
Ants (Formicidae) and Spiders (Araneae) listed from the Metropolitan Region of Salvador, Brazil
Upload
independentCategory
view
1download
0
INTRODUCTION
Wood ants of the Formica rufa-group haveachieved unquestionable ecological success. Living inmature, mainly coniferous, forests of the northern Pa-laearctic, they evolved in permanent, stable and troph-ically rich habitats. Their food basis consists of aphidhoneydew, a constant and practically endless source ofcarbohydrates, so called energy food, and diverseinvertebrates as a source of protein food essential for
reproduction and brood development. The high numer-ical strength of colonies, a high level of social structureand organisation, compound nest construction, andterritoriality combined with a system of foraging routesaccount for the top position of the members of the F. rufa-group in ant assemblages of boreal and tem-perate forests (e.g., Vepsäläinen and Pisarski 1982,Savolainen and Vepsäläinen 1988, 1989, Pisarski andVepsäläinen 1989). Wood ants that are able to developdense polycalic (multinest) colonies rank as top
TERRITORY SIZE OF WOOD ANTS (HYMENOPTERA:FORMICIDAE): A SEARCH FOR LIMITS OF EXISTENCE
OF FFOORRMMIICCAA PPOOLLYYCCTTEENNAA FÖRST., AN INHERENTLYPOLYGYNIC AND POLYCALIC SPECIES
AA NN NN AA LL EE SS ZZ OO OO LL OO GG II CC II ((WWaarrsszzaawwaa)),, 22000099,, 5599((22)):: 117799--118877
WOJCIECH CZECHOWSKI1 and KARI VEPSÄLÄINEN2
1Laboratory of Social and Myrmecophilous Insects, Museum and Institute of Zoology, Polish Academy of Sciences, Wilcza 64, 00-679 Warszawa, Poland;
e-mail: [email protected] of Biological and Environmental Sciences, P.O. Box 65, 00014
University of Helsinki, Finland; e-mail: [email protected]
Abstract.— Wood ants are often absent on islands of the Gulf of Finland, even whenseemingly suitable habitats are available. Their absence may partially be explained by thelack or rarity of ant species suitable as host during colonisation through temporaryparasitism. To search for the limits of living conditions on islands, given colonisationconstraints are overcome, we artificially established wood ant colonies on several islandsconstituting a series from suboptimal to extremely harsh living conditions. The casereported here showed that a barren < 0.2 ha islet, with aphids on its single pine tree theonly permanent and relatively rich food source, has allowed the existence of an artificiallyintroduced Formica polyctena Först. colony for 22 years. The ambient living conditionsare probably close to the limit for the species, as evidently the colony does not producesexual offspring. Thus the sustained existence of the colony is dependent on adoption offertile gynes originating in colonies that live in more optimal conditions. We suggest that thepolygynic social mode of the inherently polycalic F. polyctena is the key for its sustainedexistence on the islet, as polygyny together with receptivity to new, even alien queens keepsthe colony alive in a sink habitat insufficient for production of own sexual offspring.
Key words.— Artificial colonisation, food resources, Formica polyctena, habitat require-ments, intraspecific competition, polygyny, queen recruitment, sexual production, terri-toriality, wood ants.
PL ISSN 0003-4541 © Fundacja Natura optima duxdoi: 10.3161/000345409X463994

dominants in ant assemblages, given suitable habitatfor the species. As the mode of life of territorial antspecies requires great energy outlays, sustained exis-tence of colonies of polycalic wood ants seem – deduc-ing from their natural history – to demand an exten-sive, stable and productive environment (Pisarski andVepsäläinen 1989), usually realised in relatively oldforests (Dlusskij 1967, Collingwood 1979).
The extensive surveys by Vepsäläinen and Pisarski(1982 and unpubl.) of ant assemblages within the Tvär-minne archipelago (westernmost part of the Gulf ofFinland, Baltic Sea) showed that wood ants wereabsent from many islands that seemed suitable forthem habitat-wise. As fertilised gynes were found dur-ing their nuptial periods even on the remotest islandsof the archipelago (K. Vepsäläinen and B. Pisarski,unpubl.), the causes of the absence of wood ants theremight be traced back to difficulties in colony founding,mainly lack of Serviformica For. species that are obli-gate hosts when colonising islands through temporarysocial parasitism. In our study area, Formica (Servi-formica) fusca L. is rare on small islands, even next to the mainland (Vepsäläinen and Pisarski 1982).A small stretch of water, less than 100 m, may beenough to isolate a small island void of F. fusca fromthe rocky outcrops of a larger island with a healthy F. fusca population (K. Vepsäläinen, unpubl.).
