
The effect of introducing purified β-glucans to a wheat-based diet on total tract digestibility and...
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of The effect of introducing purified β-glucans to a wheat-based diet on total tract digestibility and...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Animal Feed Science and Technology 165 (2011) 95–104
Contents lists available at ScienceDirect
Animal Feed Science and Technology
journal homepage: www.elsevier.com/locate/anifeedsci
The effect of introducing purified �-glucans to a wheat-based diet ontotal tract digestibility and gaseous manure emissions from pigs ascompared with consumption of a �-glucan-rich, barley-based diet
C.J. O’Shea, T. Sweeney, M.B. Lynch, D.A. Gahan, B.A. Flynn, J.V. O’Doherty ∗
University College Dublin, School of Agriculture, Food Science and Veterinary Medicine, Lyons Research Farm Newcastle, Co. Dublin, Ireland
a r t i c l e i n f o
Article history:Received 9 April 2010Received in revised form 11 February 2011Accepted 17 February 2011
Keywords:�-GlucansAmmoniaBarleyOdourPigWheat
a b s t r a c t
Consumption of barley-based diets has been demonstrated to reduce manure odour andammonia emissions generated from pig production, but is associated with depressed nutri-ent digestibility when compared with wheat-based formulations. It was hypothesised thatsupplementation of a wheat-based diet with purified mixed-linkage � (1,3) (1,4)-D-glucan(�-glucans) to levels comparable with a barley-based diet may beneficially amelioratemanure odour and ammonia emissions, without depressing nutrient digestibility. A com-plete randomized design experiment was conducted to investigate the effect of dietary�-glucans source and an enzyme composite containing �-glucanase and �-xylanase onenergy and nutrient digestibility, distal gastrointestinal tract (dGIT) fermentation, manureodour and ammonia emissions from finishing boars. Twenty-eight boars (74.2 kg SD 3.6)were assigned to one of four dietary treatments (n = 7); (1) a barley-based diet (B), (2)a barley-based diet + enzyme composite (BE), (3) a wheat-based diet + purified �-glucans(WG) and (4) a wheat-based diet + purified �-glucans + enzyme composite (WGE). Thewheat-based diets containing purified �-glucans were formulated to contain concentra-tions of total �-glucans comparable with the barley-based diets. Consumption of the WGdiet increased the coefficient of total tract apparent digestibility (CTTAD) of gross energy(P = 0.001) and N (P = 0.001) compared with the B diet. Pigs offered the WG diet hadincreased manure ammonia emissions (P = 0.042) compared with the B diet. There was nosignificant difference between consumption of the WG diet and the B diet on subsequentmanure odour emissions.
In summary, the current study demonstrates that �-glucans may be added to a wheat-based diet without depressing nutrient digestibility as compared with a barley-based diet.Furthermore, manure odour from pigs offered a wheat-based diet containing �-glucansmay be improved to levels comparable with a barley-based diet. However �-glucan sup-plementation was ineffectual at attenuating manure ammonia emissions comparable withconsumption of a barley-based diet.
© 2011 Elsevier B.V. All rights reserved.
Abbreviations: � (1,3) (1,4)-D-glucan, �-glucans; ADF, acid detergent fibre; ADL, acid detergent lignin; B, barley-based diet; BE, barley-baseddiet+enzyme composite; cfu, colony forming unit; CTTAD, Coefficient of total tract apparent digestibility; dGIT, distal gastrointestinal tract; DM, drymatter; GE, gross energy; N, nitrogen; NDF, neutral detergent fibre; OM, organic matter; WG, wheat-based diet+�-glucans; WGE, wheat-based diets+�-glucans+enzyme composite; VFA, volatile fatty acids.
∗ Corresponding author. Tel.: +353 1 7167128; fax: +353 1 7161103.E-mail address: [email protected] (J.V. O’Doherty).
0377-8401/$ – see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.anifeedsci.2011.02.012

Author's personal copy
96 C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104
1. Introduction
The contribution of intensive pig production to manure malodour and ammonia emissions is caused in part throughoversupply of dietary protein (Hayes et al., 2004). Excessive protein consumption causes both urinary nitrogen (N) excretionas urea, and the conveying of undigested proteinaceous material to the distal gastrointestinal tract (dGIT) (Carpenter et al.,2004; Lynch et al., 2007a). The bacterial degradation of such nitrogenous compounds generates ammonia and pungentvolatile fatty acids which contribute to subsequent manure emissions (Mackie et al., 1998; O’Shea et al., 2009a).
Previous research from this laboratory has indicated that such environmental pollutants may be ameliorated by substi-tuting barley for wheat-based diets (Garry et al., 2007b; Lynch et al., 2007b). The beneficial influence associated with barleyhas been attributed to an increased dietary consumption of mixed-linkage � (1,3) (1,4)-D-glucan (�-glucans), a fermentablesubstrate which may stimulate a more commensal dGIT microbiota, as indicated by marker bacteria such as lactobacilliand bifidobacteria (O’Connell et al., 2005; Garry et al., 2007a). An increase in the prominence of these bacterial genera mayinduce a shift towards greater saccharolytic activity, and thus a corresponding decrease in protein end-products in the dGIT,with beneficial implications for subsequent manure odour and ammonia emissions (O’Shea et al., 2009b, 2010).
However in monogastrics, dietary barley is associated with an impaired coefficient of total tract apparent digestibility(CTTAD) (O’Connell et al., 2006; Lynch et al., 2007c). Therefore it is desirable to impart the beneficial properties of �-glucans in barley to dietary wheat without the associated implications of diminished nutrient utilisation. The addition ofa purified source of cereal �-glucans to a conventional wheat-based diet may stimulate dGIT and manure fermentation toreduce concentrations of protein metabolites associated with manure ammonia and odour emissions. To verify this, dietarysupplementation with an appropriate enzyme composite containing �-glucanase may elucidate whether induced intestinalparameters are a response to exogenous �-glucan supplementation or an additional fermentable component present innative barley (O’Connell et al., 2006).
Therefore the hypothesis of the current experiment was that dietary supplementation of a wheat-based diet with anexogenous, purified �-glucan source would decrease manure odour and ammonia emissions from finishing pigs comparablewith a barley-based diet.
2. Materials and methods
All procedures described in this experiment were conducted under experimental licence from the Irish Department ofHealth in accordance with the Cruelty to Animals Act 1876 and the European Communities (Amendments of the Cruelty toAnimals Act 1876) Regulations, 1994.
2.1. Experimental design and dietary treatments
The experiment was devised as a complete randomised block design comprising four dietary treatments as follows: (1)barley-based diet (B), (2) barley-based diet + enzyme composite (BE), (3) wheat-based diet + purified �-glucan (WG) and(4) wheat-based diet + purified �-glucan + enzyme composite (WGE). All experimental diets were formulated to be isoen-ergetic and isonitrogenous, containing similar concentrations of analysed digestible energy (DE; 13.9 MJ/kg) and calculatedapparent ileal digestible lysine content (8.6 g/kg) (INRA-AFZ, 2004) and offered in meal form. All additional ileal digestibleamino acid requirements were met relative to lysine (NRC, 1998). Wheat-based dietary treatments were supplementedwith purified mixed-linkage � (1,3) (1,4)-D-glucan obtained from oats (Cambridge Commodities Ltd., Cambridgeshire, UK)comparable with concentrations of total �-glucans typically observed in barley-based diets. The enzyme supplement wasderived from Penicillium funiculosum (IMI SD 101) and contained Endo-1, 3 (4)-�-glucanase (EC 3.2.1.6) and Endo-1, 4-�-xylanase (EC 3.2.1.8) (DSM, Belfast, Northern Ireland). Xylanase was included in the enzyme composite to expose �-glucanto the hydrolytic activity of the �-glucanase enzyme. In barley, the aleurone cell rich in �-glucans is encapsulated by xylans(Autio et al., 1996). The dietary composition and analysis are presented in Table 1.
