
The distribution of meiofauna on back‐reef sandy platforms in the Maldives (Indian Ocean)
16
ORIGINAL ARTICLE The distribution of meiofauna on back-reef sandy platforms in the Maldives (Indian Ocean) Federica Semprucci 1 , Paolo Colantoni 2 , Giuseppe Baldelli 2 , Marco Rocchi 1 & Maria Balsamo 1 1 Dipartimento di Scienze dell’Uomo, dell’Ambiente e della Natura (DiSUAN), Universita ` di Urbino ‘Carlo Bo’, Urbino, Italy 2 Dipartimento di Scienze Geologiche, Tecnologie Chimiche e Ambientali (DiGeoTeCA), Universita ` di Urbino ‘Carlo Bo’, Urbino, Italy Problem In tropical regions, most meiofauna research has been carried out in the Pacific Ocean (Ndaro & O ´ lafsson 1999; Raes et al. 2007 and references within). Given the pres- ence of heterogeneous coral reef structures and carbonate sediments, atolls offer a wide variety of micro-habitats for meiofauna. Despite this, relatively few quantitative studies have been published on the shallow sublittoral sandy sedi- ments of atolls (Netto et al. 1999a,b; Guo et al. 2007 and references within). Determinant variables that can influence the meioben- thic assemblage include: mean sediment grain size, sort- ing, oxygen content, position of the redox potential Keywords Back-reef platforms; free-living nematodes; Maldives; meiofauna; sediment grain size. Correspondence Federica Semprucci, Dipartimento di Scienze dell’Uomo, dell’Ambiente e della Natura (DiSUAN) Universita ` di Urbino ‘Carlo Bo’, loc. Crocicchia – 61029 Urbino, Italy. E-mail: [email protected] Accepted: 6 April 2010 doi:10.1111/j.1439-0485.2010.00383.x Abstract The Maldivian archipelago comprises some of the most characteristic and significant world atoll systems, but the meiobenthic assemblages of these islands continue to be largely unknown. To investigate variations in meiofaunal spatial distribution and biodiversity in back-reef platforms, three transects were studied, two at Felidhoo (the north and east sides) and one at South Male ´. The sedimentological features of the samples obtained were also analyzed to further current knowledge on the relationships that exist between sediments and meio- fauna. Our results reveal that the meiofaunal assemblage at these locations is well diversified and includes 20 major taxa. Nematodes and copepods are dom- inant, together forming 68% of the total meiofauna, followed by platyhelmin- thes, polychaetes and ostracods. The nematode assemblage is very rich and composed of 34 families and 123 genera – 96 of which (78%) set new records for the Maldives. The structures of the meiofaunal and nematode assemblages are relatively similar on the ‘large-scale’ level (i.e. when the different platforms are compared) and reveal a low b-diversity. However, significant dissimilarities were detected within each platform, emphasizing that such ‘small-scale’ differ- ences are the main factors determining the structure of the meiofauna and, in particular, the nematode assemblages. Although significant differences were not detected between the transects, greater levels of dissimilarity were recognized at North Felidhoo. Here, the building of inclined deposit layers plays a significant role in increasing the heterogeneity of the platform habitats and sediments, confirming the great importance of sediment granulometry as an environmen- tal variable. Indeed, a close relationship is observed between meiofauna (espe- cially for the nematodes) and grain size, which appears to control the structure, diversity and trophic composition of the Maldivian meiofauna assemblages, thus highlighting the high biodiversity existing in the medium- coarse sands. Marine Ecology. ISSN 0173-9565 592 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH
Transcript of The distribution of meiofauna on back‐reef sandy platforms in the Maldives (Indian Ocean)

ORIGINAL ARTICLE
The distribution of meiofauna on back-reef sandy platformsin the Maldives (Indian Ocean)Federica Semprucci1, Paolo Colantoni2, Giuseppe Baldelli2, Marco Rocchi1 & Maria Balsamo1
1 Dipartimento di Scienze dell’Uomo, dell’Ambiente e della Natura (DiSUAN), Universita di Urbino ‘Carlo Bo’, Urbino, Italy
2 Dipartimento di Scienze Geologiche, Tecnologie Chimiche e Ambientali (DiGeoTeCA), Universita di Urbino ‘Carlo Bo’, Urbino, Italy
Problem
In tropical regions, most meiofauna research has been
carried out in the Pacific Ocean (Ndaro & Olafsson 1999;
Raes et al. 2007 and references within). Given the pres-
ence of heterogeneous coral reef structures and carbonate
sediments, atolls offer a wide variety of micro-habitats for
meiofauna. Despite this, relatively few quantitative studies
have been published on the shallow sublittoral sandy sedi-
ments of atolls (Netto et al. 1999a,b; Guo et al. 2007 and
references within).
Determinant variables that can influence the meioben-
thic assemblage include: mean sediment grain size, sort-
ing, oxygen content, position of the redox potential
Keywords
Back-reef platforms; free-living nematodes;
Maldives; meiofauna; sediment grain size.
Correspondence
Federica Semprucci, Dipartimento di Scienze
dell’Uomo, dell’Ambiente e della Natura
(DiSUAN) Universita di Urbino ‘Carlo Bo’, loc.
Crocicchia – 61029 Urbino, Italy.
E-mail: [email protected]
Accepted: 6 April 2010
doi:10.1111/j.1439-0485.2010.00383.x
Abstract
The Maldivian archipelago comprises some of the most characteristic and
significant world atoll systems, but the meiobenthic assemblages of these
islands continue to be largely unknown. To investigate variations in meiofaunal
spatial distribution and biodiversity in back-reef platforms, three transects were
studied, two at Felidhoo (the north and east sides) and one at South Male. The
sedimentological features of the samples obtained were also analyzed to further
current knowledge on the relationships that exist between sediments and meio-
fauna. Our results reveal that the meiofaunal assemblage at these locations is
well diversified and includes 20 major taxa. Nematodes and copepods are dom-
inant, together forming 68% of the total meiofauna, followed by platyhelmin-
thes, polychaetes and ostracods. The nematode assemblage is very rich and
composed of 34 families and 123 genera – 96 of which (78%) set new records
for the Maldives. The structures of the meiofaunal and nematode assemblages
are relatively similar on the ‘large-scale’ level (i.e. when the different platforms
are compared) and reveal a low b-diversity. However, significant dissimilarities
were detected within each platform, emphasizing that such ‘small-scale’ differ-
ences are the main factors determining the structure of the meiofauna and, in
particular, the nematode assemblages. Although significant differences were not
detected between the transects, greater levels of dissimilarity were recognized at
North Felidhoo. Here, the building of inclined deposit layers plays a significant
role in increasing the heterogeneity of the platform habitats and sediments,
confirming the great importance of sediment granulometry as an environmen-
tal variable. Indeed, a close relationship is observed between meiofauna (espe-
cially for the nematodes) and grain size, which appears to control the
structure, diversity and trophic composition of the Maldivian meiofauna
assemblages, thus highlighting the high biodiversity existing in the medium-
coarse sands.
Marine Ecology. ISSN 0173-9565
592 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

