
Supramolecular Protein Engineering: DESIGN OF ZINC-STAPLED INSULIN HEXAMERS AS A LONG ACTING DEPOT
25
Supramolecular Protein Engineering DESIGN OF ZINC-STAPLED INSULIN HEXAMERS AS A LONG ACTING DEPOT * □ S Received for publication, January 19, 2010, and in revised form, February 18, 2010 Published, JBC Papers in Press, February 24, 2010, DOI 10.1074/jbc.C110.105825 Nelson B. Phillips ‡1 , Zhu-li Wan ‡1 , Linda Whittaker ‡ , Shi-Quan Hu ‡ , Kun Huang ‡ , Qing-xin Hua ‡ , Jonathan Whittaker ‡ , Faramarz Ismail-Beigi § , and Michael A. Weiss ‡§2 From the Departments of ‡ Biochemistry and § Medicine, Case Western Reserve University School of Medicine, Cleveland, Ohio 44106 Bottom-up control of supramolecular protein assembly can provide a therapeutic nanobiotechnology. We demonstrate that the pharmacological properties of insulin can be enhanced by design of “zinc staples” between hexamers. Paired (i, i4) His substitutions were introduced at an -helical surface. The crys- tal structure contains both classical axial zinc ions and novel zinc ions at hexamer-hexamer interfaces. Although soluble at pH 4, the combined electrostatic effects of the substitutions and bridging zinc ions cause isoelectric precipitation at neutral pH. Following subcutaneous injection in a diabetic rat, the analog effected glycemic control with a time course similar to that of long acting formulation Lantus. Relative to Lantus, however, the analog discriminates at least 30-fold more stringently between the insulin receptor and mitogenic insulin-like growth factor receptor. Because aberrant mitogenic signaling may be associated with elevated cancer risk, such enhanced specificity may improve safety. Zinc stapling provides a general strategy to modify the pharmacokinetic and biological properties of a sub- cutaneous protein depot. Supramolecular chemistry envisages the construction of novel materials and nanoscale devices ranging from molecular sensors to stimulus-responsive polymers (1, 2). Spatial organi- zation may be achieved either by templating (design from the top down) or through self-assembly of molecular components (bottom up). Connective tissue provides an example of the bot- tom-up design of a biomaterial based on hierarchical self-as- sembly of the collagen triple helix. Can such general principles be exploited in pharmacology? The present study explores the application of protein engineering (3) to the supramolecular chemistry of a therapeutic subcutaneous depot. Bottom-up control is accomplished by the pH-dependent binding of metal ions within and between self-assembled structures. A model is provided by insulin, a protein containing two chains, A (21 residues) and B (30 residues) (Fig. 1A) (4). The hormone is protected from misfolding in -cells by Zn 2 -stabilized assembly and microcrystallization (5). Storage hexamers dissociate on secretion, enabling insulin to function as a dilute zinc-free mono- mer. Zn 2 -stabilized hexamers are also employed in pharmaceu- tical formulations to delay insulin degradation (6). Treatment of diabetes mellitus increasingly employs insulin analogs with altered pharmacokinetics (7). Such pharmacokinetic “tuning” has enabled more effective glycemic control. Two classes of analogs extend the properties of wild-type formulations. More rapid acting analogs are designed to limit self-assembly (8) or accelerate disassembly (9). Longer acting analogs are designed to promote self-assembly in the subcutaneous depot (10) or retard disassembly (11, 12). These classes each exploit an inverse relationship between the size of a subcutaneous protein complex and rate of capillary absorption (8, 9, 12). Although insulin is highly soluble at pH values 7 or 5, between these limits the protein reversibly undergoes isoelec- tric precipitation (13). This property underlies the design of long acting analog insulin glargine (the active ingredient of Lantus) (12). Its B-chain is extended by paired Arg residues (Fig. 1A, top, red) whose positively charged side chains shift the pI from 5.8 to 7.0 (11, 12). Lantus is formulated at pH 4 as a clear unbuffered solution that, following subcutaneous injection, undergoes pH-dependent precipitation to form a slow release depot (11). Insulin glargine exhibits increased cross-binding to the type 1 insulin-like growth factor receptor (IGF-1R), 3 a ho- molog of the insulin receptor (IR) (12, 14). Augmented mito- genic signaling inhibits apoptosis and can drive the prolifera- tion of cancer cells (14). The impaired specificity of the analog has thus raised safety concerns (15). A recent clinical case con- trol study of Lantus reported a dose-dependent increase in IGF- 1R-related malignancies (16). This statistical analysis and its clinical implications have provoked intense debate. 