
Tannins present in Cichorium intybus enhance glucose uptake and inhibit adipogenesis in 3T3-L1...
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Tannins present in Cichorium intybus enhance glucose uptake and inhibit adipogenesis in 3T3-L1...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Chemico-Biological Interactions 174 (2008) 69–78
Contents lists available at ScienceDirect
Chemico-Biological Interactions
journa l homepage: www.e lsev ier .com/ locate /chembio int
Tannins present in Cichorium intybus enhance glucoseuptake and inhibit adipogenesis in 3T3-L1 adipocytesthrough PTP1B inhibition
V.S. Muthusamya, S. Ananda, K.N. Sangeethaa, S. Sujathaa,Balakrishnan Arunb, B.S. Lakshmia,∗
a Centre for Biotechnology, Tissue Culture and Drug Discovery Lab, Anna University, Chennai 600025, Tamilnadu, Indiab Department of Pharmacology, Nicholas Piramal Research Centre, Mumbai 400063, India
a r t i c l e i n f o
Article history:Received 21 December 2007Received in revised form 12 April 2008Accepted 15 April 2008Available online 24 April 2008
Keywords:Cichorium intybusTanninsInsulin resistanceAdipogenesisPTP1B
a b s t r a c t
Insulin resistance is a fundamental aspect for the etiology of non-insulin dependent diabetesmellitus (NIDDM) and has links with a wide array of secondary disorders including weightgain and obesity. The present study analyzes the effect of Cichorium intybus methanolic(CME) extract on glucose transport and adipocyte differentiation in 3T3-L1 cells by studyingthe radiolabelled glucose uptake and lipid accumulation assays, respectively. By performingdetannification (CME/DT), the role of tannins present in CME on both the activities was eval-uated. CME and CME/DT exhibited significant glucose uptake in 3T3-L1 adipocytes with adose-dependent response. Glucose uptake profile in the presence of PI3K and IRTK inhibitors(Wortmannin and Genistein) substantiates the mechanism used by both the extracts. CMEinhibited the differentiation of 3T3-L1 preadipocytes but failed to show glucose uptakein inhibitor treated cells. The activity exhibited by CME/DT is exactly vice versa to CME.Furthermore, the findings from PTP1B inhibition assay, mRNA and protein expression anal-ysis revealed the unique behavior of CME and CME/DT. The duality exhibited by C. intybusthrough adipogenesis inhibition and PPAR� up regulation is of interest. Current observationconcludes that the activities possessed by C. intybus are highly desirable for the treatmentof NIDDM because it reduces blood glucose levels without inducing adipogenesis in 3T3-L1adipocytes.
© 2008 Elsevier Ireland Ltd. All rights reserved.
Abbreviations: CME, Cichorium intybus methanolic extract; CME/DT, Cichorium intybus detannified methanolic extract; DMEM, Dulbecco’s Modified EagleMedium; IBMX, 3-isobutyl-1-methyl xanthine; DEX, dexamethasone; DMSO, dimethyl sulphoxide; FBS, foetal bovine serum; KRPH, krebs ringer phosphateHEPES buffer; HEPES, N-2-hydroxyethypiperazine-N′-2-ethanesulfonic acid; SDS, sodium dodecyl sulphate; LDH, lactate dehydrogenase; Rosy, rosiglitazone;PI3K, phosphatidylinositol 3-kinase; GLUT4, glucose transporter 4; PPAR�, peroxisome proliferator activator receptor gamma; PTP1B, protein tyrosinephosphatase 1B; C/EBP, CCAAT/enhancer binding protein; SREBP, sterol regulatory element binding protein; IRS, insulin receptor substrate; WT, Wortmannin;GS, Genistein; ALP, alkaline phosphatase; NBT, nitroblue tetrazolium; BCIP, 5-bromo-4-chloro-3-indolylphosphate; IDV, integrated density value.
∗ Corresponding author. Tel.: +91 44 22350772; fax: +91 44 22350299.E-mail addresses: [email protected] (V.S. Muthusamy), anand [email protected] (S. Anand), sangee22 [email protected]
(K.N. Sangeetha), [email protected] (S. Sujatha), [email protected] (B. Arun), [email protected] (B.S. Lakshmi).
0009-2797/$ – see front matter © 2008 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.cbi.2008.04.016

Author's personal copy
70 V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78
1. Introduction
Obesity combined with insulin resistance is a majorrisk factor for the prevalence of non-insulin dependentdiabetes mellitus (NIDDM). Adipocyte is the only appar-ent link between diabetes and obesity, which stores excessglucose in the form of fat by changing dramatically in itssize in accordance with metabolic needs. Since the roleof adipocyte in energy homeostasis is vital, its dysfunc-tion can negatively influence other systems [1]. The lossof insulin action selectively in adipose tissue leads to sec-ondary disorders of diabetes through the malfunctioningof adipocyte function, changes in adipogenesis, alterationsin glucose and lipid metabolism, and protein expression[2]. Now, wide varieties of pharmacological drugs are beingused for NIDDM treatment. However, currently availableanti-diabetic drugs (e.g., thiazolidinedione) promote obe-sity and hyperandrogenemia, while reducing blood glucose[3]. In the current scenario, the strategy to reduce hyper-glycemia without increasing adiposity or with reduction ofbodyweight constitutes a preferred one for the drug devel-opment of NIDDM.
Botanicals are thought of offering strong potential withminimal side affects particularly against metabolic syn-dromes as most of their efficacies are from a mixture ofactive molecules acting at the same time [4]. The plantselected for the present study is Cichorium intybus (Fam-ily: Compositeae) a salad crop known as Chicory, Frenchendive, and succory, native of Asia and this plant is mainlycultivated for its roots that contains a high content ofwell-known fructopolymer inulin. C. intybus is known tocontain anthocyanins, phenols [5], sesquiterpenes such asguaianolides, eudesmanolides, and germacranolides [6]. Itis also cultivated for its food valuable sprouts. Earlier inves-tigations have reported that the ethanolic extract of C.intybus has anti-diabetic and hypolipidemic activities instreptozotocin-induced diabetic rats [7]. It is being used bytraditional healers of South Indian villages for the treat-ment of NIDDM (through personal communication withTamil Nadu Medicinal Plants Corporation Limited, KolliHills, South India).