discontinuity (RPD) layer within the sediment, organic
content, the extent of bioturbation by the macrobenthos,
macrofaunal competition and predation, and water depth
(see Giere 2009 for review). Nematodes are usually the
dominant and ubiquitous meiofaunal taxonomic group,
with high densities, especially in fine sediments. Cope-
pods, on the other hand, become progressively more
abundant as grain size increases (Heip et al. 1985).
Accordingly, in tropical coarse sands with low silt con-
tent, harpacticoids usually constitute the most dominant
group, although polychaetes and oligochaetes may also be
present to a substantial degree (see Netto et al. 1999a).
The sediment composition may also play an important
role in controlling meiofaunal assemblage structure. Many
other, often rare, taxa may also occur in carbonate sands.
Indeed, temporary meiofauna and specialized members of
permanent meiofauna are more frequently found in these
sediments, which have been defined by Giere (2009) as ‘a
bonanza of fascinating meiofauna’. These biogenic sedi-
ments are structurally complex, relatively unsorted and
possess high porosity. The permeability of calcareous sand
has been proved to be markedly higher than that of sili-
ceous sand. High permeability favours the absorption of
nutrients, which gives rise to rich organic matter and
large quantities of microorganisms, two important feeding
resources for the meiofauna (Wild et al. 2005; Dahms
et al. 2007).
The Maldive Islands form the central part of the Cha-
gos-Maldives-Laccadive ridge in the Central Indian Ocean
and comprise some of the most characteristic and signifi-
cant atoll systems (Risk & Sluka 2000); their back-reef
platforms consist entirely of carbonate sediments, unlike
those of the Pacific Atolls. In spite of the great interest in
this archipelago, the information available about its meio-
fauna is mostly taxonomic (Gerlach 1961, 1962, 1963a,b,
1964; De Zio Grimaldi et al. 1999; Gallo et al. 2007). For
these reasons, a study of the meiofaunal and nematode
assemblages from three Maldivian back-reef sandy plat-
forms was conducted. Meiofauna were collected from
different geographical locations that are also characterized
by different geomorphological features: two transects were
placed on the Felidhoo atoll (north and east sides) and
one on South Male. The aims of the study were: (i) to
investigate the spatial patterns of the meiofaunal assem-
blages and biodiversity on a ‘small’ and ‘large’ scale in the
Maldives (i.e. within and between the platforms, respec-
tively) and (ii) to improve our understanding of the rela-
tionships that exist between meiofauna and sediment
types.
Study Area
The Maldive Islands rise up in the tropical Indian Ocean,
south-west of India, as a shallow carbonate system com-
posed of a chain of atolls which extend for approximately
800 km along the 73� meridian, between 7�06¢ N and
0�42¢ S (Fig. 1A,B). Their geological history is marked by
a complicated succession of sea-level changes, construc-
tions and erosions (Aubert & Droxler 1992) which have
given rise to carbonate sediment deposits that are over
A
B C
D
E
Fig. 1. Geographic location of the study
area and the sampling transects.
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 593

2000 m thick above an ancient and subsided volcanic
substratum (Duncan & Hargraves 1990). Seismic reflec-
tion profiles (Aubert & Droxler 1996) and the presence of
widespread terraces, notches and caves (Bianchi et al.
1996; Colantoni et al. 2003) reveal that the eroded sur-
faces, formed during the last Pleistocene glacial sea-level
low-stand, largely govern the shape of the present reef
formations. At the top of many of them, the reef flats are
wide or stunted, but are always accompanied by biogenic
sand and rubble, which accumulate and form ribbons and
low islands, especially during high energy wave-generated
events (Kench et al. 2005).
Sampling operations were carried out between 19 and 21
May 2005, during a scientific cruise organized by Albatros
Top Boat. Samples were collected in the South Male and Fe-
lidhoo Atolls (Central Eastern Maldives Archipelago) along
three transects located on their eastern rims (see Fig. 1).
The South Male transect (T1) (3�52.396¢ N–
73�27.531¢ E) ran seawards (direction 90�N) from the tip
of the narrow island of Maadhoo, which lies at the north
end of a large reef (Maadhoo Falhu) in the South Male
Atoll (Fig. 1C). This island was once one of the largest and
most populated islands of the Atoll, but it has now been
almost completely washed away (Godfrey 1996). Going off-
shore from the narrow beach, and furrowed by eroded
notches, a strip of outcropping beach rock was present. The
short foreshore, ending with scattered patches of elevated
sea grass beds (Cymodocea spp.), is followed by a large and
gentle sub-tidal sandy plain which reaches the reef flat as
far as 900 m from the shore. Samples were taken from six
stations positioned along this transect (Fig. 2A).
The North Felidhoo transect (T2) (3�33.600¢N–73�29.859¢ E) started at a sandy cay (or finolhu) on
Kudadhiggaru Falhu, a wide reef in the vicinity of the
large island of Alimathaa, near the southern edge of a pass
(kandu). Proceeding in a direction 50� N, T2 reached the
outside rim of the Felidhoo Atoll (Fig. 1D), and crossed
the sandy floor of a wide depression that is in the process
of being filled by advancing inclined coarse sandy layers.
Seven stations were analyzed along this transect (Fig. 2B).
A
B
C
Fig. 2. Section of the three transects and station locations.
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
594 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

The East Felidhoo transect (T3) (3�28.888¢N–73�42.200¢ E) crossed the Fotteyo Finolhu, a sandy
body 105 m from the reef edge on the northern side of
the pass, between Dhiggaru and Fotteyo Falhu, at the
eastern end of the Felidhoo Atoll (Fig. 1E). At this point,
the drop-off is only 1.5 m deep and is characterized by a
gentle rocky slope with blocks (Fig. 2C). Five stations
were sampled along this transect.
Material and Methods
Sample processing
At each station, samples for the quantitative analysis of
the meiofauna were collected by a SCUBA diver using a
hand-held piston corer (surface area 6 cm2) driven to a
depth of 5 cm and collected in triplicate. A fourth sam-
ple, intended for sediment analysis, was collected at each
station. Samples destined for meiofaunal analysis were
narcotized with a 7% magnesium chloride aqueous solu-
tion, fixed using a 4% formaldehyde solution (in buffered
seawater), stained with Rose Bengal and stored for subse-
quent processing.
The meiofauna were obtained by sieving the samples
through a 42-lm-mesh net, and animal extraction was
performed by flotation and multiple decantations. Centri-
fugation through a silica gel gradient (Ludox HS 30, den-
sity 1.18 gÆcm)3) was only carried out for samples with
fine sediments (Pfannkuche & Thiel 1988). The animals
were then transferred to a ‘Delfuss’ Petri dish with a
checkered bottom (200 squares, to make counting easier),
sorted into their major taxa under a Leica G26 stereomi-
croscope, and counted. About 100 nematode specimens
from each replicate were picked at random, placed in
glycerine and mounted as permanent slides. Identification
at the genus level was performed using a light microscope
equipped with Nomarski optics (Optiphoto-2 Nikon) and
aided by the pictorial keys of Platt & Warwick (1983,
1988), Warwick et al. (1998), the NeMys online identifi-
cation key (Deprez et al. 2004), and papers published by
Gerlach (1962, 1963a,b, 1964). Nematode trophic groups
were also defined according to Wieser (1953): 1A, selec-
tive deposit feeders; 1B, non-selective deposit feeders; 2A,
epistratum feeders; 2B, predators ⁄ omnivores.
Grain size analysis was performed on the collected sedi-
ments using a vibro-siever for fractions larger than 63 lm
and an X-ray analyzer for those smaller than 63 lm.
Using the percentages of each granulometric class
(defined at intervals of 1 u as the )log2 d mm; Krumbein
1934), histograms and cumulative curves were created,
from which percentiles were calculated; these measure-
ments were subsequently used to provide a statistical
characterization of the samples (according to Folk &
Ward 1957). The sediments were classified in accordance
with the Wentworth scale (Buchanan 1984).
Data analysis
Non-metric multi-dimensional scaling (nMDS) ordina-
tions derived from Bray–Curtis similarity matrices were
used to view differences in the structures of meiofaunal
and nematode assemblages between the stations located
in the different transects (on � transformed data). A two-
way nested ANOSIM (analysis of similarities) was used to
assess the statistical significance of any differences
between the transects and the stations. The SIMPER test
(cut-off of 50%) was utilized to determine the contribu-
tion of each meiofaunal taxa or nematode genus to the
total dissimilarity. Shannon’s diversity (H¢) and evenness
(J) indices (log2) were calculated to describe the nema-
tode assemblage structure. b-diversity (i.e. turnover diver-
sity, estimated as a % of Bray–Curtis dissimilarity; see
Gray 2000) was estimated using SIMPER and nMDS anal-
yses to provide a measure of genera dissimilarity between
the different transects and stations; a two-way nested
ANOSIM was used to assess the statistical significance of
any differences identified. All absolute data were pres-
ence ⁄ absence-transformed for the estimation of b-diver-
sity. An additional visual representation of diversity was
provided by a k-dominance curve, in which the abun-
dance of each genus was ranked in decreasing order of
dominance and cumulatively plotted. To evaluate the sig-
nificance of the differences in the meiofaunal and nema-
tode assemblages in relation to the sediment types
(irrespective of the station or transect of origin), a one-
way ANOSIM was used (on � transformed data). All of
the analyses referred to above were performed using the
software package PRIMER v. 5 (Clarke & Gorley 2001;
Clarke & Warwick 2001). Possible differences in the uni-
variate measures (Shannon, H¢, and Pielou, J, indices)
and in the abundance of nematode trophic groups were
evaluated using an analysis of variance (ANOVA). Prior
to statistical analysis, the logarithmic transformation
log(1 + x) was performed to normalize the data. Tukey’s
multiple-comparison tests were used when significant dif-
ferences (P < 0.05) were detected. Spearman’s correlation
analysis was utilized to test for significant correlations
between the various biological and sediment parameters
(SPSS v. 12 program).
Results
Sediment distribution
At South Male, the samples collected from St. 1 ⁄ 1(located on the foreshore) contained moderately sorted,
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 595