Here, we raise the question of the habitat require-ments of wood ants. Given that the critical foundingphase of the colony could be overcome, how extremesuboptimal conditions would still allow sustained existence of a wood-ant colony? To answer the ques-tion, we brought wood ants of the F. rufa-group toislands where they were lacking, and where the local habitat conditions seemed to be suboptimal forthem. While planning our series of introductions ofwood ants to small islands differing more or lessextremely from the normal forest habitat of F. rufa-group species, we used ‘suboptimal’ without trying tospecify suboptimality a priori, and rather expectedeach experiment in due time to render specific in-formation on local suboptimal conditions. Here wereport only on an artificial colonisation of the highlypolycal-ic species F. polyctena Först. on an islet thathas enabled long-time survival of the colony in livingconditions evidently close to the limit of existence. Wealso discuss the results in the light of territorial andfor-aging behaviour of wood ants in their more typicalhabitats.
STUDY AREA, MATERIAL AND METHODS
The Tvärminne archipelago includes dozens of rockyislands of various sizes. Their habitat spectrum main-ly embraces different stages of primary succession,
ranging from an open rocky outcrop to a mature taiga-like pine forest. Open rocks prevail on islets smallerthan 0.5 ha, whereas patches of forest appear on aboutone-hectare islands, and the forest is fully developed onislands of a few hectares (see Vepsäläinen and Pisars-ki 1982).
We chose a small islet, ca. 1800 m2, informallycalled the Skeleton Islet, one of the Kvarnskärs-grunden complex (Figs 1–4). The islet was almost treeless, with a predominance (more than 60% of thewhole surface) of an open rock outcrop in places over-grown with moss and intersected by rock crevices over-grown with pioneer vegetation (Figs 2, 4). There werethree compact patches of green separated from eachother (Fig. 2). These included two stony shore mead-ows flooded during storms (ca. 150 m2 and 450 m2,including a substantial area covered by stones; Figs 5,6). The bigger meadow constituted the lower terrace ofthe islet, separated by a fault from the rest of the area(Fig. 6). The third patch (ca. 100 m2) was an oblong veg-etated depression among rocks overgrown with herbsand grass, a dense clump of junipers (Juniperus com-munis L., Fig. 7), and a ca. 2.5 m high pine tree (Pinussylvestris L.), a shoot growing from a fallen, dead treetrunk. There were also three 2.5–3.5 m tall alder trees[Alnus glutinosa (L.) Gaertn.], with multiple trunks,and two ca. 2 m high rowans (Sorbus aucuparia L.) inthe smaller, and a 4 m high alder tree in the biggermeadow (Figs 2, 5, 6); in 2000s, some alder saplingsappeared in the bigger meadow (see Fig. 6). During the21 years of observation, growth of the alder trees washardly noticeable, and the rowans were graduallydying out. The original local myrmecofauna consistedonly of a few weak colonies of Lasius niger (L.) and L. flavus (F.) (the latter only in the bigger meadow).Later (starting from the late 1990s) we observed work-ers of Formica fusca, but did not find the nest.
On 10th June 1987, we brought a branch [ca. 50 dm3
of mound material (see Fig. 2 in Czechowski 1990)with, in proportion to the nest material volume, a highexcess of workers and worker pupae of a F. polyctenacolony from the mainland (Långholmen peninsula; seeFigs 1, 4). We established it, as colony SI-P-1, on theSkeleton Islet in a hollow among rocks within thejuniper clump, at a distance of 1.7 m from the pine tree(Figs 2, 7). Then, in late June 2001 and early June 2006,we introduced, in succession, two smaller experimen-tal colonies of F. polyctena (SI-P-2 and SI-P-2’; Fig. 8)that we took from another region of the Långholmenpeninsula. The new introductions originated farenough (ca. 260 m) from the site of the first maternalcolony (Fig. 1), to secure that the old and new ‘artifi-cial’ colonies were mutually alien societies. Here, ouraim was to introduce a competitor to the residentcolony. They were placed as far as it was possible (i.e., 22–23 m away) from SI-1 (Fig. 2). We carried out
180 W. CZECHOWSKI and K. VEPSÄLÄINEN

each colonisation along the general principles thatguarantee proper functioning of newly founded col-onies (Pisarski and Czechowski 1990, 1991; see alsoCzechowski 1990).