2.2. Animals and management
Twenty-eight finishing boars (7 replicates/dietary treatment) (progeny of Meat line boars × (Large White × Landracesows)) were blocked based on body-weight and assigned to one of four dietary treatments. Pigs were individually offeredad libitum access to feed and water and remained on their respective experimental diets until slaughter. At cessation of a14-day dietary adaption period, sixteen boars (4 replicates/dietary treatment) of a uniform body-weight (74.2 kg SD 3.6)were selected and transferred to individual metabolism crates. The pigs were allocated a further five days to adapt to themetabolism crates before collections commenced. The collection period was sub-divided into three parts to facilitate studieson manure ammonia emissions (d 1–2), manure odour emissions (d 3–5) and CTTAD and nitrogen (N) balance (d 6–10). Thedaily feed ration was offered over two meals. Fresh water was provided with meals in a 1:1 ratio and between meals wasprovided ad libitum. Feed consumed and refused was measured and recorded. The metabolism crates were located in atemperature-controlled room maintained at a constant temperature of 22 ◦C (±1.5 ◦C). Upon cessation of the collectionsfrom the metabolism crates, the pigs were reunited with their pen-mates until slaughter.
Author's personal copy
C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104 97
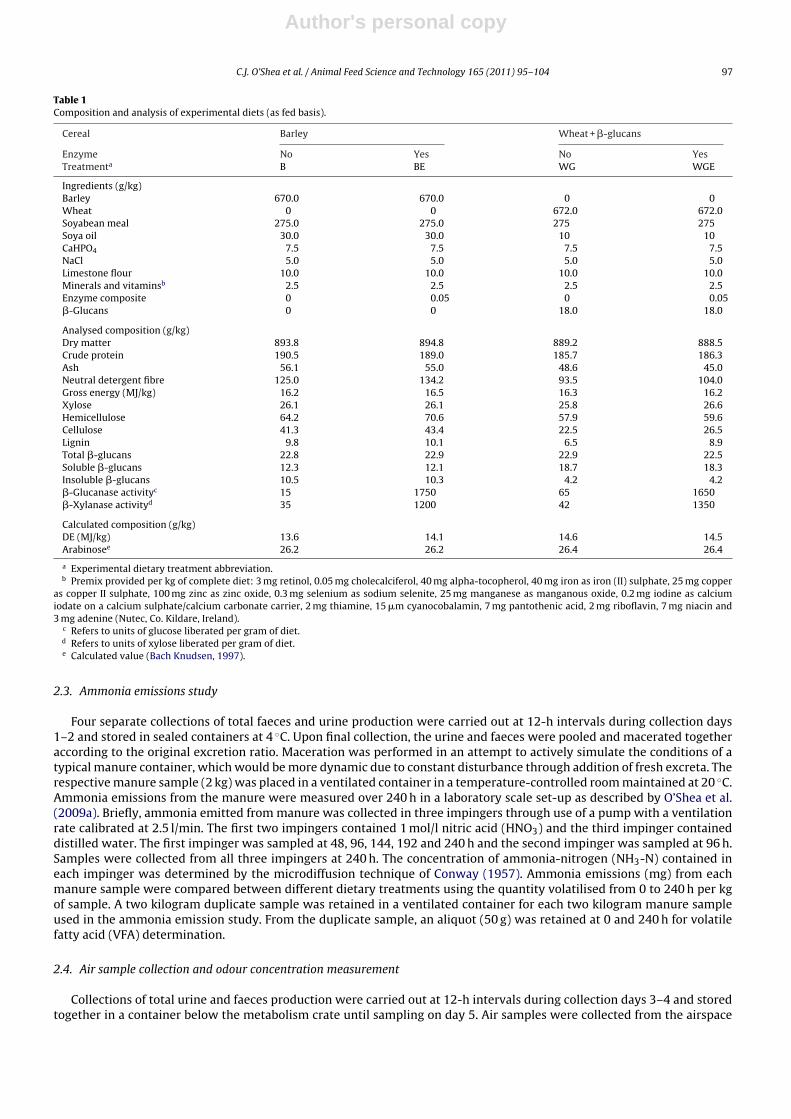
Table 1Composition and analysis of experimental diets (as fed basis).
Cereal Barley Wheat + �-glucans
Enzyme No Yes No YesTreatmenta B BE WG WGE
Ingredients (g/kg)Barley 670.0 670.0 0 0Wheat 0 0 672.0 672.0Soyabean meal 275.0 275.0 275 275Soya oil 30.0 30.0 10 10CaHPO4 7.5 7.5 7.5 7.5NaCl 5.0 5.0 5.0 5.0Limestone flour 10.0 10.0 10.0 10.0Minerals and vitaminsb 2.5 2.5 2.5 2.5Enzyme composite 0 0.05 0 0.05�-Glucans 0 0 18.0 18.0
Analysed composition (g/kg)Dry matter 893.8 894.8 889.2 888.5Crude protein 190.5 189.0 185.7 186.3Ash 56.1 55.0 48.6 45.0Neutral detergent fibre 125.0 134.2 93.5 104.0Gross energy (MJ/kg) 16.2 16.5 16.3 16.2Xylose 26.1 26.1 25.8 26.6Hemicellulose 64.2 70.6 57.9 59.6Cellulose 41.3 43.4 22.5 26.5Lignin 9.8 10.1 6.5 8.9Total �-glucans 22.8 22.9 22.9 22.5Soluble �-glucans 12.3 12.1 18.7 18.3Insoluble �-glucans 10.5 10.3 4.2 4.2�-Glucanase activityc 15 1750 65 1650�-Xylanase activityd 35 1200 42 1350
Calculated composition (g/kg)DE (MJ/kg) 13.6 14.1 14.6 14.5Arabinosee 26.2 26.2 26.4 26.4
a Experimental dietary treatment abbreviation.b Premix provided per kg of complete diet: 3 mg retinol, 0.05 mg cholecalciferol, 40 mg alpha-tocopherol, 40 mg iron as iron (II) sulphate, 25 mg copper
as copper II sulphate, 100 mg zinc as zinc oxide, 0.3 mg selenium as sodium selenite, 25 mg manganese as manganous oxide, 0.2 mg iodine as calciumiodate on a calcium sulphate/calcium carbonate carrier, 2 mg thiamine, 15 �m cyanocobalamin, 7 mg pantothenic acid, 2 mg riboflavin, 7 mg niacin and3 mg adenine (Nutec, Co. Kildare, Ireland).
c Refers to units of glucose liberated per gram of diet.d Refers to units of xylose liberated per gram of diet.e Calculated value (Bach Knudsen, 1997).
2.3. Ammonia emissions study
Four separate collections of total faeces and urine production were carried out at 12-h intervals during collection days1–2 and stored in sealed containers at 4 ◦C. Upon final collection, the urine and faeces were pooled and macerated togetheraccording to the original excretion ratio. Maceration was performed in an attempt to actively simulate the conditions of atypical manure container, which would be more dynamic due to constant disturbance through addition of fresh excreta. Therespective manure sample (2 kg) was placed in a ventilated container in a temperature-controlled room maintained at 20 ◦C.Ammonia emissions from the manure were measured over 240 h in a laboratory scale set-up as described by O’Shea et al.(2009a). Briefly, ammonia emitted from manure was collected in three impingers through use of a pump with a ventilationrate calibrated at 2.5 l/min. The first two impingers contained 1 mol/l nitric acid (HNO3) and the third impinger containeddistilled water. The first impinger was sampled at 48, 96, 144, 192 and 240 h and the second impinger was sampled at 96 h.Samples were collected from all three impingers at 240 h. The concentration of ammonia-nitrogen (NH3-N) contained ineach impinger was determined by the microdiffusion technique of Conway (1957). Ammonia emissions (mg) from eachmanure sample were compared between different dietary treatments using the quantity volatilised from 0 to 240 h per kgof sample. A two kilogram duplicate sample was retained in a ventilated container for each two kilogram manure sampleused in the ammonia emission study. From the duplicate sample, an aliquot (50 g) was retained at 0 and 240 h for volatilefatty acid (VFA) determination.
