
Social facilitation and food partitioning in the queenless ant Dinoponera quadriceps (Hymenoptera:...
10
This article was downloaded by: [Instituto de Pesquisas e Estudos Florest] On: 29 August 2012, At: 08:50 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Natural History Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tnah20 Social facilitation and food partitioning in the queenless ant Dinoponera quadriceps (Hymenoptera: Formicidae) Fabio S. Nascimento a , Danilo I.S.A. Souza b , Ivelize C. Tannure- Nascimento a & José O. Dantas b a Universidade de São Paulo, FFCLRP, Departamento de Biologia, Ribeirão Preto, São Paulo, Brazil b Universidade Federal de Sergipe, CCBS, Departamento de Biologia, São Cristóvão, Sergipe, Brazil Version of record first published: 08 Aug 2012 To cite this article: Fabio S. Nascimento, Danilo I.S.A. Souza, Ivelize C. Tannure-Nascimento & José O. Dantas (2012): Social facilitation and food partitioning in the queenless ant Dinoponera quadriceps (Hymenoptera: Formicidae), Journal of Natural History, 46:31-32, 1959-1967 To link to this article: http://dx.doi.org/10.1080/00222933.2012.700333 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and- conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
uniplan-tec -
Category
Documents
-
view
3 -
download
0
Transcript of Social facilitation and food partitioning in the queenless ant Dinoponera quadriceps (Hymenoptera:...

This article was downloaded by: [Instituto de Pesquisas e Estudos Florest]On: 29 August 2012, At: 08:50Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
Social facilitation and food partitioningin the queenless ant Dinoponeraquadriceps (Hymenoptera: Formicidae)Fabio S. Nascimento a , Danilo I.S.A. Souza b , Ivelize C. Tannure-Nascimento a & José O. Dantas ba Universidade de São Paulo, FFCLRP, Departamento de Biologia,Ribeirão Preto, São Paulo, Brazilb Universidade Federal de Sergipe, CCBS, Departamento deBiologia, São Cristóvão, Sergipe, Brazil
Version of record first published: 08 Aug 2012
To cite this article: Fabio S. Nascimento, Danilo I.S.A. Souza, Ivelize C. Tannure-Nascimento &José O. Dantas (2012): Social facilitation and food partitioning in the queenless ant Dinoponeraquadriceps (Hymenoptera: Formicidae), Journal of Natural History, 46:31-32, 1959-1967
To link to this article: http://dx.doi.org/10.1080/00222933.2012.700333
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Journal of Natural HistoryVol. 46, Nos. 31–32, August 2012, 1959–1967
Social facilitation and food partitioning in the queenless ant Dinoponeraquadriceps (Hymenoptera: Formicidae)
Fabio S. Nascimentoa*, Danilo I.S.A. Souzab, Ivelize C. Tannure-Nascimentoa andJosé O. Dantasb
aUniversidade de São Paulo, FFCLRP, Departamento de Biologia, Ribeirão Preto, São Paulo,Brazil; bUniversidade Federal de Sergipe, CCBS, Departamento de Biologia, São Cristóvão,Sergipe, Brazil
(Received 27 April 2011; final version received 8 May 2012; printed 6 August 2012)
Social facilitation occurs when an animal is more likely to behave in a certain wayin response to other animals engaged in the same behaviour. For example, an indi-vidual returning to the nest with food stimulates other ants to leave and to forage.In the present study we demonstrate the existence of new facets in the colony orga-nization of Dinoponera quadriceps: a positive feedback between the incoming foodand the activation of new foragers, and the occurrence of incipient task partitioningduring the food sharing. Lower-ranked workers located inside the nest process pro-tein resources and higher-ranked workers handle smaller pieces and distribute themto the larvae. In conclusion, D. quadriceps has a decentralized pattern of task alloca-tion with a double regulatory mechanism, which can be considered a sophisticatedaspect of division of labour in ponerine ants.
