
Systematic Detection and Characterization of Hydrogen ... - SMU
15
Systematic Detection and Characterization of Hydrogen Bonding in Proteins via Local Vibrational Modes Niraj Verma, Yunwen Tao, and Elfi Kraka* Cite This: J. Phys. Chem. B 2021, 125, 2551-2565 Read Online ACCESS Metrics & More Article Recommendations * sı Supporting Information ABSTRACT: We introduce a new software, Ef f icient Detection of Hydrogen Bonds (EDHB), that systematically detects hydrogen bonds based on the nearest neighbors algorithm. EDHB classifies inter- and intramolecular hydrogen bonds as well as hydrogen bond networks. EDHB outperforms commonly used hydrogen bond detection methods in terms of speed of execution. An important additional feature of EDHB is that information from preceding quantum chemical studies (i.e., natural bond orbital analysis data and second energy derivative information) can be used to determine the electrostatic/covalent character of the hydrogen bonds and to calculate local-mode hydrogen bond force constants as a quantitative measure of their intrinsic strength. We applied EDHB to a diverse set of 163 proteins. We identified hydrogen bond networks forming intramolecular rings of different sizes as a common feature playing an important role for specific secondary structure orientations such as α-helixes and turns. However, these networks do not have a significant influence on the hydrogen bond strength. Our comprehensive local-mode analysis reveals the interesting result that the hydrogen bond angle is the governing factor determining the hydrogen bond strength in a protein. EDHB offers a broad range of application possibilities. In addition to proteins, EDHB can be generally used to detect and characterize hydrogen bonds in protein- ligand interactions, water clusters, and other systems where a hydrogen bond plays a critical role, as well as during molecular dynamics simulations. The program is freely available at https://github.com/ekraka/EDHB. ■ INTRODUCTION Hydrogen bonds (HBs) play a crucial role in proteins forging enzyme reactions, protein folding, protein-ligand interactions, and other processes. 1-3 The early work of Pauling and Mirsky highlighted already in 1936 the importance of HBs in proteins suggesting a 5 kcal/mol stability from each HB. 4 In 1951, they discovered α-helix and β-pleated sheet as the most important conformational elements in a protein with α-helix HBs on the order of 8 kcal/mol. 5,6 Later that same year, they realized that an aqueous environment can lead to HBs of the order of 2 kcal/mol. 6 After almost 7 decades of intensive research, there are still some controversies about the strength and properties of HBs in a protein. Theoretical studies in the early 90s suggest that HBs do not contribute to thermodynamic stability as the overall energy balance of HBs is close to zero. 7 Campos et al. 8 suggested with a combined experimental and theoretical study that HBs in fact destabilize the native conformation of a protein. Recent studies have provided a better picture of HBs in proteins, although controversies about their strength still persist. 9 The protein environment adds complexity to HBs’ stability, and thus inclusion of all atoms is critical. Studies conducted on fragments of a protein structure 10 are limited by this crucial information. Pace et al. 3,11 have concluded that the polar residues buried inside the protein interior undergo stronger van der Waals forces in a HB. Further studies by Hubbard and Kamran Haider 1 have shown that the buried intramolecular HBs are stronger due to their electrostatic nature. The protein environment can induce an electric field on the order of 100 MV/cm. 12 Thus, to incorporate the protein environment, we utilized all atoms of a protein incorporating the complex environment. A HB is influenced by a multitude of factors governing its stability. Intramolecular HBs are one such factor that plays a crucial role. 1,13-16 This fact was already highlighted by Takano et al. 17 in the 90s. Donor-acceptor pairs such as N-H···O, N-H···N, O-H···N, and O-H···O lead to HBs of different strengths due to the involvement of atoms with different electronegativities. 10,18 Another important aspect is the network of HBs, 19,20 where two or more HBs share the same acceptor. 21,22 A study by Ballesteros et al. 23 shows the implications of networks in guiding secondary structures in proteins. A study by Feldblum and Arkin 24 clearly showed the existence of various networks of HBs via both experimental and computational means. The influence of the protein environment and the various factors alter the strength of a Received: December 22, 2020 Revised: February 21, 2021 Published: March 5, 2021 Article pubs.acs.org/JPCB © 2021 American Chemical Society 2551 https://dx.doi.org/10.1021/acs.jpcb.0c11392 J. Phys. Chem. B 2021, 125, 2551-2565 Downloaded via SOUTHERN METHODIST UNIV on April 17, 2021 at 16:20:56 (UTC). See https://pubs.acs.org/sharingguidelines for options on how to legitimately share published articles.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Systematic Detection and Characterization of Hydrogen ... - SMU

Systematic Detection and Characterization of Hydrogen Bonding inProteins via Local Vibrational ModesNiraj Verma Yunwen Tao and Elfi Kraka
Cite This J Phys Chem B 2021 125 2551minus2565 Read Online
ACCESS Metrics amp More Article Recommendations sı Supporting Information
ABSTRACT We introduce a new software Ef f icient Detection of Hydrogen Bonds(EDHB) that systematically detects hydrogen bonds based on the nearestneighbors algorithm EDHB classifies inter- and intramolecular hydrogen bonds aswell as hydrogen bond networks EDHB outperforms commonly used hydrogenbond detection methods in terms of speed of execution An important additionalfeature of EDHB is that information from preceding quantum chemical studies(ie natural bond orbital analysis data and second energy derivative information)can be used to determine the electrostaticcovalent character of the hydrogenbonds and to calculate local-mode hydrogen bond force constants as aquantitative measure of their intrinsic strength We applied EDHB to a diverseset of 163 proteins We identified hydrogen bond networks forming intramolecular rings of different sizes as a common featureplaying an important role for specific secondary structure orientations such as α-helixes and turns However these networks do nothave a significant influence on the hydrogen bond strength Our comprehensive local-mode analysis reveals the interesting result thatthe hydrogen bond angle is the governing factor determining the hydrogen bond strength in a protein EDHB offers a broad range ofapplication possibilities In addition to proteins EDHB can be generally used to detect and characterize hydrogen bonds in proteinminusligand interactions water clusters and other systems where a hydrogen bond plays a critical role as well as during moleculardynamics simulations The program is freely available at httpsgithubcomekrakaEDHB
INTRODUCTIONHydrogen bonds (HBs) play a crucial role in proteins forgingenzyme reactions protein folding proteinminusligand interactionsand other processes1minus3 The early work of Pauling and Mirskyhighlighted already in 1936 the importance of HBs in proteinssuggesting a 5 kcalmol stability from each HB4 In 1951 theydiscovered α-helix and β-pleated sheet as the most importantconformational elements in a protein with α-helix HBs on theorder of 8 kcalmol56 Later that same year they realized thatan aqueous environment can lead to HBs of the order of 2kcalmol6 After almost 7 decades of intensive research thereare still some controversies about the strength and propertiesof HBs in a protein Theoretical studies in the early 90s suggestthat HBs do not contribute to thermodynamic stability as theoverall energy balance of HBs is close to zero7 Campos et al8
suggested with a combined experimental and theoretical studythat HBs in fact destabilize the native conformation of aprotein Recent studies have provided a better picture of HBsin proteins although controversies about their strength stillpersist9
The protein environment adds complexity to HBsrsquo stabilityand thus inclusion of all atoms is critical Studies conducted onfragments of a protein structure10 are limited by this crucialinformation Pace et al311 have concluded that the polarresidues buried inside the protein interior undergo strongervan der Waals forces in a HB Further studies by Hubbard andKamran Haider1 have shown that the buried intramolecular
HBs are stronger due to their electrostatic nature The proteinenvironment can induce an electric field on the order of 100MVcm12 Thus to incorporate the protein environment weutilized all atoms of a protein incorporating the complexenvironmentA HB is influenced by a multitude of factors governing its
stability Intramolecular HBs are one such factor that plays acrucial role113minus16 This fact was already highlighted by Takanoet al17 in the 90s Donorminusacceptor pairs such as NminusHmiddotmiddotmiddotONminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO lead to HBs of differentstrengths due to the involvement of atoms with differentelectronegativities1018 Another important aspect is thenetwork of HBs1920 where two or more HBs share the sameacceptor2122 A study by Ballesteros et al23 shows theimplications of networks in guiding secondary structures inproteins A study by Feldblum and Arkin24 clearly showed theexistence of various networks of HBs via both experimentaland computational means The influence of the proteinenvironment and the various factors alter the strength of a
Received December 22 2020Revised February 21 2021Published March 5 2021
ArticlepubsacsorgJPCB
copy 2021 American Chemical Society2551
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
Dow
nloa
ded
via
SOU
TH
ER
N M
ET
HO
DIS
T U
NIV
on
Apr
il 17
202
1 at
16
205
6 (U
TC
)Se
e ht
tps
pub
sac
sor
gsh
arin
ggui
delin
es f
or o
ptio
ns o
n ho
w to
legi
timat
ely
shar
e pu
blis
hed
artic
les
HB and a critical assessment of its strength would bolster ourunderstanding of HBs in a proteinThe strength of a chemical bond or weak chemical
interaction such as hydrogen bonding has frequently beenassessed via bond dissociation energies (BDE)s1025minus28 bondlengths29 or bond densities30 However these measures arenot directly related to the intrinsic bond strength as theydepend on other quantities the bond dissociation energy onthe stabilization energies of the fragments the bond length onthe compressibility limit distance between the atoms and thebond-stretching frequency on the atom masses31 FurthermoreBDEs are limited to intermolecular HBs and are not accessiblefor intramolecular HBs which play an important role inproteins Vibrational spectroscopy can serve in this situation asa perfect alternative provided we utilize local vibrational modes(LVM) which were originally introduced by Konkoli andCremer32 A comprehensive review is provided by Kraka etal33 LVMs have turned out to be an excellent tool forquantifying the strength of any chemical bond or weakchemical interaction including HBs1934minus44
To systematically analyze the influencing factors of a HB wedeveloped a python-based program called Ef f icient Detection ofHydrogen Bonding (EDHB) EDHB can efficiently detect allHBs in a protein based on the geometry (nearest neighborsearch45) and in addition classify them on the basis ofsecondary structure information atom types involved intra-molecular ring size and network of HBs Furthermore EDHBcan be optionally utilized to explore and analyze HB propertiessuch as LVMs and electrostaticcovalent contributions if dataare available from a preceding quantum chemical calculationWe note that the CminusH group acts as a HB donor in
proteins46 Weak polarization in a CminusH bond leads the Hatom to be slightly positively charged creating possibilities forHB interactions However as shown by Hubbard and KamranHaider1 CminusH-based HBs have only a minor contribution tothe overall stability of a protein structure Likewise NminusHmiddotmiddotmiddotSHBs also show minor implications for the protein structureTherefore in this work we focused on strong HBs in proteinsinvolving N and O as the donor or acceptor IntramolecularHBs involving five-membered rings for NminusHmiddotmiddotmiddotO CminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotN type interactions are termed ldquounconventionalrdquo
HBs47minus49 In this work only conventional HBs wereconsideredThe paper is organized in the following way First we
describe the architecture of EDHB and the computationaltools utilized for the study The information collected byEDHB is presented and discussed in the Results andDiscussion section Conclusions and a future outlook areprovided in the Conclusions section
METHODOLOGY
Architecture of EDHB Figure 1 shows the architecture ofEDHB Based on the geometry of a protein given in atomiccoordinates EDHB forms a K-dimension tree (K-D tree45)from the coordinates of each hydrogen For each HB donor(N F and O) the nearest hydrogen is searched which ischaracterized on the basis of the distance and angle For largersystems (no of atoms gt50 000) the computation is done inparallel to reduce the computational time A graph is formedand saved as a tree structure from which information such asHB type (a network of HBs) and the intramolecular ring size isextracted Optionally if data from quantum calculations areavailable from an NBO analysis and second energy derivativesEDHB classifies orbital overlap energy (covalent character)electrostatic energies and local-mode force constants (ameasure of HB strength)
Detection of Hydrogen Bonds HBs are detected byEDHB on the basis of geometry Only three atoms involved asa donor (D)acceptor (A) pair are considered so far ienitrogen oxygen and fluorine Various references have usedvarious criteria for HB length and angle150minus52 Based onliterature data we constrained the distance between thehydrogen (H) and the A atom to 16minus24 Aring (Figure 2) TheDminusH middotmiddotmiddotA angle is constrained to a range of 90minus180deg The K-Dtree algorithm45 was implemented to identify all possible HBsThe K-D tree is a space-partitioning data structure where spaceis divided into nonoverlapping regions until each point liesexactly in one of the regions The K-D tree works like a binarytree45 where every node is a k-dimensional point The lastnode in each node is the leaf node which contains a pointEvery other node generates two planes that divide the space
Figure 1 Architecture of EDHB EDHB utilizes geometry information to construct a K-D tree that captures all hydrogen bonds and their nature interms of type and inter- and intramolecular hydrogen bonds Optionally quantum calculations can be utilized to characterize covalent electrostaticand local-mode properties
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2552
into two parts where in the left node will be all points lesserthan and on the right will be points greater than the actualvalue of the division plane For example (for 3D data) if theldquoxrdquo axis is chosen for a particular split all points with a smallerx value will appear on the left node whereas the larger x valueswill appear on the right node The same procedure is appliedto the y axis and then to the z axis This process is thenrepeated recursively until the leaf node is reached for everypoint A representation for a two-dimensional (2D) datastructure and its decomposition as a K-D tree is shown inFigure 3 The first split starts at (0 0) and splits the x axis
(Note the axis and splitting point are chosen by the ldquoslidingmidpointrdquo rule which ensures that the cells do not all becomelong and thin (0 0) is chosen here for simplicity) All pointswith negative x values lie to the left of the node and all pointswith positive x values to the right A recursive split isperformed for y followed by x and so on until the leaf node isreached Utilizing this method all HBs of a protein can bedetected with at most O(kmiddotlog(n)) time complexity where k isthe number of electronegative atoms (O N or F) and n is thenumber of hydrogen atoms