Each year from 1992 to 2008 in June or July, weinspected the condition of colony SI-P-1, its nest struc-ture, food sources and range covered by workers offthe mound; in June 2009, only the mound dimensionswere taken and the nest checked for possible sexualsoffspring. We estimated the mound volume aboveground level with the formula for the volume of half anellipsoid as follows: volume = 2/3 * π * r2 * h, where r ismound radius at bottom and h its height from groundlevel. Starting in 2002, each year in June, we paid spe-cial attention to possible presence of sexuals (adults orbrood) in the mound of colony SI-P-1. We checked thecolonies SI-P-2 and SI-P-2’, their behaviour and con-tacts with SI-P-1 for 10 days after their, establishment,and then revisited them a year later.
RESULTS
On 27th June, seventeen days after colonisation, themound of the SI-P-1 colony, already properly shaped byants, was 25 cm in height and 55 cm in diameter. Thecolony permanently stayed at the site chosen for it, andhas been living there – contrary to expectations – in a good condition for the last 22 years. The nest moundwas mainly made up of juniper needles, lichen rem-nants and crushed dry reeds, the latter deposited by
LIMITS OF EXISTENCE OF FORMICA POLYCTENA 181
Figure 1. A map of the fragment of the Tvärminne archipelago with nearest islands, including Kvarnskärgrunden (Skeleton Islet blackened,uncaptioned island between Långholmen and Joskär – Vikaskär, • – locations of maternal colonies: southern dot – maternal colony of SI-P-1,
northern dot – maternal colony of SI-P-2 and SI-P-2’) (based on a map by Maanmittaushallituksen kivipaino 1966).
waves. During the first few years, the mound grew considerably; then, in the years 1992–2009 its dimen-sions remained relatively constant, with a diameter of90–120 cm (mean ± SD = 103 ± 8 cm) and a height of50–70 cm (58 ± 5 cm) (n = 18 for both), and an estimat-ed above-ground volume of 324 ± 48 dm3 (minimum212, maximum 380 dm3) (Table 1). The mound showedno tendency of steady growth or decimation duringthese years (p = 0.13, r2 adjusted for degrees of free-dom = 0.08).
During yearly checks of the mound made in June,we never observed SI-P-1 to produce sexuals; we foundneither adults nor pupae of gynes or male. The extra-nidal activity of workers was weak. Both dynamic den-sity and the range searched by workers were amazing-ly small for a colony inhabiting such a big nest. Theactual numerical strength of the colony was only re-vealed when the ants were irritated directly close to thenest, and the ants attacked the observer in large num-bers. The foraging area of SI-P-1 included two of thethree (see Study area) green patches present on theislet: that with the nest and the smaller shore meadow.On the patch surrounding the nest, the main and rela-tively permanent carbohydrate food source was honey-dew of aphids on the only pine tree, where a distinctant route led. Chironomid flies were an ephemeral,though probably not unimportant, protein food sourcethere during their mass appearances; they often be-came trapped amongst the needles of junipers that surrounded the nest (see Fig. 7), being easy prey forants. In the shore meadow, ants mainly searched the
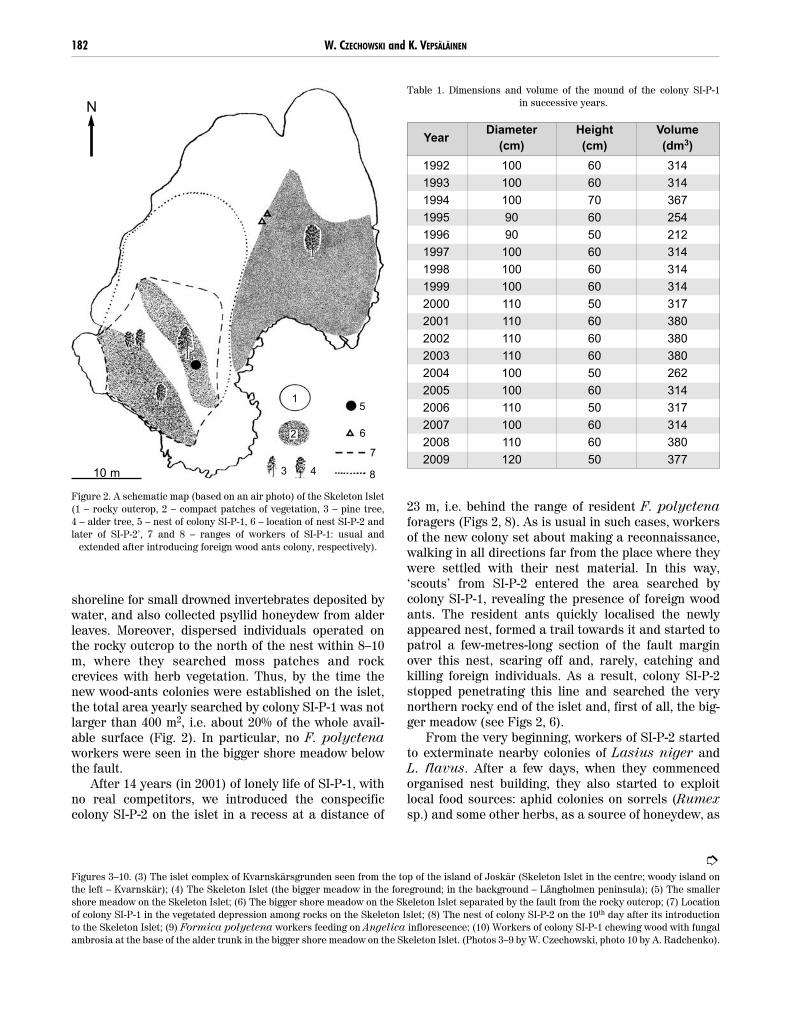