Keywords: Ponerinae; division of labour; decentralized control; foraging
Introduction
Colony organization is one of the major aspects for the ecological success of the socialinsects (Wheeler 1928; Wilson 1971; reviewed in Beshers and Fewell 2001). A perma-nent coordination between interacting individuals regulates the small units of work.Therefore, division of labour is a fragmentary set of behavioural decisions in whichthe development of colony is regulated (Oster and Wilson 1978; Pacala et al. 1996;Bourke 1999; Anderson and McShea 2001; Arnan et al. 2011). In some ants, workersare specialized in a number of limited tasks and this specialization can be determinedby physiology and development (reviewed in Robinson 2009). Also, there are general-ist individuals that respond according to the available work in the colony (Bourke andFranks 1995).
The “poneromorph” ants are the large subfamily Ponerinae and several muchsmaller subfamilies that were until recently included in it (Bolton 2003; but seeWard 2007). They share a number of features, including weak worker specialization(Traniello 1978; Peeters 1997; Anderson and McShea 2001), but this phylogeneti-cally artificial group of ants has ecological meaning because the species includedshare predaceous non-migratory habits and hence impose similar ecological impacts
*Corresponding author. Email: [email protected]
ISSN 0022-2933 print/ISSN 1464-5262 online© 2012 Taylor & Francishttp://dx.doi.org/10.1080/00222933.2012.700333http://www.tandfonline.com
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

1960 F.S. Nascimento et al.
on other small animal fauna (Scott-Santos et al. 2008). However, social facilitationoccurs when an individual arriving with food at the nest stimulates other ants toleave and to forage (Wilson 1971; Gordon and Mehdiabadi 1999). This phenomenonwas observed in several species, including Ophthalmopone berthoudi, Odontomachusbauri, Ectatomma permagnum and Gnamptogenys moelleri (Peeters and Crewe 1987;Oliveira and Hölldobler 1989; Paiva and Brandão 1989; Cogni and Oliveira 2004).In Paraponera clavata, it was shown that the return of an individual from a bait siteresulted in greatly increased foraging activity from the colony (Breed and Bennett1985). Also, temporal learning can be observed in P. clavata and Ectatoma ruidum(Fewell et al. 1992; Schatz et al. 1994). In these species, ants were able to learn thetemporal information of food availability and to anticipate the time production of thefood sources (reviewed in Fourcassié et al. 1999).
Another aspect related to the worker behaviour is task partitioning (Jeanne 1986;reviewed in Ratnieks and Anderson 1999). Task partitioning occurs when a giventask is split into a number of sequential subtasks where any kind of material canbe transferred from one worker to another (e.g. wasps: Jeanne 1999; pseudoscorpi-ons: Tizo-Pedrozo and Del Claro 2011). This type of behaviour has been consideredcommon in only relatively complex societies, because individual task flexibility andthe queuing time are more likely to occur in smaller colonies than in larger colonies(reviewed in Ratnieks and Anderson 1999; Blonder and Dornhaus 2011).
Dinoponera quadriceps is a giant queenless ant in which division of labour is a resultof their social system, which comprises a linear dominance hierarchy headed by aninseminated worker or gamergate who monopolizes egg laying, leaving other workerswith the onus to perform all non-reproductive functions in the colony. These workerscan be classified into two distinct groups: lower-ranking workers that perform forag-ing activities, guard and nest maintenance; and higher-ranking workers that engage inhelper activities (Monnin and Peeters 1999). However, it is unknown how this kind ofsocial structure affects division of labour in this species.
Because only a few models of division of labour in ponerine ants have currentlybeen tested with empirical data (Theraulaz et al. 2002). It is important to demonstratethe transition from individual behaviour to colony-level behaviour through a set ofexperimental thresholds. This study shows that a difference in the response thresh-olds associated with the two kinds of stimuli (live prey and number of hungry larvae)is a sufficient condition to produce task partitioning among workers. Therefore weaddressed complementary questions about the division of labour in D. quadriceps.(1) Are foragers partitioning food after returning to the nest or are they feeding thelarvae themselves? (2) Which factors influence the decision of individuals to forage?