4 We present a new strategy for design of long acting depots: engineered “zinc staples” between protein assemblies. The essential idea envisaged a zinc-dependent switch between an insoluble subcutaneous depot and a soluble metabolic signal. To implement this strategy, paired His substitutions (Glu A4 3 His and Thr A8 3 His; Fig. 1A, bottom) were introduced at posi- tions (i, i4) of an -helix (Fig. 1B), thus recapitulating part of a Zn 2 -binding site (17). Such paired His elements are ubiqui- tous among classical zinc fingers (supplemental Fig. S1). Posi- tions A4 and A8 were chosen as sites of modification to avoid * This work was supported, in whole or in part, by National Institutes of Health Grant DK069764 (to M. A. W.). This work was also supported by grants from the American Diabetes Association (to N. B. P. and M. A. W.) and Dietrich Diabetes Research Institute (to N. B. P. and F. I.-B.). This article is a contribution from the Cleveland Center for Membrane and Structural Biology. M. A. W. is a founding shareholder of Thermalin Diabetes, Inc. (Cleveland, OH). This article was selected as a Paper of the Week. The atomic coordinates and structure factors (code 3KQ6) have been deposited in the Protein Data Bank, Research Collaboratory for Structural Bioinformatics, Rutgers University, New Brunswick, NJ (http://www.rcsb.org/). □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental Methods and references, supplemental Figs. S1–S6, and supplemental Tables S1–S4. 1 Both authors contributed equally to this work. 2 To whom correspondence should be addressed. E-mail: michael.weiss@ case.edu. 3 The abbreviations used are: IGF, insulin-like growth factor; IGF-1R, type 1 IGF receptor; IR, insulin receptor; DKP, substitutions Asp B10 , Lys B28 , and Pro B29 . 4 Concerns have been raised about the statistical methods employed by Hemkens et al. (16) in light of inconsistent results of other studies (33). THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 16, pp. 11755–11759, April 16, 2010 © 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. APRIL 16, 2010 • VOLUME 285 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 11755 REPORT This paper is available online at www.jbc.org by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from by guest on September 3, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Supramolecular Protein Engineering: DESIGN OF ZINC-STAPLED INSULIN HEXAMERS AS A LONG ACTING DEPOT

Supramolecular ProteinEngineeringDESIGN OF ZINC-STAPLED INSULINHEXAMERS AS A LONG ACTING DEPOT*□S �
Received for publication, January 19, 2010, and in revised form, February 18, 2010Published, JBC Papers in Press, February 24, 2010, DOI 10.1074/jbc.C110.105825
Nelson B. Phillips‡1, Zhu-li Wan‡1, Linda Whittaker‡,Shi-Quan Hu‡, Kun Huang‡, Qing-xin Hua‡, Jonathan Whittaker‡,Faramarz Ismail-Beigi§, and Michael A. Weiss‡§2
From the Departments of ‡Biochemistry and §Medicine, Case WesternReserve University School of Medicine, Cleveland, Ohio 44106
Bottom-up control of supramolecular protein assembly canprovide a therapeutic nanobiotechnology.We demonstrate thatthe pharmacological properties of insulin can be enhanced bydesign of “zinc staples” between hexamers. Paired (i, i�4) Hissubstitutions were introduced at an �-helical surface. The crys-tal structure contains both classical axial zinc ions and novelzinc ions at hexamer-hexamer interfaces. Although soluble atpH 4, the combined electrostatic effects of the substitutions andbridging zinc ions cause isoelectric precipitation at neutral pH.Following subcutaneous injection in a diabetic rat, the analogeffected glycemic control with a time course similar to that oflong acting formulation Lantus�. Relative to Lantus, however,the analog discriminates at least 30-fold more stringentlybetween the insulin receptor and mitogenic insulin-like growthfactor receptor. Because aberrant mitogenic signaling may beassociated with elevated cancer risk, such enhanced specificitymay improve safety. Zinc stapling provides a general strategy tomodify the pharmacokinetic and biological properties of a sub-cutaneous protein depot.