The present study was commenced with the aim of eval-uating the methanolic (CME) and detannified methanolicextract (CME/DT) of C. intybus leaves for anti-diabetic andadipogenesis inhibition potential since the two differentactivities working together seem to be an ideal combina-tion for the treatment of insulin resistance and obesity.Tannins are plant polyphenols with the molecular weightof 500–3000 Da. Tannins are widely found in food plantsand are broadly applied for various industrial needs. In bio-logical systems, tannins are known to be a potent metalion chelators, protein precipitating agents and antioxidants[8]. The importance of tannins in plant based drug dis-covery approach is meager because of its high molecularweight and complex nature [9]. Nevertheless, Liu et al. [10]reported that tannins are significant molecules in banabaextract (Lagerstroemia speciosa) which exhibit anti-diabeticand adipogenesis inhibition activities in 3T3-L1 cells. Sincethe leaves of C. intybus have been reported for presence oftannins, the detannification analysis was initiated to eval-uate the biological significance of tannins present in the
plant. In addition, we have examined the mechanisms bywhich CME and CME/DT mediates the glucose transportand adipogenesis inhibition activities.
2. Materials and methods
2.1. Chemicals and reagents
All cell culture solutions and supplements were pur-chased from Life Technologies Inc. (Gaithersburg, MD,U.S.A.). Dulbecco’s Modified Eagle Medium (DMEM) wasobtained from GIBCO, BRL (Carlsbad, CA, U.S.A.). 2-Deoxy-d-3[H] glucose was obtained from Amersham PharmaciaBiotech (Buckinghamshire, U.K.). TRIzol reagent and MMLVreverse transcriptase, dNTP, Taq polymerase was obtainedfrom GIBCO BRL (Carlsbad, CA, U.S.A.), USA and NewEngland Biolabs (Herts, U.K.), respectively. Insulin, 3-isobutyl-1-methyl xanthine (IBMX), dexamethasone (DEX)and Genistein were obtained from Sigma (Andover, U.K.).Wortmannin was obtained from Calbiochem (Darmstadt,Germany). Insulin and Protein A Sepharose beads wereobtained from Sigma (Andover, U.K.). IR and IRS antibodieswere procured from BD Pharmingen (San Diego, CA, U.S.A.).Rosiglitazone was a kind gift from Dr. Reddy’s Laboratories,Hyderabad. Recombinant human PTP1B was obtained fromBiomol Research Laboratories, Inc. (PA, U.S.A.). Primerswere synthesized from GIBCO, BRL (Carlsbad, CA, U.S.A.).All fine chemicals were obtained from Sigma (Andover,U.K.). All other chemicals and organic solvents used wereof the highest analytical grade. Leaves of C. intybus werecollected from Trichy, Tamilnadu, India. The collected mate-rial has been authenticated and the vouchers are stored atthe Department of Life sciences, Plant breeding and tissueculture laboratory, Bharathidasan University, Trichy, Tamil-nadu, India.
2.2. Cell culture of 3T3-L1 adipocytes
3T3-L1 preadipocytes (obtained from ATCC-CL-173) wascultured in DMEM with 10% FBS and supplemented withpenicillin (120 units/ml), streptomycin (75 �g/ml), gen-tamycin (160 �g/ml) and amphotericin B (3 �g/ml) in 5%CO2 environment. 3T3-L1 preadipocytes grown in 24 wellplates until 2 days postconfluence and the cells wereinduced by the differentiation medium (combination of0.5 mmol/l of IBMX, 0.25 �mol/l of DEX and 1 mg/l ofinsulin in DMEM medium with 10% FBS) to differentiateinto adipocytes. Three days after induction, the differentia-tion medium was replaced with medium containing 1 mg/linsulin alone. The medium was subsequently replacedagain with fresh culture medium (DMEM with 10% FBS)after 2 days. By monitoring the formation of multinucle-ation in cells, the extent of differentiation was measured.
2.3. Extraction and detannification
An authenticated, dried, pulverized leaves of C. intybus(100 g) was subjected to sequential extraction using differ-ent solvents with increasing polarity (hexane, ethyl acetate,and methanol) by cold maceration technique. The extractswere filtered and concentrated to dryness in vacuo at 45 ◦C

Author's personal copy
V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78 71
to get a pasty mass of crude extract. The extracts were thenquantitatively reconstituted with DMSO and subjected for2-deoxy-d-1-3[H] glucose uptake assay.
Detannified extract was prepared by previouslydescribed gelatin procedure [11] with minor modifica-tions. In the detannification process, 100 mg/ml extractwas prepared in methanol and centrifuged for 10 min at4000 rpm and filtered with a 0.2 �m filter; 40 mg/ml ofgelatin was added with the filtered extract in the ratio of1:4, which was then centrifuged for 10 min at 4000 rpm.The supernatant was transferred to a new tube and thepellet, which contained tannin and gelatin, was discarded.This step was repeated twice to completely remove tannin.The supernatant was collected in a new tube, and mixedwith same volume of 40 mg/ml gelatin, and centrifugedat 8000 rpm for 20 min to remove residual tannin. Theresulting supernatant was transferred to a new tube andthe pellet discarded. A 5-fold volume of ethanol was addedto remove excess gelatin in the remaining extract; thissolution was then centrifuged at 8000 rpm for 10 min, andthe final tannin-free supernatant was transferred to a newtube and vacuum-dried. Methanolic (CME) and detannifiedmethanolic (CME/DT) extracts were subsequently testedfor glucose uptake activity.
2.4. Tannin estimation
The amount of tannins present in the extracts ofCME and CME/DT were estimated by Folin-Ciocalteucolorimetric method [12] using Gallic acid as a stan-dard. The quantitatively reconstituted extracts (1 mg/ml inmethanol) were incubated with Folin-phenol reagent inthe presence of alkaline medium (sodium carbonate) for30 min at room temperature. The OD measurement wastaken at 640 nm for the estimation of tannin, and from thecalibration curve the tannin content was calculated.