very coarse sands; those from St. 1 ⁄ 2 (situated amongst
the sea grasses) contained coarse and poorly sorted sand
(Fig. 2A; Table 1). Turning seawards, all of the remaining
stations (St. 1 ⁄ 3, 1 ⁄ 4, 1 ⁄ 5 and 1 ⁄ 6) were characterized by
poorly sorted, medium and coarse sands, with a ‘tail’ of
coarser material that is highlighted by their negative
asymmetries (Table 1). The sub-tidal beach of the finolhu
at North Felidhoo (St. 2 ⁄ 1 and 2 ⁄ 2) contained poorly
sorted, gravelly sands (Fig. 2B; Table 1). The deeper
depression which followed was covered by fine sands (St.
2 ⁄ 3 and 2 ⁄ 4), while the infilling inclined layers (St. 2 ⁄ 6)
were composed of moderately sorted, very coarse sands.
Towards the reef (St. 2 ⁄ 7, 2 ⁄ 8), the sea floor was covered
by coarse and medium sand (Table 1). Between the
finolhu and the reef top of East Felidhoo (St. 3 ⁄ 1 and
3 ⁄ 2), poorly sorted, medium sands were present, whilst
towards the lagoon (St. 3 ⁄ 4, 3 ⁄ 5 and 3 ⁄ 6), the sands were
coarse and poorly sorted (Fig. 2C; Table 1).
Meiofaunal and nematode assemblages
The North Felidhoo transect contained the greatest meio-
faunal abundances (on average 2463.62 ± 657.88
indÆ10 cm)2), whereas the values detected at the South
Male and East Felidhoo transects were lower and similar
(on average 1270.69 ± 375.70 and 1355.71 ± 353.34 indÆ10 cm)2, respectively) (Fig. 3). Overall, the meiofaunal
community appeared to be rich, with a total of 20 major
taxa present (Table 2). Copepods (adults and nauplii) and
nematodes were dominant in all transects and together
accounted for an average of 68% of the total meiofauna.
Other prevalent taxa were platyhelminthes, polychaetes
and ostracods, each one accounting for more than 3% of
the total assemblage (Fig. 4). Nematodes were only clearly
dominant at St. 2 ⁄ 3 and 2 ⁄ 4 (accounting for 89% and
63% of total assemblages, respectively), whereas the cope-
pods were particularly dominant at St. 2 ⁄ 6 and 2 ⁄ 1 (89%
and 77% of total assemblages, respectively), and, to a
lesser extent, at St. 1 ⁄ 6 and 1 ⁄ 1 (76% and 60%, respec-
tively) (Fig. 4A,B).
There were greater differences in the meiofaunal assem-
blage between the stations than between the transects
(ANOSIM, R = 0.61; P = 0.001, and R = 0.18; P = 0.023
for stations and transects, respectively). This phenomenon
is also visible in the nMDS plot of the meiofaunal assem-
blage (Fig. 5A), where the three transects did not appear
to be clearly separate despite the stations of the North
Felidhoo transect being the most scattered. Differences
between the meiofaunal assemblages relating to sediment
type were also found (ANOSIM, R = 0.34; P = 0.001)
(Table 3). The nematode community was well diversified
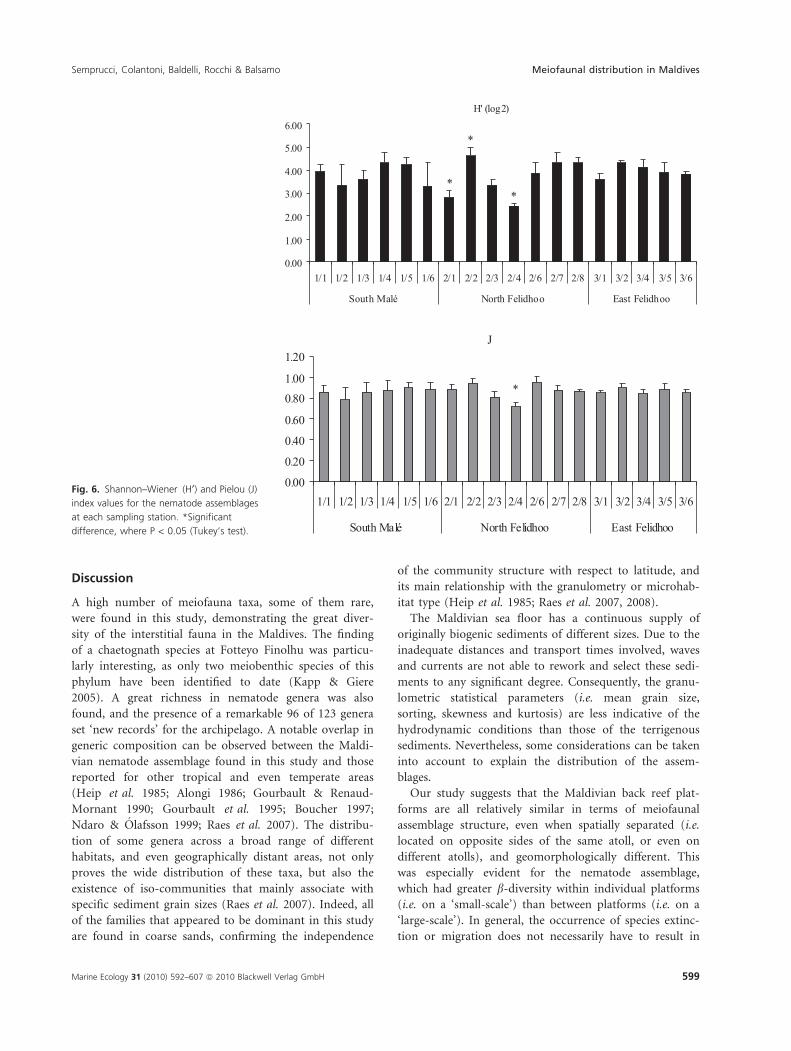
(H¢ from 4.62 to 2.16; J from 0.94 to 0.68) (Fig. 6), with
Table 1. Grain size parameters at each station.
station water depth (m) gravel % sand % pelite % mode Md mean size Mz sorting d skewness Sk1 kurtosis KG
1 ⁄ 1 0.50 15.32 83.02 1.66 0 0.00 1.08 0.10 0.98
1 ⁄ 2 0.90 6.49 86.21 7.30 1 1.03 1.78 0.27 1.31
1 ⁄ 3 0.60 3.49 94.41 2.10 2 0.99 1.22 )0.03 0.89
1 ⁄ 4 0.70 0.09 93.60 2.31 2 1.33 1.18 )0.15 0.98
1 ⁄ 5 0.50 9.77 85.61 4.63 2 0.87 1.52 )0.03 1.13
1 ⁄ 6 0.40 16.03 76.99 6.98 2 0.94 1.96 0.02 1.07
2 ⁄ 1 0.80 11.57 88.36 0.07 1 0.23 1.02 )0.12 1.07
2 ⁄ 2 1.30 14.25 82.76 2.98 1 0.53 1.37 )0.06 0.94
2 ⁄ 3 3.50 1.01 89.06 9.93 4 2.27 1.75 )0.01 1.12
2 ⁄ 4 5.00 0.11 95.18 4.71 3 2.81 0.82 0.03 1.01
2 ⁄ 6 1.50 16.61 82.03 1.36 0 )0.22 0.96 0.22 1.21
2 ⁄ 7 1.50 4.77 94.94 0.30 1 0.71 1.14 0.00 0.90
2 ⁄ 8 1.10 9.23 89.35 1.42 2 1.19 1.38 )0.20 0.98
3 ⁄ 1 0.60 2.40 92.99 4.61 3 1.67 1.33 )0.17 1.12
3 ⁄ 2 1.50 1.54 96.47 2.00 2 1.90 0.97 0.00 1.09
3 ⁄ 4 0.90 7.89 90.85 1.27 1 0.70 1.27 )0.02 1.02
3 ⁄ 5 1.20 8.47 90.75 0.77 1 0.76 1.24 )0.09 1.08
3 ⁄ 6 1.40 7.29 85.43 7.27 1 0.67 1.63 0.19 1.51
Fig. 3. Meiofaunal abundances at each sampling station.
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
596 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

