998;61:159–68.
[43] Hartfield AW, Moore NA, Clifton PG. Serotonergic and histaminergic mechanismsinvolved in intralipid drinking? Pharmacol. Biochem Behav 2003;76:251–8.
[44] Heisler LK, Cowley MA, Tecott LH, Fan W, Low MJ, Smart JL, et al. Activation ofcentral melanocortin pathways by fenfluramine. Science 2002;297:609–11.
[45] Heisler LK, Jobst EE, Sutton GM, Zhou L, Borok E, Thornton-Jones Z, et al. SerotoninReciprocally Regulates Melanocortin Neurons to Modulate Food Intake. Neuron2006;51:239–49.
[46] Hewitt KN, Lee MD, Dourish CT, Clifton PG. Serotonin 2C receptor agonistsand the behavioural satiety sequence in mice. Pharmacol Biochem Behav2000;71:691–700.
[47] Hikiji K, Inoue K, Iwasaki S, Ichihara K, Kiriike N. Local perfusion of mCPP intoventromedial hypothalamic nucleus, but not into lateral hypothalamic area andfrontal cortex, inhibits food intake in rats. Psychopharmacology 2004;174:190–6.
[48] Hutson PH, Dourish CT, Curzon G. Neurochemical and behavioural evidence formediation of the hyperphagic action of 8-OH-DPAT by 5-HT cell bodyautoreceptors. Eur J Pharmacol 1986;129:347–52.

491S.M. Steffens et al. / Physiology & Behavior 95 (2008) 484–491
[49] Jacobs BL, Fornal CA. Activity of serotonergic neurons in behaving animals.Neuropsychopharmacology 1999;21:9S–15S.
[50] Jourdan D, Piec I, Gaulier JM, Lacassie E, Alliot J. Effect of fenfluramine on caloricintake and macronutrient selection in Lou/c rats during aging. Neurobiol Aging2003;24:67–76.
[51] Kantak KM, Wayner MJ, Stein JM, Tilson HA. Effect of imipramine on serotoninturnover in the lateral hypothalamus. Pharmacol Biochem Behav 1978;9:693–6.
[52] Kantak KM, Wayner MJ, Tilson HA, Sved A. Turnover of 3H-5-hydroxytryptamineto 3H-5-hydroxyindoleacetic acid and the 3H-5-methoxyindoles in nondeprivedand 24 hr food deprived rats. Pharmacol Biochem Behav 1977;6:221–5.
[53] Kennett GA, Curzon G. Evidence that hypophagia induced by mCPP and TFMPPrequires 5-HT1C and 5-HT1B receptors, hypophagia induced by RU 24969 onlyrequires 5-HT1B receptors. Psychopharmacology 1988;96:93–100.
[54] King TS, Steger RW, Morgan WW. Effect of ovarian steroids to stimulate region-specific hypothalamic 5-hydroxytryptamine synthesis in ovariectomized rats.Neuroendocrinology 1986;42:344–50.
[55] Lakoski JM, Cunningham KA. The interaction of cocaine with central serotonergicneuronal systems: cellular electrophysiologic approaches. NIDA Res Monogr1988;88:78–91.
[56] Laviano A, Meguid MM, Gleason JR, Yang ZJ, Renvyle T. Comparison of long-termfeeding pattern between male and female Fischer 344 rats: influence of estrouscycle. Am J Physiol 1996;270:413–9.
[57] Leibowitz SF, Alexander JT. Hypothalamic Serotonin in Control of Eating Behavior,Meal Size, and Body Weight. Biol Psychiatry 1988;44:851–64.
[58] Leibowitz SF, Alexander JT, Cheung WK, Weiss GF. Effects of serotonin and theserotonin blocker metergoline on meal patterns and macronutrient selection.Pharmacol Biochem Behav 1993;45:185–94.
[59] Leibowitz SF, Weiss GF, Suh JS. Medial hypothalamic nuclei mediate serotonin'sinhibitory effect on feeding behavior. Pharmacol Biochem Behav 1990;37:735–42.
[60] Lin L, York DA. 5-HT1B receptors modulate the feeding inhibitory effects ofenterostatin. Brain Res 2005;1062:26–31.
[61] Long JA, Evans HM. The oestrous cycle in the rat and its associated phenomenon.In: Leuschner AO, editor. Memoirs of the University of California. Oxford'University of California Press; 1922. p. 1–148.
[62] López-Alonso VE, Mancilla-Díaz JM, Rito-Domingo M, González-Hernández B,Escartín-Pérez RE. The effects of 5-HT1A and 5-HT2C receptor agonists onbehavioral satiety sequence in rats. Neurosci Lett 2007;416:285–8.
[63] Lu NZ, Bethea CL. Ovarian steroid regulation of 5-HT1A receptor binding and Gprotein activation in femalemonkeys. Neuropsychopharmacology 2002;27:12–24.