2.5. Measurement of 2-deoxy-d-3[H] glucose
3T3-L1 adipocytes grown in 24-well plate (BD Falcon)were subjected to glucose uptake as reported [13] withminor modifications. After differentiation induction for thestipulated period, the cells were incubated with extracts for24 h. The cells were then stimulated with Insulin (100 nM)for 20 min followed by rinsing with Krebs Ringer phos-phate HEPES (KRPH) solution (118 mM NaCl, 5 mM KCl,1.3 mM CaCl2, 1.2 mM MgSO4, 1.2 mM KH2PO4 and 30 mMHEPES–pH 7.4). The cells were subsequently pulsed for20 min in KRPH solution containing 0.5 �Ci/ml 2-deoxy-d-3[H] glucose. The assay was terminated by aspiratingthe medium. The cells were then washed thrice with ice-cold KRPH solution and lysed in 0.1% SDS. The lysate wastransferred to a 96-well plate (Packard) with glass fiberpaper and air dried overnight. The radioactivity of the sam-ples was measured using a Top count liquid scintillationcounter (Packard, Ramsey, MN, U.S.A.). Glucose uptake wasalso checked in the presence of 100 nM of Wortmannin(WT) [14], a selective phosphatidylinositol 3-kinase (PI3K)inhibitor and 50 �M of Genistein (GS) [15], an IRTK inhibitorto understand the mechanism of action. All the assays wereperformed in duplicates for concordance. Results were
expressed as % glucose uptake with respect to solvent con-trol. Rosiglitazone (50 �M) was used as the positive control.
2.6. Adipocyte differentiation assay
3T3-L1 preadipocytes differentiating in the presenceof CME and CME/DT were assessed for induced dif-ferentiation over non-treated cells by measuring theaccumulation of triglycerides [16]. Triglyceride contentwas measured using a commercially available kit (Adi-poRed assay Reagent; Lonza Walkersville, Inc., Walkersville,MD). Cells (5000 cells/well) were grown in 96-well plates.Two days after attaining confluence, proliferation mediumwas replaced with differentiation medium containingas described above, with logarithmic doses of CME,CME/DT ranging from 1 pg/ml to 10 �g/ml and Rosiglita-zone (50 �M). Rosiglitazone was used as a positive control.Alternatively, preadipocytes were maintained with freshFBS-DMEM every other day for the whole spectrum ofinduction period. AdipoRed, a solution of the hydrophilicstain Nile Red, is a reagent that enables the quantificationof intracellular lipid. Cells were washed with PBS (pH 7.4)and 200 �l of PBS was added to the wells. 5 �l of AdipoRedreagent was added to each well. After 10 min, the plateswere placed in the fluorometer (Flouroskan Ascent, Ther-moLab systems, Helsinki, Finland) and fluorescence wasmeasured with an excitation wavelength of 485 nm andemission wavelength of 572 nm.
2.7. PTP1B Inhibition study
The phosphatase activity was assayed using 20 mM ofp-nitro phenyl phosphatase (pNPP) as substrate and car-ried out in sodium acetate buffer, in a 96-well format [17].The reaction was initiated by the addition of recombinantPTP1B enzyme in to CME, CME/DT and Sodium orthovana-date (100 ng, 1 �g and 100 �M, respectively) pretreatedwells and incubated for 30 min at 37 ◦C. By measuring theOD at 405 nm, the rate of PTP 1B-catalyzed hydrolysis ofsubstrate measured. The non-enzymatic hydrolysis of pNPPwas normalized by measuring the increase in OD at 405 nmobtained in the absence of PTP1B enzyme.
2.8. Reverse transcriptase-polymerase chain reaction
RT-PCR was performed by according to the methoddescribed previously [18]. In brief, 3T3-L1 adipocytes afterexperimental incubation (15 min for Insulin and 18 hr forRosiglitazone, CME and CME/DT) of samples were imme-diately homogenized using total RNA isolation reagentTRIzol® reagent. The RNA obtained was then converted tocDNA by reverse transcription and subjected to PCR withspecific primers for GLUT 4, PI3K, C/EBP�, PPAR�, SREBP-1c and GAPDH. PCR products were run on 1.5% Agarosegels, stained with ethidium bromide and photographed.PCR-products were consistent with the predicted sizes. Todetermine the efficacy, Insulin and Rosiglitazone were usedat 100 nM and 50 �M dose, respectively, for all the markers[19,20]. The expression levels were quantitated by scanningon a gel documentation and analysis system (Chemi Imager4400, Alpha Innotech Corporation).
Author's personal copy
72 V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78
2.9. Immunoprecipitation and Western blot analysis
Total cell lysates were prepared as reported previously[21] with minor modifications. Anti-IRS-1 antibody (phos-phorylated peptide of the tyrosine 891 mouse IRS) wasadded to the eppendorf containing the cold lysates andincubated at 4 ◦C for 1 h. 50 �l of the Protein A Sepharosebeads was activated by washing with cold lysis buffer(500 �l) and centrifuged at 10,000 × g for 30 s in twice.Finally, 50 �l of washed Protein A Sepharose slurry wasadded to total protein (125 �g) isolated form CME andCME/DT treated adipocytes and immunoprecipitated for1 h at 4 ◦C on a rocking platform. Spin the eppendorf at10,000 × g for 1 min at 4 ◦C for washing thrice with 500 �lof Lysis Buffer. After the last wash, 50 �l of 1X laemmli sam-ple buffer was added to bead pellet, vortexed and heatedto 90–100 ◦C for 10 min. Again, spin the sample for 5 minat 10,000 × g followed by 10% SDS polyacrylamide gel elu-tion. After run the gel was electrophoretically transferredonto a nitrocellulose membrane at 120 mA for 90 min.Whereas for IR�, total protein (100 �g) isolated form CMEand CME/DT treated adipocytes were mixed with SDS sam-ple buffer and heated at 90–100 ◦C for 10 min followed by10% SDS polyacrylamide gel elution and electrophoreticaltransfer to a nitrocellulose membrane. Then, the mem-brane was blocked by blocking agent (5% skimmed milk)for overnight at 4 ◦C followed by primary antibody (1:250of anti-phospho IR�) incubation for 2 h. After TBS wash-ing and secondary antibody (ALP conjugate) incubation, theblot was visualized using NBT/BCIP chromogenic agent andphotographed.
2.10. Assessment of cytotoxicity by lactatedehydrogenase (LDH) release assay
Lactate dehydrogenase (LDH) release assay was per-formed [22] using a cytotox 96 assay kit (Promega). Thisassay quantitatively measures the LDH, a stable cytosolicenzyme released during cell lysis. The assay was done with0.2 × 106 cells/0.2 ml/well, seeded in 96-well cell cultureplates. Cells on treatment with different concentrations(ranging from 10 ng/ml to 10 �g/ml) of extracts from C.intybus in 3T3-L1 adipocyte, was measured at a time pointof 24 h. 0.05% Triton X-100 was used to induce maximallysis. The plate was read at 492 nm in a scanning multiwellspectrophotometer.