Materials and methods
Five entire D. quadriceps colonies were collected in São Cristóvão, Sergipe State, Braziland transferred to the Entomology Laboratory of Universidade Federal de Sergipe.Four colonies were used in this study (Colonies 5, 8, 10 and 11; mean number of adults52 ± 9.5, Table 1). Each individual colony was transferred to a plastic tray (35 × 25 ×6 cm) used as a nest and linked by a plastic tube to the foraging area (same size). Thenest tray was divided into six interconnected chambers and covered with a glass lid toallow direct observation. Colonies were maintained under 12: 12 h of light: dark andcontrolled temperature (24–26◦C).
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

Journal of Natural History 1961
Table 1. Composition of Dinoponera quadriceps colonies used in this study.
Numberof
females
Numberof eggs
Number of smalllarvae (1st, 2nd and
3rd instars)
Number of largelarvae (4th and 5th
instars)
Numberof pupae
Colony 5 61 35 4 10 3Colony 8 47 33 3 21 16Colony 10 59 31 15 3 5Colony 11 41 51 12 17 16
The ants were fed twice a week with freshly killed dead roaches, meal worm larvaeand synthetic ant diet (Bhatkar and Whitcomb 1970). Colonies were similarly moti-vated to forage before the experiments, and were starved for 24 h before trials. Waterwas offered ad libitum in Petri dishes. Each individual was marked with a paint codeto identify caste and functional roles.
Ranking of females was determined by analysing the pattern of dominance/subordinate interactions between individuals, and other worker activities, such asbrood caring, nest maintenance, guarding and foraging (Monnin and Peeters 1999).Only workers engaged in nurse behaviour on at least four occasions during a period ofobservation were included in the analysis. Individual task specialization was calculatedusing the Shannon–Wiener index (Lehner 1979; O’Donnell & Jeanne 1990) as:
H (x) = −∑
p (x) log 2 p(x),
where p(x) is the proportion of a worker’s effort devoted to a given task (x). Theresulting index ranges from 0 (low specialization) to 1 (high task specialization).
Observations were carried out for 21 days and only one period of observationper colony was carried out per day (n = 84 trials). Each period of observation lasted60 minutes per colony and started when the last larval instar of a mealworm was putinto the foraging arena. We observed from the moment a forager discovered the preyuntil the instant that it was offered to the larvae inside the nest. Experiments onlystarted after the ants had become used to the presence of the observer (raised antennaeand opening mandibles).
Before each experiment we counted the total number of adults, number of foragersin the foraging arena and brood (eggs, larvae and pupae). To know the proportion ofants taking part in the flow of food, during the experiments we recorded the numberof ants participating in the distribution of food (foraging, sharing food and feedinglarvae). We also noted the identity of each individual dividing and receiving a pieceof food, its location inside and outside the nest, and the final destination of the prey.To evaluate the effects of brood on the foraging process, we added one or two largelarvae from a fifth colony (colony 3) every 2 days to the experimental colonies. Residentants accepted the foreign larvae in all colonies.
We evaluated whether the number of foragers arriving at a possible protein foodsource changes during the period of observation by comparing the newcomer arrivalrates between the first and second hours of the experiment by using Wilcoxon matched-rank test (STATISTICA 7.0, Statsoft Inc., Tulsa, OK, USA). The number and identity
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

1962 F.S. Nascimento et al.
of receivers were noted to estimate the distribution of food from the outside to theinside of the nest.
Results
The number of ants engaged in food transference varied significantly among colonies.The relative proportion ranged from 7% to 76% of total number of ants partic-ipating in the food sharing in the four colonies (Kruskal–Wallis3,84: H = 30.08;p < 0.001). There was a significant correlation between the number of work-ers taking part in the flow of food and the number of large larvae in colonies5, 10 and 11 (Figure 1). During the experiments, we observed at colony levela significant task specialization of nurses (Table 2 Shannon–Wiener index (H):Colony 5, H = 0.8 ± 0.1; Colony 8, H = 0.6 ± 0.12; Colony 10, H =0.82 ± 0.12; Colony 11, H = 0.7 ± 0.2 (n = 21 observation periods for eachcolony).
