
Rhizobial and actinorhizal symbiosis: What are the shared features?
16
The Plant Cell, Vol. 8, 1899-1913, October 1996 0 1996 American Society of Plant Physiologists Rhizobial and Actinorhizal Symbioses: What Are the Shared Features? Katharina Pawlowski and Ton Bisseling' Department of Molecular Biology, Dreijenlaan 3, Agricultura1 University, 6703 HA Wageningen, The Netherlands INTRODUCTION NODULE FORMATION In endophytic symbioses, microorganisms live intercellularly or intracellularly inside a plant, in a mutually beneficia1 rela- tionship. The host plant (the macrosymbiont) usually supplies the endophyte (the microsymbiont) with photosynthates, whereas the benefit of these associations for the plant is an improvement in access to important mineral nutrients whose supply would otherwise be growth limiting. With the excep- tion of cyanobacterial symbioses, in most endophytic symbioses involving higher plants, microorganisms colonize the plant root (reviewed in Quispel, 1992). In mycorrhizal sym- bioses, the fungus improves the absorption of minerals, particularly phosphorus, by the roots (Read et al., 1992; Harrison and van Buuren, 1995; see also Gianinazzi-Pearson, 1996, in this issue). In root nodule symbioses involving eubac- teria of the family Rhizobiaceae or actinomycetes of the genus frankia, the microsymbiont supplies the plant host with am- monium, the product of biological nitrogen fixation. To provide an ecological niche for the endophyte, the mac- rosymbiont has to undergo symbiosis-specific differentiation. In nitrogen-fixing root nodule symbioses, the microsymbionts are hosted in special organs, the root nodules, which develop on roots upon infection by the microsymbiont (reviewed in Benson and Silvester, 1993; Myiona et al., 1995). In root sym- bioses with fungi, plant differentiation is less dramatic. In ectomycorrhizal symbioses, for example, the fungus somewhat alters root growth, triggering the formation of short thick infected roots (Martin and Hilbert, 1991), whereas in endo- mycorrhizal symbioses, the plant root structure is altered at the cellular leve1 but not macroscopically (Koide, 1991; Gianinazzi-Pearson, 1996, in this issue). In this review, we discuss the recent progress in research on endophytic symbioses involving plant roots. Special atten- tion is given to nitrogen-fixing symbiotic interactions, and we summarize our current knowledge of nodule formation and function. The last part of this review focuses on the develop- ment of model systems and the relationships between different endophytic symbioses involving plant roots. To whom correspondence should be addressed The two microsymbiontsable to enter into root nodule symbi- oses with higher plants are quite different. Rhizobia are unicellular, Gram-negativebacteria, whereas Frankia strains are filamentous, branching, Gram-positivebacteria. Rhizobia almost exclusively enter symbioses with legumes (for reviews, see Sánchez et al., 1991; Hirsch, 1992; Mylona et ai., 1995; Long, 1996, in this issue). The legume family has three subfam- ilies, and although >90°/o of the Papilionoideae and Mimosoideae are capable of entering a symbiosis with rhizo- bia, only members of one genus of the third subfamily, the Caesalpinioideae, can doso (Doyle, 1994).Conversely, frankia strains can interact with a diverse group of dicotyledonous plants that are members of eight different families, collectively called actinorhizal plants (Benson and Silvester, 1993). 60th rhizobia and Frankia strains can produce the enzyme nitrogenase that catalyzes the reduction of dinitrogen. Nitrogenase structure is conserved among the few prokary- otes capable of producing it; the enzyme is highly sensitive to oxygen (reviewed in Burris, 1991). In symbiosis as well as under nitrogen-limiting conditions in the free-living state, frankia can form nitrogen-fixing vesicles at the ends of its hyphae (reviewed in Benson and Silvester, 1993). In thesevesi- cles, bacterial nitrogenase is protected from oxygen by a multilayered envelope consisting of hopanoids, which are bac- teria1 steroid lipids (Parsons et ai., 1987; Berry et al., 1993). In contrast, rhizobia cannot provide oxygen protection for nitrogenasethemselves and, with afew exceptions (see, e.g., de Bruijn, 1989), are capable of living on dinitrogen Only in the symbiotic state. Usually, the development of a symbiotic interaction involves some differentiation of the endophyte. This differentiation may be minimal, like the increase in heterocyst formation described for endophytic cyanobacteria(Bergman et al., 1992), but it is often quite pronounced. In most legume nodules, intracellu- lar rhizobia differentiate into nitrogen-fixingbacteroids. This process involves a marked increase in cell size, a change in morphology from rod-shaped to branched cells (Bergersen, 1974; Newcomb, 1976), and changes in surface antigens (Olsen et al., 1992; Roest et al., 1995). Symbiosis-specific differenti- ation of Frankia depends on the host plant species, which determines the morphology of the vesicles formed in most
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Rhizobial and actinorhizal symbiosis: What are the shared features?

The Plant Cell, Vol. 8, 1899-1913, October 1996 0 1996 American Society of Plant Physiologists
Rhizobial and Actinorhizal Symbioses: What Are the Shared Features?
Katharina Pawlowski and Ton Bisseling' Department of Molecular Biology, Dreijenlaan 3, Agricultura1 University, 6703 HA Wageningen, The Netherlands
INTRODUCTION NODULE FORMATION
In endophytic symbioses, microorganisms live intercellularly or intracellularly inside a plant, in a mutually beneficia1 rela- tionship. The host plant (the macrosymbiont) usually supplies the endophyte (the microsymbiont) with photosynthates, whereas the benefit of these associations for the plant is an improvement in access to important mineral nutrients whose supply would otherwise be growth limiting. With the excep- tion of cyanobacterial symbioses, in most endophytic symbioses involving higher plants, microorganisms colonize the plant root (reviewed in Quispel, 1992). In mycorrhizal sym- bioses, the fungus improves the absorption of minerals, particularly phosphorus, by the roots (Read et al., 1992; Harrison and van Buuren, 1995; see also Gianinazzi-Pearson, 1996, in this issue). In root nodule symbioses involving eubac- teria of the family Rhizobiaceae or actinomycetes of the genus frankia, the microsymbiont supplies the plant host with am- monium, the product of biological nitrogen fixation.
To provide an ecological niche for the endophyte, the mac- rosymbiont has to undergo symbiosis-specific differentiation. In nitrogen-fixing root nodule symbioses, the microsymbionts are hosted in special organs, the root nodules, which develop on roots upon infection by the microsymbiont (reviewed in Benson and Silvester, 1993; Myiona et al., 1995). In root sym- bioses with fungi, plant differentiation is less dramatic. In ectomycorrhizal symbioses, for example, the fungus somewhat alters root growth, triggering the formation of short thick infected roots (Martin and Hilbert, 1991), whereas in endo- mycorrhizal symbioses, the plant root structure is altered at the cellular leve1 but not macroscopically (Koide, 1991; Gianinazzi-Pearson, 1996, in this issue).
In this review, we discuss the recent progress in research on endophytic symbioses involving plant roots. Special atten- tion is given to nitrogen-fixing symbiotic interactions, and we summarize our current knowledge of nodule formation and function. The last part of this review focuses on the develop- ment of model systems and the relationships between different endophytic symbioses involving plant roots.
To whom correspondence should be addressed
The two microsymbionts able to enter into root nodule symbi- oses with higher plants are quite different. Rhizobia are unicellular, Gram-negative bacteria, whereas Frankia strains are filamentous, branching, Gram-positive bacteria. Rhizobia almost exclusively enter symbioses with legumes (for reviews, see Sánchez et al., 1991; Hirsch, 1992; Mylona et ai., 1995; Long, 1996, in this issue). The legume family has three subfam- ilies, and although >90°/o of the Papilionoideae and Mimosoideae are capable of entering a symbiosis with rhizo- bia, only members of one genus of the third subfamily, the Caesalpinioideae, can doso (Doyle, 1994). Conversely, frankia strains can interact with a diverse group of dicotyledonous plants that are members of eight different families, collectively called actinorhizal plants (Benson and Silvester, 1993).
60th rhizobia and Frankia strains can produce the enzyme nitrogenase that catalyzes the reduction of dinitrogen. Nitrogenase structure is conserved among the few prokary- otes capable of producing it; the enzyme is highly sensitive to oxygen (reviewed in Burris, 1991). In symbiosis as well as under nitrogen-limiting conditions in the free-living state, frankia can form nitrogen-fixing vesicles at the ends of its hyphae (reviewed in Benson and Silvester, 1993). In thesevesi- cles, bacterial nitrogenase is protected from oxygen by a multilayered envelope consisting of hopanoids, which are bac- teria1 steroid lipids (Parsons et ai., 1987; Berry et al., 1993). In contrast, rhizobia cannot provide oxygen protection for nitrogenase themselves and, with afew exceptions (see, e.g., de Bruijn, 1989), are capable of living on dinitrogen Only in the symbiotic state.
Usually, the development of a symbiotic interaction involves some differentiation of the endophyte. This differentiation may be minimal, like the increase in heterocyst formation described for endophytic cyanobacteria (Bergman et al., 1992), but it is often quite pronounced. In most legume nodules, intracellu- lar rhizobia differentiate into nitrogen-fixing bacteroids. This process involves a marked increase in cell size, a change in morphology from rod-shaped to branched cells (Bergersen, 1974; Newcomb, 1976), and changes in surface antigens (Olsen et al., 1992; Roest et al., 1995). Symbiosis-specific differenti- ation of Frankia depends on the host plant species, which determines the morphology of the vesicles formed in most

