
The body fat-lowering effect of conjugated linoleic acid: a comparison between animal and human...
7
J. Physiol. Biochem., 61 (2), 363-370, 2005 Body fat-lowering effect of conjugated linoleic acid is not due to increased lipolysis E. Sim£ M. T. Macarulla, A. Fern• V. M. RodrŸ and M. P. Portillo Department of Nutrition and Food Science, University of PaŸ Vasco, Paseo de la Universidad 7, 01006 Vitoria, Spain (Received on November 2, 2004) E. SIM‰ M. T. MACARULLA, A. FERN93 V. M. RO- DR… and M. P. PORTILLO. Body fat-lowering effect of conjugated linoleic acid is not due to increased lipolysis. J. Physiol. Biochem., 61 (2), 363-370, 2005. The ability of conjugated linoleic acid (CLA) to reduce adiposity may be due to changes in energy expenditure and/or direct effects on adipocyte lipid metabolism. The aim of the present work was to analyse if CLA supplementation modifies lipolytic activity in adipose tissue from hamsters fed on high-fat diet. Hamsters were divided into two groups and fed on diets supplemented with either 0.5 % linoleic acid (control) or 0.5% trans-lO,cis-12 CLA. After 6 weeks, animals were fasted overnight and adipose tissues were dissected and weighed. Adipocytes were isolated by colla- genase digestion and incubated in Krebs-Ringer bicarbonate buffer with or without several agents acting aL different levels of the lipolytic cascade. Adipocyte diameters were measured by microscopy. Adipose tissue DNA content was assessed by spec- trophotometry. Animals fed on CLA diet showed significantly reduced adipose tis- sue mass. No differences between both groups was found for basal lipolysis, lipoly- tic effects of isoproterenol, forskolin, dibutyryl-cAMP and isobutylmethylxanthine, and pD2 for isoproterenol. A similar total DNA amount was found in adipose tissue of both groups, showing that CLA diet had no effect on total cell number per fat pad. Although DNA content per gram tissue, ah indirect reverse index of cell size, was significantly increased in CLA fed hamsters, microscopy did not reveal differ- ences in medium mature adipocyte diameter, nor in cell size distribution between both groups. These results suggest that adipose tissue size reduction induced by trans-lO,cis-12 CLA intake is not due to changes in lipolysis. Reduced preadipocyte differentiation into mature adipocytes may account for this fat-lowering effect. Key words" Conjugated linoleic acid, Adipose tissue, Lipolysis, Hamster. Correspondence to M. P. Portillo (Tel.: +34 945 013 067; Fax: +34 945 013 014; e-mail: [email protected]).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The body fat-lowering effect of conjugated linoleic acid: a comparison between animal and human...

J. Physiol. Biochem., 61 (2), 363-370, 2005
Body fat-lowering effect of conjugated linoleic acid is not due to increased lipolysis
E. Sim£ M. T. Macarulla, A. Fern• V. M. RodrŸ and M. P. Portillo
Department of Nutrition and Food Science, University of PaŸ Vasco, Paseo de la Universidad 7, 01006 Vitoria, Spain
(Received on November 2, 2004)
E. SIM‰ M. T. MACARULLA, A. F E R N � 9 3 V. M. RO- DR… and M. P. PORTILLO. Body fat-lowering effect of conjugated linoleic acid is not due to increased lipolysis. J. Physiol. Biochem., 61 (2), 363-370, 2005.
