
The diversity of actinorhizal symbiosis
13
REVIEW ARTICLE The diversity of actinorhizal symbiosis Katharina Pawlowski & Kirill N. Demchenko Received: 24 January 2012 / Accepted: 14 February 2012 / Published online: 8 March 2012 # Springer-Verlag 2012 Abstract Filamentous aerobic soil actinobacteria of the genus Frankia can induce the formation of nitrogen-fixing nodules on the roots of a diverse group of plants from eight dicotyledonous families, collectively called actinorhizal plants. Within nodules, Frankia can fix nitrogen while being hosted inside plant cells. Like in legume/rhizobia symbio- ses, bacteria can enter the plant root either intracellularly through an infection thread formed in a curled root hair, or intercellularly without root hair involvement, and the entry mechanism is determined by the host plant species. Nodule primordium formation is induced in the root pericycle as for lateral root primordia. Mature actinorhizal nodules are cor- alloid structures consisting of multiple lobes, each of which represents a modified lateral root without a root cap, a superficial periderm and with infected cells in the expanded cortex. In this review, an overview of nodule induction mechanisms and nodule structure is presented including comparisons with the corresponding mechanisms in legume symbioses. Keywords Symbiotic nitrogen fixation . Root nodules . Infection threads . Frankia . Alnus . Casuarina . Datisca Introduction Two types of root nodule symbioses between nitrogen- fixing soil bacteria and higher plants are known: rhizobial symbioses with ca. 80% of all members of the legume family as well as one nonlegume genus, Parapsonia, and actinorhizal symbioses represented by a diverse group of more than 200 species from eight different families, collec- tively called actinorhizal plants and actinobacteria of the genus Frankia. In both cases, the microsymbionts induce the formation of special organs, root nodules, on the roots of their host plants and fix nitrogen while being stably intra- cellularly accommodated in nodule cells and being supplied with carbon sources by the plant. Both legumes and actino- rhizal plants belong to the Rosid I clade (Fig. 1), i.e., they go back to a common ancestor. Altogether, several independent evolutionary origins have been postulated for rhizobial sym- bioses (Doyle 2011) and three to four for actinorhizal sym- bioses (Fig. 1; Swensen 1996). Accordingly, it has been suggested that ca. 100 Ma ago, the common ancestor of Rosid I had acquired a unique feature upon which a root nodule symbiosis could evolve and that this evolution oc- curred several times 50–60 Ma ago (Doyle 2011; Oono et al. 2010; Soltis et al. 1995). There are two features unique to root nodule symbioses: (a) the induction of the formation of a novel plant organ by a bacterial signal factor and (b) the stable internal accommo- dation of a eubacterial symbiont in plant cells. While stable internal accommodation is not uncommon in plant–fungal interactions occurring in arbuscular mycorrhiza as well as in several pathogenic relationships (Parniske 2000), the stable internal accommodation of prokaryotic microsymbionts has, apart from root nodule symbioses, been described only for cyanobacteria in Gunnera species (Bergman et al. 1992). In Handling Editor: David Robinson K. Pawlowski (*) Department of Botany, Stockholm University, 10691 Stockholm, Sweden e-mail: [email protected] K. N. Demchenko Laboratory of Anatomy and Morphology, Komarov Botanical Institute, Russian Academy of Sciences, Prof. Popov St. 2, 197376 St. Petersburg, Russia Protoplasma (2012) 249:967–979 DOI 10.1007/s00709-012-0388-4
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The diversity of actinorhizal symbiosis

REVIEWARTICLE
The diversity of actinorhizal symbiosis
Katharina Pawlowski & Kirill N. Demchenko
Received: 24 January 2012 /Accepted: 14 February 2012 /Published online: 8 March 2012# Springer-Verlag 2012
Abstract Filamentous aerobic soil actinobacteria of thegenus Frankia can induce the formation of nitrogen-fixingnodules on the roots of a diverse group of plants from eightdicotyledonous families, collectively called actinorhizalplants. Within nodules, Frankia can fix nitrogen while beinghosted inside plant cells. Like in legume/rhizobia symbio-ses, bacteria can enter the plant root either intracellularlythrough an infection thread formed in a curled root hair, orintercellularly without root hair involvement, and the entrymechanism is determined by the host plant species. Noduleprimordium formation is induced in the root pericycle as forlateral root primordia. Mature actinorhizal nodules are cor-alloid structures consisting of multiple lobes, each of whichrepresents a modified lateral root without a root cap, asuperficial periderm and with infected cells in the expandedcortex. In this review, an overview of nodule inductionmechanisms and nodule structure is presented includingcomparisons with the corresponding mechanisms in legumesymbioses.
Keywords Symbiotic nitrogen fixation . Root nodules .
Infection threads .Frankia . Alnus .Casuarina .Datisca
Introduction
Two types of root nodule symbioses between nitrogen-fixing soil bacteria and higher plants are known: rhizobialsymbioses with ca. 80% of all members of the legumefamily as well as one nonlegume genus, Parapsonia, andactinorhizal symbioses represented by a diverse group ofmore than 200 species from eight different families, collec-tively called actinorhizal plants and actinobacteria of thegenus Frankia. In both cases, the microsymbionts inducethe formation of special organs, root nodules, on the roots oftheir host plants and fix nitrogen while being stably intra-cellularly accommodated in nodule cells and being suppliedwith carbon sources by the plant. Both legumes and actino-rhizal plants belong to the Rosid I clade (Fig. 1), i.e., they goback to a common ancestor. Altogether, several independentevolutionary origins have been postulated for rhizobial sym-bioses (Doyle 2011) and three to four for actinorhizal sym-bioses (Fig. 1; Swensen 1996). Accordingly, it has beensuggested that ca. 100 Ma ago, the common ancestor ofRosid I had acquired a unique feature upon which a rootnodule symbiosis could evolve and that this evolution oc-curred several times 50–60 Ma ago (Doyle 2011; Oono et al.2010; Soltis et al. 1995).
There are two features unique to root nodule symbioses:(a) the induction of the formation of a novel plant organ by abacterial signal factor and (b) the stable internal accommo-dation of a eubacterial symbiont in plant cells. While stableinternal accommodation is not uncommon in plant–fungalinteractions occurring in arbuscular mycorrhiza as well as inseveral pathogenic relationships (Parniske 2000), the stableinternal accommodation of prokaryotic microsymbionts has,apart from root nodule symbioses, been described only forcyanobacteria in Gunnera species (Bergman et al. 1992). In
Handling Editor: David Robinson
K. Pawlowski (*)Department of Botany, Stockholm University,10691 Stockholm, Swedene-mail: [email protected]
K. N. DemchenkoLaboratory of Anatomy and Morphology,Komarov Botanical Institute, Russian Academy of Sciences,Prof. Popov St. 2,197376 St. Petersburg, Russia
Protoplasma (2012) 249:967–979DOI 10.1007/s00709-012-0388-4

this context, it is intriguing that the evolution of rhizobialsymbioses obviously involved the recruitment of features ofthe evolutionary older arbuscular mycorrhizal (AM) symbi-osis (Markmann and Parniske 2009). AM symbioses goback at least 450 Ma and may precede the colonization ofland by plants (Parniske 2008). The rhizobial signal factorsthat elicit nodule formation, the so-called Nod factors, lip-ochitooligosaccharides, are chemically similar to the signalfactors of AM fungi, the so-called Myc factors that alsorepresent lipochitooligosaccharides (Maillet et al. 2011).Nod and Myc factors also share part of their signal trans-duction pathways. As a rule, AM symbioses do not involvethe induction of organogenesis, but only the internalizationof fungal hyphae in roots and the formation of nutrientexchange structures in root cortical cells that existed beforethe signal exchange between microsymbiont and host plant.However, there are cases known where AM symbiosesinvolve the formation of lateral rootlike structures, so-called myconodules, i.e., where Myc factor action seemsto involve the induction of plant organ formation (Duhouxet al. 2001). However, due to the facts that (a) these symbi-oses involve tropical trees and (b) AM fungi are obligatesymbionts, this particular function has not been examined.Altogether, both unique features of root nodule symbiosesseem to build up onmechanisms developed forAM symbioses.
All root nodule symbioses have to cope with the so-called oxygen dilemma of nitrogen fixation: the action ofthe enzyme complex that reduces air dinitrogen to ammonia,nitrogenase, requires a lot of energy in the form of ATP bestprovided by aerobic respiration. One part of nitrogenase isirreversibly denatured by oxygen and, therefore, the enzymecomplex has to be kept in a microaerobic or anaerobicenvironment (Shah and Brill 1977). In the case of
rhizobia/legume symbioses, the microsymbionts cannot pro-vide oxygen protection for nitrogenase, so the oxygen di-lemma has to be solved by the host plant. Few rhizobia canfix nitrogen in the saprophytic state and, if they do, onlyunder microaerobic conditions; most rhizobia can fix nitro-gen only in symbiosis, hosted within plant cells and havingdifferentiated into their symbiotic form, bacteroids (Chen etal. 2003). Frankia strains, however, can fix nitrogen in thesaprophytic state under microaerobic as well as under aero-bic conditions (Benson and Silvester 1993).
