
TAFONOMÍA DE SITIOS ARQUEOLÓGICOS, UN CASO DE ESTUDIO: MAQUIJATA, DEPARTAMENTO CHOYA, SANTIAGO DEL...
26
De la Puna a las Sierras Avances y Perspectivas en Zooarqueología Andina Editado por Andrés D. Izeta Guillermo L. Mengoni Goñalons BAR International Series 2564 2013 South American Archaeology Series No 19 Edited by Andrés D. Izeta
Transcript of TAFONOMÍA DE SITIOS ARQUEOLÓGICOS, UN CASO DE ESTUDIO: MAQUIJATA, DEPARTAMENTO CHOYA, SANTIAGO DEL...

De la Puna a las Sierras
Avances y Perspectivas en Zooarqueología Andina
Editado por
Andrés D. IzetaGuillermo L. Mengoni Goñalons
BAR International Series 2564
2013
South American Archaeology Series No 19Edited by Andrés D. Izeta

Published by
Archaeopress
Gordon House
Oxford OX2 7ED
www.archaeopress.com
©
Oxford
OX2 7BP
www.hadrianbooks.co.uk
from Hadrian Books or may be downloaded from www.archaeopress.com

1
INDICE
INDICE…………………………………………………………………………………….................1
EVALUADORES DEL VOLUMEN……………………………………………………….................3
INTRODUCCIÓN...…………………………………………………………………………………..5
1- ARQUEOLOGIA DE LOS ANIMALES EN CONTEXTOS DEL PERIODO FORMATIVO DEL VALLE DEL CAJON (CATAMARCA, ARGENTINA)
Izeta, Andrés D…………………………………………………………………………….................9
2- TAFONOMÍA DE SITIOS ARQUEOLÓGICOS, UN CASO DE ESTUDIO: MAQUIJATA, DEPARTAMENTO CHOYA, SANTIAGO DEL ESTERO
Luis M. del Papa, José Togo y Luciano J. M. De Santis…………………………………….............17
3- LESIONES EN EXTREMIDADES INFERIORES DE CAMÉLIDOS: UNA COMPARACIÓN ENTRE LOS CONJUNTOS DEL ARCAICO TARDÍO Y FORMATIVO TEMPRANO (VERTIENTE OCCIDENTAL DE LA PUNA DE ATACAMA)
Isabel Cartajena, Oscar López, Lautaro Núñez y Cristián Linares………………………….............39
4- EL VELLÓN DORADO. APROPIACIÓN SOCIAL DE VICUÑAS EN UN OASIS DE LA PUNA DE ATACAMA (CATAMARCA, ARGENTINA)
Claudio M. Revuelta…………………………………………………………………………............53
5- REGISTRO FAUNÍSTICO Y DIFERENCIACIÓN SOCIAL: EL CASO DE PIEDRAS BLANCAS, VALLE DE AMBATO, CATAMARCA (SIGLOS VI- XI d.C.)
Mariana Dantas………………………………………………………………………………............67
6- ¿ANFISBENAS PARA LOS DIOSES?: ZOOARQUEOLOGÍA DE UNA OFRENDA ANIMAL
Débora M. Kligmann, Adriana M. Albino y Elena Díaz País ………………………………............89

2
7 -HISTORIA OCUPACIONAL Y PRÁCTICAS SOCIALES: UN ANÁLISIS ARQUEOFAUNÍSTICO CONTEXTUAL. SITIO PIEDRA HORADADA 2, ANTOFAGASTA DE LA SIERRA, CATAMARCA, ARGENTINA
Silvana V. Urquiza, Andrés S. Romano y Sara M. L. López Campeny……………………............121
8- PASTORALISMO Y SUBISTENCIA DURANTE EL PERÍODO FORMATIVO EN EL ALTIPLANO CENTRAL DE BOLIVIA: EVIDENCIA ZOOARQUEOLÓGICA DE IRUCICA, BOLIVIA
José M. Capriles Flores y Juan Albarracin-Jordan …………………………………………...........145

17
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
2
PRIMERA APROXIMACIÓN A LA TAFONOMÍA DE LA REGIÓN CHACO-SANTIAGUEÑA. SITIO MAQUIJATA, SANTIAGO DEL ESTERO
Luis M. del Papa*; José Togo** y Luciano J. M. De Santis***
* CONICET, Facultad de Ciencias Naturales y Museo (UNLP), calle 64 y 120 s/n, La Plata, Argentina. E-mail: [email protected]** Facultad de Humanidades, Ciencias Sociales y de la Salud (UNSE), Avenida Belgrano (s) Nº 2180, Santiago del Estero, Argentina. E-mail: [email protected]*** Facultad de Ciencias Naturales y Museo (UNLP), calle 64 y 120 s/n, La Plata, Argentina. E-mail: [email protected]
De la Puna a las Sierras: Avances y Perspectivas en Zooarqueología Andina. Andres D. Izeta y Guillermo L. Mengoni Goñalons (Ed.) British Archaeological Reports International Series. SAMAR 19. 2012: 17-38
RESUMEN
El objetivo de este trabajo consiste en proporcionar vías de análisis referentes a la tafonomía, las
cuales no fueron desarrolladas previamente en el área de estudio. En este caso se analiza el sitio ar-
queológico Maquijata, Departamento Choya, ubicado en la falda oriental de la sierra de Guasayán
y en la boca de la quebrada homónima. En el lugar encontramos una serie de montículos, muchos
de ellos degradados o reutilizados por las poblaciones sucesivas que se asentaron en este paraje. Se
-
-
dos se evidencia la acumulación de restos por diferentes agentes y procesos, con buena preservación
de los materiales y relativamente poca distorsión del registro. Estos resultados podrán ser tenidos en
cuenta para futuro análisis de los sitios arqueológicos del área, en un marco regional.
ABSTRACT
The aim of this study is to provide some taphonomy analytic approaches that were not developed be-
fore at the study area. We analyzed the Maquijata archaeological site, Choya Department, located on
the eastern slope of the Guasayán mountain range at the entrance of the ravine of the same name. At
the site, we found several mounds; many of them were degraded or reutilized by the successive popu-
place due to the action of different agents and processes. We also observed a good preservation of the
future analysis of other archaeological sites belonging to this area, at a regional scale.
En este trabajo se llevó a cabo un análisis tafonómico para el sitio arqueológico Maquijata, Departamento Choya, ubicado en la falda oriental de la sierra de Guasayán y en la boca de la quebrada homónima. El sitio se encuentra en la región arqueológica Chaco-Santiagueña (González 1979) y corresponde a la actual provincia de Santiago del Estero (Argentina), para la cual no existen antecedentes referentes a esta temática.
Con este análisis pretendemos identificar los procesos de incorporación de los restos, los agentes acumuladores y los posibles agentes
la matriz del sitio. El estudio apunta a reducir la ambigüedad generada por las acciones naturales
del comportamiento humano y establecer tendencias respecto a la integridad que presenta la

18
Luis M. del Papa et al.
estructura arqueológica de una región, las cuales se podrán tener en cuenta para futuros análisis de sitios arqueológicos del área.
La localidad de Maquijata fue vinculada por algunos estudiosos con la entrada de la expedición de Diego de Rojas en el año 1543 a suelo santiagueño (Ledesma 1961). En dicho lugar existen una serie de montículos, muchos de ellos degradados o reutilizados por las poblaciones sucesivas que se asentaron en dicho paraje, estos montículos tienen dimensiones y altura variables, conformando un conglomerado relativamente
presencia de una población numerosa y estable, con patrones culturales estabilizados (Togo 2004).A lo largo del desarrollo de las investigaciones arqueológicas en la región se observa que el análisis de los materiales arqueofaunísticos ha tenido menor interés por parte de los investigadores en relación a otros (e.g. alfarería). La mayoría de los estudios estuvieron orientados hacia la dilucidación de la problemática de la etapa agroalfarera comprendida por los grupos portadores, principalmente, de la cerámica “Las Mercedes”, “Sunchituyoj” y “Averías”, ubicadas cronológicamente entre el 350/400 DC y la conquista española (Togo 2004, 2007).
En las décadas de 1930 y 1940, los restos arqueofaunísticos fueron estudiados con el objetivo de discutir la antigüedad de la llamada “Civilización Chaco-Santiagueña” propuesta por los hermanos Wagner, a partir de sus trabajos y de especialistas paleontólogos (Bordas 1940; Kraglievich y Rusconi 1931; Parodi Bustos 1947; Rusconi 1930, 1934; Wagner y Wagner 1936). El análisis de los materiales faunísticos estaba sujeto al método de recuperación de los mismos, ya que solo se recuperaban los que estuviesen enteros o aquellos que pudieran ser determinados por lo menos a nivel de género.
humano comienza en la década de 1970 con los trabajos de Lorandi y colaboradores (Cione et
al. 1979; Lorandi y Carrió 1970-1975; Lorandi y Lovera 1972), a partir del advenimiento de la “Nueva Arqueología” como corriente teórica en la Argentina. En dichos trabajos, que se desarrollan principalmente en la zona de la cuenca del río
tuvieron los pobladores a la hora de la obtención de los recursos, por su variedad y actividades de producción asociadas. El trabajo de Cione et al.
(1979) constituye uno de los primeros trabajos arqueofaunísticos que desarrollan la temática de la economía que tuvieron los pueblos que se asentaron en la región. Se observa una diferencia en los métodos de recuperación de los materiales, realizados por excavaciones sistemáticas con control estratigráfico, donde se evidencia un aumento significativo de restos de peces y roedores pequeños.
En la última década el interés en el estudio de los restos arqueofaunísticos de la región ha tomado mayor relevancia (del Papa 2006; Fernández Varela et al. 2001; Togo 2004), donde la recuperación de los materiales se realizó mediante excavaciones sistemáticas y el uso de mallas para el cribado de los sedimentos obteniendo así los especímenes de pequeño tamaño, además se observa el
(e.g Mammalia, Artiodactyla).
La perspectiva llevada a cabo en la presente contribución no fue desarrollada de manera sistemática en los trabajos previos para la zona de estudio, y de allí su relevancia.
Ambiente
Maquijata se encuentra ubicada a 80 km de la ciudad de Santiago del Estero, hacia el oeste, en la Sierra de Guasayán (Figura1), siendo aproximadamente la misma distancia hacia el río más cercano (río Dulce). Esta serranía se orienta N-S frente al “codo del río Dulce”, hasta Villa La Punta, con una extensión de 76 km de largo y un ancho que no sobrepasa los 4 km. La vertiente oriental se levanta en un frontón abrupto de barrera, con vegetación espesa; en cambio, por el poniente su declive no es tan pronunciado. La Sierra de Guasayán raramente excede los 300 m de altura sobre el nivel del mar.
En esta serranía y sobre todo en la parte oriental,
trecho en trecho, así como las vertientes (una de las más importantes sería la de Maquijata). Esta condición posibilitó la presencia de asentamientos humanos en la zona (Ledesma 1961).

