
Taxonomic revision of 14 south-western European species of Isotomurus (Collembola, Isotomidae), with...
29
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30 , 2, April 2001, pp115–143 115 Carapelli, A., Frati, F., Fanciulli, P. P. & Dallai, R. (2001). Taxonomic revision of 14 south- western European species of Isotomurus (Collembola, Isotomidae), with description of four new species and the designation of the neotype for I. palustris . — Zoologica Scripta , 30 , 115 –143. A detailed study of morphological characters in populations of the collembolan genus Isotomurus from southern Europe is presented. Isotomurus is a cosmopolitan genus in which species diagnosis primarily relies on pigmentation patterns and chaetotaxy. Correct diagnosis is often complicated by the high variability of these characters. This has created confusion in the systematics of the genus and has led to descriptions of several varieties, colour forms and subspecies, the taxonomic value of which needs revision. Complete screening of pigmentation patterns and the study of sensorial chaetotaxy in specimens collected in 37 localities across south-western France, Italy and Switzerland have enabled 14 species to be distinguished; one taxon is elevated to the rank of species ( Isotomurus prasinus ) and four new species are described: I. hadriaticus , I. aetnensis , I. gallicus and I. pseudopalustris . The neotype for I. palustris is designated in order to clarify the taxonomic status and the type locality for this species whose topotypic material is lost. A redescription of previously known species on the basis of the newly introduced characters is also presented, together with a key to the south-western European species of Isotomurus . Following this study, the two species I. indipendente and I. italicus came to be junior synonyms of I. palustris and I. fucicolus , respectively. A cladistic analysis of 14 morphological characters provides a phylogenetic reconstruction which is largely concordant with previous taxonomic works. Antonio Carapelli, Francesco Frati, Pietro Paolo Fanciulli & Romano Dallai, Department of Evolutionary Biology, University of Siena, Via P. A. Mattioli 4, 53100 Siena, Italy. E-mail: [email protected] Blackwell Science, Ltd Taxonomic revision of 14 south-western European species of Isotomurus (Collembola, Isotomidae), with description of four new species and the designation of the neotype for I. palustris A NTONIO C ARAPELLI , F RANCESCO F RATI , P IETRO P AOLO F ANCIULLI & R OMANO D ALLAI Accepted: 26 October 2000 Introduction Isotomurus is a cosmopolitan genus of hemiedaphic insects associated with forest litter and wet environments. Since the first description of I. palustris (Müller 1776), more than 50 species from all over the world have been described in the genus. In spite of some attempts to introduce new diagnostic characters, the taxonomy of the genus is still problematic. According to previous descriptions, species of the genus Isotomurus are distinguished on the basis of a few taxonomically significant characters, such as pigmentation pattern and the distribution of sensorial chaetotaxy (Poinsot-Balaguer 1976; Deharveng & Lek 1993; Lek & Carapelli 1998). Nevertheless, it is generally believed that the presence of many colour varieties is a common feature of the genus. Stach (1947) reviewed the taxonomy of the genus and pointed out that the European species are characterized by constant morphological traits. Body tergites are clothed with numerous short setae and several long macrochaetae; thin trichobothria are found on abdomen (abd.) II, III and IV, whereas the mucro is always quadridentate with a mucronal seta ‘usually present’. Stach (1947) described the species I. palustris as comprising several forms with different pigmentation patterns: some have a uniform pattern (such as the forms prasina and fucicola ), whereas others have a longitudinal median stripe (forms unifasciata and maculata ). Poinsot-Balaguer (1972) studied specimens of I. palustris from southern France and observed a constant trichobothrial formula characterized by 3 + 3, 3 + 3 and 1 + 1 trichobothria on abd. II, III and IV, respectively. She described the subspecies I. palustris palustris and I. palustris maculatus which were distin- guished by the different pigmentation patterns and the ciliation of abdominal macrochaetae (slightly more pronounced in I. palustris maculatus ). She also pointed out that the pigmentation pattern and trichobothria arrangement on abdominal tergites are useful characters for species diagnosis. Furthermore, she described the peculiar trichobothrial formula of I. fucicola (Reuter 1891) and studied the development of sensorial chaetotaxy in the first instar of I. palustris palustris . Later, Poinsot-Balaguer & Ferard
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Taxonomic revision of 14 south-western European species of Isotomurus (Collembola, Isotomidae), with...

© The Norwegian Academy of Science and Letters • Zoologica Scripta,
30
, 2, April 2001, pp115–143
115
Carapelli, A., Frati, F., Fanciulli, P. P. & Dallai, R. (2001). Taxonomic revision of 14 south-western European species of
Isotomurus
(Collembola, Isotomidae), with description of fournew species and the designation of the neotype for
I. palustris
. —
Zoologica Scripta
,
30
, 115–143.A detailed study of morphological characters in populations of the collembolan genus
Isotomurus
from southern Europe is presented.
Isotomurus
is a cosmopolitan genus in which species diagnosisprimarily relies on pigmentation patterns and chaetotaxy. Correct diagnosis is often complicatedby the high variability of these characters. This has created confusion in the systematics of thegenus and has led to descriptions of several varieties, colour forms and subspecies, the taxonomicvalue of which needs revision. Complete screening of pigmentation patterns and the study ofsensorial chaetotaxy in specimens collected in 37 localities across south-western France, Italyand Switzerland have enabled 14 species to be distinguished; one taxon is elevated to the rankof species (
Isotomurus prasinus
) and four new species are described:
I. hadriaticus
,
I. aetnensis
,
I.gallicus
and
I. pseudopalustris
. The neotype for
I. palustris
is designated in order to clarify thetaxonomic status and the type locality for this species whose topotypic material is lost. Aredescription of previously known species on the basis of the newly introduced characters isalso presented, together with a key to the south-western European species of
Isotomurus
.Following this study, the two species
I. indipendente
and
I. italicus
came to be junior synonymsof
I. palustris
and
I. fucicolus
, respectively. A cladistic analysis of 14 morphological characters providesa phylogenetic reconstruction which is largely concordant with previous taxonomic works.
Antonio Carapelli, Francesco Frati, Pietro Paolo Fanciulli & Romano Dallai, Department of EvolutionaryBiology, University of Siena, Via P. A. Mattioli 4, 53100 Siena, Italy. E-mail: [email protected]
Blackwell Science, Ltd
Taxonomic revision of 14 south-western European species of
Isotomurus
(Collembola, Isotomidae), with description of four new species and the designation of the neotype for
I. palustris
A
NTONIO
C
ARAPELLI
, F
RANCESCO
F
RATI
, P
IETRO
P
AOLO
F
ANCIULLI
& R
OMANO
D
ALLAI
Accepted: 26 October 2000
Introduction
Isotomurus
is a cosmopolitan genus of hemiedaphic insectsassociated with forest litter and wet environments. Since thefirst description of
I. palustris
(Müller 1776), more than 50species from all over the world have been described in thegenus. In spite of some attempts to introduce new diagnosticcharacters, the taxonomy of the genus is still problematic.According to previous descriptions, species of the genus
Isotomurus
are distinguished on the basis of a few taxonomicallysignificant characters, such as pigmentation pattern and thedistribution of sensorial chaetotaxy (Poinsot-Balaguer 1976;Deharveng & Lek 1993; Lek & Carapelli 1998). Nevertheless,it is generally believed that the presence of many colourvarieties is a common feature of the genus.
Stach (1947) reviewed the taxonomy of the genus andpointed out that the European species are characterized byconstant morphological traits. Body tergites are clothed withnumerous short setae and several long macrochaetae; thintrichobothria are found on abdomen (abd.) II, III and IV,
whereas the mucro is always quadridentate with a mucronalseta ‘usually present’. Stach (1947) described the species
I. palustris
as comprising several forms with different pigmentation patterns:some have a uniform pattern (such as the forms
prasina
and
fucicola
), whereas others have a longitudinal median stripe(forms
unifasciata
and
maculata
).Poinsot-Balaguer (1972) studied specimens of
I. palustris
from southern France and observed a constant trichobothrialformula characterized by 3 + 3, 3 + 3 and 1 + 1 trichobothriaon abd. II, III and IV, respectively. She described the subspecies
I. palustris palustris
and
I. palustris maculatus
which were distin-guished by the different pigmentation patterns and the ciliationof abdominal macrochaetae (slightly more pronounced in
I.palustris maculatus
). She also pointed out that the pigmentationpattern and trichobothria arrangement on abdominal tergites areuseful characters for species diagnosis. Furthermore, she describedthe peculiar trichobothrial formula of
I. fucicola
(Reuter 1891)and studied the development of sensorial chaetotaxy in the firstinstar of
I. palustris palustris
. Later, Poinsot-Balaguer & Ferard
ZSC055.fm Page 115 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of
Isotomurus
•
A. Carapelli
et al.
116
Zoologica Scripta,
30
, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
(1983) redescribed
Isotomurus balteatus
(Reuter 1876) fromspecimens collected in southern France, providing twoalternative trichobothrial formulas and suggesting that
I.balteatus
be considered a species group like
I. palustris
.Cassagnau (1987) studied several populations of
I. palustris
from south-western France and confirmed the reliability ofpigmentation patterns for the taxonomic identification of thevarieties
prasina
,
hortorum
,
palustris
,
maculata
and
catalana
.Due to the presence of so many colour varieties, he suggested thatthe colour pattern might be a good diagnostic criterion if its fluc-tuations were observed in a large number of specimens, butconcluded that interbreeding experiments would be necessaryto confirm its utility for species diagnosis.
Deharveng & Lek (1993) described the ‘S’ chaetotaxy of abd.V in nine
Isotomurus
species, and discussed the taxonomicinterest of a new set of morphological characters definingfive different groups of species:
I. alticolus
,
I. balteatus
,
I. cassa-gnaui
,
I. palustris
and
I. sensillatus
.Finally, Christiansen & Bellinger (1998) have proposed
four different types of dental rugosity on the dorsal side ofdens of the Nearctic species of the genus.
Two independent genetic markers (allozyme electrophoresisand DNA sequences) were recently used to support morpho-logical analysis in order to establish the correct taxonomic rankof several varieties or subspecies of the
I. palustris
group(Carapelli
et al
. 1995a,b; Frati
et al
. 1995). In particular, the fixationof alternative alleles between sympatric individuals showingdifferent pigmentation patterns and the levels of sequencedivergence in one mitochondrial and one nuclear gene confirmed,on molecular grounds, that these forms can be consideredwell-differentiated species (e.g.
I. palustris
and
I. maculatus
:Carapelli
et al
. 1995a), as already proposed by Börner (1901).In the present study, we investigated several morphological
characters of taxonomic interest in more populations ofthe genus
Isotomurus
collected in 37 localities from Italy,south-western France and Switzerland.
Materials and methods
We collected specimens of
Isotomurus
in 37 different localitiesof the Italian peninsula and islands, Switzerland and southernFrance (Fig. 1A –D). They were collected using mechanicalaspirators in forest litter, mosses and stream banks. Sorting ofspecimens was aided by coupling with parallel biochemicaland molecular analyses (Carapelli
et al
. 1995a,b). The latterapproaches were used to support morphological identification,especially when more than one species living sympatrically wassampled simultaneously.
Type material for all new species is preserved in the collectionof Professor Romano Dallai at the Department of EvolutionaryBiology of the University of Siena. Neotypes for
I. palustris
are preserved in the entomological collection at the ZoologicalMuseum of the University of Copenhagen.
The cladistic analysis was conducted using the programs
PAUP
* (Swofford 1998) and MacClade (version 3.01; Maddison& Maddison 1992). Phylogenetic reconstructions were gen-erated by parsimony analysis of the morphological data setusing branch and bound searches, with all characters weightedequally. Character state transformations were designated asunordered and unknown character states in some species weretreated as missing data.
Abbreviations used in figures and text
abd
., abdomen
al
, anterolateral setae
ant
., antennal segment
as
, anterosubmedial setae
accp
, accessory setae to the ‘p’ row of ordinary setae
cc
, cuticular crenulation of dorsal part of dens
dcA
, diameter of A corneola
L
, length of body
ms
, microsensillum
mso
, multiperforated sensorial organ
PAO
, postantennal organ
thr
, thorax
Description of species
(For the differential analysis of each species, see the dichot-omic key in Table 2).Family ISOTOMIDAE Börner, 1913Genus
Isotomurus
Börner, 1903Type species fixed:
Isotomurus palustris
(Müller, 1776)
Isotomurus aetnensis
sp. n.
Holotype.
Male, ITALY, Muto near Mt. Etna (Catania, Sicily),1000 m, Mediterranean shrub, 12 September 1995, preservedin the R. Dallai collection at the Department of EvolutionaryBiology, University of Siena.
Paratypes.
Same data as for holotype, one male and three females.
Etymology.
The name
aetnensis
is derived from the firstcollection site near Mt. Etna, where the species appears to beendemic.
Diagnosis.
This species can be distinguished by the presence ofdarker ribbon-like patches of pigmentation on the edges oftergites (especially abd. III–V) and by a dark brownish back-ground colour. Legs are uniformly coloured like tergites.
Description
Habitus.
Body slender, habitus of
Isotoma
. Body length: 2–3 mm.
Coloration.
Body brownish with darker ribbon-like patchesof pigmentation on the margins of tergites (especially abd.III–V ) and two spots on the lateral sides of abd. VI. In somespecimens, there is a very slight longitudinal median band,not visible to the naked eye. A patch of pigmentation is visible
ZSC055.fm Page 116 Friday, May 25, 2001 3:44 PM

A. Carapelli
et al.
•
Revision of south-western European species of
Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta,
30
, 2, April 2001, pp115–143
117
on the central part of the head and at the base of each antenna. Ant.III and IV slightly darker. Legs uniformly coloured like tergites.
Antennae.
Typical of the
palustris
group. Relative lengths ofthe antennal segments I : II : III : IV are 40 : 68 : 72 : 80.
Head.
8 + 8 eyes. PAO elongated. PAO/dcA: 0.6. Labrumwith 3 parallel rows of 5, 5 and 4 setae, respectively. Labiumwith 7–9 setae on each part of submentum. The maxillaryouter lobe is bipartite and with 4 sublobal hairs.
Chaetotaxy.
Ordinary setae abundant with ciliated meso-and macrochaetae. 3 + 3 multilaterally ciliated macrochaetaeon both abd
.
IV and V, arranged in a triangular fashion andin a row, respectively. Trichobothrial formula typical of the
palustris
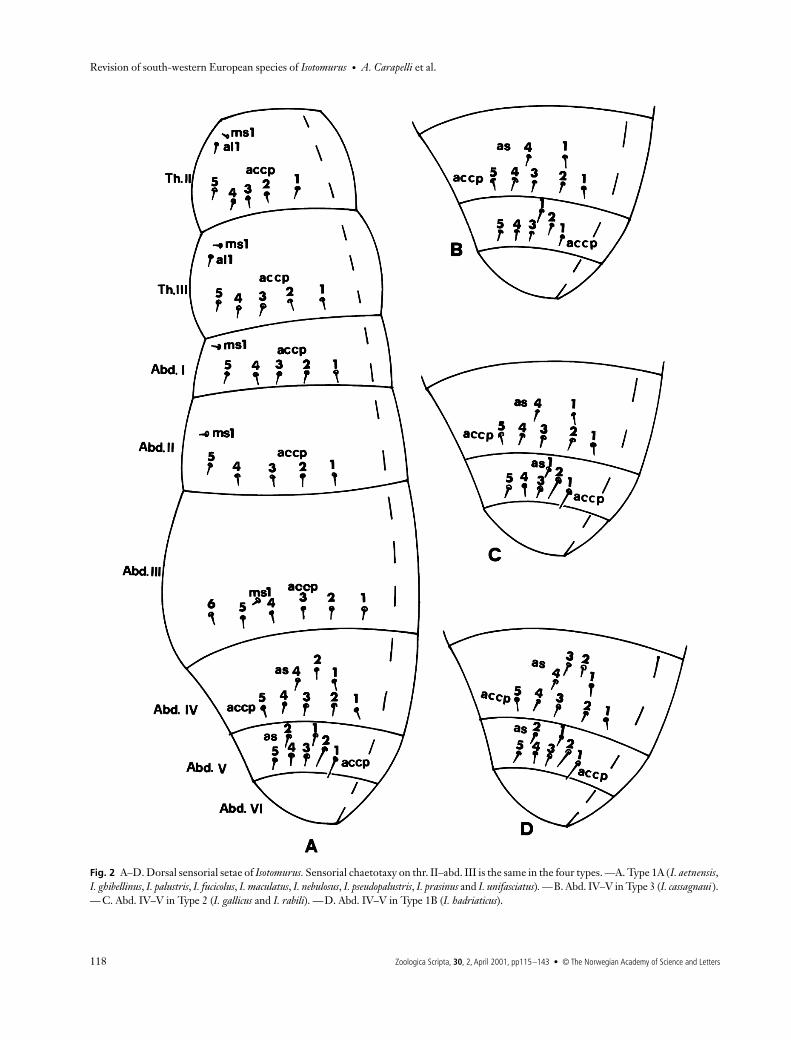
group (3 + 3 + 1). The distribution of ‘S’ setae istypical of Type 1A (Fig. 2A): ms1 + al1 + accp1–5 on thr
.