The ability of conjugated linoleic acid (CLA) to reduce adiposity may be due to changes in energy expenditure and/or direct effects on adipocyte lipid metabolism. The aim of the present work was to analyse if CLA supplementation modifies lipolytic activity in adipose tissue from hamsters fed on high-fat diet. Hamsters were divided into two groups and fed on diets supplemented with either 0.5 % linoleic acid (control) or 0.5% trans-lO,cis-12 CLA. After 6 weeks, animals were fasted overnight and adipose tissues were dissected and weighed. Adipocytes were isolated by colla- genase digestion and incubated in Krebs-Ringer bicarbonate buffer with or without several agents acting aL different levels of the lipolytic cascade. Adipocyte diameters were measured by microscopy. Adipose tissue D N A content was assessed by spec- trophotometry. Animals fed on CLA diet showed significantly reduced adipose tis- sue mass. No differences between both groups was found for basal lipolysis, lipoly- tic effects of isoproterenol, forskolin, dibutyryl-cAMP and isobutylmethylxanthine, and pD2 for isoproterenol. A similar total D N A amount was found in adipose tissue of both groups, showing that CLA diet had no effect on total cell number per fat pad. Although D N A content per gram tissue, ah indirect reverse index of cell size, was significantly increased in CLA fed hamsters, microscopy did not reveal differ- ences in medium mature adipocyte diameter, nor in cell size distribution between both groups. These results suggest that adipose tissue size reduction induced by trans-lO,cis-12 CLA intake is not due to changes in lipolysis. Reduced preadipocyte differentiation into mature adipocytes may account for this fat-lowering effect.
Key words" Conjugated linoleic acid, Adipose tissue, Lipolysis, Hamster.
Correspondence to M. P. Portillo (Tel.: +34 945 013 067; Fax: +34 945 013 014; e-mail: [email protected]).

364 E. SIM‰ M. T. MACARULLA, A. FERNANDEZ-QUINTELA et aL
Conjugated linoleic acid (CLA) is a term used to describe a group of geomet- ric and positional isomers of linoleic acid. CLA has been shown to have a variety of beneficial effects, such as a reduction in body fat. This effect has been observed in several experimental animal models, including mice, rats, hamsters, chickens and pigs (29). Some studies have found this effect in humans (13, 27). There is now strong evidence showing that trans- lO,cis-12 CLA is the active isomer in elic- iting body fat reduction (17, 19, 21).
The ability of CLA to reduce adiposity can revolve around pathways that regulate energy expenditure (20, 26, 30-32). More- over, direct effects on adipocyte triacyl- glycerol metabolism should also be con- sidered. With regard to this second possi- bility, the mechanisms by which CLA exerts its fat-lowering effect have not been completely elucidated. Several potential mechanisms include: decreased lipopro- tein lipase (5, 14, 22, 28, 34) and stearoyl CoA desaturase activities (4, 25), and increased oxidation of free fatty acids (15, 16, 22). The potential contribution of increased lipolysis has been hardly studied and conflicting results have been pub- lished (9, 22, 34).
The airo of the present work was to analyse the effects of trans-10,cis-12 CLA intake on adipose tissue cellularity, and also to determine ir changes in lipolytic activity are involved in the fat-lowering effect of this CLA isomer, in hamsters fed on a high-fat diet.
Material and Methods
Animals, diets and experimental design.- The experiment was conducted with 9-week-old, male Syrian Golden hamsters (98 • 1 g) purchased from Har-
lan Ib› (Barcelona, Spain) and was in accordance with the institution's guide for the care and use of laboratory animals. The hamsters were individually housed in polycarbonate metabolic cages (Techni- plast Gazzada, Guguggiate, Italy) and placed in an air-conditioned room (22 • 2 ~ with a 12 hours day-night rhythm. After a 6-d adaptation period, hamsters were randomly divided into two groups of 8 animals each and fed on semi-purified atherogenic diets consisting of 200 g/kg casein (Sigma, St. Louis, MO, USA), 4 g/kg L-methionine (Sigma, St. Louis, MO, USA), 200 g/kg wheat starch (Ven- casser, Bilbao, Spain), 404 g/kg sucrose (local market), 100 g/kg palm oil (Agra, Leioa, Spain), 30 g/kg cellulose (Vencass- er, Bilbao, Spain), 1 g/kg cholesterol (Sigma, St. Louis, MO, USA), 4 g/kg choline'HC1 (Sigma, St. Louis, MO, USA), and supplemented with 0.5% linoleic acid (control group), or trans- 10,cis-12 CLA (Natural Lipids Ltd., Hov- debygda, Norway). Vitamin (11 g/kg) and mineral (40 g/kg) mixes were formulated according to AIN-93 guidelines (23) and supplied by ICN Pharmaceuticals (Costa Mesa, CA, USA). The experimental diets were freshly prepared once a week, gassed with nitrogen and stored at 0-4 ~ to avoid rancidity. All animals had free access to food and water. Food intake and body weight were measured daily.
