
Target of rapamycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fat body of...
8
Target of rapamycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fat body of the tick, Haemaphysalis longicornis q Rika Umemiya-Shirafuji 1 , Damdinsuren Boldbaatar 1 , Min Liao 1 , Banzragch Battur, Md. Morshedur Rahman, Thasaneeya Kuboki, Remil Linggatong Galay, Tetsuya Tanaka, Kozo Fujisaki ⇑ Laboratory of Emerging Infectious Diseases, Department of Frontier Veterinary Medicine, Kagoshima University, 1-21-24 Korimoto, Kagoshima 890-0065, Japan article info Article history: Received 8 June 2012 Received in revised form 10 August 2012 Accepted 13 August 2012 Available online xxxx Keywords: Tick Fat body Vitellogenin Target of rapamycin S6K GATA abstract Vitellogenin (Vg) synthesis, vitellogenesis, is an essential process for the development and reproduction of ticks. Our previous finding led to the hypothesis that target of rapamycin (TOR) pathway is important for vitellogenesis in the hard tick, Haemaphysalis longicornis. The TOR pathway controls cellular activity according to nutrient availability in eukaryotes. TOR, a member of the phosphatidylinositol 3-kinase fam- ily, is a central player in this pathway. Here, we present preliminary evidence that H. longicornis TOR (HlTOR) controls vitellogenesis via activation of S6 kinase (S6K) in the fat body. RNA interference (RNAi)-mediated gene silencing of HlTOR was undertaken to elucidate the involvement of HlTOR in the vitellogenesis of the tick. HlTOR-RNAi caused inhibition of S6K phosphorylation in the fat body. HlTOR- RNAi also altered not only the expression levels of GATA mRNA and protein but also the intracellular localisation of GATA in the fat body. The expression levels of Vg mRNA and protein in the fat body of HlTOR-RNAi ticks were significantly lower than those in control ticks. In the pre-ovipositional stage, the ovaries of control ticks had brown oocytes developing, but those of HlTOR-RNAi ticks were white and immature. The haemolymph colour indicated that the amount of Vg was lower in HlTOR-RNAi ticks than in the controls. Furthermore, rapamycin inhibited S6K phosphorylation and reduced the expression levels of Vg mRNA and protein in the fat bodies. Vg proteins were not detected in rapamycin-treated fat bodies in the presence of 20-hydroxyecdysone. These results suggest that HlTOR activity is critical for vitellogenesis stimulated by 20-hydroxyecdysone. Ó 2012 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved. 1. Introduction Ticks are important haematophagous arthropods, acting as vec- tors of diverse agents of animal diseases as well as zoonoses. They need only the blood of vertebrate animals as a food source. Nutri- ents obtained during blood feeding are utilised to generate new proteins that are required for the crucial processes of tick develop- ment and reproduction. Host blood components, such as haemo- globin, are taken up by digestive cells of the midgut and then digested within the cells (Balashov, 1972; Tarnowski and Coons, 1989; Lara et al., 2005). Energy and nutrient reserves provided by digestion of a blood meal in female ticks allow the synthesis of vitellogenin (Vg), the yolk protein precursor (Grandjean, 1983). In ticks, Vg synthesis, or vitellogenesis, is commonly induced by a blood meal and is regulated by ecdysteroid hormones (James et al., 1997; Sankhon et al., 1999; Friesen and Kaufman, 2002; Thompson et al., 2005). Vg synthesised in the fat body is released into the haemolymph and then transported and incorporated into the eggs as the yolk protein, vitellin (Rosell-Davis and Coons, 1989; Rosell and Coons, 1992). Recently, Vg genes were identified from various species of ticks (Thompson et al., 2007; Granjeno-Colin et al., 2008; Horigane et al., 2010; Khalil et al., 2011). We also iden- tified multiple Vgs(HlVg-1, HlVg-2, and HlVg-3) from the hard tick, Haemaphysalis longicornis, and showed that the mRNA and protein expression of HlVg were induced by blood meals (Boldbaatar et al., 2010b). Furthermore, Boldbaatar et al. (2010a) showed that silenc- ing of S6 kinase (S6K) and GATA genes resulted in decreased tran- scription of Vg genes in female H. longicornis at engorgement. This study indicated that S6K is a key downstream regulator for GATA, which is an important transcription activator for the expres- sion of Vg in the fat body of the tick. These findings led to the hypothesis that the target of the rapamycin (TOR) pathway might be essential for vitellogenesis in the tick. However, the mechanism through which ecdysteroid hormones regulate vitellogenesis has 0020-7519/$36.00 Ó 2012 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.ijpara.2012.08.002 q Note: Nucleotide sequence data for Haemaphysalis longicornis target of rapamy- cin (HlTOR) has been submitted to the DDBJ/EMBL/GenBank databases under Accession No. AB716688. ⇑ Corresponding author. Present address: National Institute of Animal Health, National Agricultural and Food Research Organization, 3-1-5 Kannondai, Tsukuba, Ibaraki 305-0856, Japan. Tel.: +81 29 838 7713; fax: +81 29 838 7880. E-mail addresses: [email protected], [email protected] (K. Fujisaki). 1 These authors contributed equally to this work. International Journal for Parasitology xxx (2012) xxx–xxx Contents lists available at SciVerse ScienceDirect International Journal for Parasitology journal homepage: www.elsevier.com/locate/ijpara Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapamycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fat body of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.doi.org/10.1016/j.ijpara.2012.08.002
-
Upload
up-losbanos -
Category
Documents
-
view
0 -
download
0
Transcript of Target of rapamycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fat body of...

International Journal for Parasitology xxx (2012) xxx–xxx
Contents lists available at SciVerse ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
Target of rapamycin (TOR) controls vitellogenesis via activation of the S6 kinasein the fat body of the tick, Haemaphysalis longicornis q
Rika Umemiya-Shirafuji 1, Damdinsuren Boldbaatar 1, Min Liao 1, Banzragch Battur,Md. Morshedur Rahman, Thasaneeya Kuboki, Remil Linggatong Galay, Tetsuya Tanaka, Kozo Fujisaki ⇑Laboratory of Emerging Infectious Diseases, Department of Frontier Veterinary Medicine, Kagoshima University, 1-21-24 Korimoto, Kagoshima 890-0065, Japan
a r t i c l e i n f o a b s t r a c t
Article history:Received 8 June 2012Received in revised form 10 August 2012Accepted 13 August 2012Available online xxxx
Keywords:TickFat bodyVitellogeninTarget of rapamycinS6KGATA
0020-7519/$36.00 � 2012 Australian Society for Parahttp://dx.doi.org/10.1016/j.ijpara.2012.08.002
q Note: Nucleotide sequence data for Haemaphysaliscin (HlTOR) has been submitted to the DDBJ/EMBAccession No. AB716688.⇑ Corresponding author. Present address: Nationa
National Agricultural and Food Research OrganizationIbaraki 305-0856, Japan. Tel.: +81 29 838 7713; fax: +
E-mail addresses: [email protected], k8680181@ka1 These authors contributed equally to this work.
Please cite this article in press as: Umemiya-Shbody of the tick, Haemaphysalis longicornis. Int.
Vitellogenin (Vg) synthesis, vitellogenesis, is an essential process for the development and reproductionof ticks. Our previous finding led to the hypothesis that target of rapamycin (TOR) pathway is importantfor vitellogenesis in the hard tick, Haemaphysalis longicornis. The TOR pathway controls cellular activityaccording to nutrient availability in eukaryotes. TOR, a member of the phosphatidylinositol 3-kinase fam-ily, is a central player in this pathway. Here, we present preliminary evidence that H. longicornis TOR(HlTOR) controls vitellogenesis via activation of S6 kinase (S6K) in the fat body. RNA interference(RNAi)-mediated gene silencing of HlTOR was undertaken to elucidate the involvement of HlTOR in thevitellogenesis of the tick. HlTOR-RNAi caused inhibition of S6K phosphorylation in the fat body. HlTOR-RNAi also altered not only the expression levels of GATA mRNA and protein but also the intracellularlocalisation of GATA in the fat body. The expression levels of Vg mRNA and protein in the fat body ofHlTOR-RNAi ticks were significantly lower than those in control ticks. In the pre-ovipositional stage,the ovaries of control ticks had brown oocytes developing, but those of HlTOR-RNAi ticks were whiteand immature. The haemolymph colour indicated that the amount of Vg was lower in HlTOR-RNAi ticksthan in the controls. Furthermore, rapamycin inhibited S6K phosphorylation and reduced the expressionlevels of Vg mRNA and protein in the fat bodies. Vg proteins were not detected in rapamycin-treated fatbodies in the presence of 20-hydroxyecdysone. These results suggest that HlTOR activity is critical forvitellogenesis stimulated by 20-hydroxyecdysone.