IIand III; ms1 + accp1–5 on abd
.
I and II; ms1 + accp1–6 onabd. III; as1, 2 and 4 + accp1–5 on abd. IV; as1–2 + accp1–5(accp1 and accp2 longer than the others) on abd. V.
L/antennae.
2.3.
Ventral tube.
Lateral flaps with 3 + 3 setae.
Legs.
Claw with outer and lateral teeth. Empodial append-age frequently armed with a tooth. Tibiotarsus III with 4multilaterally ciliated macrochaetae.
Furca.
Retinaculum with 10–15 ordinary setae. Dorsalchaetotaxy of manubrium is composed of several ordinary,slightly ciliated setae, which are longer near the distal margin.The ventral side is completely covered by several setae, withthe exception of a median thin longitudinal stripe, and it alsobears 2 longer and ciliated microchaetae. Dentes are dorsallycovered by many setae, each one inserted in a papilla-likecuticular structure. On the ventral side, setae are less abundantand limited to the anterior part; two dorsal sensilla are foundnear the middle part of each dens. Lateral margins of denteswith longer ciliated setae; cc of type ‘A’. Mucro is quadridentateand without mucronal seta.
Fig. 1 A–D. Distribution maps of the sites where the 14 species of the genus Isotomurus studied in this work were sampled. Additional specimens of I. fucicolus and I. prasinus from Norway have been studied.
ZSC055.fm Page 117 Friday, May 25, 2001 3:44 PM
Revision of south-western European species of
Isotomurus
•
A. Carapelli
et al.
118
Zoologica Scripta,
30
, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
Fig. 2 A–D. Dorsal sensorial setae of Isotomurus. Sensorial chaetotaxy on thr. II–abd. III is the same in the four types. —A. Type 1A (I. aetnensis, I. ghibellinus, I. palustris, I. fucicolus, I. maculatus, I. nebulosus, I. pseudopalustris, I. prasinus and I. unifasciatus). —B. Abd. IV–V in Type 3 (I. cassagnaui ). — C. Abd. IV–V in Type 2 (I. gallicus and I. rabili). —D. Abd. IV–V in Type 1B (I. hadriaticus).
ZSC055.fm Page 118 Friday, May 25, 2001 3:44 PM

A. Carapelli
et al.
•
Revision of south-western European species of
Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta,
30
, 2, April 2001, pp115–143
119
Distribution and habitat.
Specimens were sampled in a grassfield nearby Mt. Etna.
Isotomurus alticolus
(Carl, 1899)
Isotoma alticola
Carl, 1899: 305, Switzerland.
Isotomurus alticolus
Handschin, 1919: 79, Switzerland.
Isotomurus alticolus
Deharveng & Lek, 1993: 255 –256, France.
Type locality.
Switzerland.
Material examined.
Several specimens from Guttanen andOberaar (Switzerland).
Description
Habitus.
Habitus of
Isotoma
. Body length: 1.8 – 2 mm. Thoracictergites narrow; antennae, claws and mucro elongated.
Coloration.
Body with yellowish or greenish background; adark longitudinal stripe occupies about one-third of eachtergite. Scattered pigmentation is also present on the lateralside of tergites. Ant. III–IV are slightly darker than the others.Central part of head and bases of antennae with irregularpigmented patches.
Head.
8 + 8 eyes. Postantennal organ large. PAO/dcA: 1.6 –1.8.Setae on labrum arranged in 3 rows of 5, 5 and 4 mesochaetae,respectively. Labium with 8 –12 setae on each part of submentum.The maxillary lobe is bipartite and with 4 sublobal hairs.
Chaetotaxy.
Ordinary setae very abundant; 3 multilaterallyciliated macrochaetae on each side of tergites are arranged ina triangular fashion on abd. IV, and in a row on abd. V. Tri-chobothrial formula typical of the
palustris
group (3 + 3 + 1).‘S’ setae arrangement sensu Deharveng (1979) (Type 5):accp1–5, al1 and two small setae on thr. II, which are prob-ably two microsensilla; accp1–6 and al1 on thr. III (no dataare available about the possible occurrence of microsensilla);accp1–6 and al1 on abd. I; seven setae (probably five accpsetae and two microsensilla) on abd. II; 10 and 24 setae diffi-cult to classify on abd. III and IV; accp1–8, as1, as3 and as4on abd. V.
L/antennae. 1.9.Legs. Claw slender; 3 ciliated macrochaetae are found on
tibiotarsus III. A longitudinal small cuticular dimple, runningfrom the distal to the median part of the tibiotarsus, is observedin all legs. Claw with outer and lateral teeth. Empodialappendage with or without tooth.
Ventral tube. Lateral flaps with 11 + 11 to 20 + 20 setae.Furca. Retinaculum with 15–18 ordinary setae. Manubrium
with several microchaetae of reduced dimensions on theventral side, and with longer ciliated setae on the dorsal one.Dentes dorsally covered with many setae inserted in littlecuticular bulges, and ventrally with a reduced number ofmicrochaetae. Lateral margins of dentes with longer ciliatedsetae; cc of type ‘A’. Dorsally, two microsensilla are presenton each dens. Mucro slender and quadridentate, mucronalseta present.
Distribution and habitat. The species has a Palearctic dis-tribution. Hygrophilic specimens, living on damp stones andmosses of Alpine brooks.
Isotomurus cassagnaui Deharveng & Lek, 1993Isotomurus cassagnaui Deharveng & Lek, 1993: 256 –257, France.
Type locality. Hautes Pyrénées, France.Material examined. Several specimens from Source de la Maure
(Pyrénées Mountains, France).
DescriptionHabitus. Habitus of Isotoma. Body length: 1.2 –1.4 mm.
Coloration. Uniform brown in colour, with violet antennaeand a blue spot on each side of abd. VI. Bases of antennae andcentral part of the head pigmented.
Antennae. Relatively short. Antennal segments ratio I : II :III : IV is 18 : 27 : 30 : 35.
Head. 8 + 8 eyes. PAO large if compared with the averageof the genus. PAO/dcA: 0.8 –1. Labrum with 3 parallel rowsof 5, 5 and 4 setae, respectively. Labium with 4 – 6 setae oneach part of submentum. The maxillary lobe is bipartite andwith 4 sublobal hairs.
Chaetotaxy. Ordinary setae abundant; micro- and mesocha-etae smooth. Macrochaetae ciliated and distributed in a tri-angular fashion on abd. IV and in a row on abd. V, and slightlyunilaterally ciliated. Trichobothrial formula typical of thepalustris group (3 + 3 + 1). The distribution of ‘S’ setae is typ-ical of Type 3 (Fig. 2B): ms1 + al1 + accp1–5 on thr. II and III;ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III;as1 and 4 + accp1–5 on abd. IV; as1 + accp1–5 (equally short)on abd. V. Some modified setae on ventrolateral region ofabd. III and IV in male.
L/antennae. 3.5– 4.0.Ventral tube. Each of the two lateral flaps of the ventral tube
has 3 setae.Legs. Claw with outer and lateral teeth. Empodial append-
age generally without tooth. Tibiotarsal macrochaetae smoothand slightly bigger than ordinary setae, but more pronouncedon tibiotarsus III.
Furca. Retinaculum with 5 –8 ordinary setae. Manubriumdorsally covered by several micro- and mesochaetae mostlypresent on the distal margin; ventrally, the setae are shorterand distributed everywhere except for the median part. Onthe ventral side of the manubrium, 2 longer setae are present.Dentes provided with many ordinary setae on the dorsal sideand with very few short setae on the ventral side; setae arelonger on the lateral margins; cc of type ‘A’. Setae are insertedin circular bulges of the dental cuticle. Dorsally, two micro-sensilla are present on each dens. Mucro is quadridentate andwithout mucronal seta.
Distribution and habitat. Hygrophilic species mostly presenton mosses of central part of Pyrénées Mountains.
ZSC055.fm Page 119 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
120 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
Isotomurus fucicolus (Reuter, 1891)Isotoma palustris fucicola Reuter, 1891: 226, Siberia.Isotomurus palustris fucicola Axelson, 1906: 131, Russia.Isotomurus fucicola Da Gama, 1964: 211, near Coimbra, Por-tugal, non Reuter, 1891.Isotomurus italicus Carapelli et al., 1995b: 94, fig. 2c. syn. nov.
Type locality. Siberia.Material examined. Several specimens from Allone (Perugia,
Italy), Circeo (Latina, Italy), Gerfalco (Grosseto, Italy),Giglio Island (Italy), Radi (Siena, Italy), Sorra (Siena, Italy)and Tjöme (Norway). Five specimens from the originalcollection (Reuter 1891; Finland Museum of Natural History,University of Helsinki).
DescriptionHabitus. Body slender, habitus of Isotoma. Body length:1.5–2 mm.
Coloration. Body uniformly brownish or yellowish in col-our. Irregular patches at the bases of antennae and behind theocular plates. Abd. VI with darker lateral spots.
Antennae. Antennae of normal length. Relative lengths ofant. I : II : III : IV are 21 : 33 : 36 : 45.
Head. 8 + 8 eyes. Postantennal organ elongated. PAO/dcA:1.5. Labrum with 3 parallel rows of 5, 5 and 4 setae, respectively.Labium with 5– 8 setae on each part of submentum. Themaxillary outer lobe is bipartite and with 4 sublobal hairs.
Chaetotaxy. Ordinary setae abundant; mesochaetae andmacrochaetae very slightly ciliated. 3 + 3 macrochaetae oneach hemitergite of abd. IV and V are distributed in a tri-angular fashion and in a row, respectively. Trichobothrialformula typical of the palustris group (3 + 3 + 1). Thedistribution of ‘S’ setae is typical of Type 1A (Fig. 2A):ms1 + al1 + accp1–5 on thr. II and III; ms1 + accp1–5 on abd.I and II; ms1 + accp1–6 on abd. III; as1, 2 and 4 + accp1–5 onabd. IV; as1–2 + accp1–5 (accp1 and accp2 longer than theothers) on abd. V.
L/antennae. 2.2.Ventral tube. Lateral flaps with 3 + 3 setae.Legs. Claw with outer and lateral teeth. Empodial appendage
with or without tooth. Tibiotarsus III with 3 –5 smooth mac-rochaetae.
Furca. Retinaculum with 5–7 ordinary setae. Manubriumwith many short setae and two longer ones on dorsal side, andwith several ventral mesochaetae, which are mostly distributedon the distal margin. Dentes dorsally with many short setaeand with a reduced number of microchaetae; setae are longeron the lateral margins; cc of type ‘A’. Setae are inserted incircular bulges of the dental cuticle. Dorsally, two microsensillaare present on each dens. Mucro quadridentate without seta.
Distribution and habitat. Species holoarctic, with one recordon the Bismark Archipelago. Specimens are usually found inforest litter, meadows and even in cultivated farmland.
Isotomurus gallicus sp. n.Isotomurus balteatus Poinsot-Balaguer & Ferard, 1983: 151–156, pl. I–II, France, non Reuter, 1876.Isotomurus balteatus Deharveng & Lek, 1993: 250, 255,table 1, non Reuter, 1876.
Holotype. Male, FRANCE, Tarn: Larroque, Forest ofGrésigne, 15 May 1994, preserved in the R. Dallai collectionat the Department of Evolutionary Biology, University ofSiena.
Paratypes. Same data as for holotype, two males and onefemale.
Etymology. The name gallicus is derived from the Latinname for France (Gallia), where the species appears to beendemic.
Diagnosis. This species can be distinguished by the presenceof a transversal dark band on the proximal part of tergites andby the trichobothrial formula 0 + 1 + 1 on abd. I to III.
DescriptionHabitus. Body short and thin, habitus of Isotoma. Body length:1.3–1.4 mm.
Coloration. Background whitish or yellowish. Anterior partof tergites with transverse dark band, more or less diffuse onthe rest of segment. Central part of head and bases of anten-nae pigmented.
Antennae. Ant. I, II and III very short. Length of antennalsegments I : II : III : IV are 11 : 17 : 18 : 24.
Head. 8 + 8 eyes. Postantennal organ very large if com-pared with other species of the genus. PAO/dcA: 1.6. Labrumwith 3 parallel rows of 5, 5 and 4 setae, respectively. Labiumwith 5–7 setae on each part of submentum. The maxillaryouter lobe is bipartite and with four sublobal hairs.
Chaetotaxy. Ordinary setae reduced if compared with spe-cies of the palustris group; mesochaetae smooth, macro-chaetae unilaterally slightly ciliated; 3 + 3 macrochaetaeon both abd. IV and V, arranged in a triangular fashion andin a row, respectively. Trichobothrial formula: 0 + 1 + 1 (or0 + 0 + 1 according to Poinsot-Balaguer & Ferard 1983). Thedistribution of ‘S’ setae is typical of Type 2 (Fig. 2C):ms1 + al1 + accp1–5 on thr. II and III; ms1 + accp1–5 on abd.I and II; ms1 + accp1–6 on abd. III; as1 and 4 + accp1–5 onabd. IV; as1 + accp1–5 (accp1 and accp2 longer than theothers) on abd. V.
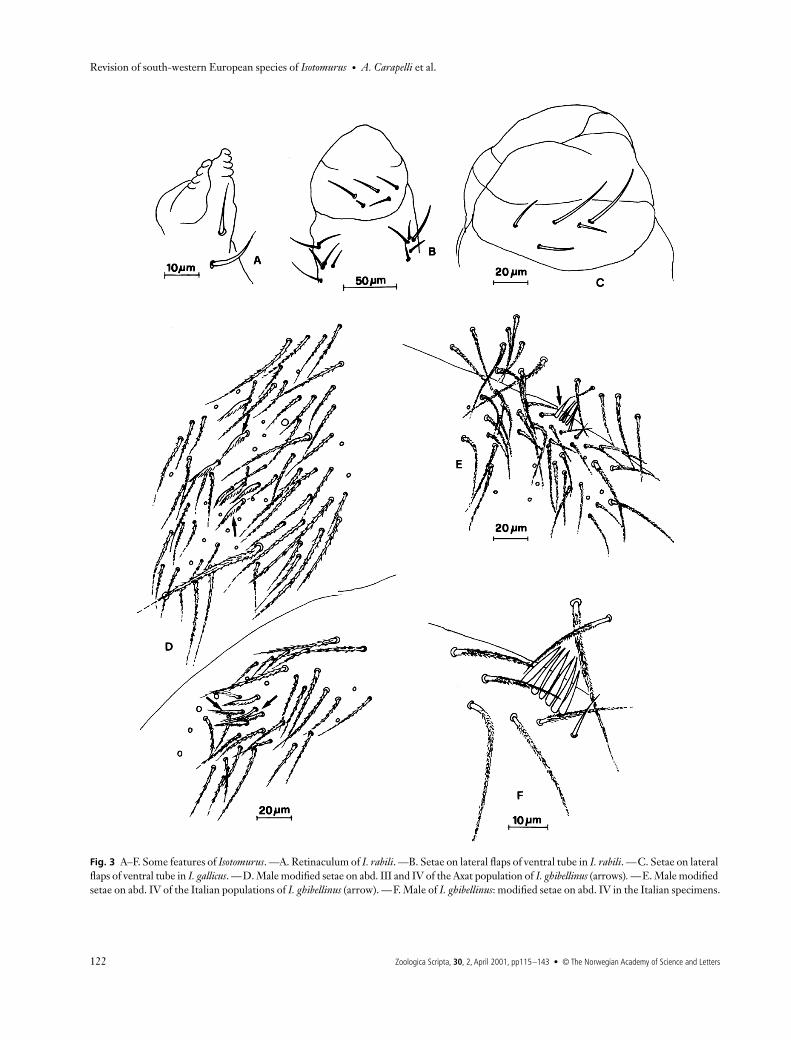
L/antennae. 3.24 –3.27.Ventral tube. Lateral flaps with 5 + 5 setae.Legs. Claw with outer and lateral teeth. Empodial append-
age with or without tooth. Tibiotarsus III with short ordinarysetae and with at least 3 smooth macrochaetae.
Furca. Retinaculum with 2 ordinary setae. Dorsal side ofmanubrium covered with several smooth setae mainly dis-tributed on the anteromedian part and on the distal margin.
ZSC055.fm Page 120 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 121
Ventrally with numerous short setae, which are absentonly in a narrow longitudinal stripe. Dental chaetotaxy madeup of several smooth microchaetae on the dorsal side andvery few on the ventral side; setae are longer on the lateralmargins; cc of type ‘A’. Setae are inserted in circular bulgesof the dental cuticle. Dorsally, two microsensilla are presenton each dens. Mucro short, quadridentate and without setae.
Distribution and habitat. The distribution of the species islimited to the south-western Massif Central (France). Speci-mens were collected on mosses and limestone ground.
Isotomurus ghibellinus Carapelli et al., 1995Isotomurus ghibellinus Carapelli et al., 1995b: 94, Italy.
