
The chimpanzee major histocompatibility complex class II DR subregion contains an unexpectedly high...
9
Immunogenetics 32: 272-280, 1990 ]11111]11110- genetics © Springer-Verlag 1990 The chimpanzee major histocompatibility complex class II DR subregion contains an unexpectedly high number of beta-chain genes Ronald E. Bontrop, Ludo A.M. Broos, Khanh Pham, Ruud M. Bakas, Nel Otting, and Margreet Jonker ITRI-TNO, Primate Center, Lange Kleiweg 151, 2288 GJ Rijswijk, The Netherlands Received April 12, 1990; revised version received July 6, 1990 Abstract. The major histocompatibility complex (MHC) class II DR subregion of the chimpanzee was studied by restriction fragment length polymorphism (RFLP) analysis. Genomic DNA obtained from a panel of 94 chimpanzees was digested with the restriction enzyme Taq I and hybridized with an HLA-DR/3 probe specific for the 3' untranslated (UT) region. Such a screening revealed the existence of 14 distinct DRB/Taq I gene-associated fragments allowing the definition of 11 haplotypes. Segregation studies proved that the number of chimpanzee class II DRB/Taq I fragments is not constant and varies from three to six depending on the haplotype. Comparison of these data with a human reference panel manifested that some MHC DRB/Taq I fragments are shared by man and chimpanzee. Moreover, the number of HLA-DRB/Taq I gene-associated fragments detected in a panel of homozygous typing cells varies from one to three and cor- responds with the number of HLA-DRB genes present for most haplotypes. However, a discrepancy is observed for the HLA-DR4, -DR7, and -DR9 haplotypes since a fourth HLA-DRB pseudogene present within these haplotypes lacks its 3' UT region and thus is not detected with the probe used. These results suggest that chimpanzees have a higher maximum number of DRB genes per haplotype than man. As a consequence, some chimpanzee haplotypes must show a dissimilar organization of the MHC DR subregion compared to their human equivalents. The implications of these findings are discussed in the con- text of the trans-species theory of MHC polymorphism. Introduction From an evolutionary point of view, the chimpanzee (Pan troglodytes) is the closest living relative of man. This Address correspondence and offprint requests to: R.E. Bontrop. phylogenetic relationship is one of several major reasons why the chimpanzee represents an important experimental animal model used by virologists and immunologists. In this context, it is significant that the major histocom- patibility complex (MHC) of the chimpanzee, recently designated MhcPatr (Klein et al. 1990), should be characterized in detail. The coding sequences of the Patr region are located on chromosome 5 (Garver et al. 1978). Serological screening allows the detection of 14 Patr-A and 7 Patr-B antigens (Balner et al. 1974) which are the analogs of the HLA-A and -B loci. The existence of class II (Patr-D) became evident by means of primary mixed leukocyte culture testing (Jonker and Balner 1980, 1981). Chimpanzee D locus-related antigens were first detected using cross-reacting HLA-DR-specific alloantisera (Balner et al. 1977; Jonker and Balner 1980). More recently, three distinct MhcPatr-DRB gene clones were isolated from a chimpanzee cDNA library and were found to share high sequence similarity with their human equivalents (Fan et al. 1989). However, the accurate description of the Parr-DR subregion is hampered by the fact that the appropriate chimpanzee B-cell sera are presently lacking, a situation encountered for other Patr class II loci too. Although the corresponding sera are absent, the heterogeneity of the Patr-DQA locus was successfully described by use of the restriction fragment length polymorphism (RFLP) technique (Bontrop et al. 1989). Thus, the RFLP method may provide an alternative ap- proach to studying the polymorphism of the Parr-DR subregion. One of the main problems in DNA HLA-DR typing is the interpretation of the complex patterns (Andersson et al. 1984; Cohen et al. 1984) caused by cross-hybridization of the commonly used fulMength HLA-DR/3 cDNA probe (Long et al. 1983) with other class II locus B genes. The Southern blot patterns of DNA obtained from HLA-DR-heterozygous individuals look particularly complex. The use of DRB gene subprobes may reduce the complexity of the Southern blot patterns
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The chimpanzee major histocompatibility complex class II DR subregion contains an unexpectedly high...

Immunogenetics 32: 272-280, 1990 ]11111]11110- genetics
© Springer-Verlag 1990
The chimpanzee major histocompatibility complex class II DR subregion contains an unexpectedly high number of beta-chain genes
Ronald E. Bontrop, Ludo A.M. Broos, Khanh Pham, Ruud M. Bakas, Nel Otting, and Margreet Jonker
ITRI-TNO, Primate Center, Lange Kleiweg 151, 2288 GJ Rijswijk, The Netherlands
Received April 12, 1990; revised version received July 6, 1990
Abstract. The major histocompatibility complex (MHC) class II DR subregion of the chimpanzee was studied by restriction fragment length polymorphism (RFLP) analysis. Genomic DNA obtained from a panel of 94 chimpanzees was digested with the restriction enzyme Taq I and hybridized with an HLA-DR/3 probe specific for the 3' untranslated (UT) region. Such a screening revealed the existence of 14 distinct DRB/Taq I gene-associated fragments allowing the definition of 11 haplotypes. Segregation studies proved that the number of chimpanzee class II DRB/Taq I fragments is not constant and varies from three to six depending on the haplotype. Comparison of these data with a human reference panel manifested that some MHC DRB/Taq I fragments are shared by man and chimpanzee. Moreover, the number of HLA-DRB/Taq I gene-associated fragments detected in a panel of homozygous typing cells varies from one to three and cor- responds with the number of HLA-DRB genes present for most haplotypes. However, a discrepancy is observed for the HLA-DR4, -DR7, and -DR9 haplotypes since a fourth HLA-DRB pseudogene present within these haplotypes lacks its 3' UT region and thus is not detected with the probe used. These results suggest that chimpanzees have a higher maximum number of DRB genes per haplotype than man. As a consequence, some chimpanzee haplotypes must show a dissimilar organization of the MHC DR subregion compared to their human equivalents. The implications of these findings are discussed in the con- text of the trans-species theory of MHC polymorphism.