� 2012 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Ticks are important haematophagous arthropods, acting as vec-tors of diverse agents of animal diseases as well as zoonoses. Theyneed only the blood of vertebrate animals as a food source. Nutri-ents obtained during blood feeding are utilised to generate newproteins that are required for the crucial processes of tick develop-ment and reproduction. Host blood components, such as haemo-globin, are taken up by digestive cells of the midgut and thendigested within the cells (Balashov, 1972; Tarnowski and Coons,1989; Lara et al., 2005). Energy and nutrient reserves providedby digestion of a blood meal in female ticks allow the synthesisof vitellogenin (Vg), the yolk protein precursor (Grandjean, 1983).
sitology Inc. Published by Elsevier
longicornis target of rapamy-L/GenBank databases under
l Institute of Animal Health,, 3-1-5 Kannondai, Tsukuba,81 29 838 7880.dai.jp (K. Fujisaki).
irafuji, R., et al. Target of rapamJ. Parasitol. (2012), http://dx.do
In ticks, Vg synthesis, or vitellogenesis, is commonly induced bya blood meal and is regulated by ecdysteroid hormones (Jameset al., 1997; Sankhon et al., 1999; Friesen and Kaufman, 2002;Thompson et al., 2005). Vg synthesised in the fat body is releasedinto the haemolymph and then transported and incorporated intothe eggs as the yolk protein, vitellin (Rosell-Davis and Coons, 1989;Rosell and Coons, 1992). Recently, Vg genes were identified fromvarious species of ticks (Thompson et al., 2007; Granjeno-Colinet al., 2008; Horigane et al., 2010; Khalil et al., 2011). We also iden-tified multiple Vgs (HlVg-1, HlVg-2, and HlVg-3) from the hard tick,Haemaphysalis longicornis, and showed that the mRNA and proteinexpression of HlVg were induced by blood meals (Boldbaatar et al.,2010b). Furthermore, Boldbaatar et al. (2010a) showed that silenc-ing of S6 kinase (S6K) and GATA genes resulted in decreased tran-scription of Vg genes in female H. longicornis at engorgement.This study indicated that S6K is a key downstream regulator forGATA, which is an important transcription activator for the expres-sion of Vg in the fat body of the tick. These findings led to thehypothesis that the target of the rapamycin (TOR) pathway mightbe essential for vitellogenesis in the tick. However, the mechanismthrough which ecdysteroid hormones regulate vitellogenesis has
Ltd. All rights reserved.
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002

2 R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx
not been characterised and it also remains unclear whether theTOR is implicated in the regulation of vitellogenesis in ticks.
TOR, a member of the phosphatidylinositol 3-kinase (PI3-K)family, is a central player in the conserved insulin/TOR pathwayand controls cellular growth by regulating several processes suchas transcription, translation, protein degradation and ribosomalbiogenesis, according to nutrient availability. TOR targets S6Kand the eukaryotic initiation factor 4E-binding protein (4EBP) tocontrol protein translation; it also regulates the localisation oftranscription factors, autophagy and intracellular amino acid levels(reviewed in Inoki et al., 2005).
In insects, some studies indicated that TOR signalling is a crucialpathway in regulating Vg gene expression. Nutritional signals acti-vate juvenile hormone (JH) biosynthesis, which is mediated by theTOR pathway in the cockroach Blattella germanica (Maestro et al.,2009). Vg gene transcription is induced by JH in the fat body of in-sects (Comas et al., 2001; Tufail et al., 2010). In other insects, it wassuggested that JH alone is not sufficient to upregulate the expres-sion of Vg genes and that JH acts as an intermediate between nutri-tion and Vg production (Fei et al., 2005; Panaitof and Scott, 2006).In Tribolium castaneum, JH is required for Vg synthesis in the fatbody, and 20-hydroxyecdysone (20E) influences Vg synthesisthrough its action on oocyte maturation (Parthasarathy et al.,2010a, 2010b). Amino acid/TOR/S6K signalling is essential for theactivation of Vg gene expression and egg development, and theexpression is regulated by the GATA factor in the anautogeny mos-quito, Aedes aegypti (Kokoza et al., 2001; Hansen et al., 2004, 2005;Park et al., 2006). In addition, evidence indicates that the smallGTPase (guanosine triphosphatase), Rheb (Ras homologue en-riched in brain), an upstream activator of TOR, is required for ami-no acid-mediated activation of the TOR pathway in the fat body ofthe same mosquito (Roy and Raikhel, 2011). These studies indi-cated that TOR-mediated amino acid signalling is an importantpathway in the reproductive cycle of the mosquito. Suganyaet al. (2010, 2011) also showed the involvement of the TOR/S6Kpathway in the regulation of yolk protein synthesis in the orientalfruit fly, Bactrocera dorsalis. Ecdysone and nutrients promote theproduction of yolk proteins in Drosophila (Bownes et al., 1988; Bus-zczak et al., 1999); however, it is unclear whether TOR regulatesvitellogenesis in the fly.
In the present study, to test the hypothesis that the TOR path-way might be essential for vitellogenesis in the tick, we isolateda homologue of TOR from H. longicornis (HlTOR), which is a proto-zoan/rickettsial vector for animals and humans in Asia and Austra-lia (Hoogstraal et al., 1968), and performed RNA interference(RNAi)-mediated gene silencing. In addition, an in vitro organ cul-ture of the fat body was used to examine whether TOR activity con-tributes to vitellogenesis in the tick.
2. Materials and methods
2.1. Ticks and animals
Adult female ticks of the parthenogenetic H. longicornis (Oka-yama strain) were used. Ticks have been maintained by feedingon the ears of Japanese white rabbits (Kyudo, Kumamoto, Japan)using cotton bags (Fujisaki, 1978) at the Laboratory of EmergingInfectious Diseases, Department of Frontier Veterinary Medicine,in Kagoshima University. Rabbits were cared for in accordancewith the guidelines approved by the Animal Care and Use Commit-tee of Kagoshima University (Approval No. A08010). They weremaintained in a temperature (23 �C)- and humidity (50–60%)-reg-ulated room under controlled lighting, with free access to tapwater and commercial regular chow throughout the experiments.
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
2.2. Cloning and sequencing of HlTOR cDNA
Total RNA was extracted from the ovaries of female ticks2 weeks after engorgement using TRI� reagent (Sigma, MO, USA),as described previously (Umemiya-Shirafuji et al., 2010). Single-stranded cDNA was generated by reverse transcription using theTranscriptor First Strand cDNA Synthesis Kit (Roche Diagnostics,Mannheim, Germany), and the cDNA was used as a template forPCRs. Based on the conserved regions of known TOR of insectsand mammals, degenerate primers were designed for PCR. ThePCR products were subjected to 1.0% agarose gel electrophoresis;the band of predictive size was purified using a QIAquick GelExtraction Kit (QIAGEN, Hilden, Germany) and cloned into thepGEM�-T vector (Promega, WI, USA). Sequence analyses were con-ducted on an Applied Biosystems 3130xl Genetic Analyzer (AppliedBiosystems, CA, USA). GENETYX� software version 7 (Genetyx, To-kyo, Japan) and BLAST analyses (National Center for BiotechnologyInformation (NCBI), National Institute of Health; http://www.ncbi.nlm.nih.gov/BLAST/) were used to confirm that the par-tial cDNA encodes TOR. The missing 50 and 30 ends were then ob-tained by 50 and 30-rapid amplification of cDNA ends (RACE)using the SMARTer RACE cDNA Amplification Kit (Takara Bio, Shi-ga, Japan) and different specific primers to complete the sequence.BLAST analysis of the translation products deduced from the full-length open reading frames (ORFs) was performed. The deducedamino acid sequences were aligned with TOR of various organismsusing a ClustalW2 program (http://www.ebi.ac.uk/Tools/clu-stalw2/index.html). The domain structure was determined usingthe Simple Modular Architecture Research Tool (SMART; http://smart.embl-heidelberg.de/).
2.3. RNA interference
A 390 bp cDNA fragment was subjected to generate double-stranded RNA (dsRNA). To minimise off-target effects, the specific-ity of the dsRNA sequence was confirmed with in silico analysis(Supplementary Tables S1 and S2). Sequences of a 390 bp fragmentand the predicted short 372 fragments did not show homologywith available sequences of H. longicornis (46,534 expressed se-quence tags (ESTs)) or other of tick species. This 390 bp fragmentcontained a C-terminal sequence encoding the partial kinase do-main and the entire FATC domains. After PCR using T7 forward(50-GTAATACGACTCACTATAGGGGAGGCGTTTGTGTACGACC-30)and reverse (50-GTAATACGACTCACTATAGGGTCACCAAAACGGG-CACCAT-30) primers to attach to the T7 promoter recognition siteson both the 50 and 30 ends (underlined), the PCR products werepurified as described in Section 2.2. The dsRNA of HlTOR was syn-thesised using the T7 RiboMAX™ Express RNAi System (Promega).Approximately 0.5 ll of HlTOR dsRNA (6.8 lM) was injected intounfed female ticks as described previously (Miyoshi et al., 2004).Control ticks were injected with 0.5 ll of luciferase dsRNA, whichwas prepared using the primers described by Hatta et al. (2010).The injected ticks were allowed to rest for 1 day at 25 �C and thenplaced on the ears of rabbits to feed. Ticks were collected from therabbits during blood feeding and at engorgement. Engorged tickswere transferred to a moist chamber at 25 �C for oviposition. Thisexperiment was replicated three times.