[64] Mancilla-Diaz JM, Escartin-Perez RE, Lopez-Alonso VE, Floran-Garduño B,Romano-Camacho J. Role of 5-HT1A and 5-HT1B receptors in the hypophagiceffect of 5-HT on the structure of feeding behavior. Med Sci Monit 2005;11:74–9.
[65] Maswood S, Stewart G, Uphouse L. Gender and estrous cycle effects of the 5-HT1Aagonist (8-OH-DPAT) on hypothalamic serotonin. Pharmacol Biochem Behav1995;51:807–13.
[66] McQueen JK, Wilson H, Sumner BE, Fink G. Serotonin transporter (SERT) mRNAand binding site densities in male rat brain affected by sex steroids. Brain Res MolBrain Res 1999;63:241–7.
[67] Messina MM, Boersma G, Overton JM, Eckel LA. Estradiol decreases the orexigeniceffect of melanin-concentrating hormone in ovariectomized rats. Physiol Behav2006;88:523–8.
[68] Ottoni EB. EthoLog 2.2: a tool for the transcription and timing of behaviourobservation sessions. Behav Res Meth Instrum Comput 2000;32:446–9.
[69] Parker GC, Balboul R, Hobday JA, Coscina DV. 5-HT receptor blockade in theposterior amygdala elicits feeding in female rats. NeuroReport 2001;12:911–4.
[70] Parker GC, Bishop C, Coscina DV. Estrous cycle and food availability affect feedinginduced by amygdale 5-HT receptor blockade. Pharmacol Biochem Behav2002;71:701–7.
[71] Parker GC, Coscina DV. Lesions of the posterior basolateral amygdala block feedinginduced by systemic 8-OH-DPAT. Pharmacol Biochem, Behav 2001;68:729–34.
[72] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 6th ed. New York:Academic Press & Elsevier Inc.; 2007.
[73] Pecins-Thompson M, Bethea CL. Ovarian steroid regulation of serotonin-1Aautoreceptor messenger RNA expression in the dorsal raphe of rhesus macaques.Neuroscience 1999;89:267–77.
[74] Rivera HM, Eckel LA. The anorectic effect of fenfluramine is increased by estradioltreatment in ovariectomized rats. Physiol Behav 2005;86:331–7.
[75] Salamanca S, Uphouse L. Estradiol modulation of the hyperphagia induced by the5-HT1A agonist, 8-OH-DPAT. Pharmacol Biochem Behav 1992;43:953–5.
[76] Samanin R, Mennini T, Bendotti C, Barone D, Caccia S, Garattini S. Evidence thatcentral 5-HT2 receptors do not play an important role in the anorectic activity ofD-fenfluramine in the rat. Neuropharmacology 1989;28:465–9.
[77] Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadianrhythms. Nature 2005;437:1257–63.
[78] Schwartz DH, McClane S, Hernandez L, Hoebel BG. Feeding increases extracellularserotonin in the lateral hypothalamus of the rat as measured by microdialysis.Brain Res 1989;479:349–54.
[79] Simansky KJ. Serotonergic control of the organization of feeding and satiety. BehavBrain Res 1996;73:37–42.
[80] Stallone D, Nicolaidis S. Increased food intake and carbohydrate preference in therat following treatment with the serotonin antagonist metergoline. Neurosci Lett1989;102:319–24.
[81] Sumner BE, Grant KE, Rosie R, Hegele-Hartung C, Fritzemeier KH, Fink G. Effects oftamoxifen on serotonin transporter and 5-hydroxytryptamine(2A) receptorbinding sites and mRNA levels in the brain of ovariectomized rats with or withoutacute estradiol replacement. Brain Res Mol Brain Res 1999;73:119–28.
[82] Swiergiel AH, Dunn AJ. Lack of evidence for a role of serotonin in interleukin-1-induced hypophagia. Pharmacol Biochem Behav 2000;65:531–7.
[83] Uphouse L, Salamanca S, Caldarola-Pastuszka M. Estrous cycle differences in theresponse to the 5-HT1A agonist 8-OH-DPAT. Pharmacol Biochem Behav1991;40:901–6.
[84] Voigt JP, Kienzle F, Sohr R. Feeding and 8-OH-DPAT-related release of serotonin inthe rat lateral hypothalamus. Pharmacol Biochem Behav 2000;65:183–9.
Copyright © 2022 FDOKUMEN







![Medial septal [beta]-amyloid 1-40 injections alter septo-hippocampal anatomy and function](https://static.fdokumen.com/doc/165x107/6329e172e9556f820801538c/medial-septal-beta-amyloid-1-40-injections-alter-septo-hippocampal-anatomy-and.jpg)