Supramolecular chemistry envisages the construction ofnovel materials and nanoscale devices ranging from molecularsensors to stimulus-responsive polymers (1, 2). Spatial organi-zation may be achieved either by templating (design from thetop down) or through self-assembly of molecular components(bottom up). Connective tissue provides an example of the bot-tom-up design of a biomaterial based on hierarchical self-as-
sembly of the collagen triple helix. Can such general principlesbe exploited in pharmacology? The present study explores theapplication of protein engineering (3) to the supramolecularchemistry of a therapeutic subcutaneous depot. Bottom-upcontrol is accomplished by the pH-dependent binding of metalions within and between self-assembled structures.Amodel is providedby insulin, a protein containing two chains,
A (21 residues) and B (30 residues) (Fig. 1A) (4). The hormone isprotected frommisfolding in �-cells by Zn2�-stabilized assemblyand microcrystallization (5). Storage hexamers dissociate onsecretion, enabling insulin to function as a dilute zinc-free mono-mer. Zn2�-stabilized hexamers are also employed in pharmaceu-tical formulations to delay insulin degradation (6). Treatment ofdiabetesmellitus increasingly employs insulin analogswith alteredpharmacokinetics (7). Suchpharmacokinetic “tuning”has enabledmore effective glycemic control. Two classes of analogs extend theproperties of wild-type formulations. More rapid acting analogsare designed to limit self-assembly (8) or accelerate disassembly(9).Longeractinganalogsaredesigned topromote self-assembly inthe subcutaneous depot (10) or retard disassembly (11, 12). Theseclasses each exploit an inverse relationship between the size of asubcutaneous protein complex and rate of capillary absorption(8, 9, 12).Although insulin is highly soluble at pH values �7 or �5,
between these limits the protein reversibly undergoes isoelec-tric precipitation (13). This property underlies the design oflong acting analog insulin glargine (the active ingredient ofLantus�) (12). Its B-chain is extended by paired Arg residues(Fig. 1A, top, red) whose positively charged side chains shift thepI from5.8 to 7.0 (11, 12). Lantus is formulated at pH4 as a clearunbuffered solution that, following subcutaneous injection,undergoes pH-dependent precipitation to form a slow releasedepot (11). Insulin glargine exhibits increased cross-binding tothe type 1 insulin-like growth factor receptor (IGF-1R),3 a ho-molog of the insulin receptor (IR) (12, 14). Augmented mito-genic signaling inhibits apoptosis and can drive the prolifera-tion of cancer cells (14). The impaired specificity of the analoghas thus raised safety concerns (15). A recent clinical case con-trol study of Lantus reported a dose-dependent increase in IGF-1R-related malignancies (16). This statistical analysis and itsclinical implications have provoked intense debate.4We present a new strategy for design of long acting depots:
engineered “zinc staples” between protein assemblies. Theessential idea envisaged a zinc-dependent switch between aninsoluble subcutaneous depot and a soluble metabolic signal.To implement this strategy, paired His substitutions (GluA43His and ThrA83His; Fig. 1A, bottom) were introduced at posi-tions (i, i�4) of an �-helix (Fig. 1B), thus recapitulating part ofa Zn2�-binding site (17). Such paired His elements are ubiqui-tous among classical zinc fingers (supplemental Fig. S1). Posi-tions A4 and A8 were chosen as sites of modification to avoid
* This work was supported, in whole or in part, by National Institutes of HealthGrant DK069764 (to M. A. W.). This work was also supported by grants fromthe American Diabetes Association (to N. B. P. and M. A. W.) and DietrichDiabetes Research Institute (to N. B. P. and F. I.-B.). This article is a contributionfrom the Cleveland Center for Membrane and Structural Biology. M. A. W. is afounding shareholder of Thermalin Diabetes, Inc. (Cleveland, OH).
� This article was selected as a Paper of the Week.The atomic coordinates and structure factors (code 3KQ6) have been deposited in
the Protein Data Bank, Research Collaboratory for Structural Bioinformatics,Rutgers University, New Brunswick, NJ (http://www.rcsb.org/).