1900 The Plant Cell
actinorhizal symbioses and their location in the infected cells (reviewed in Baker and Mullin, 1992).
60th rhizobial and actinorhizal symbioses are set in motion by an exchange of signals, a process that has been studied in detail for legume nodule induction but is poorly understood for actinorhizal symbioses (Knowlton et al., 1980; van Ghelue et al., 1997). During legume nodule induction, flavonoids secreted by plant roots induce the transcription of rhizobial nod genes, which encode proteins catalyzing the synthesis of specific lipo-chito-oligosaccharides, the Nod factors (see Long, 1996, in this issue). Nod factors induce the early steps of nodule formation by inducing cell divisions in the root cor- tex, the new cells forming the nodule primordium. Simultaneously, bacteria enter the plant root and move through the root cortex toward the nodule primordium. They infect primordium cells, and in due course, the primordium develops into a mature nodule. The processes of infection and of induc- tion of cortical cell divisions are described in detail below.
lnfection
To reach and infect cells of the nodule primordium, the en- dophytic bacteria have to cross the root epidermis and move through the cortex toward their target cells. In legumes that form determinate nodules (see “Nodule Structure and Func- tion,” below), the nodule primordium is formed in the outer cortex and the bacteria have only to cross the epidermis to reach and infect primordium cells. However, in legumes that form indeterminate nodules, nodule primordia are formed in the inner cortex. In actinorhizal plants and in farasponia, the only nonlegume that can enter a symbiosis with rhizobia, nod- ule primordia are formed in the pericycle, as are lateral root primordia. In these symbioses, then, the bacteria have to tra- verse most or all of the root cortex (Lancelle and Torrey, 1984a; Kijne, 1992; Benson and Silvester, 1993).
Bacteria can penetrate plant roots either intracellularly or intercellularly. In intracellular infections, the bacteria are al- ways surrounded by a membrane of plant origin. Either infection route can be taken in legume as well as actinorhizal symbi- ases, and as with nodule structure, the host plant determines which route is chosen.
lntracellular lnfection
lntracellular infections begin when bacteria enter the root via curled root hairs (Kijne, 1992). The bacteria induce a localized hydrolysis of the root hair cell wall (van Spronsen et al., 1994), whereas the plasma membrane invaginates and new cell wall material is deposited (reviewed in Newcomb, 1981; Kijne, 1992). In this way, a tubular structure is formed through which the bacteria enter the plant root. For both legume and actinorhizal symbioses, this tubular structure is known as the infection thread. Within the nascent infection thread, bacteria become embedded in a plant cell wall-like matrix that accumulates in
the invagination of the plasma membrane. lnfection threads grow toward the nodule primordium cells intracellularly, that is, by traversing cortical cells (reviewed in Newcomb, 1981; Berry and Sunell, 1990).
Before a legume root cortical cell is traversed by an infec- tion thread, the nucleus moves to the center of the cell, and the microtubules and cytoplasm rearrange to form a phragmoplast-like structure (van Brussel et al., 1992). These structures are polarized; the bulk of the cytoplasm and the en- domembranes are located at the outer side, and all amyloplasts are located at the inner side. The infection thread crosses the cortex to the incipient nodule primordium, growing through the more or less radially aligned phragmoplast-like structures, which form cytoplasmic bridges. In situ hybridization studies using marker genes for different phases of the cell cycle (Yang et al., 1994) have shown that cells forming these phragmoplast- like structures reenter the cell cycle and most likely become arrested in the G2 phase.
lnfection threads grow at their tips (Dart, 1974), a growth habit exhibited by only a few plant cell types such as pollen tubes and root hairs. These cells have a polarly organized cytoplasm. Thus, for root cortical cells to support infection thread growth, their cytoplasm must reorganize in a polar manner, as is in- deed the case in the cells forming cytoplasmic bridges. Cytoplasmic bridges have also been described in actinorhizal plants infected intracellularly (R.H. Berg, personal communi- cation), indicating that the penetration of cortical cells by infecting Frankia hyphae also involves a polar reorganization of the cortical cell cytoplasm.
After the infection thread reaches the nodule primordium in nodules induced by rhizobiaon members of the legume sub- families Mimosoideae and Papilionideae, the bacteria are released into the primordium cells by a process resembling endocytosis (Figure 1A; Bassett et al., 1977). During this pro- cess, rhizobia become enclosed by a plant-derived membrane, the peribacteroid membrane (PBM), which, together with the bacteria and the space between the PBM and the bacteria (peribacteroid space), forms the symbiosome (Roth and Stacey, 1989). In the symbiosomes, rhizobia differentiate into their en- dosymbiotic form, the bacteroids, and start fixing nitrogen (Newcomb, 1981).
Conversely, in severa1 other rhizobial symbioses and in all actinorhizal symbioses, bacteria are not released from the in- fection threads and, in the case of rhizobia, do not differentiate into bacteroids. Instead, they stay enclosed in infection thread-like structures within the infected cells (Figure 16; Lancelle and Torrey, 1984b; Berry and Sunell, 1990; Naisbitt et al., 1992). Furthermore, unlike the symbioses described above, in actinorhizal symbioses and in rhizobial symbioses with persistent infection threads, no compartment correspond- ing to the infection thread matrix exists (reviewed in Sprent, 1989; Berry and Sunell, 1990). lnfected cells are filled by branching infection threads. In most actinorhizal symbioses, after the infected cells have been filled with Frankia hyphae, nitrogen-fixing vesicles develop (Schwintzer et al., 1982;

Rhizobial and Actinorhizal Symbioses 1901
cytoplasmic bridgerhizobiainfection thread wallinfection thread matrixperibacteroid space
plant plasmamembrane of PBM
secreted materialfibritlar materialFranttia hypha
Figure 1. Uptake of Endosymbionts into Plant Cells.(A) Infection of a legume nodule primordium cell. The intracellular path-way of infection is shown. (Top) Within the infection thread, rhizobiaare embedded in the infection thread matrix and surrounded by thefibrillar infection thread wall. (Bottom) When an infection thread reachesa nodule primordium cell, rhizobia are released into host cells fromthe unwalled tip of the infection thread via an endocytotic process(Newcomb, 1976). Within the host cytoplasm, the bacteria are sur-rounded by the PBM, forming symbiosomes (Bassett et al., 1977). Thesymbiosomes multiply and enlarge (not shown).(B) Infection of an actinorhizal nodule primordium cell. The intercellu-lar pathway of infection is shown. (Top) Frankia hyphae grow alongthe middle lamella between cortical cells, which secrete proteinaceousand pectinaceous material into the intercellular spaces (Liu and Berry,1991). (Bottom) When they reach the nodule primordium, the Frankiahyphae disseminate rapidly throughout the small intercellular spaces(Miller and Baker, 1985). Hyphal tips enter primordium cells as thehost cell plasma membrane invaginates, and new fibrillar material isdeposited around the invading hypha. Surrounded by the invaginatedhost plasma membrane and embedded in fibrillar host-derived mate-rial, the hyphae proliferate inside the infected primordium cells (Millerand Baker, 1985).
infection route is always taken by rhizobia that form stem nod-ules on Sesbania sp or Aeschynomene sp (Tsien et al., 1983;Alazard and Duhoux, 1990). Alternatively, Frankia strains, andin some cases, rhizobia, can enter the root by penetrating themiddle lamella between intact epidermal cells (Miller and Baker,1985; de Faria et al., 1988; Racette and Torrey, 1989; Ndoyeet al., 1994). In the roots of actinorhizal plants infected inter-cellularly, bacteria move through the cortex toward the noduleprimordium intercellularly and become intracellular only whenthey are taken up into primordium cells in a process resem-bling infection thread formation (Figure 1B; Miller and Baker,1985; Racette and Torrey, 1989). The infection of primordiumcells proceeds as described above for the intracellular infec-tion route. As Frankia hyphae move intercellularly through thecortex, epidermal and cortical cells secrete electron-dense ma-terial, which is rich in pectin and proteins (Liu and Berry, 1991),into the intercellular spaces (Miller and Baker, 1985; Racetteand Torrey, 1989).
In legumes infected via intercellular penetration of theepidermis, rhizobia can proceed toward the nodule primordiumcells in different ways. In some legumes, such as Sesbaniarostrata and Neptunia sp, infection threads are formed in thecortex after intercellular penetration of the epidermis (see, forexample, Tsien et al., 1983; Ndoye et al., 1994; Subba-Rao etal., 1995). These examples represent intermediates betweenintercellular and intracellular infection routes. In other legumes,such as Aeschynomene sp and Stylosanthes sp, rhizobia movethrough the cortex intercellularly. During this process, they in-duce the collapse of cortical cells, leading to the formationof an "infection strand," which the bacteria use to reach theprimordium (Chandler et al., 1982; de Faria et al., 1988; Alazardand Duhoux, 1990). The collapse of cortical cells shows thatthe bacteria display characteristics of pathogens in this phaseof the interaction. By analogy, the secretion of material intothe intercellular spaces during intercellular growth of the bac-teria in actinorhizal symbioses could therefore be a way toobstruct pathogen resistance responses of the cortical cells.
From this point of view, it seems that the intracellular path-way of infection permits the plant more stringent control overthe microsymbiont during the infection process, because theinfection thread determines the speed and direction of bac-terial progression through the cortex.
Newcomb and Wood, 1987). Depending on the host plant spe-cies, the vesicles can be spherical, club-shaped, pear-shaped,or filamentous (reviewed in Newcomb and Wood, 1987; Bakerand Mullin, 1992).
Intercellular Infection
During intercellular infection, rhizobia can penetrate throughthe gaps in the epidermis that can form where lateral rootsor adventitious roots emerge from the main root or the stem,respectively (Chandler et al., 1982; James et al., 1992). This
Mitotic Reactivation
Upon infection, Frankia and rhizobia induce the formation ofa new organ, the nodule. Nodule development starts with themitotic reactivation of specific root cells. This process occursat the same time as the infection process. However, at leastin legumes, the induction of cell divisions does not require bac-terial infection, because purified rhizobial Nod factors are ableto mitotically reactivate root cortical cells (Spaink et al., 1991;see also Long, 1996, in this issue). As mentioned above, thelocation of the cells that become activated to form the nodule
1902 The Plant Cell
primordium is controlled by the host plant. In legumes, the primordia are formed in the inner or outer cortex of the root, whereas in Parasponia and actinorhizal plants, nodule primor- dia, like lateral root primordia, are formed in the root pericycle. In Parasponia and intracellularly infected actinorhizal plants, cell divisions are also induced in the cortex below the infected root hair. lnfection threads grow toward these dividing and en- larging cortical cells, and some of the cells become filled with endosymbiont material. The resulting structure is called a prenodule. Prenodules do not develop into nodules, so it is unclear whether they have any function in the symbiosis. Regardless, infection threads grow from the prenodule toward the developing nodule lobe (Lancelle and Torrey, 1984a; Berry and Sunell, 1990).
The mechanism by which Nod factors mitotically reactivate root cells is unclear, but severa1 lines of evidence suggest that during nodule induction, Nod factors affect the phytohormone balance in the root cortex. For example, bacteria-free nodules in which early nodulin genes are expressed can be induced on legume roots by Escherichia coli producing cytokinin (Cooper and Long, 1994) and by auxin transport inhibitors (reviewed in Hirsch, 1992). These data suggest that Nod fac-
A B
tors result in an altered phytohormone balance in the cortex, most likely an increased leve1 of cytokinin. This would distin- guish legume nodule induction from lateral root formation, which ordinarily is triggered by increases in auxin levels (Xorrey, 1950; Blakely et al., 1988).
During nodule induction, the expression of the early nodu- lin gene ENOD40 is induced in the pericycle of the root (Kouchi and Hata, 1993; Yang et al., 1993), and exogenously added Nod factors are sufficient to activate this gene (Franssen et al., 1995). This demonstrates that in addition to the epidermis and cortex, the pericycle can also respond to Nod factors. ENOD40, homologs of which have been found in nonlegumes, encodes a peptide that alters the response of plant cells to auxin (van de Sande et al., 1996). Therefore, it seems proba- ble that this early nodulin plays a role in the induction of legume nodule primordium formation. However, whether ENOD40 homologs are also involved in actinorhizal nodule formation has yet to be established.
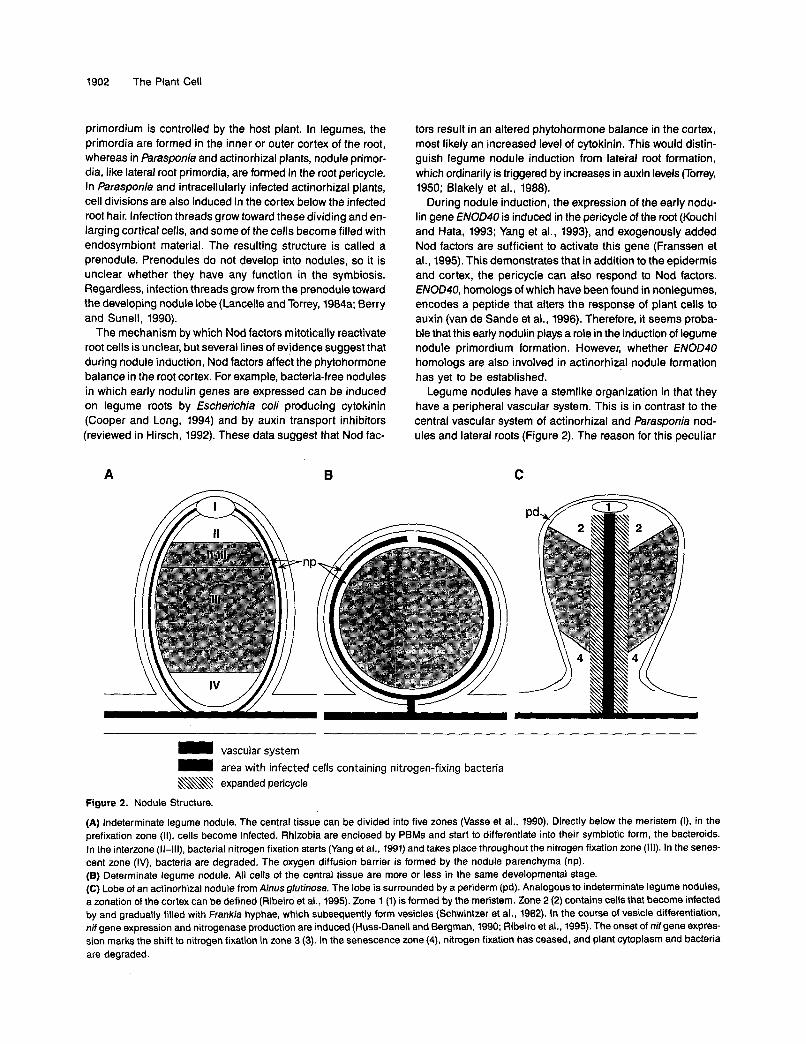
Legume nodules have a stemlike organization in that they have a peripheral vascular system. This is in contrast to the central vascular system of actinorhizal and Parasponia nod- ules and lateral roots (Figure 2). The reason for this peculiar
C
= vascular system - area with infected cells containing nitrogen-fixing bacteria &\\- expanded pericycle
Figure 2. Nodule Structure.
(A) lndeterminate legume nodule. The central tissue can be divided into five zones (Vasse et al., 1990). Directly below lhe meristem (I), in the prefixation zone (II), cells become infected. Rhizobia are enclosed by PBMs and start to differentiate into their symbiotic form, the bacteroids. In the interzone (11-lll), bacterial nitrogen fixation starts (Yang et al., 1991) and takes place throughout the nitrogen fixation zone (111). In the senes- cent zone (IV), bacteria are degraded. The oxygen diffusion barrier is formed by the nodule parenchyma (np). (6) Determinate legume nodule. All cells of the central tissue are more or less in the same developmental stage. (C) Lobe of an actinorhizal nodule from Alnus glutinosa. The lobe is surrounded by a periderm (pd). Analogous to indeterminate legume nodules, a zonation of the cortex can be defined (Ribeiro et al., 1995). Zone 1 (1) is formed by the meristem. Zone 2 (2) contains cells that become infected by and gradually filled with Frankia hyphae, which subsequently form vesicles (Schwintzer et al., 1982). In the course of vesicle differentiation, nif gene expression and nitrogenase production are induced (Huss-Dane11 and Bergman, 1990; Ribeiro et al., 1995). The onset of nif gene expres- sion mark the shift to nitrogen fixation in zone 3 (3). In the senescence zone (4), nitrogen fixation has ceased, and plant cytoplasm and bacteria are degraded.

