
Targeting of HIV-p24 particle-based vaccine into differential skin layers induces distinct arms of...
13
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particle-based vaccine into differential skin layers induces distinct arms of the immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080 ARTICLE IN PRESS G Model JVAC-11753; No. of Pages 13 Vaccine xxx (2011) xxx–xxx Contents lists available at ScienceDirect Vaccine journal homepage: www.elsevier.com/locate/vaccine Targeting of HIV-p24 particle-based vaccine into differential skin layers induces distinct arms of the immune responses Christelle Liard a , Séverine Munier b , Mauricio Arias c , Alix Joulin-Giet a , Olivia Bonduelle a , Darragh Duffy a , Robin J. Shattock c , Bernard Verrier b , Behazine Combadière a,∗,1 a Institut National de la Santé et de la Recherche Médicale, INSERM UMR-S 945 and Université Pierre et Marie Curie (UPMC University Paris 06), Laboratory of Immunity and Infection, 83/91, Boulevard de l’Hôpital, 75013 Paris Cedex 13, France b Institut de Biologie et Chimie des Protéines, UMR 5086 CNRS/UCBL, 7 Passage du Vercors, 69367 Lyon Cedex 07, France c Centre for Infection, Division of Cellular and Molecular Medicine, St George’s University of London, London, UK article info Article history: Received 16 September 2010 Received in revised form 17 April 2011 Accepted 21 April 2011 Available online xxx Keywords: Skin Nanoparticles Cellular responses Mucosal responses Vaccination abstract Skin routes of immunization such as subcutaneous (SC), intradermal (ID) and transcutaneous (TC) admin- istration are utilized for vaccination against various pathogens, without understanding their potential impact on the outcome of immune responses. We demonstrated that SC immunization induced HIV-1 p24 specific IgG in absence of antigen-specific CD8 T cells, whereas the ID route induced both cellular and humoral responses. Interestingly, TC application through empty hair follicular ducts, targeting epidermal Langerhans Cells (LCs), induced major CD8 effector cells, in the absence of IgG. However, high levels of mucosal IgA, were localized in the stratified epithelium of the vagina after TC prime. We propose that re-directing the immune responses by targeting differential skin immunization routes, offers enormous potential for innovative vaccination strategies, especially against HIV. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Confronted with the complexity of immunological events related to infectious diseases, it is clear that multiple new vaccine strategies are required for their control. Appealing approaches for innovation include (i) specific targeting of professional Antigen- Presenting Cells (APCs), (ii) improvement of APCs activation and vaccine compound uptake, (iii) modifications of vaccine route of administration [1]. Professional APCs such as Langerhans cells (LCs) in the epi- dermis and Dendritic Cells (DCs) in the dermis are found at high densities in the skin and can therefore be effective targets for immu- nization [2–4]. Thus, different skin layers, i.e. epidermis, dermis and to a lesser extent subcutis, offer access to a large variety and numbers of APCs. Interestingly, we have recently observed in a Phase I clinical trial in Humans that epidermal Influenza vaccina- tion induced preferentially Influenza-specific CD8 T cells, whereas classical intramuscular (IM) route did not [5]. One can thus ques- tion the impact of the skin route of administration on the outcome ∗ Corresponding author. Tel.: +33 140979887; fax: +33 140779734. E-mail address: [email protected] (B. Combadière). 1 Mailing address: Laboratory of immunity and infection, INSERM UMR-S 945, 91 Blvd de l’Hôpital, 75013 Paris Cedex 13, France. of immune responses and consequently on the control of infectious diseases. The major potential of skin immunization for future clinical tri- als has been illustrated in extensive studies on the development of application methods and devices that would allow targeting of vaccine compounds to a specific layer of the skin [6–8]. Nowadays, most vaccines are still administered subcutaneously (SC) using standard needles [8,9]. SC injection remains quite easy to perform, however due to vaccine loss in the fat layers, current clinical studies focus mostly on reducing antigenic doses. Recently in the vaccine field, there has also been a resurgence of interest for intradermal delivery (ID). Indeed, for some currently licensed vaccines such as Hepatitis B or Rabies vaccines, the protective immunity con- ferred by the ID vaccination route is comparable or superior to standard intramuscular or subcutaneous routes, but allows con- sequent reduction of antigenic doses (generally 1/5th of the dose used SC) [10–12]. Our interest in epidermal targeting is spurred by the fact that LCs might play a key role in the induction of CD8 T cell responses [13–15]. Recent work performed by our group and others on human skin explants has outlined the importance of hair follicles in per- cutaneous penetration processes of vaccine compounds [16,17], and the consequent impact on CD8 cellular responses in a human TC clinical trial [5]. Our early findings demonstrated that opening of hair follicles allowed the administration of nanoparticles that 0264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2011.04.080
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Targeting of HIV-p24 particle-based vaccine into differential skin layers induces distinct arms of...

G
J
Td
CDa
Lb
c
a
ARRAA
KSNCMV
1
rsiPva
ddnanPtct
B
0d
ARTICLE IN PRESSModel
VAC-11753; No. of Pages 13
Vaccine xxx (2011) xxx–xxx
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
argeting of HIV-p24 particle-based vaccine into differential skin layers inducesistinct arms of the immune responses
hristelle Liarda, Séverine Munierb, Mauricio Ariasc, Alix Joulin-Gieta, Olivia Bonduellea,arragh Duffya, Robin J. Shattockc, Bernard Verrierb, Behazine Combadièrea,∗,1
Institut National de la Santé et de la Recherche Médicale, INSERM UMR-S 945 and Université Pierre et Marie Curie (UPMC University Paris 06),aboratory of Immunity and Infection, 83/91, Boulevard de l’Hôpital, 75013 Paris Cedex 13, FranceInstitut de Biologie et Chimie des Protéines, UMR 5086 CNRS/UCBL, 7 Passage du Vercors, 69367 Lyon Cedex 07, FranceCentre for Infection, Division of Cellular and Molecular Medicine, St George’s University of London, London, UK
r t i c l e i n f o
rticle history:eceived 16 September 2010eceived in revised form 17 April 2011ccepted 21 April 2011vailable online xxx
a b s t r a c t
Skin routes of immunization such as subcutaneous (SC), intradermal (ID) and transcutaneous (TC) admin-istration are utilized for vaccination against various pathogens, without understanding their potentialimpact on the outcome of immune responses. We demonstrated that SC immunization induced HIV-1p24 specific IgG in absence of antigen-specific CD8 T cells, whereas the ID route induced both cellular andhumoral responses. Interestingly, TC application through empty hair follicular ducts, targeting epidermal
eywords:kinanoparticlesellular responsesucosal responses
Langerhans Cells (LCs), induced major CD8 effector cells, in the absence of IgG. However, high levels ofmucosal IgA, were localized in the stratified epithelium of the vagina after TC prime. We propose thatre-directing the immune responses by targeting differential skin immunization routes, offers enormouspotential for innovative vaccination strategies, especially against HIV.
© 2011 Elsevier Ltd. All rights reserved.
accination. Introduction
Confronted with the complexity of immunological eventselated to infectious diseases, it is clear that multiple new vaccinetrategies are required for their control. Appealing approaches fornnovation include (i) specific targeting of professional Antigen-resenting Cells (APCs), (ii) improvement of APCs activation andaccine compound uptake, (iii) modifications of vaccine route ofdministration [1].
Professional APCs such as Langerhans cells (LCs) in the epi-ermis and Dendritic Cells (DCs) in the dermis are found at highensities in the skin and can therefore be effective targets for immu-ization [2–4]. Thus, different skin layers, i.e. epidermis, dermisnd to a lesser extent subcutis, offer access to a large variety andumbers of APCs. Interestingly, we have recently observed in ahase I clinical trial in Humans that epidermal Influenza vaccina-
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
ion induced preferentially Influenza-specific CD8 T cells, whereaslassical intramuscular (IM) route did not [5]. One can thus ques-ion the impact of the skin route of administration on the outcome
∗ Corresponding author. Tel.: +33 140979887; fax: +33 140779734.E-mail address: [email protected] (B. Combadière).
1 Mailing address: Laboratory of immunity and infection, INSERM UMR-S 945, 91lvd de l’Hôpital, 75013 Paris Cedex 13, France.
264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2011.04.080
of immune responses and consequently on the control of infectiousdiseases.
The major potential of skin immunization for future clinical tri-als has been illustrated in extensive studies on the developmentof application methods and devices that would allow targeting ofvaccine compounds to a specific layer of the skin [6–8]. Nowadays,most vaccines are still administered subcutaneously (SC) usingstandard needles [8,9]. SC injection remains quite easy to perform,however due to vaccine loss in the fat layers, current clinical studiesfocus mostly on reducing antigenic doses. Recently in the vaccinefield, there has also been a resurgence of interest for intradermaldelivery (ID). Indeed, for some currently licensed vaccines suchas Hepatitis B or Rabies vaccines, the protective immunity con-ferred by the ID vaccination route is comparable or superior tostandard intramuscular or subcutaneous routes, but allows con-sequent reduction of antigenic doses (generally 1/5th of the doseused SC) [10–12].
Our interest in epidermal targeting is spurred by the fact thatLCs might play a key role in the induction of CD8 T cell responses[13–15]. Recent work performed by our group and others on humanskin explants has outlined the importance of hair follicles in per-
le-based vaccine into differential skin layers induces distinct arms of
cutaneous penetration processes of vaccine compounds [16,17],and the consequent impact on CD8 cellular responses in a humanTC clinical trial [5]. Our early findings demonstrated that openingof hair follicles allowed the administration of nanoparticles that

ING Model
J
2 cine x
reshf
aIciltsbcTfispte
aogvappbtewDftteAelmooHftvnb(tar
2
2
RSFprG
ARTICLEVAC-11753; No. of Pages 13
C. Liard et al. / Vac
eadily entered epidermal LCs, localized in the distal hair folliclepithelium [18,19]. As also demonstrated by Glenn et al. [20] tran-cutaneous application of heat-labile enterotoxin from E. coli inuman volunteers, using a patch system, still holds great potential
or a painless, effective and safe vaccination schedule.Challenging specific arms of immunity is however becoming
major goal to control emerging diseases such as HIV (Humanmmunodeficiency Virus). The present postulate is that an HIV vac-ine should enhance both broadly cross-reactive mucosal humoralmmunity as well as systemic and mucosal cytotoxic CD8 cel-ular immune responses, specific to HIV antigens [21,22]. Whilehe cytotoxic effector T-cell arm is needed to control viral dis-emination in both prophylactic and therapeutic approaches [23],roadly reactive anti-HIV neutralizing antibodies (IgG and IgA) arerucially required at the site of viral entry, e.g. mucosal tissues.his has become the most prominent priorities in the HIV-vaccineeld [24–26]. Hence, effective prophylactic vaccination strategieshould be able to elicit mucosa-associated immune responses torevent spreading of the virus at the site of natural primary infec-ion, whereas therapeutic vaccination should also elicit strong andfficient poly-functional effector CD8 T cells [27].