19
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
En cuanto a la fauna, su distribución estaría encuadrada en la Subregión Andino-Patagónica, Dominio Central o Subandino, Distrito Subandino (Ringuelet 1961). La fauna es fundamentalmente
seleccionada en relación con condiciones
patagónica al oeste-sur (e.g. Lama guanicoe, Dolichotis salinicola, Dolichotis patagonum, Microcavia australis, Rhea americana).
Fitogeográficamente pertenece al Dominio Chaqueño, Distrito Occidental. Éste es un bioma en mosaico caracterizado por la presencia dominante de una cobertura arbórea, salpicado o acompañado de pastizales y otras hierbas no graminoideas (Cabrera 1971). Según Morello (1958) se pueden agrupar las unidades de vegetación y ambiente en subregiones, así podemos incluir al sitio estudiado dentro de la subregión del Chaco Serrano caracterizada por bosques secos salpicados de pastizales serranos que se desarrollan en suelos esqueléticos medianamente desarrollados in situ y con menor humedad local.
Son característicos de esta subregión los suelos Entisoles, estos son azonales muy jóvenes, de origen poligénico y débil o escaso desarrollo de
con materiales acarreados por gravedad. Son suelos livianos, de granulometría media a pequeña, casi sin estructura y textura franco arenosa y areno limosa. Son suelos potencialmente productivos (Dirección General de Minería y Geología 1981).
MATERIALES Y METODOS
Se analizaron los restos arqueofaunísticos obtenidos mediante excavaciones sistemáticas por niveles artificiales de 10 cm, en cuatro
una, ubicadas en los sectores identificados como I, II y III. En el sector I del sitio se ha realizado previamente la prospección con calicata eléctrica¹ denotando probables concentraciones
excavación (Togo 2004).
En el sitio se recuperaron materiales cerámicos, donde se observa la presencia de elementos
Sunchitúyoj, en sus niveles más bajos acompañados con algunos fragmentos asimilables a Las Mercedes. En los niveles superiores también se
y Averías (Páez et al. 2009). Entre los restos se recuperaron carbones, material lítico (puntas y desechos) y huesos de aves, peces, mamíferos, etc. El fechado radiocarbónico más antiguo del sitio es de 840 ± 70 años AP y el más reciente de 210 ± 60 años AP. (Togo 2007:231).
En los primeros 20 cm la tierra presenta una coloración más clara y restos de material orgánico. Por debajo de los 20 cm el sedimento posee una coloración más rojiza. Estas diferencias se deben a que la primera capa contiene la mayor concentración de material orgánico y se considera a todo el paquete sedimentario que contiene los restos arqueológicos como una misma unidad de depositación.
En el sector I, los materiales arqueológicos se encuentran hasta los 50 cm y la mayoría de los restos cerámicos están concentrados en los primeros 20 cm (85,02 % de los materiales de la cuadrícula); a partir de allí decrece la cantidad en forma pronunciada hasta el nivel de base (9,02 % para el nivel 3 y 5,94% de los materiales de la cuadrícula para el nivel 4) (Páez et al. 2009). Se
30 cm). En este sector se obtuvieron fechados radiocarbónicos del nivel 2 (10-20 cm) de 410 ± 60 años A.P. (LP-1714; carbón vegetal) y del nivel 4 (30-40 cm) de 540 ± 60 años A.P. (LP-1707; carbón vegetal) (Togo 2007:231).
Figura 1. Ubicación del sitio.

20
Luis M. del Papa et al.
El sector III se ha excavado hasta una profundidad de 70 a 80 cm, pero los últimos niveles fueron estériles. A los 15 cm se ha encontrado una capa compacta de posible material orgánico; por encima y por debajo de la misma se recolectó abundante carbón. En los dos primeros niveles se encuentran presentes el mayor número de fragmentos cerámicos (32 % en el nivel 1, 20 % en el nivel 2), luego comienza a descender hasta el cuarto nivel (15 % en el nivel 3 y 12 % en el nivel 4), aumenta nuevamente en el quinto (18%)
% en el nivel 6) (Páez et al. 2009). De este sector se obtuvo un fechado radiocarbónico de 210 ± 60 años A.P. (LP-1732; Carbón vegetal), el mismo se considera que pudo estar contaminado (Togo 2007:231).
A pesar de las diferencias en cuanto a la cantidad
la proporción entre los grupos cerámicos se mantiene constante, materiales pertenecientes a las entidades culturales Sunchitúyoj y Famabalasto principalmente (Páez et al. 2009). El análisis se desarrollará teniendo en cuenta los sectores como unidad, dado la relación contextual de estas dos entidades.
U n a v e z r e c u p e r a d o s l o s m a t e r i a l e s arqueofaunísticos se llevó a cabo la preparación de la muestra y su análisis correspondiente siguiendo los lineamientos generales propuestos por Miotti (1998).
Se han analizado 3356 restos (incluyendo fragmentos óseos, dientes, cáscaras de huevos y fragmentos de valvas de moluscos) provenientes de dos cuadrículas: la primera correspondiente al Sector I (Mj I), cuadrícula 1, de la que se obtuvo material faunístico en los niveles 1, 2 y 4; y la segunda al Sector III (Mj III), cuadrícula 2, con restos desde el primer nivel hasta el sexto, pero en menor cantidad en los tres últimos niveles.
En cuanto a la determinación taxonómica se ha llegado a distintos niveles jerárquicos, debido al grado de fragmentación de los especímenes óseos, es decir, una representación diferencial de rasgos anatómicos utilizados como diagnósticos para su
ya que no se realizó la determinación sistemática exhaustiva para los mismos.
cálculos referentes a la abundancia taxonómica de la muestra en forma de NISP y MNI, este último según criterios de lateralidad y edad (Mengoni Goñalons 1988, 1999).
En cuanto a la representación de partes se realizaron los cálculos referentes al MNE siguiendo la metodología propuesta por Morlan (1994) y
zonas diagnósticas; para el caso de los huesos largos de camélidos y réidos, las unidades
entre el MAU% (Binford 1984; Mengoni Goñalons 1999) con los valores tomados para la densidad mineral ósea (ver párrafo siguiente). Posteriormente se realizó el recuento como MNEt (MNE total) consignando así los elementos
1999), utilizado para la razón NISP/MNEt.
Se analizó lo concerniente a la conservación de los materiales, teniendo en cuenta el grado de meteorización de los mismos para mamíferos mayores a los 5 kg siguiendo los estadios propuestos por Behrensmeyer (1978); se calculó el índice de fragmentación NISP/MNEt (Lyman 1994) para cada aquellos que pudieron estar sujetos a procesos de mayor fragmentación. La diferencia de la densidad mineral ósea en los diferentes elementos del esqueleto puede favorecer la destrucción de algunos de esos elementos o parte de ellos y la supervivencia de otros (Elkin 1995; Lyman 1984, 1994). Por ello se correlacionó a través
de partes de la muestra (expresada en MAU %) y la densidad mineral ósea (DO) propuestas para los representados (camélidos, Elkin 1995) o de una estrecha relación taxonómica (para Rhea
americana se usaron los valores de Pterocnemia
pennata, Cruz y Elkin 2003). En la correlación realizada para R. americana, el valor de DO de las vértebras fueron consignadas por el promedio de elementos cervicales y torácicos.
como culturales, con el fin de identificar el agente o actor que las produjo (Gifford-Gonzalez 1991) y medir el impacto que tuvieron sobre la

21
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
muestra. Las mismas fueron analizadas a nivel macroscópico y con lupa de mano de quince aumentos, sólo en escasos restos fue necesario el uso de lupa binocular por la ambigüedad de las marcas.
se han tenido en cuenta aquellas producidas por carnívoros (e.g. Binford 1981; Blumenschine et
al. 1996; Montalvo et al. 2007), aves Strigiformes (para vertebrados < a 1 kg, Andrews 1990; Fernández Jalvo y Andrews 1992), las producidas por roedores durante el roído de los huesos (Quintana 2007), y las ocasionadas por la acción de raíces (Lyman 1994). Por otra parte se analizaron aquellas marcas de origen antrópico como las de corte, machacado, raspado, marcas de percusión, etc. (Binford 1981; Blumenschine et al. 1996; Mengoni Goñalons 1988, 1999).
Las fracturas se producen por el resultado de la interacción de las propiedades biomecánicas del hueso y los aspectos tecnológicos y naturales asociados a su producción (Johnson 1985). Se realizó el cálculo del porcentaje de huesos fracturados por la relación especímenes fracturados / especímenes de huesos largos, descartando las categorías taxonómicas de tamaño muy pequeño y aquellos huesos largos de los que es imposible extraer médula ósea en estado fresco (e.g. aves pequeñas, Tupinambis rufescens, Caviinae; ulna de roedores, peroné de Rhea americana). Las variables que fueron consideradas en la
material óseo (fresco, seco y estadios intermedios) y su morfología. A través del índice de fractura fresca (FFI) de Outram (2002) expresado de
estado fresco (valores 0, 1 y 2 del FFI), fracturas en estado intermedio, donde se produjo una cierta deshidratación del hueso o perdida de elasticidad previa a la fractura (valores 3 y 4) y fracturas en estado seco, donde el hueso perdió la mayoría o toda su elasticidad debido principalmente a factores diagenéticos o por procesos extremos (valores 5 y 6). Las fracturas se clasificaron morfológicamente en espiral, transversal y longitudinal.
asociados a las fracturas (marcas de percusión, negativos de lascado, muescas) (Blumenschine y Selvaggio 1988; Bunn 1981, 1989; Capaldo
y Blumenschine 1994; Mengoni Goñalons 1999; Pickering y Egeland 2006). El estudio de las fracturas cobra importancia por las inferencias que podemos obtener sobre el consumo, principalmente de médula ósea o grasa ósea y las probables prácticas culinarias que intervinieron en su producción (De Nigris 2004; Gifford-Gonzalez 1993; Oliver 1993).
También ha sido importante distinguir en los conjuntos arqueofaunísticos aquellos huesos que estuvieron sometidos a la acción térmica (Cain 2005; Lanata 1988; Nicholson 1993; Shipman et al. 1984; Stiner et al. 1995). Es de destacar que este tipo de atributos no suele representar el producto total de la cocción de los alimentos sino también el descarte por cuestiones de higiene, para impedir la acción de carroñeros o como combustible (Cain 2005; Costamagno et al. 2005; De Nigris 2004; Gifford-Gonzalez 1989; Kent 1993). Incluso pueden ser el producto de fuegos naturales (Cain 2005).
Para este análisis se desechó la posibilidad del uso de los huesos como combustible, debido a la gran cantidad de madera de excelente calidad que se encontraría en las inmediaciones (quebracho, algarrobo, etc.). Los huesos quemados por fuegos naturales son alterados sólo en una parte, produciendo un quemado superficial que no llega a generar huesos de color gris o blanco (Cain 2005). La cocción de los alimentos podría estar evidenciada por la existencia de restos que presentan atributos relacionados con la
lo que sugiere que éstos pudieron ser asados con tejidos que protegen el resto del elemento (Gifford-Gonzalez 1989; Kent 1993). El descarte en lugares de combustión se podría inferir por una mayor proporción de restos, donde el quemado
y el grado de alteración es más alto por estar sometido de forma directa a grandes temperaturas. De esta actividad se espera una mayor proporción de fragmentos calcinados (De Nigris 2004; Kent 1993; Oliver 1993).
Para poder distinguir entre estas acciones se tuvieron en cuenta una serie de atributos como el color, textura y uniformidad entre otros, así
(Cain 2005).