Rhizobial and Actinorhizal Symbioses 1903
morphology of legume nodules is still a subject for specula- tion, although it has been suggested that the structural difference between legume and other root nodules may be due to the availability of a unique developmental program in legumes. Some legume cultivars can spontaneously form bacteria-free nodules (Truchet et al., 1989; Blauenfeldt et al., 1994). These nodules are rich in amyloplasts and may origi- nally have evolved as carbon storage organs (Joshi et al., 1993). The developmental program of these putative carbon storage organs may have been exploited during the evolution of nitrogen-fixing symbioses in the legumes. In other symbiotic nitrogen-fixing plants, the morphogenetic program for lateral roots was used instead. Unfortunately, this interesting hypoth- esis is hard to verify because carbon storage organs have not been found in extant or extinct legume species.
Legume and actinorhizal nodules as well as lateral roots are predominantly formed opposite to protoxylem poles (Libbenga and Bogers, 1974; Callaham and Torrey, 1977; Blakely and Evans, 1979). This suggests that the stele provides positional information that controls which cells can be activated. In the- ory, this positional information could involve compounds from the protoxylem pole and from the phloem that, respectively, facilitate or repress cell divisions in the pericycle and cortex. These postulated stele factors could act in concert with mi- totic factors.
It has indeed been demonstrated that factors from the stele stimulate phytohormone-induced cell division in stele-free root explants (Libbenga et al., 1973), and one of these factors has been identified as uridine (Smit et al., 1995). Although, in stele- free explants, extracts from the stele stimulated cell divisions all over the cortex (Libbenga et al., 1973), in stele-containing explants, cell divisions were stimulated by uridine predomi- nantly opposite protoxylem poles (Smit et al., 1995). Therefore, it is likely that compounds that are released from the stele de- termine the susceptibility of cortical cells to phytohormones and uridine (Figure 3). The fact that the development of corti- cal and/or pericycle cells is controlled by positional information from the stele is also reflected by the fact that in pea roots, the pericycle at the protoxylem poles consists of more cell layers than the pericycle opposite the phloem (Rost et al., 1988). and that in Arabidopsis roots, pericycle cells opposite protoxylem pools are morphologically distinct from their neighbors (Laskowski et al., 1995). Thus, positional information deter- mines the susceptibility of cortical cells to mitogens, and compounds from the stele, in synergy with rhizobial Nod fac- tors. control the mitotic reactivation of cortical cells.
Regulation of Nodule Number
To have a symbiotic relationship and not succumb to parasit- ism, the number of nodules must be controlled. In both legumes and actinorhizal plants, nodule formation is inhibited after a certain number of nodules have formed on the root system (Dobritsa and Novik, 1992; Wall and Huss-Danell, 1997). This autoregulation is abolished in severa1 monogenic legume mu- tants that display the so-called supernodulation phenotype
\
x /
endodermis [-I phloem
WE pericycle xylem
1-1 nodule primordium
Figure 3. Determination of the Position of Mitotically Reactivated Cells.
Mitotic reactivation normally takes place opposite protoxylem poles. Thus, factors determining the susceptibility of cortical cells to mitotic activation must be exuded by the stele, that is, positive factors from the xylem (+) and/or negative factors (-) from the phloem. These fac- tors should form a gradient, which in coordination with Nod factors produced by the bacteria and taken up by the root (arrowhead) will determine the location of cell divisions (Heidstra et al., 1994). Because uridine preferentially stimulates cortical cell divisions in pea root ex- plants opposite protoxylem poles, it is a candidate for a positive factor. However, it is not known whether uridine is in fact exuded by the protoxylem.
(reviewed in Caetano-Anollés and Gresshoff, 1991). Due to their increased nodule number, the aerial parts of supernodulating mutants can display reduced growth (Hansen et al., 1990, 1992), indicating that when the plant cannot control the nod- ule number, the relationship with rhizobia can become parasitic.
Nodule induction by intracellular infection is restricted to the zone of root hair elongation of the growing root on both legumes (Bhuvaneswari et al., 1981) and actinorhizal plants (Burggraaf et al., 1983). As the root continues to grow, new nodules form as the zone of root hair elongation migrates with the growing root tip. When nodules are formed, the formation of additional nodules in the current zone of elongation is suppressed.
Different approaches have been taken to understand how this control of nodulation is achieved. When mature nodules are excised from a root system, new nodules grow out at

1904 The Plant Cell
positions at which infection must have occurred, that is, where the elongation zone was localized before excision (Takats, 1990; Caetano-Anollés et al., 1991; Wall and Huss-Danell, 1997). This indicates that in both symbioses, autoregulation leads to an arrest of the symbiotic interaction, which can be abolished later. Furthermore, in studies using split root systems of legumes or actinorhizal plants, nodule primordium formation on one side of the root system inhibits subsequent nodule formation on the other side (Caetano-Anollés and Gresshoff, 1991; Dobritsa and Novik, 1992; Wall and Huss-Danell, 1997). These experi- ments revealed that the induction of nodule primordia leads to the formation of a translocatable signal that acts systemi- cally to suppress further primordium induction. Although, in legume roots, this signal has to affect cortical cell divisions that are involved exclusively in nodule formation, in roots of actinorhizal plants, the signal should affect only cell divisions in the pericycle that lead to the formation of nodule primordia, not those leading to lateral root development. Thus, actinorhizal plants must be able to initiate separately both processes at the leve1 of primordium induction.
Grafting experiments with legumes have shown that there is a shoot factor involved in autoregulation that is active even across species boundaries (Delves et al., 1987a, 1987b). Thus, it is likely that the translocatable signal formed in roots affects the production of a translocatable shoot factor that controls nodule number (Caetano-Anollés and Gresshoff, 1991). The molecular nature of these signals is not known, but they may represent compounds that, fike abscisic acid, are involved in the induction of a dormant stage (McCarty, 1995).
NODULE STRUMURE AND FUNCTION
Two types of legume nodules are known: determinate and in- determinate. The type of nodule that is formed depends on the host plant (Trinick, 1980). In indeterminate nodules (Fig- ure 2A), cell divisions in the nodule apical meristem continuously add new cells to the different nodule tissues. These cells differentiate, and some of them subsequently be- come infected by Rhizobium. As a consequence, a developmental gradient of infected cells is established along the nodule axis passe et al., 1990). Conversely, in determinate nodules (Figure 2B), the nodule meristem stops its activity early in nodule development, and for the most part, new cells al- ready contain bacteria because they are formed by division of infected cells (Newcomb, 1981; Rolfe and Shine, 1984).
In actinorhizal plants, the formation of nodule primordia takes place in the root pericycle. This is where lateral root primordia are formed, and like lateral roots, actinorhizal nodules have an indeterminate growth pattern (Figure 2C). Hence, ac- tinorhizal nodules also show a developmental gradient (Ribeiro et al., 1995). Whereas in most actinorhizal nodules, infected and uninfected cells are interspersed in the cortex, in nodules formed on roots of Datisca and Coriaria, the infected cells form a continuous patch on one side of the acentric stele (Newcomb
and Pankhurst, 1982; Hafeez et al., 1984). Interestingly, the only nonlegume infected by rhizobia, Parasponia (Ulmaceae), shows nodule ontogeny and tissue organization similar to that of most actinorhizal plants.
Nodules provide a niche for nitrogen-fixing bacteria. TO achieve this symbiotic relationship, the plant must balance the high energy demand for bacterial nitrogen fixation, which is met primarily by glycolytic processes requiring carbon sources and oxygen (reviewed in Day and Copeland, 1991), against the necessary protection of the nitrogenase enzyme from oxygen (reviewed in Hunt and Layzell, 1993). Furthermore, am- monium, the product of nitrogen fixation that is exported to the plant cytoplasm, is assimilated in the glutamine synthe- tase (GS)/glutamate synthase cycle, a process requiring carbon backbones (reviewed in Schubert, 1986). Subsequently, the assimilated nitrogen is transported to other parts of the pfant via the vascular system (reviewed in Streeter, 1993). Hence, nodule functioning requires the active transport of oxygen to sites of bacterial and plant respiration, but the oxygen con- centration has to be kept low to protect the nitrogenase. These requirements are met by a variety of structural and biochemi- cal speciafizations in different types of nodules, which are described in detail below.
Legume Nodules: Structure and Nodule-Specific Gene Expression
For two decades, the molecular analysis of legume nodules has been focused on the cloning and characterization of plant genes showing nodule-specific expression (nodulin genes; reviewed in SBnchez et al., 1991) in order to obtain tools to study nodule development. Severa1 nodulin cDNA clones have been isolated, representing genes that are induced during early andlor late stages of nodule development and functioning. So- called early nodulins (ENODs) are involved in the infection process and in establishing the nodule structure, whereas late nodulins have specific roles in nodule functioning (van Kammen, 1984). Studies on late nodulin genes have con- tríbuted to a better understanding of nodule functioning, and in general, their analyses support conclusions drawn from ear- lier biochemical and physiological studies (Cullimore and Bennett, 1988; Gordon, 1992). Late nodulins encoding en- zymes involved in carbon, nitrogen, and oxygen metabolism in nodules have been identified. These genes have become useful tools in experiments aimed at understanding how these metabolic processes are regulated. For example, it is now clear that the induction of GS is under both metabolic and develop- mental control in nodules (reviewed in Hirel et al., 1993), whereas in soybean, the biosynthesis of the assimilated nitro- gen transport compounds, the ureides, seems to be solely under devefopmental controf (Lee et af., 1993).
Plant genes encoding products involved in the interaction with the endosymbiont have also been identified. These 'in- clude nodulin-26, which encodes a PBM protein that is involved in metabolite exchange (Weaver et al., 1994), as well as puta-