Currently, successful vaccines are of the classical live attenu-ted type or of the inactivated pathogen type, i.e. they are madef naturally occurring “particles” carrying their own unique anti-ens. However, due to their complex nature, such vaccines canary in quality from batch to batch and moreover can inducedverse events [6,28]. Recently, particle-based vaccines have beenroposed for successful immunization [29]. Among the differentarameters that have to be considered for the choice of particle-ased vaccines, two are particularly important for skin vaccination:he size of the particles [30] and their physical and chemical prop-rties. These properties will determine skin penetration [31] andill allow their internalization by professional APCs (LCs or dermalCs) and skin recruited monocytes and neutrophils. The rationale
or the use of nanoparticles (<500 �m in diameter) as delivery sys-ems for skin vaccinations, is based on their excellent ability to beaken up by a variety of phagocytic APCs, which has been repeat-dly demonstrated both in vitro and in vivo (see references in [6]).mong the different types of nanoparticles, are found the non-lastic biodegradable particles made of polymers such as Poly (d,-lactic acid) (PLA) [6,32]. PLA particles can be reduced to an opti-al size for APC uptake (200 nm) and can be coated with proteins
r peptides. They can induce immune responses in mice, rabbitsr macaques, using different antigen models such as HIV-1 p24,IV-1 gp120 or HIV-1 Tat [33,35]. In this study, we investigated
or the first time the outcome of the immune response in regardo its intensity, nature, quality and localization after PLA-basedaccine coated with HIV-p24 antigen (HIV-1 p24 PLA). The immu-ization were performed either (i) through hair-follicle targetingy TC application, as previously shown by our group, [18,19,36],ii) after ID needle injection, and (iii) after SC needle adminis-ration. We demonstrated that routes of skin immunization have
major impact on the intensity, nature and quality of immuneesponses.
. Materials and methods
.1. Mice
Female Balb/c mice (6–8 week old) were purchased from Charlesiver Laboratories (L’Arbresle, France). Animals were housed at thepecific Pathogen Free animal facility (Centre d’Experimentation
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
onctionnelle, Pitié-Salpétriére, Paris). Mice were anesthetizedrior to immunization by intradermal (ID) or transcutaneous (TC)outes, with a mix solution of Xylazine 2% (Bayer AG, Leverkusen,ermany) and Ketamine 8% (Imalgen 1000, Merial, Lyon, France)
PRESSxx (2011) xxx–xxx
in NaCl, according to individual animal weight and duration ofsedation.
For studying the mucosal humoral response, mice werehormonally treated (intramuscular (IM) injection of 2 mg DEPO-PROVERA®, medroxyprogestérone (Pfizer, USA)) 1 week prior topriming immunization and again 1 day before boosting (∼28days post-priming). Hormonal treatment allows synchronizationof mice oestrus cycle for vaginal lavage and IgA monitoring, as pre-viously described in rabbit studies [37] and strongly recommendedby Cranage and Manoussaka [38].
All animals were handled in strict accordance with good ani-mal practice and complied with local animal experimentation andethics committee guidelines. The study was approved by the reviewboard of “Centre d’ Experimentation Fonctionnelle” with authoriza-tion number A75-13-08 (CERFA n◦50-4341; CERFA n◦50-4342).
2.2. Fluorescent nanoparticles/HIV-1 p24 nanoparticles(PLA)/protein
Poly (d, l-lactic acid) of 200 nm (PLA) nanoparticles were pre-pared by solvent diffusion as previously described [34]. Briefly, thepolymer PLA50 (Mn = 50,000 g/mol, Phusis, St Ismier, France) wasdissolved in acetone at a concentration of 2% (w/v). This organicsolution was added drop wise into an aqueous solution, whereinthe PLA precipitated to form particles. Solvents were then removedunder reduced pressure at room temperature. The size and theZeta potential of nanoparticles produced were measured at 25 ◦Cby Quasi-Elastic Light Scattering (QLS) using a ZetaSizer 3000 HS(Malvern Instruments, UK) after dilution of samples in 1 mM NaClsolution. The data were treated by the cumulant method, indicatinga poly dispersity index lower than 0.1.
Protein antigens were passively adsorbed onto PLA nanopar-ticles. In the following experiments, a recombinant HIV-1 p24protein (endotoxin level below 10 EU/mg) produced in E. coli by PXTherapeutics (Grenoble, France) was diluted in PBS at a concentra-tion of 0.2 g/l. PLA nanoparticles diluted in water at a concentrationof 0.5% (w/w) were added to the protein solution and the mixwas incubated 2 h at room temperature (RT) under gentle agi-tation. After this incubation time, formulations were centrifugedRT; during 10 min at 1000 × g. Non-adsorbed protein collected inthe supernatant was quantified using BCA Protein assay (Pierce,Bezons, France).
Nanoparticle pellets were resuspended in PBS at the concentra-tion of 0.4 mg/mL of protein.
To prepare green fluorescent particles, 6-Coumarin was dis-solved in acetone together with PLA at a concentration of 0.2%(w/v). During the precipitation process in the aqueous phase, 6-Coumarin was encapsulated in PLA nanoparticles (Coumarin-6 PLA)as described in Primard et al. [39].
2.3. Immunization protocols
Intradermal (ID) and transcutaneous (TC) administration of vac-cine were performed on the lower part of the flank. Intradermal(ID) immunization was performed with Insulin specific needles(U-100, 29GX1/2′′-0,33X12 mm, Terumo, Belgium) in animal flanksafter shaving. No erythema was observed after shaving, during andafter immunization procedure. Subcutaneous immunization (SC)was performed at the base of the tail, also using Insulin specificneedles U-100. Transcutaneous (TC) immunization was performedfollowing the protocol described by Mahe et al. [19]. Briefly, inabsence of any mechanical or chemical treatment, tape-stripping
le-based vaccine into differential skin layers induces distinct arms of
procedures, using 10 successive adhesive tapes (Lyréco, France),were completed on the lower part of the flank of anesthetized micein order to remove all the hair and part of the stratum corneumprior to the topical application of HIV-1 p24 PLA or Coumarin-6 PLA

ING Model
J
cine xx
(hPt1o
wanbH84t1pbrdtavmpippnif
2
s(c
1asuCoa
2
I(wFAbs1psa(HIf
ARTICLEVAC-11753; No. of Pages 13
C. Liard et al. / Vac
100 �L on 0.5–1 cm2). For all immunization procedure, mice withighly inflamed or damaged skin were removed from the study. TheLA solution was applied directly after tape-stripping, and allowedo completely air dry. Mice remained anesthetized approximatelyh during the procedure avoiding external contacts on the skin andral absorption by leaking.
For each route of immunization (TC, ID, and SC) control miceere treated with the same procedures using the same total
mount of control PLA/mouse (4–5 × 1011). Control PLA origi-ated from the same batch of particles as the p24-coated onesut carried no antigen. Each animal received a priming dose ofIV-1 p24 PLA, independently of vaccine route, corresponding to0 �g of p24 coated-PLA, which represented a total amount of–5 × 1011 PLA/mouse. For ID immunization, animals received 2imes at 1 day apart, 50 �L p24 coated-PLA in each flank (2 times00 �L/flank, 200 �L total per mouse). For SC vaccination 100 �L24 coated-PLA was injected 2 times (1 day apart) directly at thease of the tail (2 times 100 �L, 200 �L total per mouse). For TCoute animals received 2 times 100 �L p24 coated-PLA depositedirectly on the tape-stripped skin (total 200 �L/mouse). Becausehe volume of the vaccine is limited to a maximum of 100 �L perpplication/injection for TC and ID route and also due to the largeolume imposed by p24 coated-PLA batches, we injected each ani-al two times at one day apart to ensure a total amount of 80 �g of
24 per immunization. For boosting, all animals, regardless of prim-ng route of immunization, received subcutaneously 40 �g HIV-124 protein in Incomplete Freund Adjuvant (IFA) 28 days afterriming (100 �L injected at the base of the tail). All skin immu-izations were performed in the same body area not biasing the
mmune response in terms of the distance of the antigen depotrom both spleen and draining inguinal lymph nodes.
.4. Blood and vaginal lavages collection
Blood was collected prior to skin immunizations (pre-immuneamples) and at day 28 post-priming (PP) or day 28 post-boostPB), e.g. 56 days PP. Sera were recovered using 2 successive spinentrifugations.
Vaginal secretions were collected before vaccination but at leastweek after progesterone treatment (pre-immune samples) and
t day 28 PP and day 28 PB. 3 successive gentle lavages with 30 �Lterile PBS were performed in the mouse vaginal cavity (total vol-me recovered 90 �L). Vaginal secretions were treated with 4 �Lomplete� protease inhibitor cocktail tablets (Roche, USA) 30 minn ice and centrifuged 10 min at 1300 rpm to remove mucus. Serand vaginal lavages were stored at −80 ◦C until testing.
.5. Antibody dosage in sera and vaginal lavages
Antibodies were measured by an Enzyme-Linked-mmunosorbent Assay (ELISA). Briefly, 96 well MaxiSorp-platesNunc, Thermofisher Scientific, Rochester, NY, USA) were coatedith 50 �L recombinant HIV-1 p24 (PX-Therapeutics, Grenoble,
rance) at 1 �g/mL in PBS 1× and incubated overnight at 4 ◦C.fter washing, plates were blocked for 1 h at 37 ◦C with 300 �Llocking buffer (PBS, 1% sterile BSA, Euromedex, France). 50 �Lera samples or 45 �L vaginal lavages diluted in assay buffer (PBS×, BSA 1%, 0.05% Tween-20) were incubated overnight at 4 ◦C. Weerformed serial dilutions of 1/10 (sera) or 1/2 (vaginal lavage). Allamples were tested in duplicates. After washing, 100 �L detectionntibody, either biotin conjugated goat anti-mouse IgA (1/5000)
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
Southern biotech, Birmingham, AL, USA) for vaginal lavages ororse Radish Peroxydase (HRP)-conjugated goat anti-mouse IgG,
gG2a, or IgG1 (1/10,000) (Southern Biotech) for sera, were addedor 1 h at 37 ◦C. For IgA monitoring, after washing, Streptavidin-
PRESSx (2011) xxx–xxx 3
HRP (1/200) (R&D Systems, USA) was added for 20 min in blockingbuffer at 37 ◦C. Enzymatic activity was measured by addingtetramethylbenzidine (TMB) (Clinisciences, France), either 10 min(IgA) or 5 min (IgG, IgG1, IgG2a), in the dark. Enzymatic reactionwas stopped by addition of 50 �L stop solution (1N H2SO4, H20).Optical density (OD) of each well was monitored at 450 nm witha Multiscan Ex ELISA reader (Thermofisher) and Ascent Softwareversion 2.6. Cut-off point of the ELISA was calculated as the mean ofpre-immune samples (at the smallest serum dilution) + 3 StandardDeviations (sd). IgG and IgA titers were calculated as the last dilu-tion of sera/vaginal lavages of which OD450 is equal or superior tothe cut-off, using Prism software. Non responder animals had anOD450 beneath cut-off at the smallest dilution of serum/vaginallavage (baseline 0 of the antibody titers in non-responder mice).
2.6. Antigen-specific CD8 cells by pentamer staining
Blood samples were collected in heparin-coated tubes(100 �L/tube) (Heparine choay® 25,000 UI/5 mL, Sanofi Aven-tis, France). Red blood cells were lysed with acetic acid lysisbuffer (155 mM NH4Cl, 10 mM KHCO3, 0.1 mM EDTA, PBS 1×).Fc receptors were blocked using purified anti-CD16/CD32 mAb(clone 2.4G2, BD Biosciences) and PBMCs were stained with HIV-1gag p24 H-2Kd pentamer-PE (Proimmune, UK) (p24: 199–207AMQMLKETINE) for 10 min at RT in PBS-5% Foetal Calf Serum(FCS) (PAA laboratories, Gmbh, Austria). After 3 washes, surfacestaining was performed with anti-mouse CD8� (Ly-2)-PercP-Cy5(clone 53–6.7), anti-mouse CD3-FITC and anti-mouse B220-APC(BD Biosciences, USA) during 20 min at 4 ◦C in PBS–5% FCS. PBMCswere washed in PBS–5% FCS prior to flow cytometric analysis witha FACSCalibur (BD Biosciences). A minimum of 50,000 CD8+ eventswas acquired and analyzed using FlowJo software (Treestar, Inc.,San Carlos, CA).