Type locality. Radi (Siena, Italy).Material examined. Several specimens from Aitone (Corsica,
France), Apuane Alps (Italy), Allone (Perugia, Italy), Axat(Pyrénées, France), Bocca Serriola (Perugia, Italy) and Radi(Siena, Italy).
DescriptionHabitus. Habitus of Isotoma. Body length: 2–3 mm.
Coloration. Specimens have a whitish or yellowish back-ground. Anterior part of each tergite with a dark transverseband more or less diffuse on rest of segment. Variability wasobserved in the thickness of the band. Some specimens aredepigmented or entirely dark. Central part of head and basesof antennae with some pigmented patches. Some specimensfrom Aitone population (Corsica Island) have last abdominaltergites very dark. Claws may be pigmented.
Antennae. Lengths of antennal segments I : II : III : IV are45 : 60 : 72 : 80.
Head. 8 + 8 eyes. Postantennal organ of regular shape.PAO/dcA: 1. Labrum with 3 parallel rows of 5, 5 and 4 setae,respectively. Labium with 6–10 setae on each part of sub-mentum. The maxillary outer lobe is bipartite and with4 sublobal hairs.
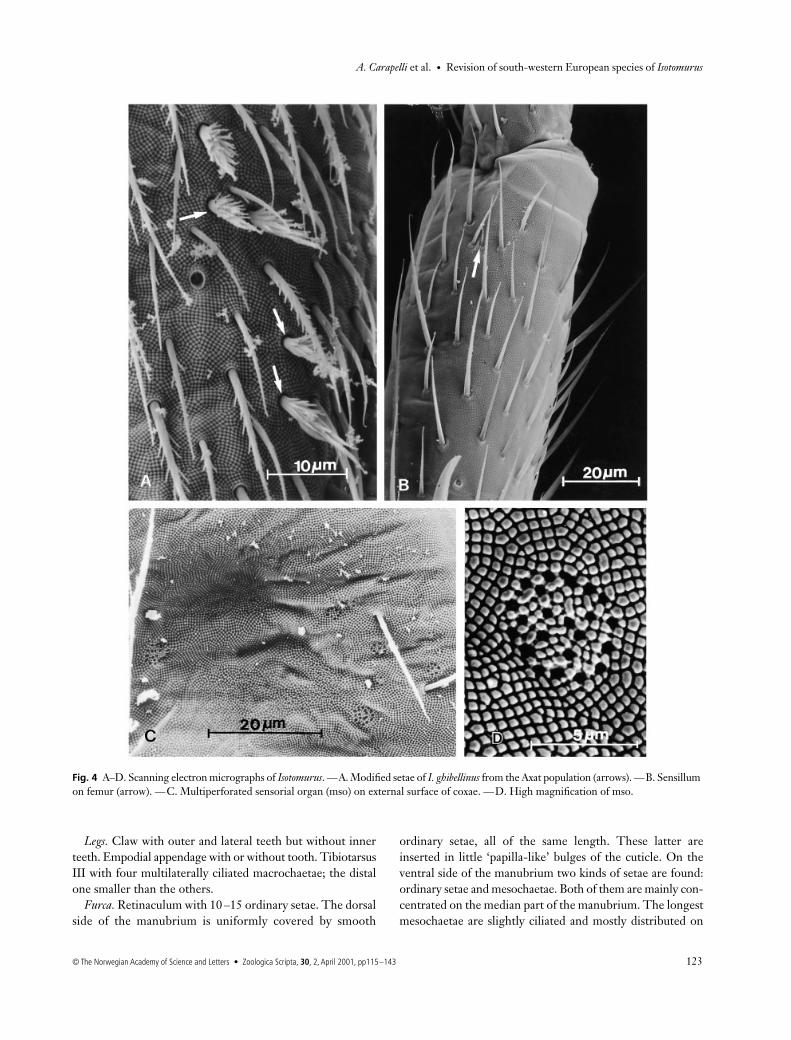
Chaetotaxy. Ordinary setae abundant; mesochaetae slightlyciliated. Macrochaetae multilaterally ciliated and arrangedon each hemitergite in a triangular fashion on abd. IV and ina row on abd. V. Trichobothrial formula typical of palustrisgroup (3 + 3 + 1). Distribution of ‘S’ setae typical of Type 1A(Fig. 2A): ms1 + al1 + accp1–5 on thr. II and III; ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III; as1, 2 and4 + accp1–5 on abd. IV; as1–2 + accp1–5 (accp1 and accp2longer than the others) on abd. V. Several modified setae onventrolateral region of abd. IV (III and IV in the Axat popu-lation) (Figs 3D and 4A) in the males.
L/antennae. 2.28.Ventral tube. Lateral flaps with 3 + 3 setae.Legs. Claw with outer and inner teeth. Empodial append-
age frequently armed with a little tooth. Tibiotarsus III with4 –5 ciliated macrochaetae.
Furca. Retinaculum with 12–16 ordinary setae. Manu-brium with several short setae and two dorsal macrochaetaeon each side. Ventrally with shorter setae densely present oncentral and distal margins. Dentes with many ordinary setaeon the ventral side, inserted in a circular area of the cuticle;dorsal chaetotaxy reduced and composed of short setae, mostlydispersed on the proximal side of dens. Two microsensilla arefound on the dorsal side. Lateral margins of dentes with severallonger setae; cc of type ‘A’. Mucro quadridentate without setae.
Distribution and habitat. This European species shows somepreferences for living on clay and limestone near freshwaterstreams.
Isotomurus hadriaticus sp. n.Holotype. Female, ITALY, S. Vitale (Ravenna), at sea level, pinewood forest, 25 May 1994, preserved in the R. Dallai collectionat the Department of Evolutionary Biology, University of Siena.
Paratypes. Same data as for the holotype, one male and twofemales.
Etymology. The species is named after the first collectionsite on the Adriatic coast of Italy, where it appears to be endemic.
Diagnosis. This species can be distinguished by the uniformdark pigmentation and by the presence of 4 as on abd. IV.
DescriptionHabitus. Body slender; habitus of Isotoma. Body length: 2–3 mm.
Coloration. Body uniformly black including antennae andlegs; no longitudinal or transverse stripe on tergites. Headwith central dark patch of pigmentation and with bases ofantennae pigmented. Furca usually white.
Antennae. Of normal length as observed in the palustrisgroup of species. Ratio of the antennal segments I : II : III : IV is24 : 33 : 43 : 50. Some specimens have ant. III and IV equallylong. Four microsensillae are present on ventral side of ant. I.
Head. 8 + 8 eyes. Postantennal organ elongated. PAO/dcA:1.38. Labrum shows 3 parallel rows of 5, 5 and 4 setae, respect-ively. Labium with 7–9 setae on each part of submentum. Themaxillary outer lobe is bipartite and with four sublobal hairs.
L/antennae. 2.35.Chaetotaxy. Ordinary setae very abundant; mesochaetae
multilaterally ciliated. On both abd. IV and V, there are threemultilaterally ciliated macrochaetae arranged in a triangularfashion and in a row, respectively. Trichobothria are thin,long and ciliated; their number and distribution are as in thepalustris group, with 3 + 3 trichobothria on abd. II and III and1 + 1 on abd. IV. The distribution of ‘S’ setae is typical ofType 1B (Fig. 2D): ms1 + al1 + accp1–5 on thr. II and III;ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III;as1– 4 + accp1–5 on abd. IV; as1–2 + accp1–5 (accp1 andaccp2 longer than the others) on abd. V.
Ventral tube. Lateral flaps with 3 + 3 setae.
ZSC055.fm Page 121 Friday, May 25, 2001 3:44 PM
Revision of south-western European species of Isotomurus • A. Carapelli et al.
122 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
Fig. 3 A–F. Some features of Isotomurus. —A. Retinaculum of I. rabili. —B. Setae on lateral flaps of ventral tube in I. rabili. —C. Setae on lateral flaps of ventral tube in I. gallicus. —D. Male modified setae on abd. III and IV of the Axat population of I. ghibellinus (arrows). —E. Male modified setae on abd. IV of the Italian populations of I. ghibellinus (arrow). —F. Male of I. ghibellinus: modified setae on abd. IV in the Italian specimens.
ZSC055.fm Page 122 Friday, May 25, 2001 3:44 PM
A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 123
Legs. Claw with outer and lateral teeth but without innerteeth. Empodial appendage with or without tooth. TibiotarsusIII with four multilaterally ciliated macrochaetae; the distalone smaller than the others.
Furca. Retinaculum with 10 –15 ordinary setae. The dorsalside of the manubrium is uniformly covered by smooth
ordinary setae, all of the same length. These latter areinserted in little ‘papilla-like’ bulges of the cuticle. On theventral side of the manubrium two kinds of setae are found:ordinary setae and mesochaetae. Both of them are mainly con-centrated on the median part of the manubrium. The longestmesochaetae are slightly ciliated and mostly distributed on
Fig. 4 A–D. Scanning electron micrographs of Isotomurus. —A. Modified setae of I. ghibellinus from the Axat population (arrows). —B. Sensillum on femur (arrow). —C. Multiperforated sensorial organ (mso) on external surface of coxae. —D. High magnification of mso.
ZSC055.fm Page 123 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
124 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
the apical part of the manubrium. The ventral side of dentesis uniformly covered by ordinary setae, which are longer onthe ventrolateral sides of each dens. The dorsal side of denteshas many smooth setae, which are mainly distributed from thebasal toward the middle part of it. These setae appear weakerthan those from the ventral side and, as in the manubrium, areinserted in a circular area. Two small sensillae are present onthe dorsal side of dentes towards the distal end; cc of type ‘A’.Mucro quadridentate without mucronal seta.
Distribution and habitat. Specimens were sampled in a swampenvironment composed of decaying organic material andfreshwater ponds, under the S. Vitale pine wood.
Isotomurus maculatus (Schäffer, 1896)Isotoma palustris maculata Schäffer, 1896: 63, Germany.Isotoma maculata Börner, 1901: 51, Central Europe.Isotomurus palustris maculata Axelson, 1906: 16, Russia.Isotomurus maculatus Kos, 1937: 185, Yugoslavia.Isotomurus palustris maculatus Poinsot-Balaguer, 1976: 646,France.Isotomurus maculatus Carapelli et al., 1995a: fig. 2a; Carapelliet al., 1995b: fig. 2a.
Type locality. Germany.Material examined. Several specimens from Adria (Ravenna,
Italy), Allone (Perugia, Italy), Antillo (Sicily, Italy), Axat(Pyrénées, France), Catania (Sicily, Italy), Fortore (Foggia,Italy), Fos sur Mer (Camargue, France), Gerfalco (Grosseto,Italy), Giglio Island (Italy), Pertosa (Salerno, Italy), Radi(Siena, Italy), Siena (Italy), Sorra (Siena, Italy) and Zomaro(Reggio Calabria, Italy). Several specimens from the originalcollection (Schäffer 1896; Zoological Museum of Hamburg).
DescriptionHabitus. Habitus of Isotoma. Body length: 2–3 mm.
Coloration. Background whitish. Irregular patches andspots of variable colour are present on tergites. Head withsome dark patches; a symmetrical pigmented spot is presenton the lateral sides of abd. VI.
Antennae. Relative lengths of antennal segments I : II : III :IV are 37.5 : 55 : 55 : 62.5.
Head. 8 + 8 eyes. PAO oval. PAO/dcA: 1. Labrum with3 parallel rows of 5, 5 and 4 setae, respectively. Labium with10–16 setae on each part of submentum.
Chaetotaxy. Ordinary setae very abundant; meso- and mac-rochaetae multilaterally ciliated. Three macrochaetae arrangedin a triangular fashion are found on each side of abd. IV.Three additional longer setae distributed in a row are presenton each hemitergite of abd. V. The trichobothrial formula istypical of the palustris group (3 + 3 + 1). The distribution of‘S’ setae is typical of Type 1A (Fig. 2A): ms1 + al1 + accp1–5on thr. II and III; ms1 + accp1–5 on abd. I and II; ms1 +accp1–6 on abd. III; as1, 2 and 4 + accp1–5 on abd. IV;
as1–2 + accp1–5 (accp1 and accp2 longer than the others) onabd. V.
L/antennae. 2.3.Ventral tube. Lateral flaps with 3 + 3 setae.Legs. Claw with outer and lateral teeth. Empodial appendage
frequently armed with a tooth. Tibiotarsus III with 3 – 4ciliated macrochaetae.
Furca. Retinaculum with 10–30 ordinary setae. Manubriumwith many short setae and two longer ones on dorsal side, andwith several ventral mesochaetae which are mostly distributedon the distal margin. Dentes dorsally with many short setae,which are longer on lateral margins, and two microsensilla;ventrally with a few microchaetae; cc of type ‘A’. Setae areinserted in circular bulges of the dental cuticle. Mucro quad-ridentate without seta.
Distribution and habitat. Eurasian distribution, with one recordin Australia. Specimens were sampled on humid environmentsof grasses, frequently exposed to sunlight.
Isotomurus nebulosus Lek & Carapelli, 1998Isotomurus nebulosus Lek & Carapelli, 1998: 1– 4.
Type locality. Arbas, Haute-Garonne, near Goueil di HerCave (Pyrénées Mountains, France).
Material examined. Several specimens from Montségur andArbas (Pyrénées Mountains, France).
DescriptionHabitus. Habitus of Isotoma. Body length: 2 mm.
Coloration. Body yellow-brownish with thin violet drawingsmostly present on last abdominal tergites. Longitudinal andtransverse stripes absent. Abd. VI with 1 + 1 ventrolateral brownspots. Violet pigmentation is present on lateral margins ofabd. III–V. Antennae (especially ant. IV), claw and empo-dium usually pigmented. Legs and furca whitish.
Antennae. Of normal length. Relative lengths of antennalsegments I : II : III : IV are 60 : 78 : 80 : 82.
Head. 8 + 8 eyes. PAO rounded. PAO/dcA: 0.6 – 0.8. Labrumwith 3 parallel rows of 5, 5 and 4 setae, respectively. Labiumwith 10–15 setae on each part of submentum. The maxillaryouter lobe is bipartite and with 4 sublobal hairs.
Chaetotaxy. Ordinary setae abundant; tergites coated withsmooth mesochaetae and long multilaterally ciliated macro-chaetae; 3 + 3 macrochaetae are present on abd. IV and abd.V where they are arranged in a triangular fashion and in arow, respectively. Trichobothrial formula typical of palustrisgroup (3 + 3 + 1). The distribution of ‘S’ setae is typical ofType 1A (Fig. 2A): ms1 + al1 + accp1–5 on thr. II and III;ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III;as1, 2 and 4 + accp1–5 on abd. IV; as1–2 + accp1–5 (accp1 andaccp2 longer than the others) on abd. V.
L/antennae. 2.25.Ventral tube. Lateral flaps with 3 + 3 setae.
ZSC055.fm Page 124 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 125
Legs. Claw with outer and lateral teeth. Empodial append-age frequently with tooth. Tibiotarsal chaetotaxy composedof smooth ordinary setae and several slightly ciliated macro-chaetae (generally 4 on the metathoracic legs).
Furca. Retinaculum with 14–32 ordinary setae. Manu-brium with many short setae and two longer ones on dorsalside, and with several ventral mesochaetae, which are mostlydistributed on the distal margin. Dentes dorsally with manyshort setae, which are longer on lateral margins, and twomicrosensilla; a few microchaetae on the ventral side; cc oftype ‘A’. Setae are inserted in circular bulges of the dentalcuticle. Mucro quadridentate without mucronal seta.
Distribution and habitat. Specimens were sampled in humidmosses on slopes from central Pyrénées, where the speciesappears to be endemic.
Isotomurus palustris (Müller, 1776)Podura palustris Müller, 1776: 184.Isotomurus palustris Börner, 1903; Folsom, 1937: 72, Pl. 25,fig. 281, Pl. 26, figs 284–290; Stach, 1947: 452–461.Isotomurus palustris palustris Poinsot-Balaguer, 1976: 641–646, fig. 1.Isotomurus indipendente Carapelli et al., 1995b: fig. 2f. syn. nov.
Neotype. Female, Mols Hills (Mols Bjerge; Randers District),Denmark, at sea level, pond shore, 13 May 2000, preservedat the Zoological Museum, Entomology Department, Uni-versity of Copenhagen, Denmark.
Material examined. Several specimens from the Haarløvcollection. Other material from: Bocca Serriola (Perugia,Italy), Cecita (Cosenza, Italy), Fos sur Mer (Camargue, France),Gerfalco (Grosseto, Italy), Kolind (Randers District, Denmark),Marie Magdalene (Randers District, Denmark), Pioch Badet(Camargue, France), Radi (Siena, Italy) and Salin de Badon(Camargue, France).
DescriptionHabitus. Habitus of Isotoma. Body length: 2.0–3.4 mm (Fig. 5A).
Coloration. Background colour from yellow-brownish togreenish. Tergites with a weak longitudinal stripe, frequentlyinterrupted or fainter, especially on abd. III. The last abdom-inal segments have a variable net-like pigmentation. Darkerpigmentation is usually observed on lateral sides of tergites,precoxa, coxa, trochanter and ant. IV. Head with central blackspot and with patches of pigmentation at the bases of anten-nae. Each part of abd. VI with a dark spot.