22
Luis M. del Papa et al.
RESULTADOS
Determinación Taxonómica
Se ha llegado a distintos niveles de inferencia taxonómica (Tabla 1), en la que se pudo determinar siete especies de mamíferos (Chaetophractus
gymnocercus, Dolichotis patagonum, Dolichotis
salinicola, y Calomys
callosus,), una especie de reptil (Tupinambis
rufescens), una de aves (Rhea americana) y en los restantes casos se han llegado a la categoría de género (Mazama, Ctenomys), Subfamilia (Caviinae, Sigmodontinae), familia (Characidae, Canidae,
Cervidae, Dasypodidae), suborden (Caviomorpha), orden (Anura, Artiodactyla) y clase (Gasteropoda, Osteichthyes, Aves, Mammalia). Con respecto a la categoría Mammalia se ha diferenciado en mamíferos pequeños (entre 1kg y 50 kg, e.g. roedores Caviomorpha, cérvidos pequeños como Mazama sp. y cánidos como gymnocercus) y mamíferos grandes (más de 50 kg, e.g. Lama sp.). La categoría mamíferos pequeños a grandes corresponde a aquellos especímenes que no pudieron asignarse a las categorías de tamaño anteriores (cercanas al tamaño de los 50 kg, e.g.
Puma concolor ).
En el sector I se observa una clara preponderancia de camélidos, no sólo por su abundancia taxonómica
Tabla 1. Número de restos, abundancia taxonómica e índice de fragmentación. *cáscaras de huevos
Mj I Mj III
Taxón N° MNI MNEt NISP/MNEt N° MNI MNEt NISP/MNEt
caracoles terrestres 2 - - - 11 11 - -
sp. 85 85 - - 1 1 - -
Osteichthyes - - - - 23 1 23 1
Characidae 1 1 1 1 - - - -
Anuro 1 1 1 1 - - - -
Chelonoidis sp. - - - - 14 1 14 1
Tupinambis rufescens 17 1 17 1 34 1 28 1,21
Aves indet. 23 6 20 1,15 119 12 111 1,07
Rhea americana 18 1 16 1,12 48 5 36 1,33
Rhea americana* 242 - - - 104 - - -
Mamíferos pequeños 11 1 10 1,1 35 2 16 2,19
Mamíferos peq. a grandes 28 1 4 7 73 1 11 6,64
Mamíferos grandes 261 3 31 8,42 356 3 33 10,79
Dasypodidae 2 1 2 1 8 2 7 1,14
Chaetophractus vellerosus 29 1 - - 19 1 - -
Canidae 2 1 1 2 - - - -
- - - - 2 1 2 1
Puma concolor - - - - 1 1 1 1
Ungulata 27 2 3 9 70 1 12 5,83
Artiodactyla 6 1 3 2 36 2 20 1,8
Lama sp. 129 4 71 - 549 8 345 1,59
Cervidae pequeños 1 1 1 1 1 1 1 1
Mazama sp. 4 1 4 1 3 1 2 1,5
Caviomorpha 15 2 14 - 46 2 46 1
Dolichotis patagonum 7 1 6 1,17 15 2 14 1,07
Dolichotis salinicola - - - - 2 1 2 1
5 1 4 1,25 20 2 16 1,25
Caviinae 4 1 3 1,33 2 2 2 1
Ctenomys sp. 3 2 3 1 - - - -
Sigmodontinae 5 1 5 1 2 1 2 1
Calomys callosus 8 2 8 1 2 2 2 1
indet 472 - - - 349 - - -
Totales 1408 - - - 1945 - - -
23
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
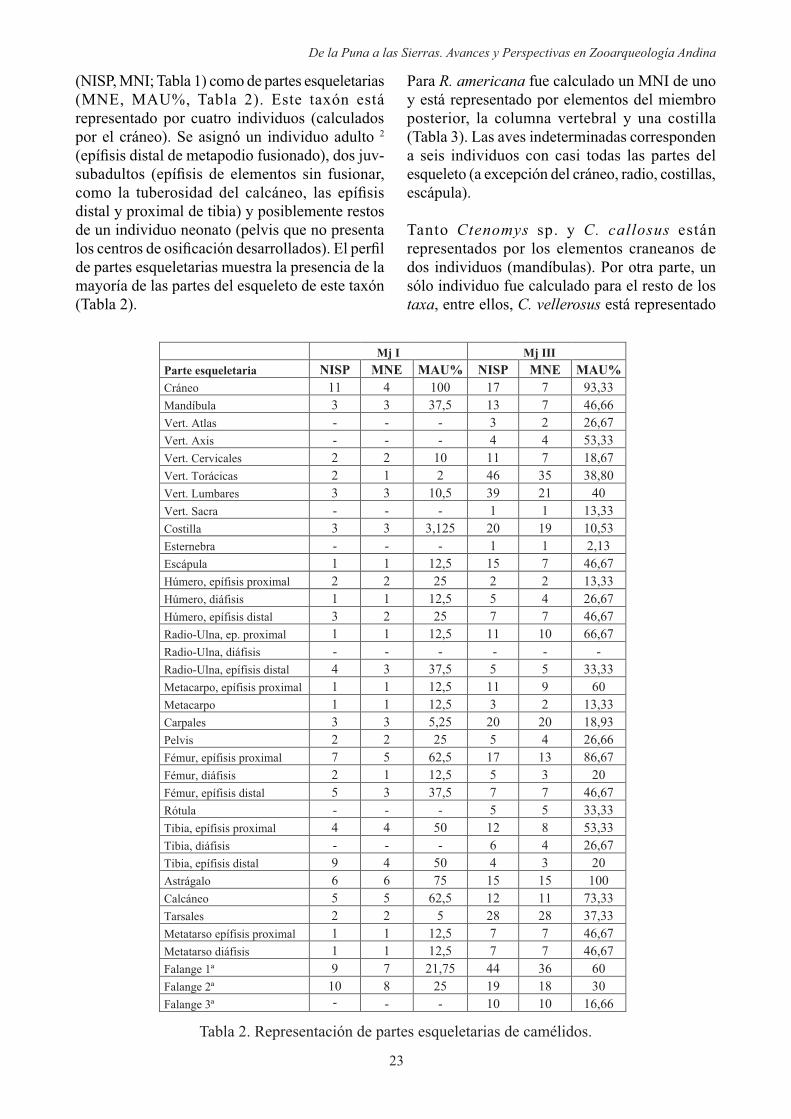
(NISP, MNI; Tabla 1) como de partes esqueletarias (MNE, MAU%, Tabla 2). Este taxón está representado por cuatro individuos (calculados por el cráneo). Se asignó un individuo adulto 2
distal y proximal de tibia) y posiblemente restos de un individuo neonato (pelvis que no presenta
de partes esqueletarias muestra la presencia de la mayoría de las partes del esqueleto de este taxón (Tabla 2).
Para R. americana fue calculado un MNI de uno y está representado por elementos del miembro posterior, la columna vertebral y una costilla (Tabla 3). Las aves indeterminadas corresponden a seis individuos con casi todas las partes del esqueleto (a excepción del cráneo, radio, costillas, escápula).
Tanto Ctenomys sp. y C. callosus están representados por los elementos craneanos de dos individuos (mandíbulas). Por otra parte, un sólo individuo fue calculado para el resto de los
, entre ellos, C. vellerosus está representado
Mj I Mj III
Parte esqueletaria NISP MNE MAU% NISP MNE MAU%
Cráneo 11 4 100 17 7 93,33Mandíbula 3 3 37,5 13 7 46,66Vert. Atlas - - - 3 2 26,67Vert. Axis - - - 4 4 53,33Vert. Cervicales 2 2 10 11 7 18,67Vert. Torácicas 2 1 2 46 35 38,80Vert. Lumbares 3 3 10,5 39 21 40Vert. Sacra - - - 1 1 13,33Costilla 3 3 3,125 20 19 10,53Esternebra - - - 1 1 2,13Escápula 1 1 12,5 15 7 46,67Húmero, epífisis proximal 2 2 25 2 2 13,33Húmero, diáfisis 1 1 12,5 5 4 26,67Húmero, epífisis distal 3 2 25 7 7 46,67Radio-Ulna, ep. proximal 1 1 12,5 11 10 66,67Radio-Ulna, diáfisis - - - - - -Radio-Ulna, epífisis distal 4 3 37,5 5 5 33,33Metacarpo, epífisis proximal 1 1 12,5 11 9 60Metacarpo 1 1 12,5 3 2 13,33Carpales 3 3 5,25 20 20 18,93Pelvis 2 2 25 5 4 26,66Fémur, epífisis proximal 7 5 62,5 17 13 86,67Fémur, diáfisis 2 1 12,5 5 3 20Fémur, epífisis distal 5 3 37,5 7 7 46,67Rótula - - - 5 5 33,33Tibia, epífisis proximal 4 4 50 12 8 53,33Tibia, diáfisis - - - 6 4 26,67Tibia, epífisis distal 9 4 50 4 3 20Astrágalo 6 6 75 15 15 100Calcáneo 5 5 62,5 12 11 73,33Tarsales 2 2 5 28 28 37,33Metatarso epífisis proximal 1 1 12,5 7 7 46,67Metatarso diáfisis 1 1 12,5 7 7 46,67Falange 1ª 9 7 21,75 44 36 60Falange 2ª 10 8 25 19 18 30Falange 3ª - - - 10 10 16,66
Tabla 2. Representación de partes esqueletarias de camélidos.

24
Luis M. del Papa et al.
por las placas de su armadura ósea, Mazama sp. por un fémur, falanges y calcáneo; los restos de D. patagonum pertenecen a elementos del cráneo, vértebras, radio, fémur y tibia; y para sólo se hallaron elementos del cráneo y miembro posterior.
La mayoría de los elementos de T. rufescens
corresponden a las vértebras, también se hallaron un especimen de cráneo y uno de cintura pélvica. El especimen de Characidae pertenece a un elemento del cráneo.
En el sector III (Tabla 1), también predominan los camélidos con ocho individuos (calculados
de metapodios y epífisis proximal de tibia fusionados), cuatro juv-subadultos (tuberosidad
de radio-ulna sin fusionar) y dos posibles neonatos (elementos apenas desarrollados, como tibia, vértebras, radio, ulnas, falanges y pelvis). En este sector están representadas todas las partes del esqueleto de dicho taxón (Tabla 2).
Las aves indeterminadas con 12 individuos (representadas todas las partes del esqueleto). Para R. americana se calcularon cinco individuos (por fíbulas) y se hallaron elementos del miembro posterior, cráneo, columna vertebral, costillas y sinsacro (Tabla 3). Tanto D. patagonum, L.
y C. callosus están representados por dos individuos, el primero de ellos calculados por la tibia y el resto por elementos craneales (maxilares y mandíbulas). D. patagonum y L.
, tiene una buena representación de las
Mj I Mj III
Parte esqueletaria NISP MNE MAU% NISP MNE MAU%
Cráneo - - - 1 1 28,57Mandíbula - - - - - -Vértebras 6 4 18 5 4 5,14costilla, porción vertebral 1 1 8 6 5 11,71costilla, porción esternal - - - - - -Escápula - - - - - -Coracoides - - - - - -Esternón - - - - - -Húmero, epífisis proximal - - - - - -Húmero, diáfisis - - - - - -Húmero, epífisis distal - - - - - -Radio - - - - - -Ulna - - - - - -Carpo-metacarpo - - - - - -Sinsacro - - - 1 1 28,57Fémur, epífisis proximal 1 1 50 - - -Fémur, diáfisis - - - - - -Fémur, epífisis distal 1 1 50 1 1 14,28Tibia-tarso, epífisis proximal 2 1 50 5 5 71,42Tibia-tarso, diáfisis - - - 6 3 42,85Tibia-tarso epífisis distal - - - 2 2 28,57Tarso-metatarso - - - 4 - -Tarso-metatarso, ep. px. - - - - 4 57,14Tarso-metatarso, diáfisis - - - - 4 57,14Tarso-metatarso, ep. dst. 2 2 100 - 4 57,14Fíbula - - - 7 - -Fíbula epífisis proximal - - - - 7 100Fíbula diáfisis - - - - 7 1001ª falange 2 2 33 3 3 14,282ª falange 1 1 16 2 2 9,423ª falange 1 1 16 - - -4ª falange 1 1 16 3 3 14,28
Tabla 3. Representación de partes esqueletarias de réidos.
25
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
distintas partes del esqueleto (cráneo, vértebras y miembro posterior).
Para el resto de los se calculó un MNI de uno. Tanto C. vellerosus como Chelonoidis sp. están representados mayormente por placas de la armadura ósea (del segundo también se hallaron vértebras). La mayoría de los elementos determinados a peces y T. rufescens corresponden a vértebras, seguidos por elementos del cráneo. D. salinicola, C. callosus y Caviinae fueron determinados por los elementos craneales.
Sólo se recuperó un especimen de P. concolor correspondiente a la epífisis distal de fémur, un metapodio y un radio de Mazama sp, una mandíbula y un fémur de P. gymnocercus.
Conservación de la Muestra
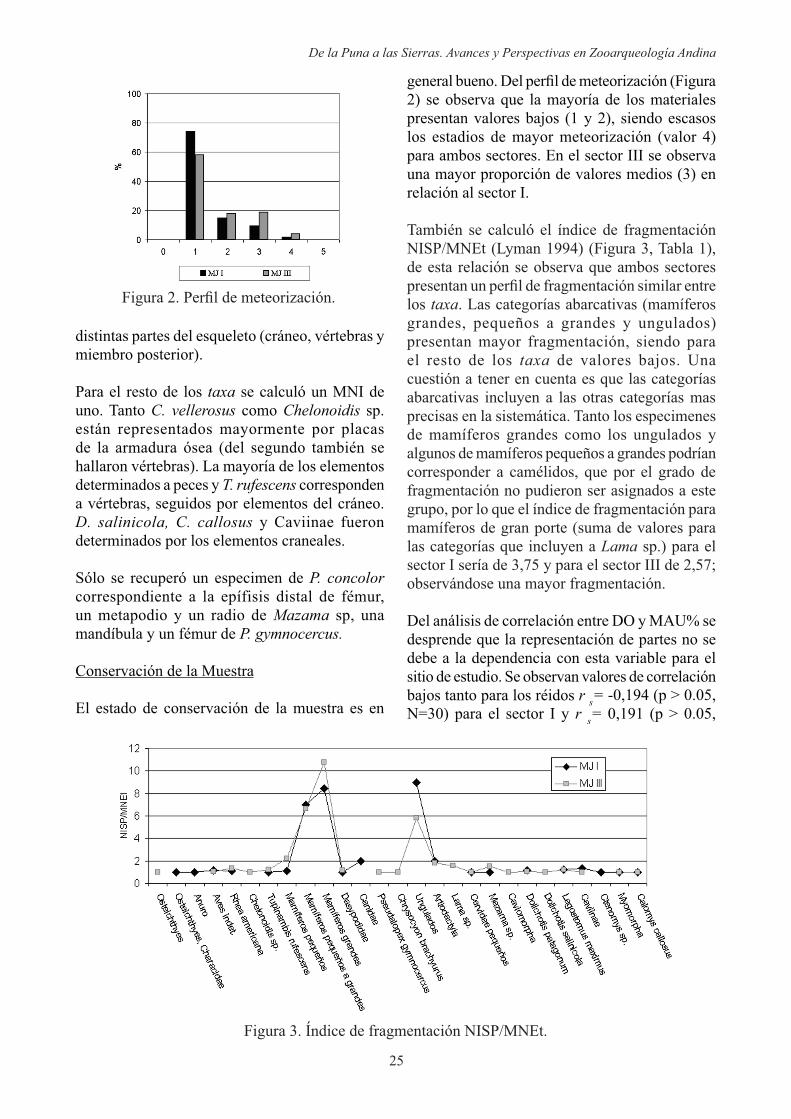
El estado de conservación de la muestra es en
2) se observa que la mayoría de los materiales presentan valores bajos (1 y 2), siendo escasos los estadios de mayor meteorización (valor 4) para ambos sectores. En el sector III se observa una mayor proporción de valores medios (3) en relación al sector I.
También se calculó el índice de fragmentación NISP/MNEt (Lyman 1994) (Figura 3, Tabla 1), de esta relación se observa que ambos sectores
los . Las categorías abarcativas (mamíferos grandes, pequeños a grandes y ungulados) presentan mayor fragmentación, siendo para el resto de los de valores bajos. Una cuestión a tener en cuenta es que las categorías abarcativas incluyen a las otras categorías mas precisas en la sistemática. Tanto los especimenes de mamíferos grandes como los ungulados y algunos de mamíferos pequeños a grandes podrían corresponder a camélidos, que por el grado de fragmentación no pudieron ser asignados a este grupo, por lo que el índice de fragmentación para mamíferos de gran porte (suma de valores para las categorías que incluyen a Lama sp.) para el sector I sería de 3,75 y para el sector III de 2,57; observándose una mayor fragmentación.
Del análisis de correlación entre DO y MAU% se desprende que la representación de partes no se debe a la dependencia con esta variable para el sitio de estudio. Se observan valores de correlación bajos tanto para los réidos r
s= -0,194 (p > 0.05,
N=30) para el sector I y r s= 0,191 (p > 0.05,
Figura 3. Índice de fragmentación NISP/MNEt.