2.4. Reverse transcription and real-time PCR
Total RNA was extracted from each organ (the fat body, midgut,ovary and salivary glands) of normal female ticks in an unfed state,after 2 and 4 days of feeding, at engorgement, and 5 and 13 daysafter engorgement to examine the expression of HlTOR mRNA.dsRNA-injected ticks and their fat bodies were also subjected tototal RNA extraction. Single-stranded cDNA was generated as
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002

R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx 3
described above. PCR was carried out using a 7300 Real-Time PCRSystem and Power SYBR� Green PCR Master Mix (AppliedBiosystems) as described previously (Umemiya-Shirafuji et al.,2010). Primer sets used are shown in Supplementary Table S3. Datawere collected with the use of the 7300 system SDS software ver-sion 1.4 (Applied Biosystems) and analysed using Microsoft Excel.The cDNA levels were normalised to the H. longicornis ribosomalprotein P0 gene (Gong et al., 2008).
2.5. Immunoblotting
The fat bodies of dsRNA-injected ticks were suspended in Tris-buffered saline containing 1% Triton-X100, a protease inhibitorcocktail (Complete Mini; Roche Diagnostics) and a phosphataseinhibitor cocktail (PhosSTOP; Roche Diagnostics). After overnightincubation at 4 �C, the extracts were centrifuged at 22,140g for30 min and the supernatants were stored at �30 �C until used.The proteins were separated by SDS–PAGE under reducing condi-tions on a 3–10% acrylamide gel (Atto, Tokyo, Japan). Immunoblot-ting was performed using the phospho-p70 S6 kinase (Thr389)antibody (Cell Signaling Technology, MA, USA), mouse polyclonalanti-HlGATA, anti-HlVg-2, and anti-HlVg-3 sera (Boldbaatar et al.,2010a, 2010b). Anti-HlACTIN serum was used as a loading control(Liao et al., 2008). The binding of antibodies was detected withhorseradish peroxidase-conjugated polyclonal goat anti-mouseimmunoglobulin (Dako, Glostrup, Denmark) and an ECL Plus Wes-tern Blotting Detection Kit (GE Healthcare, Chalfont St. Giles, UK).The gel images were analysed using a FluorChem� FC2 Imagingsystem (Cell Biosciences, CA, USA) and then processed using Micro-soft Power Point.
2.6. Immunofluorescent staining
The fat bodies of dsRNA-injected ticks were processed to makefrozen sections (approximately 10 lm thick) on a cryostat (LeicaCM 3050; Leica Microsystems, Wetzlar, Germany) as describedpreviously (Umemiya et al., 2007). The sections were placed onMAS-coated glass slides (Matsunami Glass, Osaka, Japan). Afterblocking with 5% skim milk in PBS, those were incubated with a1:100 dilution of anti-HlGATA serum for 2 h at room temperature.Normal mouse serum (1:100) was used as a negative control. Sec-tions were washed with PBS and then reacted with Alexa Fluor�
594 goat anti-mouse IgG (1:1000; Life Technologies, MD, USA)for 1 h at room temperature. After washing with PBS, they weremounted in the ProLong� Gold antifade reagent with DAPI (LifeTechnologies). Images were collected using a microscope OlympusIX71 equipped with a DP71 camera and DP Controller software,and then processed using DP Manager software (Olympus, Tokyo,Japan).
2.7. In vitro organ culture of the fat body
Female ticks were placed on the ears of rabbits to feed and col-lected at engorgement. Engorged ticks were transferred to a moistchamber at 25 �C. They were weighed and grouped before dissec-tion. There were no differences in the average body weights be-tween groups. Three females per group were dissected 6 daysafter engorgement in order to utilise their fat bodies for incubationwith rapamycin and/or 20E. The fat bodies were collected fromaround the tracheal trunks and around the ovary. The fat bodieswere transferred to each microtiter plate well containing 1 ml ofLeibovitz’s L-15 medium with L-glutamine (Life technologies, CA,USA) and incubated in the medium at 28 �C for 1 h. Subsequently,they were incubated for 1 h in a medium containing 1 lM rapamy-cin (Merck KGaA, Darmstadt, Germany) or the same volume of di-methyl sulfoxide (the rapamycin solvent) and then for 2 h in a
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
medium containing 1 lM 20E (Sigma) or the same volume of70% ethanol (the 20E solvent). After incubation, the fat bodies werecollected to extract total RNA or protein and then subjected to real-time PCR and immunoblotting as described in Sections 2.4 and 2.5.
2.8. Nucleotide sequence accession number
Nucleotide sequence data for HlTOR has been submitted to theDDBJ/EMBL/GenBank databases under Accession No. AB716688.The accession numbers of the H. longicornis genes used in thisstudy are AB491580 for HlGATA, AB359901 for HlVg-2, AB359902for HlVg-3, and EU048401 for HlP0.
2.9. Statistical analysis
Statistical analysis was performed using Student’s t-tests orMann–Whitney U tests, depending on the data distribution. Statis-tical significance was defined by P < 0.01.
3. Results
3.1. Sequence feature of HlTOR
A sequence of 7,848 bp was amplified from cDNA of ovaries offemale ticks 2 weeks after engorgement using degenerate primers.The gene contained a 7,572 bp open reading frame encoding a2,523-amino acid polypeptide with a predicted molecular massof 287 kDa. The amino acid sequence comparison by BLAST analy-sis revealed that the sequence encodes the predicted TOR protein,which has conserved domains: FAT (FKBP12-rapamycin-associatedprotein (FRAP), ataxia telangiectasia mutated (ATM), transforma-tion/transcription domain-associated protein (TRRAP)), FBP (rapa-mycin/FK506-binding protein (FKBP12)-binding domain), Kinase(phosphoinositide 3-kinase catalytic domain), and FATC (FAT car-boxyl terminal domain) (Supplementary Fig. S1). The sequence,named HlTOR in the present study, showed identities of more than55% with the TOR of the German cockroach, B. germanica, theFKBP12-rapamycin complex-associated protein of the honey bee,Apis mellifera, the TOR of the yellow fever mosquito, A. aegypti,the TOR isoform 2 of the silkworm, Bombyx mori, and the TOR ofthe fruit fly, Drosophila melanogaster. The kinase domain is highlyconserved among these species (Supplementary Table S4).
3.2. Expression profile of HlTOR mRNA in female ticks
HlTOR mRNA was expressed in the fat body, midgut, ovary andsalivary glands of female ticks (Fig. 1). Real-time PCR analysis re-vealed that the HlTOR mRNA level was highest at the slow feedingphase (2 days of feeding) and decreased after the rapid feedingphase (4 days of feeding) in the fat body and midgut. Higherexpression of HlTOR was found in the ovary and the level strikinglyincreased after engorgement. The HlTOR mRNA level also increasedin the salivary glands during blood feeding and the level peakedupon engorgement.
3.3. Knockdown of HlTOR in female ticks
It was suggested that the TOR pathway might be important forvitellogenesis in ticks (Boldbaatar et al., 2010a). Based on this pre-mise, to explore the function of HlTOR, RNAi-mediated gene silenc-ing by injection of dsRNA into female ticks was conducted. HlTORmRNA of control ticks (luciferase dsRNA-injected ticks) was abun-dant at the slow phase of feeding (1–2 days) (SupplementaryFig. S2A). HlTOR mRNA decreased after 3 days of feeding and thenincreased again during the rapid feeding phase (4–5 days of
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002
A
B
HlGATA
mR
NA
0123456
4d engorged
ControlHlTOR RNAi
369
121518
HlVg-2
mR
NA
ControlHlTOR RNAi
*
*
**
4 R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx
feeding). However, the HlTOR mRNA levels of RNAi-treated tickswere constantly lower than those of control ticks (P < 0.01). RNAiresulted in 87.0% reduction of HlTOR gene expression inHlTOR-RNAi ticks at 5 days of feeding compared with control ticks.
The fat body is the primary site of Vg production in ticks (Coonset al., 1982; Chinzei and Yano, 1985; James et al., 1999). During thereproductive phase in female ticks, it plays a vital role in reproduc-tion, synthesising and secreting vitellogenin, especially HlVg-2 andHlVg-3 (Boldbaatar et al., 2010b). Consequently, we analysed theefficiency of HlTOR-RNAi in the fat body of HlTOR dsRNA-injectedticks. RNAi resulted in 87.0% and 79.1% reduction of HlTOR geneexpression in the fat body of HlTOR-RNAi ticks at 4 days of feedingand at engorgement, respectively, when compared with controlticks (Supplementary Fig. S2B; P < 0.01). Knockdown of HlTORwas also noted in other organs, such as the midgut and ovary (datanot shown).