123 nematode genera belonging to 34 families (for a list
of all of the genera identified, see Appendix 1). Desmodo-
ridae and Chromadoridae were the richest families, with
16 and 14 genera present, respectively; the most abundant
families were Desmodoridae (26%), Chromadoridae
(13%), Xyalidae (12%) and Draconematidae (7%).
At South Male, 72 genera and 26 families of nematodes
were found; the most abundant genera were Spirinia, Mic-
rolaimus, Prochromadorella, Eubostrichus, Paradesmodora
and Chromaspirinia (SIMPER, 50%). At North Felidhoo,
93 genera and 27 families of nematodes were recorded;
Spirinia, Dracognomus, Prochromadorella, Viscosia, Stylo-
theristus, Paradesmodora and Eubostrichus were the most
abundant genera (SIMPER, 50%). Finally, at East Felid-
hoo, 98 genera and 30 families were found, which were
mainly represented by Microlaimus, Desmodora, Paradesm-
odora, Eubostrichus and Spirinia (SIMPER, 50%).
Significant differences in the nematode community
were detected between the stations (ANOSIM, R = 0.67;
P = 0.001), but not between the transects (P > 0.05)
(Fig. 5B). However, SIMPER tests revealed that the great-
est assemblage dissimilarity was at North Felidhoo, which
had the highest dissimilarity values whether comparisons
were made between the stations at North Felidhoo or
between the different transects (Table 4). Similarly, b-
diversity only revealed significant differences between sta-
tions (ANOSIM, R = 0.69; P = 0.001), indicating that the
greatest dissimilarity in genus composition was at North
Felidhoo (Table 4; Fig. 7).
Regarding the Shannon and Pielou indices of nema-
todes, no significant differences were identified between
the three transects (ANOVA, P > 0.05), even though the
H¢ index was, on average, lower at North Felidhoo,
whereas significant differences were found between the
stations (ANOVA, F17,53 = 7.30 and 3.78 P < 0.001)
(Fig. 6).
Differences in the structure of the nematode assem-
blages relating to sediment type were detected (ANOSIM,
R = 0.47; P = 0.001) (Table 3), and the main nematode
genera responsible for the majority of the average similar-
ity within each sediment type are set out in Table 5. The
assemblages were found to be significantly different in all
of the sediment type pair-wise comparisons. In particular,
the greatest differences were detected between the assem-
blages found in the very coarse versus the fine sands,
while the least differences found were between coarse ver-
sus medium sands [Group (Gr.) CS versus Gr. MS]
(Table 5). The differences between the coarse versus the
medium sands were mainly due to the greater abundance
of the genus Dracognomus in the former and the Desmo-
doridae genera in the latter.
The H¢ and J indices differed according to sediment
type (ANOVA, F3,53 = 5.63 P < 0.01; F3,53 = 10.48
P < 0.001) and had significantly lower values in fine
sands (Tukey’s test P < 0.01). This confirms the results of
the k-dominance curve, whereby a visual trend of nema-
tode diversity can be observed: the greatest dominance of
nematode genera was detected in the fine sands followed
Table 2. Presence (X) ⁄ absence (–) of the different meiofaunal taxa found at each station.
taxa
South Male North Felidhoo East Felidhoo
1 ⁄ 1 1 ⁄ 2 1 ⁄ 3 1 ⁄ 4 1 ⁄ 5 1 ⁄ 6 2 ⁄ 1 2 ⁄ 2 2 ⁄ 3 2 ⁄ 4 2 ⁄ 6 2 ⁄ 7 2 ⁄ 8 3 ⁄ 1 3 ⁄ 2 3 ⁄ 4 3 ⁄ 5 3 ⁄ 6
Cnidaria X – – – – – X – – – X – – – – X X –
Turbellaria X X X – X X X X X X X X X X X X X X
Nemertea X X X X X X X X X X X – X – X X X X
Nematoda X X X X X X X X X X X X X X X X X X
Gastrotricha X X X X X X X X X X X X X X X X X X
Kinorhyncha – – – – – – – – X – – X – – – – X X
Opistobranchia – – – – – – – – – – – – – X X – X –
Polychaeta X X X X X X X X X X X X X X X X X X
Oligochaeta X X X X – X – – X X X X X X X X X X
Copepoda X X X X X X X X X X X X X X X X X X
Ostracoda X X X X X X X X X X X X X X X X X X
Isopoda – – – – – – – X – – – – X – – – – –
Tanaidacea X X – – – – – – – – X – – – – – – –
Amphipoda – X – – – – – – – – X – – – – – – X
Cumacea X – – – – – – – – – – – – – X – – X
Halacarida X X X – X – X X X – – X X X X X X X
Tardigrada X X X X X X X X X X – X X X X X X X
Chironomida larvae X – – – – – – – X – X X
Sipunculida – – – – – – – – X – – – – – – – X –
Chaetognatha – – – – – – – – – – – – – – – – X –
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 597

by the very coarse sands, while coarse and medium sands
had very similar k-dominance curves (Fig. 8). The Spear-
man rank correlations for the nematode genera and the
main sedimentological parameters are set out in Table 6.
The overall nematode assemblage was predominantly
made up of epistrate feeders (56% of total assemblage),
followed by non-selective and selective deposit feeders
(17% and 16%, respectively) and predators ⁄ omnivores
(12%) (Fig. 9). The nematode trophic groups were also
found to be differentially distributed between the different
sediment types. In particular, 1B (non-selective deposit
feeders) showed the greatest differences (ANOVA,
F3,53 = 29.09; P < 0.001) and were the most abundant in
fine sands (Tukey’s test P < 0.001). A significant differ-
ence was also detected for 2B (predators ⁄ omnivores)
(ANOVA, F3,53 = 4.99; P < 0.01), which was the most
abundant in medium sands (Tukey’s test P < 0.01). Selec-
tive deposit feeders (1A) were significantly abundant in
coarse sands (ANOVA, F3,53 = 3.21 P < 0.05; Tukey’s test
P < 0.05), whereas no significant differences were
detected for 2A (epistratum feeders).
A
B
C
Fig. 4. Composition of meiofauna at each sampling station (Ne,
nematodes; Co, copepods; Po, polychaetes; Tu, turbellarians; Os,
ostracods; Oth, Others).
A
B
Fig. 5. nMDS plot of the meiofaunal (A) and nematode (B) assem-
blages (square root-transformed) (stress coefficient: 0.13). The
sampling grouping was based on Bray–Curtis clustering.
Table 3. Results of one-way ANOSIM (global R) on the meiofaunal and
nematode assemblages for the different sediment type comparisons.
meiofauna nematodes
R P R P
global 0.34 0.001 0.47 0.001
coarse sand versus very coarse sand 0.56 0.006 0.59 0.009
coarse sand versus medium sand 0.13 0.028 0.19 0.007
coarse sand versus fine sand 0.55 0.001 0.73 0.001
medium sand versus very coarse sand 0.73 0.005 0.99 0.001
medium sand versus fine sand 0.35 0.004 0.78 0.001
fine sand versus very coarse sand 0.98 0.012 1.00 0.012
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
598 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH
Discussion
A high number of meiofauna taxa, some of them rare,
were found in this study, demonstrating the great diver-
sity of the interstitial fauna in the Maldives. The finding
of a chaetognath species at Fotteyo Finolhu was particu-
larly interesting, as only two meiobenthic species of this
phylum have been identified to date (Kapp & Giere
2005). A great richness in nematode genera was also
found, and the presence of a remarkable 96 of 123 genera
set ‘new records’ for the archipelago. A notable overlap in
generic composition can be observed between the Maldi-
vian nematode assemblage found in this study and those
reported for other tropical and even temperate areas
(Heip et al. 1985; Alongi 1986; Gourbault & Renaud-
Mornant 1990; Gourbault et al. 1995; Boucher 1997;
Ndaro & Olafsson 1999; Raes et al. 2007). The distribu-
tion of some genera across a broad range of different
habitats, and even geographically distant areas, not only
proves the wide distribution of these taxa, but also the
existence of iso-communities that mainly associate with
specific sediment grain sizes (Raes et al. 2007). Indeed, all
of the families that appeared to be dominant in this study
are found in coarse sands, confirming the independence
of the community structure with respect to latitude, and
its main relationship with the granulometry or microhab-
itat type (Heip et al. 1985; Raes et al. 2007, 2008).
The Maldivian sea floor has a continuous supply of
originally biogenic sediments of different sizes. Due to the
inadequate distances and transport times involved, waves
and currents are not able to rework and select these sedi-
ments to any significant degree. Consequently, the granu-
lometric statistical parameters (i.e. mean grain size,
sorting, skewness and kurtosis) are less indicative of the
hydrodynamic conditions than those of the terrigenous
sediments. Nevertheless, some considerations can be taken
into account to explain the distribution of the assem-
blages.
Our study suggests that the Maldivian back reef plat-
forms are all relatively similar in terms of meiofaunal
assemblage structure, even when spatially separated (i.e.
located on opposite sides of the same atoll, or even on
different atolls), and geomorphologically different. This
was especially evident for the nematode assemblage,
which had greater b-diversity within individual platforms
(i.e. on a ‘small-scale’) than between platforms (i.e. on a
‘large-scale’). In general, the occurrence of species extinc-
tion or migration does not necessarily have to result in
Fig. 6. Shannon–Wiener (H¢) and Pielou (J)
index values for the nematode assemblages
at each sampling station. *Significant
difference, where P < 0.05 (Tukey’s test).
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 599