shoreline for small drowned invertebrates deposited bywater, and also collected psyllid honeydew from alderleaves. Moreover, dispersed individuals operated onthe rocky outcrop to the north of the nest within 8–10m, where they searched moss patches and rockcrevices with herb vegetation. Thus, by the time thenew wood-ants colonies were established on the islet,the total area yearly searched by colony SI-P-1 was notlarger than 400 m2, i.e. about 20% of the whole avail-able surface (Fig. 2). In particular, no F. polyctenaworkers were seen in the bigger shore meadow belowthe fault.
After 14 years (in 2001) of lonely life of SI-P-1, withno real competitors, we introduced the conspecificcolony SI-P-2 on the islet in a recess at a distance of
182 W. CZECHOWSKI and K. VEPSÄLÄINEN
Year Diameter Height Volume(cm) (cm) (dm3)
1992 100 60 3141993 100 60 3141994 100 70 3671995 90 60 2541996 90 50 2121997 100 60 3141998 100 60 3141999 100 60 3142000 110 50 3172001 110 60 3802002 110 60 3802003 110 60 3802004 100 50 2622005 100 60 3142006 110 50 3172007 100 60 3142008 110 60 3802009 120 50 377
Table 1. Dimensions and volume of the mound of the colony SI-P-1 in successive years.
23 m, i.e. behind the range of resident F. polyctenaforagers (Figs 2, 8). As is usual in such cases, workersof the new colony set about making a reconnaissance,walking in all directions far from the place where theywere settled with their nest material. In this way,‘scouts’ from SI-P-2 entered the area searched bycolony SI-P-1, revealing the presence of foreign woodants. The resident ants quickly localised the newlyappeared nest, formed a trail towards it and started topatrol a few-metres-long section of the fault marginover this nest, scaring off and, rarely, catching andkilling foreign individuals. As a result, colony SI-P-2stopped penetrating this line and searched the verynorthern rocky end of the islet and, first of all, the big-ger meadow (see Figs 2, 6).
From the very beginning, workers of SI-P-2 startedto exterminate nearby colonies of Lasius niger and L. flavus. After a few days, when they commencedorganised nest building, they also started to exploitlocal food sources: aphid colonies on sorrels (Rumexsp.) and some other herbs, as a source of honeydew, as
Figures 3–10. (3) The islet complex of Kvarnskärsgrunden seen from the top of the island of Joskär (Skeleton Islet in the centre; woody island onthe left – Kvarnskär); (4) The Skeleton Islet (the bigger meadow in the foreground; in the background – Långholmen peninsula); (5) The smallershore meadow on the Skeleton Islet; (6) The bigger shore meadow on the Skeleton Islet separated by the fault from the rocky outcrop; (7) Locationof colony SI-P-1 in the vegetated depression among rocks on the Skeleton Islet; (8) The nest of colony SI-P-2 on the 10th day after its introductionto the Skeleton Islet; (9) Formica polyctena workers feeding on Angelica inflorescence; (10) Workers of colony SI-P-1 chewing wood with fungalambrosia at the base of the alder trunk in the bigger shore meadow on the Skeleton Islet. (Photos 3–9 by W. Czechowski, photo 10 by A. Radchenko).
Figure 2. A schematic map (based on an air photo) of the Skeleton Islet(1 – rocky outcrop, 2 – compact patches of vegetation, 3 – pine tree, 4 – alder tree, 5 – nest of colony SI-P-1, 6 – location of nest SI-P-2 andlater of SI-P-2’, 7 and 8 – ranges of workers of SI-P-1: usual and
extended after introducing foreign wood ants colony, respectively).