2.4. Air sample collection and odour concentration measurement
Collections of total urine and faeces production were carried out at 12-h intervals during collection days 3–4 and storedtogether in a container below the metabolism crate until sampling on day 5. Air samples were collected from the airspace

Author's personal copy
98 C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104
above the surface of the amalgamated urine and faeces and were used to measure odour concentration. Air sampling wasperformed as described by Lynch et al. (2008). Samples were contained in 20 l Nalophan bags (and analysed for odourconcentration (OuE/m3) using an Ecoma T07 dynamic olfactometer (ECOMA, Honigsee, Germany) as described previouslyby Hayes et al. (2004).
2.5. Coefficient of total tract apparent digestibility and N balance studies
Daily urine production was collected separately beneath the metabolism crate in a plastic container and recorded. Toavoid N volatilisation, 20 cm3 (25%) sulphuric acid (H2SO4) was added to the container daily. A weak solution (2%) of sulphuricacid was applied to the connecting funnel twice daily. A 50 ml aliquot of urine and 100 g aliquot of faeces were retained dailyand frozen (−20 ◦C) for subsequent determination of N.
CTTAD values and N balance data were calculated as follows:
CTTAD = nutrient in−nutrient outnutrient in
Total N excretion (g/d) = faecal N excretion + urinary N excretion
N retention (g/d) = N intake − Total N excretion
N absorption = N retentionN intake
2.6. Microbial analysis
After slaughter, digesta samples (approximately 10 g ± 1 g) were recovered from the proximal and distal colon of eachanimal (7 replicates/dietary treatment) in sterile conditions. Digesta samples were stored in sterile containers (Sarstedt,Wexford, Ireland), placed in insulated containers with dry ice and transported to the laboratory within 2 h. A 1.0 g samplewas removed from each digesta sample, serially diluted (1:10) in 9.0 ml aliquots of maximum recovery diluents (MRD,Oxoid, Basingstoke, UK) and spread (0.1 ml aliquots) onto selective agars. Populations of lactobacilli, bifidobacteria andEscherichia coli were selectively isolated and enumerated as described by previous authors (Pierce et al., 2005). Lactobacilliand Bifidobacterium spp. were selected as bacterial indicators of saccharolytic fermentation (Garry et al., 2007a; O’Shea et al.,2009b), while E. coli were chosen based on previous inverse associations of this species and other genera representative ofthe Enterobacteriaceae family with carbohydrate fermentation in the dGIT (Naughton et al., 2001; van Winsen et al., 2001;Heo et al., 2009). Typical colonies of each bacterium were counted, log transformed and are presented per gram of digesta.
2.7. pH measurements
The pH of digesta samples from the proximal and distal colon was determined after slaughter. Samples (7 repli-cates/dietary treatment) were placed in universal containers (Sarstedt, Wexford, Ireland) and pH measurements were madeusing a Mettler Toledo MP 220 pH meter, which was calibrated using certified pH 4 and 7 buffer solutions (Sigma–AldrichIreland Limited, Wicklow, Ireland). Distilled water was added to some very viscous samples from the proximal colon in orderto facilitate pH measurement. The pH measurements of manure samples at 0 and 240 h were determined similarly.
2.8. VFA profiling of digesta and manure samples
Samples of digesta from the proximal and distal colon of individual pigs (7 replicates/dietary treatment) were recoveredfor VFA analysis. VFA concentrations in the digesta were determined as described previously by Porter and Murray (2001)using a modified method (Pierce et al., 2006). Briefly, a sample of 1 g was diluted with distilled water (2.5 × weight of sample)and centrifuged at 1400 × g for 4 min (Sorvall GLC – 2B laboratory centrifuge). One ml of the subsequent supernatant and1 ml of internal standard (0.5 g 3-methyl-n-valeric acid in 1l of 0.15 mol/l oxalic acid) were mixed with 3 ml of distilled water.Following centrifugation to remove the precipitate, the sample was filtered through Whatman 0.45 �m polyethersulphonemembrane filters (Sigma–Aldrich Ireland Limited, Wicklow, Ireland) into a chromatographic sample vial. A sample of 1 �lwas injected into a model 3800 Varian gas chromatograph with a 25 m × 0.53 mm i.d. megabore column (coating CP-Wax 58(FFAP) – CB (no. CP7614)) (Varian, Middelburg, The Netherlands). VFA concentrations of manure from the ammonia studyat 0 and 240 h were similarly determined with minor adaptations as described previously by O’Connell et al. (2005). Thebranched-chain fatty acids isovaleric acid and isobutyric acid, and valeric acid were designated as odorous for the purposeof this study due to their nauseating characteristics which are detectable at low concentrations by human olfactory senses(Le et al., 2005; Mackie et al., 1998).
2.9. Laboratory analysis of samples
The proximate analysis of diets and faeces for dry matter (DM) (method 934.01) and crude ash (method 942.05) werecarried out according to the Association of Official Analytical Chemists (AOAC, 1995). The DM of the diets and faeces were

Author's personal copy
C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104 99
determined after drying for 24 h at 100 ◦C. The ash content of diets and faeces was determined after ignition of a weighedsample in a muffle furnace (Nabertherm, Bremen, Germany) at 500 ◦C for 4 h. The gross energy (GE) of diets and faecessamples was determined using an adiabatic bomb calorimeter (Parr Instruments, IL, USA). The neutral detergent fibre (NDF),acid detergent fibre (ADF) and acid detergent lignin (ADL) fractions of diets and faeces were determined using a Fibertecextraction unit (Tecator, Hoganans, Sweden) as described by van Soest et al. (1991). The N concentration of diets and urinewas determined using a LECO FP 528 instrument (Leco Instruments, Cheshire, U.K.). The N concentration of fresh faeces wasanalysed by the macro-Kjeldahl technique using a Buchi digestion and distillation apparatus (Buchi Labortechnik, Flawil,Switzerland). The total �-glucan and xylose content of diets were determined colorimetrically after enzymatic hydrolysisusing a Megazyme assay kit (Megazyme International Ireland Ltd., Co. Wicklow, Ireland). The insoluble and soluble �-glucan contents were determined as described by McCleary and Glennie-Holmes (1985). The hemicellulose content of dietswas estimated as the numerical difference between NDF and ADF. The cellulose content was estimated as the numericaldifference between ADF and ADL. The lignin content is presented as ADL. The dietary �-glucanase and �-xylanase activitywere determined as described by Somogyi (1960). The activity of �-glucanase and �-xylanase refers to the units of glucoseand xylose liberated per gram of respective diet.
2.10. Statistical analysis
Experimental data were analysed as a complete randomised design using the GLM procedure of the SAS Institute (1985).The statistical model investigated the effect of dietary treatment. A contrast statement was used to compare; (1) B diet vsWG diet, (2) B diet vs BE diet and (3) WG diet vs T4 WGE diet. All data in the tables are presented as least-square means(LSM ± SEM). The individual pig served as the experimental unit. Live weight was included in the statistical model as acovariate.
3. Results
3.1. Dietary analysis
Chemical analysis indicated an average total �-glucan content of 22.9 g/kg for the barley-based diets and an average total�-glucan content of 22.7 g/kg for the wheat-based diets containing a source of exogenous �-glucans.