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Methods and references, supplemental Figs. S1–S6, andsupplemental Tables S1–S4.
1 Both authors contributed equally to this work.2 To whom correspondence should be addressed. E-mail: michael.weiss@
case.edu.
3 The abbreviations used are: IGF, insulin-like growth factor; IGF-1R, type 1 IGFreceptor; IR, insulin receptor; DKP, substitutions AspB10, LysB28, and ProB29.
4 Concerns have been raised about the statistical methods employed byHemkens et al. (16) in light of inconsistent results of other studies (33).
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 16, pp. 11755–11759, April 16, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
APRIL 16, 2010 • VOLUME 285 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 11755
REPORT This paper is available online at www.jbc.org
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

structural perturbation, preserve activity (18), and maintainclassical zinc assembly (4, 19). Choice of this A-chain �-helixwas further motivated by potentially favorable effects of itsmodification on the ratio of IR:IGF-1R binding (20, 21). Zinc-stapled insulin hexamers exemplify a structure-based approachto the design of a subcutaneous protein depot. The stringentreceptor selectivity and supramolecular assembly of the analogmay enhance the safety and efficacy of insulin therapy.
EXPERIMENTAL PROCEDURES
Materials—Insulin was provided by Novo-Nordisk (Bags-værd, Denmark). Insulin glargine was obtained from Lantus(Sanofi-Aventis, Paris, France). Reagents for peptide synthesiswere as described (22).Synthesis of Insulin Analog—Variant insulin chains were pre-
pared by solid-phase synthesis (22). Wild-type S-sulfonateB-chain derivatives were obtained by oxidative sulfitolysis ofinsulin (23). Insulin analogs were prepared by chain combina-tion and purified as described (22, 23). The paired His A-chainsubstitutionswere also incorporated into engineeredmonomerDKP-insulin (HisB103 Asp, ProB283 Lys, and LysB293 Pro)(24). Yields were similar to that of wild-type insulin. Predictedmolecular masses were confirmed by mass spectrometry.Biochemical Assays—The effects of the substitutions on pI
relative to insulin and insulin glargine were measured bynative isoelectric-focusing gel electrophoresis (supplementalMethods). Solubility was tested in dilute HCl (pH 4.0) in thepresence of 0.52 mM ZnCl2, 185 mM glycerol, and 25 mMmeta-cresol and on dilution into a buffered solution of 10 mM Tris-HCl (pH 6–9) and 140 mM NaCl by the method of DiMarchiand co-workers (20) (supplemental Methods).Crystallography—Crystals were obtained by hanging-drop
vapordiffusionat roomtemperature in thepresenceof a1:1.7 ratioof Zn2�:protein monomer and a 3.5:1 ratio of phenol:proteinmonomer in Tris-HCl (25). Diffraction was observed using syn-chrotronradiationatawavelengthbetween1.0000and1.2398ÅattheAdvancedLightSource (beamline4.2.2;Berkeley,CA); crystalswere flash-frozen to 100 K. The lattice contained one TRf dimerper asymmetric unit. The structure was determined bymolecularreplacement (supplemental Methods). Of the 86 residues in therefinedmodel of the TRf dimer in the asymmetric unit (excluding8 Gly, 2 Pro, and 6 end residues), 79 residues (92%) lie in themostfavored Ramachandran region, and 7 residues (8%) lie in gener-ously allowed regions.
1H-NMR Spectroscopy—Native self-assembly was preventedby “DKP” substitutions as described (24). Spectra of [HisA4,HisA8]DKP-insulin were acquired in D2O at 700 MHz and25 °C. The analog was made 0.5 mM in 10 mM deuterated Tris-DCl (pD 7.6, directmeter reading) in the presence or absence ofequimolar ZnCl2.Receptor Binding Assays—IR (isoformB) and IGF-1R binding
assays were performed by a microtiter plate antibody captureassay (supplemental Methods) (21). Wild-type insulin, insulinglargine, and IGF-I provided controls. Data were analyzed bynon-linear regression using a two-site sequential model (21).The percentage of tracer bound in the absence of competingligand was �15% to avoid ligand depletion artifacts.