Antennae. Antennal segments of normal length. Relativelengths of ant. I : II : III : IV are 50 : 85 : 88 : 88.
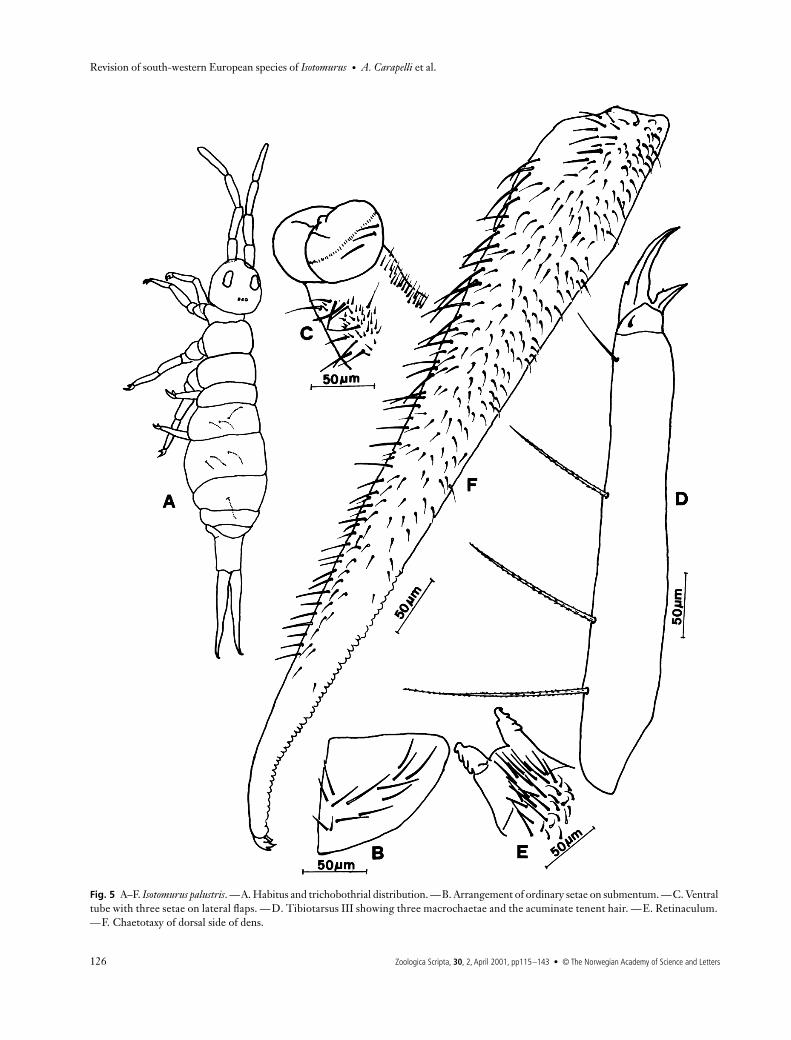
Head. 8 + 8 eyes. Postantennal organ elongated. PAO/dcA:0.8. Labrum with 3 parallel rows of 5, 5 and 4 setae, respect-ively. Labium with 8–17 setae on each part of submentum(Fig. 5B). The maxillary outer lobe is bipartite and with 4sublobal hairs.
Chaetotaxy. General chaetotaxy very abundant. Meso-chaetae and macrochaetae multilaterally ciliated. On eachhemitergite of abd. IV and V there are 3 setae arranged as atriangle and as a row, respectively. Trichobothrial formula:3 + 3 + 1. The distribution of ‘S’ setae is of Type 1A (Fig. 2A):ms1 + al1 + accp1–5 on thr. II and III; ms1 + accp1–5 on abd.I and II; ms1 + accp1–6 on abd. III; as1, 2 and 4 + accp1–5 onabd. IV; as1–2 + accp1–5 (accp1 and accp2 longer than theothers) on abd. V.
L/antennae. 2.3.Ventral tube. Lateral flaps with 3 + 3 setae (Fig. 5C).Legs. Claw with outer and lateral teeth. Empodial append-
age with or without tooth. Tibiotarsus III with 3–5 ciliatedmacrochaetae (Fig. 5D).
Furca. Retinaculum with 14–35 ordinary setae (Fig. 5E).Manubrium dorsally with several small ordinary setae andtwo longer ones on each side; ventrally the setae are longerand slightly ciliated. Dentes covered with many small setaeand two microsensilla on the dorsal side, and by a few smallsetae on the ventral side (Fig. 5F). Laterally each dens haslonger setae. Setae are inserted in circular bulges of thecuticle; cc of type ‘A’. Mucro quadridentate without seta.
Distribution and habitat. Specimens of this species are gen-erally found on clay and grasses near damp environments.
TaxonomyMüller (1776) described Podura palustris from Denmark.Since its description, the species and its ‘colour varieties’have been reported from many European localities: Poland,Lithuania, west and central Ukraine, Slovakia, Hungary,Austria, Germany, France, England, Albania, Yugoslavia,Malta, Romania, Portugal (Stach 1947; Da Gama 1964;Poinsot-Balaguer 1976; Cassagnau 1987; Deharveng & Lek1993).
A few years ago, Carapelli et al. (1995b) demonstrated thepresence of six sympatric species of Isotomurus in a locality incentral Italy (Radi, Tuscany, Siena Province). Three of thesespecies had a similar longitudinal stripe, but were geneticallydifferent from one another. We assigned one of these speciesto I. palustris, one to I. unifasciatus, which was given the rankof species, and we described the new species I. indipendente forthe third.
However, the status of I. palustris was problematic. In fact,in spite of the many times this species has been quoted infaunistic surveys, which makes it one of the most widely citedCollembola (Salmon 1964), the type material is unknown(Greenslade 1994) and no revision has been performed basedon typical or topotypical material. Henriksen (1921– 37:p. 67) pointed out that: ‘It is uncertain whether Müller everhad a collection, in any case nothing is known about whathappened to it’. He also added that this material wassearched several times and that the German entomologist
ZSC055.fm Page 125 Friday, May 25, 2001 3:44 PM
Revision of south-western European species of Isotomurus • A. Carapelli et al.
126 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
Fig. 5 A–F. Isotomurus palustris. —A. Habitus and trichobothrial distribution. —B. Arrangement of ordinary setae on submentum. —C. Ventral tube with three setae on lateral flaps. —D. Tibiotarsus III showing three macrochaetae and the acuminate tenent hair. —E. Retinaculum. —F. Chaetotaxy of dorsal side of dens.
ZSC055.fm Page 126 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 127
H. A. Hagen claimed that Müller’s material and books were:‘… destroyed during the bombardments of the City ofCopenhagen …’. This probably happened during the warbetween Denmark and England in 1807 (N. P. Kristensenand O. Karsholt, personal communication).
To clarify the status of Podura palustris, a neotype must beselected from material collected in the type locality (‘Den-mark’). We recently received from Professor N. P. Kristensensome material collected by Dr N. Haarløv in the Danishlocalities where Müller is supposed to have collected hisspecimens. Unfortunately, this material was composed ofimmature specimens, still lacking several diagnostic charactersuseful for species identification and therefore not suitable forthe designation of a neotype. Later, we personally collectedadditional material from Denmark (for locality data, seedescription of the species) which resulted in three differentIsotomurus species. One of these species showed an irregularcoloration consisting of pigmented patches. Since Müllerdescribed I. palustris as having a black dorsal stripe, this spe-cies can be excluded. It actually corresponds to I. maculatusas identified by Carapelli et al. (1995a). The remaining twospecies, both having a black dorsal stripe, could be Müller’sPodura palustris. One of these was found to correspondmorphologically and genetically to I. plumosus Petersen, 1965(Petersen 1965 pp. 350–353), and the other to a species of theI. palustris group; this results in correspondence to I. palustrispalustris sensu Poinsot-Balaguer (1976) from France (Camargue)and to I. indipendente Carapelli et al., 1995 from Italy (SienaProvince).
Consequently, in order to stabilize nomenclature, onespecimen (Zoological Museum of Copenhagen) of the latterspecies was selected as the neotype of I. palustris Müller.
The immediate consequence of the above is that I. indipendentecomes to be a junior synonym of I. palustris, and the Italianspecies recently studied by us and referred to as I. palustriscomes to be a distinct species new for science (see below).
Isotomurus prasinus (Reuter, 1891)Isotoma stuxbergi prasina Reuter, 1891: 229, Siberia.Isotoma palustris prasina Schött, 1893: 66, Palearctic.Isotomurus palustris prasina Reuter, 1895: 26, Finland, Siberia,Sweden.Isotomurus palustris prasina Axelson, 1905: 35, Tvärminne,Finland.Isotomurus palustris f. prasina Stach, 1947: 459.Isotomurus palustris prasina Poinsot-Balaguer, 1976: 646, France.Isotomurus palustris prasinus Deharveng & Lek, 1993: 255,France.
Type locality. Siberia.Material examined. Several specimens from: Malga Panna
(Trento, Italy), Montagna Spaccata (Vicenza, Italy), Tjöme(Norway).
DescriptionHabitus. Habitus of Isotoma. Body length: 2 mm.
Coloration. Body uniformly light green, usually with a darkspot on each side of abd. VI. Antennae and legs often brownish.Head with central patch of pigmentation and spots betweenbases of antennae.
Antennae. Antennal segments slender, with relative lengthsof ant. I : II : III : IV as 53 : 85 : 85 : 90.
Head. 8 + 8 eyes. Postantennal organ elongated. PAO/dcA:0.8. Labrum with 3 parallel rows of 5, 5 and 4 setae, respectively.Labium with 5–8 setae on each part of submentum. Themaxillary outer lobe is bipartite and with 4 sublobal hairs.
Chaetotaxy. Ordinary setae very abundant; abdominaltergites with ciliated meso- and macrochaetae. On abd. IVthe 3 macrochaetae are arranged in a triangular fashion oneach side of the tergite. On abd. V the 3 macrochaetae arearranged in a transversal row. Trichobothrial formula typicalof the palustris group (3 + 3 + 1). The distribution of ‘S’ setaeis typical of Type 1A (Fig. 2A): ms1 + al1 + accp1–5 on thr. IIand III; ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 onabd. III; as1, 2 and 4 + accp1–5 on abd. IV; as1–2 + accp1–5(accp1 and accp2 longer than the others) on abd. V. Severalmodified setae on ventrolateral region of abd. III and IV inmales (Fig. 6A–C).
L/antennae. 2.4.Legs. Claw with outer and lateral teeth. Empodial appendage
with or without tooth. There are 3–5 ciliated tibiotarsalmacrochaetae, which are larger on tibiotarsus III.
Ventral tube. Lateral flaps with 3 + 3 setae.Furca. Retinaculum with 10–35 ordinary setae. Dorsal
chaetotaxy of manubrium with several slightly ciliated setaemainly distributed on the distal margin. Ventral side of man-ubrium covered with several short smooth microchaetae, whichare absent only in the median part of it. Dental chaetotaxy madeup of short microchaetae on the ventral side, mainly distributedon the proximal part; dorsally each dens is completely coveredby ordinary setae inserted in little bulges of the cuticle; cc oftype ‘A’. Inner and outer margins of dentes covered with a fewlonger setae. Two microsensilla are found in the median partof the dorsal side. Mucro is quadridentate and without seta.
Distribution and habitat. Holoarctic with some reports inAsia. In Italy this species was found on mosses near freshwaterAlpine brooks. In Northern Europe often abundant in lawnsand moist grassland.
Isotomurus pseudopalustris sp. n.Isotomurus palustris Carapelli et al., 1995a: fig. 2b; Carapelliet al., 1995b: fig. 2d.
Holotype. Male, ITALY, Radi (Siena, Tuscany), 300 m,Mediterranean shrub, 4 April 1993, preserved in the R. Dallaicollection at the Department of Evolutionary Biology, Uni-versity of Siena.
ZSC055.fm Page 127 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
128 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
Fig. 6 A–D. Ventral modified setae on males of I. prasinus and I. fucicolus. —A. Abd. III and IV in the Norwegian specimens of I. prasinus. —B. Enlargement of modified setae on abd. IV in the Norwegian specimens of I. prasinus. —C. Abd. III and IV in the Italian specimens of I. prasinus. —D. Abd. IV in the Italian specimens of I. fucicolus.
ZSC055.fm Page 128 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 129
Paratypes. Same data as for holotype, three males and fivefemales.
Etymology. The name is derived from the similaritiesobserved in the pigmentation pattern of this species with I.palustris, from which it differs in having a darker longitudinalstripe and a more pronounced lateral pigmentation. It alsodiffers from I. unifasciatus in the presence of lateral pig-mentation, which is mostly absent in I. unifasciatus.
Material examined. Allone (Perugia, Italy), Baulì (Sicily),Classe (Ravenna, Italy), Gerfalco (Grosseto, Italy), GranSasso (L’Aquila, Italy), Mucone (Cosenza, Italy), Pennapied-imonte (Chieti, Italy), Radi (Siena, Italy), Seravezza (ApuaneAlps, Italy) and Sorra (Siena).
DescriptionHabitus. Habitus of Isotoma. Body length: 2 mm.
Coloration. Body with a whitish background. Longitudinaldark stripe is rarely interrupted on the last abdominal tergitesand sometimes fused with lateral brown pigmentation. Lat-eral margin of thr. II and III usually darker than the rest oftergites. Each side of abd. VI has a dark spot. Head often pig-mented with irregular patches. Antennae and legs brownish.
Antennae. Antennae of normal length. Ratio of the antennalsegments I : II : III : IV is 40 : 60 : 65 : 65.
Head. 8 + 8 eyes. Postantennal organ oval and elongated.PAO/dcA: 0.9. Labrum with 3 parallel rows of 5, 5 and 4setae, respectively. Labium with 8–12 setae on each part ofsubmentum. The maxillary outer lobe is bipartite and with4 sublobal hairs.
Chaetotaxy. Ordinary setae smooth and abundant; meso-chaetae slightly ciliated; 3 + 3 multilaterally ciliated macro-chaetae on abd. IV and V, arranged as a triangle and in a row,respectively. Trichobothrial formula: 3 + 3 + 1. Distributionof ‘S’ setae typical of Type 1A (Fig. 2A): ms1 + al1 + accp1–5on thr. II and III; ms1 + accp1–5 on abd. I and II; ms1 + accp1– 6on abd. III; as1, 2 and 4 + accp1–5 on abd. IV; as1–2 + accp1–5(accp1 and accp2 longer than the others) on abd. V.
L/antennae. 2.3.Ventral tube. Lateral flaps with 3 + 3 setae.Legs. Claw with outer and lateral teeth. Empodial appendage
with or without tooth. Tibiotarsus III with 4–5 ciliatedmacrochaetae.
Furca. Retinaculum with 10–18 ordinary setae. Manubriumwith many short setae and two longer ones on dorsal side, andwith several mesochaetae mostly distributed on the distal marginof ventral side. Dentes dorsally with many short setae, whichare longer on the lateral margins, and two microsensilla;ventrally with a few microchaetae. Setae are inserted incircular bulges of the cuticle; cc of type ‘A’. Mucro quadri-dentate and without seta.
Distribution and habitat. Species widespread probably ofcosmopolitan distribution. Specimens were sampled on
limestone ground on the banks of freshwater streams and inlitter.
Isotomurus rabili Deharveng & Lek, 1993Isotomurus rabili Deharveng & Lek, 1993: 257–259, fig. 11.
Type locality. Tarn: Larroque, Forest of Grésigne (France).Material examined. Several specimens from Tarn: Larroque,
Forest of Grésigne (France).
DescriptionHabitus. Habitus of Isotoma. Body length: 1.3–1.4 mm.
Coloration. Background colour whitish or yellowish. Thr.II–III, abd. I–II and the anterior part of abd. III with a black–violet pigmentation. Head usually whitish with a central violetspot; bases of antennae dark pigmented.
Antennae. Antennae short. Relative lengths of ant. I : II :III : IV are 20 : 27 : 30 : 40.
Head. 8 + 8 eyes. Postantennal organ large if compared withbody size. PAO/dcA: 1.6 –1.8. Labrum with 3 parallel rows of5, 5 and 4 setae, respectively. Labium with 5 –7 setae on eachpart of submentum. The maxillary outer lobe is bipartite andwith 4 sublobal hairs.
Chaetotaxy. Ordinary setae reduced; mesochaetae smooth,macrochaetae slightly unilaterally ciliated; 3 + 3 macrochaetaeon abd. IV and V are arranged in a triangular fashion and ina row, respectively. Trichobothrial formula 0 + 1 + 1 (as in I.gallicus). The distribution of ‘S’ setae is of Type 2 (Fig. 2C):ms1 + al1 + accp1–5 on thr. II and III; ms1 + accp1–5 on abd.I and II; ms1 + accp1– 6 on abd. III; as1 and 4 + accp1–5 onabd. IV; as1 + accp1–5 (accp1 and accp2 longer than the others)on abd. V.