At the end of the experimental period hamsters were fasted 12 h and sacrificed by cardiac puncture. Adipose tissue from subcutaneous and epididymal locations were dissected and weighed.
Adipocyte isolation and lipolysis.- Iso- lated fat cells were obtained from epididy- mal adipose tissue by collagenase diges- tion (1 mg/roL, 37 ~ in Krebs-Ringer bicarbonate buffer, containing 3.5 g/100
J. PhysioI. Biochem., 61 (2), 2005

CONJUGATED LINOLEIC ACID AND LIPOLYSIS 365
mL of bovine serum albumin (BSA V) and 5 mM of glucose at pH 7.4 (KRBA), under continuous vigorous shaking (90 cycles/min) according to the Rodbell method (24). Adipocytes were filtered through nylon mesh and washed three times with the same incubation buffer (KRBA) to eliminate the stroma-vascular fraction and collagenase. Measurements of lipolytic activity were performed by incu- bating isolated adipocytes in 200 q of KRBA buffer with continuous gentle shaking (30 cycles/min), at 37 ~ with several drugs acting at different levels of the lipolytic cascade: isoproterenol (10 -12 to 10 -4 M), forskolin (10 -5 M), dibutyryl- cAMP (10 -3 M) or isobutylmethylxan- thine (10 -5 M), or without drugs (basal lipolysis), for 90 min. The reaction was stopped with an ice bath and an aliquot (50 q was taken for enzymatic determi- nation (33) of glycerol released in the incubation buffer, which was used as the index of fat cell lipolysis. Total lipids were determined gravimetrically after their extraction, according to the method of Dole and Meinertz (7).
Concentration-response curves for iso- proterenol were used to determine the ECs0 value (concentration of agonist causing 50% of its maximal effect). ECs0 values were used to calculate pD2 (-log ECs0).
Lipolytic drugs were supplied by Sigma (St. Louis, MO, USA), and nicotinamide-adenine-dinucleotide oxi- dized (NAD+), adenosine-5-triphosphate (ATP), glycerol-kinase (GK), glycerol-3- phosphate-dehydrogenase (GDH) and collagenase A by Roche Diagnostics (Mannheim, Germany).
Determination of adipocyte size.- Adipocyte diameters were determined by direct microscopy using a 50 iaL aliquot of
each isolated fat cell suspension. Cells (200 in each sample) with diameters from 20 to 240 lam were counted.
The quantification of DNA content was carried out in subcutaneous adipose tissue as described previously (18), with a minor modification consisting in the removal of the fat component by washing the tissue homogenate twice with chloro- f o r m .
Statistical analysis.-Results are present- ed as means + SEM. Statistical analysis was performed using SPSS 8.0 (SPSS Inc. Chicago, IL, USA). Data were analysed by Student's t test. Significance was assessed at the P<0.05 level.
Results
Food intake, body weight and adipose tissue weights.- Animals fed trans-10,cis- 12 CLA showed significantly reduced final body weight (-7%), food intake (- 8%), food efficiency (-21%) and adipose tissue sizes (-24% and -44% for epididy- mal and subcutaneous depots, respective- ly) (Table I).
Lipolytic activity.- There were no dif- ferences in adipocyte basal lipolysis between both experimental groups. Lipolytic drugs induced similar effects in epididymal adipose tissue from control and trans-lO,cis-12 CLA fed animals. Moreover, the isoproterenol concentra- tion needed to produce 50% of maximal effect (pD2) was also similar in both groups (Table II).