Rhizobial and Actinorhizal Symbioses 1905
tive structural components of the peribacteroid space (Jacobs et al., 1987; Cheon et al., 1994). Moreover, studies on the mo- lecular biology of the oxygen transport protein leghemoglobin, the first nodulin to be characterized biochemically (reviewed in Appleby, 1984), have provided insight into the phylogeny of nodulated plants. For example, the promoter of the nodule- specific hemoglobin gene from the actinorhizal plant Casua- rina glauca, whose expression is confined to the infected cells (Gherbi et al., 1997), was found to direct infected cell-specific expression in nodules of the legume Lotus japonicus (Jacobsen-Lyon et al., 1995). These results indicate that tran- scription factors responsible for expression of (1eg)hemoglobin in the infected cells are conserved between legumes and the actinorhizal plant C. glauca (Jacobsen-Lyon et al., 1995).
In situ hybridization analyses of the expression of several early nodulin genes during nodule formation have provided some useful tools to study the mode of action of rhizobial Nod factors. A well-studied example is ENOD12, which has been used to analyze the structural requirements necessary for Nod factors to trigger the plant responses (Ardourel et al., 1994; Bauer et al., 1994; Journet et al., 1994).
Conversely, studies to date on ENOD genes have provided less information on the mechanisms by which rhizobia induce nodule development. This is in part due to the fact that as yet, none of the nodulin genes corresponds to any of the geneti- cally identified plant genes that are necessary for normal nodule development (see below).
Molecular analyses of components of the nodule oxygen diffusion barrier have also not provided significant functional insight. All legume nodules as well as nodules formed on farasponia contain an oxygen diffusion barrier to control the flow of oxygen to the infected cells. This barrier, in combina- tion with the high metabolic activity of the infected cells, results in a low concentration of free oxygen (Denison and Layzell, 1991). Within the infected cells, high levels of oxygen binding PrOteinS, such as leghemoglobins, provide efficient oxygen transport (Appleby, 1984). In legume nodules, the oxygen diffu- sion barrier is present in the nodule parenchyma, which surrounds the inner tissue and encloses the peripheral nod- Ule vascular system (Figures 2A and 26). The nodule parenchyma consists of layers of cells more or less devoid of intercellular spaces (Witty et al., 1986). Some nodule-specific genes that encode putative cell wall proteins (van de Wiel et al., 1990; Wilson et al., 1994) and a lectin (Bauchrowitz et al., 1996) are expressed in these cells. However, it is not clear whether or how these proteins contribute to the function of the oxygen diffusion barrier.
Actinorhizal Nodules: Structure and Nodule-Specific Gene Expression
Actinorhizal nodules as well as nodules induced by rhizobia on the nonlegume Parasponia consist of multiple lobes, each lobe representing a modified lateral root without a root cap and with infected cells present in the expanded cortex (Figure
2C). Most actinorhizal plants are trees or woody shrubs, which has made them recalcitrant to molecular-genetic analyses (S6guin and Lalonde, 1991). Recently, however, a number of nodule-specific plant genes in actinorhizal symbioses have been identified and characterized (Goetting-Minesky and Mullin, 1994; Jacobsen-Lyon et al., 1995; Ribeiroet al., 1995). Furthermore, it is now feasible to perform gene function studies using model systems such as Arabidopsis and yeast (Ribeiro et al., 1996), and the transformation of an actinorhizal plant species has recently been reported (Diouf et al., 1995).
The characterized actinorhizal genes that are expressed at markedly elevated levels in nodules compared with roots (so- called actinorhizin genes) include a nodule-specific hemoglo- bin gene from C. glauca (Jacobsen-Lyon et al., 1995; Gherbi et al., 1997) and some cDNAs from Alnus glufinosa (Goetting- Minesky and Mullin, 1994; Ribeiro et al., 1995). Currently, A. glutinosa is possibly the most well characterized actinorhizal plant. All of the A. glutinosa actinorhizin genes that have been identified were found to be expressed in infected cortical cells, although at different stages of their development (Pawlowski et al., 1994; Ribeiro et al., 1995; van Ghelue et al., 1996; Guan et al., 1997a, 1997b). For example, the expression of a nodule- specific subtilisin-like protease is induced early during the in- fection of cortical cells, before the onset of bacterial nitrogen fixation (Figures 4A and 46; Ribeiro et al., 1995). This gene may therefore constitute a useful reporter for the analysis of signal exchange between A. glufinosa and Frankia. Conversely, GS is expressed in infected cells at the onset of bacterial nitro- gen fixation (Guan et al., 1997b), whereas ag73, which encodes a putative acidic extracellular protein, is expressed in senes- cent infected cells (Guan et al., 1997a).
Alnus nodules contain an unusual vascular pericycle con- sisting of many layers of small cells with dense cytoplasm and numerous mitochondria, indicative of high metabolic activity (Burgess and Peterson, 1987). Sucrose synthase, GS, and eno- lase are expressed at high levels in this pericycle (Figure 4C; van Ghelue et al., 1996; Guan et al., 1997b). The pericycle there- fore seems to play a role in transport between the vascular elements and the nodule cortex. The expression of several actinorhizin genes in this tissue implies that it has properties different from those of the root pericycle (Pawlowski et al., 1994; Guan et al., 1997a). Thus, the expression patterns of the ac- tinorhizin genes define two specialized cell types in Alnus nodules, the infected cortical cells and those cells that form the pericycle.
Severa1 diverse mechanisms have evolved for oxygen con- trol in actinorhizal nodules formed by different plant species. First, the microsymbiont can, and in most cases does, con- tribute to oxygen protection of nitrogenase through the formation of vesicles. Second, the high metabolic activity of these vesicles is likely to reduce oxygen concentration (Vikman, 1992). Third, the plant in some cases provides an oxygen diffu- sion barrier andlor an oxygen transport system, that is, hemoglobin.
For instance, in actinorhizal nodules of Casuarina, in which Ffankia does not form vesicles (Berg and McDowell, 1987a),

1906 The Plant Cell
Figure 4. In Situ Localization of Gene Expression in Actinorhizal Nod-ules of A. glutinosa.
Longitudinal sections of an 8-week-old nodule were hybridized with35S-labeled antisense RNA and stained with Toluidine blue (Ribeiroet al., 1995; van Ghelue et al., 1996). Bright-field micrographs weretaken under epipolarized light, so silver grains denoting hybridizationsappear in green.(A) Expression of Frankia nifH denotes nitrogen-fixing infected cells.(B) Expression of A. glutinosa ag12, encoding a serine protease.(C) Expression of A. glutinosa sucrose synthase.
and Myrica, in which vesicles are formed (Tjepkema and Asa,1987), it seems that the plant provides facilities for both oxy-gen protection and transport similar to those described forlegume nodules. In Casuarina and Myrica, an oxygen diffu-sion barrier is achieved by lignification of the walls of infectedand adjacent uninfected cells (Berg and McDowell, 1987b; Zenget al., 1989; Zeng and Tjepkema, 1994, 1995). Both types ofactinorhizal nodules contain high amounts of nodule-specifichemoglobin, thus facilitating oxygen transport (Fleming et al.,1987; Pathirana and Tjepkema, 1995).
In Alnus nodules, Frankia forms vesicles that seem to con-tribute to oxygen protection of nitrogenase both metabolically(Vikman, 1992) and as a diffusion barrier (Kleemann et al.,1994). No oxygen diffusion barrier could be detected in thesenodules, but hemoglobin was found, although in such lowamounts that its function is unclear (Suharjo and Tjepkema,1995). In contrast, in nodules of Coriaria, Frankia also formsvesicles (Newcomb and Pankhurst, 1982), and the plant pro-vides an oxygen diffusion barrier but no hemoglobin (Silvesterand Harris, 1989; Silvester et al., 1990).
It has been assumed that because of their lateral rootlikestructure, actinorhizal nodules represent a more primitive sys-tem than legume nodules. Although these studies are only intheir infancy, the isolation of six actinorhizal nodule-specificgenes suggests that the formation of actinorhizal nodules isequally as complex as that of legume nodules. Most nodule-specific genes identified to date in both systems are expressedin infected cells, implying that they encode products involvedin the interaction with the endosymbiont. These interactionsare probably just as intricate, regardless of the4 endogenousdevelopmental program on which they are superimposed.
GENETIC ANALYSIS OF NODULE DEVELOPMENT
To obtain more insight into the processes controlling noduleformation, it is essential to integrate a genetic approach. Inthe past two decades, legume mutants showing a changednodulation phenotype have been isolated from alfalfa, pea,
The zonation of the nodule lobe is indicated in (A). Arrowheads pointat infected cells of zone 2, which are not completely filled with Frankiahyphae. These cells express sucrose synthase at low levels and ag12at high levels. In infected cells of zone 3, Frankia nifH is expressed.Arrows point at nitrogen-fixing infected cells of zone 3, which displayhigh levels of sucrose synthase expression; ag12 expression in thesecells is significantly lower than in infected cells of zone 2. The arrowlabeled with p indicates the expanded pericycle of the central vascu-lar system. Sucrose synthase is expressed at high levels in the pericycle.The nodule lobes are surrounded by a periderm consisting of cellswhose central vacuoles are filled with polyphenols and appear darkblue in the images.

