Tagging Morphogenetic Genes by Insertional Mutagenesis in the Yeast Yarrowia lipolytica

The bZIP-type transcription factor FlbB regulatesdistinct morphogenetic stages of colony formation inAspergillus nidulansmmi_6804 775..789
Oier Etxebeste,1 Erika Herrero-García,2,3
Lidia Araújo-Bazán,3 Ana B. Rodríguez-Urra,2
Aitor Garzia,2 Unai Ugalde2 and Eduardo A. Espeso3*1CIC Nanogune, Tolosa hiribidea 76, 20018 SanSebastian, Spain.2Department of Applied Chemistry, Faculty of Chemistry,University of the Basque Country, 20018 San Sebastian,Spain.3Centro de Investigaciones Biológicas (CSIC), Ramirode Maeztu, 9, 28040 Madrid, Spain.
Summary
Conidiophore formation in Aspergillus nidulansinvolves a developmental programme in which veg-etative hyphae give rise to an ordered succession ofdifferentiated cells: foot cell, stalk, vesicle, metulae,phialides and conidia. The developmental transitionrequires factors that are expressed in vegetativehyphae that activate the expression of the main regu-lator of conidiation, BrlA. One such element is thebZIP-type transcription factor FlbB. We found thatflbB - mutants show defective branching patterns andare susceptible to autolysis under high sorbitol orsucrose concentrations, revealing a role in vegetativegrowth. In addition, FlbB plays a role in conidiophoreinitiation, as its upregulation reduces conidiophorevesicle swelling and generates a reduced number ofmetulae. FlbB was located at the tip of growingmetulae, following a similar pattern as described invegetative hyphae. In wild-type strains, the transitionfrom metulae to phialides could be reversed to gen-erate vegetative hyphae, indicating the existence of aspecific control point at this stage of conidiophoreformation. The combined evidence points to FlbB as akey factor in the transition to asexual development,playing a role at various control points in which theprocess could be reversed.
Introduction
In the ascomycete Aspergillus nidulans vegetativehyphae constitute the starting point to most developmen-tal processes. These totipotent cells extend apicallythrough deposition of new cellular material at the tip(Momany, 2002).
Emergence of vegetative hyphae to the atmospherejointly with light or certain stimuli that are adverse forgrowth activates the transition from vegetative hyphae tothe formation of asexual spores (conidia; Fischer andKües, 2006). In A. nidulans, conidia-bearing structures orconidiophores are formed by the sequential appearanceof a foot cell, a stalk, a vesicle, and a number of modifiedcells called metulae or primary sterigmata. The latteremerge from different points of the vesicle, and give riseto phialides or secondary sterigmata. Phialides are mer-istematic cells, able to produce more than 10 000 mono-nucleated conidia, the last differentiated cell type in thismorphogenetic process (for a review see Adams et al.,1998).
Those stimuli responsible for conidiation induction aretransduced within vegetative hyphae by upstream devel-opmental activators (UDAs), eventually leading to theexpression of the central developmental regulator BrlA(Adams et al., 1998; Yu et al., 2006). Mutations in sixupstream genes cause an aconidial phenotype with acottony (fluffy) appearance and often delayed conidiation(Yu et al., 2006 and references within). Genetic studiesadded essential information regarding the molecular roleplayed by some of these factors and their position in theinduction genetic pathway. However, in most of the casestheir functional relationships remain unknown (Fischerand Kües, 2006 and references therein; Yu et al., 2006).
Recent investigations on the upstream activating factorFlbB, a putative bZIP transcription factor, have shown thatit is found at the hyphal tip, and in the most apical nucleus(Etxebeste et al., 2008). Moreover, localization is func-tionally dependent on another UDA, FlbE, with which FlbBforms a complex (Garzia et al., 2009).
This work extends the range of functions of FlbBbeyond early conidiation induction and adds new featuresthat include cellular dynamics at the hyphal tip. First,
Accepted 11 July, 2009. *For correspondence. E-mail [email protected]; Tel. (+34) 91 837 31 12 ext: 4227; Fax (+34) 91 536 04 32.
Molecular Microbiology (2009) 73(5), 775–789 � doi:10.1111/j.1365-2958.2009.06804.xFirst published online 29 July 2009
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd

defects in FlbB activity generate colonies with an alteredbranching pattern. Second, conidiation induction results inredistribution of FlbB from the most apical nucleus to allnuclei. Third, FlbB is present at the genesis and tips ofgrowing metulae. Finally, we show that conidiation induc-tion is reversible up to the point of phialide formation.These combined findings indicate that conidiophoredevelopment may be viewed as a succession of distinctmodules mechanistically related to vegetative growth, inwhich FlbB plays differential functions.
Results
Dynamics of FlbB localization during development
FlbB was localized in the most apical nucleus and the tipof mature vegetative hyphae (Etxebeste et al., 2008). In amore comprehensive tracking of the protein throughoutdifferent stages of development, it was first found duringgerm tube emergence exclusively at the hyphal tip(Fig. 1A). Nuclear FlbB was observed in hyphae longer
than 30 mm (n = 20), showing that nuclear localizationsucceeded apical localization. When GFP::FlbB was colo-calized with AbpA::mRFP (Araujo-Bazan et al., 2008a;Taheri-Talesh et al., 2008), FlbB accumulated at a regionapical to the ring structure where endocytosis has beenreported to occur (Fig. 1B). Consequently, FlbB localiza-tion coincided with that previously described for theSpitzenkörper, a fungal characteristic organelle governingpolarized cell growth (Virag and Harris, 2006) and whereexocytosis predominantly takes place (Taheri-Taleshet al., 2008).
In Aspergillus, many TFs are affected in their activity orlocalization during mitosis due to the cytoplasmic micro-tubule breakdown or Nuclear Pore Complex (NPC) partialdisassembly (Horio and Oakley, 2005; De Souza andOsmani, 2007). Nuclear FlbB was observed to exit thenucleus at the onset of mitosis (Fig. 2A, panels 5–7).During telophase, FlbB differentially accumulated in themost apical daughter nucleus (N1 in Fig. 2A, panels 8–10)compared with the second daughter nucleus (N2in Fig. 2A, panels 8–10). As the G1 phase advanced,
Fig. 1. FlbB localization in vegetative hyphae.A. FlbB::GFP cellular localization in different phases of vegetative growth. FlbB is located at the tip (white arrowheads) just after thegermination of a spore (left; scale bar = 10 mm) and is maintained during vegetative growth (middle, scale bar = 15 mm); it is also observed inthe most apical nucleus of mature hyphae (right, white arrow; scale bar = 15 mm).B. GFP::FlbB localization (green) in the apex of the growing hypha, delimited by the endocytic machinery marked by AbpA::mRFP (red). Scalebar = 3 mm.
Fig. 2. FlbB cellular localization during mitosis, in a DsfgA genetic background and in air-exposed hyphae.A. FlbB is maintained at the tip (T) of the growing hyphae during mitosis of Aspergillus nidulans vegetative cells. FlbB fluorescence in theapical nucleus (N) is lost during mitosis (panels 5–7) but recovered in G1 phase in the daughter apical nucleus (N1, white arrowhead; panels8–10). FlbB is also observed in the second daughter nucleus (N2, grey arrowhead) but with a minor intensity, disappearing from it along theinterphase (not shown).B. FlbB is observed in distal nuclei in a DsfgA genetic background.C. FlbB cellular localization after the induction of asexual development by air exposure. GFP::FlbB fusion is located into all the nuclei(white arrowheads). White arrow indicates FlbB accumulation at the tip of the cell. Grey arrow indicates a tip of a cell that is lacking in FlbBaccumulation.For all panels, scale bar = 10 mm.