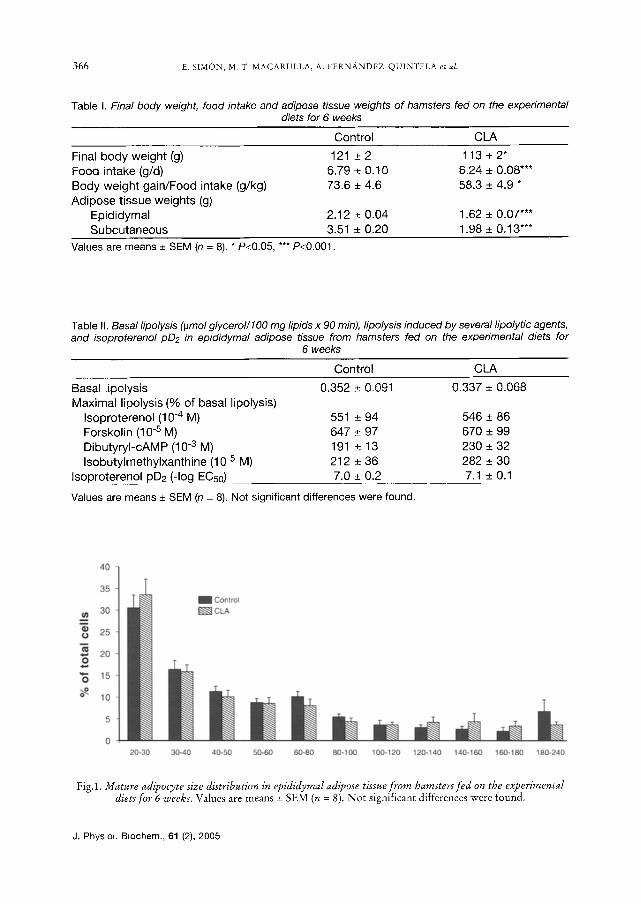
Adipocyte size and DNA quantifica- tion.- Microscopy revealed no differences neither in mature adipocyte diameter (55 -+ 3 lam in CLA group vs 58 -+ 4 tam in con- trol group), nor in cell size distribution
J. Physiol. Biochem., 61 (2), 2005
366 E. SIM‰ M. T. MACARULLA, A. FERN�93 et al.
Table I. Final body weight, food intake and adipose tissue weights of hamsters fed on the experimental diets for 6 weeks
Control CLA
Final body weight (g) 121 + 2 113 + 2* Food intake (g/d) 6.79 + 0.10 6.24 + 0.08"** Body weight gain/Food intake (g/kg) 73.6 + 4.6 58.3 _+ 4.9 * Adipose t issue weights (g)
Epididymal 2.12 + 0.04 1.62 + 0.07*** Subcutaneous 3.51 _+ 0.20 1.98 + 0.13"**
Values are means + SEM (n = 8). * P<0.05, *** P<0.001.
Table II. Basal lipolysis (#mol glycerol/lO0 mg lipids x 90 min), lipolysis induced by several lipolytic agents, and isoproterenol pD2 in epididymal adipose tissue from hamsters red on the experimental diets for
6 weeks
Control CLA
0.352 + 0.091 0.337 _+ 0.068 Basa[ lipo[ysis Maximal l ipolysis (% of basal lipolysis)
Isoproterenol (10 -4 M) Forskolin (10 -5 M) Dibutyryl-cAMP (10 -3 M) Isobutylmethylxanthine (10 -5 M)
Isoproterenol pD2 (-Iog EC50)
551 _+ 94 546 + 86 647 _+ 97 670 _+ 99 191 -+ 13 230 _ 32 212 + 36 282 _+ 30 7.0 _+ 0.2 7.1 _+ 0.1
Values are means + SEM (n = 8). Not significant differences were found.