2.11. Statistical analysis
Data were expressed in mean ± S.E. Mean differencebetween the groups was analyzed by one way analysisof variance followed by Tukey’s multiple comparison test(using SPSS version 11.0 software, SPSS, Cary, NC, U.S.A.).p < 0.05 was considered as statistically significant.
3. Results
3.1. Extraction and detannification
The shade dried leaves of C. intybus was sequen-tially extracted with hexane, ethyl acetate and methanol
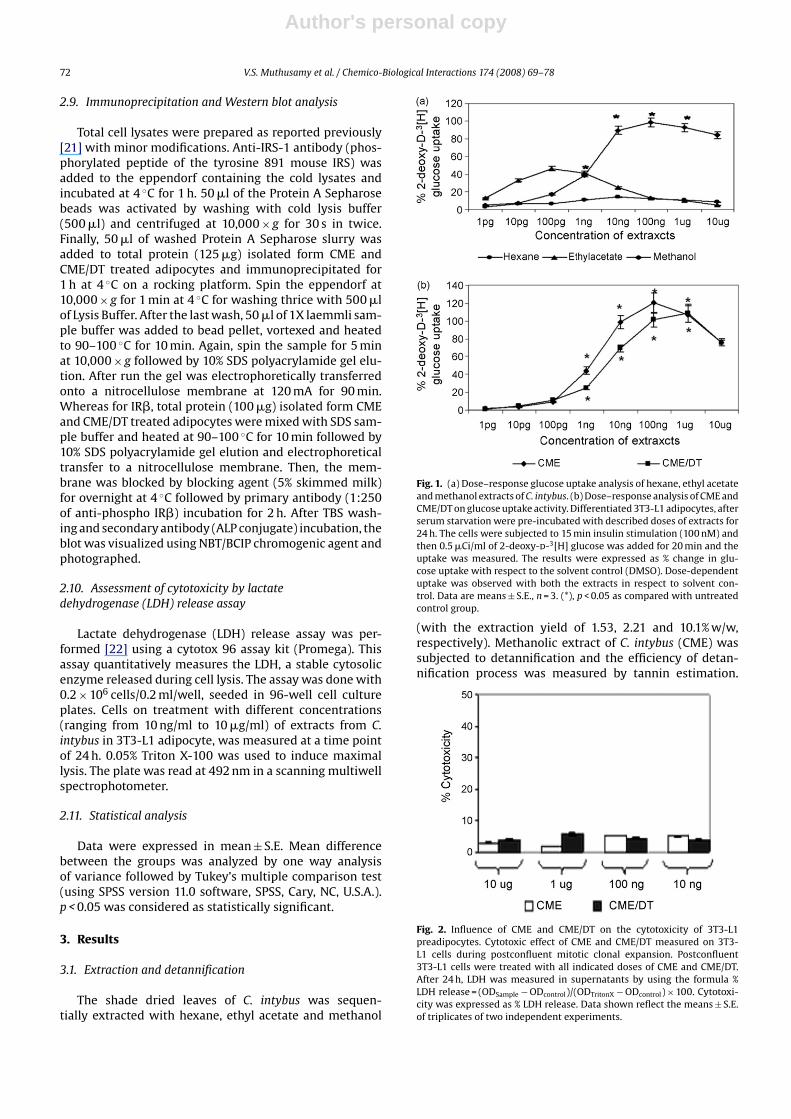
Fig. 1. (a) Dose–response glucose uptake analysis of hexane, ethyl acetateand methanol extracts of C. intybus. (b) Dose–response analysis of CME andCME/DT on glucose uptake activity. Differentiated 3T3-L1 adipocytes, afterserum starvation were pre-incubated with described doses of extracts for24 h. The cells were subjected to 15 min insulin stimulation (100 nM) andthen 0.5 �Ci/ml of 2-deoxy-d-3[H] glucose was added for 20 min and theuptake was measured. The results were expressed as % change in glu-cose uptake with respect to the solvent control (DMSO). Dose-dependentuptake was observed with both the extracts in respect to solvent con-trol. Data are means ± S.E., n = 3. (*), p < 0.05 as compared with untreatedcontrol group.
(with the extraction yield of 1.53, 2.21 and 10.1% w/w,respectively). Methanolic extract of C. intybus (CME) wassubjected to detannification and the efficiency of detan-nification process was measured by tannin estimation.
Fig. 2. Influence of CME and CME/DT on the cytotoxicity of 3T3-L1preadipocytes. Cytotoxic effect of CME and CME/DT measured on 3T3-L1 cells during postconfluent mitotic clonal expansion. Postconfluent3T3-L1 cells were treated with all indicated doses of CME and CME/DT.After 24 h, LDH was measured in supernatants by using the formula %LDH release = (ODSample − ODcontrol)/(ODTritonX − ODcontrol) × 100. Cytotoxi-city was expressed as % LDH release. Data shown reflect the means ± S.E.of triplicates of two independent experiments.

Author's personal copy
V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78 73
Fig. 3. Effect of CME and CME/DT on lipid accumulation in 3T3-L1 adipocytes after differentiation induction. (A) Two-day postconfluent 3T3-L1 cellswere differentiated according to the protocol followed by treatment with described doses of CME and CME/DT or vehicle for 2 days. Eight days afterinduction of differentiation, cells were assayed for total triglyceride content using Adipored reagent. Preadipocytes were separately maintained by protocolas indicated. Data shown reflect the means ± S.E. of triplicates of two determinations. (*), p < 0.05 as compared with preadipocytes group. Photomicrographswere documented to evaluate the morphological changes of adipocytes at magnification ×100. (B) Control (C) differentiation-induced (D) differentiationmedium + CME (100 ng/ml) (E) differentiation medium + CME/DT (1 �g/ml).
The amount of tannins present in CME was found to be2.78% w/w and after detannification (in CME/DT) the valuewas come down to 0.31% w/w. The results clearly revealedthat the detannification process is more than 85% efficient.