776 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

Role of FlbB cellular localization in conidiation 777
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

fluorescence of the distal daughter nucleus (N2) graduallyfaded and only the most apical nucleus remained fluores-cent (not shown). Apical distribution of FlbB remainedunchanged throughout the vegetative cell cycle. Theseresults showed that apical and nuclear localizations ofFlbB responded to independent mechanisms and thatnuclear localization during interphase most likely involvesthe recovery of nuclear transport upon completion ofmitosis.
Given the requirement of FlbB for conidiation, its distri-bution was analysed in a DsfgA strain, in which conidiationwas de-repressed (Seo et al., 2006). FlbB::GFP wasfound in all the nuclei of DsfgA hyphae, in marked contrastto the wild-type background (Fig. 2B). This change didnot affect nuclear localization dynamics during mitosis(not shown). Apical distribution of FlbB also remainedunaffected. When cells in internal regions of the colony,where conidiation is likely to be induced, were examinedfor FlbB::GFP localization, fluorescence accumulationwas observed in all nuclei (white arrowheads in Fig. 2C),similarly to what is observed in DsfgA hyphae (Fig. 2B).Apical accumulation of FlbB was detected at the tip ofnewly growing branches, where the endocytic machinerywas also recruited indicating the existence of active polargrowth (white arrow in Fig. 2C). Interestingly, in a numberof tips (40%), FlbB apical accumulation was not observed(grey arrow in Fig. 2C), suggesting differences betweenthe new hyphal compartments formed. The exact functionof SfgA remains unknown but its effect on FlbB nucleardistribution would appear to be directly related to conidi-ation induction since it coincides with FlbB distribution inhyphae that are exposed to the air and conidiate in thewild-type strain.
flbB - mutants are affected in hyphal branching
The location of FlbB at the Spitzenkörper region sug-gested a potential link between the FlbB/FlbE complexand the machinery involved in the control of polar growth.Thus, growth kinetic parameters were determined for wild-type strains, as well as various flbB mutants in order toevaluate the incidence of mutations.
The comparative analysis of specific growth rate (m) insurface cultures of the wild-type strain FGSC26 andderived mutants flbB100, flbB101 and flbB102 revealedthat the latter did not significantly diverge from the growthrate of the wild type (m = 0.105 � 0.003). A similarpattern was also observed between strain TN02A3(m = 0.087 � 0.006) and the derived DflbB strain BD143.Comparable values were also obtained in submergedliquid culture (Table 3).
The branching rate, as defined by the peripheral growthunit (PGU; see Experimental procedures), was signifi-cantly altered in all mutants, as shown in Fig. 3. StrainsFGSC26 and TN02A3 presented branches every 329.22and 325.30 mm, respectively, at the colony periphery(n = 20). In contrast, mutants flbB100, flbB101, flbB102and DflbB (BD143) presented branches at intervalsranging between 107.10 and 134.99 mm (n = 20). Thisnear threefold increase in branching rate at the colonyperiphery had a significant effect on morphology, as illus-trated in Fig. 3. Taken together, kinetic studies indicatedthat defective flbB function was manifested as a signifi-cant increase in branching frequency, but did not affectspecific growth rate.
flbB– mutants show an autolytic phenotype underosmotic stress
FlbB is required to promote conidiophore developmentunder aerial emergence plus light (natural stimulus) orcarbon or nitrogen starvation and salt stress (Etxebesteet al., 2008). In order to widen the scope of FlbB function,the four loss-of-function flbB strains and the two wild-typeisogenic strains were tested in diverse stress conditions.In standard minimal medium flbB - strains displayed theexpected ‘fluffy’ phenotype (Etxebeste et al., 2008;Fig. 4A, first row). Addition of either sorbitol (1.2 M) orsucrose (1 M) to the minimal medium caused that all offlbB - mutants showed reduced conidiation compared withwild-type strains and an autolysis phenotype in the centralregion of the colonies (Fig. 4A, central and lower rowsrespectively). Our previous and present data evidence amajor role of FlbB in regulating cell integrity in response to
Fig. 3. Branching pattern of wild type and flbB - mutants. Lactophenol blue-stained peripheral regions showing the branching pattern ofwild-type and flbB - strains. Below are indicated the corresponding PGU values reflecting the average length of hypha between any twobranches. Scale bar = 30 mm.
778 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789
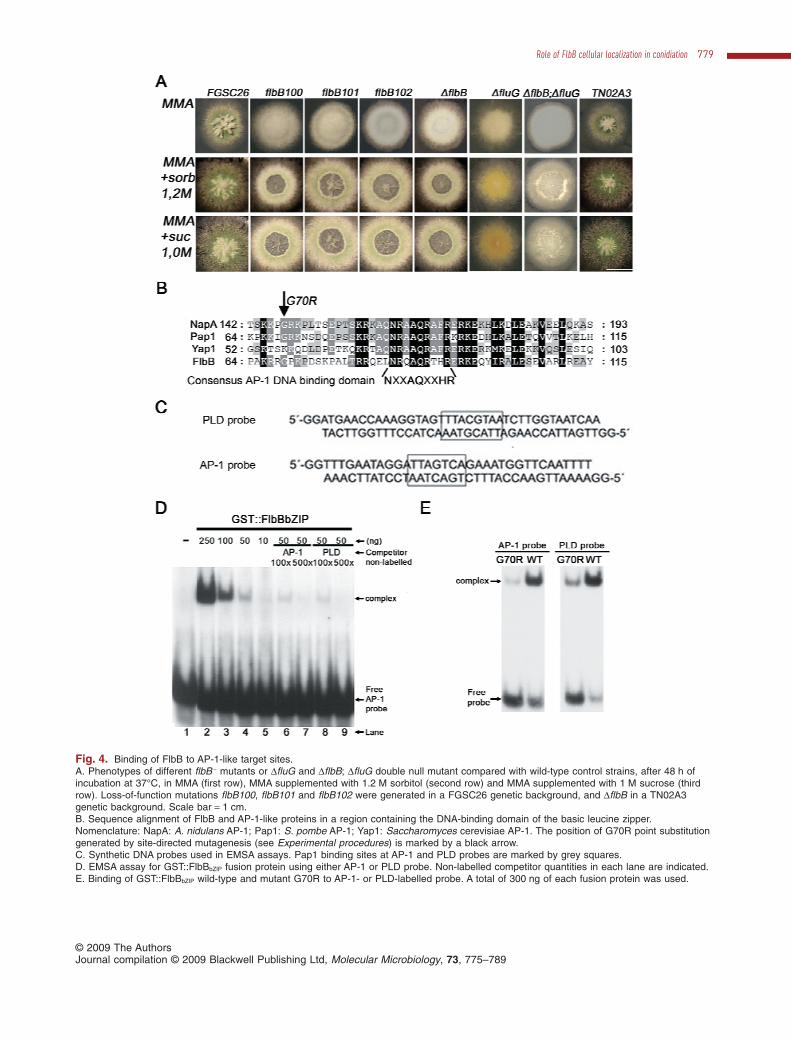
Fig. 4. Binding of FlbB to AP-1-like target sites.A. Phenotypes of different flbB - mutants or DfluG and DflbB; DfluG double null mutant compared with wild-type control strains, after 48 h ofincubation at 37°C, in MMA (first row), MMA supplemented with 1.2 M sorbitol (second row) and MMA supplemented with 1 M sucrose (thirdrow). Loss-of-function mutations flbB100, flbB101 and flbB102 were generated in a FGSC26 genetic background, and DflbB in a TN02A3genetic background. Scale bar = 1 cm.B. Sequence alignment of FlbB and AP-1-like proteins in a region containing the DNA-binding domain of the basic leucine zipper.Nomenclature: NapA: A. nidulans AP-1; Pap1: S. pombe AP-1; Yap1: Saccharomyces cerevisiae AP-1. The position of G70R point substitutiongenerated by site-directed mutagenesis (see Experimental procedures) is marked by a black arrow.C. Synthetic DNA probes used in EMSA assays. Pap1 binding sites at AP-1 and PLD probes are marked by grey squares.D. EMSA assay for GST::FlbBbZIP fusion protein using either AP-1 or PLD probe. Non-labelled competitor quantities in each lane are indicated.E. Binding of GST::FlbBbZIP wild-type and mutant G70R to AP-1- or PLD-labelled probe. A total of 300 ng of each fusion protein was used.