26
Luis M. del Papa et al.
N=30) para el sector III, como para camélidos r
s= 0,005 (p > 0.05, N=31) del primer sector
y de r s= 0,072 (p > 0.05, N=31) del sector III.
Dichos resultados indican que las correlaciones
representados la mayoría de los elementos de camélidos (Tabla 2), tanto de baja y alta DO. Si
esqueletarias diferente (Tabla 3), ya que no están representadas todos los elementos, en la muestra se encuentran aquellos de baja DO (e.g. cráneo,
distales del fémur y del tibia-tarso) junto a los de alta densidad.
M de la S Ósea
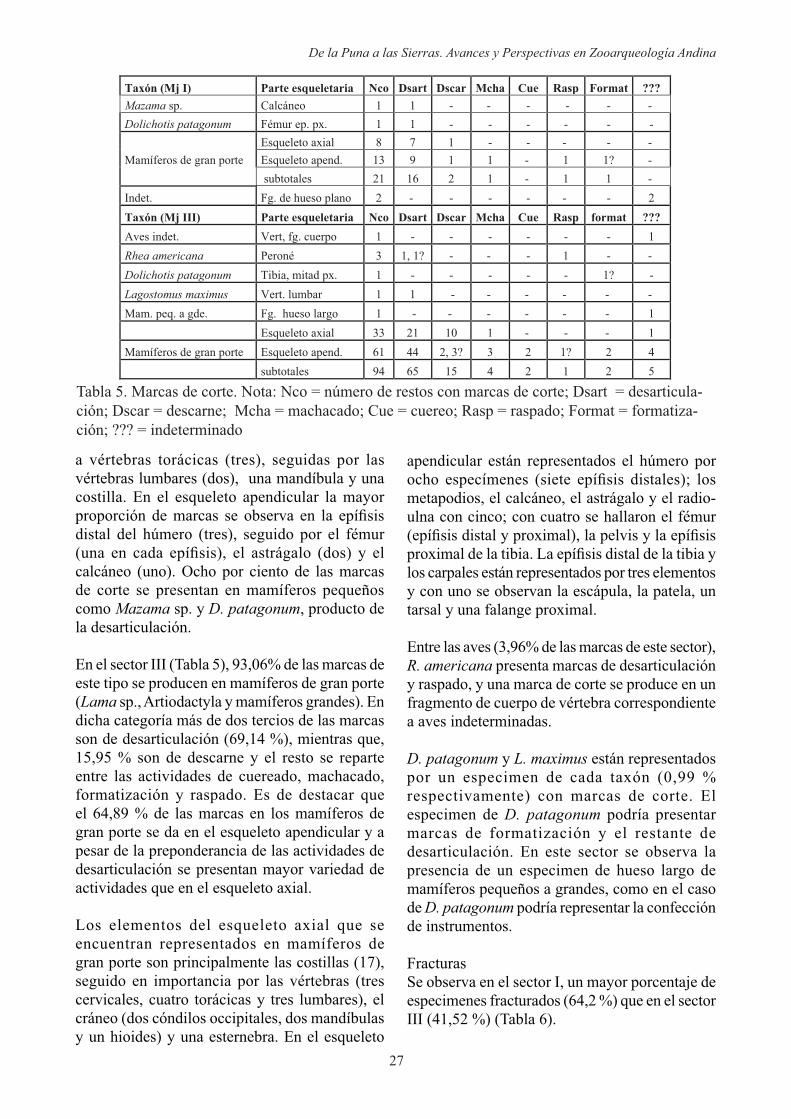
Un bajo porcentaje de especímenes (1,06 %) del Sector III presentan marcas producto de raíces (Tabla 4), principalmente en los primeros dos niveles. A pesar de la baja proporción de estas marcas, debemos considerar la posible acción de este agente como dispersor de los materiales, tanto horizontal como verticalmente. Esto se observa por el remontaje de especímenes óseos correspondientes al nivel inmediato inferior (entre nivel 1 y el 2, y entre nivel 2 y 3) y de distintas zonas de la cuadrícula del sector III, siendo los restantes niveles del sector de poca distorsión espacial, inferido por la presencia de elementos en estado articulado -región lumbar y cráneo de camélido fragmentado probablemente por el peso de los sedimentos (Figura 4)-. Dicha sección de la columna presenta marcas de corte en un elemento (última vértebra lumbar con marcas de desarticulación), por lo que podría estar representando el descarte de dicha estructura
luego del procesamiento primario.
Por la escasa evidencia de marcas de roedores sobre los especímenes (Tabla 4), representados con 0,07 % en el sector I y 0,05 % en el sector III es posible pensar en la baja incidencia de este agente
Esto se sustenta con la posible ausencia de cuevas en el registro, ya que no pudieron ser reconocidas dichas estructuras durante la excavación.
Las marcas producidas por carnívoros tienen una mayor frecuencia, aunque de escasa proporción en relación a los sectores (Tabla 4). En el sector I, las marcas producidas por este agente corresponden a
Lama sp. (uno con pozos, y otro con ahuecamiento y hoyuelos) y un fragmento de hueso largo de mamífero grande que presenta hoyuelos. En el sector III están representados los camélidos por un fragmento proximal de costilla con pozos, fragmento de diáfisis proximal de fémur y
y hoyuelos (éste último presenta marcas de corte), y una vértebra cervical con ranurado.
presenta ahuecamiento y hoyuelos, además de marcas de corte, y dos fragmentos de hueso largo de mamíferos grandes presentan pozos y hoyuelos respectivamente.
No fue posible distinguir la acción de ácidos gástricos en la muestra de microvertebrados.
Marcas de corteEn el sector I (Tabla 5), 84 % de las marcas de corte corresponden a especímenes de mamíferos de gran porte (Lama sp., mamíferos grandes, ungulados) y más de la mitad de las mismas está en el esqueleto apendicular. En su mayoría serían producto de la desarticulación del esqueleto (Binford 1981). Los elementos que presentan mayor incidencia de estas marcas en el esqueleto axial corresponden
Figura 4. Elementos en estado articulado, nive-les inferiores del Sector III.
Modificaciones óseas Mj I % MjIII %
Raíz - - 17 0,87
Carnívoros 3 0,22 7 0,36
Roedores 1 0,07 1 0,05
Corrosión gástrica* - - - -
microvertebrados (< 1 kg)
27
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
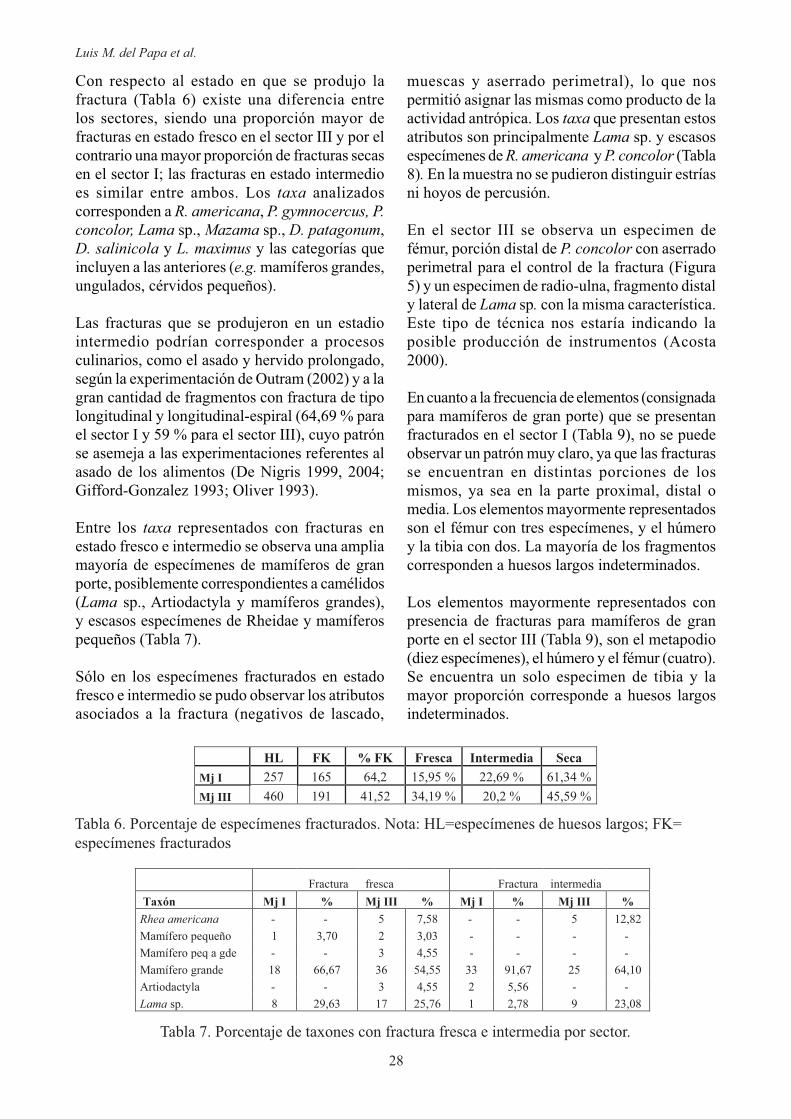
a vértebras torácicas (tres), seguidas por las vértebras lumbares (dos), una mandíbula y una costilla. En el esqueleto apendicular la mayor
distal del húmero (tres), seguido por el fémur
calcáneo (uno). Ocho por ciento de las marcas de corte se presentan en mamíferos pequeños como Mazama sp. y D. patagonum, producto de la desarticulación.
En el sector III (Tabla 5), 93,06% de las marcas de este tipo se producen en mamíferos de gran porte (Lama sp., Artiodactyla y mamíferos grandes). En dicha categoría más de dos tercios de las marcas son de desarticulación (69,14 %), mientras que, 15,95 % son de descarne y el resto se reparte entre las actividades de cuereado, machacado, formatización y raspado. Es de destacar que el 64,89 % de las marcas en los mamíferos de gran porte se da en el esqueleto apendicular y a pesar de la preponderancia de las actividades de desarticulación se presentan mayor variedad de actividades que en el esqueleto axial.
Los elementos del esqueleto axial que se encuentran representados en mamíferos de gran porte son principalmente las costillas (17), seguido en importancia por las vértebras (tres cervicales, cuatro torácicas y tres lumbares), el cráneo (dos cóndilos occipitales, dos mandíbulas y un hioides) y una esternebra. En el esqueleto
Tabla 5. Marcas de corte. Nota: Nco = número de restos con marcas de corte; Dsart = desarticula-ción; Dscar = descarne; Mcha = machacado; Cue = cuereo; Rasp = raspado; Format = formatiza-ción; ??? = indeterminado
apendicular están representados el húmero por
metapodios, el calcáneo, el astrágalo y el radio-ulna con cinco; con cuatro se hallaron el fémur
los carpales están representados por tres elementos y con uno se observan la escápula, la patela, un tarsal y una falange proximal.
Entre las aves (3,96% de las marcas de este sector), R. americana presenta marcas de desarticulación y raspado, y una marca de corte se produce en un fragmento de cuerpo de vértebra correspondiente a aves indeterminadas.
D. patagonum y están representados por un especimen de cada taxón (0,99 % respectivamente) con marcas de corte. El especimen de D. patagonum podría presentar marcas de formatización y el restante de desarticulación. En este sector se observa la presencia de un especimen de hueso largo de mamíferos pequeños a grandes, como en el caso de D. patagonum podría representar la confección de instrumentos.
FracturasSe observa en el sector I, un mayor porcentaje de especimenes fracturados (64,2 %) que en el sector III (41,52 %) (Tabla 6).
Taxón (Mj I) Parte esqueletaria Nco Dsart Dscar Mcha Cue Rasp Format ???
Mazama sp. Calcáneo 1 1 - - - - - -
Dolichotis patagonum Fémur ep. px. 1 1 - - - - - -
Esqueleto axial 8 7 1 - - - - -
Mamíferos de gran porte Esqueleto apend. 13 9 1 1 - 1 1? -
subtotales 21 16 2 1 - 1 1 -
Indet. Fg. de hueso plano 2 - - - - - - 2
Taxón (Mj III) Parte esqueletaria Nco Dsart Dscar Mcha Cue Rasp format ???
Aves indet. Vert, fg. cuerpo 1 - - - - - - 1
Rhea americana Peroné 3 1, 1? - - - 1 - -
Dolichotis patagonum Tibia, mitad px. 1 - - - - - 1? -
Vert. lumbar 1 1 - - - - - -
Mam. peq. a gde. Fg. hueso largo 1 - - - - - - 1
Esqueleto axial 33 21 10 1 - - - 1
Mamíferos de gran porte Esqueleto apend. 61 44 2, 3? 3 2 1? 2 4
subtotales 94 65 15 4 2 1 2 5
28
Luis M. del Papa et al.
Con respecto al estado en que se produjo la fractura (Tabla 6) existe una diferencia entre los sectores, siendo una proporción mayor de fracturas en estado fresco en el sector III y por el contrario una mayor proporción de fracturas secas en el sector I; las fracturas en estado intermedio es similar entre ambos. Los analizados corresponden a R. americana, P. gymnocercus, P.
concolor, Lama sp., Mazama sp., D. patagonum, D. salinicola y y las categorías que incluyen a las anteriores (e.g. mamíferos grandes, ungulados, cérvidos pequeños).
Las fracturas que se produjeron en un estadio intermedio podrían corresponder a procesos culinarios, como el asado y hervido prolongado, según la experimentación de Outram (2002) y a la gran cantidad de fragmentos con fractura de tipo longitudinal y longitudinal-espiral (64,69 % para el sector I y 59 % para el sector III), cuyo patrón se asemeja a las experimentaciones referentes al asado de los alimentos (De Nigris 1999, 2004; Gifford-Gonzalez 1993; Oliver 1993).
Entre los representados con fracturas en estado fresco e intermedio se observa una amplia mayoría de especímenes de mamíferos de gran porte, posiblemente correspondientes a camélidos (Lama sp., Artiodactyla y mamíferos grandes), y escasos especímenes de Rheidae y mamíferos pequeños (Tabla 7).
Sólo en los especímenes fracturados en estado fresco e intermedio se pudo observar los atributos asociados a la fractura (negativos de lascado,
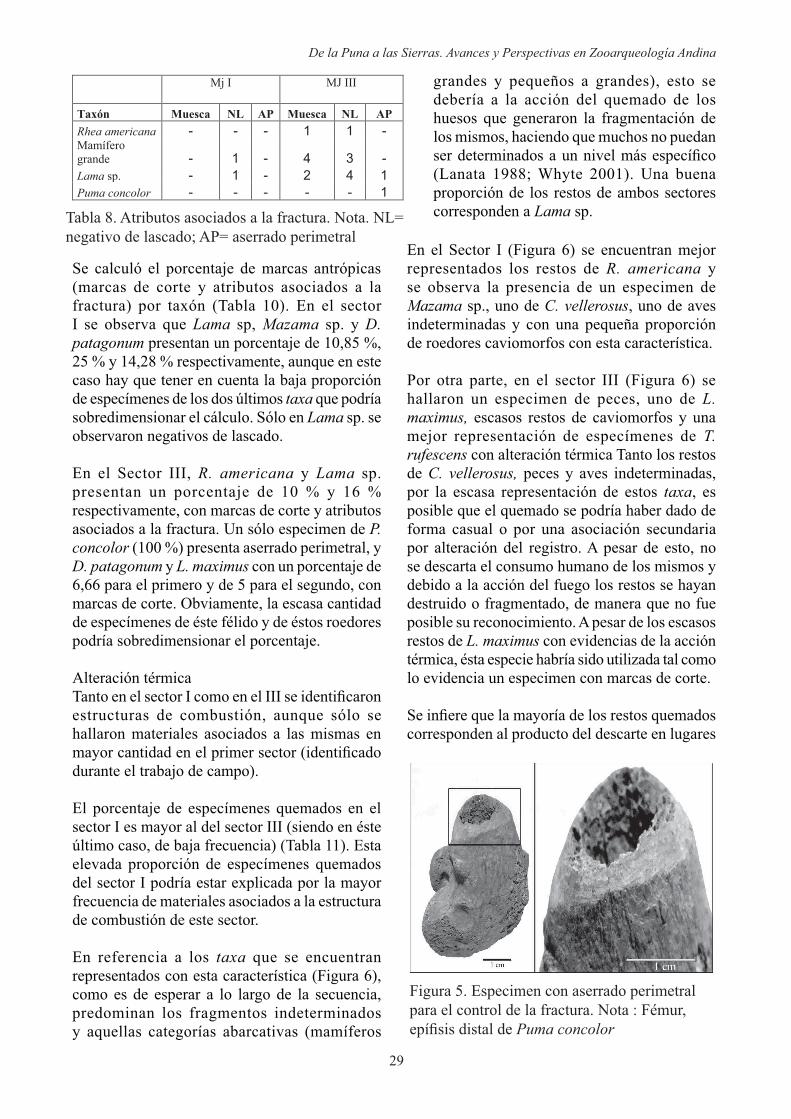
muescas y aserrado perimetral), lo que nos permitió asignar las mismas como producto de la actividad antrópica. Los que presentan estos atributos son principalmente Lama sp. y escasos especímenes de R. americana y P. concolor (Tabla 8). En la muestra no se pudieron distinguir estrías ni hoyos de percusión.
En el sector III se observa un especimen de fémur, porción distal de P. concolor con aserrado perimetral para el control de la fractura (Figura 5) y un especimen de radio-ulna, fragmento distal y lateral de Lama sp. con la misma característica. Este tipo de técnica nos estaría indicando la posible producción de instrumentos (Acosta 2000).
En cuanto a la frecuencia de elementos (consignada para mamíferos de gran porte) que se presentan fracturados en el sector I (Tabla 9), no se puede observar un patrón muy claro, ya que las fracturas se encuentran en distintas porciones de los mismos, ya sea en la parte proximal, distal o media. Los elementos mayormente representados son el fémur con tres especímenes, y el húmero y la tibia con dos. La mayoría de los fragmentos corresponden a huesos largos indeterminados.
Los elementos mayormente representados con presencia de fracturas para mamíferos de gran porte en el sector III (Tabla 9), son el metapodio (diez especímenes), el húmero y el fémur (cuatro). Se encuentra un solo especimen de tibia y la mayor proporción corresponde a huesos largos indeterminados.
HL FK % FK Fresca Intermedia Seca
Mj I 257 165 64,2 15,95 % 22,69 % 61,34 %
Mj III 460 191 41,52 34,19 % 20,2 % 45,59 %
Fractura fresca Fractura intermedia
Taxón Mj I % Mj III % Mj I % Mj III %
Rhea americana - - 5 7,58 - - 5 12,82
Mamífero pequeño 1 3,70 2 3,03 - - - -
Mamífero peq a gde - - 3 4,55 - - - -
Mamífero grande 18 66,67 36 54,55 33 91,67 25 64,10
Artiodactyla - - 3 4,55 2 5,56 - -
Lama sp. 8 29,63 17 25,76 1 2,78 9 23,08
Tabla 6. Porcentaje de especímenes fracturados. Nota: HL=especímenes de huesos largos; FK= especímenes fracturados
Tabla 7. Porcentaje de taxones con fractura fresca e intermedia por sector.
29
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
Se calculó el porcentaje de marcas antrópicas (marcas de corte y atributos asociados a la fractura) por taxón (Tabla 10). En el sector I se observa que Lama sp, Mazama sp. y D.
patagonum presentan un porcentaje de 10,85 %, 25 % y 14,28 % respectivamente, aunque en este caso hay que tener en cuenta la baja proporción de especímenes de los dos últimos que podría sobredimensionar el cálculo. Sólo en Lama sp. se observaron negativos de lascado.
En el Sector III, R. americana y Lama sp. presentan un porcentaje de 10 % y 16 % respectivamente, con marcas de corte y atributos asociados a la fractura. Un sólo especimen de P.
concolor (100 %) presenta aserrado perimetral, y D. patagonum y con un porcentaje de 6,66 para el primero y de 5 para el segundo, con marcas de corte. Obviamente, la escasa cantidad de especímenes de éste félido y de éstos roedores podría sobredimensionar el porcentaje.
Alteración térmica
estructuras de combustión, aunque sólo se hallaron materiales asociados a las mismas en
durante el trabajo de campo).
El porcentaje de especímenes quemados en el sector I es mayor al del sector III (siendo en éste último caso, de baja frecuencia) (Tabla 11). Esta elevada proporción de especímenes quemados del sector I podría estar explicada por la mayor frecuencia de materiales asociados a la estructura de combustión de este sector.
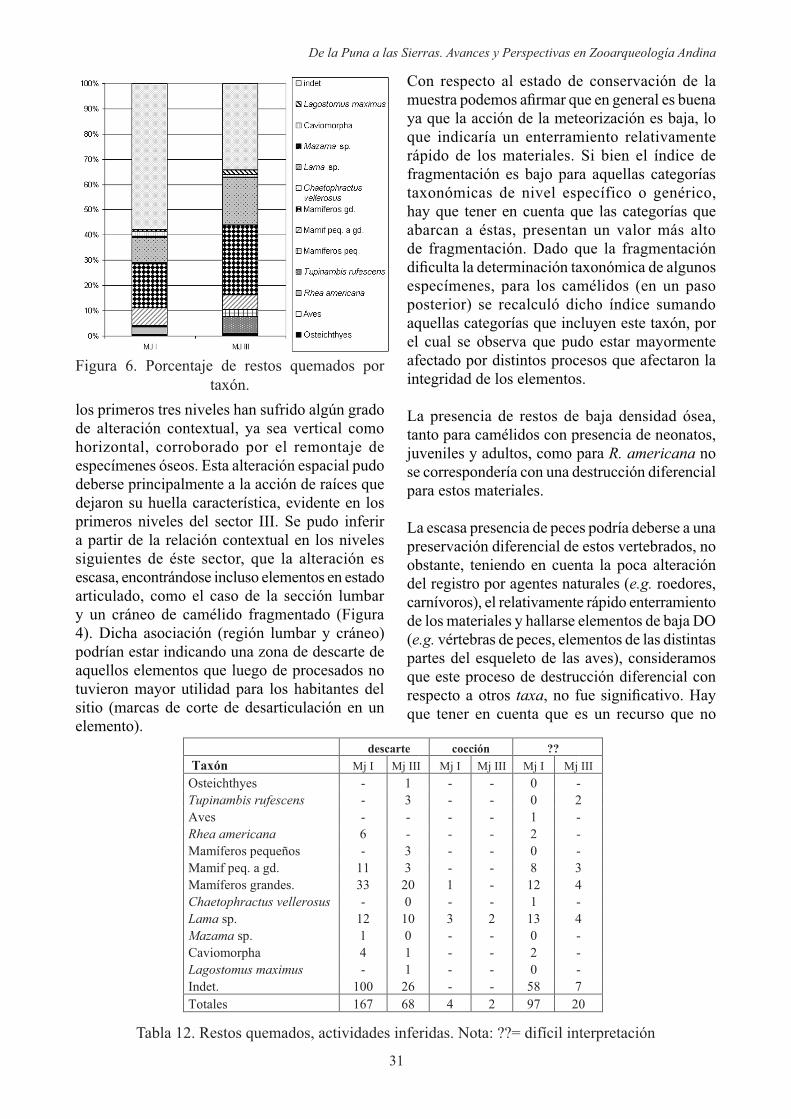
En referencia a los que se encuentran representados con esta característica (Figura 6), como es de esperar a lo largo de la secuencia, predominan los fragmentos indeterminados y aquellas categorías abarcativas (mamíferos
En el Sector I (Figura 6) se encuentran mejor representados los restos de R. americana y se observa la presencia de un especimen de Mazama sp., uno de C. vellerosus, uno de aves indeterminadas y con una pequeña proporción de roedores caviomorfos con esta característica.
Por otra parte, en el sector III (Figura 6) se hallaron un especimen de peces, uno de L.
escasos restos de caviomorfos y una mejor representación de especímenes de T.
rufescens con alteración térmica Tanto los restos de C. vellerosus, peces y aves indeterminadas, por la escasa representación de estos , es posible que el quemado se podría haber dado de forma casual o por una asociación secundaria por alteración del registro. A pesar de esto, no se descarta el consumo humano de los mismos y debido a la acción del fuego los restos se hayan destruido o fragmentado, de manera que no fue posible su reconocimiento. A pesar de los escasos restos de con evidencias de la acción térmica, ésta especie habría sido utilizada tal como lo evidencia un especimen con marcas de corte.
corresponden al producto del descarte en lugares
Mj I MJ III
Taxón Muesca NL AP Muesca NL AP
Rhea americana - - - 1 1 -
Mamífero grande - 1 - 4 3 -
Lama sp. - 1 - 2 4 1
Puma concolor - - - - - 1
Tabla 8. Atributos asociados a la fractura. Nota. NL= negativo de lascado; AP= aserrado perimetral
grandes y pequeños a grandes), esto se debería a la acción del quemado de los huesos que generaron la fragmentación de los mismos, haciendo que muchos no puedan
(Lanata 1988; Whyte 2001). Una buena proporción de los restos de ambos sectores corresponden a Lama sp.
Figura 5. Especimen con aserrado perimetral para el control de la fractura. Nota : Fémur,
Puma concolor

