
The Bone Marrow—Cardiac Axis of Myocardial Regeneration
13
The Bone Marrow—Cardiac Axis of Myocardial Regeneration Ronglih Liao, Otmar Pfister, Mohit Jain, and Frederic Mouquet Congestive heart failure remains the leading cause of morbidity and mortality in the developed world. Current therapies do not address the underlying pathophysiology of this disease, namely, the progressive loss of functional cardiomyocytes. The notion of repairing or regenerating lost myo- cardium via cell-based therapies remains highly appealing. The recent identification of adult stem cells, including both cardiac stem/progenitor cells and bone marrow stem cells, has triggered an explosive interest in using these cells for physiolo- gically relevant cardiomyogenesis. Enthusiasm for cardiac regeneration via cell therapy has further been fueled by the many encouraging reports in both animals and human studies. Further intensive research in basic science and clinical arenas are needed to make this next great frontier in cardio- vascular regenerative medicine a reality. In this review, we focus on the role of bone marrow– derived stem cells and cardiac stem/progenitor cells in cardiomyocyte homeostasis and myocardial repair and regeneration, as well as provide a brief overview of current clinical trials using cell-based therapeutic approaches in patients with heart disease. n 2007 Elsevier Inc. All rights reserved. D espite advances in the treatment of con- gestive heart failure, morbidity and mortal- ity remain inappropriately high. 1 This medical epidemic has only continued to escalate, given an overall aging population and the greater number of patients surviving an initial myocardial infarc- tion (MI). The pathophysiology of post-MI heart failure is driven by the loss of cardiomyocytes, either due to acute ischemic necrosis or chronic apoptosis, and by the inability of the remaining cardiomyocytes to adequately compensate. As such, the concept of repairing or regenerating lost myocardium via cell-based therapies (termed cardiomyoplasty) remains highly appealing. Over the past decade, much research has focused upon identifying the ideal cell type with which to promote myocardial regeneration. Thus far, sev- eral cell types have been investigated in animal models including, but not limited to, fetal cardiomyocytes, 2,3 fibroblasts, skeletal myo- blasts, 4-7 and endothelial progenitor cells [EPC]. 8,9 Results with all these cell types have generally been encouraging with regard to bene- ficial post-MI remodeling, albeit none have resulted in definitive differentiation into physio- logically significant, force-generating cardiomyo- cytes. Five years ago, striking reports suggested for the first time that bone marrow–derived stem cells (hematopoietic stem cells [HSC]) may have the potential to regenerate significant amounts of lost myocardium in mice after MI, 10,11 creating overwhelming enthusiasm and subsequent skep- ticism in the field of cardiac repair and regenera- tion. More recently, the identification of resident cardiac stem/progenitor cells by several groups, including ours, 12-15 has brought about a second wave of scientific interest. These findings have advanced our understanding of myocardial biol- ogy and physiology and have introduced the new paradigm of the heart as a non–terminally differentiated organ. Despite the inability of myocardial tissue to adequately “self-heal” after acute injury, as well as the controversy regarding From the Cardiac Muscle Research Laboratory, Cardiovascular Division, Department of Medicine, Brigham and Women's Hospital, Harvard Medical School, Boston, MA. Address reprint requests to Dr Ronglih Liao, Cardiac Muscle Research Laboratory, Cardiovascular Division, Department of Medicine, Brigham and Women's Hospital, Harvard Medical School, 77 Ave Louis Pasteur, New Research Bldg, NRB 431, Boston, MA 02115. E-mail: [email protected] 0033-0620/$ - see front matter n 2007 Elsevier Inc. All rights reserved. doi: 10.1016/j.pcad.2007.03.001 Progress in Cardiovascular Diseases, Vol. 50 No. 1 (July/August), 2007: pp 18-30 18
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The Bone Marrow—Cardiac Axis of Myocardial Regeneration

The Bone Marrow—Cardiac Axis ofMyocardial Regeneration
Ronglih Liao, Otmar Pfister, Mohit Jain, and Frederic Mouquet
Congestive heart failure remains the leading causeof morbidity and mortality in the developed world.Current therapies do not address the underlyingpathophysiology of this disease, namely, theprogressive loss of functional cardiomyocytes.The notion of repairing or regenerating lost myo-cardium via cell-based therapies remains highlyappealing. The recent identification of adult stemcells, including both cardiac stem/progenitor cellsand bone marrow stem cells, has triggered anexplosive interest in using these cells for physiolo-gically relevant cardiomyogenesis. Enthusiasm forcardiac regeneration via cell therapy has furtherbeen fueled by the many encouraging reports inboth animals and human studies. Further intensiveresearch in basic science and clinical arenas areneeded to make this next great frontier in cardio-vascular regenerative medicine a reality. In thisreview, we focus on the role of bone marrow–derived stem cells and cardiac stem/progenitorcells in cardiomyocyte homeostasis and myocardialrepair and regeneration, as well as provide a briefoverview of current clinical trials using cell-basedtherapeutic approaches in patients with heartdisease.n 2007 Elsevier Inc. All rights reserved.
D espite advances in the treatment of con-gestive heart failure, morbidity and mortal-
From the Cardiac Muscle Research Laboratory,ardiovascular Division, Department of Medicine,righam and Women's Hospital, Harvard Medicalchool, Boston, MA.Address reprint requests to Dr Ronglih Liao, Cardiacuscle Research Laboratory, Cardiovascular Division,epartment of Medicine, Brigham and Women'sospital, Harvard Medical School, 77 Ave Louis Pasteur,ew Research Bldg, NRB 431, Boston, MA 02115.-mail: [email protected]/$ - see front mattern 2007 Elsevier Inc. All rights reserved.doi: 10.1016/j.pcad.2007.03.001
CBS
MDHNE
Progress in Cardiovasc18
ity remain inappropriately high.1 This medicalepidemic has only continued to escalate, given anoverall aging population and the greater numberof patients surviving an initial myocardial infarc-tion (MI). The pathophysiology of post-MI heartfailure is driven by the loss of cardiomyocytes,either due to acute ischemic necrosis or chronicapoptosis, and by the inability of the remainingcardiomyocytes to adequately compensate. Assuch, the concept of repairing or regeneratinglost myocardium via cell-based therapies (termedcardiomyoplasty) remains highly appealing. Overthe past decade, much research has focused uponidentifying the ideal cell type with which topromote myocardial regeneration. Thus far, sev-eral cell types have been investigated in animalmodels including, but not limited to, fetalcardiomyocytes,2,3 fibroblasts, skeletal myo-blasts,4-7 and endothelial progenitor cells[EPC].8,9 Results with all these cell types havegenerally been encouraging with regard to bene-ficial post-MI remodeling, albeit none haveresulted in definitive differentiation into physio-logically significant, force-generating cardiomyo-cytes. Five years ago, striking reports suggestedfor the first time that bone marrow–derived stemcells (hematopoietic stem cells [HSC]) may havethe potential to regenerate significant amounts oflost myocardium in mice after MI,10,11 creatingoverwhelming enthusiasm and subsequent skep-ticism in the field of cardiac repair and regenera-tion. More recently, the identification of residentcardiac stem/progenitor cells by several groups,including ours,12-15 has brought about a secondwave of scientific interest. These findings haveadvanced our understanding of myocardial biol-ogy and physiology and have introduced the newparadigm of the heart as a non–terminallydifferentiated organ. Despite the inability ofmyocardial tissue to adequately “self-heal” afteracute injury, as well as the controversy regarding
ular Diseases, Vol. 50 No. 1 (July/August), 2007: pp 18-30

19CARDIAC AXIS OF MYOCARDIAL REGENERATION
the ideal cell type for therapy and a lack ofunderstanding regarding the underlying cellularmechanisms mediating cardiac regeneration, weremain cautiously optimistic that cell-basedtherapies may be sufficiently developed to effec-tively regenerate myocardium after cardiac injury.In this review, we focus on the role of bone
marrow–derived stem cells and cardiac stem/progenitor cells in cardiomyocyte homeostasisand myocardial repair and regeneration, as well asprovide a brief overview of current clinical trialsusing cell-based therapeutic approaches inpatients with heart disease.