3.2. Coefficient of apparent total tract digestibility and N balance study
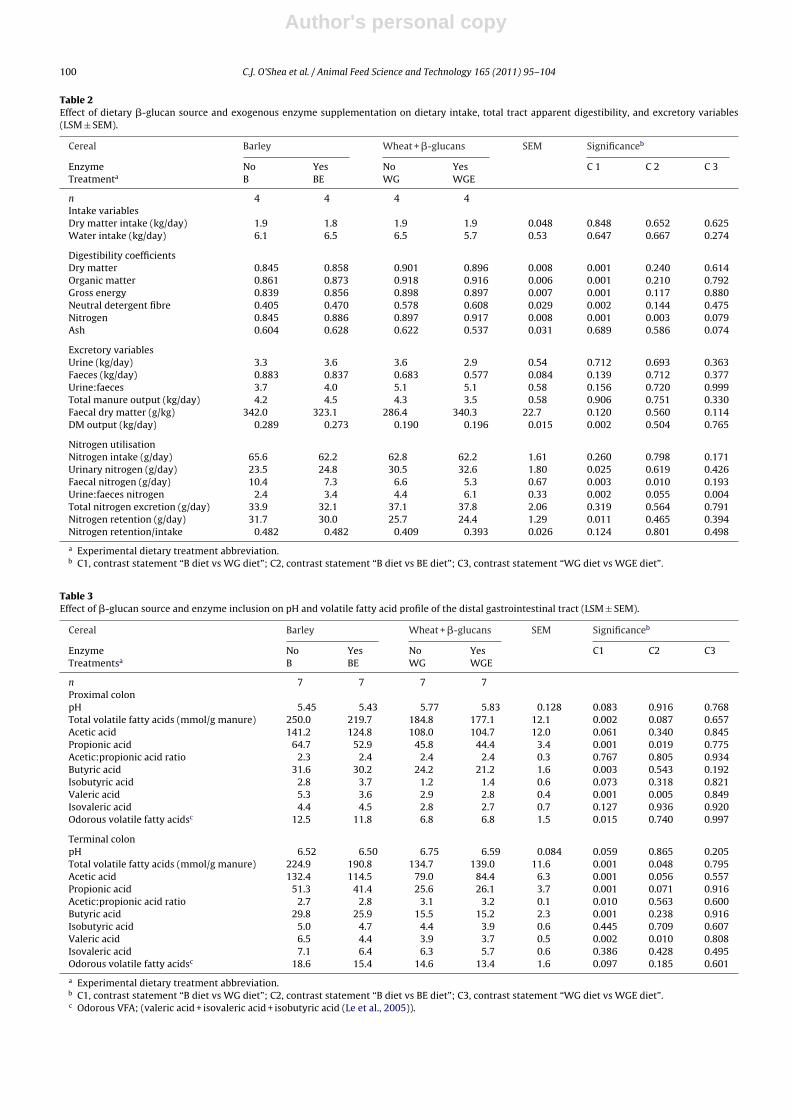
Pigs offered the WG diet had an increased CTTAD of dry matter (P = 0.001), organic matter (P = 0.001), gross energy(P = 0.001), neutral detergent fibre (P = 0.002) and N (P = 0.001) and decreased dry matter output (P = 0.002) compared withthe B diet (Table 2). Pigs offered the BE diet had increased CTTAD of N (P = 0.003) compared with pigs offered the B diet.
Offering the WG diet increased urinary N (P = 0.025), the urine:faeces N excretion ratio (P = 0.002), and decreased faecal N(P = 0.003) and N retention (P = 0.011), compared with the B diets. Pigs offered the BE diet had decreased faecal N (P = 0.010)compared with the B diet. Pigs offered the WGE diet had an increased urine:faeces N excretion ratio (P = 0.004) comparedwith the WG diet.
3.3. Selected bacterial counts and VFA profile of the distal gastrointestinal tract
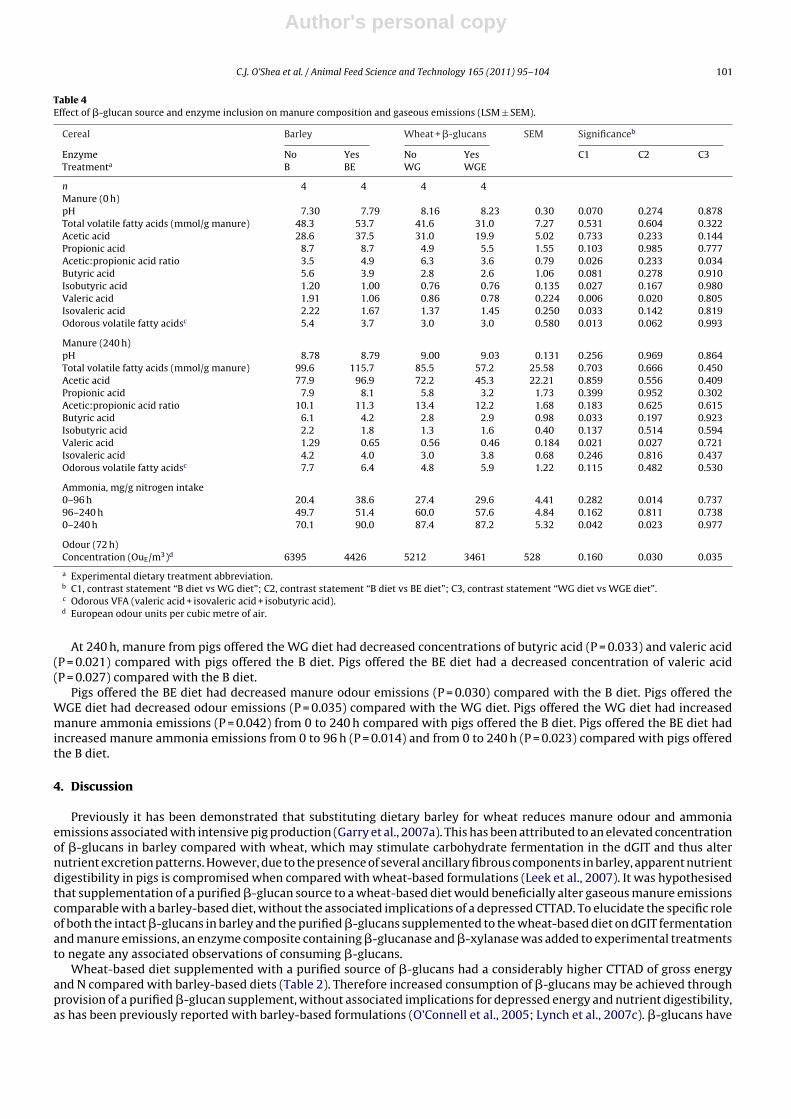
In the proximal colon, pigs offered the WG diet had decreased concentrations of total VFA (P = 0.002), propionic acid(P = 0.001), butyric acid (P = 0.003), valeric acid (P = 0.001) and overall odorous VFA (P = 0.015) compared with offering the Bdiet (Table 3). Pigs offered the BE diet had decreased concentrations of propionic acid (P = 0.019) and valeric acid (P = 0.005)compared with pigs offered the B diet.
In the terminal colon, pigs offered the WG diet had decreased concentrations of total VFA (P = 0.001), acetic acid (P = 0.001),propionic acid (P = 0.001), butyric acid (P = 0.001) and valeric acid (P = 0.002) and an increased acetic:propionic acid ratio(P = 0.010) compared with pigs offered the B diet. Pigs offered the BE diet had a decreased concentration of total VFA (P = 0.048)and a decreased concentration of valeric acid (P = 0.010) compared with pigs offered the B diet. There was no effect of dietary�-glucan source (P > 0.05) or enzyme inclusion (P > 0.05) on selected bacterial numbers in the terminal colon of finishingpigs.