Computational Time for EDHB To validate the perform-ance of the K-D tree we compared the results with classicalmethods of detecting HBs using naive nested loop andvectorized approaches The nested loop is formed by runningthree nested loops one for the donor one for the hydrogenand one for the acceptor The vectorized approach isimplemented and utilized by most of the currently availablesoftware that detects HBs such as MDTraj54 Table 1 shows acomparison of the timings between the nested loop vectorizedand K-D tree approach (an exhaustive comparison of 50different proteins is provided as Table S2) All computationswere run on an Intel(R) Xeon(R) CPU E5-2680 v4 240GHz with a 64-bit op-mode Geometry information for theproteins investigated was taken from the Protein Data Bank53
for proteins characterized by NMR solution Proteins that tookmore than 30 min for the nested approach were not consideredand are shown by ldquominusrdquo For a fair comparison the K-D treevectorized and nested approachs all were not parallelizedWhen comparing the timings for even a small protein such
as 1UAO with just 138 atoms the K-D tree is almost 1000times faster compared to nested and 4 times faster comparedto vectorized respectively As the number of atoms increasesthe performance of the nested approach highly degrades Onthe other hand the K-D tree is able to perform the samecomputation in less than a second For the protein with PDBID 2BI5 (2189 atoms) the K-D tree approach is almost 106
times faster than the nested approach For proteins as large as52 000 atoms the K-D tree is able to perform the computationin just 02 s compared to the vectorized method which takes309 s (1755 times faster)
Intramolecular Hydrogen Bond Search The intra-molecular analysis is performed via breadth first search55
(BFS) in EDHB BFS is an algorithm searching path in anetwork of connections The algorithm first traverses to all theneighbors in the first layer (Figure 4) while keeping track ofpath and history via backtracking significantly reducing thecomputational time The process is repeated recursively untilthe goal state is foundAs an example Figure 4 shows the representation of the
seven-membered intramolecular hydrogen-bonded (NminusHmiddotmiddotmiddotOtype) ring To calculate the size of the ring the initial and thegoal state are provided ie O11 and H1 respectively BFSsearches for the H1 layer by layer and when the goal state isreached it returns the path (O11minusC10minusN9minusC6minusC3minusN2minusH1) from which the ring size can be computed
Figure 2 Criteria for the identification of hydrogen bonds Thedonor acceptor can be [N O F] The angle between donorminushydrogenminusacceptor is constrained to a range of 90minus180deg
Figure 3 Iterative division of a 2D space to represent a K-D tree Theplots shown in the box are points in the 2D data structure The K-Dtree is shown (right) which is formed by alternative iterative divisionof the axis in the data
Table 1 Timing Comparison of EDHB
PDB ID nested (s) vectorized (s) KD tree (s) HBs Natoms ratio (nested) ratio (vectorized)
1UAO 047 0002 0000 6 138 1 177 41HD6 2347 0020 0001 39 526 16 552 142LDZ 36451 0079 0006 54 977 64 801 142EOT 26794 0105 0003 48 1 205 82 847 332M20 126926 0294 0005 96 2 046 247 221 572BL5 181729 0379 0006 149 2 189 295 929 685JTN minus 12602 0033 696 11 068 minus 3782KU1 minus 71505 0080 1 869 25 935 minus 8944BY9 minus 308782 0176 3 877 52 476 minus 1755
aThe protein geometries were taken from the Protein Data Bank (PDB)53 Nested vectorized and K-D-tree refer to the time taken by nestedvectorized and K-D-tree approaches respectively HBs refer to the number of HBs detected Natoms refers to the number of atoms in the proteinRatio refers to the ratio of time taken by the nestedvectorized approach to the K-D-tree approach
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2553
Hydrogen Bond Network Classification The strengthof a HB is influenced by the presenceabsence of other HBsthat share the same D or A20 Figure 5 shows a hydrogen-
bonding network The type of HB network can be defined bythe coordination number of the D A or hydrogen(s) from theD as a string
equiv minus minusHB A D DHtype c c c (1)
where Ac Dc and DHc represent the number of HBs from Aexcluding targeted HB number of HB(s) from D and numberof HB(s) from the covalently bonded hydrogen(s) to Drespectively Following this procedure the HB network typeshown in Figure 5 is 1minus2minus1Hydrogen Bond Characterization EDHB characterizes
HBs based on two properties vibrational spectroscopy andNBO analysis data Vibrational spectroscopy data are utilizedwhen the second energy derivative for a molecule is availableto calculate local-mode force constants ka obtained from thepure local stretching vibration of a HB The resultant forceconstants provide the strength of a HB The electrostatic andcovalent contributions are estimated based on results obtainedfrom the NBO analysisLocal-Mode Force Constants The vibrational spectra of
a molecule hold enormous information about the structure andproperties of the molecular interactions We utilize vibrationalspectra to obtain the relative strengths of HBs We firstperform a normal-mode analysis by the basic equation ofvibrational spectroscopy formulated as
Λ=f L MLx (2)
where L collects the vibrational eigenvectors Iμ and thediagonal eigenvalue matrix Λ contains Nvib vibrationaleigenvalues 4π2c2νμ
2 (with μ = 1 Nvib and Nvib = 3N minus KK = 6 for nonlinear molecules and 5 for linear molecules) andK zero eigenvalues corresponding to translations and rotationsof the molecule (νμ represents the harmonic vibrationalfrequencies given in cmminus1 and c is the speed of light) M is the
diagonal mass matrix and fx is the force constant matrixexpressed in Cartesian coordinates The (3N times 3N) dimen-sional L matrix collects the Nvib normal-vibrational-modevectors Iμ and K-mode vectors corresponding to translationsand rotationsExpressing eq 2 in internal coordinates q leads to the Wilson
GF formalism with5657
Λ= minusF D G Dq 1 (3)
where
= minus daggerG BM B1 (4)
=D BL (5)
Fq is the force constant matrix expressed in terms of internalcoordinates q The (Nvib times Nvib) dimensional D matrixrepresents each normal-mode vector dμ The rectangular (Nvibtimes 3N) dimensional B matrix contains the first derivatives ofthe internal coordinates with regard to the Cartesiancoordinates thus connecting both coordinate systems andthe (Nvib times Nvib) dimensional matrix G is the Wilson Gmatrix5657
In terms of internal coordinates the Lagrange equations canbe formulated as
= minusdagger minus daggerL q q q G q q F q( )12
12
q1(6)
where decoupling of the second part (potential energy) is doneby the diagonalization of Fq However the first part (kineticenergy) remains coupled due to the matrix G Konkoli andCremer32 solved the problem of mass coupling by introducingthe concept of local vibrational modes The mass-decoupledEulerminusLagrange equation was solved by setting all the atomicmasses to zero except those of the molecular fragment (egbond angle or dihedral etc) carrying out a localized vibrationAs shown in their original work the change in the localdisplacement of a specific internal coordinate is equivalent toan adiabatic relaxation of the molecule32 For any molecularfragment associated with an internal coordinate qμ thecorresponding local mode vector aμ is given by
=μμ
μ μ
minus dagger
minus daggeraK d
d K d
1
1(7)
where dμ is a row vector of matrix D Matrix K is the diagonalmatrix of force constants kQ expressed in normal coordinatesQμ with
= = daggerF K L f LxQ (8)
resulting from the Wilson GF formalism shown in eqs 3 and 4The local mode force constant kμ
a corresponding to localmode aμ is obtained by
=μ μ μdaggerk a Kaa
(9)
Local vibrational modes have been successfully applied toquantify weak chemical interactions such as hydrogenbonding1934minus38 halogen bonding39minus41 pnicogen bond-ing42minus44 chalcogen bonding58 and tetrel bonding59 and toderive new chemical descriptors such as a new aromaticityindex60minus62 or a generalized Tolman electronic parameter63minus65
as well as for the derivation of a generalized Badger Rule66 andseveral others new concepts66minus73
Figure 4 Representation of a breadth first search (BFS) On the left isa seven-membered intramolecular hydrogen bonding On the right isshown the process of how BFS traverses layer by layer in a network ofconnections to reach the goal state (H1) from the initial state (O11)
Figure 5 Network of a HB The DminusHmiddotmiddotmiddotA shown in red is thetargeted HB The dashed line in blue represents possible HBs near thetargeted HB
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2554
Covalent Characteristics of Hydrogen Bonds Thecovalent contribution to the HBs is reinforced by the twoelectron exchange integrals a phenomenon specific toelectrons where they are assumed to be at multiple places atthe same time These contributions stabilize the overallmolecular energies leading to stronger bonds with increasingcovalent contributions We utilized NBO analysis to obtain theoverlap energy of the oxygen lone pair np with the emptyantibonding σ orbital of donor hydrogen bond (LPtoBD)shown in Figure 6 This results in a concomitant polarization
of the donor hydrogen bond which is a direct consequence ofBentrsquos rule74 and can be observed in various bonds such as CminusH SiminusH SminusH NminusH etcElectrostatic Characteristics of Hydrogen Bonds The
electrostatic interaction is caused by the induction of chargesin the molecules and can be relatively estimated for a HB as
=
Eq q
rc1 2
(10)
where Ec is the electrostatic energy q1 and q2 are the atomiccharges of A and H respectively and r is the distance betweenthe two atoms75 We choose NBO charges as they are morereliable than Mulliken charges76 since they operate on the basisof electron density Localized natural atomic orbitals were usedto describe the computed electron densities and thereforeinclude bond polarizationComputational Details Geometry optimization and
normal-mode analysis were carried out in the gas phaseusing Gaussian 167778 Eight smaller proteins 1K43 1L2Y1R4G 1S4A 2EVQ 1UAO 2O0S and 5E61 shown in Figure7 were optimized with the hybrid B3LYP functional79minus82 Highaccuracy paired with relatively low computational costs hasmade B3LYP a popular DFT choice Kuhn et al83 and Bartlettet al84 successfully used this functional for the description of
small molecules and peptides We further employed empiricaldispersion correction8586 as suggested by Fadda et al87 toinclude polarization effects that can significantly impacthydrogen bond stability Based on the findings of Fadda etal87 that B3LYP paired with the 6-31G(dp) basis88minus94
provides a good description of short alanine peptides wealso used the 6-31G(dp) in our study For all geometryoptimizations conversion criteria to a maximum step size of001 au and a RMS force of 00017 au were applied Protein1R4G was cut and only the area of major interest was takenSome side chains were removed from proteins 1K43 1S4Aand 5E61 for faster calculation (details and geometryinformation are provided in the Supporting Information aspages S23minusS59)To obtain statistically relevant information in addition a set
of 155 smaller proteins was optimized and second energyderivatives were calculated with the semiempirical DFTmethod GFN2-xTB95 The GFN2-xTB is significantly fasterthan B3LYP with a low compromise regarding accuracy Asimilar distribution of second-order properties such asvibrational constants for GFN2-xTB compared to B3LYPwas observed which is described in the Results and Discussionsection The similarity between the results of the two methodsled to our choice of GFN2-xTB as the DFT method for thestatistical analysis The 155 proteins were selected based on thesmall size single chain and NMR solution as characterizingtechniques from the Protein Data Bank53 Hydrogen atomswere added utilizing Leap (a tool from Amber96) structureswere minimized for 1000 steps in Amber96 and the overallcharges were calculated from Leap prior to the GFN2-xTBcalculations The PDB IDs for all 155 proteins are provided inthe Supporting Information on page S2The local-mode force constants for each HB were calculated
using the LModeA package3397 A description of thecorresponding local vibrational mode theory is given belowLPtoBD and atomic charges were obtained from NBO98
analysis utilizing optimized geometries with the B3LYP79minus82
level of theory including empirical dispersion correction8586
along with the 6-31G(dp)88minus94 basis set
RESULTS AND DISCUSSIONThis section is organized into subsections providing results anddiscussion by analyzing patterns for a general overview of HBproperties intramolecular HBs HB type based on donorminusacceptor pairs networks of HBs and the governing factors forthe HB strength in proteins We start by comparing predictedHB properties from the two DFT methods to get statisticallyrelevant information We observed similar patterns of HBproperties from the two DFT methods Thus we provide adetailed characterization of HB properties from a higher levelof DFT method (B3LYP) followed by statistical results from alower level of DFT method (GFN2-xTB)Figure 7 shows the 8 proteins investigated with B3LYP and
the PDB IDs for 155 proteins investigated with GFN2-xTB areprovided on pages S2minusS10 The HBs detected by EDHB areshown in yellow Each HB is given a specific ID for discussion(larger images of proteins are shown in the SupportingInformation as Figures S1minusS8) These proteins accommodatea variety of secondary structures which include a majority ofHB types observed in proteinsTo generalize the outcomes from GFN2-xTB and B3LYP
we focused on the distribution of HB length and ka for theeight proteins optimized as shown in Figure 8 The HBs that
Figure 6 Overlap of the lone pair np of the acceptor to the σ of thedonor hydrogen bond The overlap energy estimates the covalentcharacteristics of a hydrogen bond
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2555
were absent in either one of the methods were removed TheGFN2-xTB and B3LYP methods differ and therefore theoptimized geometries are not exactly the same This is reflectedby Figure 8b where the median of the HB length for B3LYP(195 Aring) is slightly higher than that for GFN2-xTB (192 Aring)However the overall distribution remains the same and is alsoobserved for ka as shown in Figure 8a The comparison ofLPtoBD HB angle and electrostatic energy is shown inFigure S9 where we observe similar distributions Based onthese results we conclude that statistically relevant informationcan be obtained from the GFN2-xTB method
We analyzed quantitatively the number of different HBspresent in 155 proteins based on secondary structures shownin Table 2 Note that EDHB uses data obtained fromDictionary of Protein Secondary Structure (DSSP)99 forsecondary structure classification if the PDB ID of the proteinis available The proteins circumvent a variety of secondarystructures with α turn bend and extended strand composing204 93 86 and 67 respectively Less common HBs suchas 310 helix and isolated β-bridge are 22 and 07 respectivelyThe secondary structures with no backbone classification are
Figure 7 Eight proteins investigated Each number is shown as a unique ID associated with each HB (a) 1K43 is a beta hairpin type structure andcontains 13 HBs (1minus13) (b) A trp-cage motif 1L2Y has α helix and 19 HBs (14minus32) (c) The Sendai virus protein 1R4G was cut to keep the idealα-helix Other atoms were removed for the convenience of running ab initio calculations Twenty-eight HBs were detected for this protein (33minus60) (d) A double-strand β-helix of a DL-alternating oligonorleucine 1S4A has 18 HBs (61minus78) (e) A designed protein Chignolin 1UAO is a β-sheet type structure and looks more like a hairpin with 10 HBs (79minus88) (f) A 12-residue beta hairpin 2EVQ with 15 HBs (89minus103) (g) Adesigned peptide 2O0S is a coil type structure and has 7 HBs (104minus110) and the amyloid-forming peptide 5E61 with a near ideal β-strand has 13HBs (111minus123)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2556
termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

HB and a critical assessment of its strength would bolster ourunderstanding of HBs in a proteinThe strength of a chemical bond or weak chemical