Cardiac Stem/Progenitor Cells
For decades, the adult heart has been thought toexist as a terminally differentiated organ withlimited proliferative capacity. Cardiomyocytesundergo hypertrophy, rather then hyperplasia,in response to hemodynamic stress, in contrastto other tissues such as liver, intestine, andskeletal muscle. These long-held tenets ofmyocardial biology have recently been chal-lenged, and a new paradigm of the heart as apartially self-renewing organ has been proposed.Although evidence challenging the old belief hasfermented for some time,16,17 the identificationof resident cardiac stem/progenitor cells (sum-
Table 1. Summery of Reported Ca
Surface MarkersGeneExpression Self Renewal
In VitroDifferenti
SP Sca-1+, CD31+,CD34−, c-kit− ⁎,CD45−, Isl1−
Nkx2.5,GATA4,MEF2C,Tie2
Yes CM, CE,
c-kit Sca-1+, SP+ †Lin−, CD45−,CD31−, CD34−
Nkx2.5,GATA4,MEF2C
Yes CM, EC,
Sca-1 CD31+, CD34−,Lin−, CD45−,c-kit−‡
Nkx2.5‡,GATA4,MEF2C,Tie2
ND CM
Isl-1 Lin−, CD45−,c-kit−, CD34−,CD31−, SP−
Nkx2.5,GATA4
Yes CM
Abbreviations: CM, cardiomyocyte; EC, endothelial cell; SMC⁎ C-kit might be cleaved by cardiac tissue enzymatic digestion
et al reported 2 subpopulations of CD31+ and CD31− among carin isolation procedures used between groups.† The SP phenotype was not addressed through Hoechst exc‡ Matsuura et al reported 40% and 10% of Sca-1 cells expre
marized in Table 1) has brought this newconcept to the scientific forefront12,13,15,18,22,24
and suggests the capacity of adult myocardiumto maintain physiological homeostasis, at leastpartially, through resident cardiac stem cells.
The report by Hierlihy and colleagues18 in 2002was the first identifying the presence of a stemcell–like population in adult hearts based on theirspecific ability to efflux Hoechst dye. SuchHoechst-effluxing capacity was first introducedto identify highly enriched hematopoietic stemcell populations, termed side population (SP) stemcells, from bone marrow.25 Recently, this metho-dology has been used to identify tissue-specificstem/progenitor cells in various adult organsincluding pancreas, pituitary, testis, mammarygland, lung, liver, skeletal muscle, liver, lung aswell as heart (for review, see Asakura andRudnicki26 and Challen and Little27). Usingimmunohistochemistry analysis, Hierlihy et alfound that adult myocardium retains a specific SPcell population, capable of tissue-specific differ-entiation into cardiomyocytes, in vitro.18 In 2003,Beltrami et al thoroughly described a populationof cardiac stem cells (c-kit+ cells) found inclusters and residing among cardiomyocytes inadult hearts.12 In vitro, cardiac c-kit+ cells seem tobe clonogenic and were able to undergo selfrenewal and differentiation into cardiac cell
rdiac Stem/Progenitor Cells
ationIn VivoDifferentiation Species Reference
SMC CM, EC Mice, pig,human
Hierlihy et al, 200218
Pfister et al, 200515
Mouquet et al, 200519
Martin et al, 200413
Messina et al, 200314
SMC CM, EC, SMC Mice, rat,dog, pig,human
Urbanek et al, 200320
Beltrami et al, 200312
Linke et al, 200521
CM Mice, dog Oh et al, 200322
Matsuura et al, 200423
ND Mice, rat,human
Laugwitz et al, 200524
, smooth muscle cell; ND, not determined.. Martin et al found no expression of CD31, whereas Pfistediac SP cells. These differences may due to slight variation
lusion but through MDR-1 expression.ssing CD45 and CD34, respectively.
r

20 LIAO ET AL
lineages (cardiomyocytes, endothelial, smoothmuscle cell). More importantly, these c-kit+cells, when implanted in mouse hearts after MI,retained the capacity for differentiation intocardiomyocytes in vivo. These in vivo data are ofboth scientific and clinical significance, as theystrongly implicate the regeneration potential ofcardiac stem cells in injured hearts. In the sameyear of the report of Beltrami et al, Oh et al used adifferent stem cell marker, Sca-1, to identify yetanother population of resident cardiac progenitorcells in adult hearts.22 Similarly, these Sca1+ cellswere found to be capable of differentiation intocardiomyocytes, in vitro and in vivo, in responseto 5-azacytidine and myocardial ischemia, respec-tively.22 In addition to the initial observationidentifying SP cells in adult myocardium, severalgroups, including ours, have confirmed thepresence of such progenitor cell populations inadult hearts.13,15 Martin et al reported expressionof α-sarcomeric actinin in cardiac SP cellscocultured with other cardiac cells as well asdemonstrated the presence of SP cells in humanmyocardium.13,18 Work performed by Tomita andcolleagues documented the generation of neuro-sphere-like clusters, referred to as cardiospheres,from neonatal cardiac SP cells.28 Similar to thecardiospheres described by Messina et al,14
cardiospheres derived from cardiac SP cells havebeen shown to harbor clonogenic cells withremarkable multilineage differentiation poten-tial.28 These cardiospheres expressed cardiac,smooth muscle, and, interestingly, neuronalgenes and proteins. Data from our group demon-strated not only the capacity for biochemical, butalso functional, cardiomyogenic differentiation incardiac SP cells.15 More importantly, our studydemonstrated that among cardiac SP cells, cardi-omyogenic differentiation is restricted to cellsnegative for CD31 expression and positive for Sca-1 expression (CD31−/Sca-1+ SP cells)15 Althoughthe in vitro cardiomyogenic differentiation poten-tial of cardiac SP cells has been consistentlydemonstrated, less is known about the ability ofthese cells to undergo cardiomyogenic differentia-tion in vivo. Recently, Wada and colleaguesstudied the homing and differentiation efficiencyof intravenously injected cardiac SP cells in amyocardial cryoinjury rat model.29 Neonatal ratcardiac SP cells were found to be able to home toareas of injured myocardium and undergo differ-
entiation into cardiomyocytes, endothelial cells,smooth muscle cells, and fibroblasts. Anotherpotential marker for cardiac stem/progenitor cells,Isl-1 (LIM homeodomain transcription factor),was more recently reported by Laugwitz et al.24
These cells were found to harbor similar cardio-myogenic potential in vitro, although they werephenotypically distinct from SP cells. As moreinformation becomes available regarding cardiacstem/progenitor cells, a key question that remainsis whether these seemingly unique progenitorpopulations described above are truly distinctfrom each other or represent the same populationof progenitor cells at different stages in thedifferentiation process.
Bone Marrow–Derived AdultStem Cells
The bone marrow is known to be an excellentreservoir for many adult stem cells, and bonemarrow–derived stem cells have been used to treathematologic disorders for decades. Recent reportshave demonstrated that bonemarrow–derived stemcells are able to traverse cell lineage boundariesand transdifferentiate into hepatocytes, endothelialcells, skeletal muscle, and neurons upon properstimulation.30-32 Although the ability of bonemarrow–derived stem cells to transdifferentiateinto cardiomyocytes remains highly controversial,much of the recent progress in regenerativecardiovascular research, both in animals andhuman beings, has been achieved using bonemarrow–derived stem cell populations, includingHSC, mesenchymal stem cells (MSC), and EPC.