In three of the four colonies, the proportion of foragers increased after food offer-ing. Colony 5 experienced a small, but significant variation in the number of activeforagers in the arena (before × after: 3.14 ± 2.10 × 5.5 ± 3.27 ants; Z = 2.65; p <
0.01; n = 14). Colonies 10 and 11 also experienced a rise in forager number after for-agers returned to the colony (Colony 10: before × after: 3.92 ± 3.26 × 6 ± 2.54 ants;
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22Days
0
5
10
15
20
25
30
Num
ber
of in
divi
dual
s
LarvaeWorkers
Figure 1. Pooled results of the association between workers partitioning food (dotted line) andfeeding larvae (solid line) during 21 days of experiment. To test the effects of brood on the forag-ing process, one or two additional fifth instar larvae were added every 2 days in the experimentalcolonies.
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

Journal of Natural History 1963
Table 2. Summary of results on task partitioning and social facilitation in Dinoponeraquadriceps.
Proportionof
specializedforagers∗
Proportionof
specializednurses∗
Multipletransfer-
ences beforefeedinglarvae†
Average proportion of idleforagers exiting the nest
after:
p value
return of asuccessful
forager
no return
Colony 5 0.21 (13) 0.08 (5) 10.1 ± 3.12 0.42 0.23 <0.001Colony 8 0.23 (11) 0.14 (7) 7.38 ± 2.25 0.54 0.27 <0.001Colony 10 0.20 (12) 0.13 (8) 7.22 ± 1.71 0.33 0.25 <0.05Colony 11 0.34 (14) 0.12 (5) 7.8 ± 6.34 0.5 0.28 0.06Total 0.24 0 8.12 0.44 0.25
∗Data shown as proportion (n).†Data shown as mean ± SD.
Z = 2.00; p < 0.05; Colony 11: before × after: 4.35 ± 4.08 × 7.00 ± 4.27; Z = 3.39;p < 0.001). The number of foragers in Colony 8 also increased after experimental preyoffering, but this difference was not significant (Colony 8: before × after: 2.14 ± 1.87 ×3.35 ± 1.78 ants; Z = 1.83; p = 0.06).
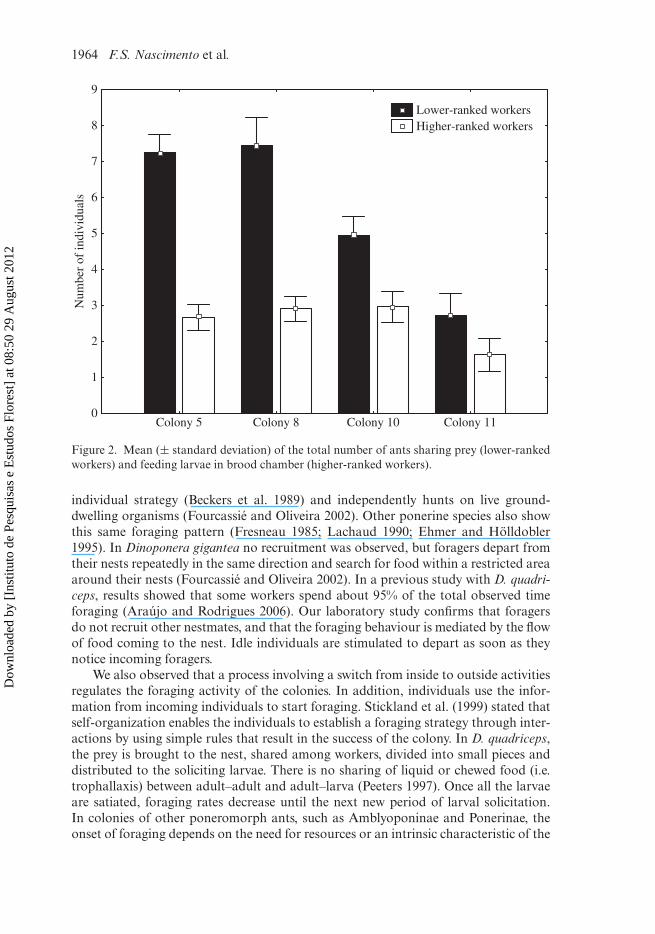
The results also showed that only a smaller proportion of specialized nurses fedthe larvae (Figure 2). The number of nurses varied from one to nine among colonies(mean ± SD: 2.75 ± 1.76 ants, n = 4), but the nurses made up a group that wassignificantly smaller than other ants taking part in the food flow (mean ± SD: 6.05 ±3.06 ants).