3.4. Impacts of HlTOR RNAi on vitellogenesis
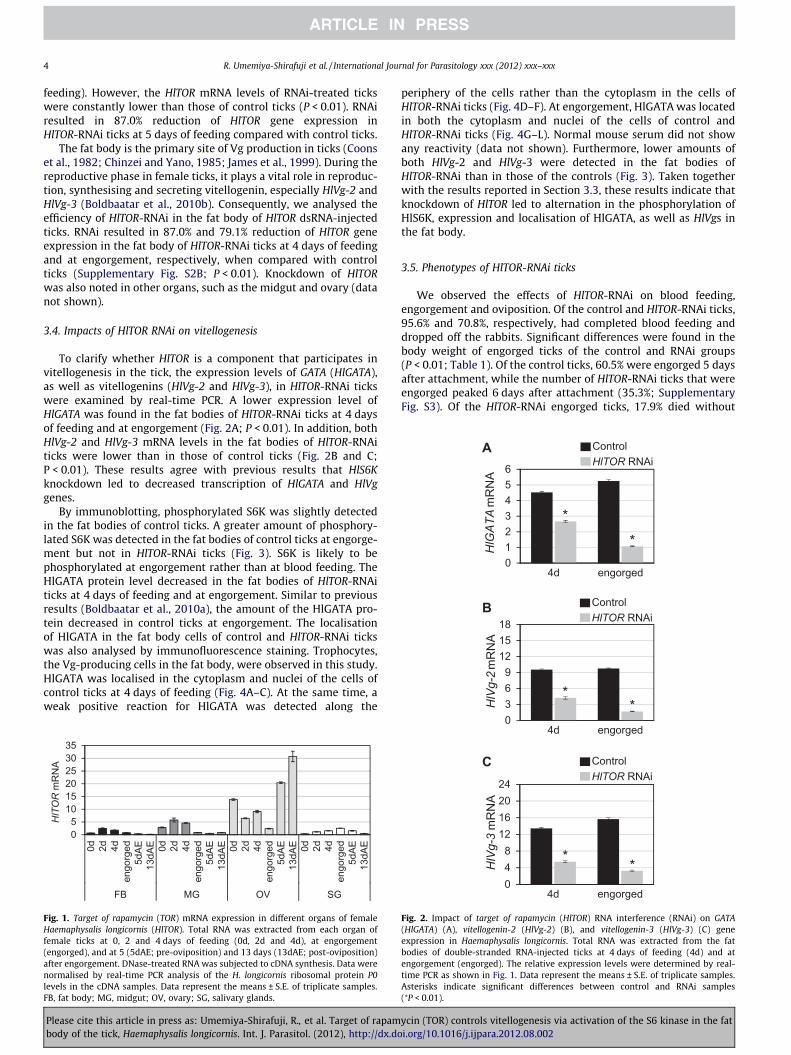
To clarify whether HlTOR is a component that participates invitellogenesis in the tick, the expression levels of GATA (HlGATA),as well as vitellogenins (HlVg-2 and HlVg-3), in HlTOR-RNAi tickswere examined by real-time PCR. A lower expression level ofHlGATA was found in the fat bodies of HlTOR-RNAi ticks at 4 daysof feeding and at engorgement (Fig. 2A; P < 0.01). In addition, bothHlVg-2 and HlVg-3 mRNA levels in the fat bodies of HlTOR-RNAiticks were lower than in those of control ticks (Fig. 2B and C;P < 0.01). These results agree with previous results that HlS6Kknockdown led to decreased transcription of HlGATA and HlVggenes.
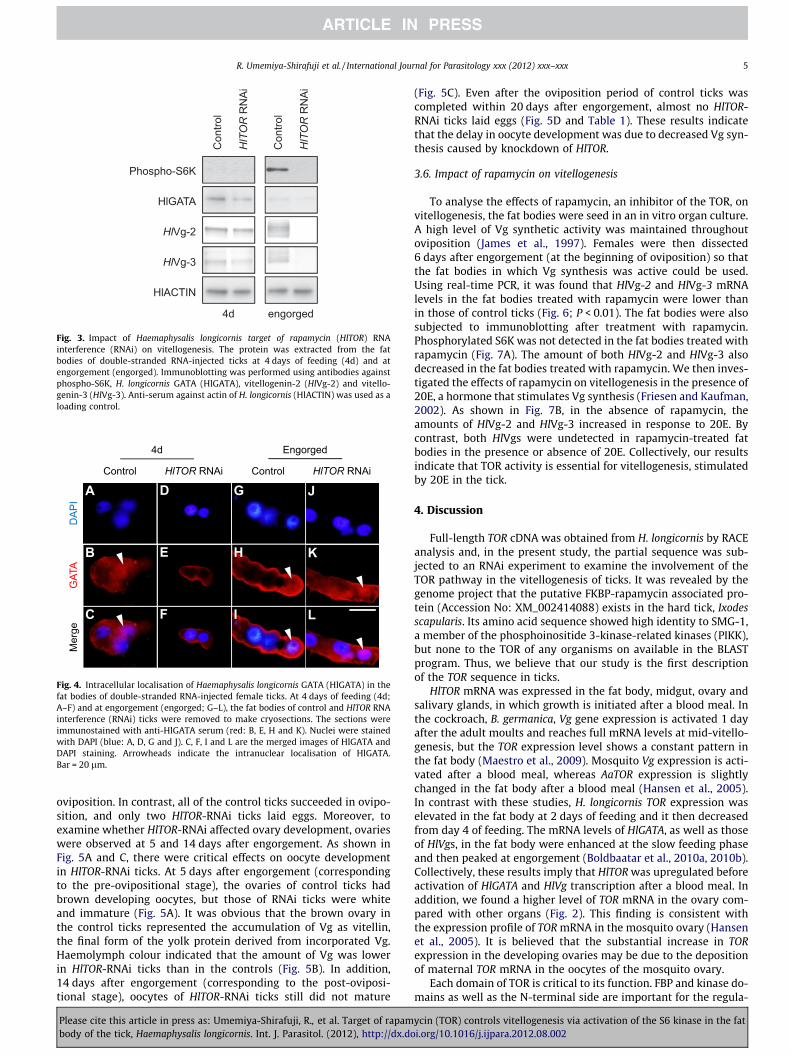
By immunoblotting, phosphorylated S6K was slightly detectedin the fat bodies of control ticks. A greater amount of phosphory-lated S6K was detected in the fat bodies of control ticks at engorge-ment but not in HlTOR-RNAi ticks (Fig. 3). S6K is likely to bephosphorylated at engorgement rather than at blood feeding. TheHlGATA protein level decreased in the fat bodies of HlTOR-RNAiticks at 4 days of feeding and at engorgement. Similar to previousresults (Boldbaatar et al., 2010a), the amount of the HlGATA pro-tein decreased in control ticks at engorgement. The localisationof HlGATA in the fat body cells of control and HlTOR-RNAi tickswas also analysed by immunofluorescence staining. Trophocytes,the Vg-producing cells in the fat body, were observed in this study.HlGATA was localised in the cytoplasm and nuclei of the cells ofcontrol ticks at 4 days of feeding (Fig. 4A–C). At the same time, aweak positive reaction for HlGATA was detected along the
HlTOR
mR
NA
05
101520253035
0d 2d 4den
gorg
ed5d
AE13
dAE 0d 2d 4d
engo
rged
5dAE
13dA
E 0d 2d 4den
gorg
ed5d
AE13
dAE 0d 2d 4d
engo
rged
5dAE
13dA
E
FB MG OV SG
Fig. 1. Target of rapamycin (TOR) mRNA expression in different organs of femaleHaemaphysalis longicornis (HlTOR). Total RNA was extracted from each organ offemale ticks at 0, 2 and 4 days of feeding (0d, 2d and 4d), at engorgement(engorged), and at 5 (5dAE; pre-oviposition) and 13 days (13dAE; post-oviposition)after engorgement. DNase-treated RNA was subjected to cDNA synthesis. Data werenormalised by real-time PCR analysis of the H. longicornis ribosomal protein P0levels in the cDNA samples. Data represent the means ± S.E. of triplicate samples.FB, fat body; MG, midgut; OV, ovary; SG, salivary glands.
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
periphery of the cells rather than the cytoplasm in the cells ofHlTOR-RNAi ticks (Fig. 4D–F). At engorgement, HlGATA was locatedin both the cytoplasm and nuclei of the cells of control andHlTOR-RNAi ticks (Fig. 4G–L). Normal mouse serum did not showany reactivity (data not shown). Furthermore, lower amounts ofboth HlVg-2 and HlVg-3 were detected in the fat bodies ofHlTOR-RNAi than in those of the controls (Fig. 3). Taken togetherwith the results reported in Section 3.3, these results indicate thatknockdown of HlTOR led to alternation in the phosphorylation ofHlS6K, expression and localisation of HlGATA, as well as HlVgs inthe fat body.
3.5. Phenotypes of HlTOR-RNAi ticks
We observed the effects of HlTOR-RNAi on blood feeding,engorgement and oviposition. Of the control and HlTOR-RNAi ticks,95.6% and 70.8%, respectively, had completed blood feeding anddropped off the rabbits. Significant differences were found in thebody weight of engorged ticks of the control and RNAi groups(P < 0.01; Table 1). Of the control ticks, 60.5% were engorged 5 daysafter attachment, while the number of HlTOR-RNAi ticks that wereengorged peaked 6 days after attachment (35.3%; SupplementaryFig. S3). Of the HlTOR-RNAi engorged ticks, 17.9% died without
C
04d engorged
048
12162024
4d engorged
HlVg-3
mR
NA
ControlHlTOR RNAi
* *
Fig. 2. Impact of target of rapamycin (HlTOR) RNA interference (RNAi) on GATA(HlGATA) (A), vitellogenin-2 (HlVg-2) (B), and vitellogenin-3 (HlVg-3) (C) geneexpression in Haemaphysalis longicornis. Total RNA was extracted from the fatbodies of double-stranded RNA-injected ticks at 4 days of feeding (4d) and atengorgement (engorged). The relative expression levels were determined by real-time PCR as shown in Fig. 1. Data represent the means ± S.E. of triplicate samples.Asterisks indicate significant differences between control and RNAi samples(⁄P < 0.01).