Frankia strains can form three different cell types. Theygrow in hyphal form and also as multilocular sporangia.Under nitrogen limitation and aerobic conditions, they dif-ferentiate so-called vesicles at the tips of hyphae or at theends of short side hyphae. These vesicles are surrounded bymultilayered envelopes containing hopanoids, bacterial ste-roid lipids. The number of layers is correlated with theoxygen tension, indicating that they impede oxygen diffu-sion. In the vesicles, nitrogenase is formed and dinitrogen isfixed (Parsons et al. 1987; Berry et al. 1993).
Frankia strains can form vesicles during saprophyticgrowth and in symbiosis. Therefore, in symbiosis, they canprovide oxygen protection for nitrogenase and contribute tosolving the oxygen dilemma of nitrogen fixation. Interest-ingly, when formed in the saprophytic state, vesicles arealways spherical and septate, while in planta, the shape ofvesicles and their position in the infected cell depends on thehost plant (Newcomb and Wood 1987; Huss-Danell 1997).For example, in infected cells of nodules of Alnus sp.,spherical septate vesicles are positioned close to the plasmamembrane. In infected cells of nodules of Casuarina sp. andAllocasuarina sp., no vesicles are formed. In infected cellsof Datisca sp. and Coriaria sp., lanceolate vesicles in radial
Discaria chacaye
Ceanothus thyrsiflorus
Elaeagnus angustifolia
Shepherdia canadensis
Cercocarpus ledifolius
Purshia tridentata
Allocasuarina verticillata
Casuarina litorea
Alnus incana
Myrica gale
Comptonia peregrina
Datisca glomerata
Datisca cannabina
Coriaria myrtifolia
Medicago sativa
98100
100
100
100
100
80
52
57
58
97
69
79
Frankiacluster III
Frankiacluster I
Frankiacluster II
Rhamnaceae
Elaeagnaceae
Rosaceae
Casuarinaceae
Myricaceae
Datiscaceae
Betulaceae
CoriariaceaeFabaceae
Ceanothus
Gymnostoma
Myrica
Fig. 1 The host specificity in actinorhizal symbioses is shown in asimplified scheme adapted from Benson and Clawson (2000); thenumbering of Frankia clusters is adapted from Normand et al.(1996). Legumes (Fabales) are indicated in black, Fagales in red,Rosales in blue and Cucurbitales in green. Thick arrows connecting aFrankia cluster with a group of plants indicate that members of this
cluster are commonly associated with the plants. Dashed arrows indi-cate that members of this cluster have been isolated from or detected inan effective or ineffective nodule of a member of the plant group atleast once. It should be pointed out that not all strains isolated from, oridentified in, an actinorhizal nodule can reinfect the correspondingplant species (Ramirez-Saad et al. 1998)
968 K. Pawlowski, K.N. Demchenko

orientation form a sphere around the central vacuole of theinfected cell (Hafeez et al. 1984; Mirza et al. 1994; Silvesteret al. 1999). Altogether, in symbiosis, Frankia vesicles canrepresent a system-specific differentiation comparable to thebacteroids of rhizobia.
Figure 1 shows that phylogenetically, Frankia strainsgroup in three different clusters (Normand et al. 1996). Nostrain from cluster II has been cultured so far, which has ledto the hypothesis that these strains represent obligate sym-bionts. However, the genome sequence of one representativeof this cluster did not reveal obvious auxotrophies or clearevidence of genome reduction that is usually found in obli-gate symbionts (Moran 2003; Persson et al. 2011).
Host specificity can be assigned summarily. For example,only members of cluster II were found in nodules of actino-rhizal Cucurbitales and of Ceanothus sp. However, hostspecificity also exists within the Frankia clusters, i.e., notall members of a cluster can nodulate all plants associatedwith that cluster. For instance, all members of cluster I testedcan nodulate Myrica sp., but only some of them can nod-ulate either Alnus sp. or Casuarina sp.
Some genera (Gymnostoma, Myrica and Ceanothus) dif-fer in microsymbiont specificity from the rest of the plantfamily they belong to (Fig. 1). Some plant groups show aparticular native geographical distribution, e.g., Casuarina-ceae are native to Australia and the western Pacific. How-ever, since Elaeagnaceae and Betulaceae occur on mostcontinents and Coriariaceae and Datiscaceae are found intemperate zones of both hemispheres, all continents exceptfor Africa contain host plants of all three Frankia clusters,indicating that their soils should contain propagules of thecorresponding Frankia strains.
Actinorhizal nodule structure
Legume nodules represent stemlike organs with a peripheralvascular system and infected cells in the central tissue. Incontrast, actinorhizal nodules as well as nodules induced byrhizobia on the nonlegume genus Parasponia are coralloidorgans composed of multiple lobes representing modifiedlateral roots (Fig. 2a) with a central vascular system, withoutroot caps and with a superficial periderm and infected cellsin the expanded cortex (Fig. 2b, c). Parasponia sp. struc-turally and developmentally resemble actinorhizal nodules(Lancelle and Torrey 1984a, b). In this context, it should bementioned that with one exception, Datisca glomerata, allactinorhizal species as well as the members of Parasponiasp. represent woody plants, and their nodules are perennial.Actinorhizal plants can be separated into three differentphylogenetic groups, Fagales (members of the Betulaceae,Casuarinaceae and Myricaceae), Rosales (members of theRosaceae, Elaeagnaceae and Rhamnaceae) and Cucurbitales
(members of the Datiscaceae and Coriariaceae; Swensen1996; Fig. 1). In nodules of Fagales and Rosales, theinfected cells in the cortex are interspersed with uninfectedcells (Fig. 2b). In nodules of the Cucurbitales, the infectedcells form an uninterrupted domain, kidney-shaped in crosssection (Fig. 2d) on one side of the acentric stele (Newcomband Pankhurst 1982; Hafeez et al. 1984; Berg et al. 1999;Fig. 2c, e, f).
Infected cells, the only plant cell type able to stably inter-nally accommodate a eubacterial microsymbiont, are onlyfound in nodules, i.e., they are formed after signal exchangewith the microsymbionts. Uninfected nodule cortical cellsdiffer in their differentiation from root cortical cells in thatthey can store starch (Laplaze et al. 2000; Schubert et al.2011). The vascular system of nodules differs from the rootvascular system in some respects. On the whole plant level,root nodules represent nitrogen sources and carbon sinks.Assimilated products of nitrogen fixation are exported viathe xylem, while the nodule is supplied with the products ofphotosynthesis, sugars, via the phloem. In comparison withthe root vascular system, the nodule vascular system has alower contribution of xylem and more pericycle at the xylempoles (Berry et al. 2011; Fig. 2h, i), as is also found in roots ofother Cucurbitales (Demchenko and Demchenko 2001).
In some actinorhizal genera, e.g., Alnus, Coriaria andDatisca, the nodule periderm can be interrupted by lenticelsto improve nodule aeration and, therefore, the supply withdinitrogen and oxygen (Silvester et al. 1990). Actinorhizalplants that often grow in waterlogged areas have a problemwith nodule aeration. In most cases, this is solved by theformation of so-called nodule roots. The nodule lobe meri-stem is modified and develops to a nodule root meristem.Nodule roots have a superficial root cap and large air spacesin the cortex, which serve as air diffusion pathways to thenodule. The length of nodule roots is inversely correlatedwith the external oxygen tension (Schwintzer et al. 1982).D. glomerata can form both nodule roots and lenticels,depending on the aeration of the soil (Pawlowski 2002;Pawlowski et al. 2007). Alnus glutinosa, which often growsin waterlogged soil, does not form nodule roots, but pro-vides oxygen access to nodules via thermo-osmotically me-diated gas transport from the aerial parts to the roots(Schröder 1989).
Infection mechanisms
During the formation of root nodules, bacteria have to (a)enter the plant root, (b) cross part of the root cortex and (c)enter the infected cells. As in rhizobia/legume symbioses,these mechanisms are determined by the host plant. Entryinto the plant root occurs intracellularly via infection thread-like structures formed in root hairs in actinorhizal Fagales
The diversity of actinorhizal symbiosis 969
and intercellularly via dissolution of the middle lamellabetween adjacent root epidermal cells in actinorhizalRosales. Due to the fact that they are infected by noncultur-able microsymbionts, the infection mechanism of Cucurbi-tales has not yet been analyzed.