3.4. Manure composition and emissions
At 0 h, manure from pigs offered the WG diet had an increased acetic:propionic acid ratio (P = 0.026) and a decreased con-centration of isobutyric acid (P = 0.027), valeric acid (P = 0.006), isovaleric acid (P = 0.033) and overall odorous VFA (P = 0.013)compared with pigs offered the B diet (Table 4). Pigs offered the BE diet had a decreased concentration of valeric acid(P = 0.020) compared with pigs offered the B diet. Pigs offered the WGE diet had a decreased acetic:propionic acid ratio(P = 0.034) compared with pigs offered the WG diet.
Author's personal copy
100 C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104
Table 2Effect of dietary �-glucan source and exogenous enzyme supplementation on dietary intake, total tract apparent digestibility, and excretory variables(LSM ± SEM).
Cereal Barley Wheat + �-glucans SEM Significanceb
Enzyme No Yes No Yes C 1 C 2 C 3Treatmenta B BE WG WGE
n 4 4 4 4Intake variablesDry matter intake (kg/day) 1.9 1.8 1.9 1.9 0.048 0.848 0.652 0.625Water intake (kg/day) 6.1 6.5 6.5 5.7 0.53 0.647 0.667 0.274
Digestibility coefficientsDry matter 0.845 0.858 0.901 0.896 0.008 0.001 0.240 0.614Organic matter 0.861 0.873 0.918 0.916 0.006 0.001 0.210 0.792Gross energy 0.839 0.856 0.898 0.897 0.007 0.001 0.117 0.880Neutral detergent fibre 0.405 0.470 0.578 0.608 0.029 0.002 0.144 0.475Nitrogen 0.845 0.886 0.897 0.917 0.008 0.001 0.003 0.079Ash 0.604 0.628 0.622 0.537 0.031 0.689 0.586 0.074
Excretory variablesUrine (kg/day) 3.3 3.6 3.6 2.9 0.54 0.712 0.693 0.363Faeces (kg/day) 0.883 0.837 0.683 0.577 0.084 0.139 0.712 0.377Urine:faeces 3.7 4.0 5.1 5.1 0.58 0.156 0.720 0.999Total manure output (kg/day) 4.2 4.5 4.3 3.5 0.58 0.906 0.751 0.330Faecal dry matter (g/kg) 342.0 323.1 286.4 340.3 22.7 0.120 0.560 0.114DM output (kg/day) 0.289 0.273 0.190 0.196 0.015 0.002 0.504 0.765
Nitrogen utilisationNitrogen intake (g/day) 65.6 62.2 62.8 62.2 1.61 0.260 0.798 0.171Urinary nitrogen (g/day) 23.5 24.8 30.5 32.6 1.80 0.025 0.619 0.426Faecal nitrogen (g/day) 10.4 7.3 6.6 5.3 0.67 0.003 0.010 0.193Urine:faeces nitrogen 2.4 3.4 4.4 6.1 0.33 0.002 0.055 0.004Total nitrogen excretion (g/day) 33.9 32.1 37.1 37.8 2.06 0.319 0.564 0.791Nitrogen retention (g/day) 31.7 30.0 25.7 24.4 1.29 0.011 0.465 0.394Nitrogen retention/intake 0.482 0.482 0.409 0.393 0.026 0.124 0.801 0.498
a Experimental dietary treatment abbreviation.b C1, contrast statement “B diet vs WG diet”; C2, contrast statement “B diet vs BE diet”; C3, contrast statement “WG diet vs WGE diet”.
Table 3Effect of �-glucan source and enzyme inclusion on pH and volatile fatty acid profile of the distal gastrointestinal tract (LSM ± SEM).
Cereal Barley Wheat + �-glucans SEM Significanceb
Enzyme No Yes No Yes C1 C2 C3Treatmentsa B BE WG WGE
n 7 7 7 7Proximal colonpH 5.45 5.43 5.77 5.83 0.128 0.083 0.916 0.768Total volatile fatty acids (mmol/g manure) 250.0 219.7 184.8 177.1 12.1 0.002 0.087 0.657Acetic acid 141.2 124.8 108.0 104.7 12.0 0.061 0.340 0.845Propionic acid 64.7 52.9 45.8 44.4 3.4 0.001 0.019 0.775Acetic:propionic acid ratio 2.3 2.4 2.4 2.4 0.3 0.767 0.805 0.934Butyric acid 31.6 30.2 24.2 21.2 1.6 0.003 0.543 0.192Isobutyric acid 2.8 3.7 1.2 1.4 0.6 0.073 0.318 0.821Valeric acid 5.3 3.6 2.9 2.8 0.4 0.001 0.005 0.849Isovaleric acid 4.4 4.5 2.8 2.7 0.7 0.127 0.936 0.920Odorous volatile fatty acidsc 12.5 11.8 6.8 6.8 1.5 0.015 0.740 0.997
Terminal colonpH 6.52 6.50 6.75 6.59 0.084 0.059 0.865 0.205Total volatile fatty acids (mmol/g manure) 224.9 190.8 134.7 139.0 11.6 0.001 0.048 0.795Acetic acid 132.4 114.5 79.0 84.4 6.3 0.001 0.056 0.557Propionic acid 51.3 41.4 25.6 26.1 3.7 0.001 0.071 0.916Acetic:propionic acid ratio 2.7 2.8 3.1 3.2 0.1 0.010 0.563 0.600Butyric acid 29.8 25.9 15.5 15.2 2.3 0.001 0.238 0.916Isobutyric acid 5.0 4.7 4.4 3.9 0.6 0.445 0.709 0.607Valeric acid 6.5 4.4 3.9 3.7 0.5 0.002 0.010 0.808Isovaleric acid 7.1 6.4 6.3 5.7 0.6 0.386 0.428 0.495Odorous volatile fatty acidsc 18.6 15.4 14.6 13.4 1.6 0.097 0.185 0.601
a Experimental dietary treatment abbreviation.b C1, contrast statement “B diet vs WG diet”; C2, contrast statement “B diet vs BE diet”; C3, contrast statement “WG diet vs WGE diet”.c Odorous VFA; (valeric acid + isovaleric acid + isobutyric acid (Le et al., 2005)).
Author's personal copy
C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104 101
Table 4Effect of �-glucan source and enzyme inclusion on manure composition and gaseous emissions (LSM ± SEM).
Cereal Barley Wheat + �-glucans SEM Significanceb
Enzyme No Yes No Yes C1 C2 C3Treatmenta B BE WG WGE
n 4 4 4 4Manure (0 h)pH 7.30 7.79 8.16 8.23 0.30 0.070 0.274 0.878Total volatile fatty acids (mmol/g manure) 48.3 53.7 41.6 31.0 7.27 0.531 0.604 0.322Acetic acid 28.6 37.5 31.0 19.9 5.02 0.733 0.233 0.144Propionic acid 8.7 8.7 4.9 5.5 1.55 0.103 0.985 0.777Acetic:propionic acid ratio 3.5 4.9 6.3 3.6 0.79 0.026 0.233 0.034Butyric acid 5.6 3.9 2.8 2.6 1.06 0.081 0.278 0.910Isobutyric acid 1.20 1.00 0.76 0.76 0.135 0.027 0.167 0.980Valeric acid 1.91 1.06 0.86 0.78 0.224 0.006 0.020 0.805Isovaleric acid 2.22 1.67 1.37 1.45 0.250 0.033 0.142 0.819Odorous volatile fatty acidsc 5.4 3.7 3.0 3.0 0.580 0.013 0.062 0.993
Manure (240 h)pH 8.78 8.79 9.00 9.03 0.131 0.256 0.969 0.864Total volatile fatty acids (mmol/g manure) 99.6 115.7 85.5 57.2 25.58 0.703 0.666 0.450Acetic acid 77.9 96.9 72.2 45.3 22.21 0.859 0.556 0.409Propionic acid 7.9 8.1 5.8 3.2 1.73 0.399 0.952 0.302Acetic:propionic acid ratio 10.1 11.3 13.4 12.2 1.68 0.183 0.625 0.615Butyric acid 6.1 4.2 2.8 2.9 0.98 0.033 0.197 0.923Isobutyric acid 2.2 1.8 1.3 1.6 0.40 0.137 0.514 0.594Valeric acid 1.29 0.65 0.56 0.46 0.184 0.021 0.027 0.721Isovaleric acid 4.2 4.0 3.0 3.8 0.68 0.246 0.816 0.437Odorous volatile fatty acidsc 7.7 6.4 4.8 5.9 1.22 0.115 0.482 0.530
Ammonia, mg/g nitrogen intake0–96 h 20.4 38.6 27.4 29.6 4.41 0.282 0.014 0.73796–240 h 49.7 51.4 60.0 57.6 4.84 0.162 0.811 0.7380–240 h 70.1 90.0 87.4 87.2 5.32 0.042 0.023 0.977
Odour (72 h)Concentration (OuE/m3)d 6395 4426 5212 3461 528 0.160 0.030 0.035
a Experimental dietary treatment abbreviation.b C1, contrast statement “B diet vs WG diet”; C2, contrast statement “B diet vs BE diet”; C3, contrast statement “WG diet vs WGE diet”.c Odorous VFA (valeric acid + isovaleric acid + isobutyric acid).d European odour units per cubic metre of air.