2.7. Intracellular cytokine assays (ICS)
Splenocytes were collected 10 days after TC, ID or SC immu-nization with HIV-1 p24 PLA. Briefly, spleens were mashed and cellsuspensions were filtered through a 70-mm nylon mesh. Red bloodcells were lysed for 5 min in acetic acid lysis buffer, and remain-ing cells were carefully washed in RPMI 1640 medium 10% FCSand numerated. Splenocytes were cultivated for 2 days at 37 ◦C atthe concentration of 5 × 106 cells/mL in RPMI 1640 medium 10%FCS, in presence of Concanavalin A (Sigma–Aldrich, St-Louis, USA)(2.5 �g/mL).
Cells were washed 3 times with fresh medium and centrifuged(8 min at 1100 rpm) followed by a trypan-blue cell count. Spleno-cytes were cultured 1 week at 37 ◦C, at the concentration of106 cells/mL, in RPMI 1640 10% FCS medium, in presence of 5 U/mLof IL-2 (Roche Diagnostics, CH). Cells were recovered in freshmedium and dead cells were removed in lymphocytes separationmedium (Eurobio, les Ullis, France) by a 20 min centrifugation atRT (2200 rpm, no brake) and then washed in culture media. Totest antigen-specific T cell effector functions, cells were restimu-lated in vitro for 16 h with a mix of two HIV-1 Consensus B Gag15-mer peptides, containing each one class II-IAd restricted HIV-1p24 peptide (p24: 21–35 NAWVKVVEEKAFSPE and p24: 125–139PVGEIYKRWIILGLN) (QBI #304743 #304769, AIDS Research andReference Reagent Program, Division of AIDS, NIAID, NIH) and aclass I-H2d restricted HIV-1 p24 peptide (p24: 199–207 AMQMLKE-TINE) (Eurogentec, Belgium) at a concentration of 2 �g/mL each orwith RPMI 1640 medium.
le-based vaccine into differential skin layers induces distinct arms of
For the final 4 h Brefeldin A was added (Sigma–Aldrich, St Louis,USA) at 5 �g/mL. After washing and a 5 min centrifugation at1500 rpm, cells were stained with conjugated antibodies againstCD3-Pacific Blue (clone 500A2), CD4-PerCp Cy5.5 (RM4-5), CD8�-

ING Model
J
4 cine x
PwwFeIdt
2
aC5I2
scwrwus
2
aoHbac(taPbtbRbUi1Ao(wr(iS
It5gsmthsA
ARTICLEVAC-11753; No. of Pages 13
C. Liard et al. / Vac
E-Cy7(Ly-2) (53–67). After fixation in PFA 4% and permeabilizationith PBS–2% FCS–0.1% saponin (Sigma–Aldrich, St Louis, USA), cellsere stained with conjugated antibodies against IFN-�−APC, IL-2-
ITC, and TNF-�-PE (BD Biosciences, San Jose, USA). At least 50,000vents per sample were analyzed with FlowJo software (Treestar,nc., San Carlos, CA). Boolean combination gating was performed toetermine the frequencies of expression profiles corresponding tohe seven different combinations of cytokines.
.8. Measurement of Th1–Th2 cytokines in supernatants by CBA
Draining lymph node (inguinal) cells were collected 10 daysfter TC, ID and SC immunization with PLA-p24 or control PLA.ells were filtered through a 70-mm nylon mesh, and numerated.× 105 inguinal DLN cells were cultured in vitro for 24 h with a class
-H2d restricted HIV-1 p24 peptide (p24: 199–207 AMQMLKETINE,�g/mL final), or left unstimulated.
After centrifugation (5 min at 1500 rpm), DLN cell-culturedupernatants were removed and stored at −80 ◦C until Th1–Th2ytokine testing. Cytokine profiles (IL-2, IL-4, IL-5, TNF� and IFN�)ere monitored using the mouse Th1–Th2 cytokine kit: Cytomet-
ic Bead Array (CBA) (BD, Biosciences). The mean concentrationas calculated by substracting the concentration of each unstim-lated sample from the concentration of peptide stimulated cellupernatant.
.9. Immunofluorescent microscopy on frozen tissue sections
Histology studies were performed on skin injected sites at 4 hfter TC, ID and SC immunization of 3 × 1010 Coumarin-6 PLAr PBS, or on vagina cryosections 28 days after TC, ID and SCIV-1 p24 PLA prime or 28 days after SC HIV-1 p24-IFA proteinoost. Skin injection sites and vagina were removed and immedi-tely embedded in O.C.TTM wax (Tissue-Tek®, Sakura, Belgium) forryopreservation. Frozen tissues were sequentially cryosectioned5 �m sections) with a Microm HM550 cryostat (Thermo Scien-ific, France). Prior to staining, tissue sections were rehydratednd fixed for 2 min with PFA 1% and then washed for 10 min inBS-1% bovine serum albumin (BSA) (Euromedex, France) followedy 30 min blocking incubation in PBS-3% BSA at room tempera-ure (RT). Endogenous biotin was blocked using the avidin/biotinlocking kit (Vector Laboratories, Burlingame, CA) for 30 min atT (15 min avidin, 15 min biotin). Slides were sequentially incu-ated for 1 h at RT with biotin rat anti-mouse CD205 (AbD Serotec,K) or with biotin rat anti-Langerin/CD207 (Euromedex, France)
n PBS–BSA 1%, 0.1% saponin. Slides were carefully washed in PBS× and then incubated 1 h RT with secondary chicken anti-rat IgG-lexa fluor 594 (Invitrogen, Europe, Paisley, UK) for CD205 staining,r with Texas Red-conjugated streptavidin (Invitrogen) for CD2070.1% saponin). After three washes in PBS, slides were mountedith Vectashield mounting medium containing DAPI (Vector Labo-
atories, UK). Slides were analyzed with a fluorescence microscopeBX51; Olympus, Rungis, France) equipped with an image process-ng and analysis system Qimaging (Media Cybernetics Inc, Silverpring, MD, USA).
The same staining protocol was used for detection of mucosalgA or CD8+CD3+T cells in the vaginal cavity, 28 days post vaccina-ion or 28 days post boost. For IgA monitoring in mucosal tissues,�m vagina cryosections were immunostained respectively withoat anti-mouse IgA (Southern Biotech) and Texas Red-conjugatedtreptavidin (Invitrogen) and mounted with Vectashield mountingedium containing DAPI. For detection of CD8 T cells in vaginal
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
issues, cryosections were immunostained with a mix of purifiedamster anti-mouse CD3 and rat anti-mouse CD8� and a mix ofecondary anti-hamster Alexa 488 and secondary chicken anti-ratlexa 594 (Invitrogen) and mounted with Vectashield mounting
PRESSxx (2011) xxx–xxx
medium containing DAPI. The number of CD8+CD3+T cells wascounted on multiple randomly chosen fields of vagina cryosectionsfrom different animals.
2.10. Statistical analyses
Prism-5 software was used for data handling, analysis,and graphic representations. Data are presented as the meanvalue ± standard deviation. For statistical analyses we used a MannWhitney Test and significance was set at p-value < 0.05. A one-wayANOVA test was used for comparisons of more than 2 groups ofmice, with a 95% significance interval. A Bonferroni correction wasapplied to compare groups 2 by 2.
3. Results
3.1. Differential intensity of cellular and humoral immuneresponses depends on skin immunization route using HIV-1 p24PLA-based vaccine
Our aim was to compare the impact of the skin routes ofimmunization on different arms of immunity. We first verified thespecific localization of the poly (d, l-lactic acid), PLA-based vaccinein the different skin layers, by using Coumarin-6 loaded PLA (greenfluorescence), at 4 h after administration by one of the 3 skin routesof immunization, namely TC (Fig. 1a), ID (Fig. 1b), and SC (Fig. 1c).As previously described by our group [18,19], TC application of flu-orescent particles, preferentially targeted the upper hair-follicularduct (infundibulum), surrounded by the epidermis rich in CD207+
(langerin) LCs, (Fig. 1a) confirming our previous findings. CD207+
LCs are by essence natural resident of the epidermal sheath ofthe hair follicle, as well described by Paus et al. [40,41]. By com-paring skin cryosections from intact skin (Fig. 1a supplementary)and tape-stripped skin, 4 h after application of Coumarin-6-PLA(Fig. 1b supplementary), we observed the same localization anddistribution of LCs along the follicular duct. This suggests that tape-stripping is not responsible for the infiltration of LCs around thehair follicles. ID needle injection targeted the upper to middle der-mis, approximately 30 and 100 �m depth from stratum corneum(SC) (Fig. 1b). Coumarin-6 coated PLA particles injected by the IDroute were surrounded by CD205+ dermal dendritic cells (DDCs)(Fig. 1b). Coumarin-6-PLA localization after SC injection was visu-alized in the fatty layer beneath the dermis (subcutis), at around300 �m from skin surface (Fig. 1c). In contrast to the epidermis anddermis layers, no resident skin APCs such as LCs (CD205+CD207+)or DDCs (CD205+) was present in the subcutis 4 h after injection.
We next evaluated the cellular and humoral immune responsesafter a single administration of PLA coated with HIV-1 p24 protein(HIV-1 p24 PLA). In regard to our previous results on preferentialinduction of CD8 T cells by TC route in human clinical trials, we firstmeasured HIV-1 p24 specific CD8 T cells using a class I-H2d HIV-1p24 pentamer (p24: 199–207 AMQMLKETINE), the main immun-odominant MHC-Class I epitope of HIV-1 p24 antigen in Balb/c mice.We found that the magnitude of HIV-1 p24 effector CD8+ T cellswas significantly higher after TC and ID routes (One way ANOVA,p-value < 0.001, Fig. 2a) compared to the absence of p24-specificCD8 cells by SC route at the peak of the response (day 7 post-immunization) (One way ANOVA, p-value < 0.001, Fig. 2a). HIV-1p24 specific CD8 T cell responses followed the conventional expan-sion and contraction phases as previously described by the groupof Ahmed and co-workers [42].
The magnitude of HIV-1 p24 specific CD8 T cell responses
le-based vaccine into differential skin layers induces distinct arms of
was also measured by ex vivo analysis of cytokine production(Fig. 2b) at day 10 post-immunization. Lymph nodes cell sus-pensions were stimulated with a class I-H2d restricted gag-p24AMQMLKETINE (AA199–207) peptide, for 24 h and cytokines were

ARTICLE IN PRESSG Model
JVAC-11753; No. of Pages 13
C. Liard et al. / Vaccine xxx (2011) xxx–xxx 5
Fig. 1. Localization of Coumarin-6 PLA administered via TC, ID or SC skin routes of vaccination. Coumarin-6 PLA (green fluorescence) localization within skin tissues after TCapplication through empty hair follicle (a), ID (b) and SC (c) injection, is shown in representative fields of 5 �m skin cryosections, 4 h post-immunization. Immunostainningswere performed using anti-CD207-Langerin (Texas Red) or anti-CD205 (Alexa 594) antibodies (RED) to identify LCs, and/or dermal DCs in the epidermis and in the dermis,respectively. (a) TC immunization with Coumarin-6-PLA applied on tape-stripped flank skin, reveals fluorescent particles in the hair follicle openings and along the follicularduct (between 20 and 40 �m depth from the skin surface) surrounded by CD207+ LCs (RED). Epidermis is lineated by a dotted white line and white arrows indicate the hairfollicle localization. (b) ID injected Coumarin-6-PLA are found confined into the dermis layer between 20 and 150 �m below the skin surface. Border of the dermis is lineatedb t CD2d ectivet indic
mo(ArF
bdmtAwg
prmttatru
3S
T
y a dotted white line. Coumarin-6-PLA particles are surrounded by dermal residenepth) beneath hypodermis. Limits of the dermis with epidermis and subcutis resphe epidermis and white arrows indicate the hair follicle localization. Scale: the bar
easured in the supernatant by CBA method. Significant levelsf HIV-1 p24 specific CD8 effector T cells producing cytokinesIL-2, TNF�, IFN�) were found after TC and ID routes (One wayNOVA, p-value < 0.001, Fig. 2b), whereas the SC immunizationoute did not induce any CD8 effector T cells in accordance withig. 2a.