30
Luis M. del Papa et al.
de combustión (Tabla 12), por la gran cantidad de fragmentos donde el quemado compromete a la mayor parte de los mismos y donde fueron sometidos a grandes temperaturas, evidenciado por la gran mayoría de elementos carbonizados (41 % de los restos quemados del sector I y 42 % del sector III) y calcinados (20 % en el sector I y 30 % para el sector III) (Tabla 11).
La cocción de los alimentos podría estar evidenciada en una muy baja proporción (Tabla 12), tanto en el sector I (4 especímenes) como en el sector III (2 especímenes), donde se observa la presencia de restos de camélidos con esta
una alteración de menor grado).
DISCUSIÓN
El eje de este trabajo fue presentar un análisis que nos permita diferenciar patrones sobre el ingreso de los materiales al registro, su depositación,
postdepositacionales. Se pudo establecer que la acción de carnívoros en el sitio es muy baja, como es de esperar en sitios a cielo abierto, en donde habitan depredadores que en general son pequeños, de hábitos solitarios y mayormente carroñeros como los cánidos, aunque no se descarta la intervención de félidos de gran tamaño. Basándonos en estudios previos para otras regiones del país (Borrero y Martín 1996; Martín 1998; Mondini 1995, 2004; Nasti 2000) esperábamos que la acción de carnívoros para el sitio de estudio se diera de una manera menos intensiva que en otras regiones del mundo. Además, el acceso secundario a los restos por parte de los carnívoros (luego de haber sido procesados por los humanos) era otro factor por el cual esperábamos una baja frecuencia de marcas (Blumenschine 1988; Lupo 1995; Marean y Spencer 1991; Yellen 1991). En el caso que aquí tratamos, el acceso secundario se encuentra evidenciado en algunos especímenes que contienen marcas de corte y de carnívoros.
La escasa evidencia de la acción de roedores por roído y que la construcción de cuevas no fueron
que no fue de gran importancia en la alteración espacial del registro. Con respecto a esto último (dispersión) se considera que los materiales de
Mj I Mj IIIParte esqueletaria NISP FK NISP FK
Húmero, fragmento proximal 1 -
Húmero, fragmento diáfisis - 2
Húmero, fragmento distal 1 2
Radio-ulna, fragmento proximal 1 -
Fémur, fragmento proximal 1 2
Fémur, fragmento diáfisis 1 1
Fémur, fragmento distal 1 1
Tibia, fragmento proximal 1 1
Tibia, fragmento distal 1 -
Metapodio, fragmento proximal - 4
Metapodio, fragmento diáfisis - 5
Metapodio, fragmento distal - 1
Fragmento de hueso largo 17 27
MJ I MJIII
Taxón NISP Mco % Fk* % Total % NISP Mco % Fk* % Total %
Aves indet. 23 - - - - - - 119 1 0,84 - - - -
R. americana 18 - - - - - - 48 3 6,25 2 4,16 5 10,41
P. concolor 0 - - - - - - 1 - - 1 100 1 100
Lama sp. 129 13 10,07 1 0,77 14 10,85 549 81 14,75 7 1,27 88 16,02
Mazama sp. 4 1 25 - - 1 25 3 - - - - - -
D. patagonum 7 1 14,28 - - 1 14,28 15 1 6,66 - - 1 6,66
5 - - - - - - 20 1 5 - - 1 5
N Q Q % Carbonizados Calcinados
Mj I 1079 268 24,83 41% 20%
Mj III 1829 90 4,92 42% 30%
Tabla 10. Porcentaje de atributos de acción antrópica por taxón, por sector. Nota: Mco= marca de cor-te; Fk*= atributos asociados a la fractura (negativos de lascado, muescas, aserrado perimetral); Total= marcas de corte junto a los atributos asociados a la fractura.
Tabla 11. Porcentaje de restos quemados. N= restos, sin caracoles ni cáscaras de huevos Q= quemados
Tabla 9. Partes esqueletarias de mamíferos de gran porte con fractura fresca. NISP FK: especí-menes fracturados
31
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
los primeros tres niveles han sufrido algún grado de alteración contextual, ya sea vertical como horizontal, corroborado por el remontaje de especímenes óseos. Esta alteración espacial pudo deberse principalmente a la acción de raíces que dejaron su huella característica, evidente en los primeros niveles del sector III. Se pudo inferir a partir de la relación contextual en los niveles siguientes de éste sector, que la alteración es escasa, encontrándose incluso elementos en estado articulado, como el caso de la sección lumbar y un cráneo de camélido fragmentado (Figura 4). Dicha asociación (región lumbar y cráneo) podrían estar indicando una zona de descarte de aquellos elementos que luego de procesados no tuvieron mayor utilidad para los habitantes del sitio (marcas de corte de desarticulación en un elemento).
Con respecto al estado de conservación de la
ya que la acción de la meteorización es baja, lo que indicaría un enterramiento relativamente rápido de los materiales. Si bien el índice de fragmentación es bajo para aquellas categorías taxonómicas de nivel específico o genérico, hay que tener en cuenta que las categorías que abarcan a éstas, presentan un valor más alto de fragmentación. Dado que la fragmentación
especímenes, para los camélidos (en un paso posterior) se recalculó dicho índice sumando aquellas categorías que incluyen este taxón, por el cual se observa que pudo estar mayormente afectado por distintos procesos que afectaron la integridad de los elementos.
La presencia de restos de baja densidad ósea, tanto para camélidos con presencia de neonatos, juveniles y adultos, como para R. americana no se correspondería con una destrucción diferencial para estos materiales.
La escasa presencia de peces podría deberse a una preservación diferencial de estos vertebrados, no obstante, teniendo en cuenta la poca alteración del registro por agentes naturales (e.g. roedores, carnívoros), el relativamente rápido enterramiento de los materiales y hallarse elementos de baja DO (e.g. vértebras de peces, elementos de las distintas partes del esqueleto de las aves), consideramos que este proceso de destrucción diferencial con respecto a otros que tener en cuenta que es un recurso que no
Figura 6. Porcentaje de restos quemados por taxón.
Tabla 12. Restos quemados, actividades inferidas. Nota: ??= difícil interpretación
descarte cocción ??
Taxón Mj I Mj III Mj I Mj III Mj I Mj III
Osteichthyes - 1 - - 0 -Tupinambis rufescens - 3 - - 0 2Aves - - - - 1 -Rhea americana 6 - - - 2 -Mamíferos pequeños - 3 - - 0 -Mamif peq. a gd. 11 3 - - 8 3Mamíferos grandes. 33 20 1 - 12 4Chaetophractus vellerosus - 0 - - 1 -Lama sp. 12 10 3 2 13 4Mazama sp. 1 0 - - 0 -Caviomorpha 4 1 - - 2 -
- 1 - - 0 -Indet. 100 26 - - 58 7Totales 167 68 4 2 97 20