40]~ 35
m Control _~ 3o
-~ 25
"6 15
~ 1o
5
o 20-30 30-40 40-50 50-60 60-80 8 0 - 1 0 0 100-120 120-140 140-160 160-180 180-240
Adipocyte diameter (#m)
Fig.1. Mature adipocyte size distribution in epididymal adipose tissue from hamsters fed on the experimental dietsfor 6 weeks. Values are means _+ SEM (n = 8). No t significant differences were found.
J. Physiol. Biochem., 61 (2), 2005
CONJUGATED LINOLEIC ACID AND LIPOLYSIS 367
1.5 I Control
'~ 1.2 [Z3 CLA
~ 0.9
'~ 0.6 Z
0.3
0 m g / g m g / d e p o t
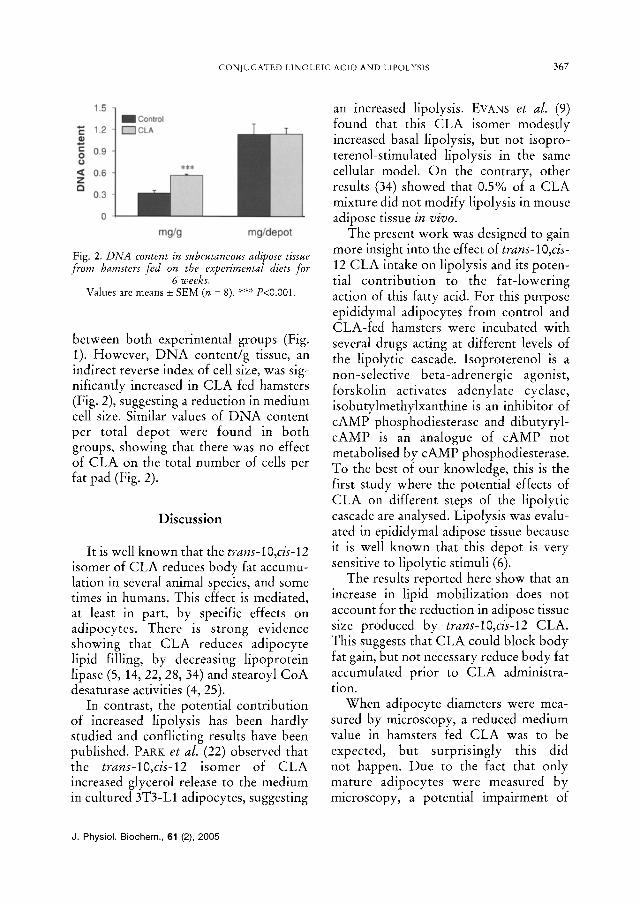
Fig. 2. DNA content in subcutaneous adipose tissue from hamsters fed on the experimental diets for
6 weeks. Values are means _+ SEM (n = 8). *** P<0.00I .
between both experimental groups (Fig. 1). However, DNA content/g tissue, ah indirect reverse index of cell size, was sig- nificantly increased in CLA fed hamsters (Fig. 2), suggesting a reduction in medium cell size. Similar values of DNA content per total depot were found in both groups, showing that there was no effect of CLA on the total number of cells per fat pad (Fig. 2).
Discussion
It is well known that the trans-10,cis-12 isomer of CLA reduces body fat accumu- lation in several animal species, and some times in humans. This effect is mediated, at least in part, by specific effects on adipocytes. There is strong evidence showing that CLA reduces adipocyte lipid filling, by decreasing lipoprotein lipase (5, 14, 22, 28, 34) and stearoyl CoA desaturase activities (4, 25).
In contrast, the potential contribution of increased lipolysis has been hardly studied and conflicting results have been published. PAV, K et al. (22) observed that the trans-lO,cis-12 isomer of CLA increased glycerol release to the medium in cultured 3T3-L1 adipocytes, suggesting
an increased lipolysis. EVANS et al. (9) found that this CLA isomer modestly increased basal lipolysis, but not isopro- terenol-stimulated lipolysis in the same cellular model. On the contrary, other results (34) showed that 0.5% of a CLA mixture did not modify lipolysis in mouse adipose tissue in vivo.