Humoral responses after skin immunization were examinedy measuring HIV-1 p24 specific IgG responses in mice sera 28ays post-priming (Fig. 2c). We found that ID and SC routes, withean specific IgG titers of 104.30 ± 100.20 and 104.28 ± 100.20 respec-
ively, induced a significant level of antibody response (One wayNOVA, p-value < 0.001) compared to TC immunized animals, ofhich the mean specific IgG titre did not differ from the control
roup.In conclusion, we demonstrated that particle-based vaccine
riming via the ID route induced both cellular and humoralesponses. However, epidermal targeting by the TC route, whichostly targets LCs, induced mainly a high CD8 T cell response, in
he absence of detectable systemic humoral responses. In contrast,he SC route induced only significant humoral immunity, in thebsence of vaccine-specific CD8 T cells. These results clearly illus-rate for the first time the impact of differential skin vaccinationoutes on the intensity of cellular and humoral immune responses,sing a particle-based vaccine.
.2. Differential quality of cellular responses induced by TC, ID or
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
C vaccination routes
We next examined whether the quality of the effector/memorycell response in regard to their multi-functionality, could be
05+DCs. (c) SC injected Coumarin-6-PLA located within the fat layer (150–500 �mly, are lineated by a dotted white line. CD207+ LCs (RED) show the localization of
ates 20 �m. Each field is representative of at least 3 independent experiments.
influenced by the skin immunization route. For monitoring ofHIV-1 p24 specific T cell responses in Balb/c mice, the HIV-1antigen p24 contains a class I-H2d HIV-1 p24 immunodominantepitope and 2 class II-IAd HIV-1 p24 sub-dominant epitopes. 10days post-immunization, we performed an in vitro short-expansionof splenocytes. Cells were concomitantly stimulated with the 2class II-IAd HIV-1 p24 sub-dominant epitopes and the class I-H2d
restricted p24 epitope. Frequencies of antigen-specific CD4 andCD8 T cells producing cytokines (IL-2, TNF�, IFN�), were analyzedby flow cytometry, using multi-parameteric intracellular cytokinestaining (ICS).
As depicted in Fig. 3a, HIV-1 p24 specific CD8+ T cells produc-ing cytokines were significantly higher in TC and ID immunizedanimals compared to SC immunized animals (One way ANOVA p-value < 0.05 and p-value < 0.1 respectively for TC and ID). However,despite the similarity observed in the intensity of antigen-specificCD8+ T cells by TC and ID routes, we detected significant differ-ences in the patterns of cytokines profiles produced by p24-specificeffector/memory T cells (Fig. 3b). Indeed, the frequency of triplecytokine-producing CD8+ T cells (TP: IL-2+, TNF�+, IFN�+) was sig-nificantly higher when HIV-1 p24 PLA were administered ID (30%)compared to TC (19%).
The total number of HIV-1 p24 specific CD4+ T cells producingcytokines (Fig. 3c) was significantly higher after ID and SC immu-nization compared to lower immune response observed by the TCroute (One way ANOVA p-value < 0.1) (Fig. 3c). Differences were
le-based vaccine into differential skin layers induces distinct arms of
observed in the polyfunctionality of HIV-1 p24 specific CD4+ Tcells between ID and SC routes, with the latter generating morepolyfunctional T cells (24%) than ID route (11%) (Fig. 3d). Despitethe lower frequency of HIV-1 p24 specific CD4+ T cells induced
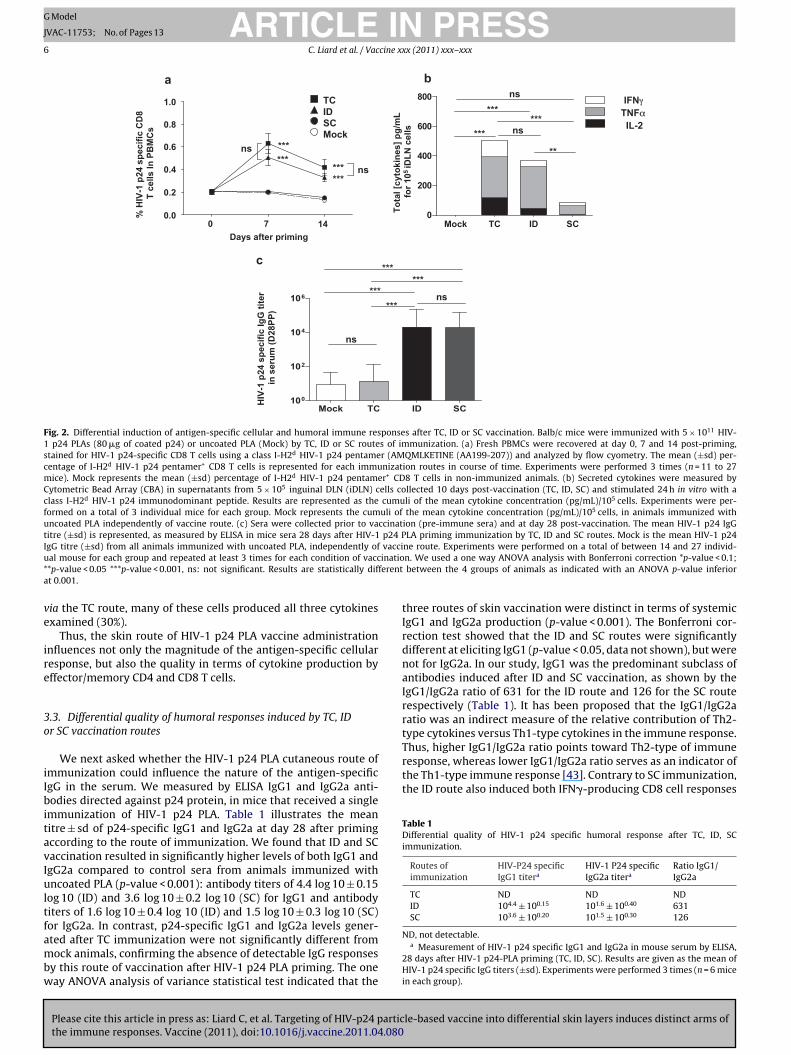
ARTICLE IN PRESSG Model
JVAC-11753; No. of Pages 13
6 C. Liard et al. / Vaccine xxx (2011) xxx–xxx
Mock TC ID SC0
200
400
600
800
Mock TC ID SC100
102
104
106
a
c
0 7 140.0
0.2
0.4
0.6
0.8
1.0
Days after priming
******
******
ns
ns%
HIV
-1 p
24 s
peci
fic C
D8
T ce
lls In
PB
MC
s
******
***ns
ns
***
IFNγTNFαIL-2ns***
Tota
l [cy
toki
nes]
pg/
mL
for 1
05 iD
LNce
lls
HIV
-1 p
24 s
peci
fic Ig
Gtit
er
in s
erum
(D28
PP)
bTCIDSCMock
**
******
ns
Fig. 2. Differential induction of antigen-specific cellular and humoral immune responses after TC, ID or SC vaccination. Balb/c mice were immunized with 5 × 1011 HIV-1 p24 PLAs (80 �g of coated p24) or uncoated PLA (Mock) by TC, ID or SC routes of immunization. (a) Fresh PBMCs were recovered at day 0, 7 and 14 post-priming,stained for HIV-1 p24-specific CD8 T cells using a class I-H2d HIV-1 p24 pentamer (AMQMLKETINE (AA199-207)) and analyzed by flow cyometry. The mean (±sd) per-centage of I-H2d HIV-1 p24 pentamer+ CD8 T cells is represented for each immunization routes in course of time. Experiments were performed 3 times (n = 11 to 27mice). Mock represents the mean (±sd) percentage of I-H2d HIV-1 p24 pentamer+ CD8 T cells in non-immunized animals. (b) Secreted cytokines were measured byCytometric Bead Array (CBA) in supernatants from 5 × 105 inguinal DLN (iDLN) cells collected 10 days post-vaccination (TC, ID, SC) and stimulated 24 h in vitro with aclass I-H2d HIV-1 p24 immunodominant peptide. Results are represented as the cumuli of the mean cytokine concentration (pg/mL)/105 cells. Experiments were per-formed on a total of 3 individual mice for each group. Mock represents the cumuli of the mean cytokine concentration (pg/mL)/105 cells, in animals immunized withuncoated PLA independently of vaccine route. (c) Sera were collected prior to vaccination (pre-immune sera) and at day 28 post-vaccination. The mean HIV-1 p24 IgGtitre (±sd) is represented, as measured by ELISA in mice sera 28 days after HIV-1 p24 PLA priming immunization by TC, ID and SC routes. Mock is the mean HIV-1 p24IgG titre (±sd) from all animals immunized with uncoated PLA, independently of vaccine route. Experiments were performed on a total of between 14 and 27 individ-u inatio* ferenta
ve
ire
3o
iIbitavIultfambw
Thus, higher IgG1/IgG2a ratio points toward Th2-type of immuneresponse, whereas lower IgG1/IgG2a ratio serves as an indicator ofthe Th1-type immune response [43]. Contrary to SC immunization,the ID route also induced both IFN�-producing CD8 cell responses
Table 1Differential quality of HIV-1 p24 specific humoral response after TC, ID, SCimmunization.
Routes ofimmunization
HIV-P24 specificIgG1 titera
HIV-1 P24 specificIgG2a titera
Ratio IgG1/IgG2a
TC ND ND NDID 104.4 ± 100.15 101.6 ± 100.40 631SC 103.6 ± 100.20 101.5 ± 100.30 126
al mouse for each group and repeated at least 3 times for each condition of vacc*p-value < 0.05 ***p-value < 0.001, ns: not significant. Results are statistically dift 0.001.
ia the TC route, many of these cells produced all three cytokinesxamined (30%).
Thus, the skin route of HIV-1 p24 PLA vaccine administrationnfluences not only the magnitude of the antigen-specific cellularesponse, but also the quality in terms of cytokine production byffector/memory CD4 and CD8 T cells.
.3. Differential quality of humoral responses induced by TC, IDr SC vaccination routes
We next asked whether the HIV-1 p24 PLA cutaneous route ofmmunization could influence the nature of the antigen-specificgG in the serum. We measured by ELISA IgG1 and IgG2a anti-odies directed against p24 protein, in mice that received a single
mmunization of HIV-1 p24 PLA. Table 1 illustrates the meanitre ± sd of p24-specific IgG1 and IgG2a at day 28 after primingccording to the route of immunization. We found that ID and SCaccination resulted in significantly higher levels of both IgG1 andgG2a compared to control sera from animals immunized withncoated PLA (p-value < 0.001): antibody titers of 4.4 log 10 ± 0.15
og 10 (ID) and 3.6 log 10 ± 0.2 log 10 (SC) for IgG1 and antibodyiters of 1.6 log 10 ± 0.4 log 10 (ID) and 1.5 log 10 ± 0.3 log 10 (SC)or IgG2a. In contrast, p24-specific IgG1 and IgG2a levels gener-
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
ted after TC immunization were not significantly different fromock animals, confirming the absence of detectable IgG responses
y this route of vaccination after HIV-1 p24 PLA priming. The oneay ANOVA analysis of variance statistical test indicated that the
n. We used a one way ANOVA analysis with Bonferroni correction *p-value < 0.1;between the 4 groups of animals as indicated with an ANOVA p-value inferior
three routes of skin vaccination were distinct in terms of systemicIgG1 and IgG2a production (p-value < 0.001). The Bonferroni cor-rection test showed that the ID and SC routes were significantlydifferent at eliciting IgG1 (p-value < 0.05, data not shown), but werenot for IgG2a. In our study, IgG1 was the predominant subclass ofantibodies induced after ID and SC vaccination, as shown by theIgG1/IgG2a ratio of 631 for the ID route and 126 for the SC routerespectively (Table 1). It has been proposed that the IgG1/IgG2aratio was an indirect measure of the relative contribution of Th2-type cytokines versus Th1-type cytokines in the immune response.
le-based vaccine into differential skin layers induces distinct arms of
ND, not detectable.a Measurement of HIV-1 p24 specific IgG1 and IgG2a in mouse serum by ELISA,
28 days after HIV-1 p24-PLA priming (TC, ID, SC). Results are given as the mean ofHIV-1 p24 specific IgG titers (±sd). Experiments were performed 3 times (n = 6 micein each group).