Discussion
In the present study we showed the existence of two new aspects in the colony organi-zation of D. quadriceps: first, a positive feedback between the incoming food and theactivation of new foragers, and second, the occurrence of an incipient task partitioningduring food sharing. Lower-ranking workers inside the nest process protein resourceswhile higher-ranking workers handle smaller pieces and distribute them among thelarvae.
In social insects, an interesting aspect is to understand how colonies allocateworkers in a particular task and how groups of individuals organize themselves intosmaller subgroups. Jeanne (1999) argued that social insects with more complex systemsof organization maximize task partitioning through worker specialization, thereforereducing queuing time of task performance and resulting in more per capita produc-tivity, whereas in smaller societies with no task partitioning, workers are more efficientby performing multiple tasks and functions.
Incipient task partitioning and specialization were also observed in other ponero-morph ants. Ectatomma ruidum workers can transfer insect prey to transporters(Schatz et al. 1996) and a few specialized Ectatomma opaciventre workers carryadult nestmates during nest migration. Usually, a Dinoponera forager exhibits an
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2
1964 F.S. Nascimento et al.
Colony 5 Colony 8 Colony 10 Colony 110
1
2
3
4
5
6
7
8
9N
umbe
r of
indi
vidu
als
Lower-ranked workersHigher-ranked workers
Figure 2. Mean (± standard deviation) of the total number of ants sharing prey (lower-rankedworkers) and feeding larvae in brood chamber (higher-ranked workers).
individual strategy (Beckers et al. 1989) and independently hunts on live ground-dwelling organisms (Fourcassié and Oliveira 2002). Other ponerine species also showthis same foraging pattern (Fresneau 1985; Lachaud 1990; Ehmer and Hölldobler1995). In Dinoponera gigantea no recruitment was observed, but foragers depart fromtheir nests repeatedly in the same direction and search for food within a restricted areaaround their nests (Fourcassié and Oliveira 2002). In a previous study with D. quadri-ceps, results showed that some workers spend about 95% of the total observed timeforaging (Araújo and Rodrigues 2006). Our laboratory study confirms that foragersdo not recruit other nestmates, and that the foraging behaviour is mediated by the flowof food coming to the nest. Idle individuals are stimulated to depart as soon as theynotice incoming foragers.
We also observed that a process involving a switch from inside to outside activitiesregulates the foraging activity of the colonies. In addition, individuals use the infor-mation from incoming individuals to start foraging. Stickland et al. (1999) stated thatself-organization enables the individuals to establish a foraging strategy through inter-actions by using simple rules that result in the success of the colony. In D. quadriceps,the prey is brought to the nest, shared among workers, divided into small pieces anddistributed to the soliciting larvae. There is no sharing of liquid or chewed food (i.e.trophallaxis) between adult–adult and adult–larva (Peeters 1997). Once all the larvaeare satiated, foraging rates decrease until the next new period of larval solicitation.In colonies of other poneromorph ants, such as Amblyoponinae and Ponerinae, theonset of foraging depends on the need for resources or an intrinsic characteristic of the
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

Journal of Natural History 1965
species, which in turn is probably influenced by colony size, because in small coloniesworkers tend to forage at an earlier age. Workers from larger colonies spent signifi-cantly longer periods of time foraging than those from small colonies (Thomas 2002),but larger colonies require more food to sustain their larvae and adults (Thomas andElgar 2003).
Ecological traits of ponerine ants such as hunting habits, prey specialization,behavioural flexibility and absence of recruitment in several species are consideredcharacteristics of primitive ants (Peeters and Crewe 1987; Peeters 1997). However,these features may result in a larger variety of morpho-physiological–behaviouraladaptations that enhance colony efficiency by reducing the time and energy of taskallocation in smaller societies (Dejean and Lachaud 2011).
In conclusion, this study shows that D. quadriceps have a decentralized pattern oftask allocation with a double regulatory mechanism, which can be considered a sophis-ticated aspect of division of labour in ants. In addition, the dynamics of hierarchyranking determine the work roles such as foraging, food distribution and brood care.