Role of FlbB cellular localization in conidiation 779
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

osmotic stress. First, loss-of-function flbB - mutants do notdisplay autolysis under nutritional stress, as reduction ofcarbon or nitrogen content (Etxebeste et al., 2008). Inaddition, this autolytic phenotype was not seen under saltstresses as in media containing KCl (0.6 M), NaH2PO4
(0.5 M), LiCl (0.2 M) or MgCl2 (0.2 M) (not shown).Second, another mutant in the upstream developmentalpathway, DfluG, did not show an autolytic phenotypeunder similar conditions, but responded with conidiation(Fig. 4A, column 6). However, the double null DflbB; DfluGmutant strain displayed a more evident autolytic response(Fig. 4A, column 7). This result highlights a specific role ofFlbB in regulating conidiation and autolysis under osmoticstress conditions.
FlbB is a putative TF that binds in vitro DNA targetsequences related to stress response
The presence of a conserved NXXAQXXHR motif at theN-terminal end of the bZIP domain among putative FlbBorthologues of filamentous fungi (not shown) relates thisTF with the Pap1 protein from Schizosaccharomycespombe (Fig. 4B). Pap1 belongs to the AP-1-related pro-teins, an important bZIP-containing family of TFs regu-lating the cell oxidative stress response (Glover andHarrison, 1995). These similarities suggested that FlbBmight recognize the same target sequences as Pap1. Totest this hypothesis, a GST fusion expressing the regionbetween residues K67 and N140 of FlbB, GST::FlbBbZIP,was generated (see Experimental procedures), andused for DNA-binding assays to the PLD and AP-1 syn-thetic probes (Toda et al., 1992; Fujii et al., 2000;Fig. 4C). The GST::FlbBbZIP fusion, but not GST alone,was able to bind these sequences confirming our pre-diction (Fig. 4D, lanes 1–5, Fig. 4E). Such DNA-bindingcapacity could be competitively inhibited by theaddition of non-radioactive AP-1 or PLD probes (Fig. 4D,lanes 6–9) indicating a comparable affinity of theFlbB-bZIP fragment for both DNA sequences. Compara-tive binding levels were observed between the GST::Fl-bBbZIP and the either AP-1- or PLD-labelled probes(Fig. 4E).
In mutant flbB100, a single-amino-acid substitutionGly70Arg generated a phenotype barely distinguishablefrom the DflbB allele. As this glycine residue was alsohighly conserved in the bZIP domain (Fig. 4B, blackarrow), the role of this residue on the DNA-binding activityof this region was tested. A G70R mutated version of theGST::FlbBbZIP fusion (see Experimental procedures) wasassayed for binding to both AP-1 and PLD probes.Figure 4E shows that the G70R substitution significantlyreduces the DNA-binding capacity. These results providean interpretation for the loss-of-function phenotype of theflbB100 allele (see Discussion) and extend the region
required for high-affinity binding to target DNA with com-parison to other AP-1 fungal-like proteins.
Since Yu et al. (2006; see also references therein) con-cluded that FlbB acts immediately before the masterconidiation regulator BrlA, in vitro binding analysis of theFlbB bZIP domain to the 3.5 kb region of the brlA pro-moter was considered. An in silico search for putativeFlbB binding sites in this region showed no such sites,and this was further confirmed experimentally (notshown).
FlbB participates in the formation of conidiophorestructures
Synchronized conidiation induction by air exposure (seeExperimental procedures) showed detectable steady-state levels of mRNA of flbB up to the time of vesicleformation (Fischer and Kües, 2006; Etxebeste et al.,2008; references therein). However, no transcript wasdetected 12 h after induction, when most conidiophoresare becoming mature. Thus, we expected to detect FlbBin forming vesicles or in primary or secondary sterigmata,but not in conidiating structures. Microscopy images indi-cated that FlbB delocalized from the tip of the formingvesicle (Fig. 5A). In a late vesicle formation, nuclei havebeen described to align at the dome of the vesicle andundergo quasi-synchronous divisions associated withmetulae production (Mims et al., 1988). A series of newfluorescent accumulations were observed at this stagewhich coincided with budding of metulae (Fig. 5B, left,and Fig. 5C). From this stage, FlbB accumulated princi-pally at the tips of elongating metulae, resembling thelocalization pattern of vegetative hyphae, with theendocytic machinery laying distally to it (Fig. 5C). Inmature conidiophores, metulae and phialides displayed adispersed localization of FlbB (Fig. 5D) and no fluores-cence was visible in conidia, which is in agreement withthe timing of flbB downregulation mentioned above.
Metula to phialide transition: a control point forconidiophore development
The presence of FlbB and the endocytic machinery at thetip of metulae suggested the re-assembly of the polargrowth apparatus as in vegetative cells. Hence we testedwhether the morphogenetic programme of metulae couldbe reverted back to vegetative polarized growth. Thus,liquid cultures in which conidiophore development wasinduced by nitrogen starvation (Etxebeste et al., 2008;Garzia et al., 2009) were transferred back to a mediumcontaining nitrate as a repressor of conidiation (seeExperimental procedures). Ten hours after the transfer tonitrate containing medium, both wild-type strains testeddisplayed conidiophores in which metulae had reverted to
780 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789
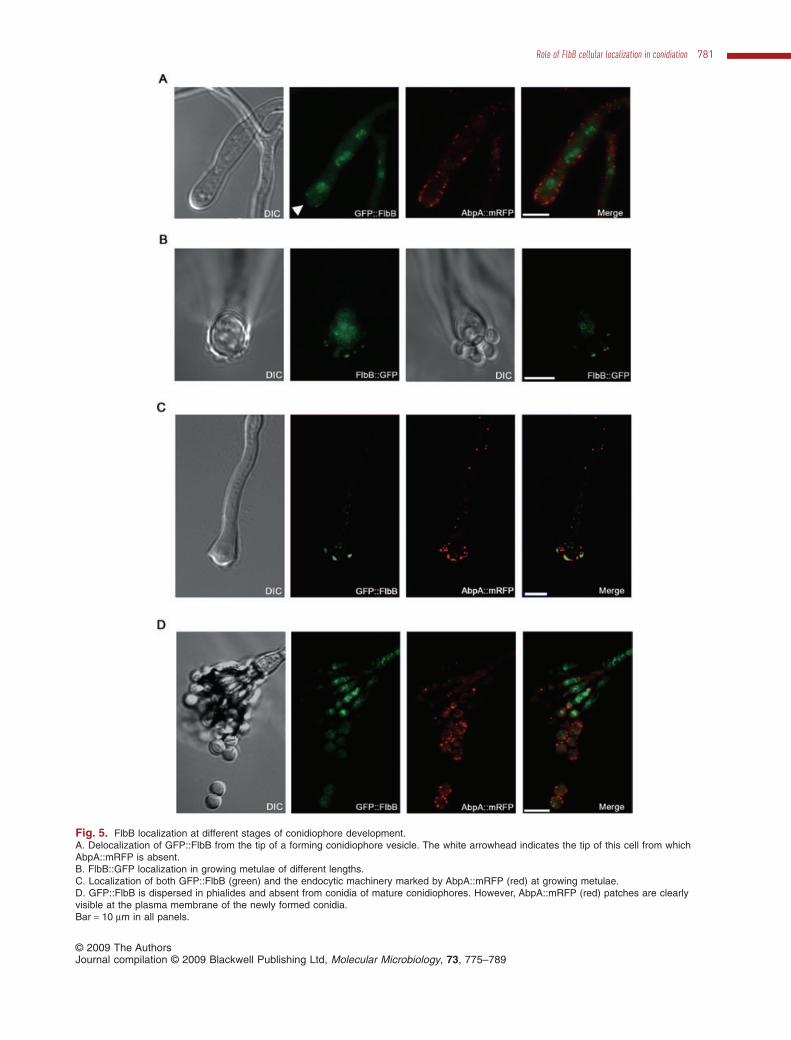
Fig. 5. FlbB localization at different stages of conidiophore development.A. Delocalization of GFP::FlbB from the tip of a forming conidiophore vesicle. The white arrowhead indicates the tip of this cell from whichAbpA::mRFP is absent.B. FlbB::GFP localization in growing metulae of different lengths.C. Localization of both GFP::FlbB (green) and the endocytic machinery marked by AbpA::mRFP (red) at growing metulae.D. GFP::FlbB is dispersed in phialides and absent from conidia of mature conidiophores. However, AbpA::mRFP (red) patches are clearlyvisible at the plasma membrane of the newly formed conidia.Bar = 10 mm in all panels.