Hematopoietic Stem Cells
Hematopoietic stem cells can be isolated frombone marrow cells through selective sorting for aparticular set of surface receptors (Lineage−, c-kit+, Sca-1+, CD34lo, CD38hi)33,34 and represent theprototypical adult stem cell population. Theability of HSC to reconstitute the hematopoieticsystem of a myeloablated host led to the firstclinical application of adult stem cells more than 3decades ago.35 Despite the failure of studies todefinitely prove differentiation of HSC intocardiomyocytes in vitro, several studies in micehave demonstrated the potential of HSC todifferentiate into cardiomyocytes or vascularcells after cardiac injury in vivo.36-38

21CARDIAC AXIS OF MYOCARDIAL REGENERATION
Mesenchymal Stem Cells
Within the bonemarrow stroma resides a subset ofnonhematopoietic cells that have the potential todifferentiate into cells of mesenchymal origin.39,40
These MSC represent approximately 0.001% to0.01% of the total nucleated marrow cell popula-tion, a concentration 10-fold lower than theirhematopoietic counterparts. Mesenchymal stemcells are self-renewing and expandable in vitrousing standard cell culture techniques. Immuno-phenotypically, MSC lack the typical hematopoie-tic antigens (CD45, CD34, CD14) but expressspecific adhesion molecules (ALCAM/CD44) andantigens (SH2/SH3/SH4/STRO-1).41,42 At first,MSC were thought to contribute solely to theformation of the stromal microenvironment in thebone marrow and maintain HSC survival andfunction. However, subsequent studies havesuggested that MSC are themselves capable ofmultipotency, with differentiation into chondro-cytes, osteoblasts, astrocytes, neurons, skeletalmuscle, and, notably, cardiomyocytes.30,43-45
Endothelial Progenitor Cells
Endothelial progenitor cells represent a subset ofhematopoietic stem cells that are able to acquirean endothelial phenotype in vitro.46-49 Endothe-lial progenitor cells express the hematopoieticstem cell markers CD133 and CD34 and theendothelial marker Flk-1 (VEGFR-2).48 Endothe-lial progenitor cells can be isolated directly fromthe bone marrow or from the peripheral circula-tion and expanded in vitro.
Cardiac Cellular Homeostasis:Physiological and Pathological States
The identification of resident cardiac stem/pro-genitor cells evokes a new understanding of themechanisms by which the adult heart maymaintain cellular homeostasis. It is still a matterof debate whether cardiac cellular homeostasis ismaintained solely by endogenous stem/progenitorcells or via extracardiac sources, notably bonemarrow–derived stem cells. In particular, theobservation of male (host) cells in male patientstransplanted with female hearts (mixed-sex donorhearts)50,51 suggests the potential role of extra-cardiac stem cells in the turnover of the cardiaccells. Interestingly, it has been proposed that the
chimerism observed in human beings maypossibly result from cardiac progenitor cellsresiding in host atria that were kept intact duringcardiac transplantation, and not from circulatingbone marrow stem cells.52 More recent data inanimal models, however, have suggested thatbone marrow–derived stem cells contribute littlein maintaining the homeostasis of cardiac cellsduring normal postnatal growth as well as normaladulthood.19,38,53
In contrast, bone marrow–derived stem cellslikely play a significant role in maintainingcardiac cell homeostasis, including the turnoverof cardiac stem/progenitor cells, cardiomyo-genesis, and angiogenesis, after myocardialinjury.19,53,54 Jackson et al38 demonstrated theability of bone marrow SP cells to undergocardiomyogenic differentiation, albeit at a verylow frequency, and angiogenesis after MI. Usinga murine model of GFP-labeled bone marrow, wealso have found that bone marrow–derived stemcells (SP cells) homed to areas of injured heart asearly as 3 days after MI.19 These bone marrow–derived cells may not only contribute to activemyocardial repair, as have been suggested byseveral groups,8,9,36,37,55-59 but also participate inthe reconstitution of the cardiac progenitor cellpool.19 This is further supported by additionalrecent work,53 which has used genetic mousemodels to demonstrate an increase in cardiac c-kit+ cells, recruited from bone marrow, afterMI53 Moreover, using a rat model of heterotropicsex-mismatched cardiac transplantation, Wang etal also demonstrated that bone marrow–derivedstem cells are attracted to areas of myocardialischemic injury and participate in cardiacrepair54 These experimental data were furthersupported by observations in human mixed-sexcardiac transplants, which suggest greater car-diac chimerism may occur in patients with MI.60
In summary, the current literature suggests thatcardiac injury may serve as a necessary andpotent stimulant for the recruitment and poten-tial cardiomyogenic differentiation of endogen-ous bone marrow–derived stem cells.
Mobilization and Homing ofMarrow–Derived Stem Cell
It is well recognized that, despite the existence ofcardiac stem/progenitor cells, this endogenous
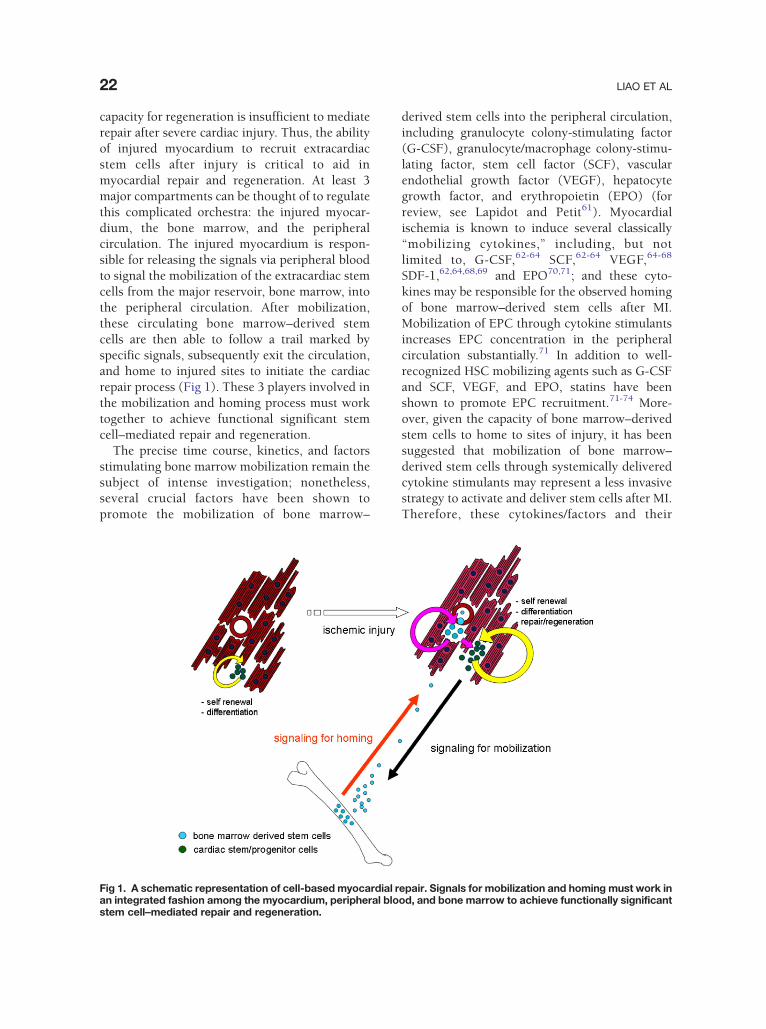
22 LIAO ET AL
capacity for regeneration is insufficient to mediaterepair after severe cardiac injury. Thus, the abilityof injured myocardium to recruit extracardiacstem cells after injury is critical to aid inmyocardial repair and regeneration. At least 3major compartments can be thought of to regulatethis complicated orchestra: the injured myocar-dium, the bone marrow, and the peripheralcirculation. The injured myocardium is respon-sible for releasing the signals via peripheral bloodto signal the mobilization of the extracardiac stemcells from the major reservoir, bone marrow, intothe peripheral circulation. After mobilization,these circulating bone marrow–derived stemcells are then able to follow a trail marked byspecific signals, subsequently exit the circulation,and home to injured sites to initiate the cardiacrepair process (Fig 1). These 3 players involved inthe mobilization and homing process must worktogether to achieve functional significant stemcell–mediated repair and regeneration.