References
Anderson C, McShea DW. 2001. Individual versus social complexity, with particular referenceto ant colonies. Biol Rev. 76:211–237.
Araújo A, Rodrigues Z. 2006. Foraging behavior of the queenless ant Dinoponera quadricepsSantschi (Hymenoptera, Formicidae). Neotr Entomol. 35:159–164.
Arnan X, Ferrandiz-Rovira M, Pladevall C, Rodrigo A. 2011. Worker size-related task parti-tioning in the foraging strategy of a seed-harvesting ant species. Behav. Ecol. Sociobiol.65:1881–1890.
Beckers R, Goss S, Deneubourg JL, Pasteels JM. 1989. Colony size, communication and antforaging strategy. Psyche 96: 239–256.
Beshers SN, Fewell JH, 2001. Models of division of labor in social insects. Annu Rev Entomol46:413–430.
Bhatkar AP, Whitcomb WH. 1970. Artificial diet for rearing various species of ants. FloridaEntomol. 53:229–232.
Blonder B, Dornhaus A. 2011. Time-ordered networks reveal limitations to information flow inant colonies, PLoS One 6: e20298.
Bolton B. 2003. Synopsis and classification of Formicidae. Mem Am Mus Entomol Inst.71:1–370.
Bourke A. 1999. Colony size, social complexity and reproductive conflict in social insects. J EvolBiol 12:245–257.
Bourke AFG, Franks NR. 1995. Social Evolution in Ants. Princeton (NJ): Princeton UniversityPress, 529 pp.
Breed MD, Bennett B. 1985. Mass recruitment to nectar sources in Paraponera clavata: a fieldstudy. Insectes Soc. 32:198–208.
Cogni R, Oliveira PS. 2004. Patterns in foraging and nesting ecology in the Neotropical antGnamptogenys moelleri (Formicidae, Ponerinae). Insectes Soc. 51:123–130.
Dejean A, Lachaud J-P. 2011. The hunting behavior of the African ponerine ant Pachycondylapachyderma. Behav Proc. 86:169–173.
Ehmer B, Hölldobler B. 1995. Foraging behavior of Odontomachus bauri on Barro ColoradoIsland, Panama. Psyche 102:215–224.
Fewell JH, Harrison JF, Stiller TM, Breed MD. 1992. A cost benefit analysis of distanceeffects on foraging and recruitment in the giant tropical ant, Paraponera clavata. Oecologia92:542–547.
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

1966 F.S. Nascimento et al.
Fourcassié V, Henriques A, Fontella C. 1999. Route fidelity and spatial orientation in the antDinoponera gigantea (Hymenoptera, Formicidae) in a primary forest: a preliminary study.Sociobiology 34:505–524.
Fourcassié V, Oliveira PS. 2002. Foraging ecology of the giant Amazonian ant Dinoponera gigan-tea (Hymenoptera: Formicidae: Ponerinae): activity schedule, diet, and special foragingpatterns. J Nat His. 36:2211–2227.
Fresneau D. 1985. Individual foraging path fidelity: a novel strategy in a ponerine ant. InsectesSoc. 32:109–116.
Gordon DM, Mehdiabadi NJ. 1999. Encounter rate and task allocation in harvester ants. BehavEcol Sociobiol. 45:370–377.
Jeanne RL. 1986. The organization of work in Polybia occidentalis: costs and benefits ofspecialization in a social wasp. Behav Ecol Sociobiol. 19:333–341.
Jeanne RL. 1999. Group size, productivity, and information flow in social wasps. In: DetrainC, Pasteels JM, Deneubourg JL. editors. Information Processing in Social Insects. Basel:Birkhäuser; p. 3–30.
Lachaud JP. 1990. Foraging activity and diet in some neotropical ponerine ants I. Ectatommaruidum Roger (Hymenoptera Formicidae). Folia Entomol Mexicana 78:241–256.