At 240 h, manure from pigs offered the WG diet had decreased concentrations of butyric acid (P = 0.033) and valeric acid(P = 0.021) compared with pigs offered the B diet. Pigs offered the BE diet had a decreased concentration of valeric acid(P = 0.027) compared with the B diet.
Pigs offered the BE diet had decreased manure odour emissions (P = 0.030) compared with the B diet. Pigs offered theWGE diet had decreased odour emissions (P = 0.035) compared with the WG diet. Pigs offered the WG diet had increasedmanure ammonia emissions (P = 0.042) from 0 to 240 h compared with pigs offered the B diet. Pigs offered the BE diet hadincreased manure ammonia emissions from 0 to 96 h (P = 0.014) and from 0 to 240 h (P = 0.023) compared with pigs offeredthe B diet.
4. Discussion
Previously it has been demonstrated that substituting dietary barley for wheat reduces manure odour and ammoniaemissions associated with intensive pig production (Garry et al., 2007a). This has been attributed to an elevated concentrationof �-glucans in barley compared with wheat, which may stimulate carbohydrate fermentation in the dGIT and thus alternutrient excretion patterns. However, due to the presence of several ancillary fibrous components in barley, apparent nutrientdigestibility in pigs is compromised when compared with wheat-based formulations (Leek et al., 2007). It was hypothesisedthat supplementation of a purified �-glucan source to a wheat-based diet would beneficially alter gaseous manure emissionscomparable with a barley-based diet, without the associated implications of a depressed CTTAD. To elucidate the specific roleof both the intact �-glucans in barley and the purified �-glucans supplemented to the wheat-based diet on dGIT fermentationand manure emissions, an enzyme composite containing �-glucanase and �-xylanase was added to experimental treatmentsto negate any associated observations of consuming �-glucans.
Wheat-based diet supplemented with a purified source of �-glucans had a considerably higher CTTAD of gross energyand N compared with barley-based diets (Table 2). Therefore increased consumption of �-glucans may be achieved throughprovision of a purified �-glucan supplement, without associated implications for depressed energy and nutrient digestibility,as has been previously reported with barley-based formulations (O’Connell et al., 2005; Lynch et al., 2007c). �-glucans have

Author's personal copy
102 C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104
previously been implicated in perturbing digestion processes due to induction of gastrointestinal viscosity (Li et al., 1996).However, the impaired nutrient digestibility associated with barley-based diets likely reflects increased concentrations ofother cell structural components such as cellulose which obdurately resist digestion (Souffrant, 2001). The absence of aneffect of �-glucanase supplementation on nutrient digestibility supports this observation.
There was no significant difference between bacterial populations across dietary treatments. Previously, consumptionof barley-based diets have stimulated populations of bifidobacteria and lactobacilli and decreased populations of E. Coli inthe dGIT when compared with wheat-based diets, and this has been attributed to the elevated concentrations of �-glucanspresent in barley (O’Connell et al., 2005; Garry et al., 2007a). The absence of a significant difference between experimentaldiets in the current study may suggest that supplementation of purified �-glucans induced comparable numbers of bifi-dobacteria and lactobacilli and E. Coli with a barley-based diet; an improvement in the proportions of these bacteria whenaligned with previous studies. During transit throughout the dGIT, carbohydrate availability progressively diminishes andfermentation may become increasingly proteolytic (Bach Knudsen et al., 1993). The current study suggests that supple-mentation of a wheat-based diet with �-glucans delivered a suitable substrate to prolong proliferation of more typicallysaccharolytic microbial populations in the colon.
Both the total concentrations and proportions of microbial metabolites in the colonic contents were influenced by thevarying dietary treatments. Compared with the barley-based diet, consumption of the wheat-based diet containing purified�-glucans resulted in reduced concentrations of total VFA. The diminished production of total VFA may indicate the absence ofsufficient additional fermentable substrate such as cellulose and hemi-cellulose in the wheat-based diet containing purified�-glucans, when compared with the barley-based diet (Table 1). In vitro fermentation of cellulose primarily yields acetic,propionic and butyric acid (Sunvold et al., 1995). Remarkably, barley-based diets also elevated concentrations of odorousVFA (isobutyric, isovaleric and valeric acid) in the proximal colon and valeric acid in the terminal colon, metabolites whichare predominately of proteolytic origin (Mortensen et al., 1988; Marounek et al., 2002) and characterized by pungent faecalodour notes at very low concentrations (Le et al., 2005). The observed increase in such indices of protein fermentation inthe colon of pigs offered the barley-based diets may reflect the increased excretion of faecal N when compared with the �-glucan-supplemented wheat-based diet, suggesting an increase in the quantities of proteinaceous material being conveyedto the dGIT. Previous authors have reported a depressive effect of diets containing elevated cellulose concentrations ondigestibility of N (Kiarie et al., 2007), of both dietary and endogenous origin (Furuya and Kaji, 1992). Therefore the provisionof purified �-glucans to a wheat-based diet may offer a more efficacious method to beneficially modulate the contents ofthe dGIT compared with a barley-based formulation, whereby a fermentable �-glucan source may be provided without theassociated implications for a depressed CTTAD of N and potential formation of odorous metabolites.
The reduction in concentrations of odorous VFA observed in the dGIT of pigs offered the �-glucan supplemented wheatdiet was correspondingly reflected in manure samples at both 0 and 240 h. Our group has previously noted that patterns ofVFA production observed in the dGIT may persist in excreted manure, and may thus offer an indication on the developmentof subsequent manure odour emissions (O’Shea et al., 2009b, 2010). In contrast to other studies, which have reported greatermanure odour emissions associated with consumption of wheat-based diets (Garry et al., 2007b), offering the wheat-baseddiet supplemented with purified �-glucans resulted in a numerical decrease in manure odour emissions compared withthe barley-based diet. The improvements in manure odour through supplementation with an exogenous, purified �-glucansource suggest that a conventional wheat-based diet may be successfully modified to result in odour emissions comparablewith manure from pigs offered a barley-based diet.