interaction such as hydrogen bonding has frequently beenassessed via bond dissociation energies (BDE)s1025minus28 bondlengths29 or bond densities30 However these measures arenot directly related to the intrinsic bond strength as theydepend on other quantities the bond dissociation energy onthe stabilization energies of the fragments the bond length onthe compressibility limit distance between the atoms and thebond-stretching frequency on the atom masses31 FurthermoreBDEs are limited to intermolecular HBs and are not accessiblefor intramolecular HBs which play an important role inproteins Vibrational spectroscopy can serve in this situation asa perfect alternative provided we utilize local vibrational modes(LVM) which were originally introduced by Konkoli andCremer32 A comprehensive review is provided by Kraka etal33 LVMs have turned out to be an excellent tool forquantifying the strength of any chemical bond or weakchemical interaction including HBs1934minus44
To systematically analyze the influencing factors of a HB wedeveloped a python-based program called Ef f icient Detection ofHydrogen Bonding (EDHB) EDHB can efficiently detect allHBs in a protein based on the geometry (nearest neighborsearch45) and in addition classify them on the basis ofsecondary structure information atom types involved intra-molecular ring size and network of HBs Furthermore EDHBcan be optionally utilized to explore and analyze HB propertiessuch as LVMs and electrostaticcovalent contributions if dataare available from a preceding quantum chemical calculationWe note that the CminusH group acts as a HB donor in
proteins46 Weak polarization in a CminusH bond leads the Hatom to be slightly positively charged creating possibilities forHB interactions However as shown by Hubbard and KamranHaider1 CminusH-based HBs have only a minor contribution tothe overall stability of a protein structure Likewise NminusHmiddotmiddotmiddotSHBs also show minor implications for the protein structureTherefore in this work we focused on strong HBs in proteinsinvolving N and O as the donor or acceptor IntramolecularHBs involving five-membered rings for NminusHmiddotmiddotmiddotO CminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotN type interactions are termed ldquounconventionalrdquo
HBs47minus49 In this work only conventional HBs wereconsideredThe paper is organized in the following way First we
describe the architecture of EDHB and the computationaltools utilized for the study The information collected byEDHB is presented and discussed in the Results andDiscussion section Conclusions and a future outlook areprovided in the Conclusions section
METHODOLOGY
Architecture of EDHB Figure 1 shows the architecture ofEDHB Based on the geometry of a protein given in atomiccoordinates EDHB forms a K-dimension tree (K-D tree45)from the coordinates of each hydrogen For each HB donor(N F and O) the nearest hydrogen is searched which ischaracterized on the basis of the distance and angle For largersystems (no of atoms gt50 000) the computation is done inparallel to reduce the computational time A graph is formedand saved as a tree structure from which information such asHB type (a network of HBs) and the intramolecular ring size isextracted Optionally if data from quantum calculations areavailable from an NBO analysis and second energy derivativesEDHB classifies orbital overlap energy (covalent character)electrostatic energies and local-mode force constants (ameasure of HB strength)
Detection of Hydrogen Bonds HBs are detected byEDHB on the basis of geometry Only three atoms involved asa donor (D)acceptor (A) pair are considered so far ienitrogen oxygen and fluorine Various references have usedvarious criteria for HB length and angle150minus52 Based onliterature data we constrained the distance between thehydrogen (H) and the A atom to 16minus24 Aring (Figure 2) TheDminusH middotmiddotmiddotA angle is constrained to a range of 90minus180deg The K-Dtree algorithm45 was implemented to identify all possible HBsThe K-D tree is a space-partitioning data structure where spaceis divided into nonoverlapping regions until each point liesexactly in one of the regions The K-D tree works like a binarytree45 where every node is a k-dimensional point The lastnode in each node is the leaf node which contains a pointEvery other node generates two planes that divide the space
Figure 1 Architecture of EDHB EDHB utilizes geometry information to construct a K-D tree that captures all hydrogen bonds and their nature interms of type and inter- and intramolecular hydrogen bonds Optionally quantum calculations can be utilized to characterize covalent electrostaticand local-mode properties
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2552
into two parts where in the left node will be all points lesserthan and on the right will be points greater than the actualvalue of the division plane For example (for 3D data) if theldquoxrdquo axis is chosen for a particular split all points with a smallerx value will appear on the left node whereas the larger x valueswill appear on the right node The same procedure is appliedto the y axis and then to the z axis This process is thenrepeated recursively until the leaf node is reached for everypoint A representation for a two-dimensional (2D) datastructure and its decomposition as a K-D tree is shown inFigure 3 The first split starts at (0 0) and splits the x axis
(Note the axis and splitting point are chosen by the ldquoslidingmidpointrdquo rule which ensures that the cells do not all becomelong and thin (0 0) is chosen here for simplicity) All pointswith negative x values lie to the left of the node and all pointswith positive x values to the right A recursive split isperformed for y followed by x and so on until the leaf node isreached Utilizing this method all HBs of a protein can bedetected with at most O(kmiddotlog(n)) time complexity where k isthe number of electronegative atoms (O N or F) and n is thenumber of hydrogen atoms
Computational Time for EDHB To validate the perform-ance of the K-D tree we compared the results with classicalmethods of detecting HBs using naive nested loop andvectorized approaches The nested loop is formed by runningthree nested loops one for the donor one for the hydrogenand one for the acceptor The vectorized approach isimplemented and utilized by most of the currently availablesoftware that detects HBs such as MDTraj54 Table 1 shows acomparison of the timings between the nested loop vectorizedand K-D tree approach (an exhaustive comparison of 50different proteins is provided as Table S2) All computationswere run on an Intel(R) Xeon(R) CPU E5-2680 v4 240GHz with a 64-bit op-mode Geometry information for theproteins investigated was taken from the Protein Data Bank53
for proteins characterized by NMR solution Proteins that tookmore than 30 min for the nested approach were not consideredand are shown by ldquominusrdquo For a fair comparison the K-D treevectorized and nested approachs all were not parallelizedWhen comparing the timings for even a small protein such
as 1UAO with just 138 atoms the K-D tree is almost 1000times faster compared to nested and 4 times faster comparedto vectorized respectively As the number of atoms increasesthe performance of the nested approach highly degrades Onthe other hand the K-D tree is able to perform the samecomputation in less than a second For the protein with PDBID 2BI5 (2189 atoms) the K-D tree approach is almost 106
times faster than the nested approach For proteins as large as52 000 atoms the K-D tree is able to perform the computationin just 02 s compared to the vectorized method which takes309 s (1755 times faster)
Intramolecular Hydrogen Bond Search The intra-molecular analysis is performed via breadth first search55
(BFS) in EDHB BFS is an algorithm searching path in anetwork of connections The algorithm first traverses to all theneighbors in the first layer (Figure 4) while keeping track ofpath and history via backtracking significantly reducing thecomputational time The process is repeated recursively untilthe goal state is foundAs an example Figure 4 shows the representation of the
seven-membered intramolecular hydrogen-bonded (NminusHmiddotmiddotmiddotOtype) ring To calculate the size of the ring the initial and thegoal state are provided ie O11 and H1 respectively BFSsearches for the H1 layer by layer and when the goal state isreached it returns the path (O11minusC10minusN9minusC6minusC3minusN2minusH1) from which the ring size can be computed
Figure 2 Criteria for the identification of hydrogen bonds Thedonor acceptor can be [N O F] The angle between donorminushydrogenminusacceptor is constrained to a range of 90minus180deg
Figure 3 Iterative division of a 2D space to represent a K-D tree Theplots shown in the box are points in the 2D data structure The K-Dtree is shown (right) which is formed by alternative iterative divisionof the axis in the data
Table 1 Timing Comparison of EDHB
PDB ID nested (s) vectorized (s) KD tree (s) HBs Natoms ratio (nested) ratio (vectorized)
1UAO 047 0002 0000 6 138 1 177 41HD6 2347 0020 0001 39 526 16 552 142LDZ 36451 0079 0006 54 977 64 801 142EOT 26794 0105 0003 48 1 205 82 847 332M20 126926 0294 0005 96 2 046 247 221 572BL5 181729 0379 0006 149 2 189 295 929 685JTN minus 12602 0033 696 11 068 minus 3782KU1 minus 71505 0080 1 869 25 935 minus 8944BY9 minus 308782 0176 3 877 52 476 minus 1755
aThe protein geometries were taken from the Protein Data Bank (PDB)53 Nested vectorized and K-D-tree refer to the time taken by nestedvectorized and K-D-tree approaches respectively HBs refer to the number of HBs detected Natoms refers to the number of atoms in the proteinRatio refers to the ratio of time taken by the nestedvectorized approach to the K-D-tree approach
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2553
Hydrogen Bond Network Classification The strengthof a HB is influenced by the presenceabsence of other HBsthat share the same D or A20 Figure 5 shows a hydrogen-
bonding network The type of HB network can be defined bythe coordination number of the D A or hydrogen(s) from theD as a string
equiv minus minusHB A D DHtype c c c (1)
where Ac Dc and DHc represent the number of HBs from Aexcluding targeted HB number of HB(s) from D and numberof HB(s) from the covalently bonded hydrogen(s) to Drespectively Following this procedure the HB network typeshown in Figure 5 is 1minus2minus1Hydrogen Bond Characterization EDHB characterizes
HBs based on two properties vibrational spectroscopy andNBO analysis data Vibrational spectroscopy data are utilizedwhen the second energy derivative for a molecule is availableto calculate local-mode force constants ka obtained from thepure local stretching vibration of a HB The resultant forceconstants provide the strength of a HB The electrostatic andcovalent contributions are estimated based on results obtainedfrom the NBO analysisLocal-Mode Force Constants The vibrational spectra of
a molecule hold enormous information about the structure andproperties of the molecular interactions We utilize vibrationalspectra to obtain the relative strengths of HBs We firstperform a normal-mode analysis by the basic equation ofvibrational spectroscopy formulated as
Λ=f L MLx (2)
where L collects the vibrational eigenvectors Iμ and thediagonal eigenvalue matrix Λ contains Nvib vibrationaleigenvalues 4π2c2νμ
2 (with μ = 1 Nvib and Nvib = 3N minus KK = 6 for nonlinear molecules and 5 for linear molecules) andK zero eigenvalues corresponding to translations and rotationsof the molecule (νμ represents the harmonic vibrationalfrequencies given in cmminus1 and c is the speed of light) M is the
diagonal mass matrix and fx is the force constant matrixexpressed in Cartesian coordinates The (3N times 3N) dimen-sional L matrix collects the Nvib normal-vibrational-modevectors Iμ and K-mode vectors corresponding to translationsand rotationsExpressing eq 2 in internal coordinates q leads to the Wilson
GF formalism with5657
Λ= minusF D G Dq 1 (3)
where
= minus daggerG BM B1 (4)
=D BL (5)
Fq is the force constant matrix expressed in terms of internalcoordinates q The (Nvib times Nvib) dimensional D matrixrepresents each normal-mode vector dμ The rectangular (Nvibtimes 3N) dimensional B matrix contains the first derivatives ofthe internal coordinates with regard to the Cartesiancoordinates thus connecting both coordinate systems andthe (Nvib times Nvib) dimensional matrix G is the Wilson Gmatrix5657
In terms of internal coordinates the Lagrange equations canbe formulated as
= minusdagger minus daggerL q q q G q q F q( )12
12
q1(6)
where decoupling of the second part (potential energy) is doneby the diagonalization of Fq However the first part (kineticenergy) remains coupled due to the matrix G Konkoli andCremer32 solved the problem of mass coupling by introducingthe concept of local vibrational modes The mass-decoupledEulerminusLagrange equation was solved by setting all the atomicmasses to zero except those of the molecular fragment (egbond angle or dihedral etc) carrying out a localized vibrationAs shown in their original work the change in the localdisplacement of a specific internal coordinate is equivalent toan adiabatic relaxation of the molecule32 For any molecularfragment associated with an internal coordinate qμ thecorresponding local mode vector aμ is given by
=μμ
μ μ
minus dagger
minus daggeraK d
d K d
1
1(7)
where dμ is a row vector of matrix D Matrix K is the diagonalmatrix of force constants kQ expressed in normal coordinatesQμ with
= = daggerF K L f LxQ (8)
resulting from the Wilson GF formalism shown in eqs 3 and 4The local mode force constant kμ
a corresponding to localmode aμ is obtained by
=μ μ μdaggerk a Kaa
(9)
Local vibrational modes have been successfully applied toquantify weak chemical interactions such as hydrogenbonding1934minus38 halogen bonding39minus41 pnicogen bond-ing42minus44 chalcogen bonding58 and tetrel bonding59 and toderive new chemical descriptors such as a new aromaticityindex60minus62 or a generalized Tolman electronic parameter63minus65
as well as for the derivation of a generalized Badger Rule66 andseveral others new concepts66minus73
Figure 4 Representation of a breadth first search (BFS) On the left isa seven-membered intramolecular hydrogen bonding On the right isshown the process of how BFS traverses layer by layer in a network ofconnections to reach the goal state (H1) from the initial state (O11)
Figure 5 Network of a HB The DminusHmiddotmiddotmiddotA shown in red is thetargeted HB The dashed line in blue represents possible HBs near thetargeted HB
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2554
Covalent Characteristics of Hydrogen Bonds Thecovalent contribution to the HBs is reinforced by the twoelectron exchange integrals a phenomenon specific toelectrons where they are assumed to be at multiple places atthe same time These contributions stabilize the overallmolecular energies leading to stronger bonds with increasingcovalent contributions We utilized NBO analysis to obtain theoverlap energy of the oxygen lone pair np with the emptyantibonding σ orbital of donor hydrogen bond (LPtoBD)shown in Figure 6 This results in a concomitant polarization
of the donor hydrogen bond which is a direct consequence ofBentrsquos rule74 and can be observed in various bonds such as CminusH SiminusH SminusH NminusH etcElectrostatic Characteristics of Hydrogen Bonds The
electrostatic interaction is caused by the induction of chargesin the molecules and can be relatively estimated for a HB as
=
Eq q
rc1 2
(10)
where Ec is the electrostatic energy q1 and q2 are the atomiccharges of A and H respectively and r is the distance betweenthe two atoms75 We choose NBO charges as they are morereliable than Mulliken charges76 since they operate on the basisof electron density Localized natural atomic orbitals were usedto describe the computed electron densities and thereforeinclude bond polarizationComputational Details Geometry optimization and
normal-mode analysis were carried out in the gas phaseusing Gaussian 167778 Eight smaller proteins 1K43 1L2Y1R4G 1S4A 2EVQ 1UAO 2O0S and 5E61 shown in Figure7 were optimized with the hybrid B3LYP functional79minus82 Highaccuracy paired with relatively low computational costs hasmade B3LYP a popular DFT choice Kuhn et al83 and Bartlettet al84 successfully used this functional for the description of
small molecules and peptides We further employed empiricaldispersion correction8586 as suggested by Fadda et al87 toinclude polarization effects that can significantly impacthydrogen bond stability Based on the findings of Fadda etal87 that B3LYP paired with the 6-31G(dp) basis88minus94