Introduction
From an evolutionary point of view, the chimpanzee (Pan troglodytes) is the closest living relative of man. This
Address correspondence and offprint requests to: R.E. Bontrop.
phylogenetic relationship is one of several major reasons why the chimpanzee represents an important experimental animal model used by virologists and immunologists. In this context, it is significant that the major histocom- patibility complex (MHC) of the chimpanzee, recently designated MhcPatr (Klein et al. 1990), should be characterized in detail. The coding sequences of the Patr region are located on chromosome 5 (Garver et al. 1978). Serological screening allows the detection of 14 Patr-A and 7 Patr-B antigens (Balner et al. 1974) which are the analogs of the HLA-A and -B loci. The existence of class II (Patr-D) became evident by means of primary mixed leukocyte culture testing (Jonker and Balner 1980, 1981). Chimpanzee D locus-related antigens were first detected using cross-reacting HLA-DR-specific alloantisera (Balner et al. 1977; Jonker and Balner 1980). More recently, three distinct MhcPatr-DRB gene clones were isolated from a chimpanzee cDNA library and were found to share high sequence similarity with their human equivalents (Fan et al. 1989). However, the accurate description of the Parr-DR subregion is hampered by the fact that the appropriate chimpanzee B-cell sera are presently lacking, a situation encountered for other Patr class II loci too.
Although the corresponding sera are absent, the heterogeneity of the Patr-DQA locus was successfully described by use of the restriction fragment length polymorphism (RFLP) technique (Bontrop et al. 1989). Thus, the RFLP method may provide an alternative ap- proach to studying the polymorphism of the Parr-DR subregion. One of the main problems in DNA HLA-DR typing is the interpretation of the complex patterns (Andersson et al. 1984; Cohen et al. 1984) caused by cross-hybridization of the commonly used fulMength HLA-DR/3 cDNA probe (Long et al. 1983) with other class II locus B genes. The Southern blot patterns of DNA obtained from HLA-DR-heterozygous individuals look particularly complex. The use of DRB gene subprobes may reduce the complexity of the Southern blot patterns

R.E. Bontrop et al.: RFLP analysis of the chimpanzee DR subregion 273
(Fei et al. 1986; Bidwell 1988). For instance the DR/3 pro- be, specific for the 3' untranslated (UT) region, detects in a panel of HLA-homozygous typing cells a number of fragments corresponding with the number of HLA-DRB genes present for most haplotypes (Fei et al. 1986). This number varies from one to three depending on the HLA- DR specificity of the cell line studied (Wake et al. 1982; Fei et al. 1986; B6hme et al. 1986). However, not all HLA-DRB genes are functional and some of them have been identified as pseudogenes (Rollini et al. 1985; Larhammer et al. 1985). In this context, it should be men- tioned that an isolated DRB1 exon present in cells with HLA-DR4, -DR7, and -DR9 specificities (Spies et al. 1985) is sometimes referred to as a fourth DRB gene pre- sent on these haplotypes. This type of pseudogene is not detected with an HLA-DR/3 3' UT region-specific probe.
The restriction enzyme Taq I targets nucleotide se- quences containing mutable CpG doublets (Barker et al. 1984) and is able to discriminate between most HLA-DR specificities when used in RFLP studies (Bidwell 1988). For this reason, the HLA-DR/3 3' UT probe was used in combination with the restriction endonuclease Taq I to study the heterogeneity of the DR subregion in a panel of chimpanzees.
Materials and methods
Animals. The ITRI-TNO Primate Center (Rijswijk, The Netherlands) harbors a self-sustaining colony of about 120 chimpanzees initially started with 36 animals caught from the wild in Sierra Leone. In total, 94 related and unrelated chimpanzees were selected for RFLP studies, including most of the 36 animals captured in the wild or their offspring. The panel of chimpanzees also included several extensive pedigrees which were used for segregation purposes. Heparinized blood was ob- tained by venipuncture from healthy chimpanzees.
DNA isolation and Southern hybridization. Genomic DNA, isolated either from heparinized blood samples (chimpanzees) or from Epstein- Barr virus-transformed human B cells, was digested with the restriction endonuclease Taq I, size-separated on an 0.8 % agarose gel, and transfer- red to Hybond (Amersham, Amersham, UK) nylon membranes. Filters were hybridized with an HLA-DR/33' UT probe (Fei et al. 1986) accord- ing to a standardized Southern blot protocol (Marcadet et al. 1987). In brief, hybridization (12 h) was performed at 42 °C in a mixture contain- ing 5 × saline-sodium phosphate-EDTA (SSPE; 0.75 M NaCL, 0.05 M NaH2PO 4 5 mM ethylenediaminetetraacetate, pH 7.7), 50% for- mamide, 0.1% Denhardt's solution, 5 % dextran sulfate, and 1% sodium dodeeyl sulfate (SDS). Filters were washed twice in 2 x SSPE at room temperature, once with 2 x SSPE, 0.5 % SDS at 65 °C, and finally once with 0.5 x SSPE at 65 °C. After drying, filters were exposed to Kodak XS film.