FIGURE 1. Sequence and structure of insulin analogs. A, insulinsequence and sites of modification in insulin glargine (Lantus) (upperpanel) and the present analog (lower panel). Wild-type A- and B-chains areshown in black and green, and disulfide bridges (A6 –A11, A7–B7, andA20 –B19) are shown as black lines. Glargine contains a 2-residue B-chainextension (ArgB31 and ArgB32) and substitution AsnA213 Gly (upper panel,red). Endogenous subcutaneous proteases may slowly remove one orboth Arg residues, in part alleviating its augmented mitogenicity (31). Thepresent analog contains substitutions GluA43 His and ThrA83 His (lowerpanel, red). Long acting analog insulin detemir (Levemir�, Novo-Nordisk)operates by attachment of an albumin-binding element (not shown) (32).B, ribbon model of insulin monomer depicting the portion of putativezinc-binding site (violet) formed by HisA4 and HisA8 (red) at the externalsurface of A1–A8 �-helix. A- and B-chain ribbons are shown in black andgreen. C and D, structures of wild-type (C) and variant (D) T3R3
f insulinhexamers. The two axial zinc ions within each hexamer are aligned at thecenter (violet), coordinated by trimer-related HisB10 side chains (light gray).The variant hexamer contains three non-classical zinc ions at T3 trimersurface (D, magenta spheres). Shown in red are side chains of HisA4, HisA8,and the third HisA4� from the adjoining hexamer. In each case, the A-chainsare shown in black, and B-chains are in green or blue (Rf-specific B1–B8�-helix). The wild-type structure was obtained from the Protein Data bank(entry 1TRZ). E, 2Fo � Fc electron density map (stereo pair contoured at 1�) showing a novel zinc-binding site formed by HisA4 and HisA8 in theT-state protomer. Distorted tetrahedral coordination is completed by res-idue A4�, which belongs to an Rf-state protomer in adjoining hexamer.
REPORT: Zinc-based Protein Design
11756 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 16 • APRIL 16, 2010
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Rodent Assay—Male Lewis rats (mean weight �300 g) wererendered diabetic by streptozotocin (26). The effects of insulinanalogs on blood glucose concentration following subcutane-ous injection were assessed using a clinical glucometer (Hypo-guard Advance Micro-Draw meter) in relation to wild-typeinsulin or buffer alone (16mgof glycerin, 1.6mg ofmeta-cresol,0.65 mg of phenol, and 3.8 mg of sodium phosphate (pH 7.4);Lilly diluent). Wild-type insulin and [HisA4, HisA8]insulin weremade zinc-free in the above buffer. [HisA4, HisA8]insulin andinsulin glargine were also dissolved in dilute HCl (pH 4) con-taining a 5.2:1 ratio of ZnCl2:insulin monomer, 25 mM meta-cresol, and 185mM glycerol. Rats were injected subcutaneouslywith 3.44 nmol of insulin or insulin analogs (�12–13.7 nmol) in100 �l of buffer per rat (for wild-type insulin, this correspondsto 2 units/kg). For neutral zinc-free formulations, blood wasobtained from the tail every 10 min from 0–90 min. For acidiczinc-containing formulations, blood was obtained at times 0, 1,2, 4, 6, 10.8, and 24 h.