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002
4d engorged
Phospho-S6K
HlGATA
HlVg-2
HlVg-3
HlACTIN
Con
trol
HlTOR
RN
Ai
Con
trol
HlTOR
RN
Ai
Fig. 3. Impact of Haemaphysalis longicornis target of rapamycin (HlTOR) RNAinterference (RNAi) on vitellogenesis. The protein was extracted from the fatbodies of double-stranded RNA-injected ticks at 4 days of feeding (4d) and atengorgement (engorged). Immunoblotting was performed using antibodies againstphospho-S6K, H. longicornis GATA (HlGATA), vitellogenin-2 (HlVg-2) and vitello-genin-3 (HlVg-3). Anti-serum against actin of H. longicornis (HlACTIN) was used as aloading control.
4d Engorged
DAP
IG
ATA
Mer
ge
Control HlTOR RNAiControl HlTOR RNAi
A
B
C
D
E
F
G
H
I
J
K
L
Fig. 4. Intracellular localisation of Haemaphysalis longicornis GATA (HlGATA) in thefat bodies of double-stranded RNA-injected female ticks. At 4 days of feeding (4d;A–F) and at engorgement (engorged; G–L), the fat bodies of control and HlTOR RNAinterference (RNAi) ticks were removed to make cryosections. The sections wereimmunostained with anti-HlGATA serum (red: B, E, H and K). Nuclei were stainedwith DAPI (blue: A, D, G and J). C, F, I and L are the merged images of HlGATA andDAPI staining. Arrowheads indicate the intranuclear localisation of HlGATA.Bar = 20 lm.
R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx 5
oviposition. In contrast, all of the control ticks succeeded in ovipo-sition, and only two HlTOR-RNAi ticks laid eggs. Moreover, toexamine whether HlTOR-RNAi affected ovary development, ovarieswere observed at 5 and 14 days after engorgement. As shown inFig. 5A and C, there were critical effects on oocyte developmentin HlTOR-RNAi ticks. At 5 days after engorgement (correspondingto the pre-ovipositional stage), the ovaries of control ticks hadbrown developing oocytes, but those of RNAi ticks were whiteand immature (Fig. 5A). It was obvious that the brown ovary inthe control ticks represented the accumulation of Vg as vitellin,the final form of the yolk protein derived from incorporated Vg.Haemolymph colour indicated that the amount of Vg was lowerin HlTOR-RNAi ticks than in the controls (Fig. 5B). In addition,14 days after engorgement (corresponding to the post-oviposi-tional stage), oocytes of HlTOR-RNAi ticks still did not mature
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
(Fig. 5C). Even after the oviposition period of control ticks wascompleted within 20 days after engorgement, almost no HlTOR-RNAi ticks laid eggs (Fig. 5D and Table 1). These results indicatethat the delay in oocyte development was due to decreased Vg syn-thesis caused by knockdown of HlTOR.
3.6. Impact of rapamycin on vitellogenesis
To analyse the effects of rapamycin, an inhibitor of the TOR, onvitellogenesis, the fat bodies were seed in an in vitro organ culture.A high level of Vg synthetic activity was maintained throughoutoviposition (James et al., 1997). Females were then dissected6 days after engorgement (at the beginning of oviposition) so thatthe fat bodies in which Vg synthesis was active could be used.Using real-time PCR, it was found that HlVg-2 and HlVg-3 mRNAlevels in the fat bodies treated with rapamycin were lower thanin those of control ticks (Fig. 6; P < 0.01). The fat bodies were alsosubjected to immunoblotting after treatment with rapamycin.Phosphorylated S6K was not detected in the fat bodies treated withrapamycin (Fig. 7A). The amount of both HlVg-2 and HlVg-3 alsodecreased in the fat bodies treated with rapamycin. We then inves-tigated the effects of rapamycin on vitellogenesis in the presence of20E, a hormone that stimulates Vg synthesis (Friesen and Kaufman,2002). As shown in Fig. 7B, in the absence of rapamycin, theamounts of HlVg-2 and HlVg-3 increased in response to 20E. Bycontrast, both HlVgs were undetected in rapamycin-treated fatbodies in the presence or absence of 20E. Collectively, our resultsindicate that TOR activity is essential for vitellogenesis, stimulatedby 20E in the tick.
4. Discussion
Full-length TOR cDNA was obtained from H. longicornis by RACEanalysis and, in the present study, the partial sequence was sub-jected to an RNAi experiment to examine the involvement of theTOR pathway in the vitellogenesis of ticks. It was revealed by thegenome project that the putative FKBP-rapamycin associated pro-tein (Accession No: XM_002414088) exists in the hard tick, Ixodesscapularis. Its amino acid sequence showed high identity to SMG-1,a member of the phosphoinositide 3-kinase-related kinases (PIKK),but none to the TOR of any organisms on available in the BLASTprogram. Thus, we believe that our study is the first descriptionof the TOR sequence in ticks.
HlTOR mRNA was expressed in the fat body, midgut, ovary andsalivary glands, in which growth is initiated after a blood meal. Inthe cockroach, B. germanica, Vg gene expression is activated 1 dayafter the adult moults and reaches full mRNA levels at mid-vitello-genesis, but the TOR expression level shows a constant pattern inthe fat body (Maestro et al., 2009). Mosquito Vg expression is acti-vated after a blood meal, whereas AaTOR expression is slightlychanged in the fat body after a blood meal (Hansen et al., 2005).In contrast with these studies, H. longicornis TOR expression waselevated in the fat body at 2 days of feeding and it then decreasedfrom day 4 of feeding. The mRNA levels of HlGATA, as well as thoseof HlVgs, in the fat body were enhanced at the slow feeding phaseand then peaked at engorgement (Boldbaatar et al., 2010a, 2010b).Collectively, these results imply that HlTOR was upregulated beforeactivation of HlGATA and HlVg transcription after a blood meal. Inaddition, we found a higher level of TOR mRNA in the ovary com-pared with other organs (Fig. 2). This finding is consistent withthe expression profile of TOR mRNA in the mosquito ovary (Hansenet al., 2005). It is believed that the substantial increase in TORexpression in the developing ovaries may be due to the depositionof maternal TOR mRNA in the oocytes of the mosquito ovary.
Each domain of TOR is critical to its function. FBP and kinase do-mains as well as the N-terminal side are important for the regula-
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002
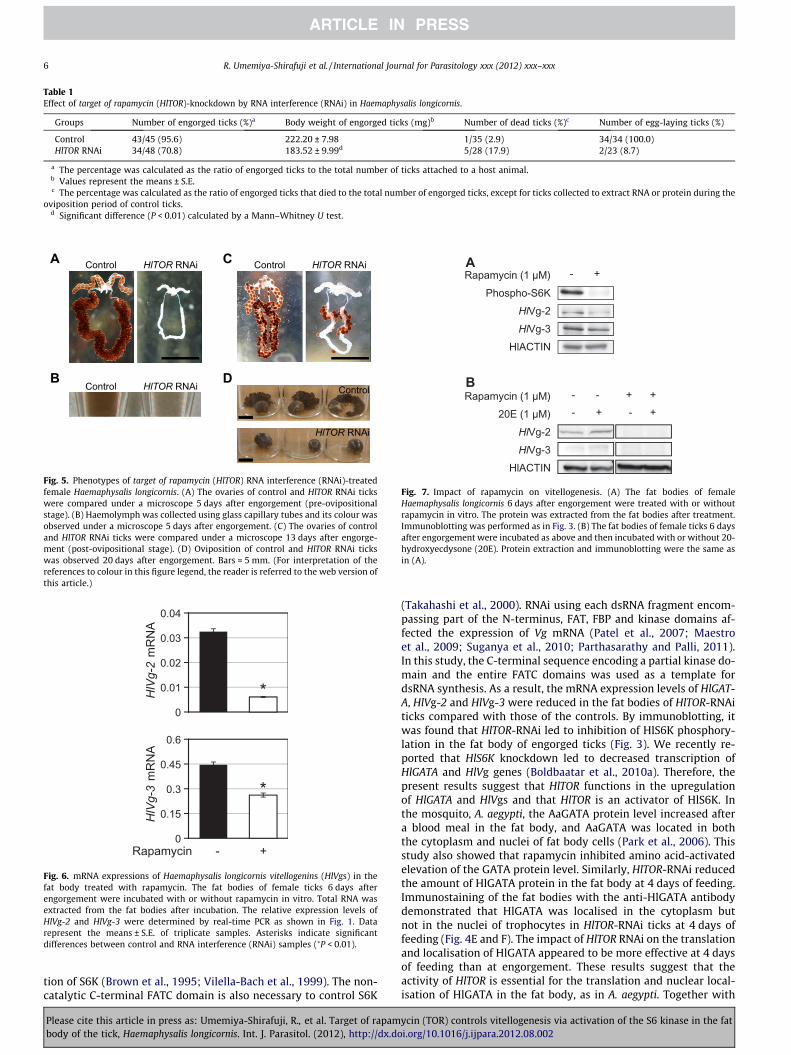
Table 1Effect of target of rapamycin (HlTOR)-knockdown by RNA interference (RNAi) in Haemaphysalis longicornis.