Results
Chimpanzee RFLP panel analysis and identification of distinct MhcPatr-DR haplotypes. High relative mass
DNA obtained from 94 chimpanzees was selected for Southern blot analysis. Taq I digestion and hybridization with an HLA-DR/3 3' UT probe showed the presence of 14 different fragments within our chimpanzee panel with a relative mass of 11.1, 9.8, 9.6, 8.4, 8.0, 7.0, 6.1, 5.8, 5.2, 4.8, 4.1, 3.2, 2.1, and 1.5 kilobases (kb) in length, respectively (Figs. 1, 4 and 5).
The hybridization patterns obtained look rather com- plicated, as is reflected by the presence of an unexpectedly high number of hybridizing fragments (Fig. 1). Most like- ly, this phenomenon is at least partly caused by heterozygosity since the majority of the ITRI-TNO Primate Center chimpanzees are known to be heterozygous for their Patr-A, -B, and -DQA1 loci too (Bontrop et al. 1989). Segregation studies were peformed to examine this possibility and all DRB/Taq I fragments were found to segregate in a Mendelian manner (Fig. 2). Thus, most chimpanzees are indeed heterozygous for their Patr-DR subregion (Figs. 1 and 4). Moreover, segrega- tion studies confirmed that the 14 distinct DRB/Taq I fragments define at least 11 different DR specificities arbitrarily designated Patr-DR1 to -DR11 (Fig. 5). As can be seen, the majority ofDRB/Taq I fragments are not uni- que to the Patr-DR specificity and are shared by distinct haplotypes (Fig. 5). This explains why some fragments are observed in double intensity in the case of DR- heterozygous animals (Fig. 1).
In more detail, the DRB/Taq I 11.1 kb fragment is observed for the Patr-DR1, -DR2, -DR4, and -DR10 specificities. Also, the DRB/Taq 19.8, 9.6, 8.0, 7.0, 6.1, 5.2, 4.8, 4.1, and 3.2 kb fragments are associated with multiple haplotypes (Fig. 5). In contrast to these so-called shared hybridizing fragments, other fragments occur that appear to be unique to the DR specificity. In this respect, the DRB/Taq I 8.4 kb fragment is solely observed for DRl-positive cells (Figs. 4 and 5), whereas the DRB/Taq I 5.8 and 1.5 kb fragments are unique to the DR9 specificity (Figs. 1 and 5). Patr-DR2-positive cells can be distinguished on the basis of the DRB/Taq I 2.1 kb fragment (Figs. 1 and 5).
The DRI3 3' UT probe does not cross-hybridize with non- DR class Il beta-chain genes. The Southern blot displayed in Figure 1 shows the hybridization profiles of three Patr- DR-homozygous chimpanzees, namely a Patr-DRlO, lO and two -DR4,4-positive individuals. In the case of each, five DRB/Taq I fragments are detected. In the human, the HLA-DR/3 3' UT probe is known to be not reactive with HLA-DQ, -DP, and -DO region beta chains (Fei et al. 1986). Whether this is the same for chimpanzees should be tested, since the presence of some of the DRB/Taq I fragments may be due to cross-hybridization. To this pur- pose, the Southern blot displayed in Figure 1 was stripped and subsequently hybridized with a mixture containing radiolabeled probes specific for the HLA-DQ/3 3' (Long
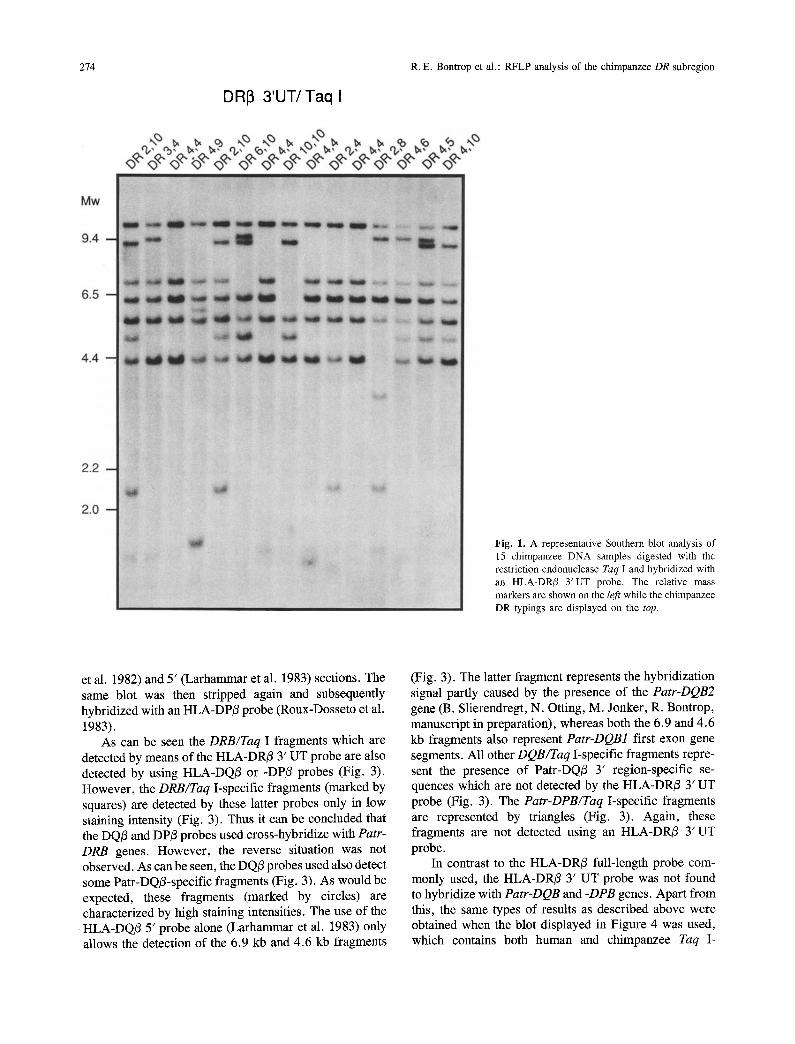
274 R.E. Bontrop et al.: RFLP analysis of the chimpanzee DR subregion
DRI3 3'UT/Taq I
Fig. 1. A representative Southern blot analysis of 15 chimpanzee DNA samples digested with the restriction endonuclease Taq I and hybridized with an HLA-DR/3 3'UT probe. The relative mass markers are shown on the left while the chimpanzee DR typings are displayed on the top.