the overall loss of that species from the surrounding area;
i.e. the same species could continue to be present in other
patches within the same area. Accordingly, an overall
higher level of species similarity, and consequent lower
b-diversity, should be observed on the larger spatial scale.
In contrast, on the small-scale level, species extinctions or
migrations might equate to the total loss thereof from an
area, resulting in the local disappearance of the species
when too few patches of a habitat remain (high b-diver-
sity). This might be due to predation or competition, or
other effects of patch dynamics that are related to spatial
heterogeneity, which on a small-scale can have a major
impact (Raes et al. 2007). However, it is interesting to
note that, even though significant differences were not
detected, North Felidhoo was the area with the greatest
degree of assemblage dissimilarity. This is most probably
due to the fact that the transects of South Male and East
Felidhoo were located on the sandy parts of the reef flats,
whereas the North Felidhoo transect was on a filling-up
karst Pleistocene depression (Fig. 2). Here, two deposi-
tional features were visible: (i) the presence of fine depos-
its at the bottom of the depression, and (ii) the presence
of advancing inclined layers of coarse sands, due to a
continuum of sediment supply from storm events. These
features would have contributed to the increased hetero-
geneity of the platform’s habitat in line with the structure
and composition of the studied assemblages. It is worth
noting that the greatest dissimilarities calculated were
always found when more weight was given to the abun-
dance of nematode genera and the common taxa (�transformed). This highlights that the dissimilarities were
mainly caused by the different contributions of the genera
to the whole community, rather than by the presence of
unique and very specific taxa restricted to a particular
atoll, platform or station (Table 4).
The influence of sediment grain size on the meiofauna
assemblages was further investigated by ordering the sam-
ples according to sediment type. When sediment types are
ordered irrespective of the station or transect of origin, a
strong relationship exists between nematodes and grain
size, in confirmation of the findings by Vincx (1989). This
is also evident when specimens are identified on the genus
level, in contrast with the observations of Vanaverbeke
et al. (2002) (Table 3). When it comes to explaining even
minor differences in the assemblage structure (see Raes
et al. 2007), the importance of small sedimentological
variations was confirmed: when only the mean grain size
(Mz) was considered, few statistically significant correla-
tions were found in relation to genus; however, when the
granulometric classes were considered, a larger and more
significant number of correlations were identified (Table 6).
Although the assemblages were found to be signifi-
cantly different in all of the pair-wise comparisons, it is
Table 4. Average dissimilarities (Av. Diss.) calculated on nematode
assemblages (SIMPER tests with 50% cut-off). Genera abundances
were square root- and presence ⁄ absence-transformed. Comparisons
were performed between assemblages inhabiting different transects
and between assemblages within the same transect.
groups
square root
Av. Diss.
presence ⁄ absence
Av. Diss.
T1 versus T2 72.0 66.2
T2 versus T3 73.4 68.2
T1 versus T3 65.8 59.1
11 versus 12 59.0 41.7
11 versus 13 54.2 45.4
12 versus 13 59.7 56.5
11 versus 14 65.1 59.3
12 versus 14 64.4 48.0
13 versus 14 60.4 57.1
11 versus 15 45.5 41.8
12 versus 15 65.8 56.5
13 versus 15 57.6 53.9
14 versus 15 59.2 53.6
11 versus 16 62.0 49.3
12 versus 16 55.4 48.2
13 versus 16 56.8 53.1
14 versus 16 58.0 41.2
15 versus 16 59.3 50.0
21 versus 22 61.9 46.9
21 versus 23 90.8 84.0
22 versus 23 77.8 70.5
21 versus 24 91.6 85.4
22 versus 24 85.3 73.1
23 versus 24 48.9 54.7
21 versus 26 69.0 55.5
22 versus 26 73.4 70.2
23 versus 26 91.5 87.5
24 versus 26 87.1 79.5
21 versus 27 76.5 64.0
22 versus 27 48.7 44.3
23 versus 27 80.6 71.0
24 versus 27 86.5 77.4
26 versus 27 73.0 66.7
21 versus 28 81.3 78.0
22 versus 28 51.7 48.4
23 versus 28 70.0 68.1
24 versus 28 78.8 71.8
26 versus 28 77.0 70.0
27 versus 28 55.3 44.7
31 versus 32 55.3 52.3
31 versus 34 46.1 40.9
32 versus 34 48.0 41.1
31 versus 35 58.4 44.1
32 versus 35 54.8 45.7
34 versus 35 49.8 47.8
31 versus 36 62.8 67.2
32 versus 36 55.1 53.9
34 versus 36 56.7 56.0
35 versus 36 61.4 54.9
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
600 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

interesting that the lowest differences were found between
assemblages in coarse versus medium sands (Gr. CS versus
Gr. MS) (Table 3). This is probably due to the fact that
these sediments were mainly occupied by interstitial fauna
(Table 3). In this dimensional range, e.g. in the nematode
assemblage, a greater degree of genera overlap was pres-
ent, highlighting the similar effects of these two sediment
types upon both fauna composition and the relative
contributions of the different taxa.
The nematode genera listed in Table 5 could be
regarded as indicators of the four sediment types found
in this Maldivian study area, which, in turn, provide use-
ful information about the relationship of some taxa with
granulometry. Genera that are known to be typical of
coarse sediment fractions and habitats exposed to high
energy hydrodynamics (Heip et al. 1985; Somerfield et al.
1995; Schratzberger & Warwick 1998; Nicholas & Hodda
1999; Raes & Vanreusel 2006; Raes et al. 2007) were here
identified as being taxa that are indicators of the very
coarse and the coarse sands. On the other hand, the num-
ber of different Desmodoridae genera present increased as
the size of the sediment interstitial spaces decreased (i.e.
Desmodoridae were more prevalent in medium than in
coarse sands). Of the Desmodoridae genera identified,
Table 5. Main nematode genera responsible for most of the average similarity within each sediment type; detected using the SIMPER test (50%
cut-off).
very coarse sand (Gr. VCS) coarse sand (Gr. CS) medium sand (Gr. MS) fine sand (Gr. FS)
Genera Contrib. % Genera Contrib. % Genera Contrib. % Genera Contrib. %
Theristus 1B 8.91 Dracognomus 2A 9.53 Eubostrichus 1A 10 Stylotheristus 1B 30.3
Dracognomus 2A 8.91 Microlaimus 2A 8.41 Microlaimus 2A 8.89 Spirinia 2A 20.11
Chromadora 2A 7.72 Prochromadorella 2A 7.35 Chromaspirinia 2B 8.88
Paracyatholaimus 2A 7.72 Spirinia 2A 7.24 Paradesmodora 2A 7.86
Halalaimus 1A 7.72 Paradesmodora 2A 6.33 Spirinia 2A 7.68
Prochromadorella 2A 6.3 Epacanthion 2B 5.6 Odontophora 1B 4.62
Viscosia 2B 6.3 Eubostrichus 1A 5.46 Desmodora 2A 3.89
Perepsilonema 1A 3.91
Fig. 7. nMDS plot of the nematode
assemblages (presence ⁄ absence-transformed)
(stress coefficient: 0.16). The sampling
grouping was based on Bray–Curtis
clustering.
Fig. 8. k-dominance curves of nematode assemblage for each sedi-
ment type.
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 601