L/antennae. 3.9.Ventral tube. Lateral flaps with 5 + 5 setae.Legs. Claw with outer and lateral teeth. Empodial appendage
with or without tooth. Tibiotarsus III with short ordinary setae,and at least 3 smooth macrochaetae.
Furca. Retinaculum with 2 ordinary setae. Dorsal setae ofmanubrium with several smooth setae mainly distributed onthe anteromedian part and on the distal margin. Ventrallywith numerous short setae, which are absent only in a narrowlongitudinal part of manubrium. Dentes with several smoothmicrochaetae, which are longer on lateral margins, and twomicrosensilla on the dorsal side, and with very few micro-chaetae on the ventral side. Setae are inserted in circular bulgesof the cuticle; cc of type ‘A’. Mucro short, quadridentate andwithout setae.
Distribution and habitat. Specimens were collected onmosses and limestone ground on the south-western MassifCentral (south of France), where the species is endemic.
Isotomurus unifasciatus (Börner, 1901)Isotoma palustris unifasciata Börner, 1901.
ZSC055.fm Page 129 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
130 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
Isotomurus palustris f. unifasciata Stach, 1947: 459.Isotomurus unifasciatus Carapelli et al., 1995b: 94, fig. 2e.
Type locality. Germany.Material examined. Allone (Perugia, Italy), Bocca Serriola
(Perugia, Italy), Fontestorbes (Pyrénées, France), Gerfalco(Grosseto, Italy), Montségur (Pyrénées, France), MalgaPanna ( Trento, Italy), Orsara (Foggia, Italy), Pennapiedimonte(Chieti, Italy), Pertosa (Salerno, Italy), Radi (Siena, Italy),Siena (Italy) and Sorra (Siena, Italy). One paratype fromBörner’s collection (1900, Bremen, Germany, British Museumof Natural History).
DescriptionHabitus. Body slender; habitus typical of Isotoma. Body length:2–3 mm.
Coloration. Body background yellowish or greenish. Tergiteswith longitudinal dark band sometimes interrupted on abd.IV and V. Some specimens are darker. A dark pigmented spotis present on lateral sides of abd. VI; dark patches are presentnear the bases of antennae. Antennal segment slightly violet.In the Siena population, the lateral sides of tergites are excep-tionally brown coloured (see the ‘Discussion’ section).
Antennae. Average length of antennal segments as in thepalustris group of species. Ratio of the antennal segmentsI : II : III : IV is 50 : 70 : 80 : 85.
Head. 8 + 8 eyes. Postantennal organ elongated. PAO/dcA:0.7. Labrum with 3 parallel rows of 5, 5 and 4 setae, respect-ively. Labium with 10–15 setae on each part of submentum.The maxillary outer lobe is bipartite and with 4 sublobalhairs.
Chaetotaxy. Abdominal tergites densely covered by a veryabundant chaetotaxy, composed of microchaetae and multi-laterally ciliated macrochaetae. On each hemitergite of abd.IV and V, there are 3 ciliated macrochaetae which are arrangedin a triangular fashion and in a row, respectively. Tricho-bothrial formula typical of palustris group (3 + 3 + 1). Thedistribution of ‘S’ setae is of Type 1A ( Fig. 2A): ms1 + al1 +accp1–5 on thr. II and III; ms1 + accp1–5 on abd. I and II; ms1 +accp1–6 on abd. III; as1, 2 and 4 + accp1–5 on abd. IV; as1–2+ accp1–5 (accp1 and accp2 longer than the others) on abd. V.
L/antennae. 2.5.Ventral tube. Lateral flaps with 3 + 3 setae.Legs. Claw with outer and lateral teeth. Empodial append-
age sometimes provided by tooth. Tibiotarsus III with 4 cili-ated macrochaetae, which are less developed in comparisonwith abdominal ones.
Retinaculum. With 10–25 ordinary setae.Furca. Dorsolateral sides of manubrium covered with several
slightly ciliated mesochaetae, which are rarer in the centralpart. Ventral side of manubrium with abundant short andsmooth microchaetae, absent only in the central part, andwith a few longer setae, usually 2, on each side. Dentes are
composed of an abundant chaetotaxy on the dorsal side, andof short, smooth, microchaetae on the ventral part. Ventralmicrochaetae are spread mostly on the proximal half of densand completely absent near the mucro. Two microsensilla areobserved dorsally on each dens, where some longer, ciliatedsetae, arranged in longitudinal rows, stand laterally on eachside. Dorsal setae are inserted in a papilla-like cuticular area;cc of type ‘A’. Mucro is quadridentate and without seta.
Distribution and habitat. European species. Specimens weresampled in mosses and grasses near freshwater streams.
Results and discussionColour patternsPigmentation patterns represent the most used and straight-forward character for species diagnosis. In Isotomurus, however,the degree of intra- and interpopulation variability constitutesan obstacle to the recognition of those features which aremost useful for this purpose.
Four species of Isotomurus have a longitudinal stripe: I. alti-colus (Carl 1899), I. palustris (Müller 1776), I. pseudopalustrissp. n. and I. unifasciatus (Börner 1901). In I. alticolus (Carl 1899),the longitudinal stripe is broader than in the other three species,and sometimes tends to diffuse laterally. According to Stach(1947), some depigmented specimens may lack the longitu-dinal stripe.
In the remaining three species with a longitudinal stripe,each tergite is occasionally coloured with a more or less diffusepigmentation which rarely overlaps with the longitudinalstripe. In addition, I. palustris and I. unifasciatus often have agreen background colour. As a result of these similarities, itis sometimes difficult to distinguish between these speciesand, given that the median longitudinal stripe is common toall of them, it is the presence and shape of the lateral accessorypigmentation which should be taken into account to tell themapart.
Isotomurus palustris is probably the most frequently describedspecies of the genus. Several different descriptions have beenproposed for this species (Folsom 1937; Stach 1947; Maynard1951). As mentioned above, a major source of uncertainty isthe extreme intraspecific variability of body colour, and it ispossible that more than one species was hidden under themorpho-type generally proposed for I. palustris.
Two of the seven colour varieties of I. palustris introducedby Stach (1947), I. palustris f. unifasciata (Börner 1901) and I.palustris f. bimaculata (Ågren 1903), have a similar pigmenta-tion pattern; these two varieties are differentiated by ‘a darkspot laterally on the sixth abdominal segment’, which isobserved in bimaculata. This model is very similar to somedrawings of I. palustris f. maculata by Stach (1947: Pl. XLVIII,figs 1 and 2) and to the ‘Type B’ drawing of I. palustris f. palus-tris by Cassagnau (1987). In our experience, there are twosouth-western European species resembling this model: I.
ZSC055.fm Page 130 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 131
palustris and I. unifasciatus. They both have a yellowish and /or greenish background with a blackish-violet longitudinalstripe, usually fainter on the last abdominal tergites andgenerally with dark lateral spots on abd. VI. Isotomurusunifasciatus has no additional markings. We refer to thisspecies as I. unifasciatus and, given the variability of the lateralpigmentation, we believe that it corresponds to either theunifasciata or the bimaculata form sensu Stach (1947). Whencompared with I. unifasciatus, I. palustris shows a weakerlongitudinal stripe that is usually fainter, especially on abd.III, and with net-like ornaments on the last abdominal seg-ments. This model is very similar to the drawing proposedby Poinsot-Balaguer (1972) for I. palustris palustris fromCamargue (southern France).
Isotomurus pseudopalustris has a yellowish or greenish back-ground, a dark median dorsal stripe and lateral brownish spots,frequently coalescing on each side of the tergites, even formingadditional stripes. This model is very similar to I. palustris f.maculata of Stach (1947: Pl. XLVIII, fig. 3) and it resemblesthe form maculata (‘Type C’) of Cassagnau (1987). Neither ofthese two descriptions are similar to the syntypes of I. maculatusfrom the original collection of Schäffer (1896), and con-sequently they should not be considered as I. maculatus.
An additional problem is presented by the population fromSiena of I. unifasciatus. Due to the presence of brownish pig-mentation on the lateral side of the tergites, these specimenshave a pigmentation pattern resembling that of I. pseudopalustris,but biochemical and molecular analysis showed that thispopulation clearly belongs to I. unifasciatus (Carapelli et al.1995a).
More than one European species of the genus shows trans-versal bands of pigmentation; this model even seems to bevery common outside the continent, since several species withsimilar body coloration have been described from other partsof the world. We collected three species with this model: I.gallicus n. sp., I. ghibellinus Carapelli et al., 1995 and I. rabiliDeharveng & Lek, 1993. Isotomurus rabili is characterized bya very peculiar pigmentation pattern: specimens have a whitish-yellow background and blackish-violet pigmentation coveringthr. II–III, abd. I–II and the anterior part of abd. III. The othertwo species, I. gallicus and I. ghibellinus, have a yellowish-whitebackground with a dark transverse band on the anterior partof each tergite. Variability among individuals is observed inthe thickness of the band; sometimes the pigmentation isdispersed over the whole tergite, although completely depig-mented specimens of I. ghibellinus, resembling I. prasinus,have been found. The taxonomic position of these two speciesand the frequency of the transversal band model are discussedlater on.
Among the species with uniform coloration, I. prasinus ispale greenish, with antennae and legs distally reddish and twospots on abd. VI. Cassagnau (1987) redescribed I. palustris f.
prasina from France. While the author considered thepigmentation pattern as a useful tool for species diagnosis inthe genus, the lack of the longitudinal stripe was not consideredsufficient to raise this taxon to the rank of species. There arerecords of specimens with a faint longitudinal stripe (Folsom1937; Maynard 1951), but we did not find this character inthe Italian populations sampled.
A very dark coloration is typical of I. hadriaticus n. sp. Thewhole body, including legs, antennae and head (except forthe furca), is homogeneously pigmented, with no markings,stripes or spots. Darker coloration, sometimes hardly visible,is present on the margins of the tergites. Isotomurus hadriaticusis also distinguished from the other species of the palustrisgroup by the presence of four (instead of five) anterosubmedialsetae on abd. IV.
Isotomurus fucicolus (Reuter 1891) and I. cassagnaui(Deharveng & Lek 1993) have a very similar uniform browncolour, but they can be distinguished by their different body sizeand some aspects of their sensorial chaetotaxy (see section:‘Distribution of trichobothria and “S” setae’).
A few years ago, Carapelli et al. (1995b) described the newspecies I. italicus from central Italy, which had a similarpigmentation pattern to I. fucicolus but different features ofthe chaetotaxy (trichobothrial formula and retinacularsetae), based on the redescription of this species provided byDa Gama (1964) and Poinsot-Balaguer (1976). Recently,we have studied five syntypes from the original Reuter’scollection and several specimens of I. fucicolus from Norwaywhich were identical to the Italian specimens of I. italicus, butdifferent from the redescriptions of Da Gama (1964) andPoinsot-Balaguer (1976) which were based on Portuguesematerial. Therefore, the species I. italicus should be regardedas a synonym of I. fucicolus; on the other hand, I. fucicolus sensuDa Gama (1964) and Poinsot-Balaguer (1972, 1976) mostlikely belongs to a different species.
The remaining species are characterized by irregularpigmentation patterns. Isotomurus aetnensis n. sp. shows abrown body colour, with fine dark ribbon-like decorations onthe marginal sides of the tergites (especially abd. III–V) andtwo spots on the lateral sides of abd. VI. In some specimens,there is a faint longitudinal median band, difficult to see bythe naked eye. A similar model of pigmentation, except forthe faint longitudinal band, is shown by I. nebulosus, wherealso antennae, claw and empodium are usually pigmented.
Isotomurus maculatus was first described by Schäffer (1896)as Isotoma palustris f. maculata and afterwards raised to therank of species and placed in the genus Isotomurus by Börner(1903). This taxon was then regarded as a subspecies byPoinsot-Balaguer (1976), and as a form by Cassagnau (1987),although his I. prasinus f. maculata is more similar to I. palustris.We have compared specimens of I. maculatus with the syntypesin the original collection of Schäffer (1896); the pigmentation
ZSC055.fm Page 131 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
132 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
patterns clearly matched, although a certain degree of intra-specific variability was observed even among members ofthe same population. In I. maculatus, the longitudinal stripeis usually well defined on the thoracic tergites, becomingbroader on the abdomen where it forms a series of irregularsymmetrical patches and spots. These patches and spots forma more or less diffuse yellowish, greenish or violet network.
The habit of considering the different pigmentation patternsas one of the most useful discriminating features at the specieslevel is confirmed in this study.
General chaetotaxy and other morphological charactersThe genus includes isotomids with quadridentate mucro andthe presence of abdominal trichobothria. All the species havemore than 10 ventral setae on the manubrium and a bifurcatepalp with four sublobal hairs (Fig. 7A).
Several characters considered in this study do not varyamong specimens. All species analysed have the same numberand arrangement of setae on the labrum: three parallel rowswith 5, 5 and 4 setae, respectively. Other common features toall species are the number of eyes (8 + 8) and the presence of3 + 3 ciliated macrochaetae on both abd. IV and V. Thesemacrochaetae are always arranged in a triangular fashion onabd. IV, and in a row on abd. V. Several multiperforatedsensorial organs (mso) (Deharveng 1980) (Fig. 4C,D) arepresent near the PAO and on the central axis of each tergite.These organs are also frequently observed on the antennaeand on the external surface of the coxae.
All specimens studied have legs with a large number ofordinary setae and an acuminate tenent hair at the distal endof the tibiotarsus. A single microsensillum (ms) is presentnear the coxal/trochanteral articulation and one on the dorsalside of the femur (Fig. 4B).
The shape of the furcula is constant, with slender dentesmeasuring twice the length of the manubrium. The dorsalside of each dens is entirely crenulate with regular cuticularrugosity corresponding to the ‘A’ type shown in Christiansen& Bellinger (1998). The dorsolateral sides of the manubriumare covered with several slightly ciliated mesochaetae, whichare not completely coalesced in the central part. The ventralside of the manubrium is covered by short and smooth micro-chaetae, absent only in the central part, and with a few longersetae (usually two for each side).
There are clearly two groups of species which are differen-tiated according to retinacular chaetotaxy. The first includesI. gallicus and I. rabili with a limited, fixed number of setae andreduced body size. These two species have only two setae onthe retinaculum (Fig. 3A), as reported for I. fucicolus (sensu DaGama 1964 and Poinsot-Balaguer 1972, 1976), which is not,however, the true I. fucicolus, as demonstrated above. Thesecond group, including the rest of the studied species, has aricher and more heterogeneous chaetotaxy, with the number
of setae ranging from 5 to 32 and some degree of intraspecificvariability (Table 1).
A high variability in the number of setae is also observed atthe base of the labium (see Table 1; Fig. 8A–C). Isotomurusnebulosus, I. alticolus, I. ghibellinus, I. palustris, I. maculatus, I.pseudopalustris and I. unifasciatus have the largest number ofsetae (with the greatest variability, from 8 + 8 to 17 + 17,observed in I. palustris); in I. rabili and I. gallicus these setaerange from 5 + 5 to 7 + 7 (Fig. 8B). Isotomurus hadriaticus andI. aetnensis have the same model (7 + 7 to 9 + 9), as do I. alticolus,I. fucicolus and I. prasinus (8 + 8 to 12 + 12) (Fig. 8C), whereasI. cassagnaui has from 4 + 4 to 6 + 6 setae (Fig. 8D). The setaeat the base of the labium are sometimes arranged asymmetricallyon both sides, which reduces the diagnostic value of the character.
The lateral flaps of the ventral tube usually have a fixed numberof setae, 3 + 3 in most species, except for I. rabili (Fig. 3B) andI. gallicus (Fig. 3C) both of which have 5 + 5 setae, and I. alticolus(11 + 11 to 20 + 20).
We have found that the degree of ciliation of the abdominalmacrochaetae varies from species to species. This ciliation isunilateral in I. fucicolus, I. gallicus and I. rabili, and multilateral in theremaining species. Tibiotarsal macrochaetae appear to be ciliatedin I. aetnensis, I. cassagnaui, I. ghibellinus, I. hadriaticus, I. palustris, I.maculatus, I. nebulosus, I. pseudopalustris, I. prasinus and I. unifasciatus.
The ratio between PAO and the diameter of the corneola(PAO/dcA) ranges from 1.5 to 2.0. Antennae are particularlyelongated in I. alticolus. Chaetotaxy of the antennae is com-posed of several ordinary setae (microchaetae) and a few sen-silla. The latter may represent a useful interspecific characterto be investigated, but the relevant interspecific variabilityand the difficulties of observation represent the main obstaclefor a correct and exhaustive description.