During intracellular infection of Fagales (Fig. 3a), thefirst response of the plant to the presence of the micro-symbiont is the deformation and branching of growing roothairs (Torrey 1976; Callaham and Torrey 1977), probablydue to blockage and reinitiation of tip growth as in legumes
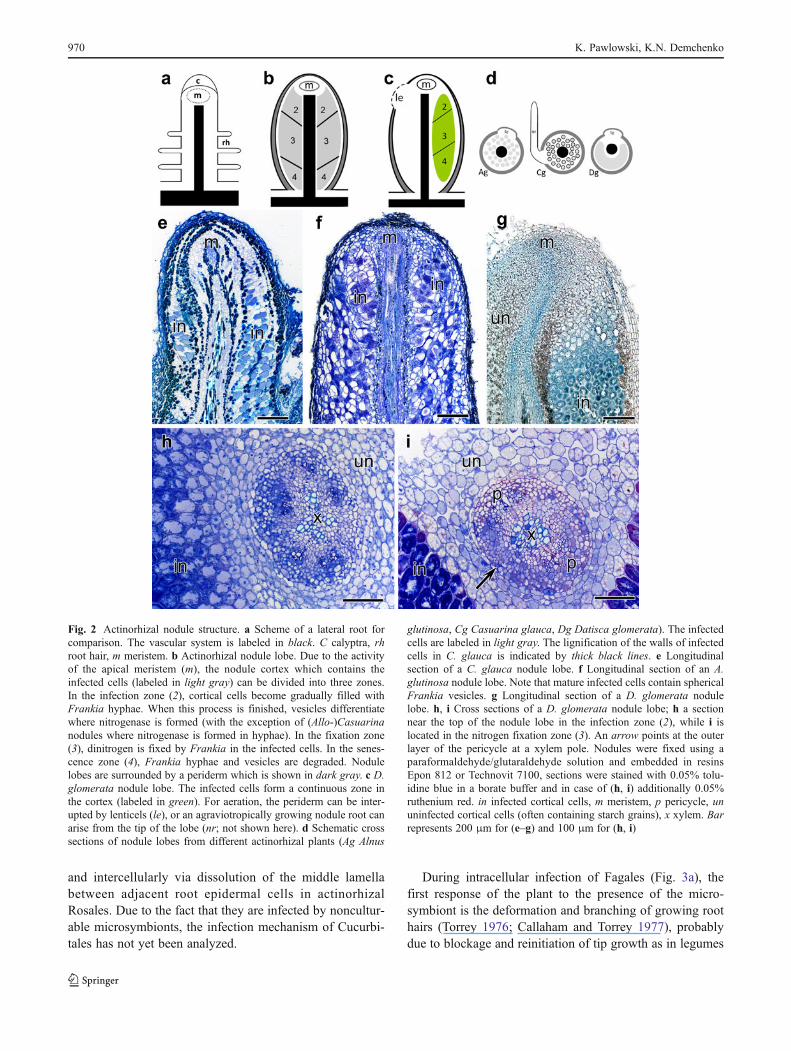
Fig. 2 Actinorhizal nodule structure. a Scheme of a lateral root forcomparison. The vascular system is labeled in black. C calyptra, rhroot hair, m meristem. b Actinorhizal nodule lobe. Due to the activityof the apical meristem (m), the nodule cortex which contains theinfected cells (labeled in light gray) can be divided into three zones.In the infection zone (2), cortical cells become gradually filled withFrankia hyphae. When this process is finished, vesicles differentiatewhere nitrogenase is formed (with the exception of (Allo-)Casuarinanodules where nitrogenase is formed in hyphae). In the fixation zone(3), dinitrogen is fixed by Frankia in the infected cells. In the senes-cence zone (4), Frankia hyphae and vesicles are degraded. Nodulelobes are surrounded by a periderm which is shown in dark gray. c D.glomerata nodule lobe. The infected cells form a continuous zone inthe cortex (labeled in green). For aeration, the periderm can be inter-upted by lenticels (le), or an agraviotropically growing nodule root canarise from the tip of the lobe (nr; not shown here). d Schematic crosssections of nodule lobes from different actinorhizal plants (Ag Alnus
glutinosa, Cg Casuarina glauca, Dg Datisca glomerata). The infectedcells are labeled in light gray. The lignification of the walls of infectedcells in C. glauca is indicated by thick black lines. e Longitudinalsection of a C. glauca nodule lobe. f Longitudinal section of an A.glutinosa nodule lobe. Note that mature infected cells contain sphericalFrankia vesicles. g Longitudinal section of a D. glomerata nodulelobe. h, i Cross sections of a D. glomerata nodule lobe; h a sectionnear the top of the nodule lobe in the infection zone (2), while i islocated in the nitrogen fixation zone (3). An arrow points at the outerlayer of the pericycle at a xylem pole. Nodules were fixed using aparaformaldehyde/glutaraldehyde solution and embedded in resinsEpon 812 or Technovit 7100, sections were stained with 0.05% tolu-idine blue in a borate buffer and in case of (h, i) additionally 0.05%ruthenium red. in infected cortical cells, m meristem, p pericycle, ununinfected cortical cells (often containing starch grains), x xylem. Barrepresents 200 μm for (e–g) and 100 μm for (h, i)
970 K. Pawlowski, K.N. Demchenko

(Heidstra et al. 1994). In legumes, this response is due to therhizobial lipochitooligosaccharide Nod factors which deter-mine the host specificity of the microsymbionts. In contrastto legumes, root hair deformation in actinorhizal plants canalso be induced by nonsymbiotic soil bacteria which mayact as infection helpers (Knowlton et al. 1980; Berry andTorrey 1983), indicating that it is not necessarily asymbiosis-specific reaction. Only few root hairs becomeinfected: a Frankia hypha associated with a deformed roothair penetrates the root hair cell wall, while the root hairplasma membrane invaginates and an infection threadlikestructure develops. Within this structure, the Frankia hyphais embedded within a plant-derived cell-wall-like pectin-richmatrix, the so-called encapsulation (Berry and Torrey 1983;Berry et al. 1986). Actinorhizal infection threadlike struc-tures differ from legume infection threads in that they do notcontain an infection thread matrix; the hyphal wall is incontact with the infection thread wall. For the sake ofsimplicity, these will be called “infection threads” through-out this manuscript.
Concomitant with infection thread formation, cell divi-sions are induced in the root cortex close to the infected roothair, giving rise to the formation of a so-called prenodule.The infection threads grow toward the prenodule and infectsome, but not all, prenodule cells by extensive branchingwithin these cells, filling them with hyphae in infectionthreads from the center outward (Schwintzer et al. 1982).Although the differentiation of infected and uninfected pre-nodule cells resembles that of their counterparts in maturenodule lobes (Laplaze et al. 2000), the prenodule is not the
precursor of a nodule lobe. In contrast to legumes, in actino-rhizal plants, cortical cell divisions do not lead to the for-mation of a nodule primordium. The cell divisions in thecortex cease, and the formation of the nodule primordium isinduced in the root pericycle. Infection threads grow fromthe infected prenodule cells toward the primordium andinfect primordium cells.
Infection thread growth and stable internalizationof bacteria in infected cells
The growth of infection threads in actinorhizal Fagalesresembles infection thread growth in legume nodules in thatcrossing of a cortical cell by an infection thread is precededby the formation of a preinfection thread (PIT; Berg 1999a).During PIT formation, the nucleus moves to the center ofthe cell, and microtubules and cytoplasm rearrange to form aphragmoplastlike structure (van Brussel et al. 1992). Thesestructures are polarized; the bulk of the cytoplasm and theendomembranes are located at the outer side. This polariza-tion of the cytoplasm is required for tip growth (Van Brusselet al. 1992). Since infection threads grow by cell-to-cellpassage (the so-called transcellular infection thread growth),infected cells often occur in files in the nodule lobes ofintracellularly infected actinorhizal plants (Fig. 4a). Thesimilarity between the infection thread growth mechanismin actinorhizal Fagales and legumes is further supported bythe fact that in both symbioses, the promoter of a nodule-specific subtilase gene from Casuarina glauca, cg12, is only
Fig. 3 Infection mechanisms. a Intracellular infection. Root hairsdeform and branch, putatively in response to Frankia equivalents ofrhizobial Nod factor. A Frankia hypha enters the plant via an infectionthread formed in a curled root hair. Concomitantly, cell divisions areinduced in the outer cortex and the infection thread grows via prein-fection thread structures (PITs) toward these dividing cells and infectssome of them, i.e., fills by extensive branching from the center out-ward. The dividing cortical cells represent the prenodule. Now, the
formation of the nodule primordium is induced in the pericycle oppo-site to a protoxylem pole, and hyphae in infection threads grow fromthe prenodule cells to the nodule primordium and infect primordiumcells. b Intercellular infection. Frankia hyphae penetrate the rootepidermis between epidermal cells and colonize the root cortex inter-cellularly. Cell divisions are induced in the root pericycle opposite aprotoxylem pole, leading to the formation of a nodule primordium.Cells of the nodule primordium are infected by Frankia hyphae
The diversity of actinorhizal symbiosis 971

active in infection thread-containing cells, indicating theconservation of a transcription factor in both symbioses(Svistoonoff et al. 2003, 2004).
During intercellular infection described for actinorhizalRosales (Fig. 3b), no root hair deformation takes place, butFrankia hyphae penetrate the middle lamella between adja-cent cells of the root epidermis and progressively colonizethe intercellular spaces of the root cortex (Racette andTorrey 1989; Miller and Baker 1985; Liu and Berry1991a, b; Valverde and Wall 1999; Fig. 3b). The penetrationof the middle lamella between epidermal cells was ascribedto the pectolytic activity which has been shown for severalFrankia strains (Séguin and Lalonde 1989). However, anal-ysis of the secretome of three Frankia genomes revealedthat these strains’ secretome made up only 4–5% of the totalproteome, and they contained comparatively low numbersof polysaccharide-degrading enzymes (Mastronunzio et al.2008). Hence, the dissolution of the middle lamella might becaused by the localized release of enzymes by the host plant.During infection, epidermal and cortical sells secrete pectin-and protein-rich material into the intercellular spaces (Liuand Berry 1991b) which might represent the equivalent ofthe cell-wall-like material encapsulating the Frankia hyphaein infection threads. Concomitantly, the nodule lobe primor-dium is initiated in the root pericycle, and Frankia hyphaeinfect primordium cells from the intercellular spaces.