3.2. Measurement of 2-deoxy-d-3[H] glucose
All sequential extracts of C. intybus were subjected toradiolabelled glucose uptake assay (Fig. 1a). CME showedmaximal glucose uptake and hence it was selected forthe detannification process. A dose-dependent increase
(Fig. 1b) in glucose uptake was observed with CME andCME/DT and concentrations of 100 ng/ml and 1 �g/ml wereobserved to be the optimal dose showing maximum glu-cose uptake activity, respectively.
3.3. Assessment of cell cytotoxicity
The cell cytotoxicity of CME and CME/DT was studied athigher doses ranging from 10 ng/ml to 10 �g/ml by meansof LDH release at 24 h. The result implies that the CME andCME/DT are non-toxic even at higher doses (Fig. 2).

Author's personal copy
74 V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78
3.4. Adipocyte differentiation assay
Two-day postconfluence, 3T3-L1 preadipocytes wereinduced for differentiation in the presence of logarith-mic doses of CME and CME/DT ranging from 1 pg/ml to10 �g/ml. Intracellular lipid content was used to mea-sure the degree of adipocyte differentiation. As illustratedin Fig. 3A, when CME is added to the preadipocytes inthe presence of differentiation medium, preadipocytesremained largely undifferentiated and the lipid content wasvery basal. Surprisingly, in CME/DT treated preadipocytesthe lipid content was significantly increased. Cytotoxic-ity assay by LDH release measurement revealed that thepreadipocytes treated with CME and CME/DT for the wholespectrum of assay duration were more than 90% viable(data not shown) and also the differences in morphologyof CME and CME/DT treated adipocytes were photomicro-graphed at magnification ×100 (Fig. 3B–E).
Fig. 4. (a) Glucose transport behavior of CME and CME/DT in the presenceof Genistein (GS) – IRTK inhibitor. (b) 3T3 adipocytes were pretreated withCME and CME/DT in addition of Wortmannin (WT) – PI3K inhibitor at theconcentration indicated and subjected to glucose uptake assay. Inhibitorssuppressed the glucose transport of CME but not CME/DT, which showeduptake similar to that of positive control Rosiglitazone. The results wereexpressed as % change in glucose uptake with respect to the solvent control(DMSO). Data are means ± S.E. of triplicates of two independent experi-ments. (*), p < 0.05 as compared with untreated control group.
Fig. 5. Effect of CME and CME/DT on inhibition of PTP1B enzyme. Theeffect of CME (100 ng/ml) and CME/DT (1 �g/ml) on PTP1B enzyme inhi-bition was observed. CME showed maximal inhibition of 92% similar topositive control sodium orthovanadate. CME/DT exhibited negligible inhi-bition of PTP1B enzyme. Data shown reflect the means ± S.E. of triplicatesof two independent experiments.
3.5. Measurement of 2-deoxy-d-3[H] glucose uptake inthe presence of PI3K and IRTK inhibitor
Our study reveals that the glucose uptake behavior ofCME, after treatment with 50 �M and 100 nM of GS andWT, respectively, in 3T3-L1 adipocytes, arrested the insulinmediated glucose uptake. But, the inhibitors did not controlthe glucose uptake activity of CME/DT (Fig. 4a and b).
3.6. PTP1B inhibition assay
As shown in Fig. 5, CME exhibits potent PTP1B inhibition(92%) similar to the positive control sodium orthovanadate(100 �M). But, CME/DT showed only basal level PTP1B inhi-bition.
3.7. Effect of CME and CME/DT on insulin signalingcascade
After demonstrating the effect of CME and CME/DT onPTP1B inhibition, the study was further extended to deter-mine the modulations of downstream markers involved inglucose uptake such as IR� and IRS1 at protein level bywestern blotting and GLUT4 and PI3K at transcript level byRT-PCR. The levels of GLUT4 and PI3K were expressed afternormalization with GAPDH (Fig. 7E) expression. Relativedensitometry scanning was performed for semiquantita-tive analysis (Figs. 6C, D, 7B and D).
Immunoprecipitation and western blot analysis (Fig. 6Aand B) revealed that the IR� and IRS1 which are impor-tant markers involved in the early stage of insulin signalingevent, were significantly phosphorylated after cells weretreated with CME on 3T3-L1 adipocytes using phosphospecific antibodies. Whereas CME/DT treated cells showedbasal level expression of IR� and IRS1. Consequently theexpression of PI3K (Fig. 7A) at 18 h post-treatment of 3T3-L1adipocytes with CME (100 ng/ml) was found to be sig-nificant. However, the adipocytes treated with CME/DTfailed to upregulate the PI3K expression, which is similarto that of Rosiglitazone – a negative control for the PI3Kmediated glucose uptake. In GLUT4 expression our find-

Author's personal copy
V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78 75
Fig. 6. Effect of CME and CME/DT on IR� and IRS1 expression. (A) 3T3-L1 adipocytes pretreated with CME for 24 h showed phosphorylation of IR� andIRS1 on par with positive control insulin (100 nM) (B) CME/DT failed to induce the phosphorylation. Bars represent the means ± S.E. (n = 3) of densitometricrepresentation of IR� and IRS1 expressions (C and D). (*), p < 0.05 as compared with untreated control group.
ing (Fig. 7C) shows that 3T3-L1 adipocytes treated withCME (100 ng/ml) and CME/DT (1 �g/ml) is upregulated andequivalent to positive control, Insulin (100 nM) and Rosigli-tazone (50 �M).
3.8. Effect of CME and CME/DT on key genes involved inadipocytes differentiation
The effect of CME (100 ng/ml) and CME/DT (1 �g/ml)extracts on C/EBP�, PPAR� and SREBP-1c expression in 3T3-L1 adipocytes is presented in Fig. 8A, C and E. CME/DTstimulated significant expression of all the three genesas compared to positive control Rosiglitazone (50 uM)whereas CME treated adipocytes showed basal expres-sion.
4. Discussion
In the present study, the glucose uptake and antiadi-pogenic activities of CME and CME/DT have been observedalong with confirming the duality (PTP1B inhibitor andPPAR� agonist) of C. intybus in exhibiting anti-diabeticactivity. The crude extracts of Aegles marmelos, Syzygiumcumini and Pterocarpus marsupium were reported for glu-cose uptake activity via PI3K and PPAR� on L6 myotubes[23,20], which supports our data for the duality observedin glucose uptake activity of C. intybus. Pushparaj et al.[7] described that the treatment with ethanolic extract ofC. intybus in streptozotocin-induced diabetic rats reducesthe blood glucose, triglycerides and cholesterol levels sig-nificantly. It is a clear evidence to justify our findings onadipogenesis inhibition.