et al. 1982) and 5' (Larhammar et al. 1983) sections. The same blot was then stripped again and subsequently hybridized with an HLA-DP/3 probe (Roux-Dosseto et al. 1983).
As can be seen the DRB/Taq I fragments which are detected by means of the HLA-DR/3 3' UT probe are also detected by using HLA-DQ/3 or -DP/3 probes (Fig. 3). However, the DRB/Taq I-specific fragments (marked by squares) are detected by these latter probes only in low staining intensity (Fig. 3). Thus it can be concluded that the DQ/3 and DP/3 probes used cross-hybridize with Patr- DRB genes. However, the reverse situation was not observed. As can be seen, the DQ/3 probes used also detect some Patr-DQ/3-specific fragments (Fig. 3). As would be expected, these fragments (marked by circles) are characterized by high staining intensities. The use of the HLA-DQ/3 5' probe alone (Larhammar et al. 1983) only allows the detection of the 6.9 kb and 4.6 kb fragments
(Fig. 3). The latter fragment represents the hybridization signal partly caused by the presence of the Patr-DQB2 gene (B. Slierendregt, N. Otting, M. Jonker, R. Bontrop, manuscript in preparation), whereas both the 6.9 and 4.6 kb fragments also represent Patr-DQB1 first exon gene segments. All other DQB/Taq I-specific fragments repre- sent the presence of Patr-DQ/3 3' region-specific se- quences which are not detected by the HLA-DR/3 3' UT probe (Fig. 3). The Patr-DPB/Taq I-specific fragments are represented by triangles (Fig. 3). Again, these fragments are not detected using an HLA-DR/3 3' UT probe.
In contrast to the HLA-DR/3 full-length probe com- monly used, the HLA-DR/3 3' UT probe was not found to hybridize with Patr-DQB and -DPB genes. Apart from this, the same types of results as described above were obtained when the blot displayed in Figure 4 was used, which contains both human and chimpanzee Taq I-

R . E . Bont rop et al.: R F L P analysis o f the ch impanzee DR subregion 275
6LI DR6,10 DR3,4 DR3,6 DR4,10 DR9,10 DR10,10 DR4,10
Fig . 2. A representat ive example o f the segregat ion
o f DRB/Taq I f ragments in a ch impanzee pedigree. The ch impanzee RFLP-de te rmined D R typings are shown.
digested DNAs (data not shown). For these reasons, it can be safely assumed that all Patr-DRB/Taq I fragments presently described are locus-specific
RFLP-determined DR frequencies and association with DQA1/Taq I fragments. The frequencies of the RFLP- determined DR specificities as found for the ITRI-TNO Primate Center chimpanzee colony are depicted in Table 1. The DQAI locus of these chimpanzees has also been studied by RFLP (Bontrop et al. 1989). Within the colony three different DQA1 gene-associated fragments were observed and designated Patr-DQA1/Taq I 6.99, 5.53, and 4.56 kb, respectively. In the chimpanzee panel tested, 63.76 % are positive for the DQA1/Taq 15.53 kb fragment whereas 22.47% and 13.77% are positive for the DQA1/Taq I 6.99 and 4.56 kb fragments, respectively. The first domain nucleotide sequence of the Patr-DQA1 gene associated with the 6.99 kb fragment closely
T a b l e 1. RFLP-de te rmined Patr-DR haplotype f requencies and associa- t ion with DQA1 locus p o l y m o r p h i s m in a panel o f 94 chimpanzees .
Haplo type Frequency Frequency Patr-DQA1/Taq I f ragment
6 .99 kb 5 .53 kb 4 .56 kb
22 .47% 63 .76% 13.77%
DR] 0 .92% - - 0 .92% DR2 27 .52% - 27 .52% -
DR3 2 .75% - - 2 .75% DR4 35 .32% - 33 .48% 1.84% DR5 6 .42% 6 .42% - -
DR6 5 .05% 2 .29% 1.84% 0 .92% DR7 0 .92% 0 .92% - - DR8 1.38% - - 1 .38%
DR9 5 .96% - - 5 .96% DRIO 12.84% 12.84% - - DR11 0 .92% - 0 .92% -
- , not observed.