Eubostrichus (Stilbonematinae), Eubostrichus cf. parasitifer
and Eubostrichus cf. exilis were the most dominant, and
were primarily correlated with medium sands and
secondly with coarse, poorly sorted sands (Table 6).
Species of Eubostrichus are sediment-dwellers which tend
to penetrate into deeper, anoxic sediments, where they
are able to exploit the high concentrations of sulfide
present around the chemocline by means of their ecto-
symbiotic bacteria (Hentschel et al. 1999; Ott et al. 2005).
Curiously, high abundances of Stilbonematinae have been
frequently reported in subtropical and tropical sediments,
and especially coralline sands (Gerlach 1963a,b; Boucher
1997; Riemann et al. 2003; Raes et al. 2007). The great
abundance of these species in carbonate sediments may
be related to the presence of large grains in generally
sheltered habitats, a condition which represents an unu-
sual combination of wide interstitial spaces and sulfide
conditions with an associated rich thiobios (Riemann
et al. 2003; Giere 2009).
In the fine sands, 50% of the total community was
made up of just two genera (Stylotheristus and Spirinia),
which include species known to be associated with fine
and silty sediments (Heip et al. 1985; Buchholz & Lamp-
adariou 2002). It is also worth noting that the genus
Spirinia, discovered by Gerlach (1963a,b) in the Maldives,
was common in all of the sediment types, except for very
coarse sands. Ndaro & Olafsson (1999) also reported a
widespread presence of Spirinia in all of the sediment
typologies of the Zanzibar lagoons, demonstrating the
general importance of this genus in carbonate sediments,
although in this study a closer relationship with sediment
grain sizes lower than 125 lm was found.
In accordance with the literature, the Shannon (H¢)and evenness (J) indices for the nematode assemblages
were highest in medium-coarse sands and lowest in fine
sands (see also: Heip et al. 1985; Steyaert et al. 1999;
Vanaverbeke et al. 2002). This is confirmed by the
k-dominance curves, which reveal a very similar pattern
for nematode genera dominance in medium-coarse sands
in accordance with the observations on the structure of
the nematode assemblage (Fig. 9). Medium and coarse
sediments are, in fact, richer in micro-niches (see Giere
2009 for an overview) and have relatively large interstitial
spaces. These provide areas for feeding and sheltering, as
well as biofilms and microalgae that grow on the grain
surfaces and are an abundant food source for the meiofa-
una. The uniform diversity pattern of this grain size
dimensional range also proves that the majority of
Table 6. Spearman correlations between the main nematode genera and sediment parameters (including all of the grain size classes).
2000 lm 1000 lm 500 lm 250 lm 125 lm 62 lm <62 lm Mz (mm) d % gravel % sand % pelite
Chromaspirinia )0.12 )0.02 0.00 0.62 0.20 )0.07 )0.05 )0.10 0.21 )0.11 0.24 )0.01
Daptonema )0.55 )0.48 )0.49 )0.07 0.43 0.56 0.26 )0.48 )0.26 )0.54 0.42 0.23
Desmodora )0.16 )0.04 0.02 0.42 0.10 0.06 )0.14 )0.02 )0.04 )0.14 0.27 )0.19
Dracognomus 0.28 0.52 0.52 0.33 )0.45 )0.56 )0.47 0.54 )0.12 0.28 0.00 )0.44
Paradesmodora )0.04 )0.04 )0.05 0.48 0.12 0.00 0.14 )0.03 0.34 )0.02 0.01 0.14
Prochromadorella 0.58 0.46 0.46 0.10 )0.45 )0.65 )0.50 0.56 )0.21 0.59 )0.26 )0.47
Spirinia )0.40 )0.51 )0.42 0.03 0.48 0.53 0.32 )0.55 0.18 )0.41 0.22 0.35
Stylotheristus )0.47 )0.59 )0.58 )0.38 0.49 0.64 0.51 )0.62 0.01 )0.49 0.17 0.51
Trefusia )0.26 0.02 0.12 0.10 )0.01 0.13 0.24 )0.11 0.35 )0.29 0.09 0.25
Zalonema )0.11 )0.28 )0.28 0.27 0.26 0.20 0.04 )0.17 )0.18 )0.10 0.12 )0.10
Eubostrichus )0.17 )0.19 )0.14 0.27 0.20 0.21 0.21 )0.22 0.32 )0.18 0.03 0.21
For values in bold, P < 0.05; for values in bold ⁄ italics, P < 0.01.
Fig. 9. Trophic structure of nematode
assemblages at each sampling station. 1A
(selective deposit feeders); 1B (non selective
deposit feeders); 2A (epigrowth feeders); 2B
(predators ⁄ omnivores).
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
602 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

nematodes are better adapted to an interstitial life strat-
egy. This was particularly evident in the comparison with
the fine sands, which were strongly dominated by few
and tolerant genera (Table 5). An intermediate condition
was present in very coarse sands, and despite this
sediment type represents a selective micro-habitat with
few genera, its indicator genera are all well adapted to the
present conditions, and overall are more equally
represented than in the fine sediments (Raes & Vanreusel
2006).
When it comes to the trophic composition of the nema-
tode assemblage, unlike the other trophic groups, the dis-
tribution of epistrate feeders (2A) was not significantly
different between the sediment types. In fact, epistrate
feeders were a broadly dominant trophic group in all of
the studied areas. They are commonly found in medium-
coarse sands with a very poor fine fraction (Alongi 1986),
and in tropical habitats they can find a high degree of ben-
thic primary production, a great abundance of diatoms,
and wide surfaces suitable for scraping off the algal and
bacterial biofilms (Boucher 1997; Raes et al. 2007). In this
study, predators ⁄ omnivores (e.g. Chromaspirinia, Epacan-
thion, Viscosia and Oncholaimus) were more abundant in
the medium than in the coarser and exposed sands. This is
probably because these species feed on organisms (other
meiofauna or small macrofauna) which take advantage of
the high organic matter content, which generally increases
as grain size decreases (Netto et al. 1999a,b). Furthermore,
some of these genera (e.g. Oncholaimidae genera) are
facultative predators and are able to exploit a wide range
of food resources (e.g. detrital material). In fact, as oppor-
tunistic feeders, predation is merely an additional mecha-
nism to obtain extra food (Moens & Vincx 1997).
The high abundances of selective deposit feeders (1A)
found in coarse sands in this study contrast with data
from other research (Heip et al. 1985; Alongi 1986; Ndaro
& Olafsson 1999). The presence of this trophic group,
typically dominant in the fine sediment fraction, was
probably high in coarse sands due to the presence of
Epsilonematidae (Perepsilonema, Epsilonema, Leptepsilo-
nema) – a family that is particularly well adapted to
high-energy habitat and larger Mz. As expected, non-
selective deposit feeders (1B) were mostly prevalent in
mud fractions that are generally rich in bacteria and par-
ticulate detritus (Alongi 1986). Indeed, the greatest 1B
abundances were detected at the station situated in the
North Felidhoo depression, characterized by high
amounts of pelite and low hydrodynamic conditions.
Acknowledgements
We are grateful to the referees who made constructive
comments and suggestions to improve the manuscript.
References
Alongi D.M. (1986) Population structure and trophic compo-
sition of the free-living nematodes inhabiting carbonate
sands of Davies Reef, Great Barrier Reef, Australia. Austra-
lian Journal of Marine and Freshwater Research, 37, 609–619.
Aubert O., Droxler A.W. (1992) Cenozoic evolution of the
Maldives carbonate system (Equatorial Indian Ocean). Bulle-
tin des Centres de Recherches Exploration-Production
Elf-Aquitaine, 162, 113–135.
Aubert O., Droxler A.W. (1996) Seismic stratigraphy and
depositional signatures of the Maldive carbonate system
(Indian Ocean). Marine and Petroleum Geology, 13, 503–536.
Bianchi C.N., Colantoni P., Geister J., Morri C. (1996) Reef
geomorphology, sediments and ecological zonation at Felidu
Atoll, Maldive Islands (Indian Ocean). Proceedings 8th Inter-
national Coral Reef Symposia, 1, 431–436.
Boucher G. (1997) Structure and biodiversity of nematode
assemblages in the SW lagoon of New Caledonia. Coral
Reefs, 16, 177–186.
Buchanan J.B. (1984) Sediment analysis. In: Holme N.A.,
McIntyre A.D. (Eds), Methods for the Study of Marine
Benthos. Blackwell Scientific Publications, Oxford: 41–65.
Buchholz T.G., Lampadariou N. (2002) Changes in composi-
tion and diversity of Malia bay nematode community
(Crete, Greece) in relationship to sediment parameters. In:
Bright M., Dworschak P.C., Stachowitsch M. (Eds), The
Vienna School of Marine Biology: A tribute to Jorg Ott.
Facultas Universitatsverlag, Vienna: 33–52.
Clarke K.R., Gorley R.N. (2001) PRIMER v5: User Manual ⁄Tutorial. Primer-E Ltd, Plymouth: 91 pp.
Clarke K.R., Warwick R.M. (2001) Changes in marine commu-
nities: an approach to statistical analysis and interpretation,
2nd edn. Plymouth Marine Laboratory, Plymouth: 172 pp.
Colantoni P., Baldelli G., Bianchi C.N., Capaccioni B., Morri
C., Sandrini M., Tassi F. (2003) A cave flooded by marine
water with hydrogen sulphide highlights the recent evolution
of the Maldives (Indian Ocean): preliminary notes. Le Grotte
d’Italia, 4, 29–37.
Dahms H.U., Harder T., Qian P.Y. (2007) Selective attraction
and reproductive performance of a harpacticoid copepod in
a response to biofilms. Journal of Experimental Marine Biol-
ogy and Ecology, 341, 228–238.
De Zio Grimaldi S., Lamarca A., D’Addabbo Gallo M., Pietan-
za R. (1999) Floractinae of Asdhu Island, Maldive, Indian
Ocean (Tardigrada, Heterotardigrada). Italian Journal of
Zoology, 66, 383–391.
Deprez T., Vanden Berghe E., Vincx M. (2004) NeMys: a mul-
tidisciplinary biological information system. In: Vanden Ber-
ghe E., Brown M., Costello M.J., Heip C., Levitus S.,
Pissierssens P. (Eds), Proceedings of ‘the colour of ocean data’
Symposium. Brussels, 25–27th November 2002, IOC Work-
shop Report 188, UNESCO, Paris: 57–63.
Duncan R.A., Hargraves R.B. (1990) 40Ar ⁄ 39Ar geochronology
of basement rocks from Mascarene Plateau, Chagos Bank, and
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 603