The precise time course, kinetics, and factorsstimulating bone marrow mobilization remain thesubject of intense investigation; nonetheless,several crucial factors have been shown topromote the mobilization of bone marrow–
Fig 1. A schematic representation of cell-based myocardial ran integrated fashion among the myocardium, peripheral blostem cell–mediated repair and regeneration.
derived stem cells into the peripheral circulation,including granulocyte colony-stimulating factor(G-CSF), granulocyte/macrophage colony-stimu-lating factor, stem cell factor (SCF), vascularendothelial growth factor (VEGF), hepatocytegrowth factor, and erythropoietin (EPO) (forreview, see Lapidot and Petit61). Myocardialischemia is known to induce several classically“mobilizing cytokines,” including, but notlimited to, G-CSF,62-64 SCF,62-64 VEGF,64-68
SDF-1,62,64,68,69 and EPO70,71; and these cyto-kines may be responsible for the observed homingof bone marrow–derived stem cells after MI.Mobilization of EPC through cytokine stimulantsincreases EPC concentration in the peripheralcirculation substantially.71 In addition to well-recognized HSC mobilizing agents such as G-CSFand SCF, VEGF, and EPO, statins have beenshown to promote EPC recruitment.71-74 More-over, given the capacity of bone marrow–derivedstem cells to home to sites of injury, it has beensuggested that mobilization of bone marrow–derived stem cells through systemically deliveredcytokine stimulants may represent a less invasivestrategy to activate and deliver stem cells after MI.Therefore, these cytokines/factors and their
epair. Signals for mobilization and homing must work inod, and bone marrow to achieve functionally significan
t
23CARDIAC AXIS OF MYOCARDIAL REGENERATION
respective receptors can be targeted to promotestem cell mobilization and homing for therapeu-tics applications. Herein, we highlight several keysignaling factors to demonstrate the potential ofmanipulating these signaling axes to achievefunctionally significant cell-based cardiac repair.
G-CSF and SCF/c-kit
Stem cell factor, also known as steel factor, is aligand for c-kit, a receptor expressed in stem celland tissue progenitor cells, including residentcardiac stem cells. Similar to G-CSF andgranulocyte/macrophage colony-stimulating fac-tor, SCF is a hematopoietic factor that is wellknown to regulate proliferation, differentiation,and survival of bone marrow–derived stemcells.75,76 Orlic et al was the first to use acombined therapy of G-CSF and SCF in amurine model of MI and demonstrateda significant improvement in left ventricularremodeling, cardiac function, and animal survi-val with 5 days of treatment.37 Improvedoutcome was associated with significant bonemarrow–derived cardiomyogenesis.37 Theseresults, however, were not reproduced when G-CSF and SCF were given as a single dose at4 hours after MI to nonhuman primates.77
Although G-CSF and SCF/c-kit represent impor-tant factors for the recruitment of bone marrow–derived stem cells after MI, actual results fromvarious groups have been controversial at bestowing to the timing of cytokine administrationand the dose used as well as the actual modelsystem. Nonetheless, the best “proof of concept”approach demonstrating the importance andinvolvement of the SCF/c-kit axis in bonemarrowmobilization and cardiac repair has taken advan-tage of a transgenic mouse model overexpressingmutant c-kit (kitw/kitw-v).53 In kitw/kitw-v mice,the mobilization and homing of bone marrow–derived stem cells to the heart are markedlyimpaired after MI, despite elevated circulatinglevels of SCF. This deficiency further results inearly cardiac failure and death. Intriguingly, thisdysfunction can be rescued by bone marrowtransplantation with wild-type cells, thus restor-ing the capacity for homing by bone marrow–derived stem cells. Although certainly essential,proper manipulation of the SCF/c-kit axis forclinical benefit remains a goal of the future.
SDF-1/CXCR4
SDF-1 and its receptor CXCR4 have recently beensuggested to also be important in regulating themobilization of stem cells. Using gain and loss offunction approaches, Moore et al has demon-strated that inhibition of SDF-1/CXCR4 by neu-tralizing antibodies retards the mobilization ofstem cells,78 whereas overexpression of CXCR4augments the migration of progenitor cells.79
More recently, several groups have suggestedthat the SDF-1/CXCR4 axis may be involved inthe mobilization of bone marrow–derived stemcells and homing to injured myocardium afterMI,80,81 whereas the mechanisms regulating thismobilization remain less clear. SDF-1 may upre-gulate secondary agents, including metalloprotei-nase-9, causing the release of SCF andsubsequently the mobilization of c-kit+ cells intothe circulation.82 Alternatively, Misao and collea-gues have suggested that the beneficial effects ofG-CSF–mediated cardiac repair after MI arethrough the upregulation of SDF-1 and subse-quently the recruitment of CXCR4+ cells.83
Altogether, these reports demonstrate the complexinterplay that exists between these mobilizing/homing signals. To take advantage of thesesecreted factors/cytokines to improve post-MIcardiac repair and regeneration, thorough inves-tigation of the timing of the release and interac-tions among signaling factors is required.
Potential Mechanisms of Stem CellMediated MyocardialRepair/Regeneration
Over the past decade, many groups have used aspectrum of bone marrow–derived stem cellpopulations, including total bone marrow, HSC,MSC, and EPC, for the treatment of post-MI heartfailure in both animal models as well as in humanclinical trials. Interestingly, although only fewgroups have observed differentiation of bonemarrow–derived stem cell into cardiomyocytes,most groups have reported a beneficial effect onpost-MI remodeling. As such, these data arecertainly both encouraging, given the improve-ment in objective measures such as cardiacstructure and function, yet disappointing, as theyfail to demonstrate physiologically relevant cardiacdifferentiation. This has brought into question the
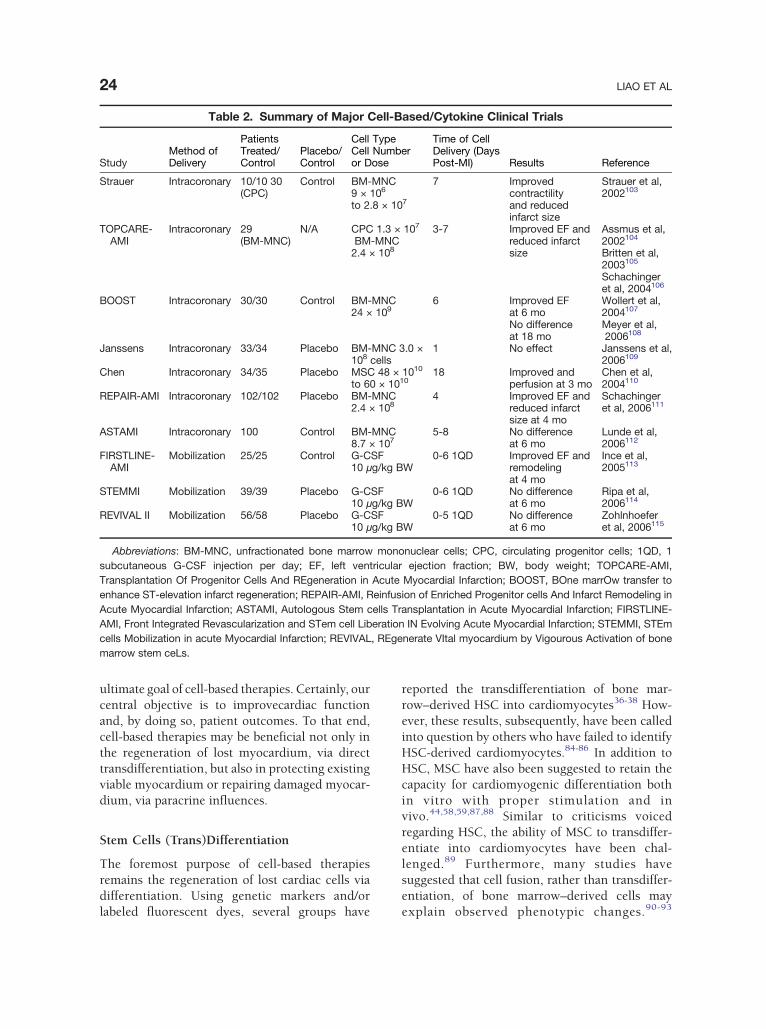
Table 2. Summary of Major Cell-Based/Cytokine Clinical Trials
StudyMethod ofDelivery
PatientsTreated/Control
Placebo/Control
Cell TypeCell Numberor Dose
Time of CellDelivery (DaysPost-MI) Results Reference
Strauer Intracoronary 10/10 30(CPC)
Control BM-MNC9 × 106
to 2.8 × 107
7 Improvedcontractilityand reducedinfarct size