Groups Number of engorged ticks (%)a Body weight of engorged ticks (mg)b Number of dead ticks (%)c Number of egg-laying ticks (%)
Control 43/45 (95.6) 222.20 ± 7.98 1/35 (2.9) 34/34 (100.0)HlTOR RNAi 34/48 (70.8) 183.52 ± 9.99d 5/28 (17.9) 2/23 (8.7)
a The percentage was calculated as the ratio of engorged ticks to the total number of ticks attached to a host animal.b Values represent the means ± S.E.c The percentage was calculated as the ratio of engorged ticks that died to the total number of engorged ticks, except for ticks collected to extract RNA or protein during the
oviposition period of control ticks.d Significant difference (P < 0.01) calculated by a Mann–Whitney U test.
A
B
Control HlTOR RNAi Control HlTOR RNAi
Control
HlTOR RNAi
Control HlTOR RNAi
C
D
Fig. 5. Phenotypes of target of rapamycin (HlTOR) RNA interference (RNAi)-treatedfemale Haemaphysalis longicornis. (A) The ovaries of control and HlTOR RNAi tickswere compared under a microscope 5 days after engorgement (pre-ovipositionalstage). (B) Haemolymph was collected using glass capillary tubes and its colour wasobserved under a microscope 5 days after engorgement. (C) The ovaries of controland HlTOR RNAi ticks were compared under a microscope 13 days after engorge-ment (post-ovipositional stage). (D) Oviposition of control and HlTOR RNAi tickswas observed 20 days after engorgement. Bars = 5 mm. (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version ofthis article.)
0
0.15
0.3
0.45
0.6
0
0.01
0.02
0.03
0.04
Rapamycin - +
HlVg-2
mR
NA
HlVg-3
mR
NA
*
*
Fig. 6. mRNA expressions of Haemaphysalis longicornis vitellogenins (HlVgs) in thefat body treated with rapamycin. The fat bodies of female ticks 6 days afterengorgement were incubated with or without rapamycin in vitro. Total RNA wasextracted from the fat bodies after incubation. The relative expression levels ofHlVg-2 and HlVg-3 were determined by real-time PCR as shown in Fig. 1. Datarepresent the means ± S.E. of triplicate samples. Asterisks indicate significantdifferences between control and RNA interference (RNAi) samples (⁄P < 0.01).
Rapamycin (1 µM)Phospho-S6K
HlVg-2HlVg-3
HlACTIN
- +
Rapamycin (1 µM)20E (1 µM)
HlVg-2HlVg-3
HlACTIN
- + - + - - + +
A
B
Fig. 7. Impact of rapamycin on vitellogenesis. (A) The fat bodies of femaleHaemaphysalis longicornis 6 days after engorgement were treated with or withoutrapamycin in vitro. The protein was extracted from the fat bodies after treatment.Immunoblotting was performed as in Fig. 3. (B) The fat bodies of female ticks 6 daysafter engorgement were incubated as above and then incubated with or without 20-hydroxyecdysone (20E). Protein extraction and immunoblotting were the same asin (A).
6 R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx
tion of S6K (Brown et al., 1995; Vilella-Bach et al., 1999). The non-catalytic C-terminal FATC domain is also necessary to control S6K
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
(Takahashi et al., 2000). RNAi using each dsRNA fragment encom-passing part of the N-terminus, FAT, FBP and kinase domains af-fected the expression of Vg mRNA (Patel et al., 2007; Maestroet al., 2009; Suganya et al., 2010; Parthasarathy and Palli, 2011).In this study, the C-terminal sequence encoding a partial kinase do-main and the entire FATC domains was used as a template fordsRNA synthesis. As a result, the mRNA expression levels of HlGAT-A, HlVg-2 and HlVg-3 were reduced in the fat bodies of HlTOR-RNAiticks compared with those of the controls. By immunoblotting, itwas found that HlTOR-RNAi led to inhibition of HlS6K phosphory-lation in the fat body of engorged ticks (Fig. 3). We recently re-ported that HlS6K knockdown led to decreased transcription ofHlGATA and HlVg genes (Boldbaatar et al., 2010a). Therefore, thepresent results suggest that HlTOR functions in the upregulationof HlGATA and HlVgs and that HlTOR is an activator of HlS6K. Inthe mosquito, A. aegypti, the AaGATA protein level increased aftera blood meal in the fat body, and AaGATA was located in boththe cytoplasm and nuclei of fat body cells (Park et al., 2006). Thisstudy also showed that rapamycin inhibited amino acid-activatedelevation of the GATA protein level. Similarly, HlTOR-RNAi reducedthe amount of HlGATA protein in the fat body at 4 days of feeding.Immunostaining of the fat bodies with the anti-HlGATA antibodydemonstrated that HlGATA was localised in the cytoplasm butnot in the nuclei of trophocytes in HlTOR-RNAi ticks at 4 days offeeding (Fig. 4E and F). The impact of HlTOR RNAi on the translationand localisation of HlGATA appeared to be more effective at 4 daysof feeding than at engorgement. These results suggest that theactivity of HlTOR is essential for the translation and nuclear local-isation of HlGATA in the fat body, as in A. aegypti. Together with
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002

R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx 7
the results of the decreased amount of HlVg proteins in the fatbodies of HlTOR-RNAi ticks, these data further support our specula-tion that HlTOR is an essential component of vitellogenesis via acti-vation of HlS6K and HlGATA in the tick. This raises the possibilitythat the molecular mechanism regulating vitellogenesis is con-served in ticks and anautogeny insects.
There was a significant difference in engorged body weight atengorgement between HlTOR-RNAi and control ticks (Table 1).With the exception of the ovary, no remarkable differences ingrowth between HlTOR-RNAi and control ticks were observed inother internal organs. TOR is essential for normal growth anddevelopment in eukaryotes. Drosophila TOR (dTOR) is required forcellular growth and proliferation during larval development (Old-ham et al., 2000; Zhang et al., 2000). In the case of adult flies, dTORmutants were approximately half the size of the wild type andwere developmentally delayed by 2 days. Ovaries of these mutantsdisplayed arrested development in the pre-vitellogenic stage. Inthe cricket, Gryllus bimaculatus, nymphs treated with dsRNAsagainst insulin-signalling components, including Insulin receptor(InR), Insulin receptor substrate (chico), Tor, S6k and Epidermalgrowth factor receptor (Egfr), displayed smaller body sizes (Dabouret al., 2011). In the current study, HlTOR-RNAi ticks had completedblood feeding normally, but their ovaries exhibited delayed devel-opment compared with those of control ticks. This can be attrib-uted to low Vg gene/protein expressions in the fat bodies, whichwere induced by silencing of HlTOR; as a result, the rate of ovipo-sitioned ticks was lower in RNAi ticks than in control ticks.
The insulin/PI3K/Akt signalling pathway is believed to beimportant for development and reproduction in ticks. Silencing ofinsulin-like growth factor-binding protein-related proteins causeda reduction of body weight at engorgement, a lethal effect for en-gorged ticks, and failure to lay eggs in the hard tick, Amblyommaamericanum (Mulenga and Khumthong, 2010). Recently, we identi-fied a homologue of Akt from H. longicornis (HlAkt) (Umemiya-Shir-afuji et al., 2012). Knockdown of HlAkt by RNAi caused theinhibition of blood feeding in female ticks and the arrest of growthin their internal organs, indicating that HlAkt is involved in thegrowth of cells/organs during blood feeding. Interestingly, HlAkt-RNAi affected the expression of HlVgs in female ticks. This resultcoincides with the report that RNAi-mediated Akt gene knock-downs effectively block stimulation of Vg gene expression by insu-lin and 20E in the mosquito (Roy et al., 2007). Since Akt is known asan upstream factor in the TOR pathway in insects (Miron et al.,2003), HlAkt may also be a component involved in vitellogenesisin the tick. Further research is needed to determine which factor(s)activates the TOR signalling pathway to initiate vitellogenesis inticks.
Vitellogenesis is controlled by JH in the cockroach (Maestroet al., 2009). Deitsch et al. (1995) discovered that yolk proteinmRNA expression is indirectly controlled by 20E in the mosquito.This expression is activated after a blood meal through the syner-gistic action of not only 20E and the amino acid/TOR-signallingpathway but also insulin signalling in the fat body (Hansen et al.,2004; Roy et al., 2007). In soft ticks, JH does not function in the reg-ulation of vitellogenin synthesis (Chinzei et al., 1991; Taylor et al.,1991). In addition, an insect-type JH has never been identified inboth hard and soft ticks. Sankhon et al. (1999) demonstrated thatVg production in fat body trophocytes of the hard tick, D. variabilis,is regulated by 20E by in vitro fat body culture. Although they mea-sured Vg in the medium produced from the fat body after additionof 20E, there is a lack of information on the direct effects of 20E onVg synthesis in the fat body cells of ticks. In the present study, Vgwas found in increasing amounts in the fat bodies, in response to20E. This increase was inhibited by rapamycin, indicating thatTOR activity is crucial for 20E-stimulated Vg synthesis in the fatbody.