The addition of an enzyme composite comprising of �-glucanase and �-xylanase reduced manure odour compared withboth the barley-based diet and the wheat-based diet containing purified �-glucans. This may reflect the substantial reduction(approx. 25%) in daily faecal N from pigs offered enzyme-supplemented diets. Faecal N is predominately microbial in origin(Kirchgessner et al., 1994), and upon further fermentation may generate noxious compounds (Zahn et al., 2001; Blachieret al., 2007). However the reduction in faecal N was not reflected in substantial reductions in odorous VFA, reflecting thecomplex and interactive properties of odorous compounds in influencing manure odour (Hobbs et al., 1997). It is likely thecomposition of faecal N as well as the quantities is relevant in determining the emission of odorous volatile compounds andthe expression of manure odour. Future investigative studies may benefit from identification and quantification of a widerrange of bacterial metabolites to greater define the effect of dietary modification in addressing manure odour emissions.
Previously, consumption of barley-based diets has consistently reduced manure ammonia emissions when comparedwith a conventional wheat-based diet (Lynch et al., 2007b). Furthermore, this effect is simultaneously negated by the provi-sion of an enzyme composite containing a �-glucanase agent (O’Connell et al., 2005; Leek et al., 2007), thus suggesting a rolefor �-glucans in ameliorating this environmental pollutant. Despite the supplementation of purified �-glucans to a wheat-based diet in the current study, manure ammonia emissions were not diminished to levels comparable with consumptionof the barley-based diet. Furthermore, the integration of a �-glucanase composite into the experimental design negated thebeneficial influence of the barley-based diet on manure ammonia emissions, but not the �-glucan supplemented wheat-based diet (Table 4). The role of dietary fermentable carbohydrate in influencing manure ammonia may be attributed to arange of polysaccharides, and not specifically �-glucans (Kerr et al., 2006). Furthermore, previous authors have describedinteractions between �-glucans with various other components such as arabinoxylans (Izydorczyk and MacGregor, 2000).It is conceivable that in a purified form, without the protection typically afforded through complex-forming with other cellcomponents, that exogenous �-glucans are degraded to a greater extent (Bach Knudsen et al., 1993; Johansen et al., 1993).Consequently, consumption of �-glucan in a purified form may not exert the same influence in influencing N excretion

Author's personal copy
C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104 103
patterns as is typically seen with dietary formulations containing native fermentable carbohydrates (Canh et al., 1998). Theincrease in urinary N from pigs offered the wheat-based diet containing purified �-glucans supports this proposition.
In summary, the current study demonstrates that an extracted, purified �-glucan source may be successfully supple-mented to a wheat-based pig diet without the implications of depressed CTTAD, as typically observed with consumption ofa barley-based diet. Furthermore, offering a purified source of �-glucans resulted in beneficial reductions in odorous VFA inthe dGIT and in excreted manure, and may reflect improvements in N digestibility. This modification of the composition ofthe dGIT and subsequent manure was accompanied by odorous emissions comparable with consumption of the barley-baseddiet. However, the influence exerted by purified dietary �-glucans in ameliorating manure ammonia emissions comparablewith an intact source of �-glucans was not substantial, and may reflect the functionality of �-glucans as they relate to othercell wall constituents in native barley.
Acknowledgements
Funding for this research was provided under the National Development Plan, through the Research Stimulus Fund,administered by the Irish Department of Agriculture, Fisheries & Food.
References
AOAC, 1995. Official methods of analysis. In: 16th Assoc. Off. Anal. Chem. , Washington DC, USA.Autio, K., Mannonen, L., Pietila, K., Koskinen, M., Siika-aho, M., Linko, M., Morgan, A., 1996. Incubation of barley kernel sections with purified cell wall
degrading enzymes. J. Inst. Brew. 102, 427–432.Bach Knudsen, K.E., 1997. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Technol. 67, 319–338.Bach Knudsen, K.E., Jensen, B.B., Hansen, I., 1993. Oat bran but not a �-glucan-enriched oat fraction enhances butyrate production in the large intestine of
pigs. J. Nutr. 123, 1235–1247.Blachier, F., Mariotti, F., Huneau, J.F., Tome, D., 2007. Effects of amino acid-derived luminal metabolites on the colonic epithelium and physiopathological
consequences. Amino Acids 33, 547–652.Canh, T.T., Sutton, A.L., Aarnink, A.J., Verstegen, M.W., Schrama, J.W., Bakker, G.C., 1998. Dietary carbohydrates alter the fecal composition and pH and the
ammonia emission from slurry of growing pigs. J. Anim. Sci. 76, 1887–1895.Carpenter, D.A., O’ Mara, F.P., O’ Doherty, J.V., 2004. The effect of dietary crude protein concentration on growth performance, carcass composition and
nitrogen excretion in entire grower-finisher pigs. Irish J. Agric. Food Res. 43, 227–236.Conway, E.J., 1957. Microdiffusion Analysis and Volumetric Error. Crosby Lockwood and Son, London, UK.Furuya, S., Kaji, Y., 1992. The effects of feed intake and purified cellulose on the endogenous ileal amino acid flow in growing pigs. Br. J. Nutr. 68, 463–472.Garry, B.P., Fogarty, M., Curran, T.P., O’Connell, M.J., O’Doherty, J.V., 2007a. The effect of cereal type and enzyme addition on pig performance, intestinal
microflora, and ammonia and odour emissions. Animal 1, 751–757.Garry, B.P., Fogarty, M., Curran, T.P., O’Doherty, J.V., 2007b. Effect of cereal type and exogenous enzyme supplementation in pig diets on odour and ammonia
emissions. Livest. Sci. 109, 212–215.Hayes, E.T., Leek, A.B., Curran, T.P., Dodd, V.A., Carton, O.T., Beattie, V.E., O’Doherty, J.V., 2004. The influence of diet crude protein level on odour and ammonia
emissions from finishing pig houses. Bioresour. Technol. 91, 309–315.Heo, J.M., Kim, J.C., Hansen, C.F., Mullan, B.P., Hampson, D.J., Pluske, J.R., 2009. Feeding a diet with decreased protein content reduces indices of protein
fermentation and the incidence of postweaning diarrhea in weaned pigs challenged with an enterotoxigenic strain of Escherichia coli. J. Anim. Sci. 87,2833–2843.
Hobbs, P.J., Misselbrook, T.H., Pain, B.F., 1997. Characterisation of odorous compounds and emissions from slurries produced from weaner pigs fed dry feedand liquid diets. J. Sci. Food Agric. 73, 437–445.
INRA-AFZ, 2004. Tables of composition and nutritional value of feed materials. In: Sauvant, D., Perez, J.M., Tran, G. (Eds.), Pigs, Poultry, Cattle, Sheep, Goats,Rabbits, Horses, Fish. Wageningen Academic Publishers, The Netherlands, pp. 13–300.
Izydorczyk, M.S., MacGregor, A.W., 2000. Evidence of intermolecular interactions of �-glucans and arabinoxylans. Carbohydr. Polym. 41, 417–420.Johansen, H.N., Wood, P.J., Knudsen, K.E.B., 1993. Molecular weight changes in the (1 → 3)(1 → 4)-�-D-glucan of oats incurred by the digestive processes in
the upper gastrointestinal tract of pigs. J. Agric. Food Chem. 41, 2347–2352.Kerr, B.J., Ziemer, C.J., Trabue, S.L., Crouse, J.D., Parkin, T.B., 2006. Manure composition of swine as affected by dietary protein and cellulose concentrations.
J. Anim. Sci. 84, 1584–1592.Kiarie, E., Nyachoti, C.M., Slominski, B.A., Blank, G., 2007. Growth performance, gastrointestinal microbial activity, and nutrient digestibility in early-weaned
pigs fed diets containing flaxseed and carbohydrase enzyme. J. Anim. Sci. 85, 2982–2993.Kirchgessner, M., Kreuzer, M., Machmüller, A., Roth-Maier, D.A., 1994. Evidence for a high efficiency of bacterial protein synthesis in the digestive tract of
adult sows fed supplements of fibrous feedstuffs. Anim. Feed Sci. Technol. 46, 293–306.Le, P.D., Aarnink, A.J.A., Ogink, N.W.M., Becker, P.M., Verstegen, M.W.A., 2005. Odour from animal production facilities: its relationship to diet. Nutr. Res.