10 m
N
2
3 4
5
6
7
8
1

LIMITS OF EXISTENCE OF FORMICA POLYCTENA 183
3 4
5 6
7 8
9 10

well as the inflorescence of Angelica sp. (Umbelli-ferae), as a source of nectar or pollen (Fig. 9). We nev-er observed workers of colony SI-P-1 visit aphids onherbs or feeding on flowers. Toward the end of theobservation period, workers of SI-P-2 came up to thetrunk of the alder tree on the big meadow (8 m awayfrom their nest), occupied at that time by L. niger ants,and started to enter into skirmishes with them.
In the next season (in 2002) colony SI-P-2 did notexist. Total destruction of herb vegetation in the wholearea of the bigger meadow, and a strip of mud deposit-ed along the bottom margin of the fault indicated thatthe nest had been washed away by a strong storm. The remnants of the nest mound were inhabited by L. niger. At the same time we noticed that colony SI-P-1, for the first time in its 15-year existence, hadsignificantly extended its searching area. The range ofworkers to the north of the nest had grown to morethan 20 m, and they controlled all small patches ofvegetation on the rocky outcrop. The total areasearched had increased to ca. 700 m2, i.e., by 75% (Fig. 2). Next year (2003), however, the situation re-turned to normal (see Fig. 2).
In 2006, five years after the establishment of colonySI-P-2, we brought one more experimental F. polycte-na colony, SI-P-2’, close to the former nest site of SI-P-2 but to an apparently safer location (Fig. 2). The sce-nario repeated itself in every way, including total elim-ination of the new colony by water in the period ofautumn-spring. The next season (in 2007) saw colonySI-P-1 not only controlling almost the whole surface ofthe islet’s rocky outcrop (like in 2002), but even form-ing a distinct 22 m long foraging route from its nestacross the bigger meadow to the local alder tree (Figs2, 6). That was the first and only year we observed thewood ants of SI-P-1 to exploit the alder tree as theirfood source, collecting honeydew of aphids and psyllidsin the crown, and chewing wood saturated with fungalambrosia in a decayed part of the trunk (Fig. 10). Itwas also the first and only time we saw them descendinto the lower terrace of the islet. Next season (2007)the situation again returned to normal: wood antssearched only a small part of the rocky outcrop, and the alder tree in the bigger meadow was used by L. niger, as before.
DISCUSSION
First of all it is surprising that a colony of F. po-lyctena has been able to survive 22 years in such a poor habitat, maintaining a fairly good condition. Themean lifespan of wood-ant queens is ca. five years(Keller and Genoud 1997, Keller 1998), and the maxi-mum age of queens is estimated as about 25 years(Gösswald 1989). We do not know how many (if any)queens we introduced to the islet during artificial
colonisation – probably only a few with the nest mate-rial – and thus the odds of any of the original queensliving are close to zero. Because the maximum longevi-ty of wood ant workers is three years (Czechowski1996), to keep the colony going, fertile replacementqueens must have been present to produce new work-ers. The fairly stable size of the mound during the 18most recent years testifies that yearly at least onequeen has been present, keeping the workforce in a steady state. Nevertheless, we never observed pro-duction of sexuals in the study nest, though in yearlycontrols (all done in June) we found winged offspring inthree other artificially established colonies on nearbyislands (W. Czechowski, unpubl.). Thus we suggestthat colony SI-P-1 has, during the years, obtainedreplacement queens from the neighbouring healthycolonies on the neighbouring larger islands or themainland. Here, an interesting connotation emerges:the very polygynic (many queens) social organisation,allowing adoption of extra, even alien gynes to thecolony, seems to be crucial for the existence of F. po-lyctena in the extreme conditions of the Skeleton Islet– a situation where the prognosis for monogynic so-cieties specialised to live in small habitat patches, e.g.those of F. rufa, are very poor.