In most legume nodules, stable internal accommodationof rhizobia in infected cells is initiated by the release ofrhizobia from infection threads into infected cells in a
process resembling endocytosis (Bassett et al. 1977). Therhizobia become enclosed by a plant-derived membrane, theperibacteroid membrane (PBM), which, together with therhizobium and the space between PBM and bacterium,forms the symbiosome (Roth and Stacey 1989). Within thesymbiosomes, rhizobia differentiate into their symbioticform, the bacteroids. In legumes where no symbiosomeformation takes place (Naisbitt et al. 1992) and in Para-sponia sp. where no symbiosomes are formed as well,researchers distinguish between “infection threads” whichgrow from cell to cell and “fixation threads” whichbranch off from infection threads to fill the infected cellsand wherein the rhizobia develop into bacteroids andsymbiotic nitrogen fixation commences (Lancelle andTorrey 1984a,b).
In actinorhizal Fagales, the process of filling infectedcells with Frankia hyphae begins with infection threadlikestructures and spreads from the cell center outward(Schwintzer et al. 1982; Fig. 4a). It has been studied indetail by Berg (1999a, b) who coined the terms “invasivehyphae” and “vegetative hyphae.” A plant cell is invaded bybeing crossed by an infection thread containing an “invasivehypha” sheathed in the infection thread wall and surroundedby the invaginated plasma membrane of the infected cell.Then, “vegetative hyphae” enclosed in primary plant cellwall material (Berg 1990) proliferate by branching from thisinfection thread, filling the cell from the center outward.When the cell has been filled, N2-fixing vesicles differenti-ate from tips of the “vegetative hyphae,” and nitrogenase is
Fig. 4 Stable internal accommodation of Frankia hyphae in infectedcells. a A file of infected cells in the cortex of a C. glauca nodule.Parallel infection threads with “invasive hyphae” (arrows) grow fromcell to cell, while infection threads with “vegetative hyphae” branchfrom the cell center outward to fill the cell. Asterisk indicates prein-fection thread structure. b Longitudinal section of a D. glomeratanodule, the infection zone. Infection threads with “invasive hyphae”enter the cells from several directions; the cells are filled with branch-ing “vegetative hyphae” from the periphery inward, while the central
vacuole is maintained. c Infection threads (red) in infected cells of a D.glomerata nodule; the infected cells are undergoing simultaneousmitoses without cytoplasm division. Total DNA was stained withpropidium iodide visualized in the red channel. De novo synthesizedDNA was labeled for 24 h with 5-ethynyl-2′-deoxyuridine, visualizedusing an Alexa Fluor 488-labeled azide in the green channel. ininfected cortical cells, un uninfected cortical cells (often containingstarch grains). Size bars represent 20 μm (a, c) and 50 μm (b),respectively
972 K. Pawlowski, K.N. Demchenko

formed in these vesicles (Huss-Danell and Bergman 1990).Hence, “vegetative hyphae” in infection threads are theequivalent of legume “fixation threads”.
During infection in actinorhizal Rosales, the plasmamembrane of the infected cells invaginates and hyphaeentering from the apoplast become embedded in cell-wall-like material, i.e., infection threads are formed within theinfected cells (Berry and Sunell 1990; Liu and Berry 1991a;Gabbarini and Wall 2008). As in intracellularly infectedactinorhizal Fagales, branching of hyphae in infectionthreads to fill the infected cell occurs from the cell centeroutwards. However, the infection threads in intercellularlyinfected actinorhizal Rosales never display transcellulargrowth, indicating that they are the equivalents of infectionthreads with “vegetative hyphae” (i.e., of “fixation threads”in legumes). The equivalent of infection threads with “inva-sive hyphae” does not seem to exist in actinorhizal Rosales.
As mentioned above, the mechanism of nodule inductionon roots of actinorhizal Cucurbitales has not yet been stud-ied. However, detailed cytological studies of mature noduleshave been performed (Newcomb and Pankhurst 1982;Hafeez et al. 1984; Mirza et al. 1994; Berg et al. 1999).The absence of prenodules would lead to the assumptionthat in these symbioses, the intercellular infection pathwaytakes place. Nevertheless, infection threads show transcel-lular growth (Berg et al. 1999). Yet, in contrast with legumesand actinorhizal Fagales, in D. glomerata transcellular in-fection thread, growth was not preceded by the formation ofPITs (Berg et al. 1999). Furthermore, infection threads inCucurbitales do not grow through the cell center duringtranscellular growth but remain in the periphery of the cell.Cells are filled with branching infection threads from theperiphery inwards and a large central vacuole is retained,while in other actinorhizal plants, the central vacuole isfragmented during infection (Berg et al. 1999; Fig. 4b, c).In both A. glutinosa and in D. glomerata nodules, severalinfection threads with invasive hyphae enter a single cell;while in A. glutinosa, these infection threads grow in paral-lel (Berg 1999a, b; Fig. 4a), in D. glomerata, they usuallycome from different directions (Berg et al. 1999; Fig. 4b, c).In summary, the infection thread growth mechanism inactinorhizal Cucurbitales is clearly different from the infec-tion thread growth mechanism in actinorhizal Fagales and inlegumes.
Signal exchange between symbionts
With one exception (Giraud et al. 2007), host specificity,internalization of bacteria in plant roots and nodule devel-opment are regulated by lipochitooligosaccharide Nod fac-tors in legume/rhizobia symbioses which bind to LysM typereceptor kinases on the host plant plasma membrane. As
mentioned above, these lipochitooligosaccharide signal fac-tors were obviously copied from AM symbioses since thesignal factors from AM fungi (Myc factors) also representlipochitooligosaccharides and use the same signal transduc-tion pathway as Nod factors, the so-called common symbi-otic pathway (Maillet et al. 2011). The relationship betweenboth signaling processes is further emphasized by the factthat in Parasponia andersonii, rhizobial Nod and fungalMyc factors are perceived by the same plant LysM typereceptor kinase (Op den Camp et al. 2011). This receptortype is also responsible for binding chitooligosaccharidesderived from fungal cell walls that elicit defense responsesin the interactions between plants and fungal pathogens(Gough and Cullimore 2011). However, no convincinghomologs of the rhizobial “common nod genes”—nodABCwhich encode the enzymes responsible for the synthesis ofthe Nod factor backbone—were found in the first sequencedFrankia genomes (Normand et al. 2007). NodC proteinsform a specific subfamily of chitin synthases, while NodBproteins form a subfamily of deacetylases. NodA-type acyltransferases are, so far, specific to rhizobia. Chitin synthasesand deacetylases were found in the Frankia genomes, butthey did not group in the NodC and NodB subfamilies,respectively (Normand et al. 2007). Nevertheless, based onthe commonality of lipochitooligosaccharide signaling inarbuscular mycorrhiza and legume/rhizobia symbioses, itcan be expected that Frankia Nod factor equivalents arealso chitin-based and perceived by plant LysM receptorkinases.
As mentioned above, two rhizobial strains are known thatinduce root nodules on Aeschynomene sp. but lack thecommon nod genes and cannot form lipochitooligosacchar-ide Nod factors (Bonaldi et al. 2011). In their symbiosis,which is specific to Aeschynomene sp. (Giraud et al. 2007),no root hair curling or infection thread formation takesplace, but the bacteria enter the plant through preexistinggaps in the epidermis at the emergence of lateral roots. NoPIT or infection thread formation has been reported. It hasbeen suggested that in these symbioses, the induction ofcytokinin signaling by rhizobial Nod factors (reviewed byOldroyd and Downie 2008) is a short cut in that bacterialcytokinins constitute the first signals triggering nodulation(Bonaldi et al. 2010). Could Frankia use such a mechanism?The similarity of infection thread growth in legumes andactinorhizal Fagales would contradict such a hypothesis.
Using RNAi inhibition of the transcription of the homo-logues of the receptor kinase SymRK/DMI2, it was con-firmed for representatives of actinorhizal Fagales (C.glauca; Gherbi et al. 2008) and actinorhizal Cucurbitales(D. glomerata; Markmann et al. 2008) that Frankia signal-ing involves the common symbiotic pathway in that theremoval of this kinase abolished both AM and nodulationby Frankia in both systems. Furthermore, transcriptome
The diversity of actinorhizal symbiosis 973

sequencing has shown for two representatives of actino-rhizal Fagales that all known compounds of the commonsymbiotic pathway are expressed in roots and nodules (A.glutinosa, C. glauca; Hocher et al. 2011). Hence, in actino-rhizal symbioses like in legume/rhizobia and AM symbio-ses, the presence of chitin-based bacterial Nod factors can beexpected, but their structure has yet to be elucidated. Giventhat the chemicals in the culture supernatants of clade IFrankia strains that induce root hair deformation on theirhost plants do not share the chemical properties of lipochi-tooligosaccharides (Céremonie et al. 1999; van Ghelue et al.1997), it is possible that Nod factors from cluster I Frankiastrains represent chitooligosaccharides without a fatty acidat the reducing end, which would also explain the absenceof homologs of the “common nod genes” in the cluster IFrankia genomes sequenced thus far (strains ACN14a andCcI3; Normand et al. 2007). Given that it was shown for theCasuarina-infective strain CcI3 that root exudates of its hostplant C. glauca changed its surface properties with regard tofatty acids and carbohydrates, it would also be possible thatthe Nod factors of cluster I Frankia strains represent part ofthe their cell surface (Beauchemin et al. 2012).