provides a good description of short alanine peptides wealso used the 6-31G(dp) in our study For all geometryoptimizations conversion criteria to a maximum step size of001 au and a RMS force of 00017 au were applied Protein1R4G was cut and only the area of major interest was takenSome side chains were removed from proteins 1K43 1S4Aand 5E61 for faster calculation (details and geometryinformation are provided in the Supporting Information aspages S23minusS59)To obtain statistically relevant information in addition a set
of 155 smaller proteins was optimized and second energyderivatives were calculated with the semiempirical DFTmethod GFN2-xTB95 The GFN2-xTB is significantly fasterthan B3LYP with a low compromise regarding accuracy Asimilar distribution of second-order properties such asvibrational constants for GFN2-xTB compared to B3LYPwas observed which is described in the Results and Discussionsection The similarity between the results of the two methodsled to our choice of GFN2-xTB as the DFT method for thestatistical analysis The 155 proteins were selected based on thesmall size single chain and NMR solution as characterizingtechniques from the Protein Data Bank53 Hydrogen atomswere added utilizing Leap (a tool from Amber96) structureswere minimized for 1000 steps in Amber96 and the overallcharges were calculated from Leap prior to the GFN2-xTBcalculations The PDB IDs for all 155 proteins are provided inthe Supporting Information on page S2The local-mode force constants for each HB were calculated
using the LModeA package3397 A description of thecorresponding local vibrational mode theory is given belowLPtoBD and atomic charges were obtained from NBO98
analysis utilizing optimized geometries with the B3LYP79minus82
level of theory including empirical dispersion correction8586
along with the 6-31G(dp)88minus94 basis set
RESULTS AND DISCUSSIONThis section is organized into subsections providing results anddiscussion by analyzing patterns for a general overview of HBproperties intramolecular HBs HB type based on donorminusacceptor pairs networks of HBs and the governing factors forthe HB strength in proteins We start by comparing predictedHB properties from the two DFT methods to get statisticallyrelevant information We observed similar patterns of HBproperties from the two DFT methods Thus we provide adetailed characterization of HB properties from a higher levelof DFT method (B3LYP) followed by statistical results from alower level of DFT method (GFN2-xTB)Figure 7 shows the 8 proteins investigated with B3LYP and
the PDB IDs for 155 proteins investigated with GFN2-xTB areprovided on pages S2minusS10 The HBs detected by EDHB areshown in yellow Each HB is given a specific ID for discussion(larger images of proteins are shown in the SupportingInformation as Figures S1minusS8) These proteins accommodatea variety of secondary structures which include a majority ofHB types observed in proteinsTo generalize the outcomes from GFN2-xTB and B3LYP
we focused on the distribution of HB length and ka for theeight proteins optimized as shown in Figure 8 The HBs that
Figure 6 Overlap of the lone pair np of the acceptor to the σ of thedonor hydrogen bond The overlap energy estimates the covalentcharacteristics of a hydrogen bond
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2555
were absent in either one of the methods were removed TheGFN2-xTB and B3LYP methods differ and therefore theoptimized geometries are not exactly the same This is reflectedby Figure 8b where the median of the HB length for B3LYP(195 Aring) is slightly higher than that for GFN2-xTB (192 Aring)However the overall distribution remains the same and is alsoobserved for ka as shown in Figure 8a The comparison ofLPtoBD HB angle and electrostatic energy is shown inFigure S9 where we observe similar distributions Based onthese results we conclude that statistically relevant informationcan be obtained from the GFN2-xTB method
We analyzed quantitatively the number of different HBspresent in 155 proteins based on secondary structures shownin Table 2 Note that EDHB uses data obtained fromDictionary of Protein Secondary Structure (DSSP)99 forsecondary structure classification if the PDB ID of the proteinis available The proteins circumvent a variety of secondarystructures with α turn bend and extended strand composing204 93 86 and 67 respectively Less common HBs suchas 310 helix and isolated β-bridge are 22 and 07 respectivelyThe secondary structures with no backbone classification are
Figure 7 Eight proteins investigated Each number is shown as a unique ID associated with each HB (a) 1K43 is a beta hairpin type structure andcontains 13 HBs (1minus13) (b) A trp-cage motif 1L2Y has α helix and 19 HBs (14minus32) (c) The Sendai virus protein 1R4G was cut to keep the idealα-helix Other atoms were removed for the convenience of running ab initio calculations Twenty-eight HBs were detected for this protein (33minus60) (d) A double-strand β-helix of a DL-alternating oligonorleucine 1S4A has 18 HBs (61minus78) (e) A designed protein Chignolin 1UAO is a β-sheet type structure and looks more like a hairpin with 10 HBs (79minus88) (f) A 12-residue beta hairpin 2EVQ with 15 HBs (89minus103) (g) Adesigned peptide 2O0S is a coil type structure and has 7 HBs (104minus110) and the amyloid-forming peptide 5E61 with a near ideal β-strand has 13HBs (111minus123)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2556
termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

into two parts where in the left node will be all points lesserthan and on the right will be points greater than the actualvalue of the division plane For example (for 3D data) if theldquoxrdquo axis is chosen for a particular split all points with a smallerx value will appear on the left node whereas the larger x valueswill appear on the right node The same procedure is appliedto the y axis and then to the z axis This process is thenrepeated recursively until the leaf node is reached for everypoint A representation for a two-dimensional (2D) datastructure and its decomposition as a K-D tree is shown inFigure 3 The first split starts at (0 0) and splits the x axis
(Note the axis and splitting point are chosen by the ldquoslidingmidpointrdquo rule which ensures that the cells do not all becomelong and thin (0 0) is chosen here for simplicity) All pointswith negative x values lie to the left of the node and all pointswith positive x values to the right A recursive split isperformed for y followed by x and so on until the leaf node isreached Utilizing this method all HBs of a protein can bedetected with at most O(kmiddotlog(n)) time complexity where k isthe number of electronegative atoms (O N or F) and n is thenumber of hydrogen atoms
Computational Time for EDHB To validate the perform-ance of the K-D tree we compared the results with classicalmethods of detecting HBs using naive nested loop andvectorized approaches The nested loop is formed by runningthree nested loops one for the donor one for the hydrogenand one for the acceptor The vectorized approach isimplemented and utilized by most of the currently availablesoftware that detects HBs such as MDTraj54 Table 1 shows acomparison of the timings between the nested loop vectorizedand K-D tree approach (an exhaustive comparison of 50different proteins is provided as Table S2) All computationswere run on an Intel(R) Xeon(R) CPU E5-2680 v4 240GHz with a 64-bit op-mode Geometry information for theproteins investigated was taken from the Protein Data Bank53
for proteins characterized by NMR solution Proteins that tookmore than 30 min for the nested approach were not consideredand are shown by ldquominusrdquo For a fair comparison the K-D treevectorized and nested approachs all were not parallelizedWhen comparing the timings for even a small protein such
as 1UAO with just 138 atoms the K-D tree is almost 1000times faster compared to nested and 4 times faster comparedto vectorized respectively As the number of atoms increasesthe performance of the nested approach highly degrades Onthe other hand the K-D tree is able to perform the samecomputation in less than a second For the protein with PDBID 2BI5 (2189 atoms) the K-D tree approach is almost 106
times faster than the nested approach For proteins as large as52 000 atoms the K-D tree is able to perform the computationin just 02 s compared to the vectorized method which takes309 s (1755 times faster)
Intramolecular Hydrogen Bond Search The intra-molecular analysis is performed via breadth first search55
(BFS) in EDHB BFS is an algorithm searching path in anetwork of connections The algorithm first traverses to all theneighbors in the first layer (Figure 4) while keeping track ofpath and history via backtracking significantly reducing thecomputational time The process is repeated recursively untilthe goal state is foundAs an example Figure 4 shows the representation of the
seven-membered intramolecular hydrogen-bonded (NminusHmiddotmiddotmiddotOtype) ring To calculate the size of the ring the initial and thegoal state are provided ie O11 and H1 respectively BFSsearches for the H1 layer by layer and when the goal state isreached it returns the path (O11minusC10minusN9minusC6minusC3minusN2minusH1) from which the ring size can be computed
Figure 2 Criteria for the identification of hydrogen bonds Thedonor acceptor can be [N O F] The angle between donorminushydrogenminusacceptor is constrained to a range of 90minus180deg
Figure 3 Iterative division of a 2D space to represent a K-D tree Theplots shown in the box are points in the 2D data structure The K-Dtree is shown (right) which is formed by alternative iterative divisionof the axis in the data
Table 1 Timing Comparison of EDHB
PDB ID nested (s) vectorized (s) KD tree (s) HBs Natoms ratio (nested) ratio (vectorized)
1UAO 047 0002 0000 6 138 1 177 41HD6 2347 0020 0001 39 526 16 552 142LDZ 36451 0079 0006 54 977 64 801 142EOT 26794 0105 0003 48 1 205 82 847 332M20 126926 0294 0005 96 2 046 247 221 572BL5 181729 0379 0006 149 2 189 295 929 685JTN minus 12602 0033 696 11 068 minus 3782KU1 minus 71505 0080 1 869 25 935 minus 8944BY9 minus 308782 0176 3 877 52 476 minus 1755
aThe protein geometries were taken from the Protein Data Bank (PDB)53 Nested vectorized and K-D-tree refer to the time taken by nestedvectorized and K-D-tree approaches respectively HBs refer to the number of HBs detected Natoms refers to the number of atoms in the proteinRatio refers to the ratio of time taken by the nestedvectorized approach to the K-D-tree approach
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2553
Hydrogen Bond Network Classification The strengthof a HB is influenced by the presenceabsence of other HBsthat share the same D or A20 Figure 5 shows a hydrogen-
bonding network The type of HB network can be defined bythe coordination number of the D A or hydrogen(s) from theD as a string
equiv minus minusHB A D DHtype c c c (1)
where Ac Dc and DHc represent the number of HBs from Aexcluding targeted HB number of HB(s) from D and numberof HB(s) from the covalently bonded hydrogen(s) to Drespectively Following this procedure the HB network typeshown in Figure 5 is 1minus2minus1Hydrogen Bond Characterization EDHB characterizes
HBs based on two properties vibrational spectroscopy andNBO analysis data Vibrational spectroscopy data are utilizedwhen the second energy derivative for a molecule is availableto calculate local-mode force constants ka obtained from thepure local stretching vibration of a HB The resultant forceconstants provide the strength of a HB The electrostatic andcovalent contributions are estimated based on results obtainedfrom the NBO analysisLocal-Mode Force Constants The vibrational spectra of
a molecule hold enormous information about the structure andproperties of the molecular interactions We utilize vibrationalspectra to obtain the relative strengths of HBs We firstperform a normal-mode analysis by the basic equation ofvibrational spectroscopy formulated as
Λ=f L MLx (2)
where L collects the vibrational eigenvectors Iμ and thediagonal eigenvalue matrix Λ contains Nvib vibrationaleigenvalues 4π2c2νμ
2 (with μ = 1 Nvib and Nvib = 3N minus KK = 6 for nonlinear molecules and 5 for linear molecules) andK zero eigenvalues corresponding to translations and rotationsof the molecule (νμ represents the harmonic vibrationalfrequencies given in cmminus1 and c is the speed of light) M is the
diagonal mass matrix and fx is the force constant matrixexpressed in Cartesian coordinates The (3N times 3N) dimen-sional L matrix collects the Nvib normal-vibrational-modevectors Iμ and K-mode vectors corresponding to translationsand rotationsExpressing eq 2 in internal coordinates q leads to the Wilson
GF formalism with5657
Λ= minusF D G Dq 1 (3)
where
= minus daggerG BM B1 (4)
=D BL (5)
Fq is the force constant matrix expressed in terms of internalcoordinates q The (Nvib times Nvib) dimensional D matrixrepresents each normal-mode vector dμ The rectangular (Nvibtimes 3N) dimensional B matrix contains the first derivatives ofthe internal coordinates with regard to the Cartesiancoordinates thus connecting both coordinate systems andthe (Nvib times Nvib) dimensional matrix G is the Wilson Gmatrix5657
In terms of internal coordinates the Lagrange equations canbe formulated as
= minusdagger minus daggerL q q q G q q F q( )12
12
q1(6)
where decoupling of the second part (potential energy) is doneby the diagonalization of Fq However the first part (kineticenergy) remains coupled due to the matrix G Konkoli andCremer32 solved the problem of mass coupling by introducingthe concept of local vibrational modes The mass-decoupledEulerminusLagrange equation was solved by setting all the atomicmasses to zero except those of the molecular fragment (egbond angle or dihedral etc) carrying out a localized vibrationAs shown in their original work the change in the localdisplacement of a specific internal coordinate is equivalent toan adiabatic relaxation of the molecule32 For any molecularfragment associated with an internal coordinate qμ thecorresponding local mode vector aμ is given by
=μμ
μ μ
minus dagger
minus daggeraK d
d K d
1
1(7)
where dμ is a row vector of matrix D Matrix K is the diagonalmatrix of force constants kQ expressed in normal coordinatesQμ with
= = daggerF K L f LxQ (8)
resulting from the Wilson GF formalism shown in eqs 3 and 4The local mode force constant kμ
a corresponding to localmode aμ is obtained by
=μ μ μdaggerk a Kaa
(9)
Local vibrational modes have been successfully applied toquantify weak chemical interactions such as hydrogenbonding1934minus38 halogen bonding39minus41 pnicogen bond-ing42minus44 chalcogen bonding58 and tetrel bonding59 and toderive new chemical descriptors such as a new aromaticityindex60minus62 or a generalized Tolman electronic parameter63minus65
as well as for the derivation of a generalized Badger Rule66 andseveral others new concepts66minus73
Figure 4 Representation of a breadth first search (BFS) On the left isa seven-membered intramolecular hydrogen bonding On the right isshown the process of how BFS traverses layer by layer in a network ofconnections to reach the goal state (H1) from the initial state (O11)
Figure 5 Network of a HB The DminusHmiddotmiddotmiddotA shown in red is thetargeted HB The dashed line in blue represents possible HBs near thetargeted HB
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2554
Covalent Characteristics of Hydrogen Bonds Thecovalent contribution to the HBs is reinforced by the twoelectron exchange integrals a phenomenon specific toelectrons where they are assumed to be at multiple places atthe same time These contributions stabilize the overallmolecular energies leading to stronger bonds with increasingcovalent contributions We utilized NBO analysis to obtain theoverlap energy of the oxygen lone pair np with the emptyantibonding σ orbital of donor hydrogen bond (LPtoBD)shown in Figure 6 This results in a concomitant polarization
of the donor hydrogen bond which is a direct consequence ofBentrsquos rule74 and can be observed in various bonds such as CminusH SiminusH SminusH NminusH etcElectrostatic Characteristics of Hydrogen Bonds The
electrostatic interaction is caused by the induction of chargesin the molecules and can be relatively estimated for a HB as
=
Eq q
rc1 2
(10)
where Ec is the electrostatic energy q1 and q2 are the atomiccharges of A and H respectively and r is the distance betweenthe two atoms75 We choose NBO charges as they are morereliable than Mulliken charges76 since they operate on the basisof electron density Localized natural atomic orbitals were usedto describe the computed electron densities and thereforeinclude bond polarizationComputational Details Geometry optimization and