There are generally three to five macrochaetae on thetibiotarsus, whose degree of ciliation is variable (Deharveng& Lek 1993); they are a little shorter on the prothoracic legs.The foot complex is composed of a slender claw with oneouter and two lateral teeth, and an empodium where thepresence of the inner tooth is variable among populations andindividuals (Stach 1947; Yosii 1963; Deharveng & Lek 1993;Christiansen & Bellinger 1998). The presence and impor-tance of teeth on the claw and empodium are a particularlycontroversial issue. Schött (1893), Schäffer (1896) and Carl(1899), describing the species I. palustris, reported untoothedclaw and empodium. Börner (1901) found an inner tooth onthe empodial appendage of I. maculatus. Handschin (1929),Folsom (1937), Kos (1937) and Maynard (1951) reported theclaw of I. palustris with lateral teeth, but without the innertooth on the empodium. According to Stach (1947), the clawsof European populations of I. palustris only have the lateralteeth, no inner tooth and an empodial appendage armed witha corner-tooth. Poinsot-Balaguer (1976) showed the claw ofI. palustris palustris with an outer tooth and the empodium
ZSC055.fm Page 132 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 133
Fig. 7 A–D. Maxillary outer lobe and chaetotaxy of dorsal side of dens. —A. Maxillary outer lobe of I. hadriaticus. —B. Dorsal dens of I. ghibellinus. — C. Dorsal dens of I. fucicolus. —D. Dorsal dens of I. cassagnaui.
ZSC055.fm Page 133 Friday, May 25, 2001 3:44 PM
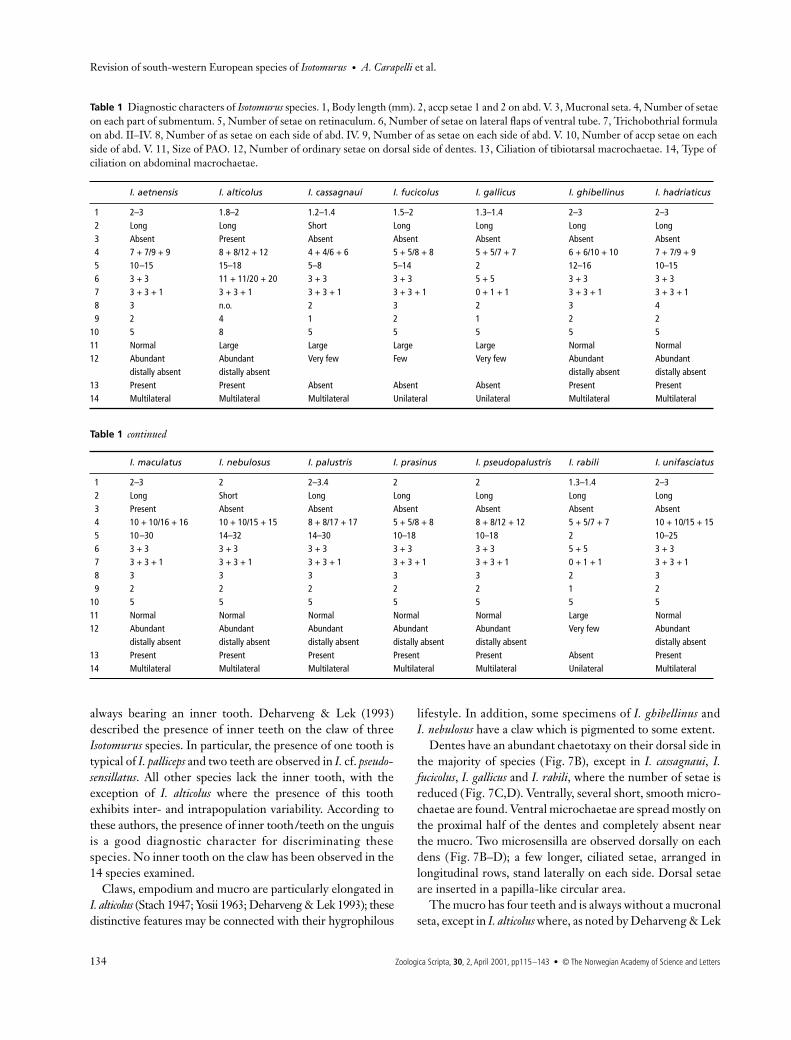
Revision of south-western European species of Isotomurus • A. Carapelli et al.
134 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
always bearing an inner tooth. Deharveng & Lek (1993)described the presence of inner teeth on the claw of threeIsotomurus species. In particular, the presence of one tooth istypical of I. palliceps and two teeth are observed in I. cf. pseudo-sensillatus. All other species lack the inner tooth, with theexception of I. alticolus where the presence of this toothexhibits inter- and intrapopulation variability. According tothese authors, the presence of inner tooth/teeth on the unguisis a good diagnostic character for discriminating thesespecies. No inner tooth on the claw has been observed in the14 species examined.
Claws, empodium and mucro are particularly elongated inI. alticolus (Stach 1947; Yosii 1963; Deharveng & Lek 1993); thesedistinctive features may be connected with their hygrophilous
lifestyle. In addition, some specimens of I. ghibellinus andI. nebulosus have a claw which is pigmented to some extent.
Dentes have an abundant chaetotaxy on their dorsal side inthe majority of species (Fig. 7B), except in I. cassagnaui, I.fucicolus, I. gallicus and I. rabili, where the number of setae isreduced (Fig. 7C,D). Ventrally, several short, smooth micro-chaetae are found. Ventral microchaetae are spread mostly onthe proximal half of the dentes and completely absent nearthe mucro. Two microsensilla are observed dorsally on eachdens (Fig. 7B–D); a few longer, ciliated setae, arranged inlongitudinal rows, stand laterally on each side. Dorsal setaeare inserted in a papilla-like circular area.
The mucro has four teeth and is always without a mucronalseta, except in I. alticolus where, as noted by Deharveng & Lek
Table 1 Diagnostic characters of Isotomurus species. 1, Body length (mm). 2, accp setae 1 and 2 on abd. V. 3, Mucronal seta. 4, Number of setae on each part of submentum. 5, Number of setae on retinaculum. 6, Number of setae on lateral flaps of ventral tube. 7, Trichobothrial formula on abd. II–IV. 8, Number of as setae on each side of abd. IV. 9, Number of as setae on each side of abd. V. 10, Number of accp setae on each side of abd. V. 11, Size of PAO. 12, Number of ordinary setae on dorsal side of dentes. 13, Ciliation of tibiotarsal macrochaetae. 14, Type of ciliation on abdominal macrochaetae.
I. aetnensis I. alticolus I. cassagnaui I. fucicolus I. gallicus I. ghibellinus I. hadriaticus
1 2–3 1.8–2 1.2–1.4 1.5–2 1.3–1.4 2–3 2–32 Long Long Short Long Long Long Long3 Absent Present Absent Absent Absent Absent Absent4 7 + 7/9 + 9 8 + 8/12 + 12 4 + 4/6 + 6 5 + 5/8 + 8 5 + 5/7 + 7 6 + 6/10 + 10 7 + 7/9 + 95 10–15 15–18 5–8 5–14 2 12–16 10–156 3 + 3 11 + 11/20 + 20 3 + 3 3 + 3 5 + 5 3 + 3 3 + 37 3 + 3 + 1 3 + 3 + 1 3 + 3 + 1 3 + 3 + 1 0 + 1 + 1 3 + 3 + 1 3 + 3 + 18 3 n.o. 2 3 2 3 49 2 4 1 2 1 2 2
10 5 8 5 5 5 5 511 Normal Large Large Large Large Normal Normal12 Abundant
distally absentAbundantdistally absent
Very few Few Very few Abundantdistally absent
Abundantdistally absent
13 Present Present Absent Absent Absent Present Present14 Multilateral Multilateral Multilateral Unilateral Unilateral Multilateral Multilateral
Table 1 continued
I. maculatus I. nebulosus I. palustris I. prasinus I. pseudopalustris I. rabili I. unifasciatus
1 2–3 2 2–3.4 2 2 1.3–1.4 2–32 Long Short Long Long Long Long Long3 Present Absent Absent Absent Absent Absent Absent4 10 + 10/16 + 16 10 + 10/15 + 15 8 + 8/17 + 17 5 + 5/8 + 8 8 + 8/12 + 12 5 + 5/7 + 7 10 + 10/15 + 155 10–30 14–32 14–30 10–18 10–18 2 10–256 3 + 3 3 + 3 3 + 3 3 + 3 3 + 3 5 + 5 3 + 37 3 + 3 + 1 3 + 3 + 1 3 + 3 + 1 3 + 3 + 1 3 + 3 + 1 0 + 1 + 1 3 + 3 + 18 3 3 3 3 3 2 39 2 2 2 2 2 1 2
10 5 5 5 5 5 5 511 Normal Normal Normal Normal Normal Large Normal12 Abundant
distally absentAbundantdistally absent
Abundantdistally absent
Abundantdistally absent
Abundantdistally absent
Very few Abundantdistally absent
13 Present Present Present Present Present Absent Present14 Multilateral Multilateral Multilateral Multilateral Multilateral Unilateral Multilateral
ZSC055.fm Page 134 Friday, May 25, 2001 3:44 PM
A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 135
(1993), this seta is present in the Alpine populations, butabsent in the Pyrenean ones.
Distribution of trichobothria and ‘S’ setaeThe distribution of trichobothria is a very stable character inEntomobryidae (Szeptycki 1979); alternative models shouldconsequently be regarded as important features in the taxo-nomy of the genus.
Two different trichobothrial arrangements on abd. II, IIIand IV were observed among the European species exam-ined. The formula 3 + 3/3 + 3/1 + 1 is typical of the species ofthe palustris group, whereas the formula 0 + 0/1 + 1/1 + 1 isfound in I. gallicus and I. rabili. These models appear to besomehow related to the size of the species, the first modeloccurring in the larger species and the second in the smaller
ones. So far, no European species has been found with adifferent model; the third formula known (3 + 3/2 + 2/1 + 1)only seems to be found among South American species(Deharveng & Lek 1993). Although the systematic relation-ships between the two groups are far from being established,Poinsot-Balaguer (1976) found that the formula 0 + 0/1 + 1/1 + 1 is typical of young specimens of I. palustris, whereas theformula 3 + 3/3 + 3/1 + 1 is found in the adults. No specimenof I. palustris with the formula 2 + 2/2 + 2/2 + 2 (observed byStach 1947) has been found in our samplings.
‘S’ setae or setulae are a basic component of the idio-chaetotaxy, comprising those chaetom elements which arepresent from the earliest instars in the Entomobryomorpha(Szeptycki 1978). The distribution of these sensorial setae hasbeen considered in the study of species of Isotomidae at
Fig. 8 A–D. Scanning electron micrographs of the ordinary setae on the submentum of Isotomurus. —A. I. pseudopalustris. —B. I. rabili. — C. I. fucicolus. —D. I. cassagnaui.
ZSC055.fm Page 135 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
136 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
different taxonomic levels (Szeptycki 1972; Deharveng 1977;Potapov 1997). Deharveng (1979) studied the complete distribu-tion of ‘S’ setae in only one species (I. alticolus); Deharveng& Lek (1993) used the arrangement of these sensorial setaeon abd. V to infer phylogenetic relationships among severalAsiatic and European species of the genus. In this analysis,we provide the complete description of the ‘S’ chaetotaxy in14 species.
The sensorial setae are smooth, hyaline and cylindrical;they are recognizable from the ordinary chaetotaxy by thedifferent consistency under the light microscope. Their classi-fication is based on the vicinity to the three rows of ordinarysetae on each tergite: a = anterior, m = medium and p = posterior(Szeptycki 1972). Deharveng & Lek (1993) described fivedifferent ‘Types’ of distribution of ‘S’ setae. We have redefinedtheir original nomenclature by introducing new characters.The species studied in this analysis are listed in bold. Itshould be pointed out that the presence of species fromDeharveng & Lek’s (1993) description is based only on theanalysis of abd. V.
Six different types of distribution of these sensorial setaewere observed.
1 Type 1A: ms1 + al1 + accp1–5 on thr. II and III;ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III;as1, 2 and 4 + accp1–5 on abd. IV; as1–2 + accp1–5 (accp1and accp2 longer than the others) on abd. V. (I. aetnensis, I.aquatilis, I. ghibellinus, I. fucicolus, I. maculatus, I. nebulosus,I. palustris, I. palustris bimaculatus, I. palliceps, I. prasinus,Isotomurus sp. (from Thailand), I. pseudopalustris and I.unifasciatus) (Fig. 2A).2 Type 1B: ms1 + al1 + accp1–5 on thr. II and III;ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III;as1– 4 + accp1–5 on abd. IV; as1–2 + accp1–5 (accp1 andaccp2 longer than the others) on abd. V. (I. hadriaticus)( Fig. 2D).3 Type 2: ms1 + al1 + accp1–5 on thr. II and III; ms1+ accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III; as1 and4 + accp1–5 on abd. IV; as1 + accp1–5 (accp1 and accp2longer than the others) on abd. V. (I. gallicus and I. rabili)( Fig. 2C).4 Type 3: ms1 + al1 + accp1–5 on thr. II and III; ms1 + accp1–5 on abd. I and II; ms1 + accp1–6 on abd. III; as1 and4 + accp1–5 on abd. IV; as1 + accp1–5 (equally short) on abd.V. (I. cassagnaui) ( Fig. 2B).5 Type 4: I. sensillatus (see fig. 1, Deharveng & Lek 1993).6 Type 5: I. alticolus (see fig. 1, Deharveng & Lek 1993).
In the four Types 1A, 1B, 2 and 3, thoracic tergites have thesame arrangement of ‘S’ setae: five accp (1–5), one al and onems. In the first abdominal tergite of I. alticolus, there is anadditional very small seta, almost qualifying as an ms. Likethe other species studied, I. alticolus has five accp and one alsetae on thr. II, but no ms.
The similar chaetotaxy of the thoracic tergites is in linewith an earlier observation that meso- and metathoracicchaetotaxy in the Entomobryomorpha underwent only min-imal changes during evolution (Szeptycki 1972). Metatergumchaetotaxy in the genus Isotomurus is considered to be themost primitive in the family Isotomidae.
The first and second abdominal tergites of Types 1A, 1B,2, 3 and 4 are characterized by five accp (1–5) and one ms;Isotomurus alticolus has six accp and one al on abd. I, and sevenaccp on abd. II.
The third abdominal tergite has six accp and one ms inTypes 1A, 1B, 2 and 3. Isotomurus alticolus has eight accp setaeand two additional setae which are difficult to classify.
Higher variability characterizes the fourth abdominaltergite. Besides the five constantly present accp (1–5), thereare four as on abd. IV in Type 1B (as1– 4), three in Type 1A(as1, as2 and as4), and two in Types 2 and 3 (as1 and as4). Iso-tomurus alticolus has many ‘S’ setae on abd. IV: Deharveng(1979) observed 24 setae which are very difficult to classify.
On abd. V all species have five accp except for I. sensillatuswhere there are four (see Deharveng & Lek 1993). Two of thefive accp (1 and 2) are longer in all species except I. cassagnaui( Type 3) in which all setae have the same length. The accp areagain more numerous (eight) in I. alticolus, but always of thesame length. The species having ‘S’ setae of Types 2 (I. gallicusand I. rabili ), 3 (I. cassagnaui) and 4 (I. sensillatus) have a singleas (1); those belonging to Types 1A and 1B have two as (1 and2), and the species belonging to Type 5 has four as.
I. balteatus, a group of similar coloured species?Since Reuter (1876) first described Isotoma balteata fromFinland, many authors have recorded this species throughoutthe world. Schött (1896), Folsom (1937) and Maynard (1951)reported Isotoma palustris var. balteata, Isotomurus palustris var.balteata and Isotomurus palustris f. balteatus, respectively, fromthe United States. All these descriptions refer to specimenswith the same pigmentation pattern, consisting of a whitishor yellowish background and a dark transverse band of vary-ing width on the anterior parts of each tergite. Yosii (1939)noted the similarity of the colour pattern of I. palustris baltea-tus from Japan and Maynard’s description for the Americanpopulations. In describing the Korean Isotomurus balteatus,Yosii (1939) considered several morphological characters,such as the number and distribution of setae on the labrumand on the lateral flaps of the ventral tube. These specimenswere identified as I. balteatus due to the similarity of thepigmentation patterns, although the author was not able tocompare his specimens with the European ones.
Stach (1947) treated the form balteata as a variety of I.palustris. Winter (1967) described the new South Americanspecies Isotomurus yamaquizensis, which has 3 + 3/3 + 3/1 + 1trichobothria on abd. II–IV and a mucronal seta. Rusek (1971)
ZSC055.fm Page 136 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 137
described Isotomurus quadrisetosus from China with a mucronalseta, 2 + 2/2 + 2/1 + 1 trichobothria and four setae on theretinaculum and the lateral flaps of the ventral tube. TheNearctic Isotomurus bimus (Christiansen & Bellinger 1992,1998) has 5 + 5 to 8 + 8 setae on the ventral tube and 3 +3/3 + 3/1 + 1 trichobothria. The Hawaiian Isotomurus opala(Christiansen & Bellinger 1992) has a model of pigmentationvery similar to I. bimus, from which it differs only in thereduction of the dark band and in the presence of elementsof a longitudinal pattern. Poinsot-Balaguer & Ferard (1983)redescribed I. balteatus from southern France. Specimens havepeculiar trichobothrial formulas (0 + 0/1 + 1/1 + 1 or sometimes0 + 0/0 + 0/1 + 1), 2–5 setae on the retinaculum and 4–6 setaeon the lateral flaps of the ventral tube. They lack the mucronal seta.