Fig. 7. Effect of CME and CME/DT on PI3K and GLUT4 mRNA transcripts. 3T3-L1 adipocytes were incubated with Insulin (100 nM), Rosiglitazone (50 �M),CME (100 ng/ml) and CME/DT (1 �g/ml) for indicated time. Lane 1–6 indicates untreated control, Insulin, Rosiglitazone, CME, CME/DT treated cells and PCRnegative control, respectively. (A) The expression levels of PI3K transcripts (248 bp), compared with control cells. Insulin and CME showed the significantexpression whereas Rosiglitazone treated cells and CME/DT treated cells showed basal expression of PI3K. (C) Illustrates the elevated levels of GLUT4transcripts (318 bp) in all the lanes (6E). The mRNA of GAPDH (588 bp) in the same sample was used as a loading reference. Bars represent the means ± S.E.(n = 3) of densitometric representation of PI3K, and GLUT4 expressions (B and D). (*), p < 0.05 as compared with untreated control group.

Author's personal copy
76 V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78
Fig. 8. Effect of CME and CME/DT on genes involved in adipogenesis. CME inhibited adipocyte differentiation by altering the expression of key genesinvolved in adipogenesis. Maximal expression of C/EBP� (224 bp), PPAR� (155 bp) and SREBP1c (222 bp) were observed with Rosiglitazone and CME/DTtreated cells whereas insulin and CME failed to upregulate the expression on comparison with untreated cells (A, C and E). Bars represent the means ± S.E.(n = 3) of densitometric representation of gene expressions (B, D and F). (*), p < 0.05 as compared with untreated control group.
Liu et al. [10] have reported that the tannins are theimperative molecules in natural products especially fortreating metabolic syndromes like diabetes. In their study,they have concluded that the tannins of banaba aqueousextract (Lagerstroemia speciosa) are the compounds respon-sible for glucose transport and adipocyte differentiationinhibitory activities. However, in our finding, significantglucose uptake was observed even after removing tanninsfrom CME. Since the observation differed from the earlierreports, we hypothesized that the tannins are sparinglyextractable with methanol than water so that their quan-tity and role might be insignificant in the biological activityof CME.
We then studied the effects of CME and CME/DTon suppressing preadipocyte differentiation. Adipogen-esis is often depicted by a cascade of genetic eventsand are controlled by various factors. The factors whichinclude insulin-like growth factor I (IGF-1), macrophagecolony-stimulating factor, fatty acids, prostaglandins andglucocorticoids promote adipogenesis by activating acascade of transcription factors that coordinate the dif-ferentiation process [24]. To stimulate adipogenesis ofpreadipocyte cell lines, a combination of adipogenic induc-ers, including a glucocorticoid agonist, an agent to increaseintracellular cAMP and high concentrations of insulin tostimulate IGF-I, is generally used [25]. In adipocyte differ-entiation assay, CME possesses inhibitory activity but notCME/DT. This is an interesting observation, which coincideswith early reports on inhibitory role of tannins in adipocytedifferentiation. This led us to investigate the mechanismused by tannins of CME in glucose uptake and adipogenesisinhibition.
Inhibitors of insulin mediated glucose transport wereincorporated with CME and CME/DT to further substanti-ate the signaling pathway. In normal as well as in diabetesconditions, the protein tyrosine kinases play a crucial rolein numerous cellular signaling pathways. It is essentialto activate the tyrosine kinase by insulin, leading to thephosphorylation of its receptors thus inducing a functionalchange in the immediate signaling molecule such as PI3K[26]. Genistein (GS) – a reported insulin receptor tyro-sine kinase (IRTK) inhibitor controls the insulin stimulatedglucose uptake [27], and Wortmannin (WT) – an identi-fied inhibitor of PI3K inhibiting the insulin-stimulated PI3Kactivity and GLUT4 translocation [28] were selected andused for the assay. The finding from the inhibitor studysuggested that the mechanism of CME is through insulinmediated PI3K dependent manner whereas the mechanismof CME/DT is through PI3K independent manner.
Current researchers have demonstrated the possibleassociation of PTP1B in insulin sensitivity and obesity [29].Insulin mediated glucose uptake is negatively regulated byprotein tyrosine phosphatases (PTPs). Among several PTPs,PTP1B has been implicated to be a key modulator of insulinsignal transduction by acting at downstream signalingcomponents, such as IRS1 and PI3K [30]. Over expressionof PTP1B protein have been reported in insulin-resistantconditions which are associated with obesity [31]. Since,PTP1B plays a pivotal role in insulin resistance and adi-pogenesis, the findings of PTP1B inhibition assay reasonsthe glucose uptake and adipogenesis inhibitory activitiesof tannins present in CME.
Insulin binding to its transmembrane insulin receptor(IR) activates IRTK, which then causes phosphorylation

Author's personal copy
V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78 77
of family of insulin receptor substrate (IRS) proteins onselective tyrosine residues [32]. From the insulin-signalingcascade, it is apparent that the activation of PI3K is nec-essary and in some cases sufficient to understand many ofinsulin’s effects on glucose and lipid metabolism. In adi-pose tissue, insulin exerts its normal cellular effects viaPI3K by first binding to its receptor followed by the activa-tion of IRS [33]. Finally, insulin mediated glucose uptake inmuscle and adipose tissue takes place through the translo-cation of an integral membrane, glucose transporter protein– GLUT4, from an intracellular compartment to the cell sur-face [34]. The IR�, IRS1, PI3K and GLUT4 expression resultsconfirm that the anti-diabetic activity of CME and CME/DTis through GLUT4 endocytosis but via bi-functional mech-anism.