Role of FlbB cellular localization in conidiation 781
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

vegetative hyphae and continued to extend without anyvisible phialides or conidia (Fig. 6A; n = 50 conidiophoresat metulae stage). Conidiophores at the phialide stagebefore the media shift to repressing medium showed noreversion, and developed conidia normally from second-ary sterigmata (n = 50 conidiophores). These results indi-cate that metulae not only resemble vegetative hyphaewith regard to FlbB and AbpA distribution, but maintain thepotential to revert to vegetative extension mode. FlbBmight play a role in the transition from vesicle formation tometula elongation and its cessation leading to phialideproduction.
Differential requirement for flbB and brlA expression inconidiophore morphogenesis
To assess a possible role of FlbB in metula formation, thedevelopmental step from metulae to phialides wasstudied. Previous studies showed that BrlA controlled theformation of phialides and conidia (Mirabito et al., 1989;Sewall, 1994; revised in Fischer and Kües, 2006). Whenwe overexpressed both transcripts of brlA, brlAa andbrlAb (see Experimental procedures), phialide-like cellscarrying a single conidium were formed directly from veg-etative hyphae. This observation was in agreement withprevious results from others (Mirabito et al., 1989; Sewall,1994). However, we found that such phenotype was inde-pendent of wild-type or DflbB mutant backgrounds(Fig. 6B). This result provides evidence that brlA mis-scheduled upregulation exerts a bypass in the morphoge-netic programme, effectively converting vegetative hyphatips into functional phialides without any requirement offlbB expression.
When flbB expression was also upregulated in a mis-scheduled fashion (Garzia et al., 2009) complex conidio-phores were produced, although the diameter of vesicleswas reduced to 40% of that of the wild type (n = 50 con-idiophores; Fig. 6C, right). Those vesicles harboured areduced number of metulae which developed normallyinto phialides and conidia (n = 50 conidiophores). Thus,mis-scheduled flbB upregulation resulted in defectivestructures, revealing a dependence of conidiophoredevelopment on FlbB. In all, our results show that forma-
tion of phialides and conidia relies on brlA expression,while conidiophore stalk, vesicle and metulae require flbBexpression in probable conjunction with other as yetunknown factors.
Discussion
The role of upstream developmental factors, which areexpressed in vegetative hyphae but are required for theonset of conidiation, has classically been envisaged tocover processes between the reception of the inducingstimulus and the activation of brlA expression, as the firstand foremost irreversible step in the process (Wieser andAdams, 1995; Han et al., 2004; Seo et al., 2006). Thecombined observations of this study qualify earlier viewsin that induction and conidiophore development appear tobe under the control of UDAs, of which FlbB is a promi-nent member.
FlbB is present from germination onward, and its puta-tive sensory role requires localization in close proximity tothe polar growth machinery. An indication of the intercon-nection between vegetative growth and conidiation asco-ordinated cellular programmes is the effect of partial ortotal loss of function of FlbB on branching frequency. Avariety of proteins have been reported to participate in thecontrol of branching (Westfall and Momany, 2002;Gatherar et al., 2004; Guest et al., 2004). FlbB could be inclose association with these factors involved in polargrowth. Identification of these possible relationships mayreveal new clues on the integration of vegetative growthand morphogenesis in this organism.
The dynamics of FlbB localization have indicated thatapical and nuclear locations are driven by distinctmechanisms. Apical localization is not altered duringmitosis and is dependent on the F-actin cytoskeleton(Garzia et al., 2009) whereas nuclear localization isvariable. During mitosis, when the NPC is partially disas-sembled (De Souza and Osmani, 2007), FlbB is dis-persed, indicating that two important mechanisms fornuclear accumulation in A. nidulans are not in operation:the nuclear import pathway and nuclear retention(Fernandez-Martinez et al., 2003; De Souza and Osmani,2007; Araujo-Bazan et al., 2008b). The FlbB nuclear
Fig. 6. Conidiophore morphological alterations due to changes in growth conditions and brlA or flbB overexpression.A. Conidiophore morphology in the two wild-type strains used to generate flbB - mutants, TN02A3 and FGSC26, after induction of asexualdevelopment by transference of mycelia from liquid MMA to media without nitrate for 20 h (Etxebeste et al., 2008; Garzia et al., 2009) andtransfer for 10 h to a nitrate containing medium which induced the undifferentiation of metulae (white arrows). Different stages ofundifferentiated metulae are shown for strain FGSC26. A magnified view of this type of cells growing directly from the vesicle is shown at thebottom of the fourth panel of the column. A conidiophore bearing all the cell types is shown as a reference (upper panel). Scale bar = 30 mm.B. Production of extremely simplified conidiating structures when the overexpression of both transcripts of brlA, brlAa and brlAb, is induced(for 7 h; lower panels), in both wild-type and DflbB backgrounds. Spores (indicated with a C) were directly produced from single phialide-likestructures formed at the tip of hyphae. Scale bar = 30 mm.C. Simplified but complete conidiophore generated after overexpression of flbB transcript (see Experimental procedures). Vesicle (V), metula(M), phialide (P) and conidiospores (C) are indicated. Scale bar = 30 mm.
782 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

Role of FlbB cellular localization in conidiation 783
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

import mechanism must be highly regulated, as selectivelocalization to the most apical nucleus shortly after exitfrom mitosis indicates. The cytoplasmic localization ofFlbB during mitosis may also be associated with a specificrole of this protein during this phase of the cell cycle,similarly to the interaction between regulators of thecentral pathway of conidiation and cell cycle regulatorssuch as NimX and NimA (Ye et al., 1999). Moreover, thetransition from vegetative growth to asexual developmentcould have an undetermined dependence on cell cycleregulators. In interphase nuclei, FlbB could participate inthe transduction of the environmental signals from the tipto the nucleus, signalling whether conidiation is to beinduced or polar growth resumed.
The specific FlbB nuclear localization in active growingcells depends on SfgA. This upstream developmental TFwas characterized together with SfgB, SfgC and SfgD, asa suppressor of fluG mutations (Seo et al., 2003; 2006). Asuppressing role of SfgA over the flb genes has beenproposed, maintaining cells in undifferentiated vegetativegrowth. It has been anticipated that the sensing of theconidiation-inducing metabolite linked to FluG activitywould inhibit this SfgA activity, allowing Flb factors topromote asexual development (Seo et al., 2006). Theabsence of SfgA in the cell would purportedly allow FlbBaccumulation in most of the nuclei, mimicking the local-ization observed after conidiation induction by exposureof hyphae to air. This feature suggests that the negativeeffect of SfgA on the Flb factors could be exerted byregulating their cellular localization. We are currentlyinvestigating this possibility.