276
DQJ3 5' + 3 ' /Taq I
R.E. Bontrop et al.: RFLP analysis of the chimpanzee DR subregion
DPI3 / Taq I
Fig. 3. Hybridization of the same blot as shown in Figure 1 with HLA-DQ~ and -DP/3 probes. The relative mass markers are shown on the left. DRB/Taq I-specific fragments have been marked with squares, DQB/Taq I-specific fragments with circles, and DPB/Taq I-specific fragments with triangles.
resembles the HLA-DQAI*OI03 allele whereas the 4.56 kb fragment is associated with the HLA-DQAI*0501 allele (M. Kenter, N. Otting, J. Leunissen, M. Jonker, R. Bontrop, manuscript in preparation). The Patr- DQA1/Taq I 5.53 kb fragment appears to be associated with a unique nonhuman primate sequence for which the human equivalent has not been found (M. Kenter et al. manuscript in preparation). The DR4 specificity is found in combination with both the DQA1/Taq 1 5.53 and 4.56 kb fragments whereas all three distinct DQA1/Taq I fragments are found in association with the DR6 specifici- ty (Table 1). All other Patr-DR specificities show unique associations with one particular DQA1/Taq I fragment (Table 1).
Human and chimpanzee RFLP-determined DR specifici- ties: a comparison. Human and chimpanzee RFLP pat- terns observed after Taq I digestion and hybridization with an HLA-DQc~ probe were found to exhibit remarkable fragment length similarities (Bontrop et al. 1989). It was investigated whether species-shared conservation of restriction sites was observed for DRB genes as well. To this purpose high relative mass DNA obtained from chim- panzees, HLA-DR-homozygous typing cells, and two heterozygous human cells was digested with the restric- tion enzyme Taq I and hybridized with the relevant DRi3 probe after size separation of the fragments on the same agarose gel. As can be seen for HLA-DR2, -DR3, -DR4, -DR5, -DR6, and -DR7-homozygous individuals, three
major HLA-DRB/Taq I fragments are observed (Figs. 4 and 5). In the case of HLA-DRw8-positive individuals only one distinct HLA-DRB/Taq I fragment is observed (Fig. 4), whereas for HLA-DRl-homozygous cells two distinct HLA-DRB/Taq I fragments are observed (Fig. 5). As ex- pected, DNA obtained from the heterozygous HLA- DRw8, -DRw13/Dw19 individual shares a fragment (8.7 kb) with the HLA-DRw8-positive typing cell and two fragments (7.0 and 2.2 kb) with the HLA-DRw13/Dw18- positive typing cell (Fig. 5). The 9.8 kb fragment of the HLA-DRw8, -DRw13/Dw19-heterozygous donor is not shared with the HLA-DRw13/Dw18 typing cell but with the HLA-DR3-posifive typing cell (Fig. 5). This HLA- DRB/Taq I 9.8 kb fragment reflects the presence of the HLA-DRB*0301 gene which is strongly associated with the HLA-DRw13/Dw19 specificity (Bontrop et al. 1986a, 1988; Termijtelen et al. 1987).
For the HLA-DR1, -DR4-heterozygous individual three major bands are detected which have the same frag- ment lengths as the reference HLA-DR4-positive cell (Fig. 5). The DRB/Taq 15.8 kb fragment is seen in double stain- ing intensity since it is derived both from HLA-DR1 and -DR4-positive individuals (Figs. 4 and 5). The HLA- DRB/Taq I 4.4 kb fragment represents the second frag- ment derived from the HLA-DR1 donor cell (Figs. 4 and 5). As has been reported earlier, the number of fragments detected after Taq I digestion and hybridization with the HLA-DR~ 3' UT probe equals the minimum number of genes found for the different HLA-DR specificities (Fei

R.E. Bontrop et al.: RFLP analysis of the chimpanzee DR subregion
DRI~ 3'UT/Yaq I
277
[ II I
HLA Patr Fig. 4. Southern blot analysis of human and chimpanzee DNA digested with the restriction enzyme Taq I and hybridized with an HLA-DRfl 3' UT probe. The relative mass markers are shown on the left while the HLA- and Patr-DR typings are shown on the top.
et al. 1986). Thus, HLA-DRw8-positive haplotypes have one DRB gene, HLA-DR1 cells have two, whereas HLA- DR3, -DR5, and -DRw6 haplotypes have three DRB genes (Figs. 4 and 5). Four hybrization signals are detected in the case of the HLA-DR4, -DR7, and -DR9 haplotypes when one uses an HLA-DRB subprobe which is specific for the 5' section of the HLA-DRB gene (Bidwelt 1988). However, this fourth signal is due to the presence of an isolated DRB1 exon (Spies et al. 1985) and is not detected with an HLA-DRfl 3' UT region-specific probe (Figs. 4 and 5).
In the case of the chimpanzee panel, three (-DR3, -DR5, -DR6, -DR8, and-DR11), four (-DR9), five (-DR2, -DR4, and -DRIO), or even six (-DR1) different DRB/Taq I fragments were detected per haplotype (Fig. 5). Some DRB/Taq I fragments are shared between man and chimpanzee (Figs. 4 and 5) but, as judged on RFLP patterns, no HLA-DR haplotype was found to be complete- ly identical to a chimpanzee equivalent (Fig. 5).
D i s c u s s i o n
The RFLP technique was selected to investigate the molecular heterogeneity of the chimpanzee DR subregion.