Peroxisome proliferator activator receptor (PPAR),CCAAT/enhancer binding protein (C/EBP), and sterol reg-ulatory element binding protein (SREBP) families are thewell-documented primary adipogenic transcription fac-tors involved in adipocyte differentiation and among themPPAR� is most extensively studied for its therapeutic utilityin the treatment of NIDDM [35]. CME controls the expres-sion of all the three adipogenic markers and similar resultswere reported for dietary flavonoids by Chien et al. [19].The unique expression of SREBP-1c on adipocytes treatedwith insulin is reasoned by the fact that lipogenesis actionof insulin is through SREBP. Cristina et al. [36] showed adecrease in adiposity with a down regulation of PPAR�,SREBP1 and other adipogenic genes in animals treated withPTP1B antisense oligonucleotide, suggesting that PTP1Breduction might have a primary role in inhibiting adipo-genesis and the development of obesity in these animals.Our results on PTP1B inhibition and mRNA expression ofadipogenic genes reason the glucose uptake activity andadipocyte differentiation activity of CME. At the same time,the expression of adipogenic markers increased signifi-cantly after detannification (in CME/DT). The activation ofPPAR� by CME/DT explains its glucose uptake activity andGLUT4 mRNA expression and enlightens the significance oftannins in the biological activity of CME.
The two activities, glucose uptake and adipogenesisinhibition, were mediated by two independent factorsinsulin receptor (IR) and IGF1 but controlled by the onlylinkage PTP1B enzyme. Therefore, the inhibition of PTP1Benzyme activity by the tannins present in CME rationalizesits bi-functional activity. In theory, CME should not exhibitadipogenesis inhibition due to the presence of PPAR� acti-vator (from non-tannin fraction) because blends of twoindependently functioning molecules will behave with theadditive mechanism of two molecules. But, our finding dif-fers from the hypothesis. This brings us to a new propositionthat the interaction of tannins present in CME on PTP1Binhibition may be very potent than the interaction of non-tannins present in CME/DT with PPAR� agonism.
5. Conclusion
Tannins are freely available in our daily diet and tillnow the usefulness of tannins are limited to only neu-traceutical level as a biological antioxidant. To the best ofour knowledge, it is for the very first time that we are
reporting the therapeutical significance of C. intybus andits tannins. The ability of existing therapy to target var-ious aspects of the insulin resistance syndrome inducesother metabolic abnormalities chiefly in lipids. Therefore,glucose-lowering drugs deficient of adipogenic activity arethe need for the current situation. The tannins of C. intybusmethanolic extract seem to have such a beneficial combi-nation. If C. intybus is identified for the presence of PTP1Binhibitor and PPAR� activator at the molecular level, thenit will also be interesting to find the possibility of par-tial PPAR� agonism/antagonism in future. The propositionwould become apparent once the CME and CME/DT arecompletely evaluated. Pharmaceutical significance of thepresent analysis depends on the lead molecule identifica-tion and in vivo animal studies, which are necessary forfurther exploration.
Acknowledgements
Authors would like to thank Dr. R.B. Narayanan, Director,Centre for Biotechnology, Anna University, Chennai, India.The valuable suggestions of Dr. M. Ramanathan and Mr.C. Saravanababu also acknowledged. Muthusamy.V.S is aCSIR-Research Associate and is highly grateful to Councilof Scientific and Industrial Research, Govt. of India for thefinancial assistance.
References
[1] Michael W. Rajala, Philipp E. Scherer, Minireview: the adipocyte-atthe crossroads of energy homeostasis, inflammation, and atheroscle-rosis, Endocrinology 44 (9) (2003) 3765–3773.
[2] B. Matthias, W. Leanne, L.G. John, S.C. Laustsen, C. Ronald Kahn,Role of insulin action and cell size on protein expression patternsin adipocytes, J. Biol. Chem. 279 (30) (2004) 31902–31909.
[3] S.E. Inzucchi, Oral antihyperglycemic therapy for type 2 diabetes:scientific review, JAMA 287 (3) (2002) 360–372.
[4] E.K. Frank, T.C. Guy, The evolving role of natural products in drugdiscovery, Nature 4 (2005) 206–221.
[5] N. Rikke, N. Karina, K. Tadao, Anthocyanins from flowers of Cichoriumintybus, Phytochemistry 60 (2002) 357–359.
[6] B.S. Rees, J.B. Harborne, The role of sesquiterpene lactones and phe-nolics in the chemical defense of the chicory plant, Phytochemistry24 (1985) 2225–2231.
[7] P.N. Pushparaj, H.K. Low, J. Manikandan, B.K.H. Tan, C.H. Tan,Anti-diabetic effects of Cichorium intybus in streptozotocin-induceddiabetic rats, J. Ethnopharmacol. 111 (2) (2007) 430–434.
[8] Sangkil Nam, David M. Smith, Q. Ping Dou, Tannic acid potentlyinhibits tumor cell proteasome activity, increases p27 and Baxexpression, and induces G1 arrest and apoptosis, Cancer Epidemiol.Biomarkers Prev. 10 (2001) 1083–1088.
[9] Ashok D. Patil, Alan J. Freyer, Drake S. Eggleston, R. Curtis Halti-wanger, Brian Tomcowicz, Ann Breen, Randall K. Johnson, Daleformis,a new phytoalexin from the roots of Dalea filiciformis: an inhibitorof endothelin converting enzyme, J. Nat. Prod. 60 (3) (1997) 306–308.
[10] F. Liu, J. Kim, Y. Li, X. Liu, J. Li, X. Chen, An extract of Lagerstroemiaspeciosa L. has insulin-like glucose uptake–stimulatory and adipocytedifferentiation–inhibitory activities in 3T3-L1 cells, J. Nutr. 131 (2001)2242–2247.
[11] K.M. Riedl, A.E. Hagerman, Tannin-protein complexes as radical scav-engers and radical sinks, J. Agric. Food Chem. 49 (2001) 4917–4923.
[12] S. McDonald, P.D. Prenzler, M. Autolovich, K. Robards, Phenolic con-tent and antioxidant activity of olive extracts, Food Chem. 73 (2001)73–84.
[13] Xueqing Liu, Jae-kyung Kim, Yunsheng Li, Jing Li, Fang Liu, XiaozhuoChen, Tannic acid stimulates glucose transport and inhibits adipocytedifferentiation in 3T3-L1 cells, J. Nutr. 135 (2) (2005) 165–171.
[14] Anne W. Harmon, Yashomati M. Patel, Naringenin inhibits phospho-inositide 3-kinase activity and glucose uptake in 3T3-L1 adipocytes,Biochem. Biophys. Res. Commun. 305 (2003) 229–234.