ARTICLE IN PRESSG Model
JVAC-11753; No. of Pages 13
C. Liard et al. / Vaccine xxx (2011) xxx–xxx 7
a c
15000
20000 ns
**
10000
15000 ns
*
**
19% 24%30% 11%
Abs
olut
e nu
mbe
rs o
f H
IV-1
p24
spe
cific
CD
4 T
cells
se
cret
ing
cyto
kine
s
TC ID SC0
5000
10000
dbTC ID SC
0
5000
Abs
olut
e nu
mbe
rs o
f H
IV-1
p24
spe
cific
CD
8 T
cells
se
cret
ing
cyto
kine
s
30%18%
73%
TP
DPSP
NA
13%57%11%
57% 77%
12%
65%13%
IL2+ TNFα+
IL2+ IFNγ+
45%9%
18%18%
NA 30%40%
30%52%
30%
52%
TP
DPSP
IL2+ TNFα+
IL2+ IFNγ+
IL2+
TNFα+IFNγ+
TNFα+ IFNγ+
33%
10%
57%
46% 64%
NA30%
15%
55%
30%
15%
56%50%
18%
35%
33%11%
18%
49%
47%4%15%
IL2+
TNFα+IFNγ+
TNFα+ IFNγ+
Fig. 3. Differential quality of HIV-1 p24 specific T cells after TC, ID or SC immunization. Splenocytes were harvested 10 days after TC, ID or SC immunization withHIV-1 p24 PLA (80 �g coated p24), cultured and stimulated in vitro as described in materials and methods section. In vitro expanded cells were stimulated witha mix of two HIV-1 Consensus B Gag 15-mer peptides (p24: 21–35 and p24: 125–139), each containing a class II-IAd restricted HIV-1 p24 peptide and a class I-H2d restricted HIV-1 p24 peptide (p24: 199–207). CD8 (a) and CD4 (b) T cell responses were measured by ICS. Histograms (upper panels) represent the mean (±sd)absolute number of HIV-1 p24-specific CD8+ T cells (a) or HIV-1 p24-specific CD4+ T cells (b). Pie charts of cytokines (IL-2, IFN�, TNF�) expression profiles are rep-resented in the lower panels, respectively for HIV-1 p24-specific CD8+ T cells (c) and HIV-1 p24-specific CD4+ T cells (d). Representative data from 2 independentexperiments (n = 6 individual animals/group). ANOVA analysis with Bonferroni correction * denotes p-value < 0.01; **p-value < 0.05, ns: not significant. Statistical signif-i CD4/C (DP)I
aerHno
3e
titmahstcaCmlpcl
cance is set at p-value < 0.05. NA: not applicable. Single positive antigen-specificD4/CD8 T cells secreting IL-2 and TNF� or IL-2 and IFN� or TNF� and IFN�
FN� (TP).
nd IFN�-producing CD4 cell responses, suggesting also the gen-ration of a Th1 type of immune response (Th1 help for the CD8esponses we do observe). We concluded that the skin route ofIV-1 p24 PLA vaccine administration influences not only the mag-itude of the cellular and humoral responses, but also the qualityf antigen-specific antibody responses.
.4. Skin administration route dictates homing of vaccine-specificffector/memory CD8 T cells to the vaginal mucosa
We have demonstrated that only ID and TC routes of vaccina-ion generated significant levels of HIV-1 p24 specific CD8 T cellsn the periphery after single priming. As suggested by the litera-ure, it would also be of great interest for an HIV vaccine to induce
emory cytotoxic T cells in the mucosae, as a first line of defencegainst infected cells [44]. Therefore, we investigated CD8 T celloming to vaginal tissues. At day 28 post-priming by TC, ID or SCkin immunizations with HIV-1 p24 PLA, vaginas were removed andissue cryosections were analyzed for the presence of CD3+CD8+Tells (Fig. 4a). We performed a double staining, green CD8-A488nd red CD3-A594 certifying the CD8 T cell’s detection. Not manyD3+CD8+ T cells were observed in vagina from SC immunized ani-als (Fig. 4a upper right) and for control animals (Fig. 4a upper
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
eft), despite the presence of CD3+ cells, mostly in the Lamina Pro-ria. However, after TC and ID immunizations, some CD3+CD8+ Tells were observed in the lamina propria of mouse vagina (Fig. 4aower pictures). Interestingly, after TC immunization CD3+CD8+ T
CD8 T cells secreting IL-2 or TNF� or INF� (SP); double positive antigen-specific, triple positive antigen-specific CD4/CD8 T cells secreting IL-2 and TNF� and
cells were also found in the Stratified Epithelium (SE) in proximityto the lumen whereas ID induced CD8 T cells were found in theLamina Propria only. We also noticed the presence of many CD3+
cells after ID and TC vaccination, randomly located in the vaginaltissues, i.e. stratified epithelium or lamina propria.
Nevertheless, it is also known that vaginal tissues contain CD8+
T cells under normal conditions. For that reason, we numeratedthe total number of CD8+ T cells in 10 fields of vaginal tissues ran-domly obtained from different animals (n = 3). Fig. 4b illustrates theaverage number CD3+CD8+ T cells numerated per mm2 of vaginaltissue, 28 days after TC, ID or SC immunization with HIV-1 p24 PLAor with uncoated PLA (Mock). We observed a significant increase inthe number of CD8 T cells found in the mucosa for ID and TC routesof skin immunization (p-value < 0.001) and at a lesser extend forthe SC route (p-value < 0.1), compared to baseline number (Mock).However, vaginal mucosa contained fewer CD3+CD8+ T cells afterSC injection compared to ID and TC immunizations (p-value < 0.05and p-value < 0,1 respectively for ID and TC route), as suggestedby histology studies (Fig. 4a). No significant difference was seenbetween the numbers of CD8 T cells numerated in the vagina fromanimals immunized TC and ID.
We demonstrated that only TC and ID routes could increasesignificantly the frequency of vaginal mucosa CD8 T cells. How-
le-based vaccine into differential skin layers induces distinct arms of
ever their specific localization in the tissues: lamina propria versusstratified epithelium might differ. Thus, differential cutaneousimmunizations shaped the antigen-induced imprinting of CD8 Tcells to genital mucosa.

ARTICLE IN PRESSG Model
JVAC-11753; No. of Pages 13
8 C. Liard et al. / Vaccine xxx (2011) xxx–xxx
Fig. 4. Effector/memory HIV-1 specific CD8 T cells migration into genital mucosa after TC and ID routes. Balb/c mice were immunized via TC, ID or SC route with 5 × 1011 HIV-1p24 PLA (80 �g of coated p24). (a) Representative fields of 5 �m vagina cryosections recovered 28 days after HIV-1 p24 PLA immunization (TC, ID, and SC). Vaginal tissuesw xa 488 + +
b D3+CDc ation.c lue < 0
3b
oHPtdo
ere immunostained using Alexa 594-labeled anti-CD3 antibodies (RED) and Alear = 20 �m. LP = Lamina Propria, SE = Stratified Epithelium, L: Lumen. * indicates Cells/mm2 of vaginal tissues, monitored 28 days after HIV-1 p24 PLA skin immunizryosections. We used a Mann–Whitney statistical test with *p-value < 0.05, ***p-va
.5. Preferential mucosal humoral responses by ID and TC routesut not SC route
In addition to CD8 T cell localization in the vagina, the inductionf mucosal IgA antibody is an important consideration for potentialIV vaccines [45]. Therefore we examined the capacity of HIV-1 p24
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
LA to elicit IgA in mouse genital tract after ID, TC or SC immuniza-ions. ELISA assays were performed on mouse vaginal lavages 28ays post-priming (Fig. 5a and c). The one-way ANOVA performedn the mean OD450 after PLA prime clearly indicated differences
-labeled anti-CD8 antibodies (GREEN). CD3 CD8 T cells (YELLOW). Scale: white8+T cells localization. (b) Each bar represents the mean number (±sd) CD8+CD3+TCD8+CD3+T cells were counted on 10 randomly chosen fields from vaginal tissues.001, ns: not significant. Representative of n = 3 mice.
in the level of detectable vaginal IgA according to the route ofadministration (p-value < 0.001) (Fig. 5a). The Boneferroni correc-tion indicated however there was no significant difference betweenTC and SC routes and ID and SC routes after PLA prime. This non-significance could probably be due to the low level of IgA detectedafter priming (mean OD450 at dilution 1/2 in vaginal lavages of 0.20
le-based vaccine into differential skin layers induces distinct arms of
and 0.17 for ID and TC respectively) (Fig. 5a).As the low level of IgA detected after priming did not allow
us to completely conclude on the influence of the skin route ofimmunization on vaginal immune responses (Fig. 5a and c), and

ARTICLE IN PRESSG Model
JVAC-11753; No. of Pages 13
C. Liard et al. / Vaccine xxx (2011) xxx–xxx 9
Fig. 5. Significant induction and specific localization of antigen specific mucosal IgA in mouse vaginal tract after TC and ID routes but not SC route of immunization. Balb/c micewere primo-immunized with 5 × 1011 HIV-1 p24 PLA (80 �g of coated p24) or uncoated PLA (Mock) via distinct cutaneous routes. Animals received a boosting subcutaneousinjection of HIV-1 p24 protein in IFA, 28 days post-priming. Vaginal lavages were collected prior to vaccination and respectively 28 days after priming and 28 days post-HIV-1 p24 protein/IFA boost (i.e. 56 days post HIV-1 p24 PLA priming). Secreted IgA in mouse genital tract were screened by ELISA 28 days post-HIV-1 p24 PLA priming (a)or 28 days post HIV-1 p24 protein/IFA boost (b). Each bar represents the mean OD450 (optical density 450 nm) in vaginal lavages (dilution 1/2) (±sd) for each skin routeof primo-vaccination. Control pre-immune (PI) lavage is the mean OD450 (±sd) from all animals prior to vaccination and mock is the mean OD450 (±sd) from all animalsprimo-immunized with uncoated PLA, independently of vaccine route of vaccination. Boost is the mean OD450 (±sd) from animals primo-immunized with uncoated PLA andboosted SC with HIV-1 p24 protein/IFA. Experiments were performed on a total of between 20 and 28 mice for each group and repeated at least 3 times for each condition ofv e < 0.0p r 28 dP ative o
aeHTmanSrTit1bn
accination. We used a one way ANOVA analysis with Bonferroni correction *p-valuresence of IgA (RED) by immunohistology, 28 days post-priming TC, ID or SC (c) oropria, SE = Stratified Epithelium, L = Lumen.* indicates IgA localization. Represent
lso because of the importance of multiple immunizations to gen-rate mucosal IgA, we decided to perform a boost vaccination usingIV-1 p24 protein (80 �g) and Incomplete Freund adjuvant (IFA).o study the impact of skin-priming routes on IgA production, ani-als were boosted by the subcutaneous route (SC). This method
llows observation of the impact of the skin priming route of immu-ization on boosted IgA immune responses in all animals. Indeed,C boost injection was performed in order to increase secondaryesponses in vivo for detection of IgA. We observed that ID andC immunizations followed by HIV-1 p24 protein/IFA SC boostnjection generated significant levels of p24-specific IgA compared
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
o control animals that received uncoated PLA particle and HIV-p24 protein/IFA SC boost (Fig. 5b). TC and ID primed and SC
oosted animals induced similar level of HIV-1 p24 IgA in vagi-al lavages. However, no significant HIV-1 p24 IgA was detected
5, ***p-value < 0.001, ns: not significant Vagina cryosections were analyzed for theays post HIV-1 p24 protein/IFA SC boost (d). Scale: white bar = 20 �m. LP = Laminaf n = 3 mice.
in vaginal lavages of animals primed and boosted subcutaneously(Fig. 5b).