The present work was designed to gain more insight into the effect of trans-10,cis- 12 CLA intake on lipolysis and its poten- tial contribution to the fat-lowering action of this fatty acid. For this purpose epididymal adipocytes from control and CLA-fed hamsters were incubated with several drugs acting at different levels of the lipolytic cascade. Isoproterenol is a non-selective beta-adrenergic agonist, forskolin activates adenylate cyclase, isobutylmethylxanthine is an inhibitor of cAMP phosphodiesterase and dibutyryl- cAMP is an analogue of cAMP not metabolised by cAMP phosphodiesterase. To the best of our knowledge, this is the first study where the potential effects of CLA on different steps of the lipolytic cascade are analysed. Lipolysis was evalu- ated in epididymal adipose tissue because it is well known that this depot is very sensitive to lipolytic stimuli (6).
The results reported here show that an increase in lipid mobilization does not account for the reduction in adipose tissue size produced by trans-lO,cis-12 CLA. This suggests that CLA could block body fat gain, but not necessary reduce body fat accumulated prior to CLA administra- tion.
When adipocyte diameters were mea- sured by microscopy, a reduced medium value in hamsters fed CLA was to be expected, but surprisingly this did not happen. Due to the fact that only mature adipocytes were measured by microscopy, a potential impairment of
J. Physiol. Biochem., 61 (2), 2005

368 e. SIM‰ M. T. MACARULLA, A. FERN~NDEZ QU1NTELA e~ al.
preadipocyte differentiation, induced by CLA feeding, could be suggested. In order to obtain more information con- cerning this issue, D N A content was mea- sured. This measurement was carried out in the subcutaneous depot because it is well known that this anatomical location is the most sensitive to preadipocyte dif- ferentiation (10). The amount of D N A per gram of tissue, a reverse indicator of cell size, was significantly increased by CLA feeding, but total D N A content, an indi- cator of total cell number, was not modi- fied.
Taking both microscopy and D N A results into consideration, the possible conclusion could be that adipose tissue from trans-lO,cis-12 CLA-fed animals contained a lower amount of mature adipocytes but a greater amount of preadipocytes, assessed by D N A analysis although not counted by microscopy. This decrease in mature adipocytes may be responsible for the reduced adipose tis- sue size. In fact, because adipocyte pre- cursor cells (i.e. preadipocytes) ate present throughout life (1, 11), inhibition of fat- tening may be mediated not only by reducing body fat accumulation in differ- entiated adipocytes, but also by inhibiting the differentiation of preadipocytes into adipocytes. This hypothesis is supported by several papers, carried out with murine (2, 8, 12) of human (3) preadipocytes, which have showed that trans-10,cis-12 CLA inhibits adipocyte differentiation.
In summary, the present study demon- strates that the reduction in adipose tissue size produced by trans-lO,cis-12 CLA in hamsters is not due to changes in lipolytic activity. Reduced preadipocyte differenti- ation into mature adipocytes could account for this fat-lowering effect.
Acknowledgernents
This study was supported by grants from the Ministerio de Ciencia y TecnologŸ (BFI2002- 00273), the Government of the Basque Country (Biogune, Programa Etortek) and University of the Basque Country (00101.125-E-14788/2002 and 00101.125-15340/2003). Palm oil was a generous gift from Agra-Unilever (Leioa, Spain).
E. SIM‰ M. T. MACARULLA, A. FERN�93 V. M. RO- DR… y M. P. PORTILLO. La reducci£ de la adiposidad por el • linoleico con juga- do no se debe a incremento de la lipolisis. J. Physiol. Biochem., 61 (2), 363-370, 2005.