Deharveng & Lek (1993) placed I. balteatus in an inde-pendent group together with I. rabili, since both species havetwo retinacular setae, 0 + 0/1 + 1/1 + 1 trichobothria, 6 + 6‘S’ setae on abd. V and no mucronal seta. As the material ofthe original description of I. balteatus is unknown, and prob-ably lost (Greenslade 1994), the correct taxonomic positionof the balteatus-like species described from all over the worldis controversial, and several issues need to be clarified includ-ing: which one of the above-mentioned species with similarpigmentation patterns matches the original description ofReuter; what is the original trichobothrial formula; and whatis the importance of ‘body colour’ for the identification of thisspecies. We had the opportunity of examining old alcohol-preserved specimens of the Linnaniemi (1912) collectionfrom Finland. These specimens are reasonably closely relatedto those of the original description from ‘Fennia orientali’. Inspite of the age of these specimens, the trichobothrial formula(3 + 3/3 + 3/1 + 1) and the mucronal seta are clearly visible. Ifwe consider this model as the typical one for I. balteatus, thenthe specimens described by Poinsot-Balaguer & Ferard(1983) should belong to a different species.
We sampled two different European species with a pig-mentation pattern similar to I. balteatus. The first one, whichwe described as I. ghibellinus, has the typical distribution oftrichobothria (3 + 3/3 + 3/1 + 1) and the same ‘S’ setae on thetergites as the species of the palustris group, but it is differen-tiated from I. balteatus sensu Reuter in the absence of themucronal seta. It also has modified setae in the males, whosetaxonomic significance is discussed in the next section. Thesecond species is identical to I. balteatus sensu Poinsot-Balaguer& Ferard (1983) or sensu Deharveng & Lek (1993). We renamethis species I. gallicus, as it is differentiated from the true I.balteatus in its different trichobothrial formula (0 + 0/1 + 1/1 + 1instead of 3 + 3/3 + 3/1 + 1) and the absence of the mucronal seta.
The existence of species with the same pigmentation pat-tern from different parts of the world makes the diagnosis ofthe balteatus-like species a difficult task. It appears from thisexample that a similar pigmentation pattern has evolved,
possibly independently, in different species of Isotomurusfrom several parts of the world. On the other hand, thesimilarity of pigmentation patterns does not seem to be usefulfor reconstructing phylogenetic relationships (see section:‘Phylogenetic analysis’). The use of other characters, such assensorial chaetotaxy and the mucronal seta, has helped indistinguishing three different species in Europe among thosewith an identical pigmentation pattern. However, the studyof additional characters may provide further data to differen-tiate one species from another.
Taxonomic significance of male modified setaeThe occurrence of male modified setae is a well-knownfeature which has been observed in several Onychiuridae(Fjellberg 1998) and (more rarely) in some Isotomidae genera(Deharveng 1976; Fjellberg 1994).
Four species among those studied here (I. cassagnaui, I. fuci-colus, I. ghibellinus and I. prasinus) possess modified setae onthe III and/or IV abdominal sternite of the males. These hya-line setae were originally found by Deharveng & Lek (1993:figs 5, 6) in I. cassagnaui ( long and not distally subdivided) andI. prasinus (short and apically frayed). We have observed thesesetae also in I. fucicolus and in I. ghibellinus. In I. fucicolus, thereare four spine-like setae on abd. IV (Fig. 6D). In I. ghibellinus,interestingly, the shape and arrangement of these modifiedsetae are different when comparing Italian specimens (allfour populations examined) with specimens from the Frenchpopulation of Axat (Pyrénées). In the Italian populations,they are smooth and grouped on the ventral side of abd. IVonly (Fig. 3E,F). In the specimens from Axat, they are foundscattered on the ventral side of both abd. III and IV and areunilaterally fringed (Fig. 3D; Fig. 4A). The study of Italianspecimens of I. prasinus also revealed a difference in the shapeof male modified setae in comparison to French specimens.In the Italian specimens from Malga Panna, in fact, they arefound ventrally on abd. III and IV and appear very short andapically pointed (Fig. 6C). Their shape is clearly differentfrom those described by Deharveng & Lek (1993) for Frenchspecimens. We also studied new material from Norway whichwas presumably collected near the original topotypic localityfor the species. These Norwegian specimens have apicallypointed setae similar to those of the Italian specimens,although their size is larger (Fig. 6A,B). Italian specimens,however, were much smaller than Norwegian ones and thedifferent size of the modified setae might reflect a differencein developmental stage.
It is difficult to establish the taxonomic significance of thischaracter, which does not occur in all other species of Iso-tomurus studied so far. If the shape and arrangement of thesesetae can be considered diagnostic at the species level, thenwe would have to erect a new species within I. ghibellinuspopulations. The same line of reasoning would lead us to
ZSC055.fm Page 137 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
138 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
erect a new species also for I. prasinus sensu Deharveng & Lek(1993), assuming that the Italian populations are the true I. prasinus,as suggested by the similarity of the male modified setae withScandinavian specimens. However, the observed differencesin male modified setae could also be an example of intraspecificvariability due to particular stages of the reproductive cycle(epitoky, see Cassagnau 1985, 1992; Fjellberg 1988; Zettel& Zettel 1994). Although these latter aspects are yet to bedemonstrated, we prefer to follow a conservative approachand consider these differences as part of the intraspecificvariability until new evidence (morphological and /or molecular)is found to support the description of new species.
Phylogenetic analysis
The assessment of the phylogenetic relationships amongthese 14 species of Isotomurus did not represent the major aimof this study. However, the availability of a number ofcharacters (Table 1), which appeared to be suitable for infer-ring phylogenetic relationships, allowed the construction ofa matrix (Tables 3 and 4) which was used for a cladistic ana-lysis. The matrix includes 14 morphological characters, mostof which are generally used in the taxonomy of the genus(Deharveng & Lek 1993). We voluntarily excluded from thecompilation all the features of pigmentation, which areindeed very useful for species diagnosis (Cassagnau 1987;Deharveng & Lek 1993; Carapelli et al. 1995a,b; this paper),but are generally more affected by parallelism and con-vergence. In addition, given the presence of two differentarrangements of the modified setae of males of I. ghibellinus,we have not included this character in the analysis.
The species Hydroisotoma schäfferi, closely related to Iso-tomurus, was used as an outgroup taxon. Among the 14 char-acters used in the analysis, 12 were parsimony informative:the two uninformative characters (2 and 10) were autapomor-phies in I. cassagnaui and I. alticolus, respectively. Most of thecharacters were binary (1, 2, 3, 4, 5, 10, 11, 13 and 14), butsome were coded as multistate (Table 3); all character statesobserved in the outgroup species were coded as 0. Characters4, 13 and 14 were homoplastic in the tree of Fig. 9.
Three equally parsimonious trees were obtained (25 steps;CI = 0.88; RI = 0.90), whose consensus is presented in Fig. 9;they only differ in the position of I. fucicolus. Due to the lownumber of characters and character states, the tree is partiallyunresolved, but some interesting considerations can be made.Isotomurus alticolus is the most basal species, as it retains sev-eral plesiomorphic character states (see Table 3). Isotomurusalticolus is also distinguished in terms of the number of as(character 9) and accp (character 10) setae on abd. V, whichrepresent two autapomorphies of this species. The evolutionof as setae on abd. V (character 9) implies that the ancestralstate of all Isotomurus species is 2 as setae, and that 4 setae
have evolved as an autapomorphic feature in I. alticolus. Thisreconstruction is concordant with the hypothesis byDeharveng & Lek (1993: p. 249) who described theancestral pattern of the genus as having 2 as setae. A furtherreduction from 2 to 1 as setae is hypothesized in the lineageleading to I. cassagnaui, I. gallicus and I. rabili. All Isotomurusspecies, with the exception of I. alticolus, are joined in a mono-phyletic cluster identified by two synapomorphies: characters6 (number of setae on lateral flaps of ventral tube) and 9(reduction of as setae on abd. V).
The reduced (normal) size of PAO (character 11) and theciliation of tibiotarsal macrochaetae (character 13) define amonophyletic group including nine species. Within thisgroup, five species (I. maculatus, I. nebulosus, I. palustris, I. pseu-dopalustris and I. unifasciatus) are joined by the number ofsetae on each side of the submentum (character 4). In theremaining species, a robust cluster of Pyrenean plus south-western French species is defined by four synapomorphiccharacters: the reduced body length (character 1), thenumber of as setae on abd. IV (character 8) and on abd. V(character 9), and the reduced number of ordinary setae onthe dorsal side of the dens (character 12). Four synapo-morphic features are shared by I. gallicus and I. rabili,including the number of setae on the retinaculum (character5) and on the lateral flaps of the ventral tube (character 6), thetrichobothrial formula (character 7) and the ciliation ofabdominal macrochaetae (character 14).
As a consequence of the limited number of charactersavailable, many suboptimal trees were present. In fact, 804trees were found which were only one step longer (26 steps)than the best trees. In this situation, the possibility that thebest trees are affected by homoplasy is high, and it is neces-sary to analyse more carefully the evolution of some criticalcharacters.
One crucial character in the taxonomy of Isotomurus isthe number of trichobothria in abdominal tergites II–IV(Deharveng & Lek 1993). Two different formulas areobserved in the species studied here: 0 + 1 + 1 (I. gallicus andI. rabili) and 3 + 3 + 1 (all remaining species). In addition,a different formula was found in Hydroisotoma schäfferi(0 + 0 + 1). From the selected topology, it appears that, whilethe formula 0 + 1 + 1 is one of the three synapomorphic char-acters uniting I. gallicus and I. rabili, the group of species withthe 3 + 3 + 1 type is paraphyletic (Fig. 9). To assess the robustnessof this hypothesis, we explored alternative topologies wherethe species with 3 + 3 + 1 trichobothria form a monophyleticclade, and we found that the five equally parsimonious treesshowing this clade are two steps longer than our best trees (27steps; CI = 0.81; RI = 0.83). We therefore believe that thereis strong evidence that the formula 3 + 3 + 1 is the plesiomorphicstate, perhaps evolved in the ancestral species of this Europeangroup. The number of trichobothria in abd. II–IV increases
ZSC055.fm Page 138 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 139
Table 2 Key to the south-western European species of the genus Isotomurus. To make this key easier to use for the direct identification of species, priority has been given to different features of pigmentation, regardless of their phylogenetic significance. Additional characters of chaetotaxy, only visible on mounted specimens, are also provided to aid species diagnosis when pigmentation patterns are identical. The key and colour pictures of the 14 species are available at the web site: http://www.unisi.it/ricerca/dip/collemboli/isotopage/intro.htm
1 Uniform body colour, without bands. 2
Non-uniform body colour. 5
2 Light greenish or yellowish colour. 5 + 5 to 8 + 8 setae on submentum. I. prasinusModified setae on ventrolateral region of abd. III in the male.Four anterosubmedial setae (as1, as3, as4, as5) on abd. IV.
Uniform dark colour. 3
3 Black colour. 7 + 7 to 9 + 9 setae on submentum. I. hadriaticusFive anterosubmedial setae (as1, as2, as3, as4, as5) on abd. IV.
Brownish colour. 4
4 4 + 4 to 6 + 6 setae on submentum. I. cassagnauiModified setae in ventrolateral region of abd. III and IV in the male.Reduced body size. Two anterosubmedial setae (as1, as4) on abd. IV.One anterosubmedial seta (as1) on abd. V; accp4 and 5 in abd. V as long as other accp setae.Very few microchaetae on the dorsal side of dentes.
5 + 5 to 8 + 8 setae on submentum. Four anterosubmedial setae (as1, as3, as4, as5) on abd. IV. I. fucicolusThree anterosubmedial setae (as1, as1 and as4) on abd. IV.Two anterosubmedial setae (as1 and as2) on abd. V; accp4 and 5 of abd. V longer than other accp setae.Modified setae in ventrolateral region of abd. IV in the male.
5 With median longitudinal band. 6
With no longitudinal band. 9
6 Very wide longitudinal band (about one-third of width of tergites). I. alticolus11 + 11 to 20 + 20 setae on lateral flaps of ventral tube.8 + 8 to 12 + 12 setae on submentum.Four anterosubmedial setae on abd. V.
Longitudinal band less than one-fifth of width of tergites and sometimes lacking on the last abdominal tergites; 7generally with dark lateral spots on abd. VI.3 + 3 setae on lateral flaps of ventral tube.Four anterosubmedial setae (as1, as3, as4, as5) on abd. IV.
7 Longitudinal band interrupted or fainter, especially on abd. III, I. palustriswith net-like ornaments on lateral parts of each tergite.
Continuous and very pronounced longitudinal band sometimes lacking on abd. V and VI. 8
81 Abundant brownish pigmentation on dorsolateral side of all tergites. I. pseudopalustris
No or very limited dorsolateral pigmentation. I. unifasciatus
9 With transverse bands. 10
With irregular pigmentation in patches and spots. 12
10 Thr. II to abd. II completely covered in dark pigmentation; anterior part of abd. III and IV with dark pigmentation. I. rabiliTrichobothrial formula on abd. II–IV: 0 + 0/1 + 1/1 + 1.5 + 5 setae on lateral flaps of ventral tube. 5 + 5/7 + 7 setae on submentum.Two anterosubmedial setae (as1, as4) on abd. IV.First two accp setae on abd. V as long as other accp setae.One anterosubmedial seta (as1) on abd. V. Only 2 setae on retinaculum.Very few microchaetae on the dorsal side of dentes.
Only anterior parts of tergites with dark pigmentation. 11
11 Trichobothrial formula on abd. II–IV: 3 + 3/3 + 3/1 + 1. 6 + 6 to 10 + 10 setae on submentum. I. ghibellinusFour anterosubmedial setae (as1, as3, as4, as5) on abd. IV.3 + 3 setae on lateral flaps of ventral tube.Several modified setae in ventrolateral region of abd. III in the male.
Trichobothrial formula on abd. II–IV: 0 + 0/1 + 1/1 + 1. I. gallicus5 + 5 setae on lateral flaps of ventral tube. 5 + 5 to 7 + 7 setae on submentum.Two anterosubmedial setae (as1, as4) on abd. IV. One anterosubmedial seta (as1) on abd. V.Only 2 setae on retinaculum. Very few microchaetae on the dorsal side of dentes.
12 Irregular symmetrical patches of pigmentation on tergites. I. maculatus3 + 3 setae on lateral flaps of ventral tube. 10 + 10 to 16 + 16 setae on submentum.
With irregular stripes on posterior part of abd. III, IV and V. 13
13 Violet irregular stripes. 10 + 10/15 + 15 setae on submentum. Tibiotarsal macrochaetae slightly ciliated. I. nebulosus
Dark brown irregular stripes. 7 + 7 to 9 + 9 setae on submentum. Tibiotarsal macrochaetae ciliated. I. aetnensis
1This key does not apply to the population of I. unifasciatus from Siena, where specimens resemble those of I. pseudopalustris (see critical remarks on this topic in the ‘Discussion’ section).
ZSC055.fm Page 139 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
140 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
during ontogeny in Isotomurus palustris, with juvenile stagesshowing also the 0 + 1 + 1 type (Poinsot-Balaguer 1976).Therefore, it has been suggested that species having 0 + 1 + 1trichobothria in the adults have evolved by neoteny (N. Poinsot-Balaguer, personal communication). This is in agreementwith our hypothesis of evolution of this character, where the0 + 1 + 1 type is the derived state. It is also remarkable, in thisrespect, that the reduction of the number of trichobothria isassociated with a reduction of body size (character 1) and withthe presence of two setae on the retinaculum along the samelineage leading to I. gallicus, I. rabili and, perhaps, I. fucicolussensu Da Gama (1964) and Poinsot-Balaguer (1972, 1976).
The phylogenetic hypothesis produced by this cladisticanalysis is also in large agreement with the taxonomic workby Deharveng & Lek (1993), who divided several European
species of Isotomurus into four groups. Their I. palustris group,in fact, corresponds to a monophyletic clade grouping ninespecies (I. aetnensis, I. ghibellinus, I. hadriaticus, I. maculatus, I.nebulosus, I. palustris, I. prasinus, I. pseudopalustris and I. unifasciatus),and defined by the reduction of the size of PAO (character11). Similarly, the group I. balteatus sensu Deharveng & Lek(1993) corresponds, in our analysis, to a monophyletic cladegrouping I. gallicus and I. rabili, and defined by four synapo-morphies (characters 5, 6, 7 and 14). The two remainingEuropean groups in Deharveng & Lek’s (1993) study (I. alti-colus and I. cassagnaui) contained only one species: their exist-ence is compatible with our analysis and, in particular, issupported by two autapomorphic characters (9 and 10) for I.alticolus and one(2) for I. cassagnaui.
In conclusion, our analysis has yielded a phylogenetichypothesis which is largely concordant with previous taxo-nomic studies on the genus. It represents a good startingpoint for further research on Isotomurus phylogeny if newmorphological characters can be added and if biochemical(allozyme) or molecular (DNA sequences) data sets can be used.