normal-mode analysis were carried out in the gas phaseusing Gaussian 167778 Eight smaller proteins 1K43 1L2Y1R4G 1S4A 2EVQ 1UAO 2O0S and 5E61 shown in Figure7 were optimized with the hybrid B3LYP functional79minus82 Highaccuracy paired with relatively low computational costs hasmade B3LYP a popular DFT choice Kuhn et al83 and Bartlettet al84 successfully used this functional for the description of
small molecules and peptides We further employed empiricaldispersion correction8586 as suggested by Fadda et al87 toinclude polarization effects that can significantly impacthydrogen bond stability Based on the findings of Fadda etal87 that B3LYP paired with the 6-31G(dp) basis88minus94
provides a good description of short alanine peptides wealso used the 6-31G(dp) in our study For all geometryoptimizations conversion criteria to a maximum step size of001 au and a RMS force of 00017 au were applied Protein1R4G was cut and only the area of major interest was takenSome side chains were removed from proteins 1K43 1S4Aand 5E61 for faster calculation (details and geometryinformation are provided in the Supporting Information aspages S23minusS59)To obtain statistically relevant information in addition a set
of 155 smaller proteins was optimized and second energyderivatives were calculated with the semiempirical DFTmethod GFN2-xTB95 The GFN2-xTB is significantly fasterthan B3LYP with a low compromise regarding accuracy Asimilar distribution of second-order properties such asvibrational constants for GFN2-xTB compared to B3LYPwas observed which is described in the Results and Discussionsection The similarity between the results of the two methodsled to our choice of GFN2-xTB as the DFT method for thestatistical analysis The 155 proteins were selected based on thesmall size single chain and NMR solution as characterizingtechniques from the Protein Data Bank53 Hydrogen atomswere added utilizing Leap (a tool from Amber96) structureswere minimized for 1000 steps in Amber96 and the overallcharges were calculated from Leap prior to the GFN2-xTBcalculations The PDB IDs for all 155 proteins are provided inthe Supporting Information on page S2The local-mode force constants for each HB were calculated
using the LModeA package3397 A description of thecorresponding local vibrational mode theory is given belowLPtoBD and atomic charges were obtained from NBO98
analysis utilizing optimized geometries with the B3LYP79minus82
level of theory including empirical dispersion correction8586
along with the 6-31G(dp)88minus94 basis set
RESULTS AND DISCUSSIONThis section is organized into subsections providing results anddiscussion by analyzing patterns for a general overview of HBproperties intramolecular HBs HB type based on donorminusacceptor pairs networks of HBs and the governing factors forthe HB strength in proteins We start by comparing predictedHB properties from the two DFT methods to get statisticallyrelevant information We observed similar patterns of HBproperties from the two DFT methods Thus we provide adetailed characterization of HB properties from a higher levelof DFT method (B3LYP) followed by statistical results from alower level of DFT method (GFN2-xTB)Figure 7 shows the 8 proteins investigated with B3LYP and
the PDB IDs for 155 proteins investigated with GFN2-xTB areprovided on pages S2minusS10 The HBs detected by EDHB areshown in yellow Each HB is given a specific ID for discussion(larger images of proteins are shown in the SupportingInformation as Figures S1minusS8) These proteins accommodatea variety of secondary structures which include a majority ofHB types observed in proteinsTo generalize the outcomes from GFN2-xTB and B3LYP
we focused on the distribution of HB length and ka for theeight proteins optimized as shown in Figure 8 The HBs that
Figure 6 Overlap of the lone pair np of the acceptor to the σ of thedonor hydrogen bond The overlap energy estimates the covalentcharacteristics of a hydrogen bond
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2555
were absent in either one of the methods were removed TheGFN2-xTB and B3LYP methods differ and therefore theoptimized geometries are not exactly the same This is reflectedby Figure 8b where the median of the HB length for B3LYP(195 Aring) is slightly higher than that for GFN2-xTB (192 Aring)However the overall distribution remains the same and is alsoobserved for ka as shown in Figure 8a The comparison ofLPtoBD HB angle and electrostatic energy is shown inFigure S9 where we observe similar distributions Based onthese results we conclude that statistically relevant informationcan be obtained from the GFN2-xTB method
We analyzed quantitatively the number of different HBspresent in 155 proteins based on secondary structures shownin Table 2 Note that EDHB uses data obtained fromDictionary of Protein Secondary Structure (DSSP)99 forsecondary structure classification if the PDB ID of the proteinis available The proteins circumvent a variety of secondarystructures with α turn bend and extended strand composing204 93 86 and 67 respectively Less common HBs suchas 310 helix and isolated β-bridge are 22 and 07 respectivelyThe secondary structures with no backbone classification are
Figure 7 Eight proteins investigated Each number is shown as a unique ID associated with each HB (a) 1K43 is a beta hairpin type structure andcontains 13 HBs (1minus13) (b) A trp-cage motif 1L2Y has α helix and 19 HBs (14minus32) (c) The Sendai virus protein 1R4G was cut to keep the idealα-helix Other atoms were removed for the convenience of running ab initio calculations Twenty-eight HBs were detected for this protein (33minus60) (d) A double-strand β-helix of a DL-alternating oligonorleucine 1S4A has 18 HBs (61minus78) (e) A designed protein Chignolin 1UAO is a β-sheet type structure and looks more like a hairpin with 10 HBs (79minus88) (f) A 12-residue beta hairpin 2EVQ with 15 HBs (89minus103) (g) Adesigned peptide 2O0S is a coil type structure and has 7 HBs (104minus110) and the amyloid-forming peptide 5E61 with a near ideal β-strand has 13HBs (111minus123)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2556
termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

Hydrogen Bond Network Classification The strengthof a HB is influenced by the presenceabsence of other HBsthat share the same D or A20 Figure 5 shows a hydrogen-
bonding network The type of HB network can be defined bythe coordination number of the D A or hydrogen(s) from theD as a string
equiv minus minusHB A D DHtype c c c (1)
where Ac Dc and DHc represent the number of HBs from Aexcluding targeted HB number of HB(s) from D and numberof HB(s) from the covalently bonded hydrogen(s) to Drespectively Following this procedure the HB network typeshown in Figure 5 is 1minus2minus1Hydrogen Bond Characterization EDHB characterizes
HBs based on two properties vibrational spectroscopy andNBO analysis data Vibrational spectroscopy data are utilizedwhen the second energy derivative for a molecule is availableto calculate local-mode force constants ka obtained from thepure local stretching vibration of a HB The resultant forceconstants provide the strength of a HB The electrostatic andcovalent contributions are estimated based on results obtainedfrom the NBO analysisLocal-Mode Force Constants The vibrational spectra of
a molecule hold enormous information about the structure andproperties of the molecular interactions We utilize vibrationalspectra to obtain the relative strengths of HBs We firstperform a normal-mode analysis by the basic equation ofvibrational spectroscopy formulated as
Λ=f L MLx (2)
where L collects the vibrational eigenvectors Iμ and thediagonal eigenvalue matrix Λ contains Nvib vibrationaleigenvalues 4π2c2νμ
2 (with μ = 1 Nvib and Nvib = 3N minus KK = 6 for nonlinear molecules and 5 for linear molecules) andK zero eigenvalues corresponding to translations and rotationsof the molecule (νμ represents the harmonic vibrationalfrequencies given in cmminus1 and c is the speed of light) M is the
diagonal mass matrix and fx is the force constant matrixexpressed in Cartesian coordinates The (3N times 3N) dimen-sional L matrix collects the Nvib normal-vibrational-modevectors Iμ and K-mode vectors corresponding to translationsand rotationsExpressing eq 2 in internal coordinates q leads to the Wilson
GF formalism with5657
Λ= minusF D G Dq 1 (3)
where
= minus daggerG BM B1 (4)
=D BL (5)
Fq is the force constant matrix expressed in terms of internalcoordinates q The (Nvib times Nvib) dimensional D matrixrepresents each normal-mode vector dμ The rectangular (Nvibtimes 3N) dimensional B matrix contains the first derivatives ofthe internal coordinates with regard to the Cartesiancoordinates thus connecting both coordinate systems andthe (Nvib times Nvib) dimensional matrix G is the Wilson Gmatrix5657
In terms of internal coordinates the Lagrange equations canbe formulated as
= minusdagger minus daggerL q q q G q q F q( )12
12
q1(6)
where decoupling of the second part (potential energy) is doneby the diagonalization of Fq However the first part (kineticenergy) remains coupled due to the matrix G Konkoli andCremer32 solved the problem of mass coupling by introducingthe concept of local vibrational modes The mass-decoupledEulerminusLagrange equation was solved by setting all the atomicmasses to zero except those of the molecular fragment (egbond angle or dihedral etc) carrying out a localized vibrationAs shown in their original work the change in the localdisplacement of a specific internal coordinate is equivalent toan adiabatic relaxation of the molecule32 For any molecularfragment associated with an internal coordinate qμ thecorresponding local mode vector aμ is given by
=μμ
μ μ
minus dagger
minus daggeraK d
d K d
1
1(7)
where dμ is a row vector of matrix D Matrix K is the diagonalmatrix of force constants kQ expressed in normal coordinatesQμ with
= = daggerF K L f LxQ (8)
resulting from the Wilson GF formalism shown in eqs 3 and 4The local mode force constant kμ
a corresponding to localmode aμ is obtained by
=μ μ μdaggerk a Kaa
(9)
Local vibrational modes have been successfully applied toquantify weak chemical interactions such as hydrogenbonding1934minus38 halogen bonding39minus41 pnicogen bond-ing42minus44 chalcogen bonding58 and tetrel bonding59 and toderive new chemical descriptors such as a new aromaticityindex60minus62 or a generalized Tolman electronic parameter63minus65
as well as for the derivation of a generalized Badger Rule66 andseveral others new concepts66minus73
Figure 4 Representation of a breadth first search (BFS) On the left isa seven-membered intramolecular hydrogen bonding On the right isshown the process of how BFS traverses layer by layer in a network ofconnections to reach the goal state (H1) from the initial state (O11)
Figure 5 Network of a HB The DminusHmiddotmiddotmiddotA shown in red is thetargeted HB The dashed line in blue represents possible HBs near thetargeted HB
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2554
Covalent Characteristics of Hydrogen Bonds Thecovalent contribution to the HBs is reinforced by the twoelectron exchange integrals a phenomenon specific toelectrons where they are assumed to be at multiple places atthe same time These contributions stabilize the overallmolecular energies leading to stronger bonds with increasingcovalent contributions We utilized NBO analysis to obtain theoverlap energy of the oxygen lone pair np with the emptyantibonding σ orbital of donor hydrogen bond (LPtoBD)shown in Figure 6 This results in a concomitant polarization
of the donor hydrogen bond which is a direct consequence ofBentrsquos rule74 and can be observed in various bonds such as CminusH SiminusH SminusH NminusH etcElectrostatic Characteristics of Hydrogen Bonds The
electrostatic interaction is caused by the induction of chargesin the molecules and can be relatively estimated for a HB as
=
Eq q
rc1 2
(10)
where Ec is the electrostatic energy q1 and q2 are the atomiccharges of A and H respectively and r is the distance betweenthe two atoms75 We choose NBO charges as they are morereliable than Mulliken charges76 since they operate on the basisof electron density Localized natural atomic orbitals were usedto describe the computed electron densities and thereforeinclude bond polarizationComputational Details Geometry optimization and
normal-mode analysis were carried out in the gas phaseusing Gaussian 167778 Eight smaller proteins 1K43 1L2Y1R4G 1S4A 2EVQ 1UAO 2O0S and 5E61 shown in Figure7 were optimized with the hybrid B3LYP functional79minus82 Highaccuracy paired with relatively low computational costs hasmade B3LYP a popular DFT choice Kuhn et al83 and Bartlettet al84 successfully used this functional for the description of
small molecules and peptides We further employed empiricaldispersion correction8586 as suggested by Fadda et al87 toinclude polarization effects that can significantly impacthydrogen bond stability Based on the findings of Fadda etal87 that B3LYP paired with the 6-31G(dp) basis88minus94
provides a good description of short alanine peptides wealso used the 6-31G(dp) in our study For all geometryoptimizations conversion criteria to a maximum step size of001 au and a RMS force of 00017 au were applied Protein1R4G was cut and only the area of major interest was takenSome side chains were removed from proteins 1K43 1S4Aand 5E61 for faster calculation (details and geometryinformation are provided in the Supporting Information aspages S23minusS59)To obtain statistically relevant information in addition a set
of 155 smaller proteins was optimized and second energyderivatives were calculated with the semiempirical DFTmethod GFN2-xTB95 The GFN2-xTB is significantly fasterthan B3LYP with a low compromise regarding accuracy Asimilar distribution of second-order properties such asvibrational constants for GFN2-xTB compared to B3LYPwas observed which is described in the Results and Discussionsection The similarity between the results of the two methodsled to our choice of GFN2-xTB as the DFT method for thestatistical analysis The 155 proteins were selected based on thesmall size single chain and NMR solution as characterizingtechniques from the Protein Data Bank53 Hydrogen atomswere added utilizing Leap (a tool from Amber96) structureswere minimized for 1000 steps in Amber96 and the overallcharges were calculated from Leap prior to the GFN2-xTBcalculations The PDB IDs for all 155 proteins are provided inthe Supporting Information on page S2The local-mode force constants for each HB were calculated
using the LModeA package3397 A description of thecorresponding local vibrational mode theory is given belowLPtoBD and atomic charges were obtained from NBO98
analysis utilizing optimized geometries with the B3LYP79minus82
level of theory including empirical dispersion correction8586
along with the 6-31G(dp)88minus94 basis set
RESULTS AND DISCUSSIONThis section is organized into subsections providing results anddiscussion by analyzing patterns for a general overview of HBproperties intramolecular HBs HB type based on donorminusacceptor pairs networks of HBs and the governing factors forthe HB strength in proteins We start by comparing predictedHB properties from the two DFT methods to get statisticallyrelevant information We observed similar patterns of HBproperties from the two DFT methods Thus we provide adetailed characterization of HB properties from a higher levelof DFT method (B3LYP) followed by statistical results from alower level of DFT method (GFN2-xTB)Figure 7 shows the 8 proteins investigated with B3LYP and
the PDB IDs for 155 proteins investigated with GFN2-xTB areprovided on pages S2minusS10 The HBs detected by EDHB areshown in yellow Each HB is given a specific ID for discussion(larger images of proteins are shown in the SupportingInformation as Figures S1minusS8) These proteins accommodatea variety of secondary structures which include a majority ofHB types observed in proteinsTo generalize the outcomes from GFN2-xTB and B3LYP
we focused on the distribution of HB length and ka for theeight proteins optimized as shown in Figure 8 The HBs that
Figure 6 Overlap of the lone pair np of the acceptor to the σ of thedonor hydrogen bond The overlap energy estimates the covalentcharacteristics of a hydrogen bond
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2555