Rhizobial and Actinorhizal Symbioses 1907
chick pea, and soybean (reviewed in Caetano-AnollBs and Gresshoff, 1991). These studies have demonstrated that mu- tants affected in almost every step of nodule development can be obtained. For example, one class of mutants has lost the ability to form root nodules altogether. Some of these mutants are affected in the very first plant response to bacterial Nod factors, showing no root hair deformation (Had- phenotype; Duc and Messager, 1989; Sagan et al., 1994). One of these mutants is blocked in the first measurable response to Nod factors, oscillating changes of intracellular calcium in root hairs (Ehrhardt et al., 1996). Other mutants show root hair deforma- tion but do not support infection thread formation (Inf- phenotype; Postma et al., 1988; Markwei and LaRue, 1992; Sagan et al., 1994). There are also mutants that can form in- fection threads but are affected in the induction of cortical cell divisions (Noi- phenotype; Duc and Messager, 1989; Sagan et al., 1994). Other mutants, such as the aforementioned su- pernodulating mutants, can form nodules but are affected in the number of nodules they form (Jacobsen and Feenstra, 1984; Duc and Messager, 1989), and another class comprises mutants that form nodules defective in nitrogen fixation and are thus disturbed at later steps of nodule development (Fix- phenotype; Duc and Messager, 1989; Sagan et al., 1993).
In addition to their value in elucidating nodule formation and function, these mutants have also provided the means to study relationships between plants and other symbiotic microorgan- isms, particularly endomycorrhizal fungi. For example, it has been shown that some Nod- mutants are also unable to sup- port an endomycorrhizal symbiosis (Myc- phenotype). Conversely, all Fix- mutants examined thus far display a Myc+ phenotype (Duc et al., 1989; Gianinazzi-Pearson et al., 1991; Bradbury et al., 1993; Sagan et al., 1995). These observations imply that some early events in the establishment of symbioses are shared in very different systems (see Gianinazzi-Pearson, 1996, in this issue).
Unfortunately, the above-mentioned nodulation mutants have been generated in legume species that are not easily amena- ble to positional cloning strategies, despite the development of molecular maps (Kiss et al., 1993) and YAC libraries (Funke et al., 1994). Two legume species, Medicago truncatula and L. japonicus, have been proposed as model systems (Barker et al., 1990; Handberg and Stougaard, 1992). Both plant spe- cies represent small diploid plants with a small genome (0.9 pg per haploid genome for M. truncatula, 0.5 pg for L. japoni- cus) and short generation times. Moreover, L. japonicus forms determinate nodules, whereas those formed on M. truncatula are indeterminate. M. truncatula is nodulated by Rhizobium meliloti, a well-characterized rhizobial strain from which large numbers of site-directed mutants have been generated; the Rhizobium strain nodulating L. japonicus has not been exam- ined in detail. For both systems, mutant plants affected in diffbrent steps of nodule development have been identified (Webb et al., 1994; Benaben et al., 1995; Sagan et al., 1995). Transposon mutagenesis is also feasible (Thykjaer et al., 1995). We await exciting developments in the molecular genetics of these model systems over the next few years.
Because the majority of actinorhizal plants are trees or woody shrubs with long generation times, no genetic systems for these symbioses are available. The only actinorhizal plant identified thus far that may offer the potential for genetic analysis is Datisca glomerata. Datisca is the only herbaceous actinorhizal plant genus identified to date. D. glomerata has a generation time of 6 months and a small genome of 1 to 2 pg (P. Okubara and A.M. Berry, personal communication). Moreover, some actinorhizal plants can be transformed (Diouf et al., 1995; C. Franche, D. Bogusz, and E. Duhoux, personal communica- tion), providing a more directed approach toward functional studies of nodule-specific genes. Recent achievements in legume transformation (Puonti-Kaelas et al., 1990; Thomas et al., 1992; Sato et al., 1993) have provided the impetusfor similar experiments in this system (Cheon et al., 1993; Lee et al., 1993).
CONCLUDING REMARKS
In this review, aspects of the formation of nitrogen-fixing root nodules, induced by rhizobia and Frankia strains, have been described. The similarities in the infection processes used in the different systems suggest that actinorhizal and rhizobial symbioses are related, but this hypothesis appears to be con- tradicted by the striking differences in legume and actinorhizal nodule ontogeny and structure. Nevertheless, rhizobial Nod factors, which trigger the formation of legume nodules, also appear to be essential for the induction of the formation of actinorhiza-like nodules on the nonlegume Parasponia (Marvel et al., 1987), indicating that nodule structure is entirely a host plant-determined trait and is not dependent on the nature of the signal molecules of the microsymbiont.
Recent studies of the molecular phylogeny of plants involved in nitrogen-fixing symbioses have revealed that in fact all plants entering nitrogen-fixing root nodule symbioses, namely, le- gumes, farasponia, and actinorhizal plants, form part of a single clade, the rosid I clade (Soltis et al., 1995). These data suggest a single evolutionary origin of the susceptibility to gain the ability to enter nitrogen-fixing root nodule symbioses.
Interestingly, independent studies concerning the molecu- lar phylogeny of legumes and of actinorhizal plants have led to the hypothesis that on the basis of a common predisposir tion, symbiosis has developed independently three or four times in both symbiotic systems. This hypothesis is supported by the differences in nodule morphology and infection mech- anisms within both systems (Doyle, 1994; Swensen, 1996). Furthermore, the specificity for the microsymbiont does not follow a clear pattern. Phylogenetic analyses show that the different actinorhizal families are not, for the most part, more closely related to each other than they are to legumes, and that Parasponia, which enters a symbiosis with rhizobia, isin fact more closely related to actinorhizal members of the Rham- naceae and Elaeagnaceae than to legumes (Swensen, 1996). At present, it is not possible to define the phylogenetic branches within this group of plants on which the symbiotic syndrome

1908 The Plant Cell
arose. All actinorhizal plant families containing symbiotic spe- CieS also contain nonsymbiotic genera. For example, Alnus spp (Betulaceae) can enter actinorhizal symbioses, whereas Bet- ula cannot. Therefore, the establishment and loss of the ability to enter actinorhizal symbioses may have occurred multiple times during evolution (Swensen, 1996).
Because actinorhizin genes are now available, additional approaches toward comparing the different root nodule-form- ing symbioses are feasible. For example, analysis of the expression of legume leghemoglobin and C. glauca symbiotic hemoglobin genes has revealed that infected cell-specific tran- scription factors seem to be conserved in both systems. This result implies that icfected cell-specific transcription factors may already have been present in the common ancestor of Casuarina and legumes. Further research is needed to clarify this point.
There seems to be a relationship between root nodule and endomycorrhizal symbioses, because mutants defective in both interactions have been identified (see above, and Gianinazzi- Pearson, 1996, in this issue). In certain mutants, all responses to rhizobia and mycorrhizal fungi seem to be blocked, imply- ing that some recognition and/or signal transduction components are shared in both symbioses. The involvement of common mechanisms in the mycorrhizal and rhizobial plant interactions has led to the hypothesis that rhizobia made use of mycorrhizal signals to establish their symbiotic relationship with plants (see Gianinazzi-Pearson, 1996, in this issue). Thus, the mycorrhizal signal molecules could resemble rhizobial Nod factors, which is not unlikely because Nod factors have a chi- tin backbone reminiscent of funga1 cell wall components (LaRue and Weeden, 1994). Because most plants can enter into endomycorrhizal symbioses, this hypothesis implies that Nod factor-like molecules should be recognized by nonle- gumes, as is in fact suggested by recent results (Rbhrig et al., 1995). The generation of Nod-IMyc- mutants in a model le- gume system (Sagan et al., 1995) has now provided the means to clone and characterize the genes whose products are in- volved in both types of interaction. However, although most angiosperms are able to establish symbioses with mycorrhizal fungi (see Gianinazzi-Pearson et al., 1996, this issue), only cer- tain members of the rosid I clade can enter root nodule symbioses. Therefore, it seems very probable that apart from the possible exploitation of the signal transduction pathway developed for the interaction with symbiotic fungi, to form nitrogen-fixing root nodules, plants needed a unique feature, which arose only once during angiosperm evolution.
Rhizobium-legume symbioses have often been compared with pathogenic interactions (Djordjevic et al., 1987; Long and Staskawicz, 1993). The occurrence of certain steps in the in- fection process that seem to be recruited from or are vestiges of a pathogenic interaction. For instance, infection thread de- velopment may be derived from a defense response papilla formation (Beswetherick and Bishop, 1993; Clark et al., 1993). The induction of cortical cell collapse by rhizobia during inter- cellular infection of certain legumes, shows that the borders between symbiotic and pathogenic interactions are not clearly
defined. This point is further exemplified by the interaction be- tween Alnus and the saprophytic fungus Penicillium nodositatum (Capellano et al., 1987; Sequerra et al., 1994, 1995). P nodositatum follows an intracellular infection process identical to that occurring in the interaction with Frankia, which leads to the formation of an actinorhiza-like nodule. The result- ing myconodules resemble actinorhizal nodules induced by Frankia strains not capable of symbiotic nitrogen fixation. Thus, the same host plant mechanisms are exploited by both sym- biotic and saprophytic organisms.
Now that legumes are accessible to genetic analysis, com- parisons of the effect of mutations on symbiotic and pathogenic interactions can be made. With the advent of molecular bio- logical analyses of actinorhizal symbioses, integrated studies of interactions between one plant and different microorgan- isms will become easier. Eventually, such studies will lead to a better understanding of plant-microbe interactions.
ACKNOWLEDGMENTS
We thank R.H. Berg (Biology Department, University of Memphis, Mem- phis, TN), A.M. Berry and !? Okubara (Department of Environmental Horticulture, University of California, Davis, CA), E. Duhoux, C. Franche, and D. Eogusz (ORSTOM-CIRA'DiForêt, Nogent sur Marne, France), K. Huss-Dane11 and L. Wall (Department of Plant Physiology, Umea University, Umel, Sweden), and B. Solheim and M. van Ghelue (In- stitute of Biology and Geology, University of Tromsd, Tromsd, Norway) for sharing unpublished information. We are indebted to S.M. Swensen (Biology Department, lthaca College, Ithaca, NY) for helpful discussions.
REFERENCES
Alazard, D., and Duhoux, E. (1990). Development of stem nodules in a tropical forage legume, Aeschynomene afraspera. J. Exp. Bot.
Appleby, C.A. (1984). Leghemoglobin and rhizobial respiration. Annu. Rev. Plant Physiol. 35, 443-478.
Ardourel, M., Demont, N., Debellb, F., Maillet, F., de Billy, F., Promb, J.-C., Dbnarib, J., and Truchet, G. (1994). Rhizobium meliloti lipooligosaccharide nodulation factors: Different structural require- ments for bacterial entry into target root hair cells and induction of plant symbiotic developmental responses. Plant Cell6, 1357-1374.
Baker, D.D., and Mullin, B.C. (1992). Actinorhizal symbioses. In Eio- logical Nitrogen Fixation, G. Stacey, R.H. Eurris, and H.J. Evans, eds (New York: Chapman and Hall), pp. 259-292.
Barker, D., Blanchi, S., Blondon, F., Dattbe, Y., Duc, G., Essad, S., Flament, P., Gallusci, P., GBnier, O., Guy, P., Muel, X., Tourneur, J., Dharib, J., and Huguet, T. (1990). Medicago troncatula, a model plant for studying the molecular genetics of the Rhizobium-legume symbiosis. Plant MOI. Biol. Rep. 8, 40-49.
Bassett, E., Goodman, R.N., and Novacky, A. (1977). Ultrastructure of soybean nodules. 1. Release of rhizobiafrom the infection thread. Can. J. Microbiol. 23, 573-582.
41, 1199-1206.