Thus, only TC and ID skin routes chosen for the first immuniza-tion, but not SC route, generated mucosal HIV-1 p24 specific IgA invaginal lavages, which became highly detectable after HIV-1 p24protein/IFA boost. These results demonstrate again the differentialimpact of the skin-priming route of immunization on consequenthumoral responses.
In order to investigate the vaginal localization of IgA responses,we analyzed cryosections of vagina tissues 28 days after priming(Fig. 5c) and 28 days after boosting (i.e. 56 days after priming)
le-based vaccine into differential skin layers induces distinct arms of
(Fig. 5d). First, the presence of IgA was detected at low levels afteran ID or TC priming step whereas IgA was absent after SC HIV-1 p24PLA immunization and in control animals (Fig. 5c). Theses resultsare in accordance with the measurement of HIV-1 p24 specific IgA

ING Model
J
1 cine x
bl
warslneba
iiv
4
nbsn(aetwtoCeHetl[scwtoCic
TSi
Sw
os
2mHo(
ARTICLEVAC-11753; No. of Pages 13
0 C. Liard et al. / Vac
y ELISA 28 days after priming (Fig. 5a). Observed mucosal IgA wereocalized mainly in the Lamina Propria (Fig. 5c).
After boosting SC with HIV-1 p24 protein/IFA, high levels of IgAere detected in the vagina from animals that were primed by ID
nd TC in contrast with low levels by the SC route (Fig. 5d). Theseesults are again in accordance with the measurement of antigen-pecific IgA by ELISA 28 days after boosting. Strikingly, differentialocalization of IgA was observed in regard of the route of immu-ization. Indeed, high levels of IgA were detected in both, stratifiedpithelium and Lamina Propria after TC priming followed by SCoost whereas IgA were only detected in the vaginal lamina propriafter ID priming followed by SC boost (Fig. 5d).
We demonstrated that pertinent HIV-1 p24 PLA priming admin-stration by different skin routes of immunization impacted on thentensity of IgA in vaginal lavages as well as their localization in theaginal mucosa.
. Discussion
In the present study, using poly (lactic) Acid (PLA)-basedanoparticles (PLA) coated with HIV-1 p24 protein as a particle-ased antigenic model, we demonstrated for the first time that apecific route of vaccine administration determined the intensity,ature, quality and tissue localization of the immune responsessummarized in Table 2). Indeed, “priming” the immune system inway to differentially induce the immune arms could be an inter-sting approach for innovative vaccination strategies. We foundhat the TC route of immunization using the hair follicular path-ay strongly favors the generation of cytotoxic CD8 T cells, in
he absence of IgG induction. These results are in accordance withur recent Phase I clinical trial which showed the superiority ofD8 T cell responses after Influenza vaccine TC application (via thempty hair follicles), compared to the IM route of vaccination [5].owever, we do not exclude that the TC route using follicular pen-tration could induce IgG. Indeed, we have previously publishedhat MVA (Modified Vaccinia Virus Ankara) vaccination by hair fol-icle TC route induced detectable level of neutralizing antibodies19]. Thus, the capacity of this route of immunization to inducepecific IgG responses might also depend on the nature of the vac-ine preparation. Other methods of TC immunization using patchesith an adjuvant such as Cholera Toxin or Lymphotoxin allowed
he induction of high amounts of IgG in the serum. The inductionf antibodies has been described in mice after vaccination with
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
holera Toxin [20], Yersinia Pestis peptides and Cholera Toxin [46],nactivated Influenza virus + oleic or retinoic acid [47] or in humanlinical trials using Cholera Toxin based vaccines [48,49]. Thus, the
able 2ummary of results on differential induction of immune responses by skin routes ofmmunization using particle-based vaccine.
Skin vaccine route Blooda Vaginab
CD8 IgG CD8 IgA
TC +++ none + (d) +++(d)
ID +++ IgG1 » IgG2a ++ (c) +++(c)
SC none IgG1 » IgG2a none none
ummary of the different arms of immunity after TC, ID or SC primo-immunizationith HIV-1 p24-PLA.a Peripheral immune response in the blood was measured by pentamer staining
f HIV-1 p24 specific CD8 T cells at day 7 post-priming (TC, ID, SC) and HIV-1 p24pecific IgG were measured by ELISA 28 days post-priming (TC, ID, SC).
b Presence of CD8 T cells was assessed in the vaginal cavity on tissue cryosections8 days after HIV-1 p24 PLA priming. HIV-1 p24 specific IgA in vaginal lavages wereeasured by ELISA, 28 days after HIV-1 p24 PLA priming (TC, ID, SC), followed byIV-1 p24 protein/IFA SC boost. IgA localization in vaginal mucosa was determinedn tissue crysosections at the same time point. Immune responses were localized
c) in the Lamina Propria (LP), (d) in the LP and stratified epithelium (SE).
PRESSxx (2011) xxx–xxx
TC immunization techniques as well as the type of vaccine prepa-ration used in the different studies cited before could explain thedissimilarities observed in the results.
Our study further showed the superiority of TC route in theinduction of antigen-specific CD8 responses compared to a SCimmunization. The discrepancy of immune responses was how-ever lost when vaccine was injected by the ID route. Dissimilaritieswere however observed in the quality of the specific immuneresponse measured by the proportion of multiple cytokine pro-ducing antigen-specific CD8 T cells. We analyzed on purpose 3specific cytokines: IFN�, TNF� (inflammatory cytokines) and IL-2 (T cell proliferation). Indeed, the simultaneous secretion of thesethree cytokines (polyfonctionality) by virus specific T cells has beenshown to correlate with protection in vivo in various viral mod-els as well described in the review from Seder [50] and especiallydescribed for CD8 T cells by Appay and colleagues [51]. Betts andcolleagues have shown particularly the importance of having T cellssecreting simultaneously these 3 cytokines (polyfonctionality) forgenerating efficient immune responses against HIV infection [51].However, other cytokines could be assessed, notably the produc-tion of IL-4 by T cells which has been shown to correlate with theloss of CD8 T cells effector functions (CTL) and AIDS progression[52]. However, cytokine beads array allowing simultaneous analy-sis of IL-2, IL-4, IL-5, IFN� and TNF�, showed that only IL-2, IFN�and TNF� were detectable after ex vivo stimulation of T cells withHIV-1 p24 peptides (data not shown for IL-4 and IL-5).
In addition, TC primo-immunization was capable of generatingmucosal IgA, which were located in the vaginal epithelium layer.Cellular and molecular mechanisms responsible for the productionof IgA in distant tissues such as the mucosa remain unknown. Thegroup of Belyakov and colleagues proposed that skin LCs would beable to migrate to the mucosal DLN (illiac) or Peyer patches follow-ing a skin epicutaneous immunization, where they could prime Tand B cells with mucosal homing properties (expression of certainreceptors such as CCR9, CCR10, �4�7) [53]. The skin immunizationsite (flank) could also favor the targeting of the antigen to local DLNsas suggested by the group of Guy and colleagues [54]. In addition,two recent in vitro studies also explain the importance of IL-15 forthe proliferation and antibody class switching of B follicular cells.The secretion of IL-15 is interestingly one of the main feature ofskin epidermal LCs in vitro and differentiate them from DDCs [14].
This later result is crucial for designing new vaccine strate-gies against HIV-1, as recently proposed in the overview of HIVfield by Haase [21]. The present postulate is that an HIV vaccineshould enhance both, broadly cross-reactive mucosal humoral andcytotoxic cellular immune responses specific to HIV antigens. Thecytotoxic effector T-cell arm is particularly needed to control viraldissemination in both prophylactic and therapeutic approaches[23]. To induce broadly reactive anti-HIV neutralizing antibodiesat the site of viral entry e.g. mucosal tissues has today become themost urgent priority in the HIV-vaccine field [24–26]. Other stud-ies by Nakanishi et al. monitored the infiltration of effector specificCD8 T cells after a viral infection (using pentamere staining), in thevaginal tissues between 3 and 5 days after infection [55]. Kaufmanet al. monitored effector and memory specific CD8 T cells (pen-tamere and multiparametric flow cytometry) in mouse vagina at2 weeks after IM vaccination [56]. Due to the lack of specific toolssuch as class I-H2d HIV-1 p24 tetramer available for immunohis-tology studies, we decided to simply evaluate the changes in CD8T cell’s frequency in mucosal tissues. Still, we did perform flowcytometry analyses of antigen-specific CD8 T cells from the vaginaby pentamer staining. These experiments were not conclusive due
le-based vaccine into differential skin layers induces distinct arms of
to the low frequencies of specific CD8 T cells detected (data notshown). Thus, we have no evidence demonstrating that infiltratedCD8 T cells observed in the vagina 28 days after priming were spe-cific for HIV-p24. However we did observe a significant increase in

ING Model
J
cine xx
tde
grtmnInMpiVrrttcccsatg[aieiT
tTsatapToidlhtin
bmCvtfmtpAblosar
ARTICLEVAC-11753; No. of Pages 13
C. Liard et al. / Vac
he total number of CD3+CD8+ present in the vaginal mucosa 28ays after priming by TC and ID routes, as shown by the numberingxperiment and the corresponding histogram (Fig. 4b).
Other studies have shown that vaccine route, vector and anti-enic dose play an important role in the dynamics of CD8 T cellesponses after ID, Intravenous (IV) and IM routes [57]. However,hese routes of immunization were very different in terms of tissue
icroenvironment and antigen targeting. For example, IV immu-ization allows direct systemic delivery of the antigen whereas
D route enables targeting of antigen to the skin draining lymphodes. In our work, we voluntarily chose to study skin tissues only.oreover, we used a unique vaccine model at a single dose for
riming of the immune responses to avoid any bias on the dynam-cs of CD8 T cells. In a recent study by our group using Modifiedaccinia Ankara (MVA) vector [58], we compared the ID and IMoutes of injection, in terms of intensity and quality of the immuneesponses (humoral and cellular). We provided in vivo evidencehat the ID route was more potent at inducing high levels of neu-ralizing antibodies and multi-cytokine producing specific CD8 Tells compared to the IM route. In a similar approach, Eyles andolleagues made a first attempt to compare 2 skin routes of vac-ine application, i.e. ID and epidermal delivery routes (patch), usingubunit vaccines against plague administered with cholera toxindjuvant. However, they focused only on monitoring of the sys-emic humoral responses, which was quite limiting, despite havingood protection experiments against a virulent strain of Y. pestis46]. Moreover, this further questions the nature and quality ofll arms of the immune system. Contrary to these previous stud-es which focused on a single arm of the immune response, wextended our comparison to both, humoral (systemic and mucosalmmunity) and antigen-specific T cell responses (CD8 and CD4cells).