Strauer et al,2002103
TOPCARE-AMI
Intracoronary 29(BM-MNC)
N/A CPC 1.3 × 107
BM-MNC2.4 × 108
3-7 Improved EF andreduced infarctsize
Assmus et al,2002104
Britten et al,2003105
Schachingeret al, 2004106
BOOST Intracoronary 30/30 Control BM-MNC24 × 109
6 Improved EFat 6 mo
Wollert et al,2004107
No differenceat 18 mo
Meyer et al,2006108
Janssens Intracoronary 33/34 Placebo BM-MNC 3.0 ×108 cells
1 No effect Janssens et al,2006109
Chen Intracoronary 34/35 Placebo MSC 48 × 1010
to 60 × 101018 Improved and
perfusion at 3 moChen et al,2004110
REPAIR-AMI Intracoronary 102/102 Placebo BM-MNC2.4 × 108
4 Improved EF andreduced infarctsize at 4 mo
Schachingeret al, 2006111
ASTAMI Intracoronary 100 Control BM-MNC8.7 × 107
5-8 No differenceat 6 mo
Lunde et al,2006112
FIRSTLINE-AMI
Mobilization 25/25 Control G-CSF10 μg/kg BW
0-6 1QD Improved EF andremodelingat 4 mo
Ince et al,2005113
STEMMI Mobilization 39/39 Placebo G-CSF10 μg/kg BW
0-6 1QD No differenceat 6 mo
Ripa et al,2006114
REVIVAL II Mobilization 56/58 Placebo G-CSF10 μg/kg BW
0-5 1QD No differenceat 6 mo
Zohlnhoeferet al, 2006115
Abbreviations: BM-MNC, unfractionated bone marrow mononuclear cells; CPC, circulating progenitor cells; 1QD, 1subcutaneous G-CSF injection per day; EF, left ventricular ejection fraction; BW, body weight; TOPCARE-AMI,Transplantation Of Progenitor Cells And REgeneration in Acute Myocardial Infarction; BOOST, BOne marrOw transfer toenhance ST-elevation infarct regeneration; REPAIR-AMI, Reinfusion of Enriched Progenitor cells And Infarct Remodeling inAcute Myocardial Infarction; ASTAMI, Autologous Stem cells Transplantation in Acute Myocardial Infarction; FIRSTLINE-AMI, Front Integrated Revascularization and STem cell Liberation IN Evolving Acute Myocardial Infarction; STEMMI, STEmcells Mobilization in acute Myocardial Infarction; REVIVAL, REgenerate VItal myocardium by Vigourous Activation of bonemarrow stem ceLs.
24 LIAO ET AL
ultimate goal of cell-based therapies. Certainly, ourcentral objective is to improvecardiac functionand, by doing so, patient outcomes. To that end,cell-based therapies may be beneficial not only inthe regeneration of lost myocardium, via directtransdifferentiation, but also in protecting existingviable myocardium or repairing damaged myocar-dium, via paracrine influences.
Stem Cells (Trans)Differentiation
The foremost purpose of cell-based therapiesremains the regeneration of lost cardiac cells viadifferentiation. Using genetic markers and/orlabeled fluorescent dyes, several groups have
reported the transdifferentiation of bone mar-row–derived HSC into cardiomyocytes36-38 How-ever, these results, subsequently, have been calledinto question by others who have failed to identifyHSC-derived cardiomyocytes.84-86 In addition toHSC, MSC have also been suggested to retain thecapacity for cardiomyogenic differentiation bothin vitro with proper stimulation and invivo.44,58,59,87,88 Similar to criticisms voicedregarding HSC, the ability of MSC to transdiffer-entiate into cardiomyocytes have been chal-lenged.89 Furthermore, many studies havesuggested that cell fusion, rather than transdiffer-entiation, of bone marrow–derived cells mayexplain observed phenotypic changes.90-93

25CARDIAC AXIS OF MYOCARDIAL REGENERATION
Regardless of the mechanism responsible, fusionvs transdifferentiation, it is generally agreed thatthe number of reported cardiomyocytes derivedfrom exogenously delivered bone marrow stemcells remains relatively low and cannot physicallyaccount for the observed functional improve-ments. As such, one alternative proposed mechan-ism is stem cell–mediated paracrine effects(discussed in the following section). In contrastto cardiomyogenesis, by and large, most groupshave observed bone marrow–derived stem cells tocontribute to angiogenesis, an observation madeover 10 years by Asahara and colleagues46,94 and,more recently, by many other laboratories.8,95-98
Finally, an alternative mechanism by which bonemarrow–derived stem cell populations may con-tribute to myocardial repair is via maintenance ofcardiac-specific stem cell pool after injury. Indeed,our group has recently found that bone marrow–derived SP cells home to injured myocardiumafter MI and undergo phenotypic changes to adapta cardiac SP cell phenotype. Such cardiac SP cellsmay in turn contribute to the capacity of the heartfor long-term endogenous cardiac repair.
Paracrine Influence
As suggested above, the beneficial effects of stemcell therapy on post-MI cardiac function remaindisproportionate to the degree of cardiomyogenicdifferentiation. Such observations have led to thehypothesis that potential paracrine effects mayhold a prominent role in stem cell therapy. Suchparacrine influences may include secretion offactors that either attenuate apoptosis of endo-genous cardiomyocytes99,100 and endothelialcells,8 promote angiogenesis,55,101 and/or activateresident cardiac stem/progenitor cells. Uemuraand colleagues suggested that hypoxia-inducedapoptosis may be attenuated in cardiomyocytescocultured with total bone marrow cells invitro.100 Moreover, MSC also may produceangiogenic factors such as VEGF and basicfibroblast growth factor, as well as chemotacticfactors, including monocyte chemoattractant pro-tein 1 and placental growth factor, that serve torecruit monocytes and promote angiogenesis.102
Although the “paracrine hypothesis” of stem celltherapy seems rational given prior observations,to date, it remains unexplored with largelyindirect supportive evidence.
Current Translational Approaches forBone Marrow–Based Therapies
Although the ideal cell type for stem cell–basedtherapies remains to be determined, to date, bonemarrow–derived stem cell, isolated from wholebone marrow aspirate, remains the most com-monly used cell type for human studies. This islargely due to its easy accessibility and well-characterized properties by hematologists formore than 30 years. Among bone marrowpopulations, total mononuclear bone marrowcells and circulating EPC have recently or arecurrently being used in many phase I and/or IItrials (Table 2). Current methods of deliveryinclude direct intramyocardial injection, via bothendocardial catheter-based and epicardial surgi-cal-based approaches and, more recently, percu-taneous (catheter-based) intracoronary injection.Alternatively, indirect mobilization has also beenattempted with peripheral delivery of cytokines,notably G-CSF.