High relative mass DNA was digested with the restriction enzyme Taq I and hybridized with an HLA-DR/3 3' UT- specific probe. As demonstrated, hybridization patterns obtained with this particular probe exhibit reduced com- plexity due to negative cross-hybridization with class II B genes from other loci. More importantly, this probe does not hybridize with isolated DRB1 exons as they are present on HLA-DR4, -DR7, and -DR9-positive cells (Spies et al. 1985). Concordingly, when hybridizations were performed with an HLA-DRB1 domain-specific pro- be instead of the HLA-DR/3 3' UT probe, similar numbers of fragments were detected for most haplotypes. However, one extra hybridization signal was observed for HLA-DR4, -DR7, and -DR9-positive cells due to the presence of isolated DRB1 domains (Bidwell 1988). The value of the HLA-DR/3 3' UT probe is therefore best il- lustrated by the fact that the number of fragments detected in a panel of human homozygous typing cell lines represents the minimum number of HLA-DRB genes pre- sent per haplotype (Fei et al. 1986). In man the number of DRB genes per haplotype varies from one to four (Fei et al. 1986; B6hme et al. 1986; Bontrop et al. 1986b; Bidwell 1988), suggesting that the physical length of the HLA-DR subregion is not constant.

278 R.E. Bontrop et al.: RFLP analysis of the chimpanzee DR subregion
man DR1 DR2 DR3 DR3
~ DR4 DR5
..(= DR6
DR6 DR7 DR8
chimpanzee DR1
DR2 DR3 DR4
~. DR5 N DR6
~" DR7 J ~
DR8 DR9 DR10 DR11
fragment (kb)
Fig. 5. Comparison of human and chim- panzee MHC DRB/Taq I fragments and haplotypes. The HLA-DRB/Taq I fragment patterns were obtained by use of homo- zygous typing cells, whereas the Patr- DRB/Taq I haplotypes were determined by segregation analyses.
Segregation studies showed that the maximum number of DRB/Taq I fragments detected with the HLA-DRI3 3' UT probe is for some chimpanzee haplotypes much higher than for their human equivalents (Fig. 4). This number of DRB/Taq I fragments varied from three (Patr- DR3, -DR5, -DR6, -DR7, and -DR8), four (Parr-DR9), five (Patr-DR2, -DR4, and -DRIO) to even six (Patr- DR1). Other great ape species such as the gorilla and orangutan (data not shown), but also the rhesus monkey (Slierendregt et al. 1990), have a maximum number which is higher than three DRB/Taq I fragments per haplotype as detected with the HLA-DR/3 3' UT probe. Thus, some monkeys and nonhuman primates appear to have haplotypes with a larger number of DRB genes than man. It is realized that the actual counting of Patr-DRB genes presently described is based on the use of a single enzyme. One can argue that the use of a single endonuclease (in this case, Taq I) does not allow an accurate estimation of the gene copy number. Although an excellent correlation appears to exist between the number of DRB/Taq I fragments detected with the HLA-DR/3 3' UT probe and the minimum number of HLA-DRB genes present per haplotype within man, this is not necessarily the case for other species like the chimpanzee. At present we are se- quencing the polymerase chain reaction-amplified polymorphic Patr-DRB first domains of various RFLP- typed chimpanzees. This means that the Patr-DRB genes present within these chimpanzees are analyzed in a head- to-tail fashion. Comparison of the number of Patr-DRB
gene segments detected by means of sequencing and the numbers ofDRB/Taq I fragments detected in RFLP-typed chimpanzees will allow us to draw more firm conclusions about the exact copy number of Patr-DRB genes.
There is growing evidence that the evolution of MHC alleles does not start at the inception of a species, but is rather a trans-species process in which a group of major alleles is passed on in the phylogeny from one species to another (Arden and Klein 1982; Mayer et al. 1988; Figueroa et al. 1988; Lawlor et al. 1988; Gyllensten and Erlich 1989). If one extends this hypothesis, it is likely that not only individual MHC alleles, but also entire DR subregions evolve in a trans-species manner (Fan et al. 1989). If this indeed happened then not only the great apes but also the hominids must have inherited at least some haplotypes with DR subregions containing more than four DRB genes from a common ancestor. Man and great ape species like the gorilla and chimpanzee have a combina- tion of certain DRB genes in common. (J. Klein, personal communication). However, to our knowledge human haplotypes containing more than four DRB genes which are equivalent to some of the chimpanzee haplotypes presently described have not (yet) been detected within the human population. One explanation may be that such haplotypes are deficient in the best genetically studied human population, the Caucasoid one. Therefore, it can not formally be excluded that HLA-DR haplotypes giving rise to more than four DRB/Taq I fragments are present in other ethnic groups and are there to be discovered. An

R.E. Bontrop et al.: RFLP analysis of the chimpanzee DR subregion 279
alternative explanation is that DR haplotypes with more than four DRB genes may have been lost during evolution of the hominid lineage, for instance due to unequal cross- ing-over events. The implication of such a phenomenon would be that the organization of the HLA-DR subregion with HLA-DRB1, -DRB2 , -DRB 3 , -DRB4 genes, etc., may have been shuffled around for other species. This might explain why DRB/Taq I RFLP profiles of man and chimpanzee are rather dissimilar (Fig. 4), whereas this is not observed for the MHC DQ region genes (Bontrop et al. 1989).
The identification of 14 distinct DRB/Taq I fragments within our chimpanzee colony (Fig. 5) allowed the defini- tion of at least 11 Patr-DR haplotypes. This number of chimpanzee DR haplotypes probably reflects a minimum quantity. In the human population, members of the HLA- DRw52 (-DR3, -DR5, and -DRw6) and HLA-DRw53 (-DR4, -DR7, and -DR9) supergroups are difficult to distinguish by RFLP analysis (Fei et al. 1986; Tilanus et al. 1986). However, other techniques enable researchers to differentiate between the members of the HLA-DRw52 and -DRw53 supertypic specificities. It is expected that two-dimensional gel electrophoresis of class II molecules and sequence determination will allow us to generate a further subdivision among these 11 chimpanzee DR specificities defined so far.