It has been clearly established that vaccine efficacy and protec-ion seems to go beyond serum antibodies and also needs to harnesscell immunity and mucosal immunity [59]. Indeed, it was clearly
hown in a clinical trial by the group of Belshe that one dose livettenuated vaccine against Influenza induced a superior protectionhan 2 doses of inactivated influenza vaccine despite lower serumntibodies [60]. It has thus been speculated that this enhancedrotection might be triggered by the cellular arm of immunity.herefore immunomonitoring of vaccines efficacy should also focusn the generation of cytotoxic CD8 T cells as well as mucosalmmunity. We did not perform any protection model studies sinceepending of the challenge model used (tumor injection, viral chal-
enge, bacterial challenge), the requested immune responses wouldave been different. However, this study is the first demonstra-ion of the impact of priming route on the cellular and humoralmmune responses; it opens new avenues for the design of vacci-ation strategies.
It might also be expected that the nature of the antigen couldias the outcome of the resulting immune responses. Indeed, a plas-id or a viral vector would be certainly more potent to enhance
D8 T cell responses by acting on APCs activation and maturationia The Toll-Like Receptors (TLR) [61]. There is no clear demonstra-ion of the nanoparticles’ mode of action and particularly of theate of the antigen once inside the APC (i.e. protein degradation
echanisms, cellular compartment targeting, epitope conserva-ion, implication of cross presentation mechanisms etc.). A recentublication suggested the importance of particle size for optimalPC uptake and processing [30]. Particles of 20–200 nm size canenefit from both CD11c+ DCs transport from the injection site to
ymphoid tissues (LN) as well as from free drainage via the blood
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
r lymphatic vessels to lymphoid resident CD8�+ DCs. This sub-et of lymphoid tissue resident DCs is known to be very potentt cross-presenting antigens in mice and in vitro in Humans, asecently described [62,63]. Moreover, nanoparticles with a defi-
PRESSx (2011) xxx–xxx 11
nite size (∼200 nm) such as Poly-Lactic based particles (PLA) coatedwith HIV-1 p24 (HIV-1 p24 PLA) have been demonstrated to inducegood humoral and cellular responses, as well as mucosal immunity,in mice, rabbits and macaques [33–35,64]. Eventually, it has alsobeen shown that that the immunoglobulin subclass induced afterparticle-based vaccination could be determined by the nanopar-ticle size, with smaller beads (less than 120 nm ∅) preferentiallyinducing a Th1 response and larger beads (greater than 120 nm∅),such as those we used, inducing a Th2 response [65]. This could alsoexplain why we observed mostly the IgG1 specific immunoglobulinsubtype in the serum.
Interestingly, each of the skin vaccine routes of application relieson the presence of specific DC subsets that might play a specific rolein the nature and quality of the immune responses [7,66]. Indeed,epidermal LCs were especially capable of inducing cytotoxic T cellsin human and mice in vitro studies [13,14]. LCs seemed to be morepotent than dermal DCs at inducing cytotoxic effector CD8+ T cellswith increased lytic activies (Perforin, granzyme B). Our recentclinical trial in humans studying LC targeting by TC transfollicu-lar vaccination confirmed the preferential induction of CD8 T cellscompared to the IM route [5,36]. LCs have also been shown to beindispensable for anti-tumor immunity in mice [15] and humans[67].
We provide here evidence, in accordance with a growing bodyof literature in Humans and mice that TC transfollicular immuniza-tion, certainly due to the presence of epidermal LCs, is an importantskin route of vaccination to harness CD8 T cell responses. More-over, we found that the TC route induced specific CD4 T cells inthe absence of significant serum IgG after priming. This is worthmentioning as a recent work by Nakanishi et al. has suggested thatvirus specific CD4+ T helper cells controlled the migration of CTL toinfected mucosal sites [55]. Interestingly, we have also shown thatthe TC route was able to induce mucosal IgA in the vaginal epithe-lium, which is of great interest to prevent viral transmission at themucosal portal of entry by inducing specific immunity that couldbe effective at the early stages of infection [22].
Human dermal DCs have been shown in vitro to preferentiallyprime CD4+ T helper cells, and especially T follicular Helper CD4cells, that in turn induce B cells to become antibody secreting cells.[68]. These in vitro data were confirmed in langerin DTR (diphteriatoxin receptor) mice after skin immunization, where dermal DCslocalized in close proximity to B cell follicles in the outer paracortexof the lymph node whereas LCs were settled in the inner paracortexin the T cell zone of the lymph node [69]. Our results are also inaccordance with the literature, as the ID route was really potent atinducing specific systemic IgG [9].
In Humans the immunological potential of ID immunization hasnow been extensively demonstrated with several licensed vaccinessuch as vaccines against influenza [70–72] and rabies [73]. Theseclinical studies highlight the benefits obtained in terms of dosesparing, immunogenicity and long term protection when using theID immunization route, compared to conventional intramuscularor subcutaneous routes. Interestingly, the ID route induced bothIgG and CD8 T cells with either HIV-1 p24 PLA particles or with anMVA vector [58].
This study opens a new area of vaccine research, based on skinAPCs targeting, to design a “tailor-cut” immune response in orderto fight a specific pathogen. Our results suggest that certain vac-cination concepts need to be renewed and propose new strategiesto struggle against specific infectious diseases that mobilize theprecise arms of immune response required.
le-based vaccine into differential skin layers induces distinct arms of
Conflit of interest
The authors have no conflicting financial interests.

ING Model
J
1 cine x
A
Mgs(pR
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLEVAC-11753; No. of Pages 13
2 C. Liard et al. / Vac
cknowledgments
This work was supported by EU-FP6 health program,uNanoVac “Mucosal HIV vaccines” and EU-FP7 health pro-
ram, CUT’HIVAC “Cutaneous HIV vaccination”. C. Liard wasupported by a grant from Fondation pour la Recherche MedicaleFRM). B. Combadière is a recipient of INSERM-Interface AP/HProgram. Darragh Duffy was a recipient of Agence National de laecherche contre le SIDA (ANRS) grant.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.vaccine.2011.04.080.
eferences
[1] Sumida SM, McKay PF, Truitt DM, Kishko MG, Arthur JC, Seaman MS,et al. Recruitment and expansion of dendritic cells in vivo potentiatethe immunogenicity of plasmid DNA vaccines. J Clin Invest 2004;114(9(November)):1334–42.
[2] Merad M, Ginhoux F, Collin M. Origin, homeostasis and function of Langerhanscells and other langerin-expressing dendritic cells. Nat Rev Immunol 2008;8(12(December)):935–47.
[3] Ochoa MT, Loncaric A, Krutzik SR, Becker TC, Modlin RL. Dermal dendriticcells” comprise two distinct populations: CD1+ dendritic cells and CD209+macrophages. J Invest Dermatol 2008;128(9 (September)):2225–31.
[4] Zaba LC, Krueger JG, Lowes MA. Resident and “inflammatory” dendritic cells inhuman skin. J Invest Dermatol 2009;129(2 (February)):302–8.
[5] Combadiere B, Vogt A, Mahe B, Costagliola D, Hadam S, Bonduelle O, et al.Preferential amplification of CD8 effector-T cells after transcutaneous applica-tion of an inactivated influenza vaccine: a randomized phase I trial. PLoS One2010;5(5):e10818.
[6] Combadiere B, Mahe B. Particle-based vaccines for transcutaneous vaccination.Comp Immunol Microbiol Infect Dis 2008;31(2–3 (March)):293–315.
[7] Romani N, Thurnher M, Idoyaga J, Steinman RM, Flacher V. Targeting of antigensto skin dendritic cells: possibilities to enhance vaccine efficacy. Immunol CellBiol 2010;88(4 (May)):424–30.
[8] Nicolas JF, Guy B, Intradermal. epidermal and transcutaneous vaccina-tion: from immunology to clinical practice. Expert Rev Vaccines 2008;7(8(October)):1201–14.
[9] Laurent PE, Bonnet S, Alchas P, Regolini P, Mikszta JA, Pettis R, et al. Evalua-tion of the clinical performance of a new intradermal vaccine administrationtechnique and associated delivery system. Vaccine 2007;25(52 (December17)):8833–42.
10] Egemen A, Aksit S, Kurugol Z, Erensoy S, Bilgic A, Akilli M. Low-dose intrader-mal versus intramuscular administration of recombinant hepatitis B vaccine:a comparison of immunogenicity in infants and preschool children. Vaccine1998;16(16 (October)):1511–5.
11] Sabchareon A, Chantavanich P, Pasuralertsakul S, Pojjaroen-Anant C,Prarinyanupharb V, Attanath P, et al. Persistence of antibodies in childrenafter intradermal or intramuscular administration of preexposure primary andbooster immunizations with purified Vero cell rabies vaccine. Pediatr Infect DisJ 1998;17(11 (November)):1001–7.
12] Mitragotri S. Immunization without needles. Nat Rev Immunol 2005;5(12(December)):905–16.
13] Ratzinger G, Baggers J, de Cos MA, Yuan J, Dao T, Reagan JL, et al. Maturehuman Langerhans cells derived from CD34+ hematopoietic progenitors stim-ulate greater cytolytic T lymphocyte activity in the absence of bioactiveIL-12p70, by either single peptide presentation or cross-priming, than dodermal-interstitial or monocyte-derived dendritic cells. J Immunol 2004;173(4(August 15)):2780–91.
14] Klechevsky E, Morita R, Liu M, Cao Y, Coquery S, Thompson-Snipes L, et al.Functional specializations of human epidermal Langerhans cells and CD14+dermal dendritic cells. Immunity 2008;29(3 (September 19)):497–510.
15] Stoitzner P, Green LK, Jung JY, Price KM, Tripp CH, Malissen B, et al. Tumorimmunotherapy by epicutaneous immunization requires langerhans cells. JImmunol 2008;180(3 (February 1)):1991–8.
16] Wosicka H, Cal K. Targeting to the hair follicles: current status and potential. JDermatol Sci 2010;57(2 (February)):83–9.
17] Knorr F, Lademann J, Patzelt A, Sterry W, Blume-Peytavi U, Vogt A. Follicu-lar transport route – research progress and future perspectives. Eur J PharmBiopharm 2009;71(2 (February)):173–80.
18] Vogt A, Combadiere B, Hadam S, Stieler KM, Lademann J, Schaefer H, et al.40 nm, but not 750 or 1500 nm, nanoparticles enter epidermal CD1a+ cellsafter transcutaneous application on human skin. J Invest Dermatol 2006;126(6
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
(June)):1316–22.19] Mahe B, Vogt A, Liard C, Duffy D, Abadie V, Bonduelle O, et al. Nanoparticle-
based targeting of vaccine compounds to skin antigen-presenting cells byhair follicles and their transport in mice. J Invest Dermatol 2009;129(5(May)):1156–64.
[
PRESSxx (2011) xxx–xxx
20] Glenn GM, Rao M, Matyas GR, Alving CR. Skin immunization made possible bycholera toxin. Nature 1998;391(6670 (February 26)):851.
21] Haase AT. Targeting early infection to prevent HIV-1 mucosal transmission.Nature 2010;464(7286 (March 11)):217–23.
22] Virgin HW, Walker BD. Immunology and the elusive AIDS vaccine. Nature2010;464(7286 (March 11)):224–31.
23] Barouch DH. Challenges in the development of an HIV-1 vaccine. Nature2008;455(7213 (October 2)):613–9.
24] Belyakov IM, Berzofsky JA. Immunobiology of mucosal HIV infection and thebasis for development of a new generation of mucosal AIDS vaccines. Immunity2004;20(3 (March)):247–53.
25] Montefiori DC, Morris L, Ferrari G, Mascola JR, Neutralizing. other antiviralantibodies in HIV-1 infection and vaccination. Curr Opin HIV AIDS 2007;2(3(May)):169–76.