were absent in either one of the methods were removed TheGFN2-xTB and B3LYP methods differ and therefore theoptimized geometries are not exactly the same This is reflectedby Figure 8b where the median of the HB length for B3LYP(195 Aring) is slightly higher than that for GFN2-xTB (192 Aring)However the overall distribution remains the same and is alsoobserved for ka as shown in Figure 8a The comparison ofLPtoBD HB angle and electrostatic energy is shown inFigure S9 where we observe similar distributions Based onthese results we conclude that statistically relevant informationcan be obtained from the GFN2-xTB method
We analyzed quantitatively the number of different HBspresent in 155 proteins based on secondary structures shownin Table 2 Note that EDHB uses data obtained fromDictionary of Protein Secondary Structure (DSSP)99 forsecondary structure classification if the PDB ID of the proteinis available The proteins circumvent a variety of secondarystructures with α turn bend and extended strand composing204 93 86 and 67 respectively Less common HBs suchas 310 helix and isolated β-bridge are 22 and 07 respectivelyThe secondary structures with no backbone classification are
Figure 7 Eight proteins investigated Each number is shown as a unique ID associated with each HB (a) 1K43 is a beta hairpin type structure andcontains 13 HBs (1minus13) (b) A trp-cage motif 1L2Y has α helix and 19 HBs (14minus32) (c) The Sendai virus protein 1R4G was cut to keep the idealα-helix Other atoms were removed for the convenience of running ab initio calculations Twenty-eight HBs were detected for this protein (33minus60) (d) A double-strand β-helix of a DL-alternating oligonorleucine 1S4A has 18 HBs (61minus78) (e) A designed protein Chignolin 1UAO is a β-sheet type structure and looks more like a hairpin with 10 HBs (79minus88) (f) A 12-residue beta hairpin 2EVQ with 15 HBs (89minus103) (g) Adesigned peptide 2O0S is a coil type structure and has 7 HBs (104minus110) and the amyloid-forming peptide 5E61 with a near ideal β-strand has 13HBs (111minus123)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2556
termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

Covalent Characteristics of Hydrogen Bonds Thecovalent contribution to the HBs is reinforced by the twoelectron exchange integrals a phenomenon specific toelectrons where they are assumed to be at multiple places atthe same time These contributions stabilize the overallmolecular energies leading to stronger bonds with increasingcovalent contributions We utilized NBO analysis to obtain theoverlap energy of the oxygen lone pair np with the emptyantibonding σ orbital of donor hydrogen bond (LPtoBD)shown in Figure 6 This results in a concomitant polarization
of the donor hydrogen bond which is a direct consequence ofBentrsquos rule74 and can be observed in various bonds such as CminusH SiminusH SminusH NminusH etcElectrostatic Characteristics of Hydrogen Bonds The
electrostatic interaction is caused by the induction of chargesin the molecules and can be relatively estimated for a HB as
=
Eq q
rc1 2
(10)
where Ec is the electrostatic energy q1 and q2 are the atomiccharges of A and H respectively and r is the distance betweenthe two atoms75 We choose NBO charges as they are morereliable than Mulliken charges76 since they operate on the basisof electron density Localized natural atomic orbitals were usedto describe the computed electron densities and thereforeinclude bond polarizationComputational Details Geometry optimization and
normal-mode analysis were carried out in the gas phaseusing Gaussian 167778 Eight smaller proteins 1K43 1L2Y1R4G 1S4A 2EVQ 1UAO 2O0S and 5E61 shown in Figure7 were optimized with the hybrid B3LYP functional79minus82 Highaccuracy paired with relatively low computational costs hasmade B3LYP a popular DFT choice Kuhn et al83 and Bartlettet al84 successfully used this functional for the description of
small molecules and peptides We further employed empiricaldispersion correction8586 as suggested by Fadda et al87 toinclude polarization effects that can significantly impacthydrogen bond stability Based on the findings of Fadda etal87 that B3LYP paired with the 6-31G(dp) basis88minus94
provides a good description of short alanine peptides wealso used the 6-31G(dp) in our study For all geometryoptimizations conversion criteria to a maximum step size of001 au and a RMS force of 00017 au were applied Protein1R4G was cut and only the area of major interest was takenSome side chains were removed from proteins 1K43 1S4Aand 5E61 for faster calculation (details and geometryinformation are provided in the Supporting Information aspages S23minusS59)To obtain statistically relevant information in addition a set
of 155 smaller proteins was optimized and second energyderivatives were calculated with the semiempirical DFTmethod GFN2-xTB95 The GFN2-xTB is significantly fasterthan B3LYP with a low compromise regarding accuracy Asimilar distribution of second-order properties such asvibrational constants for GFN2-xTB compared to B3LYPwas observed which is described in the Results and Discussionsection The similarity between the results of the two methodsled to our choice of GFN2-xTB as the DFT method for thestatistical analysis The 155 proteins were selected based on thesmall size single chain and NMR solution as characterizingtechniques from the Protein Data Bank53 Hydrogen atomswere added utilizing Leap (a tool from Amber96) structureswere minimized for 1000 steps in Amber96 and the overallcharges were calculated from Leap prior to the GFN2-xTBcalculations The PDB IDs for all 155 proteins are provided inthe Supporting Information on page S2The local-mode force constants for each HB were calculated
using the LModeA package3397 A description of thecorresponding local vibrational mode theory is given belowLPtoBD and atomic charges were obtained from NBO98
analysis utilizing optimized geometries with the B3LYP79minus82
level of theory including empirical dispersion correction8586
along with the 6-31G(dp)88minus94 basis set
RESULTS AND DISCUSSIONThis section is organized into subsections providing results anddiscussion by analyzing patterns for a general overview of HBproperties intramolecular HBs HB type based on donorminusacceptor pairs networks of HBs and the governing factors forthe HB strength in proteins We start by comparing predictedHB properties from the two DFT methods to get statisticallyrelevant information We observed similar patterns of HBproperties from the two DFT methods Thus we provide adetailed characterization of HB properties from a higher levelof DFT method (B3LYP) followed by statistical results from alower level of DFT method (GFN2-xTB)Figure 7 shows the 8 proteins investigated with B3LYP and
the PDB IDs for 155 proteins investigated with GFN2-xTB areprovided on pages S2minusS10 The HBs detected by EDHB areshown in yellow Each HB is given a specific ID for discussion(larger images of proteins are shown in the SupportingInformation as Figures S1minusS8) These proteins accommodatea variety of secondary structures which include a majority ofHB types observed in proteinsTo generalize the outcomes from GFN2-xTB and B3LYP
we focused on the distribution of HB length and ka for theeight proteins optimized as shown in Figure 8 The HBs that
Figure 6 Overlap of the lone pair np of the acceptor to the σ of thedonor hydrogen bond The overlap energy estimates the covalentcharacteristics of a hydrogen bond
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2555
were absent in either one of the methods were removed TheGFN2-xTB and B3LYP methods differ and therefore theoptimized geometries are not exactly the same This is reflectedby Figure 8b where the median of the HB length for B3LYP(195 Aring) is slightly higher than that for GFN2-xTB (192 Aring)However the overall distribution remains the same and is alsoobserved for ka as shown in Figure 8a The comparison ofLPtoBD HB angle and electrostatic energy is shown inFigure S9 where we observe similar distributions Based onthese results we conclude that statistically relevant informationcan be obtained from the GFN2-xTB method
We analyzed quantitatively the number of different HBspresent in 155 proteins based on secondary structures shownin Table 2 Note that EDHB uses data obtained fromDictionary of Protein Secondary Structure (DSSP)99 forsecondary structure classification if the PDB ID of the proteinis available The proteins circumvent a variety of secondarystructures with α turn bend and extended strand composing204 93 86 and 67 respectively Less common HBs suchas 310 helix and isolated β-bridge are 22 and 07 respectivelyThe secondary structures with no backbone classification are
Figure 7 Eight proteins investigated Each number is shown as a unique ID associated with each HB (a) 1K43 is a beta hairpin type structure andcontains 13 HBs (1minus13) (b) A trp-cage motif 1L2Y has α helix and 19 HBs (14minus32) (c) The Sendai virus protein 1R4G was cut to keep the idealα-helix Other atoms were removed for the convenience of running ab initio calculations Twenty-eight HBs were detected for this protein (33minus60) (d) A double-strand β-helix of a DL-alternating oligonorleucine 1S4A has 18 HBs (61minus78) (e) A designed protein Chignolin 1UAO is a β-sheet type structure and looks more like a hairpin with 10 HBs (79minus88) (f) A 12-residue beta hairpin 2EVQ with 15 HBs (89minus103) (g) Adesigned peptide 2O0S is a coil type structure and has 7 HBs (104minus110) and the amyloid-forming peptide 5E61 with a near ideal β-strand has 13HBs (111minus123)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2556
termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

were absent in either one of the methods were removed TheGFN2-xTB and B3LYP methods differ and therefore theoptimized geometries are not exactly the same This is reflectedby Figure 8b where the median of the HB length for B3LYP(195 Aring) is slightly higher than that for GFN2-xTB (192 Aring)However the overall distribution remains the same and is alsoobserved for ka as shown in Figure 8a The comparison ofLPtoBD HB angle and electrostatic energy is shown inFigure S9 where we observe similar distributions Based onthese results we conclude that statistically relevant informationcan be obtained from the GFN2-xTB method
We analyzed quantitatively the number of different HBspresent in 155 proteins based on secondary structures shownin Table 2 Note that EDHB uses data obtained fromDictionary of Protein Secondary Structure (DSSP)99 forsecondary structure classification if the PDB ID of the proteinis available The proteins circumvent a variety of secondarystructures with α turn bend and extended strand composing204 93 86 and 67 respectively Less common HBs suchas 310 helix and isolated β-bridge are 22 and 07 respectivelyThe secondary structures with no backbone classification are
Figure 7 Eight proteins investigated Each number is shown as a unique ID associated with each HB (a) 1K43 is a beta hairpin type structure andcontains 13 HBs (1minus13) (b) A trp-cage motif 1L2Y has α helix and 19 HBs (14minus32) (c) The Sendai virus protein 1R4G was cut to keep the idealα-helix Other atoms were removed for the convenience of running ab initio calculations Twenty-eight HBs were detected for this protein (33minus60) (d) A double-strand β-helix of a DL-alternating oligonorleucine 1S4A has 18 HBs (61minus78) (e) A designed protein Chignolin 1UAO is a β-sheet type structure and looks more like a hairpin with 10 HBs (79minus88) (f) A 12-residue beta hairpin 2EVQ with 15 HBs (89minus103) (g) Adesigned peptide 2O0S is a coil type structure and has 7 HBs (104minus110) and the amyloid-forming peptide 5E61 with a near ideal β-strand has 13HBs (111minus123)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2556
termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

termed as backbone and involve 201 of all HBs The sidechains involve 311 of all HBs
General Trends of Hydrogen Bonds in Proteins Theestimated contribution of an individual HB in general is in therange of 2minus8 kcalmol depending on factors such as geometryand environment It has been suggested that the strength of aHB in a protein depends on the HB length29 However weobserve that the strength can vary depending on theenvironment of the HB Furthermore shorter bonds are notalways stronger bonds68 The environment creates an electricfield that perturbs the electrostatic and covalent interactions ofthe HB Thus we analyzed the covalent character by LPtoBD
Figure 8 Comparison of B3LYP and GFN2-xTB on eight small proteins Hydrogen bonds that do not exist in either one of the methods wereremoved (a) Plot of ka vs HB length The line shows the corresponding quadratic fit We observe a similar distribution of ka values from eithermethod (b) Box plot for the HB length The median distribution of GFN2-xTB is slightly higher than that of B3LYP
Table 2 Statistics for Secondary Structure
secondary structures percentage of HBs number of HBs
side chain 317 674α 204 435backbone 201 427turn 93 207bend 86 182extended strand 67 143310 helix 22 46isolated β-bridge 07 14total 2128
Figure 9 Scatter matrix plot for all continuous data from EDHB results on 155 proteins HB length LPtoBD ka HB angle and Elec are thehydrogen bond length (in Aring) LPtoBD (in kcalmol) ka for stretching vibration of HB (in mdynAring) the angle created by acceptor hydrogen anddonor respectively and the electrostatic energy (in e2Aring) The diagonal elements of the matrix show the histogram of the distributions Each of theoff-diagonal elements is the plot of x vs y where x y are the HB properties named as xlabel and ylabel respectively
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2557
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565
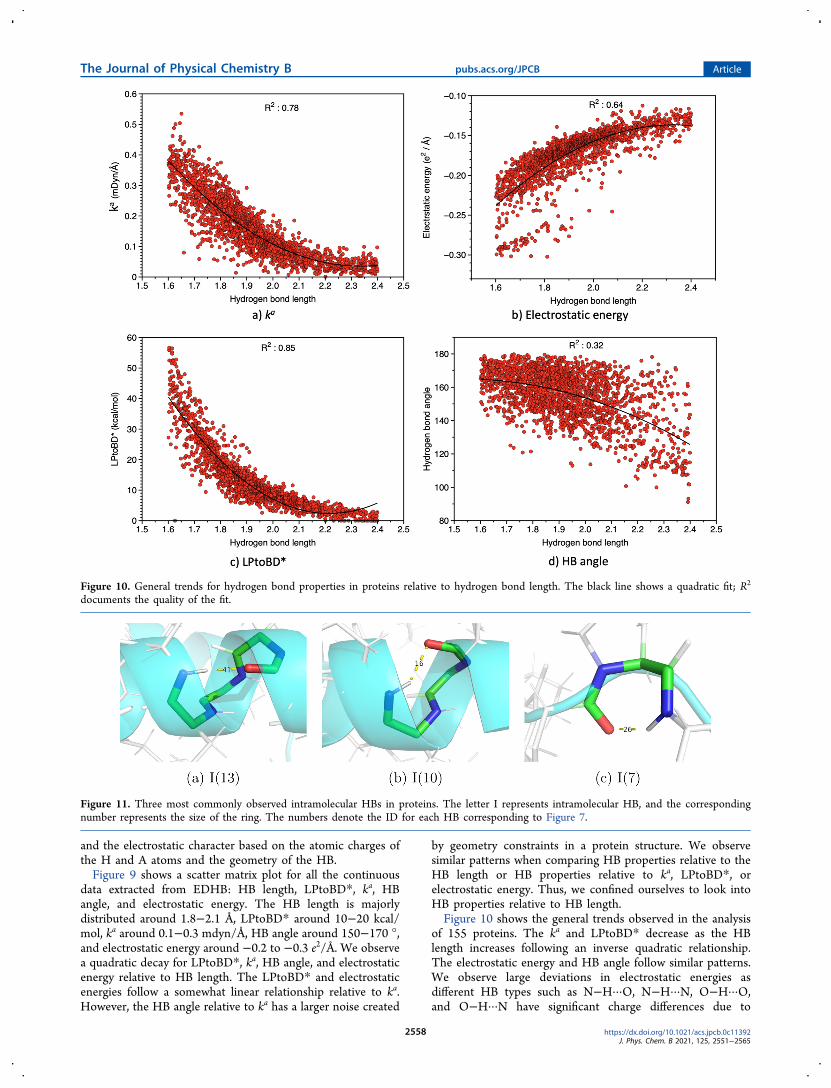
and the electrostatic character based on the atomic charges ofthe H and A atoms and the geometry of the HBFigure 9 shows a scatter matrix plot for all the continuous
data extracted from EDHB HB length LPtoBD ka HBangle and electrostatic energy The HB length is majorlydistributed around 18minus21 Aring LPtoBD around 10minus20 kcalmol ka around 01minus03 mdynAring HB angle around 150minus170 degand electrostatic energy around minus02 to minus03 e2Aring We observea quadratic decay for LPtoBD ka HB angle and electrostaticenergy relative to HB length The LPtoBD and electrostaticenergies follow a somewhat linear relationship relative to kaHowever the HB angle relative to ka has a larger noise created
by geometry constraints in a protein structure We observesimilar patterns when comparing HB properties relative to theHB length or HB properties relative to ka LPtoBD orelectrostatic energy Thus we confined ourselves to look intoHB properties relative to HB lengthFigure 10 shows the general trends observed in the analysis
of 155 proteins The ka and LPtoBD decrease as the HBlength increases following an inverse quadratic relationshipThe electrostatic energy and HB angle follow similar patternsWe observe large deviations in electrostatic energies asdifferent HB types such as NminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN OminusHmiddotmiddotmiddotOand OminusHmiddotmiddotmiddotN have significant charge differences due to
Figure 10 General trends for hydrogen bond properties in proteins relative to hydrogen bond length The black line shows a quadratic fit R2
documents the quality of the fit
Figure 11 Three most commonly observed intramolecular HBs in proteins The letter I represents intramolecular HB and the correspondingnumber represents the size of the ring The numbers denote the ID for each HB corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2558
variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

variations in the electronegativity for N and O atoms The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs have a high electrostatic characteraccompanied by a short hydrogen bond length In general Nas the acceptor increases the electrostatic character Whenacting as the donor O being more electronegative comparedto N makes the corresponding H more positively chargedThis results in a strong electrostatic interaction between the Hand the acceptor Thus we observe an island of data pointsthat correspond to NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type hydrogenbonds as shown in Figure 10b The HB angle follows a similarpattern however it has significant noise that might be causeddue to the geometric constraints of a protein structureIntramolecular Hydrogen Bonds Figure 11 shows the