La capacidad del • linoleico conjugado (ALC) para reducir la adiposidad puede deber- se a cambios en el gasto energ› y/o a efec- tos directos sobre el metabolismo lipŸ de los adipocitos. En el presente trabajo se estudia si la ingesti£ de ALC modifica la actividad lipolŸ del tejido adiposo en h• ali- mentados con una dieta hipergrasa. Los anima- les se divididieron en dos grupos y se alimenta- ron durante 6 semanas con dietas suplementa- das al 0,5% con • linoleico (control) o con el is£ trans-lO,cis-12 del ALC, respectiva- mente. Los animales se sacrificaron tras una noche de ayuno y, previa disecci£ se pesaron sus dep£ adiposos. Los adipocitos se aisla- ron mediante digesti£ con colagenasa y se incubaron en tamp£ Krebs-Ringer bicarbo- nato sin y con varias sustancias que act¨ a diferentes niveles de la cascada lipolŸ Se midieron sus di• medios mediante microscopŸ y se determin£ el contenido en DNA del tejido adiposo por espectrofotome- trŸ Los animales que ingirieron ALC presen- taron una reducci£ del tama¡ de los dep£ tos adiposos. No se encontraron diferencias significativas entre ambos grupos en la lipolisis basal, en los efectos lipolŸ del isoprotere- nol, forskolina, dibutiril-cAMP e isobutilme- tilxantina, ni en la pD2 del isoproterenol. El contenido total de DNA del tejido adiposo fue similar en ambos grupos, lo que indica que el ALC no afect£ al n¨ total de adipocitos por dep£ adiposo. Aunque el contenido de DNA por gramo de tejido, un Ÿ inverso
J. Physiol. Biochem., 61 (2), 2005

CONjUGATED LINOLEIC ACID AND LIPOLYS[S 369
del tama¡ celular, aument£ significativamen- te, el an• microsc£ no mostr£ diferen- cias en el di• medio de los adipocitos maduros ni en su distribuci£ por tama¡ celular. Estos resultados sugieren que la reduc- ci£ del tama¡ del tejido adiposo inducida por la ingesti£ de A L C no se debe a cambios en la lipolisis, sino posiblemente a una menor diferenciaci£ de los preadipocitos a adipoci- tos maduros.
Palabras clave: �93 linoleico conjugado, Tejido adiposo, Lipolisis, H•
R e f e r e n c e s
1. Ailhaud, G. (1992): Diabetes Metab. Rev., 8, 3-7. 2. Brodie, A. E., Manning, V. A., Ferguson, K. R.,
Jewell, D. E. and Hu, Y. (1999): J. NuŸ 129, 602-606.
3. Brown, J. M., Boysen, M. S., Jensen, S. S., Morri- son, R. F., Storkson, J., Lea-Currie, R., Pariza, M., Mandrup, S. and Mclntosh, M. K. (2003): J. Lipid Res., 44, 1287-1300.
4. Choi, Y., Kim, Y. C., Han, Y. B., Park, Y., Pariza, M. W. and Ntambi, J. M. (2000):J. Nutr., 130, 1920-1924.
5. Cl› L., Poirier, H., Niot, I., Bocher, V., Guerre-Millo, M., Krief, S., Staels, B. and Besnard, P. (2002): J. Lipid Res., 43, 1400-1409.