26] Haynes BF, Shattock RJ. Critical issues in mucosal immunity for HIV-1 vaccinedevelopment. J Allergy Clin Immunol 2008;122(1 (July)):3–9, quiz 10-1.
27] Jespers V, Harandi AM, Hinkula J, Medaglini D, Le Grand R, Stahl-Hennig C,et al. Assessment of mucosal immunity to HIV-1. Expert Rev Vaccines 2010;9(4(April)):381–94.
28] Hilleman MR. Vaccines in historic evolution and perspective: a narrative ofvaccine discoveries. J Hum Virol 2000;3(2 (March–April)):63–76.
29] Levine MM, Sztein MB. Vaccine development strategies for improvingimmunization: the role of modern immunology. Nat Immunol 2004;5(5(May)):460–4.
30] Manolova V, Flace A, Bauer M, Schwarz K, Saudan P, Bachmann MF. Nanopar-ticles target distinct dendritic cell populations according to their size. Eur JImmunol 2008;38(5 (May)):1404–13.
31] Bos JD, Meinardi MM. The 500 Dalton rule for the skin penetration of chemicalcompounds and drugs. Exp Dermatol 2000;9(3 (June)):165–9.
32] Nandedkar TD. Nanovaccines: recent developments in vaccination. J Biosci2009;34(6 (December)):995–1003.
33] Guillon C, Mayol K, Terrat C, Compagnon C, Primard C, Charles MH, et al. For-mulation of HIV-1 Tat and p24 antigens by PLA nanoparticles or MF59 impactsthe breadth, but not the magnitude, of serum and faecal antibody responses inrabbits. Vaccine 2007;25(43 (October 23)):7491–501.
34] Lamalle-Bernard D, Munier S, Compagnon C, Charles MH, Kalyanaraman VS,Delair T, et al. Coadsorption of HIV-1 p24 and gp120 proteins to surfactant-free anionic PLA nanoparticles preserves antigenicity and immunogenicity. JControl Release 2006 Sep 28;115(1):57–67.
35] Ataman-Onal Y, Munier S, Ganee A, Terrat C, Durand PY, Battail N, et al.Surfactant-free anionic PLA nanoparticles coated with HIV-1 p24 proteininduced enhanced cellular and humoral immune responses in various animalmodels. J Control Release 2006;112(2 (May 15)):175–85.
36] Vogt A, Mahe B, Costagliola D, Bonduelle O, Hadam S, Schaefer G, et al. Transcu-taneous anti-influenza vaccination promotes both CD4 and CD8 T cell immuneresponses in humans. J Immunol 2008;180(3 (February 1)):1482–9.
37] Cranage MP, Fraser CA, Stevens Z, Huting J, Chang M, Jeffs SA, et al. Repeatedvaginal administration of trimeric HIV-1 clade C gp140 induces serum andmucosal antibody responses. Mucosal Immunol 2010;3(1 (January)):57–68.
38] Cranage MP, Manoussaka M. Modern mucosal vaccines, adjuvants and micro-bicides. Expert Rev Anti Infect Ther 2009;7(1 (February)):21–3.
39] Primard C, Rochereau N, Luciani E, Genin C, Delair T, Paul S, et al.Traffic of poly(lactic acid) nanoparticulate vaccine vehicle from intestinalmucus to sub-epithelial immune competent cells. Biomaterials 2010;31(23(August)):6060–8.
40] Paus E, Nustad K, Bormer OP. Epitope mapping and affinity estimation of83 antibodies against prostate-specific antigen. Tumour Biol 1999;20(Suppl1):52–69.
41] Paus R, Christoph T, Muller-Rover S. Immunology of the hair follicle: ashort journey into terra incognita. J Investig Dermatol Symp Proc 1999;4(3(December)):226–34.
42] Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differenti-ation: implications for vaccine development. Nat Rev Immunol 2002;2(4(April)):251–62.
43] Finkelman FD, Holmes J, Katona IM, Urban Jr JF, Beckmann MP, Park LS, et al.Lymphokine control of in vivo immunoglobulin isotype selection. Annu RevImmunol 1990;8:303–33.
44] Belyakov IM, Kuznetsov VA, Kelsall B, Klinman D, Moniuszko M, Lemon M,et al. Impact of vaccine-induced mucosal high-avidity CD8+ CTLs in delay ofAIDS viral dissemination from mucosa. Blood 2006;107(8 (April 15)):3258–64.
45] Shattock RJ, Moore JP. Inhibiting sexual transmission of HIV-1 infection. NatRev Microbiol 2003;1(1 (October)):25–34.
46] Eyles JE, Elvin SJ, Westwood A, Lebutt CS, Alpar HO, Somavarapu S, et al.Immunisation against plague by transcutaneous and intradermal applicationof subunit antigens. Vaccine 2004;22(31–32 (October 22)):4365–73.
47] Skountzou I, Quan FS, Jacob J, Compans RW, Kang SM. Transcutaneousimmunization with inactivated influenza virus induces protective immuneresponses. Vaccine 2006;24(35–36 (August 28)):6110–9.
48] Glenn GM, Taylor DN, Li X, Frankel S, Montemarano A, Alving CR. Transcuta-neous immunization: a human vaccine delivery strategy using a patch. Nat Med
le-based vaccine into differential skin layers induces distinct arms of
2000;6(12 (December)):1403–6.49] Glenn GM, Villar CP, Flyer DC, Bourgeois AL, McKenzie R, Lavker RM, et al.
Safety and immunogenicity of an enterotoxigenic Escherichia coli vaccine patchcontaining heat-labile toxin: use of skin pretreatment to disrupt the stratumcorneum. Infect Immun 2007;75(5 (May)):2163–70.

ING Model
J
cine xx
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLEVAC-11753; No. of Pages 13
C. Liard et al. / Vac
50] Seder RA, Darrah PA, Roederer M. T-cell quality in memory and protection:implications for vaccine design. Nat Rev Immunol 2008;8(4 (April)):247–58.
51] Appay V, Douek DC, Price DA. CD8+ T cell efficacy in vaccination and disease.Nat Med 2008;14(6 (June)):623–8.
52] Erard F, Dunbar PR, Le Gros G. The IL4- induced switch of CD8+ T cells to aTH2 phenotype and its possible relationship to the onset of AIDS. Res Immunol1994;145(8-9 (October–December)):643–5 [discussion 5–6].
53] Belyakov IM, Ahlers JD, Nabel GJ, Moss B, Berzofsky JA. Generation of function-ally active HIV-1 specific CD8+ CTL in intestinal mucosa following mucosal,systemic or mixed prime-boost immunization. Virology 2008;381(1 (Novem-ber 10)):106–15.
54] Guy B, Fourage S, Hessler C, Sanchez V, Millet MJ. Effects of the nature of adju-vant and site of parenteral immunization on the serum and mucosal immuneresponses induced by a nasal boost with a vaccine alone. Clin Diagn LabImmunol 1998;5(5 (September)):732–6.
55] Nakanishi Y, Lu B, Gerard C, Iwasaki A. CD8(+) T lymphocyte mobilizationto virus-infected tissue requires CD4(+) T-cell help. Nature 2009;462(7272(November 26)):510–3.
56] Kaufman DR, Liu J, Carville A, Mansfield KG, Havenga MJ, Goudsmit J, et al. Traf-ficking of antigen-specific CD8+ T lymphocytes to mucosal surfaces followingintramuscular vaccination. J Immunol 2008;181(6 (September 15)):4188–98.
57] Estcourt MJ, Letourneau S, McMichael AJ, Hanke T. Vaccine route, dose and typeof delivery vector determine patterns of primary CD8+ T cell responses. Eur JImmunol 2005;35(9 (September)):2532–40.
58] Abadie V, Bonduelle O, Duffy D, Parizot C, Verrier B, Combadiere B. Originalencounter with antigen determines antigen-presenting cell imprinting of thequality of the immune response in mice. PLoS One 2009;4(12):e8159.
59] Rappuoli R. Bridging the knowledge gaps in vaccine design. Nat Biotechnol2007;25(12 (December)):1361–6.
60] Belshe RB, Edwards KM, Vesikari T, Black SV, Walker RE, Hultquist M, et al. Liveattenuated versus inactivated influenza vaccine in infants and young children.N Engl J Med 2007;356(7 (February 15)):685–96.
Please cite this article in press as: Liard C, et al. Targeting of HIV-p24 particthe immune responses. Vaccine (2011), doi:10.1016/j.vaccine.2011.04.080
61] Pulendran B, Ahmed R. Translating innate immunity into immunologicalmemory: implications for vaccine development. Cell 2006;124(4 (February24)):849–63.
62] Poulin LF, Salio M, Griessinger E, Anjos-Afonso F, Craciun L, Chen JL, et al.Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equiv-
[
PRESSx (2011) xxx–xxx 13
alents of mouse CD8alpha+ dendritic cells. J Exp Med 2010;207(6 (June 7)):1261–71.
63] Villadangos JA, Shortman K. Found in translation: the human equivalent ofmouse CD8+ dendritic cells. J Exp Med 2010;207(6 (June 7)):1131–4.
64] Nasir A. Nanotechnology in vaccine development: a step forward. J Invest Der-matol 2009;129(5 (May)):1055–9.
65] Mottram PL, Leong D, Crimeen-Irwin B, Gloster S, Xiang SD, Meanger J, et al.Type 1 and 2 immunity following vaccination is influenced by nanoparticlesize: formulation of a model vaccine for respiratory syncytial virus. Mol Pharm2007;4(1 (January–February)):73–84.
66] Casares S, Inaba K, Brumeanu TD, Steinman RM, Bona CA. Antigen presentationby dendritic cells after immunization with DNA encoding a major histocom-patibility complex class II-restricted viral epitope. J Exp Med 1997;186(9(November 3)):1481–6.
67] Fay JW, Palucka AK, Paczesny S, Dhodapkar M, Johnston DA, Burkeholder S, et al.Long-term outcomes in patients with metastatic melanoma vaccinated withmelanoma peptide-pulsed CD34(+) progenitor-derived dendritic cells. CancerImmunol Immunother 2006;55(10 (October)):1209–18.
68] Ueno H, Schmitt N, Palucka AK, Banchereau J. Dendritic cells and humoralimmunity in humans. Immunol Cell Biol 2010;88(4 (May)):376–80.
69] Kissenpfennig A, Henri S, Dubois B, Laplace-Builhe C, Perrin P, Romani N, et al.Dynamics and function of Langerhans cells in vivo: dermal dendritic cellscolonize lymph node areas distinct from slower migrating Langerhans cells.Immunity 2005;22(5 (May)):643–54.
70] Belshe RB, Newman FK, Cannon J, Duane C, Treanor J, Van Hoecke C, et al. Serumantibody responses after intradermal vaccination against influenza. N Engl JMed 2004;351(22 (November 25)):2286–94.
71] Kunzi V, Dornseiff M, Horwath J, Hartmann K. Safe vaccination of childrenwith a virosomal adjuvanted influenza vaccine. Vaccine 2009;27(8 (February18)):1261–5.
72] Van Damme P, Oosterhuis-Kafeja F, Van der Wielen M, Almagor Y, Sharon O,Levin Y. Safety and efficacy of a novel microneedle device for dose sparing intra-
le-based vaccine into differential skin layers induces distinct arms of
dermal influenza vaccination in healthy adults. Vaccine 2009;27(3 (January14)):454–9.
73] Vien NC, Feroldi E, Lang J. Long-term anti-rabies antibody persistence follow-ing intramuscular or low-dose intradermal vaccination of young Vietnamesechildren. Trans R Soc Trop Med Hyg 2008;102(3 (March)):294–6.




