Besides air contact, other nutritional or abiotic stressconditions such as carbon or nitrogen starvation, orelevated saline concentrations require the FlbB/FlbEcomplex activity to induce conidiation (Etxebeste et al.,2008; Garzia et al., 2009). FlbB activity prevents cell lysisunder nitrogen starvation in submerged cultures (Etx-ebeste et al., 2008); here we show that the presence of afully functional FlbB protein is required for cell integrity inresponse to osmotic stress. We therefore hypothesizethat FlbB’s role in regulating conidiophore developmentalso involves preventing uncontrolled autolysis in inducedcells. Cells not subjected to conidiation induction stimuliwould be exposed to nutrient starvation within the colonyand eventually undergo autolysis. FluG has been associ-ated with such a response in ageing liquid cultures (Emriet al., 2008). Our data support evidence that FlbB isrequired for preventing cell lysis under osmotic stress.
Although related to the AP-1 family of transcriptionfactors, FlbB seems to be highly specialized in mediatingconidiation response to different stresses. The AP-1-likehomologue from A. nidulans, NapA, has been describedas having an important role in the oxidative stressresponse (Asano et al., 2007). No evidence of an FlbB-
dependent oxidative response has been observed using arange of H2O2 concentrations. In addition, the DnapAmutant grew and conidiated as the wild-type strain, thusdiscarding a hypothetic role of this TF in radial growth ofthe colony and the induction of conidiation. Those fea-tures and the restrictive phylogenetic distribution of FlbB,limited to filamentous fungi (Etxebeste et al., 2008),suggest that NapA is widely directed to ‘housekeeping’functions under oxidative stress conditions whereas FlbBis rather directed to specifically co-ordinate conidiophoreformation in filamentous fungi.
Our previous results (Etxebeste et al., 2008) and thoseobtained here characterizing the bZIP domain of FlbBstrongly suggest that FlbB is a TF for the upstream devel-opmental pathway. The nearly identical aconidial pheno-type caused by both null and flbB100 alleles supports thetranscriptional role of FlbB, as the G70R substitution inFlbB100 reduces the ability of the bZIP domain to bind thedescribed target sequences. The effect of the G70R sub-stitution on the phenotype shows that other residues notcontained within the sequence NXXAQXXHR are alsorequired to increase DNA-binding ability. Additional rolesfor this glycine residue could be hypothesized as theregion where G70 lays might be involved in interactionswith other transcriptional regulatory elements (Schneiderand Schepartz, 2001) or being part of the putative NuclearLocalization Signal (NLS; Pokorska et al., 2000). Thedimerization through the leucine zipper region of FlbB withother bZIP proteins also becomes a possibility (Vinsonet al., 2006). By these means FlbB would increase itsrange of action in gene regulation.
FlbB localization at the tip of metulae during conidiationresembles that described for vegetative hyphae. Thetransit from metula to phialide necessarily requires a celldivision process significantly different from that whichoccurs during hyphal growth. The cues that controlmetula–phialide transition remain unknown but our obser-vations support the importance of nutrient availabilitysensing mechanisms as a strong signal for conidiation(Etxebeste et al., 2008; Garzia et al., 2009) and here weshow that metula differentiation is aborted by providingnitrogen, resulting in reversion to vegetative growth. Thissupports the view that metulae are totipotent cells whichundergo a special budding programme to form phialides,but still preserve the potential to extend as vegetativecells. This is consistent with findings that mutations infactors involved in polarity establishment and mainte-nance during vegetative growth lead to morphologicalabnormalities in metulae formation, elongation or division,as the Ras GTPase-activating protein GapA (Harispeet al., 2007), the GTPase RacA (Virag et al., 2007) andthe auxiliary proteins for nuclear migration and positioningApsA or ApsB (Veith et al., 2005). Prior stages of devel-opment, vesicle swelling and metula emergence, also
784 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

require controlled doses of FlbB, since its upregulationnotably reduced both vesicle diameter and numbers ofmetulae. Hence, the correct generation of conidiophoresrequires an accurate spatial and temporal pattern of FlbBexpression. StuA and MedA regulate, respectively, thespatial and temporal expression pattern of genes requiredfor conidiophore development (Miller et al., 1992; Miller,1993; Busby et al., 1996). Any possible dependence ofFlbB expression and function on those factors remains tobe investigated.
A renewed outlook of development emerges from thisstudy, according to which conidiophore development canbe analysed as a succession of morphogenetic modulesbearing some elements in common with vegetativegrowth, such as FlbB. This modular outlook may facili-tate a new conception of conidiophore production as areversible mechanism where the activities of the keyregulators BrlA and FlbB are co-ordinated to est-ablish different control points that guarantee accuratedevelopment.
Experimental procedures
Strains, oligonucleotides and culture conditions
Aspergillus nidulans strains employed in this study are listedin Table 1. Strains were cultivated in Aspergillus minimalmedium, MMA (Kafer, 1965), adequately supplemented. Sor-bitol (1.2 M) or sucrose (1.0 M) was added for osmotic stressconditions. alcA-driven overexpression experiments wereperformed essentially as described in Garzia et al. (2009).Strains overexpressing both brlAa and brlAb transcripts wereobtained by transformation of strains BD185 (gfp::flbB) andBD177 (DflbB), respectively, with plasmid pAlcA(p)::brlAa/b::pyroA* which was generated as described below. Strain
BD184, expressing both GFP::FlbB and AbpA::mRFPfusions, was obtained by meiotic recombination betweenstrains BD185 (GFP::FlbB) and BD181 (AbpA::mRFP). StrainBD183 (DsfgA; gfp::flbB) was generated by transformation ofstrain BD185 with the fusion PCR product (Yang et al., 2004)containing the native 5′ (oligonucleotides sfgA-PP1 and sfgA-PP2; 1.25 kb; Table 2) and 3′ (oligonucleotides sfgA-GSP3and sfgA-GFP4; 1.46 kb) untranslated regions of sfgA fusedto Aspergillus fumigatus pyrG gene (oligonucleotides sfgA-SMP1 and sfgA-GFP2; 1.90 kb). The double null (DflbB;DfluG) strain was obtained by transformation of strain BD85.6(DfluG) with the null flbB::pyrG deletion cassette (Etxebesteet al., 2008).
FlbB cellular localization during vegetative growth wasanalysed by inoculating 20 ml of a conidiospore suspension inPetri dishes containing 1 ml of adequately supplementedWatch Minimal Medium (WMM; Penalva, 2005). FlbB in vivolocalization was monitored after 14–18 h of growth at 37°C. Toanalyse FlbB cellular localization during conidiophore devel-opment after air exposure induction, mycelia were grown onthe vertical surface of a piece of solid medium placed in a Petridish. After 16–24 h of incubation at 37°C, nitrogen-free liquidmedium at the same temperature was added until the solidmedium was covered, to provide an aqueous medium for themicroscopic analysis of fluorescence.
Liquid cultures were carried out in 100 ml conical shakeflasks, containing 30 ml of medium, inoculated with 105
spores ml-1 and incubated at 180 r.p.m. at 37°C. Biomasswas collected by filtration at different incubation times, driedovernight at 80°C and weighed in a precision balance.
Light and fluorescence microscopy
Microscopic analyses were performed as described by Etx-ebeste et al. (2008) and Garzia et al. (2009). For fluorescentAbpA and FlbB observation, images were taken using 500and 1000 ms exposures respectively.
Table 1. Aspergillus nidulans strains used in this study.