Lehner PN. 1979. Handbook of Ethological Methods. New York (NY): Garland STPM Press.Monnin T, Peeters C. 1999. Dominance hierarchy and reproductive conflicts among subordi-
nates in a monogynous queenless ant. Behav Ecol. 10:323–332.O’Donnell S, Jeanne RL. 1990. Forager specialization and the control of nest repair in Polybia
occidentalis Oliver (Hymenoptera: Vespidae). Behav Ecol Sociobiol. 27:359–364.Oliveira PS, Hölldobler B. 1989. Orientation and communication in the Neotropical ant
Odontomachus bauri Emery (Hymenoptera, Formicidae, Ponerinae). Ethology 83:154–166.Oster GF, Wilson EO. 1978. Caste and ecology in the social insects. Princeton (NJ): Princeton
University Press. 352 pp.Pacala SW, Gordon DM, Godfray HCJ. 1996. Effects of group size on information transfer and
task allocation. Evol Ecol. 10:127–165.Paiva RVS, Brandão CRF. 1989. Estudos sobre a organização social de Ectatomma Permagnum
Forel, 1908 (Hymenoptera: Formicidae). Rev Bras Biol. 49:783–792.Peeters C. 1997. Morphologically primitive ants: comparative review of social characters, and
the importance of queen-worker dimorphism. In: Choe J, Crespi B. editors. The evolu-tion of social behaviour in insects and arachnids. Cambridge: Cambridge University Press;p. 372–391.
Peeters C, Crewe RM. 1987. Foraging and recruitment in ponerinae ants: solitary hunting in thequeenless Ophthalmopone berthoudi (Hymenoptera: Formicidae). Psyche 94:201–214.
Ratnieks FLW, Anderson C. 1999. Task partitioning in insect societies. Insectes Soc. 46:95–108.Robinson EJH. 2009. Physiology as a caste-defining feature. Insectes Soc. 56:1–6.Schatz B, Beugnon G, Lachaud JP. 1994. Time–place learning by an invertebrate, the ant
Ectatomma ruidum Roger. Anim Behav. 48:236–238.Schatz B, Beugnon G, Lachaud JP. 1996. Polyethism within hunters of the ponerine ant,
Ectatomma ruidum Roger (Hymenoptera, Ponerinae). Insectes Soc. 43:111–118.Scott-Santos CP, Esteves FA, Brandão CRF. 2008. Catalogue of “poneromorph” ant type spec-
imens (Hymenoptera, Formicidae) deposited in the Museu de Zoologia da Universidadede São Paulo, Brazil. Pap Avulsos Zool (São Paulo) 50:693–699.
Stickland TR, Britton NF, Franks NR. 1999. Models of information flow in ant foraging: thebenefits of both attractive and repulsive signals. In: Detrain C, Deneubourg JL, PasteelsJM. editors. Information Processing in Social Insects. Basel: Birkhäuser; p. 83–100.
Theraulaz G, Bonabeau E, Sole RV, Schatz B, Deneubourg JL. 2002. Task partitioning in aponerine ant. J Theor Biol. 215:481–489.
Thomas ML. 2002. Nest site selection and longevity in the ponerine ant Rhytidoponera metallica(Hymenoptera: Formicidae). Insectes Soc. 49:147–152.
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2

Journal of Natural History 1967
Thomas ML, Elgar MA. 2003. Colony size affects division of labour in the ponerine antRhytidoponera metallica. Naturwissenschaften 90:88–92.
Tizo-Pedroso E, Del-Claro K. 2011. Is there division of labor in cooperative pseudoscorpions?An analysis of the behavioral repertoire of a tropical species. Ethology 117:498–507.
Traniello JFA. 1978. Caste in a primitive ant: absence of age polyethism in Amblyopone. Science202:770–772.
Ward PS. 2007. Phylogeny, classification, and species-level taxonomy of ants (Hymenoptera:Formicidae). Zootaxa 1668:549–563.
Wheeler WM. 1928. The social insects: their origin and evolution. London: Kegan Paul, Trench,Trubner and Co. Ltd. 378 pp.
Wilson EO. 1971. The insect societies. Cambridge (MA): Belknap Press of Harvard UniversityPress.
Dow
nloa
ded
by [
Inst
ituto
de
Pesq
uisa
s e
Est
udos
Flo
rest
] at
08:
50 2
9 A
ugus
t 201
2




