RESULTS AND DISCUSSION
The pI of [HisA4, HisA8]insulin was found to be 6.6. Althoughhighly soluble at pH 7.4 in the absence of zinc ions, the analogprecipitated at Zn2�:insulin molar ratios above 0.4. Solubilitywas largely regained at pH values �8.5 at room temperature,presumably due to deprotonation of N-terminal �-aminogroups. Such precipitation requires native protein assembly as1H-NMR studies of an engineeredmonomeric analog (24) indi-cated that the paired His element does not bind equimolarZn2� in this context (supplemental Fig. S2). Lattice contactsbetween wild-type hexamers in insulin crystals provided amodel for structural mechanisms of assembly-dependent zincbinding. The proximity of A4 and A8 side chains across suchcontact surfaces suggested that tetrahedral zinc-binding sitescould be formed between successive [HisA4, HisA8]insulin hex-amers. Novel A4–A8-mediated zinc coordinationmight there-fore be possible near sites of bound interfacial water moleculesin wild-type crystal lattices (Fig. 2A, blue spheres, shown in ste-reo in supplemental Fig. S3A).To test this hypothesis, we determined the crystal structure of
[HisA4,HisA8]insulin.Crystalsweregrown in thepresenceofZn2�
and phenol to yield T3R3f hexamers (27). The structure wasobtained by molecular replacement at 1.9 Å resolution (Table 1).Themode of hexamer assembly of the analog (Fig. 1D) is identicalto that of wild-type insulin (Fig. 1C). Respective conformations ofT and Rf protomers are essentially identical to those of wild-typeinsulin (supplemental Fig. S4 and supplemental Table S1). Notransmitted perturbations occur at receptor-binding surfaces (4).Wild-type and variant hexamers each contain two axial zinc
ions, one per T3 and R3f trimer (Fig. 1, C and D, overlaid violet
spheres). Coordination at each site is mediated by trimer-re-lated HisB10 side chains with distorted tetrahedral geometry(Fig. 2, C and D, light gray at center of hexamers, and sup-plemental Table S2). Stereo views of the variant TRf dimer inthe asymmetric unit and of the zinc-binding surface of the Tprotomer are provided in supplemental Fig. S5. In theR3
f trimer,the fourth ligand is a chloride ion; in the T3 trimer, this site(more exposed than in the R3
f trimer) exhibits partial occu-pancy by either a chloride ion or a bound water molecule
(supplemental Fig. S6). These features are consistent with wild-type structures (27). As is also observed in wild-type crystalsgrown under similar conditions, the R3
f trimer contains threebound phenol molecules (not shown). The A4 and A8 substitu-tions thus do not block the TR transition (22), a classical modelfor the reorganization of insulin on receptor binding (4).The variant T3R3
f hexamer contains three additional trimer-related zinc ions at the T-state surfaces (Fig. 1, B and D,magenta spheres; see also supplemental Fig. S5). These novelzinc ions are coordinated in part byHisA4 andHisA8 at an inter-facial site. Representative electron density at the peripheralzinc-binding site defines a distorted tetrahedral site (Fig. 1E,supplemental Table S2). Coordination is completed by a chlo-ride ion and a “stapled” HisA4 side chain belonging to an Rf
protomer of an adjoining hexamer (Fig. 1E, labeledA4�, and Fig.2B, brown arrows). Views of the opposing T and Rf faces ofadjoining hexamers are shown in Fig. 2C (90° rotated from theorientation shown in Fig. 2B). Binding of the chloride ion isstabilized by a network of threewatermolecules bound to theRf
protomer (Fig. 2D, blue spheres in stereo pairs); HisA8 in Rf isdisplaced from the zinc-binding site. The three non-classicalzinc ions thus bridge the T3 and Rf
3 trimers of adjacent hexam-ers (Fig. 2, B–D, magenta spheres, and in stereo in supple-mental Fig. S3B), in part displacing water molecules ordinar-ily bound at the wild-type interface (Fig. 2A, blue spheres).N-Zn2� bond distances and angles are similar to those of theaxial metal ion-binding sites (supplemental Table S2). Side-chain conformations of HisA4 and HisA8 differ between Tand Rf protomers (supplemental Table S3).Studies of hormone binding to IR and IGF-1Rwere undertaken
to assess relative affinities and receptor binding selectivity (Fig. 2Eand supplemental Table S4). Ligands were characterized as zinc-freemonomers. Relative to the binding of human insulin to IR andIGF-IR (Fig. 2E, solid and dotted green lines, respectively), insulinglargine (solid and dotted blue lines) exhibits 2-fold reduced affin-ity for IR and 3-fold enhanced affinity for IGF-1R. By contrast,[HisA4, HisA8]insulin exhibits native-like affinity for IR (Fig. 2E,solid red line) but 6-fold reduced affinity for IGF-1R (dotted redline shifted to right). Thus, although the receptor binding selectiv-ity of insulin glargine is impaired by �6-fold, that of [HisA4,HisA8]insulin is enhanced by 7.5 (�2.5)-fold. This represents animprovement of �30-fold relative to insulin glargine.The potency and duration of action of [HisA4, HisA8]insulin