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
In summary, we present preliminary evidence that HlTOR acti-vates HlS6K and regulates the transcription and intracellular local-isation of HlGATA factor, which is a crucial step for vitellogenesis inticks. Data in this study also suggests that HlTOR activity is re-quired for the facilitation of HlVg synthesis by 20E. In the absenceof a completed H. longicornis genome sequence, it is difficult todetermine the specificity of our RNAi phenotype observations.We may be able to confirm the specificity of knockdown pheno-types by minimising the size of the RNAi treatments (�100 bp)to determine whether the HlTOR knockdown effects are similarto those observed here. The present study will contribute to betterunderstanding of the molecular mechanisms involved in tickreproduction. Female hard ticks produce thousands of eggs, andsome pathogens are transovarially transmitted. Investigating theregulatory network underlying vitellogenesis would provide valu-able information to develop new methods for controlling ticks aswell as tick-borne pathogens.
Acknowledgements
We would like to thank Dr. DeMar Taylor (University of Tsu-kuba, Japan) for advice on the manuscript. We are also gratefulto Toru Tanaka, Suguru Kawano, Aiko Kume and Hiroki Maedafor their technical assistance. This work was supported by theBio-oriented Technology Research Advancement Institution(BRAIN), a Grant-in-Aid for Scientific Research (A), and a Grant-in-Aid for Research Activity Start-up of the Japan Society for thePromotion of Science (JSPS). R. Umemiya-Shirafuji was supportedby a Grant-in-Aid for JSPS Fellows.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.ijpara.2012.08.002.
References
Balashov, Y.S., 1972. Bloodsucking ticks (Ixodoidea) – vectors of disease of man andanimals (English translation). Misc. Pub. Entomol. Soc. America 8, 161–376.
Boldbaatar, D., Battur, B., Umemiya-Shirafuji, R., Liao, M., Tanaka, T., Fujisaki, K.,2010a. GATA transcription, translation and regulation in Haemaphysalislongicornis tick: analysis of the cDNA and an essential role for vitellogenesis.Insect Biochem. Mol. Biol. 40, 49–57.
Boldbaatar, D., Umemiya-Shirafuji, R., Liao, M., Tanaka, T., Xuan, X., Fujisaki, K.,2010b. Multiple vitellogenins from the Haemaphysalis longicornis tick are crucialfor ovarian development. J. Insect Physiol. 56, 1587–1598.
Bownes, M., Scott, A., Shirras, A., 1988. Dietary components modulate yolk proteingene transcription in Drosophila melanogaster. Development 103, 119–128.
Brown, E.J., Beal, P.A., Keith, C.T., Chen, J., Shin, T.B., Schreiber, S.L., 1995. Control ofp70 S6 kinase by kinase activity of FRAP in vivo. Nature 377, 441–446.
Buszczak, M., Freeman, M.R., Carlson, J.R., Bender, M., Cooley, L., Segraves, W.A.,1999. Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 126, 4581–4589.
Chinzei, Y., Taylor, D., Ando, K., 1991. Effects of juvenile hormone and its analogs onvitellogenin synthesis and ovarian development in Ornithodoros moubata(Acari: Argasidae). J. Med. Entomol. 28, 506–513.
Chinzei, Y., Yano, I., 1985. Fat body is the site of vitellogenin synthesis in the softtick, Ornithodoros moubata. J. Comp. Physiol. B. 155, 671–678.
Comas, D., Piulachs, M.D., Belles, X., 2001. Induction of vitellogenin genetranscription in vitro by juvenile hormone in Blattella germanica. Mol. Cell.Endocrinol. 183, 93–100.
Coons, L.B., Tarnowski, B., Ourth, D.D., 1982. Rhipicephalus sanguinius: localization ofvitellogenin synthesis by immunological methods and electron microscopy.Exp. Parasitol. 54, 331–339.
Dabour, N., Bando, T., Nakamura, T., Miyawaki, K., Mito, T., Ohuchi, H., Noji, S., 2011.Cricket body size is altered by systemic RNAi against insulin signalingcomponents and epidermal growth factor receptor. Dev. Growth Differ. 53,857–869.
Deitsch, K.W., Chen, J.S., Raikhel, A.S., 1995. Indirect control of yolk protein genes by20-hydroxyecdysone in the fat body of the mosquito, Aedes aegypti. InsectBiochem. Mol. Biol. 25, 449–454.
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002

8 R. Umemiya-Shirafuji et al. / International Journal for Parasitology xxx (2012) xxx–xxx
Fei, H., Martin, T.R., Jaskowiak, K.M., Hatle, J.D., Whitman, D.W., Borst, D.W., 2005.Starvation affects vitellogenin production but not vitellogenin mRNA levels inthe lubber grasshopper, Romalea microptera. J. Insect Physiol. 51, 435–443.
Friesen, K.J., Kaufman, W.R., 2002. Quantification of vitellogenesis and its control by20-hydroxyecdysone in the ixodid tick, Amblyomma hebraeum. J. Insect Physiol.48, 773–782.
Fujisaki, K., 1978. Development of acquired resistance precipitating antibody inrabbits experimentally infested with females of Haemaphysalis longicornis(Ixodoidea: Ixodidae). Natl. Inst. Anim. Health Q (Tokyo) 18, 27–38.
Gong, H., Liao, M., Zhou, J., Hatta, T., Huang, P., Zhang, G., Kanuka, H., Nishikawa, Y.,Xuan, X., Fujisaki, K., 2008. Gene silencing of ribosomal protein P0 is lethal tothe tick Haemaphysalis longicornis. Vet. Parasitol. 151, 268–278.
Grandjean, O., 1983. Blood digestion in Ornithodorus moubata Murray sensu strictoWalton females (Ixodoidea: Argasidae) II. Modifications of midgut cells relatedto the digestive cycle and to the triggering action of mating. Ann. Parasitol.Hum. Comp. 58, 493–514.
Granjeno-Colin, G., Hernandez-Ortiz, R., Mosqueda, J., Estrada-Mondaca, S.,Figueroa, J.V., Garcia-Vazquez, Z., 2008. Characterization of a vitellogeningene fragment in Boophilus microplus ticks. Ann. N. Y. Acad. Sci. 1149, 58–61.
Hansen, I.A., Attardo, G.M., Park, J.H., Peng, Q., Raikhel, A.S., 2004. Target ofrapamycin-mediated amino acid signaling in mosquito anautogeny. Proc. Natl.Acad. Sci. USA 101, 10626–10631.
Hansen, I.A., Attardo, G.M., Roy, S.G., Raikhel, A.S., 2005. Target of rapamycin-dependent activation of S6 kinase is a central step in the transduction ofnutritional signals during egg development in a mosquito. J. Biol. Chem. 280,20565–20572.
Hatta, T., Tsuji, N., Miyoshi, T., Islam, M.K., Alim, M.A., Yamaji, K., Anisuzzaman,Fujisaki, K., 2010. Leucine aminopeptidase, HlLAP, from the ixodid tickHaemaphysalis longicornis, plays vital roles in the development of oocytes.Parasitol. Int. 59, 286–289.
Hoogstraal, H., Roberts, F.H., Kohls, G.M., Tipton, V.J., 1968. Review of Haemaphysalis(Kaiseriana) longicornis Neumann (resurrected) of Australia, New Zealand, NewCaledonia, Fiji, Japan, Korea, and Northeastern China and USSR, and itsparthenogenetic and bisexual populations (Ixodoidea, Ixodidae). J. Parasitol.54, 1197–1213.
Horigane, M., Shinoda, T., Honda, H., Taylor, D., 2010. Characterization of avitellogenin gene reveals two phase regulation of vitellogenesis byengorgement and mating in the soft tick Ornithodoros moubata (Acari:Argasidae). Insect Mol. Biol. 19, 501–515.
Inoki, K., Ouyang, H., Li, Y., Guan, K.L., 2005. Signaling by target of rapamycinproteins in cell growth control. Microbiol. Mol. Biol. Rev. 69, 79–100.
James, A.M., Zhu, X.X., Oliver, J.H., 1997. Vitellogenin and ecdysteroid titers in Ixodesscapularis during vitellogenesis. J. Parasitol. 83, 559–563.
James, A.M., Zhu, X.X., Oliver, J.H., 1999. Localization of vitellogenin production inthe blacklegged tick, Ixodes scapularis (Acari: Ixodidae). Invertebr. Reprod. Dev.35, 81–87.
Khalil, S.M.S., Donohue, K.V., Thompson, D.M., Jeffers, L.A., Ananthapadmanaban, U.,Sonenshine, D.E., Mitchell, R.D., Roe, R.M., 2011. Full-length sequence,regulation and developmental studies of a second vitellogenin gene from theAmerican dog tick, Dermacentor variabilis. J. Insect Physiol. 57, 400–408.