Strain Genotype Source
FGSC26 biA1; veA1 Kafer (1965)TN02A3 pyrG89; DnkuA::argB; argB2; pyroA4; veA1 Nayak et al. (2006)BD181 abpA::rfp/pyrG; pyrG89, pabaA1, yA2; DnkuA::argB; argB2; veA1 Araujo-Bazan et al. (2008a)flbB100f biA1; flbB100; veA1 Etxebeste et al. (2008)flbB101f biA1; flbB101; veA1 Etxebeste et al. (2008)flbB102f biA1; flbB102; veA1 Etxebeste et al. (2008)BD85.6 pabaA1, pyrG89; DfluG::trpC This studyBD143 pyrG89; DnkuA::argB; argB2; DflbB::pyrG, pyroA4; veA1 Etxebeste et al. (2008)BD145 pabaA1, pyrG89; DfluG::trpC; DflbB::pyrG This studyBD167 pyrG89; DnkuA::argB; argB2; flbB::gfp/pyrG, pyroA4; veA1 Etxebeste et al. (2008)BD177 pyrG89, pabaA1; DnkuA::argB; DflbB::riboB, pyroA4; riboB2, veA1 Garzia et al. (2009)BD181 pabaA1. yA2, pyrG89; abpA::mrfp::pyrG veA1 Araujo-Bazan et al. (2008a)BD183 pyrG89; DnkuA::argB; DsfgA::pyrG; argB2; gfp::flbB, pyroA4; veA1 This studyBD184 pyrG89, pabaA1, yA2; DnkuA::argB; gfp::flbB, pyroA4; abpA::rfp/pyrG; veA1 This studyBD185 pyrG89; DnkuA::argB; argB2; gfp::flbB, pyroA4; veA1 Etxebeste et al. (2008)BD188 pyrG89; DnkuA::argB; argB2; alcA(p)::pyroA, pyroA4; veA1 Garzia et al. (2009)BD190 pyrG89; DnkuA::argB; argB2; alcA(p)::flbB::pyroA, pyroA4; veA1 Garzia et al. (2009)BD194 pyrG89, pabaA1; DnkuA::argB; argB2; DflbB::riboB, alcA(p)::pyroA, pyroA4; riboB2, veA1 Garzia et al. (2009)BD218 pyrG89, pabaA1; DnkuA::argB; argB2; DflbB::riboB, alcA(p)::brlAa/b::pyroA, pyroA4; riboB2, veA1 This studyBD219 pyrG89; DnkuA::argB; argB2; gfp::flbB, alcA(p)::brlAa/b::pyroA, pyroA4; veA1 This study
Role of FlbB cellular localization in conidiation 785
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

Measurement of growth kinetic parameters
Two fundamental parameters to analyse the impact of FlbBfunction on growth kinetics were considered. The specificgrowth rate (m) was calculated from dry weight measure-ments of biomass collected at different incubation times inliquid cultures (Trinci, 1969). Confirmatory measurements insolid media were also performed through equation Kr = w·m(Trinci, 1971; Trinci et al., 1994), from experimental values ofradial extension rate (Kr) and the width of the peripheralgrowth zone (w, determined according to Trinci, 1971;Table 3). The branching frequency at the peripheral region ofmature colonies was determined as the PGU, defined by theaverage hyphal length between any two branching points atthe peripheral region. This was determined by measuringlactophenol blue-stained hyphae backwards from the apicaltip and recording the number of branches (Reeslev andKjoller, 1995; Vesentini et al., 2006). Images were recordedwith a digital camera coupled to a Nikon Optiphot microscope
under bright field optics. Experiments were repeated threetimes and mean values are given.
GST fusion proteins and EMSA analyses
An EcoRI–BamHI cDNA fragment of flbB encoding aminoacids 67–140 and containing the whole bZIP domain wasamplified using oligonucleotides flbBbZIP(+1) and flbBbZIP(-1), andinserted into plasmid pGEX-2T (Pharmacia) to give plasmidpGEX-2T::FlbBbZIP. Gly70Arg FlbB bZIP mutant version wasgenerated using standard mutagenic PCR techniques:pGEX-2T::FlbBbZIP plasmid was amplified with mutant oligo-nucleotides bZIPG70R(+1) and bZIPG70R(-1) and the PCR productwas digested with DpnI before transformation in Escherichiacoli. The presence of the mutation causing the Gly70Argsubstitution at the recombinant plasmid was confirmed bysequencing and it was named as pGEX-2T::FlbBbZIPG70R.Fusion proteins were expressed in E. coli DH1 strain after
Table 2. Oligonucleotides used in this study.
Oligonucleotide Sequence (5′-3′)
flbBbZIP(-1) CGGAATTCTCAGTTTTCCTCCGATATGGAGTTGGCCPLD-A GGATGAACCAAAGGTAGTTTACGTAATCTTGGTAATCAAPLD-B GGTTGATTACCAAGATTACGTAAACTACCTTTGGTTCATAP1-A GGTTTGAATAGGATTAGTCAGAAATGGTTCAATTTTAP1-B GGAAAATTGAACCATTTCTGACTAATCCTATTCAAAbZIPG70R(+1) CCAAACGAAGACGACCAAAACCCGATAGCbZIPG70R(-1) GCTATCGGGTTTTGGTCGTCTTCGTTTGGbrlAp1Up CCCCCGGGGTCTTCGAGCGAGGGGGCbrlAp1Dw GCTCTAGAGGTCGTGCATTCCTCAGGGCbrlAp2Up GCTCTAGACCACAATGGCTGATACCGGACCbrlAp2Dw CGGGATCCGAGGAAGTGGTAAACTGGCGGATGGbrlAa/b(+1) GAAGATCTCCCACCGCAGAAAGGCbrlAa/b(-1) GAAGATCTCTGTTTCATTCATCCCAGCCsfgA-PP1 GGCAGAAACGAACCCTTGGAACCsfgA-PP2 CATGGAGAGAAAGATCGACGGTGGsfgA-SMP1 CCACCGTCGATCTTTCTCTCCATGGGACATCAGATGCTGGATTACTAAGGsfgA-GFP2 GCGAGGTGTATGGTCCGGTCAGTCTGAGAGGAGGCACTGATGCGsfgA-GSP3 TGACCGGACCATACACCTCGCsfgA-GSP4 GGTAATCCGAGTTGGCTTGACTACGG
Table 3. Radial growth rate (Kr), width of the peripheral growth zone (w), surface specific growth rate (ms) and submerged specific growth rate(m) of A. nidulans strains in this study.
Kr (mm h-1)a w (mm)b ms (h-1)c m (h-1)d
FGSC26 0.287 � 0.005e 2.744 � 0.088 0.105 � 0.003 0.1046 � 0.004flbB102 0.2584 � 0.005 2.376 � 0.164 0.109 � 0.0074 0.1035 � 0.0047flbB101 0.2434 � 0.007 2.544 � 0.161 0.096 � 0.0063 0.0997 � 0.005flbB100 0.2765 � 0.011 2.926 � 0.181 0.095 � 0.0065 0.102 � 0.0103TN02A3 0.2564 � 0.006 2.952 � 0.196 0.087 � 0.0060 0.0978 � 0.0047DflbB 0.2233 � 0.007 2.56 � 0.187 0.088 � 0.0064 0.0928 � 0.0055
a. Kr was measured as the linear regression of colony radii on time.b. In order to measure the width of the peripheral growth zone (w), colonies were cut diagonally at different distances from their margin with asterile razor. The growth of the severed hyphae was followed by taking photographs at 15 min intervals with a digital camera coupled to a NikonOptiphot microscope. The width of the peripheral growth zone was determined as the shortest hyphal length with an unaltered growth rate (Trinci,1971; Trinci et al., 1994).c. The surface specific growth rate (ms) was calculated through the equation Kr = w·m (Trinci, 1971).d. The submerged specific growth rate (m) was calculated from dry weight measurements of biomass collected at different incubation times inliquid cultures (Trinci, 1969).e. Values are means of five replicates � s.e.m.