Another unexpected finding was the very small for-aging area used by a colony of that size, especially inreference to its possibilities: the colony normallyutilised the same part of the islet amounting to aboutone-fifth of the available area. On islands of the Tvär-minne archipelago, wood ants, if they occur, normallyoccupy pine forests, usually the richest local habitataccessible to them. On the nearby 10 ha island ofJoskär (see Fig. 1) in the 1980s, habitat mosaic of pineforest with an undergrowth and rocky outcrop withscattered pine trees was monopolised by two largecolonies of F. polyctena. Together their territoriescovered ca. 2.3 ha (Pisarski and Vepsäläinen 1989,Savolainen and Vepsäläinen 1989), and the coloniescontinuously expanded their territory by establishingremote daughter nests (K. Vepsäläinen and B. Pisar-ski, unpubl.). At the same time, the F. polyctena col-ony on the Skeleton Islet had, for years, settled for a foraging area of less than 400 m2 with one small pine tree. Not only natural wood ant colonies, but alsoall ‘artificial’ ones, successfully introduced to biggerand partly forested islands with well formed local antassemblages, had their foraging areas much biggerthan that of SI-P-1. For example, the areas occupied by a F. polyctena colony on Porsgrundet and twocolonies on Vikaskär islands (see Fig. 1) (all col-onies smaller than SI-P-1) reached ca. 690, 540 and 940 m2 respectively, in 1995, eight years after colonisa-tion, and each colony included from 10 to more than 20 pine trees visited for aphids (W. Czechowski,unpubl.).
184 W. CZECHOWSKI and K. VEPSÄLÄINEN

One could suggest that perhaps the growth of SI-P-1 to a colony producing sexual offspring would be constrained by the fixed, only possible nest site, as pre-dicted by the theory of central place foraging. Thisexplanation is here, however, implausible, becausewood ants may easily maintain a territory with a 60-mforaging radius from the nest, and visit trees withaphids beyond 100 m from their mound (e.g., Savo-lainen and Vepsäläinen 1989, Vepsäläinen and Savolai-nen 1994). By walking only about 40 m from the mound,workers of colony SI-P-1 could have reached the mostdistant tip of the islet. Our two attempts to introduce anadditional F. polyctena colony onto the Skeleton Islet,though unsuccessful, provided some insight to possiblereasons, why the healthy colony was stuck to such a small territory. They showed that, apart from theplace already occupied by colony SI-P-1, there werepractically no other possible nest sites suitable forwood ants, emphasising the hard local habitat condi-tions for wood ants. Also, we observed workers of theshort-lived colony SI-P-2 collecting honeydew fromaphids living on herbs and nectar (or pollen; seeCzechowski et al. 2008) from inflorescences, both atyp-ical and probably too small resources to allow exis-tence of a wood ant colony – a behaviour to whichworkers of SI-P-1 never seemed to commit themselves.
Our study emphasises the multifarious factors crit-ical in successful colonisation of islands by ants in gen-eral (Vepsäläinen and Pisarski 1982). The first one isdispersal capacity, evidently not a problem to woodants, not only within the most inshore islands of theTvärminne archipelago (see Introduction). On smallislands, the second requirement for wood ants, avail-ability of suitable temporary hosts during colonyfounding, is rarely fulfilled. The most critical, final fac-tors are availability of nest sites, marginally present onthe Skeleton Islet – and a reliable, continuous supply offood resources. An ample parallel to the present casestudy is the colonisation of small treeless skerries byLasius flavus. Together with that of L. niger, its dis-persal potential and number of propagules is by far thehighest among all local ant species. Whereas L. nigersuccessfully colonises smallest skerries with a mini-mum amount of soil in narrow crevices of open rock, L. flavus is able to establish itself on primary-succes-sion islands only after they have obtained a healthypopulation of aphids (their base food source) living on the roots of some grass species and pine saplings(Vepsäläinen and Pisarski 1982). [After Seifert’s (1991)described Lasius platythorax, which was previouslyconfused with L. niger, we (W. Czechowski, A. Rad-chenko and K. Vepsäläinen, unpubl.) checked thespecies identity of the pioneer species on the skerriesin Tvärminne (including the Skeleton Islet): the name L. niger maintains]. As comes to wood ants, withlaboratory colonies of F. polyctena, Lange (1960)
demonstrated that colonies fed with only carbohy-drates decreased in size by one third while coloniesthat obtained mainly carbohydrates supplemented byprotein food doubled their workforce. As it seems, onthe Skeleton Islet F. polyctena has been able to har-vest enough of protein food to maintain relatively sta-ble colony size, but not enough to enable production ofsexuals.
An illustrative series of events in the context of ter-ritoriality developed after the new introductions ofpotential competitors on the islet, and the strong work-force (relative to the small foraging area usuallysearched by them) of SI-P-1 became evident. Then theestablished colony substantially expanded their forag-ing area, only to retreat back to the smaller area afterthe new colony had died due to environmental vicissi-tudes. Such behaviour fits well the general picture onintra- and interspecific competition between wood-antcolonies and species, where – in presence of competi-tors – fierce territorial fights are a regular phenome-non in spring. Such “wood-ant wars” thus take placeduring the early season of activity, when the food re-quirements are higher than the food supply (Mabelis1979, 1984). Notably, the consequence of such encoun-ters is a buffer zone between neighbouring colonies,void of wood ants, and territories that in most situa-tions are much larger than usually needed by thecolonies.