Rev. 18, 3–30.Leek, A.B.G., Callan, J.J., Reilly, P., Beattie, V.E., O’Doherty, J.V., 2007. Apparent component digestibility and manure ammonia emission in finishing pigs fed
diets based on barley, maize or wheat prepared without or with exogenous non-starch polysaccharide enzymes. Anim. Feed Sci. Technol. 135, 86–99.Li, S., Sauer, W.C., Huang, S.X., Gabert, V.M., 1996. Effect of beta-glucanase supplementation to hulless barley- or wheat-soybean meal diets on the
digestibilities of energy, protein, beta-glucans, and amino acids in young pigs. J. Anim. Sci. 74, 1649–1656.Lynch, M.B., O’Shea, C.J., Sweeney, T., Callan, J.J., O’Doherty, J.V., 2008. Effect of crude protein concentration and sugar-beet pulp on nutrient digestibility,
nitrogen excretion, intestinal fermentation and manure ammonia and odour emissions from finisher pigs. Animal 2, 425–434.Lynch, M.B., Sweeney, T., Callan, B.F.J.J., O’Doherty, J.V., 2007a. The effect of high and low dietary crude protein and inulin supplementation on nutrient
digestibility, nitrogen excretion, intestinal microflora and manure ammonia emissions from finisher pigs. Animal 1, 1112–1121.Lynch, M.B., Sweeney, T., Callan, J.J., O’Doherty, J.V., 2007b. The effect of dietary barley level on volatile fatty acid concentration and manure ammonia
emissions in finishing pigs. Livest. Sci. 109, 236–239.Lynch, M.B., Sweeney, T., Callan, J.J., O’Doherty, J.V., 2007c. Effects of increasing the intake of dietary �-glucans by exchanging wheat for barley on nutrient
digestibility, nitrogen excretion, intestinal microflora, volatile fatty acid concentration and manure ammonia emissions in finishing pigs. Animal 1,812–819.
Mackie, R.I., Stroot, P.G., Varel, V.H., 1998. Biochemical identification and biological origin of key odor components in livestock waste. J. Anim. Sci. 76,1331–1342.
Marounek, M., Adamec, T., Skivanova, V., Latsik, N.I., 2002. Nitrogen and in vitro fermentation of nitrogenous substrates in caecal contents of the pig. ActaVet. Brno 71, 429–433.
McCleary, B.V., Glennie-Holmes, M., 1985. Enzymatic quantification of (1,3) (1,4)-�-D-glucan in barley and malt. J. Inst. Brew. 91, 285–295.

Author's personal copy
104 C.J. O’Shea et al. / Animal Feed Science and Technology 165 (2011) 95–104
Mortensen, P.B., Holtug, K., Rasmussen, H.S., 1988. Short-chain fatty acid production from mono- and disaccharides in a fecal incubation system: implicationsfor colonic fermentation of dietary fibre in humans. J. Nutr. 118, 321–325.
Naughton, P.J., Mikkelsen, L.L., Jensen, B.B., 2001. Effects of nondigestible oligosaccharides on Salmonella enterica Serovar Typhimurium and nonpathogenicEscherichia coli in the pig small intestine in vitro. Appl. Environ. Microbiol. 67, 3391–3395.
NRC, 1998. Nutrient Requirements of Swine, 10th rev. Natl. Acad. Press, Washington, DC, USA.O’Connell, J.M., Callan, J.J., Byrne, C., Sweeney, T., O’Doherty, J.V., 2005. The effect of cereal type and exogenous enzyme supplementation in pig diets on
nutrient digestibility, intestinal microflora, volatile fatty acid concentration and manure ammonia emissions. Anim. Sci. 81, 357–364.O’Connell, J.M., Callan, J.J., O’Doherty, J.V., 2006. The effect of dietary crude protein level, cereal type and exogenous enzyme supplementation on nutrient
digestibility, nitrogen excretion, faecal volatile fatty acid concentration and ammonia emissions from pigs. Anim. Feed Sci. Technol. 127, 73–88.O’Shea, C.J., Lynch, B., Lynch, M.B., Callan, J.J., O’Doherty, J.V., 2009a. Ammonia emissions and dry matter of separated pig manure fractions as affected by
crude protein concentration and sugar beet pulp inclusion of finishing pig diets. Agric. Ecosyst. Environ. 131, 154–160.O’Shea, C.J., Lynch, M.B., Callan, J.J., O’Doherty, J.V., 2009b. Dietary supplementation with chitosan at high and low crude protein concentrations promotes
Enterobacteria spp in the caecum and colon and increases manure odour emissions from finisher boars. In: XI International Symposium on DigestivePhysiology of Pigs, Livestock Science , Montbrio del Camp, Costa Daurauda, Spain, p. 149.
O’Shea, C.J., Sweeney, T., Lynch, M.B., Gahan, D.A., Callan, J.J., O’Doherty, J.V., 2010. Effect of �-glucans contained in barley- and oat-based diets andexogenous enzyme supplementation on gastrointestinal fermentation of finisher pigs and subsequent manure odor and ammonia emissions. J. Anim.Sci. 88, 1411–1420.
Pierce, K.M., Sweeney, T., Brophy, P.O., Callan, J.J., McCarthy, P., O’Doherty, J.V., 2005. Dietary manipulation post weaning to improve piglet performanceand gastro-intestinal health. Anim. Sci. 81, 347–356.
Pierce, K.M., Sweeney, T., Callan, J.J., Byrne, C., McCarthy, P., O’Doherty, J.V., 2006. The effect of inclusion of a high lactose supplement in finishing diets onnutrient digestibility, nitrogen excretion, volatile fatty acid concentrations and ammonia emission from boars. Anim. Feed Sci. Technol. 125, 45–60.
Porter, M.G., Murray, R.S., 2001. The volatility of components of grass silage on oven drying and the inter-relationship between dry-matter content estimatedby different analytical methods. Grass Forage Sci. 56, 405–411.
Somogyi, M., 1960. Modifications of two methods for the assay of amylase. Clin. Chem. 6, 23–35.Souffrant, W.B., 2001. Effect of dietary fibre on ileal digestibility and endogenous nitrogen losses in the pig. Anim. Feed Sci. Technol. 90, 93–102.Statistical Analysis Systems Institute, 1985. Statistical Analysis Systems, 6.12. SAS Institute Inc., Cary, NC, USA.Sunvold, G.D., Hussein, H.S., Fahey Jr., G.C., Merchen, N.R., Reinhart, G.A., 1995. In vitro fermentation of cellulose, beet pulp, citrus pulp, and citrus pectin
using fecal inoculum from cats, dogs, horses, humans, and pigs and ruminal fluid from cattle. J. Anim. Sci. 73, 3639–3648.van Soest, P.J., Robertson, J.B., Lewis, B.A., 1991. Methods for dietary fibre, neutral detergent fiber and non starch polysaccharides in relation to animal
nutrition. J. Dairy Sci. 74, 3583–3597.van Winsen, R.L., Urlings, B.A.P., Lipman, L.J.A., Snijders, J.M.A., Keuzenkamp, D., Verheijden, J.H.M., van Knapen, F., 2001. Effect of fermented feed on the
microbial population of the gastrointestinal tracts of pigs. Appl. Environ. Microbiol. 67, 3071–3076.Zahn, J.A., DiSpirito, A.A., Do, Y.S., Brooks, B.E., Cooper, E.E., Hatfield, J.L., 2001. Correlation of human olfactory responses to airborne concentrations of
malodorous volatile organic compounds emitted from swine effluent. J. Environ. Qual. 30, 624–634.




