Rhizobial and Actinorhizal Symbioses 1909
Bauchrowltr, M.A., Bahr, D.G., and hrchet, 0. (1996). Lectin genes are expressed throughout root nodule development and during nitrogen-fixation in the Rhizobium-Medicago symbiosis. Plant J. 9,
Bauer, P., Crespl, M.D., Szecsl, J., Alllson, L.A., Schultze, M., Ratet, P., Kondomsl, E., and Kondorosl, A. (1994). Alfalfa ENOD12genes are differentially regulated during nodule development by Nod fac- tors and Rhizobium invasion. Plant Physiol. 105, 53-62.
Benaben, V., Duc, G., Lefebvre, V., and Huguet, T. (1995). TE7, an inefficient symbiotic mutant of Medicago truncatula Gaertn cv Jemalong. Plant Physiol. 107, 53-62.
Benson, D.R., and Sllvester, W.B. (1993). Biology of Frankia strains, actinomycete symbionts of actinorhizal plants. Microbiol. Rev. 57,
Berg, R.H., and McDowell, L. (1987a). Endophyte differentiation in Casuarina actinorhizae. Protoplasma 136, 104-1 17.
Berg, R.H., and McDowell, L. (1987b). Cytochemistry of the wall of infected cells in Casuarina actinorhizae. Can. J. Bot. 66,2038-2047.
Bergersen, F.J. (1974). Formation and function of bacteroids. In The Biology of Nitrogen Fixation, A. Quispel, ed (Amsterdam: North- Holland Publishing Co.), pp. 473-498.
Bergman, B., Johansson, C., and WderbPck, E. (1992). The Nostoc-Gunnera symbiosis. New Phytol. 122, 379-400.
Berry, A.M., and Sunell, L.A. (1990). The infection process and nod- ule development. In The Biology of Frankia and Actinorhizal Plants, C.R. Schwintzer and J.D. Tjepkema, eds (New York Academic Press),
Berry, A.M., Harriot, O.T., Moreau, R.A., Osman, S.F., Benson, D.R., and Jones, A.D. (1993). Hopanoid lipids compose the Frankia vesicle envelope, presumptive barrier of oxygen diffusion to nitrogenase. Proc. Natl. Acad. Sci. USA 90, 6091-6094.
Beswetherlck, Y.T., and Blshop, C.D. (1993). An ultrastructural study of tomato roots inoculated with pathogenic and nonpathogenic necw trophic fungi and a saprophytic fungus. Plant Pathol. 42, 577-588.
Bhuvaneswarl, T.V., Bhagwat, A.A., and Bauer, W.D. (1981). Tran- sient susceptibility of root cells in four common legumes to nodulation by rhizobia. Plant Physiol. 68, 1144-1149.
Blakely, L.M., and Evans, T.A. (1979). Cell dynamics studies on the pericycle of radish seedling roots. Plant Sci. Lett. 14, 79-83.
Blakely, L.M., Blakely, R.M., Colowlt, P.M., and Elllott, D.S. (1988). Experimental studies on lateral root formation in radish seedling rmts. Plant Physiol. 87, 414-419.
Blauenfeldt, J., Joshl, P.A., Gmsshoff, P.M., and Caetano-Anollts, 0. (1994). Nodulation of white clover (Tifolium repens) in the absence of Rhizobjum. Protoplasma 179, 106-1 10.
Bradbury, S.M., Peterson, R.L., and Bowley, S.R. (1993). Further evidence for a correlation between nodulation genotypes in alfalfa (Medicago sativa L.) and mycorrhiza formation. New Phytol. 124,
Burgess, D., and Peterson, R.L. (1987). Effect of nutrient conditions on root nodule development in Alnus laponica. Can. J. Bot. 65,
Burggraaf, A.J.P., van der Linden, J., and Tak, T. (1983). Studies on the localization of infectible cells on Alnus glutinose roots. Plant Soil 74, 175-188.
Burrls, R.H. (1991). Nitrogenases. J. Biol. Chem. 266, 9339-9342. Caetano-Anollts, G., and Gresshoff, P.M. (1991). Plant genetic con-
31-43.
293-319.
pp. 61-81.
665-673.
1658-1670.
trol of nodulation. Annu. Rev. Microbiol. 45, 345-382.
Caetano-Anollts, G., Papamzzl, E.T., and Gresshoff, P.M. (1991). Mature nodules and root tips control nodulation in soybean. J. Plant Physiol. 137, 389-396.
Callaham, D., and Torrey, J.G. (1977). Prenodule formation and pri- mary nodule development in roots of Comptonia (Myricaceae). Can. J. Bot. 51, 2306-2318.
Capellano, A., Dequatre, E., Valla, G., and Molroud, A. (1987). Root- nodules formation by Bnicillium sp. on Alnus glutinose and Alnus incana. Plant Soil 104, 45-51.
Chandler, M.R., Date, R.A., and Roughley, R.J. (1982). lnfection and root nodule development in Stylosanthes species by Rhizobium. J.
Cheon, C.4, Lee, N.G., Slddlque, A.B.M., Bal, A.K., and Verma, D.P.S. (1993). Roles of plant homologs of Rablp and Rab7p in the biogenesis of the peribacteroid membrane, a subcellular compart- ment formed de novo during root nodule symbiosis. EMBO J. 12, 4125-4135.
Cheon, C.4, Hong, Z., and Verma, D.P.S. (1994). Nodulin-24 follows a nove1 pathway for integration into the peribacteroid membrane in soybean root nodules. J. Biol. Chem. 269, 6598-6602.
Clark, T.A., Zeyen, R.Y., Smlth, A.G., Bushnell, W.R., Szabo, L.Y., and Vance, CP. (1993). Host response gene transcript accumula- tion in relation to visible cytological events during Erysiphe gfaminis attack in isogenic barley lines differing at the Ml-a locus. Physiol. MOI. Plant Pathol. 43, 283-298.
Cooper, J.B., and Long, S.R. (1994). Morphogenetic rescue of Rhizo- bium meliloti nodulation mutants by trans-zeatin secretion. Plant Cell
Culllmofe, J.V., and Bennett, M.J. (1988). The molecular biology and biochemistry of plant glutamine synthetase from root nodules of Phaseolus vu/gafis L. and other legumes. J. Plant Physiol. 132, 387-393.
Dart, P.J. (1974). The infection process. In The Biology of Nitrogen Fixation, A. Quispel, ed (Amsterdam: North-Holland Publishing Co.),
Day, D.A., and Copeland, L. (1991). Carbon metabolism and com- partmentation in nitrogen-fixing legume nodules. Plant Physiol. Biochem. 29, 185-201.
de Bruljn, F.J. (1989). The unusual symbiosis between the diazotrophic stem-nodulating bacterium Azorhjzobium cauljnodans ORS571 and its host, the tropical legume Sesbanie mstreta: Plant and bacterial aspects. In Plant-Microbe Interactions, T. Kosuge and E.W. Nester, eds (New York: Macmillan), pp. 457-493.
de Farla, S.M., Hay, G.T., and Sprent, J.I. (1988). Entry of rhizobia into roots of Mimosa scabmlla Bentham occurs between epidermal cells. J. Gen. Microbiol. 134, 2291-2296.
Delves, A.C., Hlgglns, A., and Gresshoff, P.M. (1987a). Shoot con- trol of nodulation in a number of mutant soybeans (Glycine max L. Merr.). Aust. J. Plant Physiol. 14, 689-694.
Delves, A.C., Hlgglns, A., and Gresahoff, P.M. (1987b). Supernodu- lation in interspecific grafts between Glycine max (soybean) and Glycine soja. J. Plant Physiol. 128, 473-478.
Denison, R.F., and Layzell, D.B. (1991). Measurement of legume nodule respiration and O2 permeability by noninvasive spectm- photometry of leghemoglobin. Plant Physiol. 96, 137-143.
Diout, D., Ghe*l, H., Prln, Y., Franche, C, Duhoux, E., and Bogusz, D. (1995). Hairy root nodulation of Casuarina glauca: A system for
EXP. Bot. 33, 47-57.
6, 215-225.
pp. 381-429.

1910 The Plant Cell
the study of symbiotic gene expression in an actinorhizal tree. MOI. Plant-Microbe Interact. 8, 325-349.
Djordjevlc, MA., Gabrlel, D.W., and Rolfe, 8.0. (1987). Rhizobium- The refined parasite of legumes. Annu. Rev. Phytopathol. 25,
Dobrltsa, S.V., and Novlk, S.N. (1992). Feedback regulation of nod- ule formation in Hippophae rhamnoides. Plant Soil 144, 45-50.
Doyle, J.J. (1994). Phylogeny of the legume family: An approach to understanding the origins of nodulation. Annu. Rev. Ecol. Syst. 25,
Duc, E., and Messager, A. (1989). Mutagenesis of pea (Pisum seti- vum L.) and the isolation of mutants for nodulation and nitrogen fixation. Plant Sci. 60, 207-213.
Duc, E., Rouvelot, A., Glanlnaul-Pearson, V., and Glanlnazzl, S. (1989). First report of non-nodulating plant mutants (Myc-) obtained in pea (Pisum sativum L.) and fababean (Vicie feba L.). Plant Sci.
Ehrhardt, D.W., Wals, R., and Long, S.R. (1996). Calcium spiking in plant root hairs responding to Rhizobium nodulation signals. Cell
Flemlng, A.I., Wlttenberg, J.B., Wlttenberg, B.A., Dudman, W.F., and Appleby, C.A. (1987). The purification, characterization and ligand-binding kinetics of hemoglobins from root nodules of the non- leguminous Cesuerina giauca-Fmnkia symbiosis. Biochim. Biophys. Acta 911, 209-220.
Franssen, H., Mylona, I?, Pawlowskl, K., van de Sande, K., Helstra, H., Geurts, R., Kozlk, A., Matvlenko, M., Yang, W.-C., Hadrl, A.-E., Martlnez-Abarca, F., and Blssellng, T. (1995). Plant genes involved in root nodule development on legumes. Philos. Trans. R. SOC. Lond. B 350, 101-107.
Funke, R.P., Kolchlnsky, A., and Gresshoff, P.M. (1994). High EDTA concentrations cause entrapment of small DNA molecules in the compression zone of pulsed field gels, resulting in smaller than ex- pected insert sizes in YACs prepared from size selected DNA. Nucleic Acids Res. 22, 2708-2709.
Gherbl, H., Duhoux, E., Franche, C., Pawlowskl, K., Berry, A., and Bogusz, D. (1997). Cloning of a full-length symbiotic hemoglobin cDNA and in situ localization of the corresonding mRNA in Casua- rina glauca root nodule. Physiol. Plant., in press.
Glanlnaui-Pearson, V. (1996). Plant cell responses to arbuscular mycorrhizal fungi: Getting to the roots of the symbiosis. Plant Cell
Glaninaul-Pearson, V., Glanlnaul, S., Gulllemln, J.P., Rouvelot, A., and Duc, 0. (1991). Genetic and cellular analysis of resistance to vesicular arbuscular (VAI mycorrhizal fungi in pea mutants. In Advances in Molecular Genetics of Plant-Microbe Interactions, H. Hennecke and D.P.S. Verma, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp. 336-342.
Goetllng-Mlnesky, M.P., and Mullln, B.C. (1994). Differential gene expression in an actinorhizal symbiosis: Evidence for a nodule- specific cysteine proteinase. Proc. Natl. Acad. Sci. USA 91,
Gordon, A.J. (1992). Carbon metabolism in the legume nodule. In Car- bon Partitioning within and between Organisms, C.J. Pollock and J.F. Farrar, eds (Oxford, UK: BlOS Scientific Publishers), pp. 133-162.
Guan, C., Akkermans, A.D.L., vsn Kammen, A., Blssellng, T., and Pawlowskl, K. (1997a). ag73 is expressed in Alnus glutinosa nod- ules in infected cells during endosymbiont degradation and in the nodule pericycle. Physiol. Plant., in press.
145-168.
325-349.
60, 215-222.
85, 673-681.
8, 1871-1883.
9891-9895.
Guan, C., Ribeiro, A,, Akkermans, A.D.L., Jlng, X., van Kammen, A., Blssellng, T., and Pawlowskl, K. (1997b). Nitrogen metabolism in actinorhizal nodules of Alnus glutinosa: Expression of glutamine synthetase and acetylornithine transaminase. Plant MOI. Biol., in press.
Hafeez, F., Akkermans, A.D.L., and Chaudhary, A.H. (1984). Ob- servations on the ultrastructure of frankla sp. in root nodules of Datisce cannabina L. Plant Soil 79, 383-402.
Handberg, K., and Stougaard, J. (1992). Lotus japonicus, an autoga- mous, diploid legume species for classical and molecular genetics. Plant J. 2, 487-496.
Hansen, A.P., Peoples, M.B., Brown, P.H., Carroll, B.J., and Grssshoft, P.M. (1990). Nitrogen partitioning during early develop- ment of supernodulating soybean (Glycine max (L.) Merrill) mutants and their wild-type parent. J. Exp. Bot. 41, 1239-1244.
Hansen, A.P., Yoneyama, T., and Kouchl, H. (1992). Short-term ni- trate effects on hydroponically-grown soybean cv. Bragg and its supernodulating mutant: li. Distribution and respiration of recently- fixed 13C-labeled photosynthate. J. Exp. Bot. 43, 9-14.
Harrlson, M.J., and Van Buuren, M.L. (1995). Aphosphate transporter from the mycorrhizal fungus Glomus vemifoftne. Nature 378,626-629.
Heldstra, R., Geurts, I?., Frsnssen, H., Spalnk, H.P., van Kammen, A,, and Blssellng, T. (1994). Root hair deformation activity of nodu- lation factors and their fate on Vicia setive. Plani Physiol. 105,787-797.
Hlrel, E., Mlao, G.H., and Verme, D.P.S. (1993). Metabolic and de- velopmental control of glutamine synthetase genes in legumes and non-legume plants. In Control of Plant Gene Expression, D.P.S. Verma, ed (Boca Raton, FL: CRC Press), pp. 443-458.
Hlrsch, A.M. (1992). Developmental biology of legume nodulation. New
Hunt, S., and Leyzell, 0.6. (1993). Gas exchange of legume nodules and the regulation of nitrogenase activity. Annu. Rev. Plant Physiol. Plant Moi. Biol. 44, 483-511.
Huss-Danell, K., and Begman, E. (1990). Nitrogenase in Fmnkiafrom root nodules of Alnus incana (L.) Moench: lmmunolocalization of the Fe- and MoFe-proteins during vesicle differentiation. New Phytol. 116, 443-455.
Jacobs, F.A., Zhang, M., Fortln, M.G., and Verma, D.P.S. (1987). Severa1 nodulins of soybean share structural domains but differ in their subcellular locations. Nucleic Acids Res. 15, 127l-1280.
Jacobsen, E., and Feenstra, W.J. (1984). A new pea mutant with effi- cient nodulation in the presence of nitrate. Plant Sci. Lett. 33,337-344.
Jacobsen-Lyon, K., dstergaard Jensen, E., J#rgensen, J.-E., Mamker, K.A., Peacock, W.J., and Dennls, E.S. (1995). Symbi- otic and nonsymbiotic hemoglobin genes of Casuarina glauca. Plant Cell 7, 213-223.
Phytol. 122, 2l1-237.
James, E.K., Sprent, J.I., Sutherland, J.M., Yclnroy, S.G., snd Mlnchln, F.R. (1992). The structure of nitrogen fixing root nodules on the aquatic mimosoid legume Neptunia plena. Ann. Bot. 69,
Joshl, P.A., Caetano-Anoll&, O., Graham, E.T., and Gmsshoft, EM. (1993). Ultrastructure of transfer cells in spontaneous nodules of al- falfa (Medicago setive). Protoplasma 172, 64-76.
Journet, E.P., Plchon, M., Dedleu, A,, de Bllly, F., Ttuchet, G., and Barker, D.G. (1994). Rhizobium meliloti Nod factors elicit cell-specific transcription of the ENODl2 gene in transgenic alfalfa. Plant J. 6,
173-180.
241-249.