three most commonly observed intramolecular HBs theproperties of which are summarized in Table 3 and arefrequently observed in the backbone Figure 11a shows I(13)(intramolecular HB with a ring size of 13 atoms) that forms aHB observed in the α-helix The orientation of the fourconsecutive amino acids (RnminusRn+4 where Rn is the residue atthe nth position in the sequence of the protein) makes the HBalmost linear significantly increasing the LPtoBD Figure 11bshows I(10) formed in between RnminusRn+3 A study by Perutz100
in the 1950s confirmed the presence of I(10) in 310 helixesHowever we observe I(10) over all backbone types majorly inα-helixes and in smaller amounts from β-sheets and coil I(10)has a larger deviation in the HB angle depending on itsorientation exhibiting variation in electrostatic energyLPtoBD and HB angle Figure 11c shows I(7) formed inbetween RnminusRn+2 Contrary to I(13) and I(10) I(7) is not justconfined to the backbone but extends to side chains as wellFurther investigation from the 155 proteins provided
statistically relevant data I(10) was the most commonlyobserved intramolecular HB constituting 186 of all intra-molecular HBs followed by I(13) (161) and I(7) (130)The majority of I(13) is observed in α-helices constituting437 of all I(13) The percentage of other intramolecular HBscontributing more than 3 is provided in Table S3Figure 12a shows ka for I(10) I(13) and I(7) relative to HB
lengths I(13) have a higher mean of HB angle as shown inFigure 12b leading to an increase in the LPtoBD andtherefore ka This is reflected by the exponential fit curve thattends to be consistently higher than the exponential fit forI(10) and I(7) We observe I(7) have lower ka values relativeto HB length due to the smaller HB angle (Figure 12b)
Table 3 Intramolecular HBs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
41 NminusHmiddotmiddotmiddotO 190 minus017 157 021 I(13) 130 bminusb 0minus0minus016 NminusHmiddotmiddotmiddotO 213 minus014 143 007 I(10) 148 bminusb 0minus0minus026 NminusHmiddotmiddotmiddotO 200 minus016 146 014 I(7) 616 bminusb 0minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 12 Statistics of three major intramolecular HBs for (a) ka and (b) HB angle The lines in (a) shows the quadratic fit curve for thecorresponding intramolecular hydrogen bonds The plots show that the hydrogen bond stability follows the order I(13) gt I(10) gt I(7) with respectto the same HB length
Figure 13 Various HB types based on the donorminusacceptor pairs The numbers denote the HB ID corresponding to Figure 7
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2559
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565
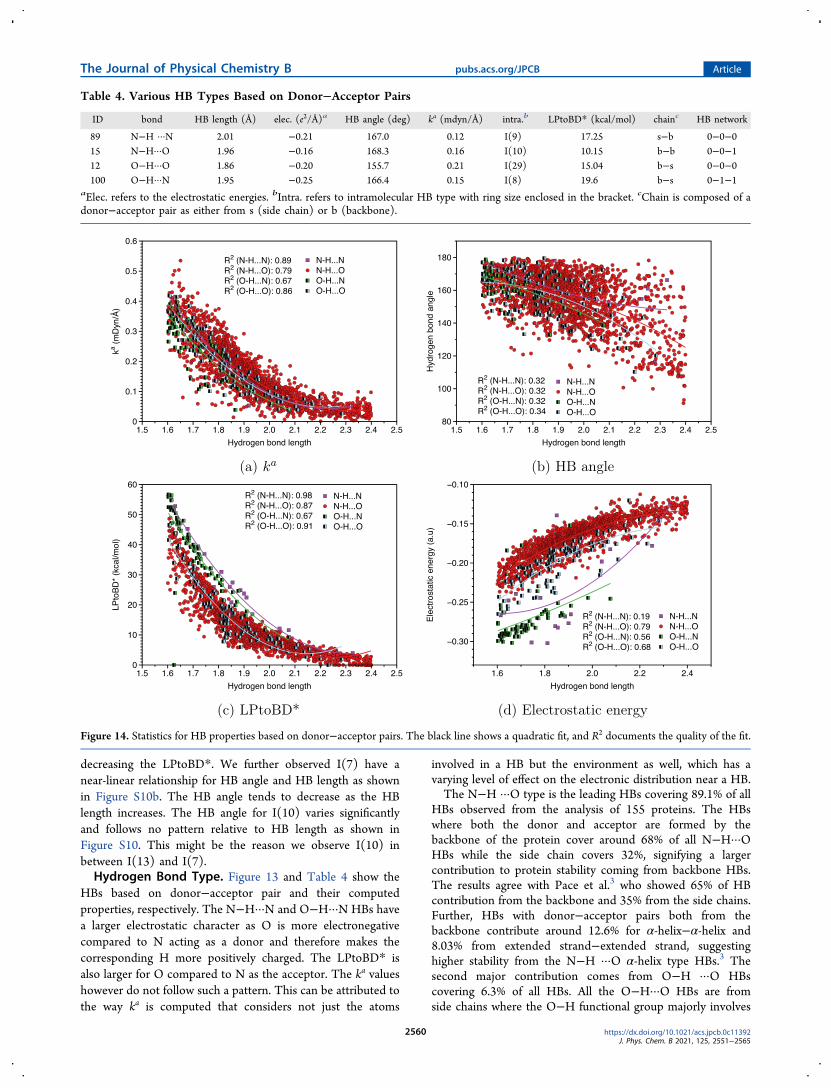
decreasing the LPtoBD We further observed I(7) have anear-linear relationship for HB angle and HB length as shownin Figure S10b The HB angle tends to decrease as the HBlength increases The HB angle for I(10) varies significantlyand follows no pattern relative to HB length as shown inFigure S10 This might be the reason we observe I(10) inbetween I(13) and I(7)Hydrogen Bond Type Figure 13 and Table 4 show the
HBs based on donorminusacceptor pair and their computedproperties respectively The NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs havea larger electrostatic character as O is more electronegativecompared to N acting as a donor and therefore makes thecorresponding H more positively charged The LPtoBD isalso larger for O compared to N as the acceptor The ka valueshowever do not follow such a pattern This can be attributed tothe way ka is computed that considers not just the atoms
involved in a HB but the environment as well which has avarying level of effect on the electronic distribution near a HBThe NminusH middotmiddotmiddotO type is the leading HBs covering 891 of all
HBs observed from the analysis of 155 proteins The HBswhere both the donor and acceptor are formed by thebackbone of the protein cover around 68 of all NminusHmiddotmiddotmiddotOHBs while the side chain covers 32 signifying a largercontribution to protein stability coming from backbone HBsThe results agree with Pace et al3 who showed 65 of HBcontribution from the backbone and 35 from the side chainsFurther HBs with donorminusacceptor pairs both from thebackbone contribute around 126 for α-helixminusα-helix and803 from extended strandminusextended strand suggestinghigher stability from the NminusH middotmiddotmiddotO α-helix type HBs3 Thesecond major contribution comes from OminusH middotmiddotmiddotO HBscovering 63 of all HBs All the OminusHmiddotmiddotmiddotO HBs are fromside chains where the OminusH functional group majorly involves
Table 4 Various HB Types Based on DonorminusAcceptor Pairs
ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
89 NminusH middotmiddotmiddotN 201 minus021 1670 012 I(9) 1725 sminusb 0minus0minus015 NminusHmiddotmiddotmiddotO 196 minus016 1683 016 I(10) 1015 bminusb 0minus0minus112 OminusHmiddotmiddotmiddotO 186 minus020 1557 021 I(29) 1504 bminuss 0minus0minus0100 OminusHmiddotmiddotmiddotN 195 minus025 1664 015 I(8) 196 bminuss 0minus1minus1
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
Figure 14 Statistics for HB properties based on donorminusacceptor pairs The black line shows a quadratic fit and R2 documents the quality of the fit
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2560
the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

the residues SER (442) THR (400) and TYR (86) asdonors The contributions from OminusHmiddotmiddotmiddotN and NminusHmiddotmiddotmiddotN are34 and 11 respectively The small fraction of OminusHmiddotmiddotmiddotNHBs is majorly dominated by side chain HBs (908)compared to backbone HBs (92) However the smallfraction of NminusHmiddotmiddotmiddotN bonds are either from the side chains(440) or the backbone HBs (560) signifying almost equalcontribution from both the side chain and the backboneFigure 14 shows the distribution of ka HB angle LPtoBD
and electrostatic energy relative to the HB length for theproteins investigated with GFN2-xTB In general OminusH middotmiddotmiddotNHBs have the lowest quadratic fit curve signifying lower HBstrength relative to HB length when compared with thequadratic fit curve for other HBs However the HB strength ofNminusHmiddotmiddotmiddotO NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotO does not seem to havesignificant differences The distribution in the HB angle has asimilar pattern for all the HBs The LPtoBD has a higher
quadratic fit curve for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN HBs relative toHB length and so does the electrostatic energy The OminusHmiddotmiddotmiddotOand NminusHmiddotmiddotmiddotO HBs do not have significant differencesHowever NminusHmiddotmiddotmiddotO HBs have the lowest electrostatic energycompared to other HB types
Hydrogen Bond Networks HBs in general are comprisedof single acceptor and donor pair However there are instanceswhere two or more HBs share the same acceptor forming anetwork of HBs These HBs have been analyzed2122 and theirimplications for the bending of helices have been discussed23
The most common example can be found in α-helices where aside chain interacts with the backbone amide carbonyl group22
However these HBs are also commonly observed at the startor the end of β-sheets A study by Feldblum and Arkin24
clearly shows the existence of HB networks with bothexperimental and computational means and highlighted that
Figure 15 Various scenarios of HBs leading to various HB networks derived from eq 1 The network 1minus0minus0 is shown in four different plots (a)(c) (d) and (f) The network 1minus1minus0 is shown as (b) and the network 2minus0minus0 is shown as (e)
Table 5 Various HB Properties Based on the Network of a HB
no ID bond HB length (Aring) elec (e2Aring)a HB angle (deg) ka (mdynAring) intrab LPtoBD (kcalmol) chainc HB network
1 34 NminusH middotmiddotmiddotO 219 minus014 1355 002 I(10) 09 bminusb 1minus0minus036 NminusHmiddotmiddotmiddotO 204 minus015 1648 007 I(13) 907 bminusb 1minus0minus0
2 33 NminusHmiddotmiddotmiddotN 239 minus017 1542 003 I(8) 444 bminusb 0minus0minus153 NminusHmiddotmiddotmiddotO 225 minus011 1265 011 I(6) 159 sminusb 1minus1minus054 NminusHmiddotmiddotmiddotO 19 minus015 1495 022 I(7) 1205 sminusb 1minus0minus0
3 44 NminusHmiddotmiddotmiddotO 2 minus017 1585 02 I(13) 893 bminusb 1minus0minus058 OminusHmiddotmiddotmiddotO 177 minus021 1672 028 I(18) 1215 bminuss 1minus0minus0
4 5 NminusHmiddotmiddotmiddotO 22 minus015 1288 004 I(34) 135 bminusb 1minus0minus06 NminusHmiddotmiddotmiddotO 183 minus018 1748 025 I(37) 2158 bminusb 1minus0minus0
5 104 NminusHmiddotmiddotmiddotO 229 minus014 160 009 I(10) 097 bminusb 2minus0minus0105 NminusHmiddotmiddotmiddotO 195 minus016 1657 026 I(13) 1083 bminusb 2minus0minus0106 NminusHmiddotmiddotmiddotO 202 minus016 1507 014 I(16) 606 bminusb 2minus0minus0
6 90 NminusHmiddotmiddotmiddotO 214 minus015 1626 01 I(10) 301 bminusb 1minus0minus092 NminusHmiddotmiddotmiddotO 21 minus015 1634 015 I(16) 628 bminusb 1minus0minus0
aElec refers to the electrostatic energies bIntra refers to intramolecular HB type with ring size enclosed in the bracket cChain is composed of adonorminusacceptor pair as either from s (side chain) or b (backbone)
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2561
such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

such HBs do not necessarily decrease the strength of thecanonical HB (the HB with the shortest HB length)Figure 15 and Table 5 show six different networks of HBs in
different scenarios Figure 15a 15d and 15f show a 1minus0minus0 (eq1) network of HBs in α-helix extended strand and turnrespectively The HB angle for the two HBs varies from 128degto 165deg The HB with a higher angle has higher LPtoBDsignifying more covalent characteristics The electrostaticenergy for both HBs remains almost equivalent Howeverthe covalent contribution to the linear HB makes it stronger asaccepted by ka values A similar setup with an additional HBfrom the donor of a canonical HB (1minus1minus0) is shown in Figure15b Although the angle is lower compared to 1minus0minus0 asshown in Figure 15a there is a significant LPtoBD This is aconsequence of the extended electron density causing a higherconcentration of electrons for the canonical HB We observe araise in covalent energy for the canonical HB and thus in itsstrength The electrostatic energies do not have significantdifferences Figure 15c shows the network 1minus0minus0 with one ofthe HB as OminusHmiddotmiddotmiddotO from a side chain The LPtoBD is similarto 1minus1minus0 as shown in Figure 15b The electronegative natureof O atom increases the electrostatic energy The overallcontribution of electrostatic and covalent energies leads tosimilar ka values Figure 15e shows three HBs with the sameacceptor as network 2minus0minus0 Despite the HB length of 195 Aringfor the canonical HB LPtoBD is relatively high (1083 kcalmol) which contributes to its high ka value of 026 mdynAringThe elongated electron density of the three HBs stabilizes theenergy and therefore guides the protein to form a uniquesecondary structureAs stated by Feldblum and Arkin24 the network of HBs does
not necessarily decrease the strength of canonical HBs Wethus observe no pattern in the strength among variousnetworks of HBs The relationship of ka LPtoBD HBangle and electrostatic energy is shown in Figure S11 where
the quadratic fit curve relative to the HB length overlap forvarious networks
What Governs Hydrogen Bond Stability in ProteinsIn this section we detail the statistical results obtained forproperties of a HB based on the angle and amino acids inorder to investigate what drives strong HBs in proteins Figure16a shows the quadratic fit curve (middle) for all HBs relativeto the HB length We shifted the curve to plusmn005 to obtain twofit curves The two-shifted fit curve characterizes the strongand weak HBs (based on ka values) for the same HB lengthThus the points above the +005-shifted fit curve shouldcorrespond to strong HBs while those below the minus005-shiftedcurve should correspond to weak HBs relative to the HBlength Figure 16bminusf show the histogram for the two datapoints above the fit curve and below the fit curve for the HBangle LPtoBD electrostatic energy ka and HB lengthExcept for ka and HB angle other HB properties tend tooverlap denoting no significant differences The HB angle thusplays a strong role in distinguishing strong and weak HBswhere we observe a clear distinction for above fit curve pointsand below fit curve pointsWe further analyzed the amino acids involved in hydrogen
bonding ARG is the major donor for HBs contributing to 24of all HBs followed by LYS (13) Both have N in the sidechain to act as an HB donor Similarly they act as majoracceptors contributing to 10 and 9 for ARG and LYSrespectively The contribution of other residues is shown inTable S4 Table S5 shows the contribution of amino acids interms of a network of HBs and intramolecular HBs Howeverno intuitive relationship was found that clearly distinguishesamino acids based on HB properties
CONCLUSIONS
In this work we developed a new software Ef f icient Detectionof Hydrogen Bonds (EDHB) that systematically detects HBsbased on a nearest neighbors algorithm EDHB classifies inter-
Figure 16 Quadratic fit curve for all HBs relative to HB length is shifted by plusmn005 to obtain two fit curves The two-shifted fit curve characterizesthe strong and weak HBs for relatively the same HB length (a) shifted fit curves of HBs based on ka denoting strong (above fit curve) and weakHBs (below fit curve) (bminusf) histograms for each HB property generated for these HBs
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2562
and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

and intramolecular HBs as well as HB networks We couldshow that EDHB outperforms other commonly used methodsto detect HBs in terms of speed of execution An importantfeature of EDHB is that information from preceding quantumchemical studies (ie NBO data and second energy derivativeinformation) can be used to derive the electrostaticcovalentcharacter of the HBs and to calculate local-mode forceconstants as a quantitative measure of the intrinsic HB strengthvia the local-mode analysis program LModeA33
Utilizing EDHB we analyzed various HB properties in aprotein system for a diverse set of 163 proteins General trendsof HB strengths follow an inverse quadratic relationship withthe HB length as observed by local-mode force constantselectrostatic energies and covalent characteristics The mostcommon intramolecular HB is observed to be I(13) followedby I(10) and I(7) In general the strength of I(13) is higherthan that of I(10) followed by I(7) Analyzing various HBsbased on donorminusacceptor pairs highlights higher electrostaticand covalent characters for NminusHmiddotmiddotmiddotN and OminusHmiddotmiddotmiddotN type HBsNetworks of HBs are common in protein structures and playan important role in specific secondary structure orientationssuch as α-helixes and turns however they do not significantlyinfluence the HB strength A comprehensive analysis of thedistribution of local-mode force constants comes to theinteresting conclusion that the HB angle is the governingfactor determining the HB strengthAll EDHB scripts are provided at httpsgithubcom
ekrakaEDHB We further note that a linux binary file is alsoavailable for quick implementation on high-performancecomputers (HPC) and servers The applicability of EDHB indiscerning patterns of hydrogen bonding is extendable but notlimited to proteinminusligand interaction protein folding proteinminusprotein interaction molecular dynamics water ice clustersand improving force fields for proteins EDHB can be extendedto any molecular system provided the geometric information isknown reaching out to a broad computational chemistryaudience
ASSOCIATED CONTENTsı Supporting InformationThe Supporting Information is available free of charge athttpspubsacsorgdoi101021acsjpcb0c11392
Coordinates of the optimized geometry for the eightproteins PDB IDs for 155 proteins that were optimizedvia GFN2-xTB enlarged pictures of the eight proteinsEDHB time comparison EDHB output for the eightproteins various statistical plots made from the outputfrom the GFN2-xTB method (PDF)
AUTHOR INFORMATIONCorresponding AuthorElfi Kraka minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States orcidorg0000-0002-9658-5626 Email ekrakagmailcom