In just a few years, cell-based therapy hasevolved at an explosive pace, from early in vitrocell studies to animal models of MI and now toseveral early-phase clinical trials. Overall, mostinitial nonrandomized clinical trials, althoughdesigned for safety rather than efficacy, haveencouragingly suggested a moderate improve-ment in heart function after stem cell therapy. Thefirst randomized trial of intracoronary bonemarrow–derived stem cells, the BOOST Itrial,107 demonstrated an early benefit in leftventricular ejection fraction at 6 months post–celltherapy as assessed by cardiac magnetic resonanceimaging. However, because of continued improve-ment in the control group, the benefit in treatedpatients relative to the control group was lost at18 months follow-up.108 Two larger clinical trialsinvestigating intracoronary delivery of bonemarrow cells have recently been published.111,112
In the ASTAMI trial, a randomized trial of 100patients112 with acute MI, bone marrow mono-nuclear cells were delivered 6 days post–percuta-nous transluminal coronary angioplasty. At6 months follow-up, no improvement in leftventricular ejection fraction or infarct size wasobserved. In contrast somewhat, the REPAIR-AMI,111 a randomized, placebo-controlled trialwith more than 200 patients after acute MI,suggested a small, albeit significantly important,

26 LIAO ET AL
improvement in left ventricular ejection fractionby ventriculography. Many potential explanationsfor observed differences, including the severity ofventricular dysfunction, the timing of cell deliv-ery, and the method of cell isolation (quality of thecells), have been proposed and are currentlyundergoing investigation in animal models.Mobilization of stem cells from the bone marrowrepresents an alternative cell-based therapy thathas also recently been investigated in clinicaltrials. The FIRSTLINE-AMI113 has demonstratednot only the safety and feasibility of bone marrowmobilization using G-CSF in MI patients afterreperfusion, but also suggested a potentialimprovement in left ventricular ejection fractionand an attenuation of left ventricular dilation.Importantly, this trial showed that treatment withG-CSF did not augment the post–percutaneouscoronary intervention restenosis rate. However,subsequent randomized, placebo-controlled clin-ical trials REVIVAL II115 and STEMMI114 havefailed to reproduce the benefits previously seen inearly human studies. Although these trials failedto demonstrate positive outcomes, no adverseevents, including vessel restenosis, were observed.The reasons for these negative results remain to bedetermined, although inappropriate cytokinedosing and inadequate timing of the cytokineadministration have been proposed as potentialexplanations.
Conclusions
Myocardial infarction results in cell death andreplacement of cardiomyocytes with noncontrac-tile scar tissue. The optimal goal for cell-basedcardiac repair is to restore cardiac structure andfunction through regeneration of functionallycompetent cardiomyocytes. To rebuild a normaland functional cardiac tissue requires not onlyhighly integrated cardiomyogenesis and angio-genesis, but also a proper matrix network systemto ensure synchronized contraction and relaxa-tion with native myocardium. Although the taskof achieving such goal is daunting, the ther-apeutic potential of myocardial regenerationremains enormous. Further intensive research,in basic science and clinical arenas, and carefullyconstructed clinical trials are needed to makethis next great frontier in cardiovascular medi-cine a reality.
Acknowledgments
This work was supported by research grantsfunding from the National Institutes of Health(RL). OP is a recipient of a Swiss NationalScience Foundation Grant (PBBSB-101349), andFM is a recipient of a Federation Francaise deCardiologie grant.
References1. Braunwald E, Bristow MR: Congestive heart failure:
fifty years of progress. Circulation 102:IV14-IV23,2000
2. Scorsin M, et al: Comparison of the effects offetal cardiomyocyte and skeletal myoblast trans-plantation on postinfarction left ventricular func-tion. J Thorac Cardiovasc Surg 119:1169-1175,2000
3. Koh GY, et al: Stable fetal cardiomyocyte grafts inthe hearts of dystrophic mice and dogs. J Clin Invest96:2034-2042, 1995
4. Jain M, et al: Cell therapy attenuates deleteriousventricular remodeling and improves cardiac perfor-mance after myocardial infarction. Circulation103:1920-1927, 2001
5. Taylor DA, et al: Regenerating functional myocar-dium: improved performance after skeletal myoblasttransplantation. Nat Med 4:929-933, 1998
6. Murry CE, Wiseman RW, Schwartz SM, et al: Skeletalmyoblast transplantation for repair of myocardialnecrosis. J Clin Invest 98:2512-2523, 1996
7. Chiu RC, Zibaitis A, Kao RL: Cellular cardiomyo-plasty: myocardial regeneration with satellite cellimplantation. Ann Thorac Surg 60:12-18, 1995
8. Kocher AA, et al: Neovascularization of ischemicmyocardium by human bone-marrow–derived angio-blasts prevents cardiomyocyte apoptosis, reducesremodeling and improves cardiac function. Nat Med7:430-436, 2001
9. Kawamoto A, et al: Intramyocardial transplantation ofautologous endothelial progenitor cells for therapeu-tic neovascularization of myocardial ischemia. Cir-culation 107:461-468, 2003
10. Orlic D: Adult BM stem cells regenerate mousemyocardium. Cytotherapy 4:521-525, 2002
11. Schachinger V, et al: Intracoronary bone marrow-derived progenitor cells in acute myocardial infarc-tion. N Engl J Med 355:1210-1221, 2006
12. Lunde K, et al: Intracoronary injection of mono-nuclear bone marrow cells in acute myocardialinfarction. N Engl J Med 355:1199-1209, 2006
13. Martin CM, et al: Persistent expression of the ATP-binding cassette transporter, Abcg2, identifies car-diac SP cells in the developing and adult heart. DevBiol 265:262-275, 2004
14. Messina E, et al: Isolation and expansion of adultcardiac stem cells from human and murine heart.Circ Res 95:911-921, 2004

27CARDIAC AXIS OF MYOCARDIAL REGENERATION
15. Pfister O, et al: CD31− but not CD31+ cardiac sidepopulation cells exhibit functional cardiomyogenicdifferentiation. Circ Res 97:52-61, 2005
16. McDonnell TJ, Oberpriller JO: The response of theatrium to direct mechanical wounding in the adultheart of the newt, Notophthalmus viridescens. Anelectron-microscopic and autoradiographic study.Cell Tissue Res 235:583-592, 1984
17. Rumyantsev PP, Borisov A: DNA synthesis in myo-cytes from different myocardial compartments ofyoung rats in norm, after experimental infarction andin vitro. Biomed Biochim Acta 46:S610-S615, 1987
18. Hierlihy AM, Seale P, Lobe CG, et al: The post-natalheart contains a myocardial stem cell population.FEBS Lett 530:239-243, 2002
19. Mouquet F, et al: Restoration of cardiac progenitorcells after myocardial infarction by self-proliferationand selective homing of bone marrow–derived stemcells. Circ Res 97:1090-1092, 2005
20. Urbanek K, et al: Intense myocyte formation fromcardiac stem cells in human cardiac hypertrophy.Proc Natl Acad Sci U S A 100:10440-10445,2003
21. Linke A, et al: Stem cells in the dog heart are self-renewing, clonogenic, and multipotent and re-generate infarcted myocardium, improving cardiacfunction. Proc Natl Acad Sci U S A 102:8966-8971, 2005
22. Oh H, et al: Cardiac progenitor cells from adultmyocardium: homing, differentiation, and fusion afterinfarction. Proc Natl Acad Sci U S A 100:12313-12318, 2003
23. Matsuura K, et al: Adult cardiac Sca-1–positive cellsdifferentiate into beating cardiomyocytes. J BiolChem 279:11384-11391, 2004