32
Luis M. del Papa et al.
se encuentra al alcance de las poblaciones que habitaron la zona (el río más cercano se encuentra a ca. 80 km), un ejemplo sería el caso del sitio arqueológico Villa la Punta (VP) muy cercano a Maquijata (Figura 1) en donde no esta representado este tipo de recurso, tal vez por la antigüedad del sitio (VP, 1.550 ± 60 años A.P. y 1.580 ± 60 años A.P., Togo 2007) o por corresponder a una etapa cultural anterior (contexto Las Mercedes). De
a la acción antrópica y su obtención pudo ser por distintas estrategias. Una de ellas, tal vez por
extensiones en busca de éste u otros recursos, es decir, salidas logísticas; y la otra opción sería por intercambio con poblaciones que habitaban las zonas ribereñas.
Del análisis se desprende que los diferentes representados en la muestra entraron al registro de diferentes maneras. Fue posible discriminar de los 18 presentes en el conjunto, aquellos que fueron utilizadas, ya sea como alimento, producción de instrumentos o ambos; resultando en un NTAXA = 7 para el Sector I y de un NTAXA = 8 en el Sector III, siendo un NTAXA general (ambos sectores) de 8, con claras evidencias de haber sido utilizadas.
Con respecto a la utilización antrópica, podemos decir que tanto R. americana, Lama sp., Mazama
sp., D. patagonum, y P. concolor presentan algún tipo de evidencia al respecto, ya sea por marcas de cortes, formatización, o fractura intencional, no sólo debido al consumo sino también como materia prima para la confección de instrumentos (e.g. el especimen de P. concolor con fractura de tipo transversal y aserrado perimetral, Figura 5)3. Otros muestran evidencias de alteración térmica como es el caso de T. rufescens y Osteichthyes4 con escasos restos, lo cual trae problemas a la hora de interpretar la utilización antrópica, principalmente del primero, ya que el quemado de los huesos se puede producir por una asociación secundaria con estructuras de combustión.
A partir de este análisis pudimos establecer que en el sitio se practicaron una gran variedad de actividades y la utilización de diversos .
Se observa fácilmente la preponderancia de los
camélidos en relación al resto de los recursos utilizados (Tabla 1), dejando relegados en segundo lugar a R. americana y en tercer lugar a los roedores caviomorfos con evidencias de uso (D.
patagonum y ). A pesar de la variedad de recursos, los pobladores desarrollaron una estrategia de obtención de presas de mayor retorno energético, en relación al tamaño de las mismas.
Otras especies pudieron ser potencialmente explotadas, si bien no presentan evidencias de ello. Con respecto al taxón aves es posible que sea uno de los recursos que los habitantes pudieron conseguir no sólo por su aporte proteínico sino también por sus plumas y huevos, como en el caso de R. americana. Se destaca la información brindada por los cronistas para esta región que dan cuenta de su uso (Diego Fernández, González de Prado y Pedro Sotelo de Narváez, en Palavecino 1940).
No se descarta el posible uso como recurso de D. salinicola, Dasipodidae (C. vellerosus) y Caviinae. Aunque estos están pobremente representados, su tamaño, la evidencia presentada por investigadores para otras regiones (Acosta y Pafundi 2005; Quintana 2005; Quintana et
al. 2002; Salemme 1988; Santiago 2004) y los estudios desarrollados en poblaciones actuales (Altrichter 2006) dejan abierta la posibilidad de su uso antrópico. Es probable que su utilización haya sido como recurso ocasional o circunstancial. Esto también pudo ser el caso de Chelonoidis sp., quelonio que se encuentra representada en los tres primeros niveles del sector III con escasos restos, aunque para este taxón se debe tener en cuenta la probable distorsión del registro por raíces y su posible depositación natural.
La presencia de cánidos (e.g. P. gymnocercus) no se debería a una actividad esencial para la subsistencia, dado los escasos restos hallados. Hay que tener en cuenta que sus actividades serían principalmente nocturnas y de hábitos solitarios (Redford y Eisenberg 1992), lo que representaría un gran costo de obtención como recurso; aunque no se descarta el uso de trampas (lo que facilitaría su obtención y reduciría los costos de búsqueda), de este modo se esperaría una mejor representación de este taxón. Sin embargo, al hallarse restos óseos con marcas de corte y de carnívoros, nos lleva a pensar que P.