786 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

addition of 0.1 mM IPTG (isopropil-b-D-tiogalactopyranoside;Sigma) to LB medium. The GST fusion protein purificationstep was carried out essentially as described by Pharmacia’sprotocol, but using an elution solution containing 40 mM ofreduced glutathion (plus DTT, 2 mM; Tris-HCl, 50 mM,pH = 8.0). Protein concentration was estimated by Bradfordassay (Bradford, 1976).
Synthetic double-stranded oligonucleotides PLD and AP1were prepared as described by Tilburn et al. (1995), byannealing equimolar amounts of single-stranded oligonucle-otides, PLD-A and PLD-B, or AP1-A and AP1-B respectively.Binding reactions were performed as described (Perez-Esteban et al., 1993; Espeso and Penalva, 1994).
For EMSA studies the brlA promoter was divided into twooverlapping fragments that were amplified by PCR usingoligonucleotide pair brlAp1Up and brlAp1Dw, or brlAp2Upand brlAp2Dw. The amplified overall fragment covered 3.5 kbupstream from brlAa starting ATG codon (Adams et al., 1988;Prade and Timberlake, 1993). Each PCR product was intro-duced into pGEM-T-easy vector (Promega), generatingpGEM-brlAp1 (4.4 kb) and pGEM-brlAp2 (5.2 kb) plasmidsrespectively. The former was XbaI and AvaI double-digested,and the latter XbaI, AvaI and BamHI triple-digested. Thedifferent DNA fragments from the restriction enzyme diges-tion were radio-labelled using Klenow enzyme, purified andassayed by EMSA following Perez-Esteban et al. (1993) andEspeso and Penalva (1994).
Generation of the plasmid for brlA overexpressionexperiments
A DNA fragment corresponding to the region -927 to +1652from brlA locus (being position +1 the brlA cap site describedby Prade and Timberlake, 1993) and coding for both tran-scripts of brlA (brlAa and brlAb) was PCR amplified usingwild-type genomic DNA as template and oligonucleotidesbrlAa/b(+1) and brlAa/b(-1), both adding BglII restrictionsites at each 5′-end. PCR product was digested with BglII andligated to a BamHI-digested pALC-pyroA* plasmid (Garziaet al., 2009) to give pALC::brlAa/b::pyroA*, where properinsertion and orientation of the fragment was determined.
Isolation and manipulation of nucleic acids
The isolation and manipulation of DNA and RNA samplesas well as Southern and Northern blot experiments wereperformed as described in Etxebeste et al. (2008) and Garziaet al. (2009).
Acknowledgements
Eduardo A. Espeso wishes to thank Ministerio de Educacióny Ciencia for support through Grant BFU2006-04185. UnaiUgalde wishes to thank Ministerio de Educación y Ciencia forGrant BFU2004-03499/BMC, and University of The BasqueCountry (UPV/EHU) for Grant GIU08/32. Oier Etxebeste is acontract researcher of the CIC Nanogune. Erika Herrero-García was a contract researcher under the research pro-gramme of Gipuzkoako Foru Aldundia/Diputación Foral de
Gipuzkoa. Aitor Garzia and Ana B. Rodríguez-Urra held pre-doctoral fellowships from Basque Government and UPV/EHUrespectively. Lidia Araújo-Bazán held a predoctoral fellowshipfrom the Ministerio de Educación y Ciencia.
References
Adams, T.H., Boylan, M.T., and Timberlake, W.E. (1988) brlAis necessary and sufficient to direct conidiophore develop-ment in Aspergillus nidulans. Cell 54: 353–362.
Adams, T.H., Wieser, J.K., and Yu, J.H. (1998) Asexualsporulation in Aspergillus nidulans. Microbiol Mol Biol Rev62: 35–54.
Araujo-Bazan, L., Penalva, M.A., and Espeso, E.A. (2008a)Preferential localization of the endocytic internalizationmachinery to hyphal tips underlies polarization of the actincytoskeleton in Aspergillus nidulans. Mol Microbiol 67:891–905.
Araujo-Bazan, L., Fernandez-Martinez, J., Rios, V.M.d.l, Etx-ebeste, O., Albar, J.P., Penalva, M.A., et al. (2008b) NapAand NapB are the Aspergillus nidulans Nap/SET familymembers and NapB is a nuclear protein specifically inter-acting with importin-a. Fungal Genet Biol 45: 278–291.
Asano, Y., Hagiwara, D., Yamashino, T., and Mizuno, T.(2007) Characterization of the bZip-type transcriptionfactor NapA with reference to oxidative stress response inAspergillus nidulans. Biosci Biotechnol Biochem 71: 1800–1803.
Bradford, M.M. (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal Biochem 72: 248–254.
Busby, T.M., Miller, K.Y., and Miller, B.L. (1996) Suppressionand enhancement of the Aspergillus nidulans medusamutation by altered dosage of the bristle and stuntedgenes. Genetics 143: 155–163.
De Souza, C.P.C., and Osmani, S.A. (2007) Mitosis, not justopen or closed. Eukaryot Cell 6: 1521–1527.
Emri, T., Molnar, Z., Szilagyi, M., and Pocsi, I. (2008) Regu-lation of autolysis in Aspergillus nidulans. Appl BiochemBiotechnol 151: 211–220.
Espeso, E.A., and Penalva, M.A. (1994) In vitro binding of thetwo-finger repressor CreA to several consensus and non-consensus sites at the ipnA upstream region is contextdependent. FEBS Lett 342: 43–48.
Etxebeste, O., Ni, M., Garzia, A., Kwon, N.J., Fischer, R., Yu,J.H., et al. (2008) Basic-zipper-type transcription factorFlbB controls asexual development in Aspergillus nidulans.Eukaryot Cell 7: 38–48.
Fernandez-Martinez, J., Brown, C.V., Diez, E., Tilburn, J.,Arst, H.N., Penalva, M.A., and Espeso, E.A. (2003)Overlap of nuclear localisation signal and specific DNA-binding residues within the zinc finger domain of PacC.J Mol Biol 334: 667–684.
Fischer, R., and Kües, U. (2006) Asexual sporulation inmycelial fungi. In The Mycota, Vol. I: Growth, Differentiationand Sexuality. Kües, U., and Fischer, R. (eds). Berlin:Springer-Verlag, pp. 263–292.
Fujii, Y., Shimizu, T., Toda, T., Yanagida, M., and Hakoshima,T. (2000) Structural basis for the diversity of DNA recogni-
Role of FlbB cellular localization in conidiation 787
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

tion by bZIP transcription factors. Nat Struct Mol Biol 7:889–893.
Garzia, A., Etxebeste, O., Herrero-Garcia, E., Fischer, R.,Espeso, E.A., and Ugalde, U. (2009) Aspergillus nidulansFlbE is an upstream developmental activator of conidiationfunctionally associated with the putative transcription factorFlbB. Mol Microbiol 71: 172–184.
Gatherar, I.M., Pollerman, S., Dunn-Coleman, N., andTurner, G. (2004) Identification of a novel gene hbrBrequired for polarised growth in Aspergillus nidulans.Fungal Genet Biol 41: 463–471.
Glover, J.N.M., and Harrison, S.C. (1995) Crystal structure ofthe heterodimeric bZIP transcription factor c-Fos-c-Junbound to DNA. Nature 373: 257–261.
Guest, G.M., Lin, X., and Momany, M. (2004) Aspergillusnidulans RhoA is involved in polar growth, branching, andcell wall synthesis. Fungal Genet Biol 41: 13–22.