For most HLA-DR haplotypes the presence of DRB pseudogenes has been demonstrated (Rollini et al. 1985; Larhammar et al. 1985). This situation may also be en- countered for some Patr-DR specificities. For instance, the chimpanzee Hugo (Patr-DR2,2) expresses at least three types of DR molecules as shown by immuno- precipitation studies (Bontrop et al. 1990). This is in con- cordance with an earlier study reporting the isolation of three types of cDNA clones encoding DR molecules from a Hugo chimpanzee cDNA library (Fan et al. 1989). Two of these cDNA clones encode HLA-DR2-1ike molecules while the third cDNA clone encodes an HLA-DRw52-1ike molecule. The current data show that these three DR molecules are encoded by a Patr-DR2,2-homozygous chimpanzee which possesses five different DRB genes (Fig. 4). Thus, the remaining two Patr-DRB genes are possibly pseudogenes. This example again emphasizes that the combination of DRB genes as observed in the human population may be completely different from that observed in the nonhuman primates.
In the chimpanzee colony some preferential RFLP- determined DQA1-DR associations were observed (Table 1). The distribution of the DQA1-DR haplotypes in the present colony more or less resembles that found for the
founder animals. It will be of interest to determine to what extent known DR-DQ linkage disequilibria as observed for different human ethnic populations can be traced back in the chimpanzee colony.
The usefulness of nonhuman primates in immunology
has been hampered by difficulties in characterizing their MHC region products (Watkins et al. 1988). However, typing for the DR subregion of the chimpanzee by means of the RFLP technique is feasible. Moreover, RFLP studies performed on closely related species such as nonhuman primates and man may provide detailed insight into the evolutionary development of the MHC.
Acknowledgments. The expert technical help of Mrs. M.C. Noort is appreciated and Erik van der Reijden is acknowledged for preparing the figures.
References
Andersson, M., B6hme, J., Andersson, G., M611er, E., Thorsby, E., Rask, L., and Peterson, P.: Genomic hybridization with class II transplantation antigen cDNA probes as a complementary technique in tissue typing. Hum Immunol 11: 57-67, 1984
Arden, B. and Klein, J.: Biochemical comparison of major histocom- patibility complex molecules from different subspecies of Mus musculus: evidence for trans-specific evolution of alleles. Proe Natl Acad Sci USA 79: 2342-2346, 1982
Balner, H., Gabb, B. W., D'Amaro, J., van Vreeswijk, W., and Visser, T.P.: Evidence for two linked loci controlling the serologically defined leucocyte antigens of chimpanzees (ChL-A). Tissue An- tigens 4: 313-328, 1974
Balner, H., van Vreeswijk, W., D'Amaro, J., Roger, J., Schreuder, I., and van Rood, J.J.: Chimpanzees share B-cell as well as SD antigens with man. In W.F. Bodmer, J.R. Batchelor, J.G. Bodmer, H. Festenstein, and P. J. Morris (eds.): Histocompatibility Testing 1977, p 412, Munksgaard, Copenhagen, 1977
Barker, D., Schafer, M., and White, R.: Restriction sites containing CpG show a higher frequency of polymorphism in human DNA. Cell 36: 131-135, 1984
Bidwell, J. : DNA-RFLP analysis and genotyping of HLA-DR and -DQ antigens. Immunol Today 9: 18-23, 1988
B6hme, J., Andersson, M., Andersson, G, M611er, E., Peterson, P. A., and Rask, L.: HLA-DR beta genes vary in number between dif- ferent DR specificities whereas the number of DQ beta genes is constant. J Immunol I35: 2149-2155, 1986
Bontrop, R.E., Tilanus, M.G.J. , Mikulski, M., van Eggermond, M. C. J. A., Termijtelen, A., and Giphart, M. J.: Polymorphisms within the HLA-DR3 haplotypes. I. HLA-DR polymorphisms detected at the protein and DNA levels are reflected by T-cell recognition. Immunogenetics 23: 401-405, 1986a
Bontrop, R.E., Schreuder, G.M. Th., Elferink, D.G., Mikulski, M. M. A., Geerse, R., and Giphart, M.J.: Molecular, serologic and functional evidence for an apparent HLA-DR triplet. J lmmunol 137." 211-216, 1986b
Bontrop, R. E., Tilanus, M. G. J., Mikulski, M. M. A., Elferink, D. G., Termijtelen, A., de Vries, R. R. P., van Rood, J. J., and Giphart, M. J.: Polymorphism and complexity of HIM-DR: evidence for in- tra-HLA-DR region crossing-over events. Immunogenetics 27: 40-45, 1988
Bontrop, R. E., Otting, N., Broos, L. A. M., Noort, M. C., Kenter, M., and Jonker, M.: RFLP analysis of the HIM-, ChLA-, and RhLA-DQ alpha chain gene regions: conservation of restriction sites during evolution. Immunogenetics 30: 432-439, 1989
Bontrop, R. E., Elferink, B. G., Otting, N., Jonker, M., and de Vries, R. R. P.: MHC-class II restricted antigen presentation across a species barrier: conservation of restriction determinants. JExp Med 172: 53-59, 1990