24. Laugwitz KL, et al: Postnatal isl1+ cardioblasts enterfully differentiated cardiomyocyte lineages. Nature433:647-653, 2005
25. Goodell MA, Brose K, Paradis G, et al: Isolation andfunctional properties of murine hematopoietic stemcells that are replicating in vivo. J Exp Med183:1797-1806, 1996
26. Asakura A, Rudnicki MA: Side population cells fromdiverse adult tissues are capable of in vitro hemato-poietic differentiation. Exp Hematol 30:1339-1345,2002
27. Challen GA, Little MH: A side order of stem cells: theSP phenotype. Stem Cells 24:3-12, 2006
28. Tomita Y, et al: Cardiac neural crest cells contributeto the dormant multipotent stem cell in the mamma-lian heart. J Cell Biol 170:1135-1146, 2005
29. Wada H, Matsuura K, Oyama T, et al: Homing anddifferentiation of intravenously transplanted cardiacside population cells in injured myocardium. Circula-tion 111:801, 2005
30. Ferrari G, et al: Muscle regeneration by bonemarrow–derived myogenic progenitors. Science279:1528-1530, 1998
31. Krause DS, et al: Multi-organ, multi-lineage engraft-ment by a single bone marrow–derived stem cell.Cell 105:369-377, 2001
32. Mezey E, Chandross KJ, Harta G, et al: Turning bloodinto brain: cells bearing neuronal antigens generatedin vivo from bone marrow. Science 290:1779-1782,2000
33. Osawa M, et al: In vivo self-renewal of c-Kit+ Sca-1+Lin(low/−) hemopoietic stem cells. J Immunol156:3207-3214, 1996
34. Spangrude GJ, Heimfeld S, Weissman IL: Purifica-tion and characterization of mouse hematopoieticstem cells. Science 241:58-62, 1988
35. Goldman JM, Horowitz MM: The international bonemarrow transplant registry. Int J Hematol 76:393-397, 2002 (Suppl 1)
36. Orlic D, et al: Bone marrow cells regenerate infarctedmyocardium. Nature 410:701-705, 2001
37. Orlic D, et al: Mobilized bone marrow cells repair theinfarcted heart, improving function and survival. ProcNatl Acad Sci U S A 98:10344-10349, 2001
38. Jackson KA, et al: Regeneration of ischemic cardiacmuscle and vascular endothelium by adult stemcells. J Clin Invest 107:1395-1402, 2001
39. Friedenstein AJ, Petrakova KV, Kurolesova AI, et al:Heterotopic of bone marrow. Analysis of precursorcells for osteogenic and hematopoietic tissues.Transplantation 6:230-247, 1968
40. Tavassoli M, Crosby WH: Transplantation of marrowto extramedullary sites. Science 161:54-56, 1968
41. Conget PA, Minguell JJ: Phenotypical and functionalproperties of human bone marrow mesenchymalprogenitor cells. J Cell Physiol 181:67-73, 1999
42. PittengerMF, et al:Multilineagepotential of adult humanmesenchymal stem cells. Science 284:143-147, 1999
43. Azizi SA, Stokes D, Augelli BJ, et al: Engraftment andmigration of human bone marrow stromal cellsimplanted in the brains of albino rats—similaritiesto astrocyte grafts. Proc Natl Acad Sci U S A95:3908-3913, 1998
44. Makino S, et al: Cardiomyocytes can be generatedfrom marrow stromal cells in vitro. J Clin Invest103:697-705, 1999
45. Pereira RF, et al: Cultured adherent cells frommarrow can serve as long-lasting precursor cellsfor bone, cartilage, and lung in irradiated mice. ProcNatl Acad Sci U S A 92:4857-4861, 1995
46. Asahara T, et al: Isolation of putative progenitorendothelial cells for angiogenesis. Science 275:964-967, 1997
47. Gehling UM, et al: In vitro differentiation of endothe-lial cells from AC133-positive progenitor cells. Blood95:3106-3112, 2000
48. Peichev M, et al: Expression of VEGFR-2 and AC133by circulating human CD34(+) cells identifies apopulation of functional endothelial precursors.Blood 95:952-958, 2000
49. Shi Q, et al: Evidence for circulating bone marrow–derived endothelial cells. Blood 92:362-367, 1998
50. Quaini F, et al: Chimerism of the transplanted heart.N Engl J Med 346:5-15
51. Murry CE, Whitney ML, Laflamme MA, et al: Cellulartherapies for myocardial infarct repair. Cold SpringHarb Symp Quant Biol 67:519-526, 2002

28 LIAO ET AL
52. Schwartz RS, Curfman GD: Can the heart repairitself? N Engl J Med 346:2-4, 2002
53. Fazel S, et al: Cardioprotective c-kit+ cells are fromthe bone marrow and regulate the myocardialbalance of angiogenic cytokines. J Clin Invest116:1865-1877, 2006
54. Wang Y, et al: Combining pharmacological mobiliza-tion with intramyocardial delivery of bone marrowcells over-expressing VEGF is more effective forcardiac repair. J Mol Cell Cardiol 40:736-745, 2006
55. Kamihata H, et al: Implantation of bone marrowmononuclear cells into ischemic myocardiumenhances collateral perfusion and regional functionvia side supply of angioblasts, angiogenic ligands,and cytokines. Circulation 104:1046-1052, 2001
56. Kawamoto A, et al: Therapeutic potential of ex vivoexpanded endothelial progenitor cells for myocardialischemia. Circulation 103:634-637, 2001
57. Mangi AA, et al: Mesenchymal stem cells modifiedwith Akt prevent remodeling and restore perfor-mance of infarcted hearts. Nat Med 9:1195-1201,2003
58. Shake JG, et al: Mesenchymal stem cell implantationin a swine myocardial infarct model: engraftment andfunctional effects. Ann Thorac Surg 73:1919-1925,2002 [discussion 1926]
59. Toma C, Pittenger MF, Cahill KS, et al: Humanmesenchymal stem cells differentiate to a cardio-myocyte phenotype in the adult murine heart.Circulation 105:93-98, 2002
60. Hocht-Zeisberg E, et al: Cellular repopulation ofmyocardial infarction in patients with sex-mis-matched heart transplantation. Eur Heart J25:749-758, 2004
61. Lapidot T, Petit I: Current understanding of stem cellmobilization: the roles of chemokines, proteolyticenzymes, adhesion molecules, cytokines, and stro-mal cells. Exp Hematol 30:973-981, 2002
62. Leone AM, et al: Endogenous G-CSF and CD34(+)cell mobilization after acute myocardial infarction. IntJ Cardiol 111:202-208, 2006
63. Dewald O, et al: Of mice and dogs: species-specificdifferences in the inflammatory response followingmyocardial infarction. Am J Pathol 164:665-677, 2004
64. Wojakowski W, et al: Mobilization of CD34/CXCR4+,CD34/CD117+, c-met+ stem cells, and mononuclearcells expressing early cardiac, muscle, and endothe-lial markers into peripheral blood in patients withacute myocardial infarction. Circulation 110:3213-3220, 2004
65. Namiki A, et al: Hypoxia induces vascular endothelialgrowth factor in cultured human endothelial cells.J Biol Chem 270:31189-31195, 1995
66. Couffinhal T, et al: Mousemodel of angiogenesis. AmJ Pathol 152:1667-1679, 1998
67. Banai S, et al: Angiogenic-induced enhancement ofcollateral blood flow to ischemic myocardium byvascular endothelial growth factor in dogs. Circula-tion 89:2183-2189, 1994
68. Wang Y, et al: Changes in circulating mesenchymalstem cells, stem cell homing factor, and vascular
growth factors in patients with acute ST elevationmyocardial infarction treated with primary percuta-neous coronary intervention. Heart 92:768-774, 2006
69. Askari AT, et al: Effect of stromal-cell–derived factor1 on stem-cell homing and tissue regeneration inischaemic cardiomyopathy. Lancet 362:697-703,2003
70. Vogt C, Noe G, Rich IN: The role of the blood islandduring normal and 5-fluorouracil–perturbed hemo-poiesis. Blood Cells 17:105-121, 1991 [discussion121-5]
71. Heeschen C, et al: Erythropoietin is a potentphysiologic stimulus for endothelial progenitor cellmobilization. Blood 102:1340-1346, 2003
72. Kalka C, et al: Vascular endothelial growth factor(165) gene transfer augments circulating endothelialprogenitor cells in human subjects. Circ Res86:1198-1202, 2000
73. Llevadot J, et al: HMG-CoA reductase inhibitormobilizes bone marrow–derived endothelial progeni-tor cells. J Clin Invest 108:399-405, 2001