Rhizobial and Actinorhizal Symbioses 191 1
Kljne, J.W. (1992). The Rhizobium infection process. In Biological Nitro- gen Fixation, G. Stacey, R.H. Burris, and H.J. Evans, eds(New York: Chapman and Hall), pp. 349-398.
Kiss, G.B., Csanbdi, G., KBlmBn, K., Kalb, I?, and b k & z , L. (1993). Construction of a basic genetic map for alfalfa using RFLP, RAPD, isozyme and morphological markers. MOI. Gen. Genet. 238,129-137.
Kleemann, O., Alskog, G., Berry, A.M., and Huss-Danell, K. (1994). Lipid wmposition and nitrogenase activity of symbidic Frenkia (Alnus incana) in response to different oxygen concentrations. Protoplasma
Knowlton, S., Berry, A., and Torrey, J.G. (1980). Evidence that as- sociated soil bacteria may influence root hair infection of actinorhizal plants by frankia. Can. J. Microbiol. 26, 971-977.
Kolde, R.T. (1991). Nutrient supply, nutrient demand and plant response to mycorrhizal infection. New Phytol. 117, 365-386.
Kouchi, H., and Hata, S. (1993). lsolation and characterization of nove1 cDNAs representing genes expressed at early stages of soybean nodule development. MOI. Gen. Genet. 238, 106-119.
Lancelle, S.A., and Torrey, J.G. (1984a). Early development of Rhizo- bium-induced root nodules of Parasponia rigida. 1. lnfection and early nodule initiation. Protoplasma 123, 26-37.
Lancelle, S.A., and Torrey, J.G. (1984b). Early development of Rhizobium-induced root nodules of Parasponia rigida. II. Nodule mor- phogenesis and symbiotic development. Can. J. Bot. 63, 25-35.
LaRue, T.A., and Weeden, N.F. (1994). The symbiosis genes of the host. In Proceedings of the First European Nitrogen Fixation Con- ference, G.B. Kiss and G. Endre, eds (Szeged, Hungary: Officina Press), pp. 147-151.
Laskowskl, MiJ., Williams, M.E., Nusbaum, H.C., and Sussex, I.M. (1995). Formation of lateral root meristems is a two-stage process. Development 121, 3303-3310.
Lee, N.-G., Steln, B., Suzukl, H., andVerma, D.P.S. (1993). Expres- sion of antisense nodulin-35 RNA in Vigna aconitifolia transgenic root nodules retards peroxisome development and affects nitrogen availability to the plant. Plant J. 3, 599-606.
Libbenga, K.R., and Bogers, R.J. (1974). Root-nodule morphogene- sis. In The Biology of Nitrogen Fixation, A. Quispel, ed (Amsterdam: North-Holland Publishing Co.), pp. 430-472.
Libbenga, K.R., Van Iren, F., Bogers, R.J., and Schraag-Lamers, M.F. (1973). The role of hormones and gradients in the initiation of cortex proliferation and nodule formation in Pisum sativum L. Planta
Liu, O., and Berry, A.M. (1991). Localization and characterization of pectic polysaccharides in roots and root nodules of Ceanothus spp. during intercellular infection by Frankia. Protoplasma 164, 93-101.
Long, S.R. (1996). Rhizobium symbiosis: Nod factors in perspective. Plant Cell 8, 1885-1898.
Long, S.R., and Staskawlu, B.J. (1993). Prokaryotic plant parasites. Cell 73, 921-935.
Markwei, C.M., and LaRue, T.A. (1992). Phenotypic characterization of sym8 and sym9, two genes conditioning non-nodulation in Pi- sum sativum Sparkle. Can. J. Microbiol. 38, 548-554.
Martin, EM., and Hilbert, J.L. (1991). Morphological, biochemical and molecular changes during ectomycorrhiza development. Experientia
Manrel, D.J., Torrey, J.G., and Ausubel, F.M. (1987). Rhizobium sym- biotic genes required for nodulation of legume and nonlegume hosts. Proc. Natl. Acad. Sci. USA 84, 1319-1323.
183, 107-115.
114, 29-39.
47, 321-331.
McCarty, DA. (1995). Genetic control and integration of maturation and germination pathways in seed development. Annu. Rev. Plant Physiol. Plant MOI. Biol. 46, 71-93.
Miller, I.M., and Baker, D.D. (1985). The initiation, development and structure of root nodules in Elaeagnus angusMolia L. (Elaeagnaceae). Protoplasma 128, 107-119.
Mylona, P., Pawlowskl, K., and Bisseling, T. (1995). Symbiotic nitro- gen fixation. Plant Cell 7, 869-885.
Naisbitt, T., James, E.K., and Sprent, J.I. (1992). The evolutionary significance of the legume genus Chamaecrista, as determined by nodule structure. New Phytol. 122, 487-492.
Ndoye, I., de Billy, F., Vasse, J., Dreyfus, B., and Ttuchet, G. (1994). Root nodulation of Sesbania rostrata. J. Bacteriol. 176, 1060-1068.
Newcomb, W. (1976). A correlated light and electron microscopic study of symbiotic growth and differentiation in Pisum sativum root nod- ules. Can. J. Bot. 54, 2163-2186.
Newcomb, W. (1981). Nodule morphogenesis and differentiation. Int. Rev. Cytol. 13 (suppl.), S247-S297.
Newcomb, W., and Pankhurst, C.E. (1982). Fine structure of ac- tinorhizal nodules of Coriarja arborea (Coriariaceae). N. Z. J. Bot.
Newcomb, W., and Wood, S.M. (1987). Morphogenesis and fine struc- ture of frankia (Actinomycetales): The microsymbiont of nitrogen-fixing actinorhizal root nodules. Int. Rev. Cytol. 109, 1-88.
Olsen, P., Collihs, M., and Rice, W. (1992). Surface antigenspresent on vegetative Rhizobium meliloti cells may be diminished or absent when cells are in the bacteroid form. Can. J. Microbiol. 38,506-509.
Parsons, R., Silvester, W.B., Harris, S., Gruijters, W.T.M., and Bullivant, S. (1987). Frankia vesicles provide inducible and abso- lute oxygen protection for nitrogenase. Plant Physiol. 83, 728-731,
Pathirana, S.M., and Tjepkema, J.D. (1995). Purification of hemo- globin from the actinorhizal root nodules of Myrica gale L. Plant Physiol. 107, 827-831.
Pawlowski, K., Guan, C., Ribeiro, A., van Kammen, A., Akkrmans, A.D.L., and Bisseling, T. (1994). Genes involved in Alnus glutinosa nodule development. In Proceedings of the First European Nitro- gen Fixation Conference, G.B. Kiss and G. Endre, eds (Szeged, Hupgary: Officina Press), pp. 220-224.
Postma, J.G., Jacobsen, E., and Feenstra, W.J. (1988). Three pea mutants with an altered nodulation studied by genetic analysis and grafting. J. Plant Physiol. 132, 424-430.
Puontl-Kaelas, J., Eriksson, T., and Engstrom, P. (1990). Produc- tion of transgenic pea (Pisum satium L.) plants by Agrobacferium tumefaciens-mediated gene transfer. Theor. Appl. Genet. 80,
Quirpel, A. (1992). A search for signals in endophytic microorgan- isms. In Molecular Signals in Plant-Microbe Communications, D.P.S. Verma, ed (Boca Raton, FL: CRC Press), pp. 471-491.
Racette, S., and Torrey, J.G. (1989). Root nodule initiation in Gym- nostoma (Casuarinaceae) and Shepherdia (Elaeagnaceae) induced by Frankia strain HFPGpll. Can. J. Bot. 67, 2873-2879.
Read, D.J., Lewis, D.H., Fllter, A.H., and Alexander, I. J. (1992). Mycor- rhizas in Ecosystems. (Oxford, UK: CAB International).
Ribeiro, A., Akkermans, A.D.L., van Kammen, A., Bisseling, T., and Pawlowski, K. (1995). A nodule-specific gene encoding a subtilisin-like protease is expressed in early stages of actinorhizal nodule development. Plant Cell 7, 785-794.
20, 93-103.
246-252.