280 R.E. Bontrop et al. : RFLP analysis of the chimpanzee DR subregion
Cohen, D., LeGall, I., Marcadet, A., Font, M., Lalouel, J., and Dausset, J.: Clusters of HLA-class II restriction fragments describe allelic series. Proc Natl Acad Sci USA 81: 7870-7874, 1984
Fan, W., Kasahara, M., Gutknecht, J., Klein, D., Mayer, W.E., Jonker, M., and Klein, J.: Shared class II MHC polymorphisms between humans and chimpanzees. Hum Immunol 26: 107-121, 1989
Fei, H., Tilanus, M., van Eggermond, M., and Giphart, M. J.: Reduced complexity of RFLP for HLA-DR typing by the use of a DR/3 3' cDNA probe. Tissue Antigens 28: 129-135, 1986
Figueroa, F., GiJnther, E., and Klein, J.: MHC polymorphisms predating speeiation. Nature 335: 265-267, 1988
Garver, J.J., Pearson, P.L., Estop, A., Dijksman, T.M., Wijnen, L. M. M., Westefeld, A., and Khan, P.M.: Gene assignments to the presumptive homologs of human chromosomes 1,6, 11, 12 and X in the Pongidae and Cercopethecoidea (HGM4). Cytogenet Cell Genet 22: 564-569, 1978
Gyllensten, U.F. and Erlich, H.A.: Ancient roots for polymorphism at the HLA-DQa locus in primates. Proc Natl Acad Sci USA 86: 9986-9990, 1989
Jonker, M. and Balner, H.: Current knowledge of the D/DR region of the major histocompatibility complex of rhesus monkeys and chim- panzees. Hum lmmunol 1: 305-316, 1980
Jonker, M., and Balner, H.: Mixed leukocyte reactivity in chimpanzees. II. Family studies and identification of D antigens. Tissue Antigens 17: 179-194, 1981
Klein, J., Bontrop, R.E., Dawkins, R.L., Erlich, H.A., Gyllensten, U., Heise, E. R., Jones, P. P., Parham, P., Wakelaud, E. K., and Watkins, D. I.: Nomenclature for the major histocompatibility com- plexes of different species: a proposal. Immunogenetics 31: 217-219, 1990
Larhammar, D., Hylding-Nielsen, J. J., Servenius, B., Andersson, G., Rask, L., and Peterson, P. A.: Exon-intron organization and com- plete nucleotide sequence of a human major histocompatibility an- tigen DC$ gene. Proc Natl Acad Sci USA 80: 7313-7317, 1983
Larhammar, D., Servenius, B., Rask, L., and Peterson, P.A.: Characterization of an HLA-DR beta pseudogene. Proc Nat! Acad Sci USA 82: 1475-1479, 1985
Lawlor, D. A., Ward, F. E., Ennis, P. D., Jackson, A. P., and Parham, P.: HLA-A and -B polymorphisms predate the divergence of humans and chimpanzees. Nature 335: 268-271, 1988
Long, E. O., Wake, C. T., Strubin, M., Gross, N., Acolla, R. S., Car- rel, S., and Mach, B.: Isolation of distinct cDNA clones encoding
HLA-DR beta chains by use of an expression assay. Proc Natl Acad Sci USA 79: 7465-7469, 1982
Long, E.O., Wake, C.T., Gorsld, J., and Mach, B.: Complete se- quence of an HLA-DR beta chain deduced from a cDNA clone and identification of multiple non-allelic DR beta chain genes. EMBO J 2: 389-394, 1983
Marcadet, A., O'Connell, P., and Cohen, D.: Standardized Southern blot workshop technique. In B. Dupont (ed.): Immunobiology of HLA, Vol 1, pp. 553-560, Springer, New York, 1987
Mayer, W.E., Jonker, M., Klein, D., Ivanyi, P., van Seventer, G., and Klein, J.: Nucleotide sequences of chimpanzee MHC class I alleles: evidence for trans-species mode of evolution. EMBO J 7: 2765-2774, 1988
Rollini, P., Mach, B., and Gorsld, J.: Linkage map of three HLA-DR chain genes: evidence for a recent duplication event. ProcNatlAcad Sci USA 82: 7197-7201, 1985
Roux-Dosseto, M., Auffray, C., Lillie, J. W., Boss, J. M., Cohen, D., DeMars, R., Mawas, C., Seidman, J.G., and Strominger, J.: Genetic mapping of a human class II antigen ~-chain cDNA clone to the SB region of the HLA complex. Proc Natl Acad Sci USA 80: 6036-6040, 1983
Slierendregt, B. L., Otting, N., Jonker, M., and Bontrop, R. E. : RFLP analysis of the rhesus monkey MHC class II DR subregion. Hum lmmunol, in press, 1990
Spies, T., Sorrentino, R., Boss, J. M., Okada, K., and Strominger, J.: Structural organization of the DR subregion of the human major histocompatibility complex. Proc Natl Acad Sci USA 82: 5165-5169, 1985
Termijtelen, A., Tilanus, M. G. J., Engelen, I., Koning, F., and van Rood, J. J.: Molecular localization of LB-Q1, a DRw52-1ike T cell recognition epitope and identification at the genomic level of associated shared hybridizing fragments. Hum Immunol 19: 255-276, 1987
Tilanus, M. G. J., Morolli, B., van Eggermond, M. C. J. A., Schreuder, G. M. Th., de Vries, R. R. P., and Giphart, M.J.: Dissection of HLA class II haplotypes in HLA-DR4 homozygous individuals. Im- munogenetics 23: 333-340, 1986
Wake, C.T., Long, E.O., and Mach, B.: Allelic polymorphism and complexity of the genes for HLA-DR beta chains-direct analysis by DNA-DNA hybridization. Nature 300: 372-375, 1982
Watkins, D.I., Kannagi, M., Stone, M.E., and Letvin, N.: Major histocompatibility complex class I molecules of nonhuman primates. Eur J Immunol 18: 1425-1432, 1988




