Differentiation of infected cells
Infected cells of root nodules represent the only cell typethat can stably internally accommodate a eubacterial micro-symbiont (Parniske 2000). Root cells that existed before thecontact with the microsymbiont can support infection threadgrowth in that they can form PITs and be crossed by infec-tion threads both in legumes and in actinorhizal Fagales.However, in legumes and in all actinorhizal symbioses, onlycells that were formed after signal exchange between macro-and microsymbiont can be filled with symbiosomes or withbranching infection threads.
Studies on the model legume Medicago truncatula haveshown that Nod factor signaling is involved in the stableinternal accommodation of rhizobia in symbiosomes ininfected cells in so far as the removal of a component ofthe Nod factor signal transduction chain interfered withsymbiosome formation (Limpens et al. 2005a, b). This isconsistent with the facts that rhizobia in infection threadsproduce Nod factors (D'Haeze et al. 1998; Schlaman et al.1991; Sharma and Signer 1990; Timmers et al. 1998) andthat their receptors are expressed by the plant in the infec-tion zone of nodules (Arrighi et al. 2006). Studies on speciesof the woody legume genus Chamaecrista have shown thatthere is a continuum between stable internal accommodationof rhizobia in branching infection threads (“fixationthreads”) and in symbiosomes (Naisbitt et al. 1992). Fur-thermore, molecular studies on transcription factor conser-vation showed that the promoters of soybean leghemoglobin
and of the symbiotic hemoglobin from C. glauca are bothspecifically active in cells that stably accommodate themicrosymbiont. This is in spite of the fact that these cellsare part of different tissues in both symbioses and usedifferent mechanisms for the internal accommodation ofthe microsymbiont (Jacobsen-Lyon et al. 1995; Franche etal. 1998). Hence, it is very likely that at least in actinorhizalFagales, the internalization of bacteria in infected cellsby branching infection threads depends on Nod factorsignaling.
However, studies on the leguminous tree Sesbania ros-trata revealed that when rhizobia did not produce Nodfactors in infection threads, infection thread growth, butnot symbiosome development, was affected (Den Herderet al. 2007). Taking into account the two Nod factor-freesymbioses between Aeschynomene sp. and bradyrhizobia(Giraud et al. 2007), it is clear that like nodule organogen-esis, the stable accommodation of bacteria in infected cellscan also be induced by signaling processes that do notinvolve Nod factors. In this context, it should be mentionedthat the activity of the promoter of the legume marker geneENOD11, the symbiotic expression of which in the modellegumeM. truncatula is induced by (a) Nod factor signalingand (b) infection (Boisson-Dernier et al. 2005), was con-served in roots and nodules of the actinorhizal Fagales C.glauca and A. verticillata. However, here the Nod factor-dependent activation of the promoter before physical contactbetween the symbionts did not take place (Svistoonoffet al. 2010). Surprisingly, the induction of ENOD11::GUStranscription in arbuscule-containing cells, which in M.truncatula depended on the same promoter part as theinfection-dependent induction during nodulation, was miss-ing in the two actinorhizal Fagales. In the intercellularlyinfected Discaria trinervis (Rosales), the pattern ofENOD11 promoter activity during nodulation was evenmore divergent (Imanishi et al. 2011). Hence, the connectionbetween Nod/Myc factor signaling and infection mecha-nisms is complex.
The question of which factor enables the stable internal-ization of a eubacterial microsymbiont is one of the centralenigmas of root nodule symbioses. In this context, thesymbioses of actinorhizal Cucurbitales like D. glomerataare of particular interest, since here, the infected cells arespatially clearly separated from the uninfected cells(Fig. 2c). The lobes of their nodules do not show the lateralsymmetry typical for (modified) lateral roots in that cells onone side of the acentric stele cannot become infected. There-fore, in these symbioses, cells that will become infected can,to some extent, be distinguished from cells that cannotbecome infected. However, the only difference found sofar between both cell types was that only those cells thatare to become infected seem to express a gene encoding aglycine- and histidine-rich protein that is also expressed in
974 K. Pawlowski, K.N. Demchenko

the root and nodule periderm (Pawlowski et al. 2003) andwhich has a nodule-specific homologue in actinorhizalFagales (A. glutinosa; Pawlowski et al. 1997). However,there is reason to believe that the infection mechanism inactinorhizal Cucurbitales is different from that in actinorhizalFagales and Cucurbitales. Apart from the above-mentioneddifferences in infection thread growth, nodule cortical cells ofDatisca sp. become multinucleate in the course of being filledwith branching infection threads (Newcomb and Pankhurst1982; Hafeez et al. 1984; Berg et al. 1999; Fig. 4c), which isnot the case in Fagales, Rosales or legumes. However, endor-eduplication and ploidy-dependent cell enlargement has beenshown to be part of the differentiation of infected cells oflegume nodules (Vinardell et al. 2003). Altogether, the systemwhich offers a unique opportunity for the molecular andcytological analyses of the differentiation of infected cells,D. glomerata, also seems to have a unique infection threadgrowth mechanism.
Phytohormones in nodulation
The involvement of phytohormones in legume nodule in-duction has been well examined. The application of Nodfactors blocks auxin transport which leads to local auxinaccumulation (reviewed by Ferguson and Mathesius 2003)and also leads to local cytokinin accumulation (Murray et al.2007). Cytokinin signaling is essential for eliciting legumenodule organogenesis (reviewed by Oldroyd and Downie2008). However, while auxin levels are increased in noduleprimordia compared to roots, in mature nodules auxin levelsdecrease (Fedorova et al. 2000).
In actinorhizal symbioses, some Frankia strains havebeen shown to produce the auxin analogue, phenoxy aceticacid (PAA; Hammad et al. 2003; Perrine-Walker et al. 2010)as well as auxin itself (Perrine-Walker et al. 2010). Thedistribution of auxin exporters and importers in nodulesleads to further accumulation of auxin in infected cells(Péret et al. 2007), and auxin levels are strongly enhancedin mature nodules compared to roots. However, no evidencefor involvement of cytokinin signaling in nodule inductionhas been shown thus far. A homologue of the histidinekinase involved in nodule induction (LHK1/CRE1; Murrayet al. 2007) was found to be expressed in roots and nodulesof C. glauca but not in A. glutinosa (Hocher et al. 2011).Hence, phytohormone effects seem to differ in both symbi-oses with regard to auxin which seems to play a larger rolein actinorhizal than in legume symbioses. This is not sur-prising in that the hormone balance in legume roots seems todiffer from that in other plants. Only in legumes can rootnodule primordia be induced in the root cortex instead of inthe pericycle like lateral root primordia. So, it would not besurprising if the phytohormone involvement in the induction
of nodules on nonlegumes were different from that onlegumes and that auxin would play a major role in inducingthe formation of an organ primordium in the root pericycle.Previously, it has been suggested that in inducing legumenodules, rhizobia might have exploited a program evolved forthe development of root storage organs (Joshi et al. 1993). Aninvolvement of cytokinin in the induction of storage organswould, therefore, not be surprising (Arnholdt-Schmitt 1999).
It is interesting that legumes as well as Betulaceae, Myr-icaceae, Casuarinaceae, and Elaeagnaceae can form clusterroots, which are defined as a densely packed group ofdeterminate rootlets which develop, grow, cease growth,undergo an exudative burst and absorb phosphate in a syn-chronous way. Cluster roots exude large quantities of malateand citrate during phosphate deficiency, increasing theavailability of mineral-bound phosphate (Gilbert et al.2000). Thus, the ability to form cluster roots and the abilityto form root nodules are often correlated (Skene 1998;Shane and Lambers 2005). Natural and synthetic auxins caninduce some aspects of cluster root development (Laskowskiet al. 1995; Gilbert et al. 2000), and the importance of auxinfor inducible root responses to nutrient stress has been estab-lished (Landsberg 1996). In other words, the roots of legumesand four out of eight actinorhizal families have a pericycle thatcan easily be stimulated in response to nutrient stress and canform rootlets that exudate malate.
Malate is the carbon source provided by legumes (Oldroydet al. 2011) and at least one actinorhizal species (A. gluti-nosa; Jeong et al. 2004) to their respective microsymbionts,rhizobia and Frankia, in infected cells. Intracellular bacteriaare separated from the cytosol by a derivative of the plantplasma membrane (rhizobia) or the invaginated plant plasmamembrane (Frankia) and, therefore, topologically located inthe apoplast. Hence, it is likely that the same dicarboxylateexporter is active in infected cells of nodules and in clusterroots. Given that not all plant families that form cluster rootscan also form root nodules (e.g., Proteaceae and Moraceaecannot) and that cluster roots have not been reported for fouractinorhizal families including all actinorhizal Cucurbitales, itis intriguing to speculate which mechanism evolved first:cluster root formation or nodulation. This question might beapproached via the analysis of the evolution of plasma mem-brane dicarboxylate exporters. They have to be involved indelivering carbon sources to intracellular nitrogen-fixing bac-teria and presumably also in exuding dicarboxylates into therhizosphere. Thus far, only one such transporter has beencharacterized (Jeong et al. 2004).