the Maldives Ridge. Proceedings of the Ocean Drilling Program
Scientific Results, Vol. 115, College Station, TX: 43–51.
Folk L.R., Ward W.C. (1957) Brazos River (Texas); a study in
significance on grain size parameters. Journal of Sedimentary
Research, 27, 3–26.
Gallo M., D’Addabbo R., De Leonardis C., Sandulli R., De Zio
Grimaldi S. (2007) The diversity of Indian Ocean Heterotar-
digrada. Journal of Limnology, 66, 60–64.
Gerlach S. (1961) }Uber Gastrotrichen aus dem Meeressand der
Maldiven (Indischer Ozean). Zoologischer Anzeiger, 167,
471–475.
Gerlach S. (1962) Freilebende Meeresnematoden von den
Maldiven. Kieler Meeresforschungen, 18, 81–108.
Gerlach S. (1963a) Freilebende Meeresnematoden von den
Maldiven II. Kieler Meeresforschungen, 19, 67–103.
Gerlach S. (1963b) Robbea tenax sp. n., ein merkwurdiger
mariner Nematode von den Maldiven. Internationale Revue
der Gesamten Hydrobiologie, 48, 153–158.
Gerlach S. (1964) Neue Cyatholaimidae (Nematoda Chromad-
orida) von den Maldiven. Veroffentlichungen des Instituts fur
Meeresforschung in Bremerhaven, 9, 70–78.
Giere O. (2009) Meiobenthology: the microscopic motile fauna of
aquatic sediments, 2nd edn. Springer, Berlin: 527 pp.
Godfrey T. (1996) Dive Maldives. A guide to the Maldives
Archipelago. Atoll Editions , Apollo Bay, Australia: 180 pp.
Gourbault N., Renaud-Mornant J. (1990) Micro-meiofaunal
community structure and nematode diversity in a lagoonal
ecosystem (Fangataufa, Eastern Tuamotu Archipelago).
PSZNI: Marine Ecology, 11, 173–189.
Gourbault N.E., Warwick R.M., Helleouet M. (1995) A survey
of intertidal meiobenthos (especially Nematoda) in coral
sandy beaches of Moorea (French Polynesia). Bulletin of
Marine Science, 57, 476–488.
Gray J.S. (2000) The measurement of marine species diversity,
with an application to the benthic fauna of the Norwegian
continental shelf. Journal of Experimental Marine Biology and
Ecology, 250, 23–49.
Guo Y., Helleouet M.N., Boucher G. (2007) Spatial patterns of
meiofauna and diversity of nematode species assemblages in
the Uvea lagoon (Loyalty Islands, South Pacific). Journal of
Marine Biological Association of the United Kingdom, 88,
934–940.
Heip C., Vincx M., Vranken G. (1985) The ecology of marine
nematodes. Oceanography and Marine Biology: An Annual
Review, 23, 399–489.
Hentschel U., Berger E.C., Bright M., Felbeck H., Ott J.A.
(1999) Metabolism of nitrogen and sulpur in ectosymbiotic
bacteria of marine nematodes (Nematoda, Stilbonematinae).
Marine Ecology Progress Series, 183, 149–158.
Kapp H., Giere O. (2005) Spadella interstitialis, a meiobenthic
chaetognath from Mediterranean calcareous sands. Meiofauna
Marina, 14, 109–114.
Kench P.S., Mc Lean R.F., Nichol S.L. (2005) New model of
reef-island evolution: Maldives, Indian Ocean. Geology, 33,
145–148.
Krumbein W.C. (1934) Size frequency distributions of sedi-
ments. Journal of Sedimentary Research, 4, 65–70.
Moens T., Vincx M. (1997) Observations on the feeding
ecology of estuarine nematodes. Journal of Marine
Biological Association of the United Kingdom, 77,
211–227.
Ndaro S.G.M., Olafsson E. (1999) Soft-bottom fauna with
emphasis on nematode assemblage structure in a tropical
intertidal lagoon in Zanzibar, eastern Africa: I. spatial vari-
ability. Hydrobiologia, 405, 133–148.
Netto S.A., Attrill M.J., Warwick R.M. (1999a) Sublittoral mei-
ofauna and macrofauna of Rocas Atoll (NE, Brazil): indirect
evidence of a topographically controlled front. Marine Ecol-
ogy Progress Series, 179, 175–186.
Netto S.A., Warwick R.M., Attrill M.J. (1999b) Meiobenthic
and macrobenthic community structure in carbonate sedi-
ments of Rocas Atoll (North-east, Brazil). Estuarine Coastal
and Shelf Science, 48, 39–50.
Nicholas W.L., Hodda M. (1999) The free-living nematodes of
temperate, high energy, sandy beach: faunal composition
and variation over space and time. Hydrobiologia, 394,
113–127.
Ott J.A., Bright M., Bulgheresi S. (2005) Marine microbial
thiotrophic ectosymbioses. Oceanography and Marine Biology
An Annual Review, 42, 95–118.
Pfannkuche O., Thiel H. (1988) Sample processing. In: Higgins
R.P., Thiel H. (Eds), Introduction to the study of meiofauna.
Smithsonian Institute, Washington, DC: 134–145.
Platt H.M., Warwick R.M. (1983) Free-living marine nema-
todes. Part I. British Enoplids. Synopses of the British Fauna
(New Series) 28. Cambridge University Press, Cambridge:
307 pp.
Platt H.M., Warwick R.M. (1988) Free-living marine nema-
todes. Part II. British Chromadorids. Synopses of the British
Fauna (New Series) 38. Brill, Leiden: 502 pp.
Raes M., Vanreusel A. (2006) Microhabitat type determines
the composition of nematode communities associated with
sediment-clogged cold-water coral framework in the Porcu-
pine Seabight (NE Atlantic). Deep-Sea Research Part I, 53,
1880–1894.
Raes M., De Troch M., Ndaro S.G.M., Muthumbi A., Guilini
K., Vanreusel A. (2007) The structuring role of microhabitat
type in coral degradation zones: a case study with marine
nematodes from Kenya and Zanzibar. Coral Reefs, 26,
113–126.
Raes M., Decraemer W., Vanreusel A. (2008) Walking with
worms: coral-epifaunal nematodes. Journal of Biogeography,
35, 2207–2222.
Riemann F., Thiermann F., Bock L. (2003) Leptonemella
species (Desmodoridae, Stilbonematinae), benthic marine
nematodes with ectosymbiotic bacteria, from littoral sand of
the North Sea island of Sylt: taxonomy and ecological
aspects. Helgoland Marine Research, 57, 118–131.
Risk M.J., Sluka R. (2000) The Maldives: A nation of atolls. In:
McClanahan T.R., Sheppard C.R.C., Obura D.O. (Eds),
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
604 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