74. Walter DH, et al: Statin therapy accelerates reen-dothelialization: a novel effect involving mobilizationand incorporation of bone marrow–derived endothe-lial progenitor cells. Circulation 105:3017-3024, 2002
75. Ulich TR, et al: Hematologic effects of stem cellfactor in vivo and in vitro in rodents. Blood78:645-650, 1991
76. Anderson DM, et al: Molecular cloning of mast cellgrowth factor, a hematopoietin that is active in bothmembrane bound and soluble forms. Cell63:235-243, 1990
77. Norol F, et al: Influence of mobilized stem cells onmyocardial infarct repair in a nonhuman primatemodel. Blood 102:4361-4368, 2003
78. Petit I, et al: G-CSF induces stem cell mobilization bydecreasing bone marrow SDF-1 and up-regulatingCXCR4. Nat Immunol 3:687-694, 2002
79. Kahn J, et al: Overexpression of CXCR4 on humanCD34+ progenitors increases their proliferation,migration, and NOD/SCID repopulation. Blood103:2942-2949, 2004
80. KuciaM, et al: Cells expressing early cardiac markersreside in the bone marrow and are mobilized into theperipheral blood after myocardial infarction. Circ Res95:1191-1199, 2004
81. Abbott JD, et al: Stromal cell-derived factor-1alphaplays a critical role in stem cell recruitment to theheart after myocardial infarction but is not sufficientto induce homing in the absence of injury. Circulation110:3300-3305, 2004
82. Heissig B, et al: Recruitment of stem and progenitorcells from the bone marrow niche requires MMP-9mediated releaseof kit-ligand.Cell 109:625-637, 2002
83. Misao Y, et al: Importance of recruitment of bonemarrow–derived CXCR4(+) cells in post-infarct car-diac repair mediated by G-CSF. Cardiovasc Res71:455-465, 2006
84. Murry CE, et al: Haematopoietic stem cells do nottransdifferentiate into cardiac myocytes in myocar-dial infarcts. Nature 428:664-668, 2004

29CARDIAC AXIS OF MYOCARDIAL REGENERATION
85. Balsam LB, et al: Haematopoietic stem cells adoptmature haematopoietic fates in ischaemic myocar-dium. Nature 428:668-673, 2004
86. Limbourg FP, et al: Haematopoietic stem cellsimprove cardiac function after infarction withoutpermanent cardiac engraftment. Eur J Heart Fail7:722-729, 2005
87. Tomita S, et al: Autologous transplantation of bonemarrow cells improves damaged heart function.Circulation 100:II247-II256, 1999
88. Hakuno D, et al: Bone marrow–derived regeneratedcardiomyocytes (CMG Cells) express functionaladrenergic and muscarinic receptors. Circulation105:380-386, 2002
89. Davani S, et al: Mesenchymal progenitor cellsdifferentiate into an endothelial phenotype, enhancevascular density, and improve heart function in a ratcellular cardiomyoplasty model. Circulation 108:II253-II258, 2003 (Suppl 1)
90. Terada N, et al: Bone marrow cells adopt thephenotype of other cells by spontaneous cell fusion.Nature 416:542-545, 2002
91. Ying QL, Nichols J, Evans EP, et al: Changingpotency by spontaneous fusion. Nature 416:545-548, 2002
92. Alvarez-Dolado M, et al: Fusion of bone-marrow–derived cells with Purkinje neurons, cardiomyocytesand hepatocytes. Nature 425:968-973, 2003
93. Vassilopoulos G, Russell DW: Cell fusion: an alter-native to stem cell plasticity and its therapeuticimplications. Curr Opin Genet Dev 13:480-485, 2003
94. Asahara T, et al: Bone marrow origin of endothelialprogenitor cells responsible for postnatal vasculo-genesis in physiological and pathological neovascu-larization. Circ Res 85:221-228, 1999
95. Urbich C, et al: Relevance of monocytic features forneovascularization capacity of circulating endothelialprogenitor cells. Circulation 108:2511-2516, 2003
96. Camargo FD, Green R, Capetanaki Y, et al:Single hematopoietic stem cells generate skeletalmuscle through myeloid intermediates. Nat Med 9:1520-1527, 2003
97. Zhao Y, Glesne D, Huberman E: A human peripheralblood monocyte–derived subset acts as pluripotentstem cells. Proc Natl Acad Sci U S A 100:2426-2431,2003
98. Wang JS, Shum-Tim D, Chedrawy E, et al: Thecoronary delivery of marrow stromal cells formyocardial regeneration: pathophysiologic and ther-apeutic implications. J Thorac Cardiovasc Surg122:699-705, 2001
99. Takahashi T, et al: Ischemia- and cytokine-inducedmobilization of bone marrow–derived endothelialprogenitor cells for neovascularization. Nat Med5:434-438, 1999
100. Uemura R, Xu M, Ahmad N, et al: Bone marrow stemcells prevent left ventricular remodeling of ischemicheart through paracrine signaling. Circ Res98:1414-1421, 2006
101. Fuchs S, et al: Transendocardial delivery of auto-logous bone marrow enhances collateral perfusion
and regional function in pigs with chronic experi-mental myocardial ischemia. J Am Coll Cardiol37:1726-1732, 2001
102. Kinnaird T, et al: Marrow-derived stromal cellsexpress genes encoding a broad spectrum ofarteriogenic cytokines and promote in vitro and invivo arteriogenesis through paracrine mechanisms.Circ Res 94:678-685, 2004
103. Strauer BE, et al: Repair of infarcted myocardium byautologous intracoronary mononuclear bone marrowcell transplantation in humans. Circulation106:1913-1918, 2002
104. Assmus B, et al: Transplantation of progenitor cellsand regeneration enhancement in acute myocardialinfarction (TOPCARE-AMI). Circulation 106:3009-3017, 2002
105. Britten MB, et al: Infarct remodeling after intracor-onary progenitor cell treatment in patients with acutemyocardial infarction (TOPCARE-AMI): mechanisticinsights from serial contrast-enhanced magneticresonance imaging. Circulation 108:2212-2218, 2003
106. Schachinger V, et al: Transplantation of progenitorcells and regeneration enhancement in acute myo-cardial infarction: final one-year results of theTOPCARE-AMI Tr ia l . J Am Col l Card io l44:1690-1699, 2004
107. Wollert KC, et al: Intracoronary autologous bone-marrow cell transfer after myocardial infarction: theBOOST randomised controlled clinical trial. Lancet364:141-148, 2004
108. Meyer GP, et al: Intracoronary bone marrow celltransfer after myocardial infarction: eighteen months'follow-up data from the randomized, controlledBOOST (BOne marrOw transfer to enhance ST-elevation infarct regeneration) trial. Circulation113:1287-1294, 2006
109. Janssens S, et al: Autologous bone marrow–derivedstem-cell transfer in patients with ST-segmentelevation myocardial infarction: double-blind, rando-mised controlled trial. Lancet 367:113-121, 2006
110. Chen SL, et al: Improvement of cardiac function aftertransplantation of autologous bone marrowmesenchymal stem cells in patients with acutemyocardial infarction. Chin Med J (Engl) 117:1443-1448, 2004
111. Schachinger V, et al: Intracoronary bone marrow-derived progenitor cells in acute myocardial infarc-tion. N Engl J Med 355:1210-1221, 2006
112. Lunde K, et al: Intracoronary injection of mono-nuclear bone marrow cells in acute myocardialinfarction. N Engl J Med 355:1199-1209, 2006
113. Ince H, et al: Prevention of left ventricular remodelingwith granulocyte colony-stimulating factor afteracute myocardial infarction: final 1-year results ofthe Front-Integrated Revascularization and StemCell Liberation in Evolving Acute Myocardial Infarc-tion by Granulocyte Colony-Stimulating Factor(FIRSTLINE-AMI) Trial. Circulation 112:I73-I80, 2005
114. Ripa RS, et al: Stem cell mobilization induced bysubcutaneous granulocyte-colony stimulating factorto improve cardiac regeneration after acute ST-

30 LIAO ET AL
elevation myocardial infarction: result of the double-blind, randomized, placebo-controlled stem cells inmyocardial infarction (STEMMI) trial. Circulation113:1983-1992, 2006
115. Zohlnhofer D, et al: Stem cell mobilization bygranulocyte colony-stimulating factor in patientswith acute myocardial infarction: a randomizedcontrolled trial. JAMA 295:1003-1010, 2006




