We conclude by suggesting that the small islet withits harsh conditions represent the extreme limit to sus-tained existence of wood ants. This conclusion obtainssupport from our artificial introductions of F. polycte-na to two small, totally treeless islets, where one of thecolonies succumbed in three, the other in five years (W. Czechowski, unpubl.). Notwithstanding the longexistence of the introduced colony SI-P-1, a number offacts indicate that, applying the concept of sink–sourcehabitats (see Wiens 1989), the islet is a sink habitat toF. polyctena: (1) Even though the artificially estab-lished colony has now lived 22 years on the original siteof introduction, we have never observed them producesexual offspring; thus their further existence seems todepend on adoption of gynes originating in healthy col-onies of close-by source habitats. It seems that the onlyway to explain the lack of sexual production is theshortcoming of a final boost of resources that wouldcarry the colony beyond the threshold to a maturecolony; (2) Even though the whole islet is smaller thanthe area of a wood-ant territory typical of a colony ofthis size, the workforce of the colony used maximally20% of the available area – one more sign of scantyresources almost everywhere on the islet, not worth ofcontinuous searching without reward; (3) The shortexistence of the two additional introduced colonies onthe islet, and their evidently strongly limiting and atyp-ical food resources also indicate that colony SI-P-1 is
LIMITS OF EXISTENCE OF FORMICA POLYCTENA 185

186 W. CZECHOWSKI and K. VEPSÄLÄINEN
highly dependent on a spatially very limited food source,mainly aphids on the single pine tree next to the mound.
A highly relevant future task would be to study clos-er the use by workers of aphids on the only pine tree on our study islet. Evidently, the fickle appearance ofchironomids, although at times superabundant, andremnants of fish brought on the islet by gulls, seem notto satisfy the safe and right timing of high-grade pro-tein-rich food needed for production of sexuals. Thenutritional requirements for production of sexuals inwood ants are strict and complicated; for a concisereview of the intricate determination of sexual casts,and the role of food therein, see pages 174–175 in Höll-dobler and Wilson (1990). Thus the only reliable poten-tial source of protein there would be aphids, given thatthe aphid population is able to produce enough of sur-plus individuals that could be carried to the nest asprey. As is known, wood ants do predate also on aphidsthat they tend mainly for honeydew (e.g., Skinner 1980,Rosengren and Sundström 1987), but the results of thisstudy point toward too little of protein. Thus we sug-gest that our study colony – though successful whenrated by longevity – has hit the local resource ceilingbefore exceeding zero fitness, the ultimate minimumcriterion of success. A recent study supports our con-clusion about the key role of food as a limiting resource:through comparisons of wood ant colonies in pre- andpost-clearcut patches with those in forest interiors, Sor-vari and Hakkarainen (2009) suggested that the observ-ed decrease of worker size, lower body-fat contents ofworkers and lower nest temperatures after logging aredue to a decrease in the availability of tree-living aphids,the main food resource (see also Punttila et al. 1991).Summarising, ample food resources are mandatory inmaintaining stable, optimal nest temperatures for pro-duction of sexual offspring (Rosengren et al. 1987).
ACKNOWLEDGEMENTS
We are grateful to Bálint Markó and Pekka Punttilafor their helpful comments on our manuscript. Wethank Bálint Markó and Piotr Ślipiński for checking thestate of SI-P-1 in 2009. We carried out the field re-search within the program of scientific cooperationbetween the Polish Academy of Sciences and the Acad-emy of Finland. Tvärminne Zoological Station, Univer-sity of Helsinki, provided working facilities and a pleas-ant environment for the study.
REFERENCES
Collingwood, C. A. 1979. The Formicidae (Hymenoptera) ofFennoscandia and Denmark. Fauna Entomologica Scandi-navica, 8, 174 pp.
Czechowski, W. 1990. Aggression of Formica aquilonia
Yarr. to Camponotus ligniperdus (Latr.) (Hymenoptera,Formicidae) under conditions of artificial colonization.Memorabilia Zoologica, 44: 83–91.
Czechowski, W. 1996. Colonies of hybrids and mixed colonies;interspecific nest takeover in wood ants (Hymenoptera,Formicidae). Memorabilia Zoologica, 50, 116 + xx pp.
Czechowski, W., Markó, B. and A. Radchenko. 2008. Rubbishdumps reveal the diet of ant colonies: Myrmica schenckiEm. and Myrmica rubra (L.) (Hymenoptera: Formicidae)as facultative pollen-eaters. Polish Journal of Ecology, 56:737–741.