Summary
Independent evolutionary origins have been proposed forthe actinorhizal symbioses in the three phylogenetic groups
The diversity of actinorhizal symbiosis 975

of its host plants (Swensen 1996). This is supported by theinfection mechanisms which are consistent within the threegroups. Many parallels exist between legume/rhizobia sym-bioses and actinorhizal Fagales, but parallels betweenlegumes and actinorhizal Rosales or Cucurbitales are lessobvious. This is interesting in that the newest phylogeneticstudies showed Fabales as a sister group to (Rosales,(Fagales and Cucurbitales)), i.e., the similarities betweenlegumes and actinorhizal Fagales cannot be explained bytheir phylogenetic position (Wang et al. 2009). Even inactinorhizal Fagales, phytohormone effects on nodule for-mation seem to differ from those in legumes, consistent withthe differences in nodule structure in both systems. Actino-rhizal Cucurbitales offer a unique opportunity to examinethe differentiation of cells that can stably accommodate aeubacterial microsymbiont; however, it seems that at leasttheir infection thread growth mechanism is different fromthat established in legumes and in actinorhizal Fagales. Thefact that the mechanism of stably internalizing microsym-biont involves multinucleation might be correlated with theendoreduplication of infected cells in legume nodules.
Acknowledgments We thank the Swedish Research councils VRand FORMAS and the Russian Foundation for Basic Research (11-04-02022) as well as the Ministry of Education and Sciences of theRussian Federation (GC # 16.552.11.7047, 14.740.11.1226) for theirsupport. We are indebted to Dr. Maria A. Osipova for her help withpreparing Fig. 3.
References
Arnholdt-Schmitt B (1999) On the physiology of yield production incarrots.—Implications for breeding towards nutrient efficiency.Gartenbauwissenschaft 64:26–32
Arrighi J-F, Barre A, Ben Amor B, Bersoult A, Campos Soriano L,Mirabella R, de Carvalho-Niebel F, Journet E-P, Ghérardi M,Huguet T, Geurts R, Dénarié J, Rougé P, Gough C (2006) TheMedicago truncatula lysine motif-receptor-like kinase gene fam-ily includes NFP and new nodule-expressed genes. Plant Physiol142:265–27
Bassett B, Goodman RN, Novacky A (1977) Ultrastructure of soybeannodules. I: release of rhizobia from the infection thread. Can JMicrobiol 23:573–582
Beauchemin NJ, Furnholm T, Lavenus J, Svistoonoff S, Doumas P,Bogusz D, Laplaze L, Tisa LS (2012) Casuarina root exudatesalter the physiology, surface properties, and plant infectivity ofFrankia sp. strain CcI3. Appl Environ Microbiol 78:575–580
Benson DR, Clawson ML (2000) Evolution of the actinorhizal plantsymbiosis. In: Prokaryotic nitrogen fixation: a model system foranalysis of a biological process. Horizon Scientific Press,Symondham, UK, pp 207-224
Benson DR, Silvester WB (1993) Biology of Frankia strains, actino-mycete symbionts of actinorhizal plants. Microbiol Rev 57:293–319
Berg RH (1990) Cellulose and xylans in the interface capsule insymbiotic cells of actinorhizae. Protoplasma 159:35–43
Berg RH (1999a) Cytoplasmic bridge formation in the nodule apex ofactinorhizal root nodules. Can J Bot 77:1351–1357
Berg RH (1999b) Frankia forms infection threads. Can J Bot 77:1327–1333
Berg RH, Langenstein B, Silvester WB (1999) Development in theDatisca-Coriaria nodule type. Can J Bot 77:1334–1350
Bergman B, Johansson C, Söderbäck E (1992) The Nostoc–Gunnerasymbiosis. New Phytol 122:379–400
Berry AM, Sunell LA (1990) The infection process and nodule devel-opment. In: Schwintzer CR, Tjepkema JD (eds) The biology ofFrankia and actinorhizal plants. Academic Press, San Diego, pp61–81
Berry AM, Torrey JG (1983) Root hair deformation in the infectionprocess of Alnus rubra. Can J Bot 61:2863–2876
Berry AM, McIntyre L, McCully ME (1986) Fine structure of root hairinfection leading to nodulation in the Frankia–Alnus symbiosis.Can J Bot 64:292–305
Berry AM, Harriott OT, Moreau RA, Osman SF, Benson DR, JonesAD (1993) Hopanoid lipids compose the Frankia vesicle enve-lope, presumptive barrier of oxygen diffusion to nitrogenase. ProcNatl Acad Sci USA 90:6091–6094
Berry AM, Mendoza-Herrera A, Guo Y-Y, Hayashi J, Persson T,Barabote R, Demchenko K, Zhang S, Pawlowski K (2011) Newperspectives on nodule nitrogen assimilation in actinorhizal sym-bioses. Funct Plant Biol 38:645–652
Boisson-Dernier A, Andriankaja A, Chabaud M, Niebel A, Journet EP,Barker DG, de Carvalho-Niebel F (2005) MtENOD11 gene acti-vation during rhizobial infection and mycorrhizal arbuscule de-velopment requires a common AT-rich-containing regulatorysequence. Mol Plant Microbe Interact 18:1269–1276
Bonaldi K, Gourion B, Fardoux J, Hannibal L, Cartieaux F, Boursot M,Vallenet D, Chaintreuil C, Prin Y, Nouwen N, Giraud E (2010)Large-scale transposon mutagenesis of photosynthetic Bradyrhi-zobium sp. strain ORS278 reveals new genetic loci putativelyimportant for nod-independent symbiosis with Aeschynomeneindica. Mol Plant Microbe Interact 23:760–770
Bonaldi K, Gargani D, Prin Y, Fardoux J, Gully D, Nouwen N,Goormachtig S, Giraud E (2011) Nodulation of Aeschynomeneafraspera and A. indica by photosynthetic Bradyrhizobium sp.strain ORS285: the nod-dependent versus the nod-independentsymbiotic interaction. Mol Plant Microbe Interact 24:1359–1371
Callaham D, Torrey JG (1977) Prenodulae formation and primarynodule development in roots of Comptonia (Myricaceae). Can JBot 51:2306–2318
Céremonie H, Debelle F, Fernandez MP (1999) Structural and func-tional comparison of Frankia root hair deforming factor andrhizobia Nod factor. Can J Bot 77:1293–1301
Chen WM, Moulin L, Bontemps C, Vandamme P, Béna G,Boivin-Masson C (2003) Legume symbiotic nitrogen fixationby beta-proteobacteria is widespread in nature. J Bacteriol185:7266–7272
Demchenko KN, Demchenko NP (2001) Changes of root structure inconnection with the development of lateral root primordia inwheat and pumpkins. In: Recent advances of plant root structureand function: proceedings of the 5th International Symposium onStructure and Function of Roots; Stara Lesna, Slovakia, Slovakia,1998. Gasparikova O, Ciamporova M, Mistrik I, Baluska F (eds),Developments in plant and soil sciences, Kluwer. Vol 90: 39–47
Den Herder J, Vanhee C, De Rycke R, Corich V, Holsters M,Goormachtig S (2007) Nod factor perception during infectionthread growth fine-tunes nodulation. Mol Plant Microbe In-teract 20:129–137
D’Haeze W, Gao M, De Rycke R, Van Montagu M, Engler G, HolstersM (1998) Roles for azorhizobial Nod factors and surface poly-saccharides in intercellular invasion and nodule penetration, re-spectively. Mol Plant-Microbe Interact 11:999–1008
Doyle JJ (2011) Phylogenetic perspectives on the origins of nodula-tion. Mol Plant Microbe Interact 24:1289–1295
976 K. Pawlowski, K.N. Demchenko

Duhoux E, Rinaudo G, Diem HG, Auguy F, Fernandez D, Bogusz D,Franche C, Dommergues Y, Huguenin B (2001) AngiospermGymnostoma trees produce root nodules colonized by arbuscularmycorrhizal fungi related to Glomus. New Phytol 149:115–125
Fedorova EE, Zhiznevskaya GY, Kalibernaya ZV, Artemenko EN,Izmailov SF, Gus’kov AV (2000) IAA metabolism during devel-opment of symbiosis between Phaseolus vulgaris and Rhizobiumphaseoli. Russ J Plant Physiol 47:203–206
Ferguson BJ, Mathesius U (2003) Signaling interactions during noduledevelopment. J Plant Growth Regul 22:47–72
Franche C, Diouf D, Laplaze L, Auguy F, Frutz T, Rio M, Duhoux E,Bogusz D (1998) Soybean (lbc3), Parasponia, and Trema hemo-globin gene promoters retain symbiotic and nonsymbiotic speci-ficity in transgenic Casuarinaceae: implications for hemoglobingene evolution and root nodule symbioses. Mol Plant-MicrobeInteract 11:887–894