ConclusionsIn spite of the many attempts to clarify the taxonomy of thedifferent species of the genus Isotomurus, there are still severalcontroversial issues. The study of pigmentation patterns andof sensorial chaetotaxy turned out to be useful, but does notsolve all the problems. The occurrence of similar features,such as body colour or chaetotaxy, in geographically isolatedpopulations may be due to parallel evolution. Preliminarysurveys combining biochemical (allozymes) and molecular(DNA sequences) techniques (Carapelli et al. 1995a,b; Fratiet al. 1995) suggest caution in describing different popula-tions of the same species. Such data show that two of themorphological species, I. ghibellinus and I. pseudopalustris, maybe composed of genetically well-differentiated species which
Table 3 Character matrix for the Isotomurus species. Character numbers correspond to those in Table 4 and the cladogram in Fig. 9. ‘?’ denotes missing data. H. schäfferi is the outgroup.
Taxon 1 2 3 4 5 6 7 8 9 10 11 12 13 14
I. aetnensis 0 0 1 0 0 1 2 2 2 0 1 3 1 0I. alticolus 0 0 0 1 0 0 2 ? 3 1 0 3 1 0I. cassagnaui 1 1 1 0 0 1 2 1 1 0 0 1 0 0I. fucicolus 0 0 1 0 0 1 2 2 2 0 0 2 0 1I. gallicus 1 0 1 0 1 2 1 1 1 0 0 1 0 1I. ghibellinus 0 0 1 0 0 1 2 2 2 0 1 3 1 0I. hadriaticus 0 0 1 0 0 1 2 3 2 0 1 3 1 0I. maculatus 0 0 1 1 0 1 2 2 2 0 1 3 1 0I. nebulosus 0 0 1 1 0 1 2 2 2 0 1 3 1 0I. palustris 0 0 1 1 0 1 2 2 2 0 1 3 1 0I. prasinus 0 0 1 0 0 1 2 2 2 0 1 3 1 0I. pseudopalustris 0 0 1 1 0 1 2 2 2 0 1 3 1 0I. rabili 1 0 1 0 1 2 1 1 1 0 0 1 0 1I. unifasciatus 0 0 1 1 0 1 2 2 2 0 1 3 1 0H. schäfferi 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Characters States
1 Body length (mm) 0: > 1.5; 1: ≤ 1.52 Length of accp 1 & 2 on abd. V 0: long; 1: short3 Mucronal seta 0: present; 1: absent4 Setae on each part of submentum 0: ≤ 10; 1: > 105 Setae on retinaculum 0: > two; 1: two6 Setae on each side of lat. flaps of ventr. tube 0: > 10; 1: three; 2: five7 Trichobothria on each hemitergite of abd. II–IV 0: 0/0/1; 1: 0/1/1; 2: 3/3/18 as setae on each hemitergite of abd. IV 0: five; 1: two; 2: three; 3: four9 as setae on each hemitergite of abd. V 0: three; 1: one; 2: two; 3: four
10 accp setae on each hemitergite of abd. V 0: five; 1: eight11 Size of PAO 0: large; 1: normal12 Setae on dorsal side of dentes 0: many and uniformly distributed; 1: very few;
2: few; 3: abundant but distally absent13 Ciliation of tibiotarsal macrochaetae 0: absent; 1: present14 Type of ciliation of abdominal macrochaetae 0: multilateral; 1: unilateral
Table 4 Characters and character states used in the analysis. Character numbers correspond to those in Table 3.
ZSC055.fm Page 140 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 141
are not yet differentiated in pigmentation and chaetotaxiccharacters; interbreeding analysis may be relevant in thisrespect (Cassagnau 1987).
AcknowledgementsWe wish to thank N. Poinsot-Balaguer, L. Deharveng,P. Cassagnau, A. Bedos, S. Lek, M. Niklasson, H. Petersen,J. Zettel and U. Zettel for providing specimens and for
helpful discussions on the problem of the genus Isotomurus.We are deeply indebted to Professor N. P. Kristensen for thematerial of the Haarløv’s collection, to Professor H. Strümpelfor providing specimens from Schäffer’s original collectionand to Professor P. Vilkaama for the original Reuter’s collection.A. Fjellberg provided Scandinavian specimens used for com-parison, and Dr O. Karsholt provided information on theoriginal Müller’s collection. Special thanks go to Professor
Fig. 9 The consensus of the three most parsimonious trees of the Isotomurus species under study (length = 25 steps; CI = 0.88; RI = 0.90) (numbers refer to character numbers as given in Table 3). Symbols as follows: –, synapomorphy.
ZSC055.fm Page 141 Friday, May 25, 2001 3:44 PM

Revision of south-western European species of Isotomurus • A. Carapelli et al.
142 Zoologica Scripta, 30, 2, April 2001, pp115–143 • © The Norwegian Academy of Science and Letters
F. Giusti and Dr G. Manganelli who, with their comments,greatly improved the manuscript. We also wish to thankL. De Carlo for help with sampling and F. Nardi for editingthe homepage and sampling. This work was supported by grantsfrom M.U.R.S.T. (Programmi di ricerca di rilevante interessenazionale: ‘Meccanismi microevolutivi dell’adattamento edell’evoluzione’), the University of Siena (P.A.R. 1999) andC.N.R. (Progetto strategico: ‘Biodiversità ed ecosistemi’) to F.F.
ReferencesÅgren, H. (1903). Zur kenntnis der Apterygotenfauna Süd-Schwedens.
Stettinen Entomologische Zeitung, 64, 113 –176.Axelson, W. M. (1905). Zur Kenntnis der Apterygotenfauna von
Tvärminne. Festschrift für Palmén, 15, 1–46.Axelson, W. M. (1906). Beitrag zur Kenntnis der Collembolenfauna
in der Umgebung Revals. Acta Societatis pro Fauna et Flora Fennica,28, 1–22.
Börner, C. (1901). Zur kenntnis der Apterygotenfauna von Bremenund der Nachbardistrikte. Beitrag zu einer Apterygoten FaunaMitteleuropas. Band der Abhandlungen des NaturwissenschaftlichenVerein zu Bremen, 17, 1 –140.
Börner, C. (1903). Neue Altweltliche Collembolen, nebst Bemerkungenzur systematik der Isotominen und Entomobryinen. Sitzung vomGesellschaft naturforschender Freunde, Berlin, 129 –182.
Carapelli, A., Fanciulli, P. P., Frati, F. & Dallai, R. (1995a). The useof genetic markers for the diagnosis of sibling species in the genusIsotomurus ( Insecta, Collembola). Bollettino di Zoologia, 62, 71–76.
Carapelli, A., Frati, F., Fanciulli, P. P. & Dallai, R. (1995b). Geneticdifferentiation of six sympatric species of Isotomurus (Colembola,Isotomidae); is there any difference in their microhabitat preference?European Journal of Soil Biology, 31 (2), 87– 99.
Carl, J. (1899). Uber Schweizerische Collembola. Revue Suisse deZoologie, 6, 273–362.
Cassagnau, P. (1985). Le polymorphisme des femmes d’Hydroisotomaschaefferi (Krausbauer): un nouveau cas d’épitoquie chez les collem-boles. Annales de la Société Entomologique de France (N.S.), 21 (3),287 – 296.
Cassagnau, P. (1987). A propos des types de coloration chez Isotomuruspalustris (Müller) (Collemboles). Revue d’Ecologie et de Biologie duSol, 24, 85– 89.
Cassagnau, P. & Lauga-Reyrel, F. (1992). Sur le polymorphisme etle cycle sexuel du collembole Superodontella lamellifera (Axelson)dans les Pyrénées. Annales de la Société Entomologique de France(N.S.), 28 (4 ), 371–384.
Christiansen, K. A. & Bellinger, P. F. (1992). Insects of Hawaii,Vol. 15: Collembola. Honolulu: University of Hawaii Press.
Christiansen, K. A. & Bellinger, P. F. (1998). The Collembola of NorthAmerica North of the Rio Grande, 2nd edn. Grinnell, Iowa: GrinnellCollege.
Da Gama, M. M. (1964). Colêmbolos de Portugal continental.Memorias e Estudos do Museu Zoologico da Universitade de Coimbra,292, 1– 252.
Deharveng, L. (1976). Presénce d’un caractère sexuel secondairechez Anurophorus serratus n. sp. (Collembola: Isotomidae). NouvelleRevue D’Entomologie, VI, 2, 109 –111.
Deharveng, L. (1977). Étude chaetotaxique des Collemboles Isotomidae.Premiers résultats. Bulletin du Muséum National d’Histoire Naturelle,455, 597– 619.
Deharveng, L. (1979). Chétotaxie sensillaire et phylogenèse chez lescollemboles Arthropleona. In P. Cassagnau (Ed.) Travaux duLaboratoire d’Ecobiologie des Arthropodes Edaphiques, Vol. 1, 3 (pp. 1–15).Toulouse: Université Paul Sabatier.
Deharveng, L. (1980). Note sur un type d’organites tégumentairesoriginaux rencontrés chez les Isotomidae (Collembola). InR. Dallai (Ed.) First International Seminary on Apterygota,Siena, 13–16 September, 1978 (pp. 59 – 62). Siena: Atti Accademiadei Fisiocritici Siena.
Deharveng, L. & Lek, S. (1993). Remarques sur la morphologie etla taxonomie du genre Isotomurus Börner, 1903 et description dedeux espèces nouvelles de France (Collembola: Isotomidae).Annales de la Société Entomologique de France ( N.S.), 29, 245 – 259.
Fjellberg, A. (1988). Proisotoma rainieri Folsom, 1937 (= Isotoma kisoanaYosii, 1939) syn. nov. An epitokous species of Collembola (Isotomidae).Canadian Journal of Zoology, 66, 965 – 967.
Fjellberg, A. (1994). The Collembola of the Norwegian Artic Islands.Meddelelser Nr. 133. Oslo: Norsk Polarinstitutt.
Fjellberg, A. (1998). The Collembola of Fennoscandia and Denmark.Part I: Poduromorpha. Fauna Entomologica Scandinavica, 35.
Folsom, J. W. (1937). Nearctic Collembola or Springtails of the FamilyIsotomidae. United States National Museum. Bulletin 168. WashingtonD.C.: Smithsonian Institution.
Frati, F., Carapelli, A., Fanciulli, P. P. & Dallai, R. (1995). Thegenus Isotomurus: where molecular markers help to evaluate theimportance of morphological characters for the diagnosis of species.Polskie Pismo Entomologiczne, 64, 41–51.
Greenslade, P. (1994). Collembola. In W. W. K. Houston (Ed.)Zoological Catalogue of Australia, Vol. 22. Protura, Collembola,Diplura. Melbourne, Australia: CSIRO.
Handschin, E. (1919). Ueber die Collembolenfauna der nivalstufe.Revue Suisse de Zoologie, 27, 65 – 98.
Handschin, E. (1929). Urinsekten oder Apterygota (Protura, Col-lembola, Diplura und Thysanura). Die Tierwelt Deutschlands, 16,1–150.
Henriksen, K. L. (1921–37). Oversigt over Dansk Entomologis Historie.Entomologiske Maddelelser, 15, 1– 558.
Kos, F. (1937). Über die polymorphe Aufspaltung der Isotomurini.Prirodoslovne Razprave, 3, 167–237.
Lek, S. & Carapelli, A. (1998). Isotomurus nebulosus, a new Pyreneanspecies of the palustris group (Insecta, Collembola, Isotomidae).Spixiana., 21 (2), 125 –128.
Linnaniemi, W. M. (1912). Die Apterygotenfauna Finlands. II.Spezieller Teil. Acta Societatis Scientiarum Fennicae, 40, 1– 361.
Maddison, W. P. & Maddison, D. R. (1992). MacClade: Analysis ofPhylogeny and Character Evolution (Computer Program and Manual).Sunderland, Massachusetts: Sinauer Associates, Inc.
Maynard, E. A. (1951). The Collembola of New York State. Ithaca, NewYork: Comstock Publishing Co. Inc.
Müller, O. F. (1776). Zoologiae Danicae prodromus, seu animaliumDaniae et Norvegiae indigenarum characteres, nomina et synonymaimprimis popularium, Havniae XXXII. Copenhagen: Hallageriis.
Petersen, H. (1965). The Collembola of the Hansted Reserve, Thy,North Jutland. Taxonomy and ecology (with the description ofa new species and subgenus and a record of a single species ofProtura). Entomologiske Meddelelseries, 30, 313 – 395.
Poinsot-Balaguer, N. (1972). Etude systématique et écologiquedes collemboles Isotomidae de Provence. Annales de la SociétéEntomologique de France, 8, 669– 691.
ZSC055.fm Page 142 Friday, May 25, 2001 3:44 PM

A. Carapelli et al. • Revision of south-western European species of Isotomurus
© The Norwegian Academy of Science and Letters • Zoologica Scripta, 30, 2, April 2001, pp115–143 143
Poinsot-Balaguer, N. (1976). Contribution a l’étude de quelques espècesdu groupe Isotomurus palustris (Müller) (Collembole, Isotomidae).Annales de la Société Entomologique de France ( N.S.), 12, 639 – 652.
Poinsot-Balaguer, N. & Ferard, M. (1983). Contribution a l’etudedu groupe Isotomurus palustris (Müller) (Collembole, Isotomidae)Redescription de Isotomurus balteatus (Reuter, 1876 no. Comb.).Nouvelle Revue D’Entomologie, 2, 147 –156.
Potapov, M. (1997). Towards a new systematics of Isotomidae (Collem-bola). Examples from Pseudanurophorus Stach, 1922 with descriptionof a new furcate species from NE Asia. Pedobiologia, 41, 29 – 34.
Reuter, O. M. (1876). Catalogous praecursorius Poduriarum Fenniae.Meddelanden af Societas pro Fauna et Flora Fennica, 1, 78 – 76.
Reuter, O. M. (1891). Podurider från nordvestra Sibirien, samlade afJ. R., Sahlberg. Öfversigt af Finska Vetenskaps-Societetens Förhandlingar,33, 226 – 229.
Reuter, O. M. (1895). Apterygogenea fennica. Acta Societatis proFauna et Flora Fennica, 11, 1–35.
Rusek, J. (1971). Zweiter Beitrag zur Kenntnis der Collembola (Aptery-gota) Chinas. Acta Entomologica Bohemoslovaca, 68, 108 –137.
Salmon, J. T. (1964). An index to the Collembola. Royal Society of NewZealand Bulletin, 7, 1–644.
Schäffer, C. (1896). Die Collembolen der Umgebung von Hamburgund benachbarter Gebiete. Mitteilungena aus dem NaturhistorischenMuseum in Hamburg, 13, 149 – 216.
Schött, H. (1893). Zur Systematik und Verbreitung palaearctischerCollembola. Kongliga Svenska Vetenskapsakademiens Handlinger, 25,1–100.
Schött, H. (1896). North American Apterygogenea. Proceedings of theCalifornia Academy of Science, 6, 169 –196.
Stach, J. (1947). The Apterygotan fauna of Poland in relation to theworld-fauna of this group of insects. Family Isotomidae. ActaMonographica Musei Historiae Naturalis. Poland: Polish Academyof Sciences and Letters.
Swofford, D. L. (1998). PAUP* — Phylogenetic Analysis Using Parsimony(*And Other Methods), Version 4.0b4a [Computer Software and Manual].Sunderland, Massachusetts: Sinauer Associates.
Szeptycki, A. (1972). Morpho-systematic studies on Collembola. Bodychaetotaxy in the first instar of several genera of the Entomo-bryomorpha. Acta Zoologica Cracoviensia, 17, 341– 372.
Szeptycki, A. (1978). Some remarks of the chaetotaxy of Entomobryidae(Collembola). In R. Dallai ( Ed.) First International Seminary onApterygota, Siena, 13–16 September, 1978 (pp. 63 – 67). Siena: AttiAccademia dei Fisiocritici Siena.
Szeptycki, A. (1979). Chaetotaxy of the Entomobryidae and itsphylogenetic significance. Polska Akademia Nauk, Warsawa-Krakow:Panstwowe Wydawnictwo Naukowe.
Winter, C. (1967). Beitrag zur Kenntnis der neotropischen Collem-bolen fauna. Isotomidae. Senckenbergiana Biologica, 48, 37 – 69.
Yosii, R. (1939). Isotomid Collembola of Japan. Tenthredo, ActaEntomologica, 2, 348 – 392.
Yosii, R. (1963). On some Collembola of Hindukush, with notes onIsotoma Bourlet and its allies. In Results of the Kyoto UniversityScientific Expedition to the Karakoam and Hindukush, 1955, IV InsectFauna of Afghanistan and Hindukush, Art. I (pp. 3 – 42). Kyoto:Kyoto University.
Zettel, U. & Zettel, J. (1994). Seasonal and reproductional polymor-phism in Ceratophysella sigillata (Uzel) (Collembola, Hypogastruridae).Acta Zoologica Fennica, 195, 154 –156.
ZSC055.fm Page 143 Friday, May 25, 2001 3:44 PM




