Dlusskij, G. M. 1967. Murav’i Roda Formika. (Hymenoptera,Formicidae, G. Formica). Nauka, Moskva, 236 pp.
Gösswald, K. 1989. Die Waldameise. Band 1, BiologischeGrundlagen, Ökologie und Verhalten. Aula-Verlag, Wies-baden, 660 pp.
Hölldobler, B. and E. O. Wilson. 1990. The Ants. Harvard Uni-versity Press, Cambridge, MA, 733 pp.
Keller, L. 1998. Queen lifespan and colony characteristics inants and termites. Insectes Sociaux, 45: 235–246.
Keller, L. and M. Genoud. 1997. Extraordinary lifespans inants: a test of evolutionary theories of ageing. Nature, 389:958–960.
Lange, R. 1960. Modellversuche über den Nahrungsbedarf vonVölkern der Kahlrückigen Waldameise Formica polycte-na Först. Zeitschrift für Angewandte Entomologie 46:200–208.
Maanmittaushallituksen kivipaino. 1966. Tvärminne perus-kartta 1:20000 No: 201111. Helsinki.
Mabelis, A. A. 1979. Wood ant wars. The relationship betweenaggression and predation in the red wood ant (Formicapolyctena Först.). Netherlands Journal of Zoology, 29:451–620.
Mabelis, A. 1984. Aggression in wood ants (Formica polycte-na Foerst., Hymenoptera, Formicidae). Aggressive Behav-ior, 10: 47–53.
Pisarski, B. and W. Czechowski. 1990. The course of artificialcolonization of red wood ants in the Gorce National Park.Memorabilia Zoologica, 44: 37–46.
Pisarski, B. and W. Czechowski. 1991. Modalites de colonisa-tion des fourmis de grouppe Formica rufa au Parc Na-tional de Gorce (Pologne). Actes des Colloques InsectesSociaux, 6: 237–242.
Pisarski, B. and K. Vepsäläinen. 1989. Competition hierar-chies in ant communities (Hymenoptera, Formicidae).Annales Zoologici, 42: 321–328.
Punttila, P., Haila, Y., Pajunen, T. and H. Tukia. 1991. Coloni-sation of clearcult forests by ants in southern Finland: a quantitative survey. Oikos, 61: 250–262.
Rosengren, R., Fortelius, W. Lindström, K. and A. Luther1987. Phenology and causation of nest heating and ther-moregulation in red wood ants of the Formica rufa groupstudied in coniferous forest habitats in southern Finland.Annales Zoologici Fennici, 24: 147–155.
Rosengren, R. and L. Sundström. 1987. The foraging system ofa red wood ant colony (Formica s. str.) – collecting anddefending food through an extended phenotype. In: FromIndividual to Collective Behavior in Social Insects. LesTreilles Workshop. Birkhäuser Verlag, Basel. ExperientiaSupplementum, 54: 117–137.
Savolainen, R. and K. Vepsäläinen. 1988. A competition

hierarchy among boreal ants: impact on resource parti-tioning and community structure. Oikos, 51: 135–155.
Savolainen, R. and K. Vepsäläinen. 1989. Niche differentiationof ant species within territories of the wood ant Formicapolyctena. Oikos, 56: 3–16.
Seifert, B. 1991. Lasius platythorax n. sp., a widespread sibling species of Lasius niger (Hymenoptera: Formici-dae). Entomologica Generalis, 16: 69–81.
Skinner, G. J. 1980. The feeding habits of the wood-ant, For-mica rufa (Hymenoptera: Formicidae), in limestonewoodland in north-west England. Journal of Animal Ecol-ogy, 49: 417–433.
Sorvari, J. and H. Hakkarainen. 2009 (preprint). Forest clear-cutting causes small workers in the polydomous wood antFormica aquilonia. Annales Zoologici Fennici, 46.
Vepsäläinen, K. and B. Pisarski. 1982. Assembly of island antcommunities. Annales Zoologici Fennici, 19: 327–335.
Vepsäläinen, K. and R. Savolainen. 1994. Ant-aphid interac-tion and territorial dynamics of wood ants. MemorabiliaZoologica, 48: 251--259.
Wiens, J. A. 1989. The Ecology of Bird Communities. Vol. 1,Processes and Variations. Cambridge University Press,Cambridge UK, 328 pp.
LIMITS OF EXISTENCE OF FORMICA POLYCTENA 187
Received: May 11, 2009Accepted: June 15, 2009
Copyright © 2022 FDOKUMEN




