1912 The Plant Cell
Rlbelro, A., Praekelt, U., Akkermans, A.D.L., Meacock, P.A., van Kammen, A., Blssellng, T., and Pawlowskl, K. (1996). Identifica- tion of agfhil, whose product is involved in biosynthesis of the thiamine precursor thiazole, in actinorhizal nodules of Alnus glutinosa. Plant J. 10, 361-368.
Roest, H.P., Goosen-de Roo, L., Wljffelman, C.A., de Maagd, R.A., and Lugtenberg, B.B.J. (1995). Outer membrane protein changes during bacteroid development are independent of nitrogen fixation and differ between indeterminate and determinate nodulating host plants of Rhizobium leguminosarum. MOI. Plant-Microbe Interact.
Rlihrlg, H., Schmidt, J., Walden, R., Czaja, I., Mlklasevlcs, E., Wleneke, U., ScheII, J., and John, M. (1995). Growth of tobacco protoplasts stimulated by synthetic lipo-chitooligosaccharides. Science 269, 841-843.
Rolfe, B.G., and Shlne, J. (1984). Rhizobium-Leguminosae symbio- sis: The bacterial point of view. In Genes lnvolved in Microbe-Plant Interactions, D.P.S. Verma and T.H. Hohn, eds (New York: Springer- Verlag), pp. 95-128.
Rost, T.L., Jones, T.J., and Falk, R.H. (1988). Distribution and rela- tionship of cell division and maturation events in Pisum sativum (Fabaceae) seedling roots. Am. J. Bot. 75, 1571-1583.
Roth, L.E., and Stacey, 0. (1989). Bacterium release into host cells of nitrogen-fixing soybean nodules: The symbiosome membrane comes from three sources. Eur. J. Cell Biol. 49, 13-23.
Sagan, M., Measager, A., and Duc, G. (1993). Specificity of the Rhizo- bium-legume symbiosis obtained after mutagenesis in pea (Pisum sativum L.). New Phytol. 125, 757-761.
Sagan, M., Huguet, T., and Duc, G. (1994). Phenotypic characteriza- tion and classification of nodulation mutants of pea (fisum sativum L.). Plant Sci. 100, 59-70.
Sagan, M., Morandl, D., Tarenghi, E., and Duc, G. (1995). Selection of nodulation and mycorrhizal mutants in the model plant Medicago truncatula (Gaertn.) after y-ray mutagenesis. Plant Sci. 111, 63-71.
Sdnchez, F., Padllla, J.E., Perez, H., and Lara, M. (1991). Control of nodulin genes in root-nodule development and metabolism. Annu. Rev. Plant Physiol. Plant Mo\. 6\01. 42, 507-528.
Sato, S., Newell, C., Kolacz, K., Ikdo, L., Flner, J., and Hlnchee, M. (1993). Stable transformation via particle bombardment in two different soybean regeneration systems. Plant Cell Rep. 12,408413.
Schubert, K.R. (1986). Products of biological nitrogen fixation in higher plants: Synthesis, transport, and metabolism. Annu. Rev. Plant Phys- iol. 37, 539-574.
Schwintrer, C.R., Berry, A.M., and Disney, L.D. (1982). Seasonal patterns of root nodule growth, endophyte morphology, nitrogenase activity and shoot development in Myrica gale. Can. J. Bot. 60,
SBguln, A., and Lalonde, M. (1991). Expression of actinorhizins in the development of the Frankie-Alnus symbiosis. In Developments in Plant and Soil Sciences: Nitrogen Fixation, M. Polsinelli, R. Materassi, and M. Vincenzini, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp. 601-613.
Sequerra, J., Capellano, A., Faure-Raynaud, M., and Molroud, A. (1994). Root hair infection process and myconodule formation on Alnus incana by &nicilliUm nodositatum Valla. Can. J. Bot. 72, 955-975.
Sequerra, J., Capellano, A., Glanlnaul-Pearson, V., and Molroud, A. (1995). Ultrastructure of cortical root cells of Ahus incana infected by hnicillium nodositatum. New Phytol. 130, 545-555.
8, 14-22.
746-757.
Sllvester, W.B., and Harrls, S.L. (1989). Nodule structure and nitrogenase activity of Coriaria arborea in response to varying pO2. Plant Soil 118, 97-109.
Silvester, W.B., Hanls, S.L., and Tjepkema, J.D. (1990). Oxygen regu- lation and hemoglobin. In The Biology of Frankia and Actinorhizal Plants, C.R. Schwintzer and J.D. Tjepkema, eds (New York: Aca- demic Press), pp. 157-176.
Smlt, O., de Koster, C.C., Schrlpsema, J., Spalnk, H.P., van Brussel, A.A., and Kljne, J.W. (1995). Uridine, a cell division factor in pea roots. Plant MOI. Biol. 29, 869-873.
Soltis, D.E., Soltls, RS., Morgan, D.R., Swensen, S.M., Mullln, B.C., Dowd, J.M., and Martln, P.G. (1995). Chloroplast gene sequence data suggest a single origin of the pfedisposition for symbiotic nitro- gen fixation in angiosperms. Proc. Natl. Acad. Sci. USA 92,
Spalnk, H.P., Sheeley, D.M., van Bruasel, A.A.N., Glushka, J., York, W.S., Tak, T., Gelger, O., Kennedy, E.P., Relnhold, V.N., and Lugtenberg, B.J.J. (1991). A nove1 highly unsaturated fatty acid moi- ety of lipo-oligosaccharide signals determines host specificity of Rhizobium. Nature 354, 125-130.
Sprent, J.I. (1989). Which steps are essential for the formation of func- tional legume nodules? New Phytol. 111, 129-153.
Streeter, J.G. (1993). Translocation-A key factor limiting the efficiency of nitrogen fixation in legume nodules. Physiol. Plant. 87, 616-623.
Subba-Rao, N.S., Mateos, P.F., Baker, D., Pankratr, H.S., Palma, J., Dano, F.B., and Sprent, J.I. (1995). The unique root-nodule sym- biosis between Rhizobium and the aquatic legume, Neptunia natans (L. F.) Druce. Planta 196, 311-320.
Suharjo, U.K.J., and Tjepkema, J.D. (1995). Occurrence of hemo- globin in the nitrogen-fixing root nodules of Alnusglutinosa. Physiol. Plant. 95, 247-252.
Swensen, S. (1996). The evolution of actinorhizal symbioses: Evidence for multiple origins of the symbiotic association. Am. J. Bot. 83,
Takats, S.T. (1990). Early autoregulation of symbiotic root nodulation in soybeans. Plant Physiol. 94, 865-869.
Thomas, M.R., Rose, R.J., and Nolan, K.E. (1992). Genetic trans- formation of Medicago truncatula using Agrobacterium with genetically modified RI and disarmed TI plasmids. Plant Cell flep.
Thykjaer, T., Stiller, J., Handberg, K., Jones, J., and Stougaard, J. (1995). The maize transposable element Ac is mobile in the le- gume Lotus japonicus. Plant MOI. Biol. 27, 981-993.
Tjepkema, J.D., and Asa, D.J. (1987). Total and CO-reactive heme content of actinorhizal nodules and the roots of some non-nodulated plants. Plant Soil 100, 225-236.
Torrey, J.G. (1950). The induction of lateral roots by indoleacetic acid and root decapitation. Am. J. Bot. 37, 257-264.
Trlnick, M.J. (1980). Relationships amongst the fast-growing rhizobia of Lablab purpureus, Leucaena leucocephala, Mimosa spp., Acacia farnesiana and Sesbania grandiflora and their affinities with other rhizobial groups. J. Appl. Bacteriol. 49, 39-53.
nuchet, O., Barker, D.G., Camut, S., de Bllly, F., Vasse, J., and Huguet, T. (1989). Alfalfa nodulation in the absence of Rhizobium. Moi. Gen. Genet. 219, 65-68.
Tsien, H.C., Dreyfus, B.L., and Schmidt, E.L. (1983). lnitial stages in the morphogenesis of nitrogen-fixing stem nodules of Sesbania rostrata. J. Bacteriol. 156, 888-897.
2647-2651.
1503-1512.
11, 113-117.

Rhizobial and Actinorhizal Symbioses 191 3
van Brussel, A.A.N., Bakhulzen, R., van Spronsen, P.C., Spalnk, H.P., Tak, T., and Lugtenberg, B.J.J. (1992). lnduction of pre-infection thread structures in the leguminous host plant by mitogenic lipo- oligosaccharides of Rhizobium. Science 257, 70-71.
van de Sande, K., Pawlowski, K., Czaja, I., Wleneke, U., Schell, J., Schmldt, J., Walden, R., Matvlenko, M., Welllnk, J., van Kammen, A., Franssen, H., and Blssellng, T. (1996). Modifica- tion of phytohormone response by a peptide encoded by ENOD40 of legumes and a nonlegume. Science 273, 370-373.
van de Wlel, C., Scheres, B., Franssen, H., van Llemp, M.J., van Lammeren, A., vsn Kammen, A., and Blsseling, T. (1990). The early nodulin transcript ENOD2 is located in the nodule parenchyma (inner cortex) of pea and soybean root nodules. EMBO J. 9, 1-7.
van Ghelue, M., Rlbelro, A., Solhelm, E., Akkermans, A.D.L., Bisseling, T., and Pawlowski, K. (1996). Sucrose synthase and ene lase expression in actinorhizal nodules of Ahus glutinosa: Comparison with legume nodules. MOI. Gen. Genet. 250,437-446.
van Ghelue, M., Lwaas, E., RingO, E., and Solhelm, B. (1997). Early interactions between Anus glutinosa (L.) Gaertn. and Frankia strain Arl3. Physiol. Plant., in press.
van Kammen, A. (1984). Suggested nomenclature for plant genes in- volved in nodulation. Plant MOI. Biol. Rep. 2, 43-45.
van Spmnsen, P.C., van Brussel, A.A.N., and Kljne, J.W. (1994). Nod factors produced by Rhizobium leguminosarum biovar viciae induce ethylene-related changes in root cortical cells of Vicia sativa ssp. nigra. Eur. J. Cell Biol. 68, 463-469.
Vasse, J., de Bllly, F., Camut, S., and Tkuchet, O. (1990). Correlation between ultrastructural differentiation of bacteroids and nitrogen fix- ation in alfalfa nodules. J. Bacteriol. 172, 4295-4306.
Vlkman, P.A. (1992). The symbiotic uesicle is a major site for respira- tion in frankia from Alnus incana root nodules. Can. J. Microbiol.
Wall, L.G., and Huss-Danell, K. (1997). Regulation of nodulation in
38, 779-784.
Nnus incena-Frankia symbiosis. Physiol. Plant., in press.
Weaver, C.D., Shower, N.H., Louls, C.F., and Roberts, D.M. (1994). Nodulin-26, a nodule-specific symbiosome membrane protein from soybean, is an ion channel. J. Biol. Chem. 269, 17858-17862.
Webb, K.J., Skbt, L., and J@gensen, 8. (1994). Plant genes involved in nodule development and nitrogen fixation. In Proceedings of the First European Nitrogen Fixation Conference, G.B. Kiss and G. Endre, eds (Szeged, Hungary: Officina Press), pp. 234-238.
Wllson, C.W., Long, F., Maruoka, E.M., and Cooper, J.B. (1994). A new proline-rich early nodulin from Medicago truncatule is highly expressed in nodule meristematic cells. Plant Cell 6, 1265-1275.
Wltty, J.F., Mlnchln, F.R., Sk#t, L., and Sheely, J.E. (1986). Nitro- gen fixation and oxygen in root nodules. Oxf. Surv. Plant MOI. Cell Biol. 3, 275-315.
Yang, W.4 , Horvath, B., Hontelez, J., van Kammen, A,, and Blsseling, T. (1991). ln situ localization of Rhizobium mRNAs in pea root nodules: nifA and nifH localization. MOI. Plant-Microbe Inter- act. 7, 276-281.
Yang, W.-C., Katlnakls, P., Hendrlks, P., Smolders, A., de Vrles, F., Spee, J., van Kammen, A.. Blssellng, T., and Franssen, H. (1993). Characterization of GmENOD40, a gene showing nove1 patterns of cell-specific expression during soybean nodule development. Plant
Yang, W.-C., de Blank, C., Mesklene, I., Hlrt, H., Bakker, J., van Kammen, A., Franssen, H., and Blssellng, T. (1994). Rhizobium Nod factors reactivate the cell cycle during infection and nodule primordium formation, but the cycle is only completed in primor- dium formation. Plant Cell 6, 1415-1426.
Zeng, S., and Tjepkema, J.D. (1994). The wall of the infected cell may be the major diffusion barrier in nodules of Myrica gale L. Soil Biol. Biochem. 5, 633-639.
Zeng, S., and Tjepkema, J.D. (1995). The resistance of the diffusion barrier in nodules of Myrica gale L. changes in response to temper- ature but not to partia1 pressure of 02. Plant Physiol. 107,
Zeng, S., Tjepkema, J.D., and Berg, R.H. (1989). Gas diffusion path- way in nodules of Casuarine cunninghmiana. Plant Soilll8,119-123.
J. 3, 573-585.
1269-1275.

DOI 10.1105/tpc.8.10.1899 1996;8;1899-1913Plant Cell
K. Pawlowski and T. BisselingRhizobial and Actinorhizal Symbioses: What Are the Shared Features?
This information is current as of March 25, 2015
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists




