AuthorsNiraj Verma minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Yunwen Tao minus Department of Chemistry Southern MethodistUniversity Dallas Texas United States
Complete contact information is available athttpspubsacsorg101021acsjpcb0c11392
NotesThe authors declare no competing financial interest
ACKNOWLEDGMENTS
The authors thank SMU for its generous supercomputerresources This work was financially supported by the NationalScience Foundation Grants CHE 1464906 and DeanrsquosResearch Council Fellowship Southern Methodist University
REFERENCES(1) Hubbard R E Kamran Haider M Encyclopedia of Life SciencesJohn Wiley amp Sons Ltd Chichester UK 2010(2) Thapa B Raghavachari K Energy Decomposition Analysis ofProtein-Ligand Interactions Using Molecules-in-Molecules Fragmen-tation-Based Method J Chem Inf Model 2019 59 3474minus3484(3) Pace C N Fu H Fryar K L Landua J Trevino S RSchell D Thurlkill R L Imura S Scholtz J M Gajiwala K et alContribution of Hydrogen Bonds to Protein Stability Protein Sci2014 23 652minus661(4) Mirsky A E Pauling L On the Structure of Native Denaturedand Coagulated Proteins Proc Natl Acad Sci U S A 1936 22 439minus447(5) Pauling L Corey R B Branson H R The Structure ofProteins Two Hydrogen-Bonded Helical Configurations of thePolypeptide Chain Proc Natl Acad Sci U S A 1951 37 205minus211(6) Pauling L Corey R B Configurations of Polypeptide ChainsWith Favored Orientations Around Single Bonds Two New PleatedSheets Proc Natl Acad Sci U S A 1951 37 729minus740(7) Sippl M J Ortner M Jaritz M Lackner P Floumlckner HHelmholtz Free Energies of Atom Pair Interactions in ProteinsFolding Des 1996 1 289minus298(8) Campos L A Cuesta-Loacutepez S Loacutepez-Llano J Falo FSancho J A Double-Deletion Method to Quantifying IncrementalBinding Energies in Proteins from Experiment Example of aDestabilizing Hydrogen Bonding Pair Biophys J 2005 88 1311minus1321(9) Herschlag D Pinney M M Hydrogen Bonds Simple after AllBiochemistry 2018 57 3338minus3352(10) Scheiner S Contributions of NHmiddotmiddotmiddotO and CHmiddotmiddotmiddotO HydrogenBonds to the Stability of β-Sheets in Proteins J Phys Chem B 2006110 18670minus18679(11) Pace C N Scholtz J M Grimsley G R Forces StabilizingProteins FEBS Lett 2014 588 2177minus2184(12) Fried S D Boxer S G Measuring Electric Fields andNoncovalent Interactions Using the Vibrational Stark Effect AccChem Res 2015 48 998minus1006(13) Sticke D F Presta L G Dill K A Rose G D HydrogenBonding in Globular Proteins J Mol Biol 1992 226 1143minus1159(14) Alber T Dao-Pin S Wilson K Wozniak J A Cook S PMatthews B W Contributions of Hydrogen Bonds of Thr 157 to theThermodynamic Stability of Phage T4 Lysozyme Nature 1987 33041minus46(15) Green S M Meeker A K Shortle D Contributions of thePolar Uncharged Amino Acids to the Stability of StaphylococcalNuclease Evidence for Mutational Effects on the free Energy of theDenatured State Biochemistry 1992 31 5717minus5728(16) Serrano L Kellis J T Cann P Matouschek A Fersht A RThe Folding of an Enzyme J Mol Biol 1992 224 783minus804(17) Takano K Yamagata Y Funahashi J Hioki Y KuramitsuS Yutani K Contribution of Intra- and Intermolecular HydrogenBonds to the Conformational Stability of Human LysozymeBiochemistry 1999 38 12698minus12708(18) Sarkhel S Desiraju G R N-HmiddotmiddotmiddotO O-HmiddotmiddotmiddotO and C-HmiddotmiddotmiddotOHydrogen Bonds in Protein-Ligand Complexes Strong and WeakInteractions in Molecular Recognition Proteins Struct Funct Genet2004 54 247minus259
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2563
(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

(19) Tao Y Zou W Jia J Li W Cremer D Different Ways ofHydrogen Bonding in Water - Why Does Warm Water Freeze Fasterthan Cold Water J Chem Theory Comput 2017 13 55minus76(20) Tao Y Zou W Kraka E Strengthening of HydrogenBonding with the Push-Pull Effect Chem Phys Lett 2017 685 251minus258(21) Baker E Hubbard R Hydrogen Bonding in GlobularProteins Prog Biophys Mol Biol 1984 44 97minus179(22) Gray T Matthews B Intrahelical Hydrogen Bonding ofSerine Threonine and Cysteine Residues within α-Helices and itsRelevance to Membrane-bound Proteins J Mol Biol 1984 175 75minus81(23) Ballesteros J A Deupi X Olivella M Haaksma E EPardo L Serine and Threonine Residues Bend α-Helices in the χ1 =g- Conformation Biophys J 2000 79 2754minus2760(24) Feldblum E S Arkin I T Strength of a Bifurcated H bondProc Natl Acad Sci U S A 2014 111 4085minus4090(25) Alkorta I Elguero J Frontera A Not Only Hydrogen BondsOther Noncovalent Interactions Crystals 2020 10 180(26) Summers T J Daniel B P Cheng Q DeYonker N JQuantifying Protein Contact Networks through Interaction EnergiesJ Chem Inf Model 2019 59 5034minus5044(27) Lucarini M Pedrielli P Pedulli G F Cabiddu S FattuoniC Bond Dissociation Energies of O-H Bonds in Substituted Phenolsfrom Equilibration Studies J Org Chem 1996 61 9259minus9263(28) Deepak R N V K Sankararamakrishnan R UnconventionalN-HmiddotmiddotmiddotN Hydrogen Bonds Involving Proline Backbone Nitrogen inProtein Structures Biophys J 2016 110 1967minus1979(29) Remer L C Jensen J H Toward a General Theory ofHydrogen Bonding The Short Strong Hydrogen Bond [HOHmiddotmiddotmiddotOH] J Phys Chem A 2000 104 9266minus9275(30) Klein R A Electron Density Topological Analysis of HydrogenBonding in Glucopyranose and Hydrated Glucopyranose J AmChem Soc 2002 124 13931minus13937(31) Cremer D Kraka E From Molecular Vibrations to BondingChemical Reactions and Reaction Mechanism Curr Org Chem2010 14 1524minus1560(32) Konkoli Z Cremer D A New Way of Analyzing VibrationalSpectra I Derivation of Adiabatic Internal Modes Int J QuantumChem 1998 67 1minus9(33) Kraka E Zou W Tao Y Decoding Chemical Informationfrom Vibrational Spectroscopy Data Local Vibrational Mode TheoryWIREs Comput Mol Sci 2020 10 (5) No e1480(34) Kraka E Freindorf M Cremer D Chiral Discrimination byVibrational Spectroscopy Utilizing Local Modes Chirality 2013 25185minus196(35) Zhang X Dai H Yan H Zou W Cremer D B-HπInteraction A New Type of Nonclassical Hydrogen Bonding J AmChem Soc 2016 138 4334minus4337(36) Kalescky R Kraka E Cremer D Local Vibrational Modes ofthe Formic Acid Dimer - The Strength of the Double HydrogenBond Mol Phys 2013 111 1497minus1510(37) Kalescky R Zou W Kraka E Cremer D Local VibrationalModes of the Water Dimer - Comparison of Theory and ExperimentChem Phys Lett 2012 554 243minus247(38) Freindorf M Kraka E Cremer D A Comprehensive Analysisof Hydrogen Bond Interactions Based on Local Vibrational ModesInt J Quantum Chem 2012 112 3174minus3187(39) Oliveira V Kraka E Cremer D The Intrinsic Strength of theHalogen Bond Electrostatic and Covalent Contributions Describedby Coupled Cluster Theory Phys Chem Chem Phys 2016 1833031minus33046(40) Oliveira V Kraka E Cremer D Quantitative Assessment ofHalogen Bonding Utilizing Vibrational Spectroscopy Inorg Chem2017 56 488minus502(41) Oliveira V Cremer D Transition from Metal-ligand Bondingto Halogen Bonding Involving a Metal as Halogen Acceptor a Studyof Cu Ag Au Pt and Hg complexes Chem Phys Lett 2017 68156minus63
(42) Setiawan D Kraka E Cremer D Strength of the PnicogenBond in Complexes Involving Group Va Elements N P and As JPhys Chem A 2015 119 1642minus1656(43) Setiawan D Kraka E Cremer D Description of PnicogenBonding with the help of Vibrational Spectroscopy - The MissingLink Between Theory and Experiment Chem Phys Lett 2014 614136minus142(44) Setiawan D Cremer D Super-pnicogen Bonding in theRadical Anion of the Fluorophosphine Dimer Chem Phys Lett 2016662 182minus187(45) Samet H Fault Detection and Diagnosis in Industrial Systems 1sted Addison-Wesley Boston MA 1990(46) Derewenda Z S Lee L Derewenda U The Occurence of C-H middotmiddotmiddot O Hydrogen Bonds in Proteins J Mol Biol 1995 252 248minus262(47) Adhikary R Zimmermann J Liu J Forrest R P Janicki TD Dawson P E Corcelli S A Romesberg F E Evidence of anUnusual N-HmiddotmiddotmiddotN Hydrogen Bond in Proteins J Am Chem Soc2014 136 13474minus13477(48) Newberry R W Raines R T A Prevalent IntraresidueHydrogen Bond Stabilizes Proteins Nat Chem Biol 2016 12 1084minus1088(49) Mejiacutea S Hernaacutendez-Peacuterez J M Sandoval-Lira J Sartillo-Piscil F Looking Inside the Intramolecular C-HmiddotmiddotmiddotO Hydrogen Bondin Lactams Derived from a-Methylbenzylamine Molecules 2017 22361(50) Jeffrey G A An Introduction to Hydrogen Bonding OxfordUniversity Press Oxford UK 1997 p 303(51) Langkilde A Kristensen S M Lo Leggio L Moslashlgaard AJensen J H Houk A R Navarro Poulsen J C Kauppinen SLarsen S Short Strong Hydrogen Bonds in Proteins A Case Study ofRhamnogalacturonan Acetylesterase Acta Crystallogr Sect D BiolCrystallogr 2008 D64 851minus63(52) Harris T K Mildvan A S High-Precision Measurement ofHydrogen Bond Lengths in Proteins by Nuclear Magnetic ResonanceMethods Proteins Struct Funct Genet 1999 35 275minus282(53) Berman H M Westbrook J Feng Z Gilliland G Bhat TN Weissig H Shindyalov I N Bourne P E The Protein DataBank Nucleic Acids Res 2000 28 235minus242(54) McGibbon R T Beauchamp K A Harrigan M P KleinC Swails J M Hernaacutendez C X Schwantes C R Wang L-PLane T J Pande V S MDTraj A Modern Open Library for theAnalysis of Molecular Dynamics Trajectories Biophys J 2015 1091528minus1532(55) Cormen T H Leiserson C E Rivest R L Stein CIntroduction to Algorithms 2nd ed MIT Press Cambridge MA 2001pp 531minus539(56) Wilson E B Decius J C Cross P C Molecular VibrationsMcGraw-Hill New York 1955(57) Wilson E B Jr A Method of Obtaining the Expanded SecularEquation for the Vibration Frequencies of a Molecule J Chem Phys1939 7 1047(58) Oliveira V Cremer D Kraka E The Many Facets ofChalcogen Bonding Described by Vibrational Spectroscopy J PhysChem A 2017 121 6845minus6862(59) Sethio D Oliveira V Kraka E Quantitative Assessment ofTetrel Bonding Utilizing Vibrational Spectroscopy Molecules 201823 2763(60) Kalescky R Kraka E Cremer D Description of Aromaticitywith the Help of Vibrational Spectroscopy Anthracene andPhenanthrene J Phys Chem A 2014 118 223minus237(61) Setiawan D Kraka E Cremer D Quantitative Assessment ofAromaticity and Antiaromaticity Utilizing Vibrational Spectroscopy JOrg Chem 2016 81 9669minus9686(62) Li Y Oliveira V Tang C Cremer D Liu C Ma J ThePeculiar Role of the Au3 Unit in Aum Clusters σ-Aromaticity of theAu5Zn+ Ion Inorg Chem 2017 56 5793minus5803(63) Cremer D Kraka E Generalization of the Tolman ElectronicParameter The Metal-Ligand Electronic Parameter and the Intrinsic
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2564
Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565

Strength of the Metal-Ligand Bond Dalton Transac 2017 46 8323minus8338(64) Kalescky R Kraka E Cremer D New Approach to TolmanrsquosElectronic Parameter Based on Local Vibrational Modes Inorg Chem2014 53 478minus495(65) Setiawan D Kalescky R Kraka E Cremer D DirectMeasure of Metal-Ligand Bonding Replacing the Tolman ElectronicParameter Inorg Chem 2016 55 2332minus2344(66) Kraka E Larsson J A Cremer D In ComputationalSpectroscopy Methods Experiments and Applications Grunenberg JEd Wiley New York 2010(67) Kalescky R Kraka E Cremer D Identification of theStrongest Bonds in Chemistry J Phys Chem A 2013 117 8981minus8995(68) Kraka E Setiawan D Cremer D Re-evaluation of the BondLength - Bond Strength Rule The Stronger Bond is not Always theShorter Bond J Comput Chem 2016 37 130minus142(69) Setiawan D Kraka E Cremer D Hidden Bond AnomaliesThe Peculiar Case of the Fluorinated Amine Chalcogenides J PhysChem A 2015 119 9541minus9556(70) Humason A Zou W Cremer D 1111-Dimethyl-16-methano[10]annulene-An Annulene with an Ultralong CC Bond or aFluxional Molecule J Phys Chem A 2015 119 1666minus1682(71) Kalescky R Zou W Kraka E Cremer D QuantitativeAssessment of the Multiplicity of Carbon-Halogen Bonds Carbeniumand Halonium Ions with F Cl Br and I J Phys Chem A 2014 1181948minus1963(72) Kraka E Cremer D Characterization of CF Bonds withMultiple-Bond Character Bond Lengths Stretching Force Constantsand Bond Dissociation Energies ChemPhysChem 2009 10 686minus698(73) Verma N Tao Y Marcial B L Kraka E CorrelationBetween Molecular Acidity (pKa) and Vibrational Spectroscopy JMol Model 2019 25 48(74) Bent H A An Appraisal of Valance-bond Structures andHybridization in Compounds of the First-row Elements Chem Rev1961 61 275minus311(75) Halliday D Resnick R Walker J Fundamentals of Physics 5thed Wiley New York 1997(76) Mulliken R S Electronic Population Analysis on LCAO-MOMolecular Wave Functions I J Chem Phys 1955 23 1833minus1840(77) Frisch M J Trucks G W Schlegel H B Scuseria G ERobb M A Cheeseman J R Scalmani G Barone V PeterssonG A Nakatsuji H et al Gaussian 16 revision A03 Gaussian IncWallingford CT 2016(78) Dennington R Keith T A Millam J M Gauss View version509 Semichem Inc Shawnee Mission KS 2016(79) Vosko S H Wilk L Nusair M Accurate Spin-dependentElectron Liquid Correlation Energies for Local Spin DensityCalculations A Critical Analysis Can J Phys 1980 58 1200minus1211(80) Frisch M J Gaussian 09 revision A1 Gaussian IncWallingford CT 2009 (b) Stephens P J Devlin F J ChabalowskiC F Frisch M J J Phys Chem 1994 98 11623(81) Lee C Yang W Parr R G Development of the Colle-Salvetti Correlation-energy Formula into a Functional of the ElectronDensity Phys Rev B Condens Matter Mater Phys 1988 37 785minus789(82) Becke A D Density-functional Thermochemistry III TheRole of Exact Exchange J Chem Phys 1993 98 5648minus5652(83) Kuhn B Mohr P Stahl M Intramolecular HydrogenBonding in Medicinal Chemistry J Med Chem 2010 53 2601minus2611(84) Bartlett G J Newberry R W VanVeller B Raines R TWoolfson D N Interplay of Hydrogen Bonds and n-pi Interactionsin Proteins J Am Chem Soc 2013 135 18682minus18688(85) Chai J-D Head-Gordon M Long-range Corrected HybridDensity Functionals with Damped Atom-atom Dispersion Correc-tions Phys Chem Chem Phys 2008 10 6615
(86) Chai J-D Head-Gordon M Systematic Optimization ofLong-range Corrected Hybrid Density Functionals J Chem Phys2008 128 084106(87) Fadda E Woods R J Contribution of the EmpiricalDispersion Correction on the Conformation of Short AlaninePeptides Obtained by Gas-phase QM Calculations Can J Chem2013 91 859minus865(88) Rassolov V A Ratner M A Pople J A Redfern P CCurtiss L A 6-31G Basis Set for Third-row Atoms J ComputChem 2001 22 976minus984(89) Rassolov V A Pople J A Ratner M A Windus T L 6 -31G basis set for atoms K through Zn J Chem Phys 1998 1091223minus1229(90) Binning R C Curtiss L A Compact contracted basis sets forthird-row atoms Ga-Kr J Comput Chem 1990 11 1206minus1216(91) Francl M M Pietro W J Hehre W J Binkley J SGordon M S DeFrees D J Pople J A Self-Consistent MolecularOrbital Methods XXIII A Polarization-type Basis set for Second-RowElements J Chem Phys 1982 77 3654minus3665(92) Hariharan P C Pople J A The Influence of PolarizationFunctions on Molecular Orbital Hydrogenation Energies TheoreticaChimica Acta 1973 28 213minus222(93) Hehre W J Ditchfield R Pople J A Self-ConsistentMolecular Orbital Methods XII Further Extensions of Gaussian-Type Basis Sets for Use in Molecular Orbital Studies of OrganicMolecules J Chem Phys 1972 56 2257minus2261(94) Ditchfield R Hehre W J Pople J A Self-ConsistentMolecular-Orbital Methods IX An Extended Gaussian-Type Basisfor Molecular-Orbital Studies of Organic Molecules J Chem Phys1971 54 724minus728(95) Bannwarth C Caldeweyher E Ehlert S Hansen A PrachtP Seibert J Spicher S Grimme S Extended Tight-BindingQuantum Chemistry MethodsWIREs Comput Mol Sci 2020 11 (2)e1493(96) Case D A Belfon K Ben-Shalom I Y Brozell S RCerutti D S Cheatham T E III et al AMBER 2018 University ofCalifornia San Francisco CA 2018(97) Zou W Tao Y Freindorf M Makos M Z Verma NKraka E Local Vibrational Mode Analysis (LModeA) Computationaland Theoretical Chemistry Group (CATCO) Southern MethodistUniversity Dallas TX 2020(98) Pounds A J Valency and Bonding A Natural Bond OrbitalDonor-Acceptor Perspective (Frank Weinhold and Clark Landis) JChem Educ 2007 84 43(99) Kabsch W Sander C Dictionary of Protein SecondaryStructure Pattern Recognition of Hydrogen-bonded and GeometricalFeatures Biopolymers 1983 22 2577minus2637(100) Perutz M F New X-Ray Evidence on the Configuration ofPolypeptide Chains Polypeptide Chains in Poly-γ-benzyl-L-gluta-mate Keratin and Haeligmoglobin Nature 1951 167 1053minus1054
The Journal of Physical Chemistry B pubsacsorgJPCB Article
httpsdxdoiorg101021acsjpcb0c11392J Phys Chem B 2021 125 2551minus2565
2565




