33
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
gymnocercus podría haber ingresado a la muestra, tal vez, por su aproximación al sitio en busca de comida por carroña y de esta manera los grupos humanos tuvieron un fácil acceso para la caza de este animal.
Se considera que tanto los anuros como los roedores Ctenomyidae y Sigmodontinae (e.g. C.
callosus) habrían sido introducidos por la acción de Strigiformes, especialmente aquellas que no modifican sustancialmente la superficie ósea, ya que no se hallaron evidencias de la acción de corrosión gástrica (e.g. Tyto alba y ) (Andrews 1990; Gómez 2007) y que el rango de tamaño de las presas coincide con la selección que producen éstos depredadores (Pardiñas 1999; Quintana et al. 2002).
Por último, en lo referente a la presencia de los diferentes en el registro, los moluscos terrestres ( sp.), son característicos de los sedimentos de la zona centro-norte de la Argentina, y no presentan evidencias de haber sido utilizados por el hombre, sobre todo como objetos decorativos o suntuarios en forma de collar.
CONCLUSIONES
Se considera imprescindible el estudio de los sitios arqueológicos desde una perspectiva tafonómica, para luego poder hacer inferencias respecto a la utilización de los recursos y el empleo de los diversos índices que servirán para interpretar la economía de los grupos que habitaron Santiago del Estero.
Se pudo inferir en el sitio bajo estudio, que fue habitado por pobladores que portaban tecnología cerámica asimilables a Sunchitúyoj y Famabalasto principalmente (Páez et al. 2009), que desarrollaron una estrategia especializada en la obtención de productos de camélidos, complementado en primer lugar con los de R.
americana y en segundo lugar los de roedores caviomorfos (D. patagonum y ). También se establece el posible uso de manera circunstancial (en algunos casos por su presencia durante ciertas estaciones del año) de T. rufescens, huevos de R. americana (en primavera-verano) y de Mazama sp.
Para algunos
cuya obtención pudo depender de otros factores o necesidades, entre ellos P. concolor con evidencias de su posible uso como materia prima para la confección de instrumentos y la obtención de peces, tal vez por intercambio con pobladores ribereños o salidas logísticas en su busca.
Si bien en la muestra nos fue posible reconocer en algunos casos la acción humana en la utilización de ciertos recursos, es de destacar que se han reconocido otros agentes que pudieron actuar como formadores del registro, como es el caso de los carnívoros u otros depredadores (Strigiformes) y la posible muerte natural de los organismos; así como el reconocimiento de los agentes
los primeros niveles del sector III, por raíces). A pesar de que la representación de la acción de estos agentes o procesos es relativamente baja para la muestra, el reconocimiento de los mismos nos proporciona información que se tendrá en cuenta en el futuro análisis de sitios arqueológicos del área.
Agradecimientos. A Reinaldo Moralejo por su ayuda con el software de estadística y la confección de la Figura 1. A César García Esponda por la traducción del resumen al inglés y a M. Amelia Martínez por la lectura de las versiones preliminares del trabajo. A Andrés Izeta y Guillermo Mengoni Goñalons por invitarnos a participar de este volumen y a dos evaluadores anónimos cuyos comentarios ayudaron a mejorar la producción del trabajo. Lo aquí expresado es responsabilidad de los autores.Este trabajo forma parte del proyecto de beca doctoral del CONICET otorgada al primer autor.
REFERENCIAS CITADAS
Acosta, A. 2000 Huellas de corte relacionadas con la manufactura de artefactos óseos en el nordeste de la provincia de Buenos Aires. Relaciones de
la Sociedad Argentina de Antropología XXV: 159-177.
Acosta, A. y L. Pafundi. 2005 Zooarqueología y tafonomía de Cavia aperea en el humedal del Paraná inferior. Intersecciones en Antropología
6: 59-74.
Altrichter, M. 2006 Wildlife in the life of local

34
Luis M. del Papa et al.
people of the semi-arid Argentine Chaco. Biodiversity and Conservation 15: 2719-2736.
Andrews, P. 1990 Owls, Caves and Fossils. Natural History Museum Publications, Londres.
Behrensmeyer, A. F. 1978 Taphonomic and ecologic information from bone weathering. Paleobiology 4: 150-162.
Binford, L. R. 1981 Bones: Ancient Men and
Modern Myths. Academic Press, New York.
Binford, L. R.1984 Faunal Remains from Klasies
River Mouth. Academic Press, Orlando.
Blumenschine, R. J. 1988 An experimental model
on archaeological bone assemblages. Journal of
Archaeological Science 15: 483-502.
Blumenschine, R. J., C. W. Marean y S. D. Capaldo. 1996 Blind test of inter-analyst correspondence
percussion marks, and carnivore tooth marks on bone surfaces. Journal of Archaeological Science 23: 493-507.
Blumenschine, R. J. y M. M. Selvaggio. 1988 Percussion marks on bone surfaces as a new diagnostic of hominid behavior. Nature 333: 763-765.
Bordas, A. 1940. Síntesis Paleontológica. En Los
Aborígenes de Santiago del Estero. Sociedad Argentina de Antropología, pp.117-121. Buenos Aires.
Borrero, L. A. y F. M. Martín. 1996 Tafonomía de carnívoros: un enfoque regional. En Arqueología.
Sólo Patagonia, editado por J. Gomez Otero, pp. 189-206. CENPAT-CONICET, Puerto Madryn, Chubut.
Bunn, H. T. 1981 Archaeological evidence for meat-eating by Plio-Pleistocene hominids at Koobi For and Olduvai Gorge. Nature 291: 574-577.
Bunn, H. T. 1989 Diagnosing Plio-Pleistocene hominid activity with bone fracture evidence. En
editado por R. Bonnichsen y
M. H. Sorg, pp. 299-315, University of Maine, Orono.
Cabrera, A. L. 1971 Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de
Botánica XIV (1-2), Buenos Aires.
Cain, C. R. 2005 Using Burned animal bone to look at Middle Stone Age occupation and behavior. Journal of Archaeological Science 32: 873-884.
Capaldo, S. D. y R. J. Blumenschine. 1994 A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing in bovid long bones. American Antiquity 59: 724-748.
Cione, A., A. M. Lorandi y E. P. Tonni. 1979. Patrón de Subsistencia y Adaptación Ecológica en la Aldea Prehispánica “El Veinte”, Santiago del Estero. Relaciones de la Sociedad Argentina
de Antropología XIII: 102-116.
Costamagno, S., I. Théry-Parisot, J. P. Brugal y R. Guibert. 2005. Taphonomic consequences of the use of bones as fuel: experimental data and archaeological consequences. En Biosphere
to Lithosphere. New Studies in Vertebrate
Taphonomy, editado por T. O’Connor, pp: 51-62, 9th ICAZ, Durham, 2002. Oxbow Books, Oxford.
Cruz, I. y D. Elkin. 2003 Structural Bone Density of the Lesser Rhea (Pterocnemia pennata) (Aves: Rheidae). Taphonomic and Archaeological Implications. Journal of Archaeological Science
30: 37-44.
De Nigris, M. E. 1999 Lo crudo y lo cocido: sobre
Arqueología 9: 239-364.
De Nigris, M. E. 2004 El consumo en
grupos cazadores recolectores. Un ejemplo
zooarqueológico de patagonia meridional. Sociedad Argentina de Antropología. Colección Tesis Doctorales, Buenos Aires.
del Papa, L. M. 2006 Primeras aproximaciones en el estudio del uso de animales en el sitio arqueológico Maquijata, provincia de Santiago del Estero. Trabajo presentado en las VII