Han, K.H., Seo, J.A., and Yu, J.H. (2004) Regulators ofG-protein signalling in Aspergillus nidulans: RgsA down-regulates stress response and stimulates asexual sporula-tion through attenuation of GanB (Ga) signalling. MolMicrobiol 53: 529–540.
Harispe, L., Portela, C., Scazzocchio, C., Penalva, M.A., andGorfinkiel, L. (2007) Ras GTPase-activating protein regu-lation of actin cytoskeleton and hyphal polarity in Aspergil-lus nidulans. Eukaryot Cell 7: 141–153.
Horio, T., and Oakley, B.R. (2005) The role of microtubules inrapid hyphal tip growth of Aspergillus nidulans. Mol BiolCell 16: 918–926.
Kafer, E. (1965) Origins of translocations in Aspergillusnidulans. Genetics 52: 217–232.
Miller, B. (1993) The medA gene of Aspergillus nidulans.J Cell Biochem 53 (Suppl. 17C): 44.
Miller, K.Y., Wu, J., and Miller, B.L. (1992) StuA is required forcell pattern formation in Aspergillus. Genes Dev 6: 1770–1782.
Mims, C.W., Richardson, W.D., and Timberlake, W.E. (1988)Ultrastructural analysis of conidiophore development in thefungus Aspergillus nidulans using freeze-substitution. Pro-toplasma 244: 132–141.
Mirabito, P.M., Adams, T.H., and Timberlake, W.E. (1989)Interactions of three sequentially expressed genes controltemporal and spatial specificity in Aspergillus development.Cell 57: 859–868.
Momany, M. (2002) Polarity in filamentous fungi: establish-ment, maintenance and new axes. Curr Opin Microbiol 5:580–585.
Nayak, T., Szewczyk, E., Oakley, C.E., Osmani, A., Ukil, L.,Murray, S.L., et al. (2006) A versatile and efficient gene-targeting system for Aspergillus nidulans. Genetics 172:1557–1566.
Penalva, M.A. (2005) Tracing the endocytic pathway ofAspergillus nidulans with FM4-64. Fungal Genet Biol 42:963–975.
Perez-Esteban, B., Orejas, M., Gomez-Pardo, E., andPenalva, M.A. (1993) Molecular characterization of afungal secondary metabolism promoter: transcription of theAspergillus nidulans isopenicillin N synthetase gene ismodulated by upstream negative elements. Mol Microbiol9: 881–895.
Pokorska, A., Drevet, C., and Scazzocchio, C. (2000) The
analysis of the transcriptional activator PrnA reveals a tri-partite nuclear localisation sequence. J Mol Biol 298: 585–596.
Prade, R.A., and Timberlake, W.E. (1993) The Aspergillusnidulans brlA regulatory locus consists of overlapping tran-scription units that are individually required for conidio-phore development. EMBO J 12: 2439–2447.
Reeslev, M., and Kjoller, A. (1995) Comparison of biomassdry weights and radial growth rates of fungal colonies onmedia solidified with different gelling compounds. ApplEnviron Microbiol 61: 4236–4239.
Schneider, T.L., and Schepartz, A. (2001) Hepatitis B virusprotein pX enhances the monomer assembly pathwayof bZIP·DNA complexes. Biochemistry 40: 2835–2843.
Seo, J.A., Guan, Y., and Yu, J.H. (2003) Suppressor muta-tions bypass the requirement of fluG for asexual sporula-tion and sterigmatocystin production in Aspergillusnidulans. Genetics 165: 1083–1093.
Seo, J.A., Guan, Y., and Yu, J.H. (2006) FluG-dependentasexual development in Aspergillus nidulans occurs viaderepression. Genetics 172: 1535–1544.
Sewall, T.C. (1994) Cellular effects of misscheduled brlA,abaA, and wetA expression in Aspergillus nidulans. Can JMicrobiol 40: 1035–1042.
Taheri-Talesh, N., Horio, T., Araujo-Bazan, L., Dou, X.,Espeso, E.A., Penalva, M.A., et al. (2008) The tip growthapparatus of Aspergillus nidulans. Mol Biol Cell 19: 1439–1449.
Tilburn, J., Sarkar, S., Widdick, D.A., Espeso, E.A., Orejas,M., Mungroo, J., et al. (1995) The Aspergillus PacC zincfinger transcription factor mediates regulation of both acid-and alkaline-expressed genes by ambient pH. EMBO J 14:779–790.
Toda, T., Shimanuki, M., Saka, Y., Yamano, H., Adachi, Y.,Shirakawa, M., et al. (1992) Fission yeast Pap1-dependenttranscription is negatively regulated by an essential nuclearprotein, Crm1. Mol Cell Biol 12: 5474–5484.
Trinci, A.P.J. (1969) A kinetic study of the growth of Aspergil-lus nidulans and other fungi. J Gen Microbiol 57: 11–24.
Trinci, A.P.J. (1971) Influence of the width of the peripheralgrowth zone on the radial growth rate of fungal colonies onsolid media. J Gen Micrtobiol 67: 325–344.
Trinci, A.P.J., Wiebe, M.G., and Robson, G.D. (1994) Themycelium as an integrated entity. In The Mycota I: Growth,Differentiation and Sexuality. Wessels, J.G.H., and Mein-hardt, F. (eds). Berlin-Heidelberg: Springer-Verlag, pp.175–193.
Veith, D., Scherr, N., Efimov, V.P., and Fischer, R. (2005)Role of the spindle-pole-body protein ApsB and the cortexprotein ApsA in microtubule organization and nuclearmigration in Aspergillus nidulans. J Cell Sci 118: 3705–3716.
Vesentini, D., Dickinson, D.J., and Murphy, R.J. (2006) Fun-gicides affect the production of extracellular mucilaginousmaterial (ECMM) and the peripheral growth unit (PGU) intwo wood-rotting basidiomycetes. Mycol Res 110: 1207–1213.
Vinson, C., Acharya, A., and Taparowsky, E.J. (2006) Deci-phering B-ZIP transcription factor interactions in vitro andin vivo. Biochim Biophys Acta 1759: 4–12.
788 O. Etxebeste et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789

Virag, A., and Harris, S.D. (2006) The Spitzenkörper: amolecular perspective. Mycol Res 110: 4–13.
Virag, A., Lee, M.P., Si, H., and Harris, S.D. (2007) Regulationof hyphal morphogenesis by Cdc42 and Rac1 homologuesin Aspergillus nidulans. Mol Microbiol 66: 1579–1596.
Westfall, P.J., and Momany, M. (2002) Aspergillus nidulansseptin AspB plays pre- and postmitotic roles in septum,branch, and conidiophore development. Mol Biol Cell 13:110–118.
Wieser, J., and Adams, T.H. (1995) flbD encodes a Myb-likeDNA-binding protein that coordinates initiation of Aspergil-lus nidulans conidiophore development. Genes Dev 9:491–502.
Yang, L., Ukil, L., Osmani, A., Nahm, F., Davies, J., DeSouza, C.P.C., et al. (2004) Rapid production of genereplacement constructs and generation of a green fluores-cent protein-tagged centromeric marker in Aspergillusnidulans. Eukaryot Cell 3: 1359–1362.
Ye, X.S., Lee, S.L., Wolkow, T.D., McGuire, S.L., Hamer,J.E., Wood, G.C., and Osmani, S.A. (1999) Interactionbetween developmental and cell cycle regulators isrequired for morphogenesis in Aspergillus nidulans. EMBOJ 18: 6994–7001.
Yu, J.H., Mah, J.H., and Seo, J.A. (2006) Growth and devel-opmental control in the model and pathogenic Aspergilli.Eukaryot Cell 5: 1577–1584.
Role of FlbB cellular localization in conidiation 789
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 775–789
Copyright © 2022 FDOKUMEN