Gabbarini LA, Wall LG (2008) Analysis of nodulation kinetics inFrankia-Discaria trinervis symbiosis reveals different factors in-volved in the nodulation process. Physiol Plant 133:776–785
Gherbi H, Markmann K, Svistoonoff S, Estevan J, Autran D, GiczeyG, Auguy F, Péret B, Laplaze L, Franche C, Parniske M, BoguszD (2008) SymRK defines a common genetic basis for plant rootendosymbioses with arbuscular mycorrhiza fungi, rhizobia, andFrankia bacteria. Proc Natl Acad Sci USA 105:4928–4932
Gilbert GA, Knight JD, Vance CP, Allan DL (2000) Proteoid rootdevelopment of phosphorus deficient lupin is mimicked by auxinand phosphonate. Ann Bot 85:921–928
Giraud E, Moulin L, Vallenet D, Barbe V, Cytryn E, Avarre JC, JaubertM, Simon D, Cartieaux F, Prin Y, Bena G, Hannibal L, Fardoux J,Kojadinovic M, Vuillet L, Lajus A, Cruveiller S, Rouy Z,Mangenot S, Segurens B, Dossat C, Franck WL, Chang WS,Saunders E, Bruce D, Richardson P, Normand P, Dreyfus B,Pignol D, Stacey G, Emerich D, Verméglio A, Médigue C,Sadowsky M (2007) Legumes symbioses: absence of nod genesin photosynthetic bradyrhizobia. Science 316:1307–1312
Gough C, Cullimore J (2011) Lipo-chitooligosaccharide signaling inendosymbiotic plant–microbe interactions. Mol Plant MicrobeInteract 24:867–878
Hafeez F, Akkermans ADL, Chaudhary AH (1984) Observations onthe ultrastructure of Frankia sp. in root nodules of Datisca can-nabina L. Plant Soil 79:383–402
Hammad Y, Nalin R, Marechal J, Fiasson K, Pepin R, Berry AM,Normand P, Domenach A-M (2003) A possible role for phenylacetic acid (PAA) on Alnus glutinosa nodulation by Frankia.Plant Soil 254:193–205
Heidstra R, Geurts R, Franssen H, Spaink HP, Kammen A, van BisselingT (1994) Root hair deformation activity of nodulation factors andtheir fate on Vicia sativa. Plant Physiol 105:787–797
Hocher V, Alloisio N, Auguy F, Fournier P, Doumas P, Pujic P, GherbiH, Queiroux C, Da Silva C, Wincker P, Normand P, Bogusz D(2011) Transcriptomics of actinorhizal symbioses reveals homo-logs of the whole common symbiotic signaling cascade. PlantPhysiol 156:700–711
Huss-Danell K (1997) Tansley Review No. 93. Actinorhizal symbiosesand their N2 fixation. New Phytol 136:375–405
Huss-Danell K, Bergman B (1990) Nitrogenase in Frankia from rootnodules of Alnus incana (L.) Moench: immunolocalization of theFe− and MoFe proteins during vesicle differentiation. New Phytol116:443–455
Imanishi L, Vayssières A, Franche C, Bogusz D, Wall L, Svistoonoff S(2011) Transformed hairy roots of Discaria trinervis: a valuabletool for studying actinorhizal symbiosis in the context of intercel-lular infection. Mol Plant Microbe Interact 24:1317–1324
Jacobsen-Lyon K, Østergaard-Jensen E, Jørgensen J-E, Marcker KA,Peacock J, Dennis E (1995) Symbiotic and nonsymbiotic hemo-globin genes of Casuarina glauca. Plant Cell 7:213–223
Jeong J, Suh SJ, Guan C, Tsay Y-F, Moran N, Oh CJ, An CS,Demchenko K, Pawlowski K, Lee Y (2004) A nodule-specificdicarboxylate transporter from Alnus glutinosa. Plant Physiol134:969–978
Joshi PA, Caetano-Anollés G, Graham ET, Gresshoff PM (1993)Ultrastructure of transfer cells in spontaneous nodules of alfalfa(Medicago sativa). Protoplasma 172:64–76
Knowlton S, Berry A, Torrey JG (1980) Evidence that associated soilbacteria may influence root hair infection of actinorhizal plants byFrankia. Can J Microbiol 26:971–977
Lancelle SA, Torrey JG (1984a) Early development of Rhizobium-induced root nodules of Parasponia rigida. I. Infection and earlynodule initiation. Protoplasma 123:26–37
Lancelle SA, Torrey JG (1984b) Early development of Rhizobium-induced root nodules of Parasponia rigida. II. Nodule morpho-genesis and symbiotic development. Can J Bot 63:25–35
Landsberg EC (1996) Hormonal regulation of iron–stress response insunflower roots: a morphological and cytological investigation.Protoplasma 194:69–80
Laplaze L, Duhoux E, Franche C, Frutz T, Svistoonoff S, Bisseling T,Bogusz D, Pawlowski K (2000) Casuarina glauca prenodulecells display the same differentiation as the corresponding nodulecells. Mol Plant-Microbe Interact 13:107–112
Laskowski MJ, Williams ME, Nusbaum HC, Sussex IM (1995) For-mation of lateral root meristems is a two stage process. Dev121:3303–3310
Limpens E, Mirabella R, Fedorova E, Franken C, Franssen H, BisselingT, Geurts R (2005a) Formation of organelle-like N2-fixing symbio-somes in legume root nodules is controlled by DMI2. Proc NatlAcad Sci USA 102:10375–10380
Limpens E, Mirabella R, Fedorova E, Franken C, Franssen H, BisselingT, Geurts R (2005b) Formation of organelle-like N2-fixing symbio-somes in legume root nodules is controlled by DMI2. Proc NatlAcad Sci USA 102:10375–10380
Liu Q, Berry AM (1991a) The infection process and nodule initiationin the Frankia-Ceanothus root nodule symbiosis: a structural andhistochemical study. Protoplasma 163:82–92
Liu Q, Berry AM (1991b) Localization and characterization of pecticpolysaccharides in roots and root nodules of Ceanothus spp.during intercellular infection by Frankia. Protoplasma 164:93–101
Maillet F, Poinsot V, André O, Puech-Pagès V, Haouy A, Gueunier M,Cromer L, Giraudet D, Formey D, Niebel A, Martinez EA,Driguez H, Bécard G, Dénarié J (2011) Fungal lipochitooligosac-charide symbiotic signals in arbuscular mycorrhiza. Nature469:58–63
Markmann K, Giczey G, Parniske M (2008) Functional adaptation of aplant receptor-kinase paved the way for the evolution of intracel-lular root symbioses with bacteria. PLoS Biol 6:e68
Markmann K, Parniske M (2009) Evolution of root endosymbiosiswith bacteria: how novel are nodules? Trends Plant Sci 14:77–86
Mastronunzio JE, Tisa LS, Normand P, Benson DR (2008) Compara-tive secretome analysis suggests low plant cell wall degradingcapacity in Frankia symbionts. BMC Genom 9:47
Miller IM, Baker DD (1985) The initiation development and structureof root nodules in Elaeagnus angustifolia (Elaeagnaceae). Proto-plasma 128:107–119
Mirza S, Pawlowski K, Hafeez FY, Chaudhary AH, Akkermans ADL(1994) Ultrastructure of the endophyte and localization of nifHtranscripts in root nodules of Coriaria nepalensis Wall. by in situhybridization. New Phytol 126:131–136
Moran NA (2003) Tracing the evolution of gene loss in obligatebacterial symbionts. Curr Opin Microbiol 6:512–518
Murray JD, Karas BJ, Sato S, Tabata S, Amyot L, Szczyglowski K(2007) A cytokinin perception mutant colonized by Rhizobium inthe absence of nodule organogenesis. Science 315:101–104
The diversity of actinorhizal symbiosis 977

Naisbitt T, James EK, Sprent JI (1992) The evolutionary significanceof the legume genus Chamaecrista, as determined by nodulestructure. New Phytol 122:487–492
Newcomb W, Pankhurst CE (1982) Fine structure of actinorhizalnodules of Coriaria arborea (Coriariaceae). N Zeal J Bot20:93–103
Newcomb WR, Wood S (1987) Morphogenesis and fine structure ofFrankia (Actinomycetales): the microsymbiont of nitrogen-fixingactinorhizal root nodules. Int Rev Cytol 109:1–88
Normand P, Orso S, Cournoyer B, Jeannin P, Chapelon C, Dawson J,Evtushenko L, Misra A (1996) Molecular phylogeny of the genusFrankia and related genera and emendation of the family Frank-iaceae. Internat J Syst Bacteriol 46:1–9
Normand P, Lapierre P, Tisa LS, Gogarten JP, Alloisio N, Bagnarol E,Bassi CA, Berry AM, Bickhart DM, Choisne N, Couloux A,Cournoyer B, Cruveiller S, Daubin V, Demange N, FrancinoMP, Goltsman E, Huang Y, Kopp OR, Labarre L, Lapidus A,Lavire C, Marechal J, Martinez M, Mastronunzio JE, Mullin BC,Niemann J, Pujic P, Rawnsley T, Rouy Z, Schenowitz C, SellstedtA, Tavares F, Tomkins JP, Vallenet D, Valverde C, Wall LG, WangY, Medigue C, Benson DR (2007) Genome characteristics offacultatively symbiotic Frankia sp. strains reflect host range andhost plant biogeography. Genome Res 17:7–15
Oldroyd GE, Downie JA (2008) Coordinating nodule morphogenesiswith rhizobial infection in legumes. Annu Rev Plant Biol 59:519–546