35
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
Jornadas de Jóvenes Investigadores en Ciencias Antropológicas, Buenos Aires.
Dirección General de Minería y Geología. 1981 Mapa de suelos de la provincia de Santiago del
Estero.
Elkin, D. C. 1995 Volume density of South American camelids skeletal parts. Internacional
Journal of Osteoarchaeology 5: 29-37.
Fernández-Jalvo, Y. y P. Andrews 1992 Small mammal taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. Journal of Archaeological
Science 19:407-428.
Fernández Varela, V., J. Peña, D. Olivera y J. Togo. 2001 Los Recursos Animales en las Estrategias Económicas de Sociedades Prehispánicas de Santiago del Estero (R. A.) Trabajo presentado en el XIV Congreso Nacional de Arqueología Argentina, Rosario.
Gifford-Gonzalez, D. 1989 Ethnographic
Cases form East Africa. En . editado por R. Bonnichsen y M. Sorg, pp. 179-246, University of Maine, Orono.
Gifford-Gonzalez, D. 1991 Bones Are not Enough: Analogues, Knowledge, and Interpretive Strategies in Zooarchaeology. Journal of Anthropological
Archaeology 10: 215-254.
G i f f o r d - G o n z a l e z , D . 1 9 9 3 G a p s i n zooarchaeological analyses of butchery. Is gender an issue? En From Bones to Behavior, editado por J. Hudson, pp. 181-199, Southern Illinois University Press, Carbondale.
Gómez, G. N. 2007 Predators categorization based on taphonomic análisis of micromammal bones: a comparison to proposed models. En Taphonomy
and Zooarchaeology in Argentina, editado por M. A. Gutiérrez, L. Miotti, G. Barrientos, G. Mengoni Goñalons y M. Salemme, pp. 89- 103. BAR International Series 1601, Oxford.
González, A. R. 1979. Dinámica Cultural del N.O. Argentino. Evolución e Historia en las Culturas del N.O. Argentino. Antiquitas N° 28-29: 1-15.
Johnson, E. 1985 Current developments in bone tecnology. En Advances in Archaeological
Method and Theory vol. 8, editado por M.B. Schiffer, pp. 157-235, Academic Press. Orlando.
Kent, J. D. 1982. The Domestication and exploitation of South American camelids: methods of analysis and their application to circum-lacustrine archaeological sites in Bolivia and Peru. Ph.D. dissertation, Washington University, St. Louis.
Kent, S. 1993 Variability in Faunal Assemblages:
and Mode of Cooking on Faunal Remains at a Sedentary Kalahari Community. Journal of
Anthropological Archaeology 12: 323-383.
Kraglievich, L. y C. Rusconi. 1931. Restos de vertebrados vivientes y extinguidos hallados por los Señores E. R. Wagner y hermano en túmulos precolombinos de Santiago del Estero. Physis Tomo X: 229-241.
Lanata, J.L. 1988. ¿Huesos quemados, huesos contados? Los datos de un experimento. En De
, editado por N.R. Ratto y A.F. Haber, pp.75-90, Facultad de Filosofía y Letras, UBA, Buenos Aires.
Ledesma, R. 1961 Maquijata. Instituto de Lingüistica, Folklore y Arqueología, Facultad de Filosofía y Letras, Universidad Nacional de Tucumán, Santiago del Estero.
Lorandi, A. M. y N. Carrió. 1970-1975. Informe sobre las investigaciones arqueológicas en Santiago del Estero. Actas y Trabajos del primer
Congreso de Arqueología Argentina. Rosario.
Lorandi, A. M. y D. M. Lovera. 1972. Economía y Patrón de Asentamiento en la Provincia de Santiago del Estero. Relaciones de la Sociedad
Argentina de Antropología VI: 173-191.
Lupo, K. D. 1995 Hadza Bone Assemblages and Hyena Attrition: An Ethnographic Example of the
the Intensity of Scavenger Ravaging. Journal of
Anthropological Archaeology 14: 288-314.
Lyman, R. L. 1984 Bone density and differential

36
Luis M. del Papa et al.
survivorship of fossil classes. Journal of
Anthropological Archaeology 3: 259-299.
Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Marean, C. W, y L. M. Spencer. 1991 Impact of carnivore ravaging on zooarchaeological measures of element abundance. American
Antiquity 56: 645-658.
Martín, F. M. 1998 Madrigueras, dormideros y letrinas: Aproximación a la tafonomía de zorros. En Arqueología de la Patagonia Meridional
, compilado por L.A. Borrero, pp. 73-96, Ediciones Búsqueda de Ayllu, Concepción del Uruguay.
Mengoni Goñalons, G. L. 1988 Análisis de materiales faunísticos de sitios arqueológicos. Xama 1: 71-120.
Mengoni Goñalons, G. L. 1999 Cazadores de
guanacos de la estepa patagónica. Sociedad Argentina de Antropología. Colección tesis Doctorales. Buenos Aires.
Miotti, L. 1998 Zooarqueología de la Meseta
Central y Costa de Santa Cruz. Un enfoque de
las estrategias adaptativas aborígenes y los
paleoambientes. Revista del Museo de Historia Natural de San Rafael, Tomo X (1-4).
Mondini, M. 1995 Artiodactyl prey transport by foxes in Puna rock shelters. Current Anthropology 36: 520-524.
Mondini, M. 2004 La comunidad de predadores en la Puna durante el Holoceno, Interacciones bióticas entre humanos y carnívoros. Relaciones
de la Sociedad Argentina de Antropología XXIX: 183-209.
Montalvo, C. I., M. E. M. Pessino y V. H. González. 2007 Taphonomic análisis of remains of mammals eaten by pumas (Puma concolor Carnivora, Felidae) in central Argentina. Journal
of Archaeological Science 34: 2151-2160.
monte. Opera Lilloana 2, Tucumán, Miguel Lillo.
Morlan, R. E. 1994 Bison Bone Fragmentation and Survivorship: a Comparative Method. Journal
of Archaeological Science 21: 797-807.
in high-altitude deserts. Current Athropology 41: 179-283.
Nicholson, R. A. 1993 A Morphological Investigation of Burnt Animal Bone and Evaluation of its Utility in Archaeology. Journal
of Archaeological Science 20: 411-428.
Oliver, J. O. 1993 Carcass Processing by the Hadza: Bone Breakage from Butchery to Consumption. En From Bones to Behavior, editado por J. Hudson, pp. 200-207, Southern Illinois University Press, Carbondale.
Outram, A. K. 2002 Bone fracture and within-bone nutrients: an experimentally based method for investigating levels of marrow extraction. En Consuming Passions and Patterns of Consumption, editado por P. Miracle y N. Milner, pp. 51-62. McDonald Institute for Archaeological Research, Cambridge.
Páez, R., J. A. Togo y P. Villar Benítez. 2009 Primera Aproximación al sitio de Maquijata, sierras de Guasayán, Departamento de Choya, Santiago del Estero. En Entre Pasados y Presentes
II. Estudios Contemporáneos en Ciencias
Antropológicas, editado por T. Bourlot, D. Buzzoto, C. Crespo, A. C. Helcht y N. Kuperszmit, pp. 275-288. Fundación de Historia Natural Félix de Azara.
Palavecino, E. 1940 Síntesis histórica. En Los
Aborígenes de Santiago del Estero. Sociedad Argentina de Antropología, pp. 47-71. Buenos Aires.
Pardiñas, U. F. J . 1999 Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia. Arqueología 9: 265-308.
Parodi Bustos, R. 1947 La presencia del género Paleolama en los túmulos indígenas de Santiago del Estero. En Argentina. Tomo CXLIII: 3-9. Buenos Aires.
Pickering, T. R y C. P. Egeland. 2006 Experimental

37
De la Puna a las Sierras. Avances y Perspectivas en Zooarqueología Andina
patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. Journal of Archaeological
Science 33: 459-469.
Quintana, C. A. 2005 Despiece de microroedores en el Holoceno Tardío de las Sierras de Tandilla (Argentina). Archaeofauna 14: 227-241.
Quintana, C. A. 2007 Marcas de dientes de roedores en huesos de sitios arqueológicos de las sierras de Tandilia, Argentina. Archaeofauna 16: 185-191.
Quintana, C. A., F. Valverde y D. L. Mazzanti. 2002 Roedores y Lagartos Como Emergentes De
El Holoceno Tardío En Sierras De La Región Pampeana Argentina. Latin American Antiquity 13 (4): 455-473.
Redford K. J. y J. F. Eisenberg. 1992 Mammals
of the Neotropics. The southern Cone. Vol. 2, University of Chicago Press.
Ringuelet, R. A. 1961 Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22.
Rusconi, C. 1930 Las especies fósiles de pecaríes (Tayassuidae) y sus relaciones con las del Brasil y Norte América. Natural de Buenos Aires, Vol. XXVI: 228-241, Buenos Aires.
Rusconi, C. 1934. Nuevos restos de vertebrados vivientes y extinguidos de los túmulos prehispánicos de Santiago del Estero. En Wagner E y D Wagner. 1934. La Civilización Chaco-
Santiagueña y su correlación con el Viejo y Nuevo
Mundo. Tomo I, Impresora Argentina.
Salemme, M. C. 1988 Paleoetnozoología del sector bonaerense de la región pampeana, con especial atención a los mamíferos. Tesis Doctoral no publicada, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata.
Santiago, F. C. 2004 Los roedores en el “menú” de los habitantes de Cerro Aguará (provincia de Santa Fe): su análisis arqueofaunístico. Intersecciones
en Antropología 5: 3-18.
Shipman, P, G. F. Foster y M. Schoeninger. 1984 Burnt bones and teeth: an experimental study of colour, morphology, crystal structure and shrinkage. Journal of Archaeological Science
11: 307-325.
Stiner, M. C., S. L. Kuhn, S. Weiner y O. Bar-Yosef. 1995 Differential burning, recrystalization, and fragmentation of archaeological bone. Journal
of Archaeological Science 22: 223-237.
Togo, J. 2004 Arqueología Santiagueña: Estado actual del Conocimiento y Evolución de un Sector de la Cuenca del Río Dulce. Tesis Doctoral no publicada, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata.
Togo, J. 2007 Los fechados radiocarbónicos de Santiago del Estero. En Actas de resúmenes ampliados del XVI Congreso de Arqueología Argentina, Tomo III, pp. 227-232.
Wagner, E. y D. Wagner. 1936 Las Llanuras de
Santiago del Estero. En Historia de la Nación Argentina, Tomo I, Editorial El Ateneo, Buenos Aires.
Whyte, T. R. 2001 Distinguishing Remains of Human Cremations from Burned Animal Bones. Journal of Field Archaeology 8 (3-4): 437-448.
Yellen, J. E. 1991 Small mammals: post-discard patterning of ¡Kung San faunal remains. Journal
of Anthropological Archaeology 10: 152-192.
NOTAS
¹ Las calicatas eléctricas permiten realizar sondeos eléctricos, que de acuerdo a los valores de resistividad aparente del subsuelo (1/conductividad eléctrica de los materiales) denotan anomalidades del terreno. A partir de la detección de dichas anomalidades (probables concentraciones de materiales arqueológicos) se escogió la zona a excavar (para más detalle de la metodología, ver Togo 2004: 499-503).2 Edades estimadas según criterios de Kent (1982), las categorías utilizadas aquí corresponden a neonatos (elementos que apenas esbozan sus estructuras, menores a un año de edad), juv-subaldulto (categoría amplia, entre uno y tres años

38
Luis M. del Papa et al.
de edad) y adultos (más de tres años).3 El aserrado perimetral permite adelgazar la capa ósea, y así poder fracturar el hueso de manera controlada (Acosta 2000).4 Los peces a pesar de encontrarse quemados en
por un factor antrópico, dado la distancia hacia el río más próximo (80 km) y las estrategias referidas a su incorporación (ver discusión).




