Kokoza, V.A., Martin, D., Mienaltowski, M.J., Ahmed, A., Morton, C.M., Raikhel, A.S.,2001. Transcriptional regulation of the mosquito vitellogenin gene via a bloodmeal-triggered cascade. Gene 274, 47–65.
Lara, F.A., Lins, U., Bechara, G.H., Oliveira, P.L., 2005. Tracing heme in a living cell:hemoglobin degradation and heme traffic in digest cells of the cattle tickBoophilus microplus. J. Exp. Biol. 208, 3093–3101.
Liao, M., Boldbaatar, D., Gong, H., Huang, P., Umemiya, R., Harnnoi, T., Zhou, J.,Tanaka, T., Suzuki, H., Xuan, X., Fujisaki, K., 2008. Functional analysis of proteindisulfide isomerases in blood feeding, viability and oocyte development inHaemaphysalis longicornis ticks. Insect Biochem. Mol. Biol. 38, 285–295.
Maestro, J.L., Cobo, J., Belles, X., 2009. Target of rapamycin (TOR) mediates thetransduction of nutritional signals into juvenile hormone production. J. Biol.Chem. 284, 5506–5513.
Miron, M., Lasko, P., Sonenberg, N., 2003. Signaling from Akt to FRAP/TOR targetsboth 4E-BP and S6K in Drosophila melanogaster. Mol. Cell. Biol. 23, 9117–9126.
Miyoshi, T., Tsuji, N., Islam, M.K., Kamio, T., Fujisaki, K., 2004. Gene silencing of acubilin-related serine proteinase from the hard tick Haemaphysalis longicornisby RNA interference. J. Vet. Med. Sci. 66, 1471–1473.
Mulenga, A., Khumthong, R., 2010. Silencing of three Amblyomma americanum (L.)insulin-like growth factor binding protein-related proteins prevents ticks fromfeeding to repletion. J. Exp. Biol. 213, 1153–1161.
Oldham, S., Montagne, J., Radimerski, T., Thomas, G., Hafen, E., 2000. Genetic andbiochemical characterization of dTOR, the Drosophila homolog of the target ofrapamycin. Genes Dev. 14, 2689–2694.
Panaitof, S.C., Scott, M.P., 2006. Effect of juvenile hormone on vitellogenin geneexpression in the fat body of burying beetles, Nicrophorus orbicollis. Arch. InsectBiochem. Physiol. 63, 82–91.
Please cite this article in press as: Umemiya-Shirafuji, R., et al. Target of rapambody of the tick, Haemaphysalis longicornis. Int. J. Parasitol. (2012), http://dx.do
Park, J.H., Attardo, G.M., Hansen, I.A., Raikhel, A.S., 2006. GATA factor translation isthe final downstream step in the amino acid/target-of-rapamycin-mediatedvitellogenin gene expression in the anautogenous mosquito Aedes aegypti. J.Biol. Chem. 281, 11167–11176.
Parthasarathy, R., Palli, S.R., 2011. Molecular analysis of nutritional and hormonalregulation of female reproduction in the red flour beetle, Tribolium castaneum.Insect Biochem. Mol. Biol. 41, 294–305.
Parthasarathy, R., Sheng, Z.T., Sun, Z.Y., Palli, S.R., 2010a. Ecdysteroid regulation ofovarian growth and oocyte maturation in the red flour beetle, Triboliumcastaneum. Insect Biochem. Mol. Biol. 40, 429–439.
Parthasarathy, R., Sun, Z.Y., Bai, H., Palli, S.R., 2010b. Juvenile hormone regulation ofvitellogenin synthesis in the red flour beetle, Tribolium castaneum. InsectBiochem. Mol. Biol. 40, 405–414.
Patel, A., Fondrk, M.K., Kaftanoglu, O., Emore, C., Hunt, G., Frederick, K., Amdam,G.V., 2007. The making of a queen: TOR pathway is a key player in dipheniccaste development. PLoS One 2, e509.
Rosell, R., Coons, L.B., 1992. The role of the fat body, midgut and ovary invitellogenin production and vitellogenesis in the female tick, Dermacentorvariabilis. Int. J. Parasitol. 22, 341–349.
Rosell-Davis, R., Coons, L.B., 1989. Relationship between feeding, mating,vitellogenin production and vitellogenesis in the tick Dermacentor variabilis.Exp. Appl. Acarol. 7, 95–105.
Roy, S.G., Hansen, I.A., Raikhel, A.S., 2007. Effect of insulin and 20-hydroxyecdysonein the fat body of the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol.Biol. 37, 1317–1326.
Roy, S.G., Raikhel, A.S., 2011. The small GTPase Rheb is a key component linkingamino acid signaling and TOR in the nutritional pathway that controls mosquitoegg development. Insect Biochem. Mol. Biol. 41, 62–69.
Sankhon, N., Lockey, T., Rosell, R.C., Rothschild, M., Coons, L., 1999. Effect ofmethoprene and 20-hydroxyecdysone on vitellogenin production in culturedfat bodies and backless explants from unfed female Dermacentor variabilis. J.Insect Physiol. 45, 755–761.
Suganya, R., Chen, S.L., Lu, K.H., 2010. Target of rapamycin in the oriental fruit flyBactrocera dorsalis (Hendel): its cloning and effect on yolk protein expression.Arch. Insect Biochem. Physiol. 75, 45–56.
Suganya, R., Chen, S.L., Lu, K.H., 2011. CDNA cloning and characterization of S6kinase and its effect on yolk protein gene expression in the oriental fruit flyBactrocera dorsalis (Hendel). Arch. Insect Biochem. Physiol. 78, 177–189.
Takahashi, T., Hara, K., Inoue, H., Kawa, Y., Tokunaga, C., Hidayat, S., Yoshino, K.,Kuroda, Y., Yonezawa, K., 2000. Carboxyl-terminal region conserved amongphosphoinositide-kinase-related kinases is indispensable for mTOR functionin vivo and in vitro. Genes Cells 5, 765–775.
Tarnowski, B.I., Coons, L.B., 1989. Ultrastructure of the midgut and blood mealdigestion in the adult tick Dermacentor variabilis. Exp. Appl. Acarol. 6, 263–289.
Taylor, D., Chinzei, Y., Miura, K., Ando, K., 1991. Vitellogenin synthesis, processingand hormonal regulation in the tick, Ornithodoros parkeri (Acari: Argasidae).Insect Biochem. 21, 723–733.
Thompson, D.M., Khalil, S.M.S., Jeffers, L.A., Ananthapadmanaban, U., Sonenshine,D.E., Mitchell, R.D., Osgood, C.J., Apperson, C.S., Roe, R.M., 2005. In vivo role of20-hydroxyecdysone in the regulation of the vitellogenin mRNA and eggdevelopment in the American dog tick, Dermacentor variabilis (Say). J. InsectPhysiol. 51, 1105–1116.
Thompson, D.M., Khalil, S.M., Jeffers, L.A., Sonenshine, D.E., Mitchell, R.D., Osgood,C.J., Michael Roe, R., 2007. Sequence and the developmental and tissue-specificregulation of the first complete vitellogenin messenger RNA from ticksresponsible for heme sequestration. Insect Biochem. Mol. Biol. 37, 363–374.
Tufail, M., Naeemullah, M., Elmogy, M., Sharma, P.N., Takeda, M., Nakamura, C.,2010. Molecular cloning, transcriptional regulation, and differential expressionprofiling of vitellogenin in two wing-morphs of the brown planthopper,Nilaparvata lugens Stal (Hemiptera: Delphacidae). Insect Mol. Biol. 19, 787–798.
Umemiya, R., Matsuo, T., Hatta, T., Sakakibara, S., Boldbaatar, D., Fujisaki, K., 2007.Cloning and characterization of an autophagy-related gene, ATG12, from thethree-host tick Haemaphysalis longicornis. Insect Biochem. Mol. Biol. 37, 975–984.
Umemiya-Shirafuji, R., Matsuo, T., Liao, M., Boldbaatar, D., Battur, B., Suzuki, H.,Fujisaki, K., 2010. Increased expression of ATG genes during nonfeeding periodsin the tick Haemaphysalis longicornis. Autophagy 6, 473–481.
Umemiya-Shirafuji, R., Tanaka, T., Boldbaatar, D., Fujisaki, K., 2012. Akt is anessential player in regulating cell/organ growth at the adult stage in the hardtick Haemaphysalis longicornis. Insect Biochem. Mol. Biol. 42, 164–173.
Vilella-Bach, M., Nuzzi, P., Fang, Y., Chen, J., 1999. The FKBP12-rapamycin-bindingdomain is required for FKBP12-rapamycin-associated protein kinase activityand G1 progression. J. Biol. Chem. 274, 4266–4272.
Zhang, H., Stallock, J.P., Ng, J.C., Reinhard, C., Neufeld, T.P., 2000. Regulation ofcellular growth by the Drosophila target of rapamycin dTOR. Genes Dev. 14,2712–2724.
ycin (TOR) controls vitellogenesis via activation of the S6 kinase in the fati.org/10.1016/j.ijpara.2012.08.002




