Author's personal copy
78 V.S. Muthusamy et al. / Chemico-Biological Interactions 174 (2008) 69–78
[15] T. Akiyama, J. Ishida, S. Nakagawa, H. Ogawara, S. Watanabe, N. Itoh, M.Shibuya, Y. Fukami, Genistein, a specific inhibitor of tyrosine-specificprotein kinases, J. Biol. Chem. 262 (1987) 5592–5595.
[16] Suresh Ambati, Hye-Kyeong Kim, Jeong-Yeh Yang, Ji Lin, Mary AnneDella-Fera, Clifton A. Baile, Effects of leptin on apoptosis and adi-pogenesis in 3T3-L1 adipocytes, Biochem. Pharmacol. 73 (3) (2007)374–384.
[17] T.R. Burke, X. Ye, S. Yan, Z. Wang, L. Jia, Z. Chen, D. Zhang, Bar-ford, Small molecule interactions with protein-tyrosine phosphatasePTP1B and their use in inhibitor design, Biochemistry 35 (1996)15989–15996.
[18] L.R. Hall, R.K. Mehlotra, A.W. Higgins, M.A. Haxhiu, E. Pearlman, Anessential role for interleukin-5 and eosinophils in helminth inducedairway hyper-responsiveness, Infect. Immun. 66 (1998) 4425–4430.
[19] Po-Jung Chien, Ying-Chen Chen, Shou-Chin Lu, Fuu Sheu, Dietaryflavonoids suppress adipogenesis in 3T3-L1 preadipocytes, J. FoodDrug Anal. 13 (2) (2005) 168–175.
[20] R. Anandharajan, S. Jaiganesh, N.P. Shankernarayanan, R.A.Viswakarma, A. Balakrishnan, In vitro glucose uptake activityof Aegles marmelos and Syzygium cumini by activation of Glut-4PI3kinase and PPARgamma in L6 myotubes, Phytomedicine 13 (2006)434–441.
[21] A. Klip, A. Guma, T. Ramlal, P.J. Bilan, L. Lam, L.A. Leiter, Stimula-tion of hexose transport by metformin in L6 muscle cells in culture,Endocrinology 130 (1992) 2535–2544.
[22] V. Senthil, S. Ramadevi, V. Venkatakrishnan, P. Giridharan, B.S.Lakshmi, R.A. Vishwakarma, A. Balakrishnan, Withanolide inducesapoptosis in HL-60 leukemia cells via mitochondria mediatedcytochrome c release and caspase activation, Chem. Biol. Interact.167 (2007) 19–30.
[23] R. Anandharajan, K. Pathmanathan, N.P. Shankernarayanan, RamA. Vishwakarma, Arun Balakrishnan, Upregulation of Glut-4 andPPAR gamma by an isoflavone from Pterocarpus marsupium on L6myotubes: a possible mechanism of action, J. Ethnopharmacol. 97(2005) 253–260.
[24] A. Ormond, MacDougald, Susanne Mandrup, Adipogenesis: forcesthat tip the scales, Trends Endocrinol. Metab. 13 (1) (2002) 5–11.
[25] Y.M. Patel, M.D. Lane, Mitotic clonal expansion during preadipocytedifferentiation Calpain-mediated turnover of p27, J. Biol. Chem. 275(2000) 17653–17660.
[26] F. Kanai, K. Ito, M. Todaka, H. Hayashi, S. Kamohara, K. Ishii, Insulin-stimulated GLUT4 translocation is relevant to the phosphorylation ofIRS-1 and the activity of PI3-kinase, Biochem. Biophys. Res. Commun.195 (1993) 762–768.
[27] A.W. Harmon, Y.M. Patel, J.B. Harp, Genistein inhibitsCCAAT/enhancer-binding protein beta (C/EBP�) activity and3T3-L1 adipogenesis by increasing C/EBP homologous proteinexpression, Biochem. J. 367 (2002) 203–208.
[28] J.F. Clarke, P. Young, K. Yonezawa, M. Kasuga, G.D. Holman, Inhibi-tion of the translocation of GLUT1 and GLUT4 in 3T3-L1 cells bythe phosphatidylinositol 3-kinase inhibitor, Wortmannin, Biochem.J. 300 (1994) 631–635.
[29] O. Ukkola, M. Santaniemi, Protein tyrosine phosphatase 1B: a newtarget for the treatment of obesity and associated co-morbidities, J.Intern. Med. 251 (2002) 467–547.
[30] Rebecca J. Gum, Lori L. Gaede, Sandra L. Koterski, Matthew Heindel,Jill E. Clampit, Bradley A. Zinker, James M. Trevillyan, Roger G. Ulrich,Michael R. Jirousek, Cristina M. Rondinone, Reduction of protein tyro-sine phosphatase 1B increases insulin-dependent signaling in ob/obmice, Diabetes 52 (2003) 21–28.
[31] M.R. Calera, G. Vallega, P.F. Pilch, Dynamics of protein-tyrosine phos-phatases in rat adipocytes, J. Biol. Chem. 275 (2000) 6308–6312.
[32] Yehiel Zick, Insulin resistance: a phosphorylation-based uncouplingof insulin signaling, Trends Cell Biol. 11 (11) (2001) 437–441.
[33] Peter R. Shepherd, Dominic J. Withers, Kenneth Siddle, Phospho-inositide 3-kinase: the key switch mechanism in insulin signaling,Biochem. J. 333 (1998) 471–490.
[34] Geoffrey D. Holman, Ignacio V. Sandoval, Moving the insulin-regulated glucose transporter GLUT4 into and out of storage, TrendsCell Biol. 11 (4) (2001) 173–179.
[35] J.A. Bonini, J.R. Colca, C. Dailey, M. White, C. Hofmann, Compensatoryalterations for insulin signal transduction and glucose transport ininsulin-resistant diabetes, Am. J. Physiol. Endocrinol. Metab. 269(1995) E759–E765.
[36] Cristina M. Rondinone, James M. Trevillyan, Jill Clampit, RebeccaJ. Gum, Cathy Berg, Paul Kroeger, Leigh Frost, Bradley A. Zinker,Regina Reilly, Roger Ulrich, Madeline Butler, Brett P. Monia, Michael R.Jirousek, Jeffrey F. Waring, Protein tyrosine phosphatase 1B reductionregulates adiposity and expression of genes involved in lipogenesis,Diabetes 51 (2002) 2405–2411.