Oldroyd GE, Murray JD, Poole PS, Downie JA (2011) The rules ofengagement in the legume–rhizobial symbiosis. Annu Rev Genet45:119–144
Oono R, Schmitt I, Sprent JI, Denison RF (2010) Multiple evolution-ary origins of legume traits leading to extreme rhizobial differen-tiation. New Phytol 187:508–520
Op den Camp R, Streng A, De Mita S, Cao Q, Polone E, Liu W,Ammiraju JS, Kudrna D, Wing R, Untergasser A, Bisseling T,Geurts R (2011) LysM-type mycorrhizal receptor recruited for rhi-zobium symbiosis in nonlegume Parasponia. Science 331:909–912
Parniske M (2000) Intracellular accommodation of microbes by plants:a common developmental program for symbiosis and disease?Curr Opin Plant Biol 3:320–328
Parniske M (2008) Arbuscular mycorrhiza: the mother of plant rootendosymbioses. Nat Rev Microbiol 6:763–775
Parsons R, Silvester WB, Harris S, Gruijters WTM, Bullivant S (1987)Frankia vesicles provide inducible and absolute oxygen protec-tion for nitrogenase. Plant Physiol 83:728–731
Pawlowski K (2002) Actinorhizal symbioses. In: Leigh GJ (ed) Nitro-gen fixation at the millennium. Elsevier, Amsterdam
Pawlowski K, Twigg P, Dobritsa S, Guan C,Mullin BC (1997) A nodule-specific gene family from Alnus glutinosa encodes glycine- andhistidine-rich proteins expressed in the early stages of actinorhizalnodule development. Mol Plant-Microbe Interact 10:656–664
Pawlowski K, Swensen S, Guan C, Hadri A-E, Berry AM, Bisseling T(2003) Distinct patterns of symbiosis-related gene expression inactinorhizal nodules from different plant families. Mol Plant-Microbe Interact 16:796–807
Pawlowski K, Jacobsen KR, Alloisio N, Ford Denison R, Klein M,Tjepkema JD, Winzer T, Sirrenberg A, Guan C, Berry AM (2007)Truncated hemoglobins in actinorhizal nodules of Datisca glom-erata. Plant Biol 9:776–785
Péret B, Swarup R, Jansen L, Devos G, Auguy F, Collin M, Santi C,Hocher V, Franche C, Bogusz D, Bennett M, Laplaze L (2007)Auxin influx activity is associated with Frankia infection duringactinorhizal nodule formation in Casuarina glauca. Plant Physiol144:1852–1862
Perrine-Walker F, Doumas P, Lucas M, Vaissayre V, Beauchemin NJ,Band LR, Chopard J, Crabos A, Conejero G, Péret B, King JR,Verdeil JL, Hocher V, Franche C, Bennett MJ, Tisa LS, Laplaze L
(2010) Auxin carriers localization drives auxin accumulation inplant cells infected by Frankia in Casuarina glauca actinorhizalnodules. Plant Physiol 154:1372–1380
Persson T, Benson DR, Normand P, Vanden Heuvel B, Pujic P, ChertkovO, Teshima H, Bruce BC, Detter C, Tapia R, Han S, Han J, WoykeT, Pitluck S, Pennacchio L, Nolan M, Ivanova N, Pati A, Land ML,Pawlowski K, Berry AM (2011) The genome ofCandidatus Frankiadatiscae Dg1, the uncultured microsymbiont from nitrogen-fixingroot nodules of the dicot Datisca glomerata. J Bacteriol 193:7017–7018
Racette S, Torrey JG (1989) Root nodule initiation in Gymnostoma(Casuarinaceae) and Shephardia (Elaeagnaceae) induced byFrankia strain HFPGpI1. Can J Bot 67:2873–2879
Ramirez-Saad H, Janse JD, Akkermans ADL (1998) Root nodules ofCeanothus caeruleus contain both the N2-fixing Frankia endo-phyte and a phylogenetically related Nod-/Fix- actinomycete. CanJ Bot 44:140–148
Roth LE, Stacey G (1989) Bacterium release into host cells of nitrogen-fixing soybean nodules: the symbiosome membrane comes fromthree sources. Eur J Cell Biol 49:13–23
Schlaman HRM, Horvath B, Vijgenboom E, Okker RJH, LugtenbergBJJ (1991) Suppression of nodulation gene expression in bacte-roids of Rhizobium leguminosarum biovar viciae. J Bacteriol173:4277–4287
Schröder P (1989) Aeration of the root system in Alnus glutinosa L.Gaertn. Annales des Sciences Forestières 46:310–314
Schubert M, Koteyeva NK, Wabnitz PW, Santos P, Büttner M, SauerN, Demchenko K, Pawlowski K (2011) Plasmodesmata distribu-tion and sugar partitioning in nitrogen-fixing root nodules ofDatisca glomerata. Planta 233:139–152
Schwintzer CR, Berry AM, Disney LD (1982) Seasonal patterns ofroot nodule growth, endophyte morphology, nitrogenase activityand shoot development in Myrica gale. Can J Bot 60:746–757
Séguin A, Lalonde M (1989) Detection of pectolytic activity and pelhomologous sequences in Frankia. Plant Soil 118:221–229
Shah VK, Brill WJ (1977) Isolation of an iron–molybdenum cofactorfrom nitrogenase. Proc Natl Acad Sci USA 74:3249–3253
Shane MW, Lambers H (2005) Cluster roots: a curiosity in context.Plant Soil 274:101–125
Sharma SB, Signer ER (1990) Temporal and spatial regulation of thesymbiotic genes of Rhizobium meliloti in planta revealed bytransposon Tn5-gusA. Genes Dev 4:344–356
Silvester WB, Harris SL, Tjepkema JD (1990) Oxygen regulation andhemoglobin. In: Schwintzer CR, Tjepkema JD (eds) The biologyof Frankia and actinorhizal plants. Academic Press, San Diego,pp 157–176
Silvester WB, Langenstein B, Berg RH (1999) Do mitochondria pro-vide the oxygen diffusion barrier in root nodules of Coriaria andDatisca? Can J Bot 77:1358–1366
Skene KR (1998) Cluster roots: some ecological considerations. J Ecol86:1060–1064
Soltis DE, Soltis PS, Morgan DR, Swensen SM, Mullin BC, Dowd JM,Martin PG (1995) Chloroplast gene sequence data suggest asingle origin of the predisposition for symbiotic nitrogen fixationin angiosperms. Proc Natl Acad Sci USA 92:2647–2651
Svistoonoff S, Laplaze L, Auguy F, Runions J, Duponnois R, HaseloffJ, Franche C, Bogusz D (2003) cg12 expression is specificallylinked to infection of root hairs and cortical cells during Casua-rina glauca and Allocasuarina verticillata actinorhizal noduledevelopment. Mol Plant Microbe Interact 16:600–607
Svistoonoff S, Laplaze L, Liang J, Ribeiro A, Gouveia MC, Auguy F,Fevereiro P, FrancheC, Bogusz D (2004) Infection-related activationof the cg12 promoter is conserved between actinorhizal and legume–rhizobia root nodule symbiosis. Plant Physiol 136:3191–3197
Svistoonoff S, Sy MO, Diagne N, Barker DG, Bogusz D, Franche C(2010) Infection-specific activation of the Medicago truncatula
978 K. Pawlowski, K.N. Demchenko

Enod11 early nodulin gene promoter during actinorhizal rootnodulation. Mol Plant Microbe Interact 23:740–747
Swensen SM (1996) The evolution of actinorhizal symbioses—evidence for multiple origins of the symbiotic association.Am J Bot 83:1503–1512
Timmers ACJ, Auriac M-C, de Billy F, Truchet G (1998) Nod factorinternalization and microtubular cytoskeleton changes occur con-comitantly during nodule differentiation in alfalfa. Development125:339–349
Torrey JG (1976) Initiation and development of root nodules of Casu-arina (Casuarinaceae). Am J Bot 63:335–344
Valverde C, Wall LG (1999) Time course of nodule development in theDiscaria trinervis (Rhamnaceae) Frankia symbiosis. New Phytol141:345–354
van Brussel AAN, Bakhuizen R, van Spronsen PC, Spaink HP, Tak T,Lugtenberg BJ, Kijne JW (1992) Induction of pre-infection thread
structures in the leguminous host plant by mitogenic lipo-oligosaccharides of Rhizobium. Science 257:70–72
van Ghelue M, Lovaas E, Ringo E, Solheim B (1997) Early interactionsbetween Alnus glutinosa and Frankia strain ArI3—production andspecificity of root hair deformation factor(s). Physiol Plant 99:579–587
Vinardell JM, Fedorova E, Cebolla A, Kevei Z, Horvath G, KelemenZ, Tarayre S, Roudier F, Mergaert P, Kondorosi A, Kondorosi E(2003) Endoreduplication mediated by the anaphase-promotingcomplex activator CCS52A is required for symbiotic cell differ-entiation in Medicago truncatula nodules. Plant Cell 15:2093–2105
Wang H, Moore MJ, Soltis PS, Bell CD, Brockington SF, Alexandre R,Davis CC, Latvis M, Manchester SR, Soltis DE (2009) Rosidradiation and the rapid rise of angiosperm-dominated forests. ProcNatl Acad Sci USA 106:3853–3858
The diversity of actinorhizal symbiosis 979




