Coral Reefs of the Indian Ocean: Their Ecology and Conserva-
tion. Oxford University Press, New York: 325–351.
Schratzberger M., Warwick R.M. (1998) Effects of physical
disturbance on nematode communities in sand and mud: a
microcosm experiment. Marine Biology, 130, 643–650.
Somerfield P.J., Rees H.L., Warwick R.M. (1995)
Interrelationships in community structure between shallow-
water marine meiofauna and macrofauna in relation to
dredging disposal. Marine Ecology Progress Series, 127,
103–112.
Steyaert M., Garner N., Gansbeke D., Vincx M. (1999) Nema-
tode communities from the North Sea: environmental
controls on species diversity and vertical distribution within
the sediment. Journal of Marine Biological Association of the
United Kingdom, 79, 253–264.
Vanaverbeke J., Gheskiere T., Steyaert M., Vincx M. (2002)
Nematode assemblages from subtidal sandbanks in the
Southern Bight of the North Sea: effect of small
sedimentological differences. Journal of Sea Research, 48,
197–207.
Vincx M. (1989) Free-living marine nematodes from the
Southern Bight of North Sea. Academia Analecta Klasse
Wetenschappen, 51: 39–70.
Warwick R.M., Platt H.M., Somerfield P.J. (1998) Free-living
marine nematodes. Part III. British Monhysterids. Synopses of
the British Fauna (New Series) 53. Field Studies Council,
Shrewsbury, UK: 296 pp.
Wieser W. (1953) Die Beziehung zwischen Mundhohlengestalt,
Ernahrungsweise und Vorkommen bei freilebenden marinen
Nematoden. Eine okologisch-morphologische Studie. Arkiv
fur Zoologie, 4, 439–484.
Wild C., Rasheed M., Jantzen C., Cook P., Stuck U., Huettel
M., Boetius A. (2005) Benthic metabolism and degradation
of natural particulate organic matter carbonate and silicate
reef sands of the northern Red Sea. Marine Ecology Progress
Series, 298, 69–78.
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 605

Appendix 1
List of all nematode genera found in the Maldives Archi-
pelago. The genera marked with an asterisk indicate those
recorded for the first time in the Maldives.
Nematode genera
*Actinonema Cobb, 1920
*Aegialoalaimus de Man, 1907
*Alaimella Cobb, 1920
*Ammotheristus Lorenzen, 1977
*Amphimonhystera Allgen, 1929
Anticoma Bastian, 1865
*Ascolaimus Ditlevsen, 1919
*Astomonema Ott, Rieger and Enderes, 1982
*Atrochromadora Wieser, 1959
Axonolaimus de Man, 1889
Bolbonema Cobb, 1920
*Calomicrolaimus Lorenzen, 1976
Camacolaimus de Man, 1889
*Campylaimus Cobb, 1920
*Ceramonema Cobb, 1920
*Chitwoodia Gerlach, 1956
*Chromadora Bastian, 1865
*Chromadorella Filipjev, 1918
*Chromadorina Filipjev, 1918
*Chromadorita Filipjev, 1922
*Chromaspirina Filipjev, 1918
*Cobbia de Man, 1907
*Comesa Gerlach, 1956
Comesomatidae gen. 1
*Coninckia Gerlach, 1956
*Cricolaimus Southern, 1914
Croconema Cobb, 1920
*Cyartonema Cobb, 1920
Cyatholaimidae gen. 1
*Cyatholaimus Bastian, 1865
*Daptonema Cobb, 1920
Dasynemoides Chitwood, 1936
*Demonema Cobb, 1894
Desmodora de Man, 1889
*Desmoscolex Claparede, 1863
*Dichromadora Kreis, 1929
*Didelta Cobb, 1920
*Disconema Filipjev, 1918
*Dracognomus Allen and Noffsinger, 1978
*Dracograllus Allen and Noffsinger, 1978
*Draconema Cobb, 1913
*Epacanthion Wieser 1953
*Epsilonema Steiner, 1927
Eubostrichus Greeff, 1869
*Eurystomina Filipjev, 1921
*Gammanema Cobb, 1920
*Gomphionema Wieser and Hopper, 1966
*Graphonema Cobb, 1898
Halalaimus de Man, 1888
*Halichoanolaimus de Man, 1886
*Innocuonema Inglis, 1969
Appendix 1. (Continued)
*Latronema Wieser, 1954
*Leptepsilonema Clasing, 1983
*Leptolaimus de Man, 1876
Leptonemella Cobb, 1920
*Linhomoeus Bastian, 1865
*Linhystera Juario, 1974
Litinium Cobb, 1920
Longicyatholaimus Micoletzky, 1924
*Marylynnia Hopper, 1977
*Megadesmolaimus Wieser, 1954
*Mesacanthion Filipjev, 1927
Metachromadora Filipjev, 1918
Metacyatholaimus Stekhoven, 1942
*Metalinhomoeus de Man, 1907
*Metepsilonema Steiner, 1927
*Metoncholaimus Filipjev, 1918
*Microlaimus de Man, 1980
*Molgolaimus Ditlevsen, 1921
Monoposthia de Man, 1889
*Monoposthioides Hopper, 1963
*Neochromadora Micoletzky, 1924
*Odontanticoma Platonova, 1976
Odontophora Butschli, 1874
Onchium Cobb, 1920
*Oncholaimellus de Man, 1890
*Oncholaimus Dujardin, 1845
Oxystomina Filipjev, 1921
*Paracanthonchus Micoletzky, 1924
*Paracomesoma Hope and Murphy, 1972
*Paracyatholaimoides Gerlach, 1953
Paracyatholaimus Micoletzky, 1922
Paradesmodora Stekhoven, 1950
*Paralinhomoeus de Man, 1907
*Paralongicyatholaimus Stekhoven, 1942
*Paramonohystera Steiner, 1916
*Parapinnanema Inglis, 1969
*Parodontophora Timm, 1963
*Paroxystomina Micoletzky, 1924
*Perepsilonema Lorenzen, 1973
*Perspiria Wieser and Hopper, 1967
*Polygastrophora de Man, 1922
*Polysigma Cobb, 1920
*Pomponema Cobb, 1917
*Praeacanthonchus Micoletzky, 1924
*Prochromadorella Micoletzky, 1924
*Pselionema Cobb, 1933
*Pseudocella Filipjev, 1927
*Ptycholaimellus Cobb, 1920
*Rhynchonema Cobb, 1920
Robbea Gerlach, 1956
*Sabatieria Rouville, 1903
*Sigmophoranema Hope and Murphy, 1972
*Siphonolaimus de Man, 1893
*Spilophorella Filipjev, 1917
Spirinia Gerlach, 1963
Stilbonema Cobb, 1920
*Stylotheristus Lorenzen, 1977
Meiofaunal distribution in Maldives Semprucci, Colantoni, Baldelli, Rocchi & Balsamo
606 Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH

Appendix 1. (Continued)
*Subsphaerolaimus Lorenzen, 1978
*Symplocostoma Bastian, 1865
*Synonema Cobb, 1920
Tarvaia Allgen, 1934
*Terschellingia de Man, 1888
*Thalassironus de Man, 1889
*Thalassomonhystera Jacobs, 1987
*Theristus Bastian, 1865
*Trefusia de Man, 1893
*Tricoma Cobb, 1893
*Trissonchulus Cobb, 1920
*Tubolaimoides Gerlach, 1963
*Viscosia de Man, 1890
*Xennella Cobb, 1920
Zalonema Cobb, 1920
Semprucci, Colantoni, Baldelli, Rocchi & Balsamo Meiofaunal distribution in Maldives
Marine Ecology 31 (2010) 592–607 ª 2010 Blackwell Verlag GmbH 607



