6. Dieudonne, M. N., Pecquery, R. and Giudicelli, Y. (1992): Eur. J. Biochem., 205, 867-873.
7. Dole, V. P. and Meinertz, H. (1960): J. Biol. Chem., 235, 2595-2599.
8. Evans, M., Park, Y., Pariza, M., CurtŸ L., Kue- bler, B. and Mclntosh, M. (2001): Lipids, 36, 1223-1232.
9. Evans, M., Lin, X., Odle, J., and Mclntosh, M. (2002):J. Nutr., 132, 450-455.
10. GarcŸ P., Bellido, D. and Salas-Salvad£ J. (2004): Rey. Esp. Obes., 2, supp. 1, 57-68.
11. Gregoire, F. M., Smas, C. M. and Sul, H. S. (1998): Physiol. Rey., 78, 783-809.
12. Kang, K., Liu, W., Albright, K. J., Park, Y. and Pariza, M. W. (2003): Biochem. Biophys. Res. Commun., 303, 795-799.
13. Larsen, T.M., Toubro, S. and Astrup, A. (2003): J. Lipid Res., 44, 2234-2241.
14. Lin, Y., Kreeft, A., Schuurbiers, J. A. and Drai- jer, R. (2001):J. Nutr. Biochern., 12, 183-189.
I5. Macarulla, M. T., Fern• A., Zabala, A., Navarro, V., EchevarrŸ E., Churru- ca, I., RodrŸ V. M. and Portillo, M. P. (2005): Nutrition, 21,512-529.
16. MartŸ J. C., Gr› S., Siess, M. H., Genty, M., Chardingy, J. M., Berdeaux, O., Juan› P. and S›233 J. L. (2000): Lipids, 35, 91-98.
17. Martin, J. C. and Valeille, K. (2002): Reprod. Nutr. Dev., 42, 525-536.
18. MartŸ J. A., Macarulla, M. T., Marcos, R. and Larralde, J. (1992): Br. J. Nutr., 68,493-503.
19. Navarro, V., Zabala, A., Macarulla, M. T., Fer- n• A., RodrŸ V. M., Sim£ E. and Portillo, M. P. (2003): J. PhysioL Biochem., 59, 193-199.
20. Onuki, K., Haramizu, S., Ishihara, K. and Fushi- ki, T. (2001): Biosci. Biotechnol. Biochem., 65, 2200-2204.
21. Pariza, M. W., Park, Y. and Cook, M. E. (2001): Prog. Lipid. Res., 40, 283-298.
22. Park, Y., Albright, K. J., Liu, W., Storkson, J. M., Cook, M. E. and Pariza, M. W. (1997): Lipids, 32, 853-858.
23. Reeves, P. G., Nielsen, F. H. and Fahey, G. C. Jr. (1993):]'. Nutr., 123, 1939-1951.
24. Rodbell, M. (1964)J. Biol. Chem., 239, 375-380. 25. Smith, S. B., Hively, T. S., Cortese, G. M., Han,
J. J., Chung, K. Y., Castaneda, P., Gilbert, C. D., Adams, V. L. and Mersmann H. J. (2002): J. Anim. Sci., 80, 2110-2115.
26. Takahashi, Y., Kushiro, M., Shinohara, K. and Ide, T. (2002): Comp. Biochern. Physiol., 133, 395-404.
27. Terpstra, A.H.M. (2004): Aro. J. Clin. Nutr., 79, 352-361.
28. Tsuboyama-Kasaoka, N., Takahashi, M., Tane- mura, K., Kim, H. J., Tange, T., Okuyama, H., Kasai, M., Ikemoto, S. and Ezaki, O. (2000): Dia- betes, 49, 1534-1542.
29. Wang, Y. W. and Jones, P. J. H. (2004): Int. J. Obes., 28: 941-955.
30. Warren, J. M., Simon, V. A., Bartolini, G., Erick- son, K. L., Mackey, B. E. and Kelley, D. S. (2003): Lipids, 38, 497-504.
31. West, D. B., Delany, J. P., Carnet, P. M., Blohm, F., Truett, A. A. and Scimeca, J. (1998): Aro. J. Physiol., 275, R667-672.
32. West, D. B., Blohm, F .Y., Truett, A. A. and DeLany, J. P. (2000): J. Nutr., 130, 2471-2477.
33. Wieland, O. (1957): Biochern. Z., 239, 313-319. 34. Xu, X., Storkson, J., Kim, S., Sugimoto, K., Park,
Y. and Pariza, M. W. (2003): J. Nutr., 133, 663- 667.
J. Physiol. Biochem. 61 (2), 2005




