were tested in diabetic rats in relation to insulin glargine (Fig. 2F).Glycemic control by long acting insulin analogs in rodents (5–10h) is less prolonged than in humans (18–24 h), presumably due tosmaller depot sizes (28). [HisA4, HisA8]insulin and insulin glarginewere dissolved (like Lantus) in dilute HCl (pH 4.0) with a Zn2�:insulin ratio of 5.2:1. The time course and extent of glycemic con-trol were similar on injection of the two analogs (Fig. 2F, red andblue lines). A rapid acting control was provided by zinc-freehuman insulin in Lilly diluent (Fig. 2F, green line). Because the ratsate only at night, the effects of daytime insulin injections wereinfluenced by diurnal fasting; controls were provided by injectionof diluent alone (Fig. 2F, brown line). Control studies were alsoundertaken of [HisA4, HisA8]insulin in neutral zinc-free Lilly dilu-ent; its time course was similar to that of wild-type insulin control
REPORT: Zinc-based Protein Design
APRIL 16, 2010 • VOLUME 285 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 11757
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

(not shown).Zinc-free glarginewasnot tested atneutral pHdue toits sparing solubility.Isoelectric precipitationof [HisA4,HisA8]insulin at pH7.4 is due
to the combined effects of two electrostatic changes: removal of awild-type negative charge by substitution GluA4 3 His and the
binding of three interfacial zinc ions per hexamer. Although insu-lin glargine contains 12 additional positive charges per hexamer,the net change in the formal charge of the [HisA4, HisA8] hexamermay be between 9 and 12, depending on the fractional Cl� occu-pancyat the fourth coordination site and indirect effects of protein
FIGURE 2. Zinc-stapled hexamers and biological activity. A, wild-type hexamer-hexamer packing. Left, in each hexamer, the upper trimer has T3 conformation, andthe lower trimer has R3
f conformation. Axial zinc ions are purple, and interfacial water molecules near residues A4 and A8 are blue. A-chains are shown in green, andB-chains are black. T and R protomers differ in B1–B9 secondary structure, extended (T) or helical (R); residues B1 and B2 are disordered in the “frayed” Rf-state. Right,expansion of boxed region at left. The violet sphere toward the bottom is the axial zinc ion of T3 trimer in the bottom hexamer. Brown arrows indicate Rf-state residuesGluA4� in the upper R3
f trimer. B, zinc-stapled hexamer-hexamer packing of [HisA4, HisA8]insulin; the upper trimer has T3 conformation, and the lower trimer has R3f
conformation. Axial zinc ions are violet, and A4–A8–A4�-coordinated zinc ions are magenta. A-chains are shown in green, and B-chains are in black. Right, expansion ofthe boxed region. Three novel zinc ions (magenta) are observed at the hexamer-hexamer interface. Brown arrows indicate the Rf-state side-chain HisA4� (from thebottom trimer of the top hexamer), which completes the interfacial zinc-binding sites. C, Corey-Pauling-Koltun models showing T and Rf faces of [HisA4, HisA8]insulinhexamer (left and right). The view shown is rotated by 90° relative to panel B. The three non-classical zinc ions (magenta spheres) are shown bound to HisA4 and HisA8
(sticks). White crosses indicate the position of Cl� ions; the coloring scheme is otherwise as in panel B. D, stereo pair showing a non-classical zinc ion (magenta sphere),a Cl� ion (gray), and three bound water molecules (blue) in relation to HisA4� (brown sticks) in the Rf protomer and HisA4-HisA8 (red) in the T protomer. The bound watermolecules participate in a hydrogen-bond network within Rf involving the side-chain carboxylate of GluB4�, para-OH of TyrB26�, and carbonyl oxygen of ProB28�
(labeled). E, competitive displacement assays probing high affinity binding of insulin or insulin analogs to IR (left-hand three curves) and low affinity cross-binding toIGF-1R (right-hand three curves). In each group, wild-type insulin is shown in green, glargine is in blue, and [HisA4, HisA8]insulin is in red. The enhanced receptor bindingselectivity of [HisA4, HisA8]insulin results from the leftward shift of its IR binding titration and rightward shift of its IGF-1R binding titration. Relative affinities anddissociation constants are provided in supplemental Table S4. Assays were performed in the absence of zinc ions. X, insulin; f, insulin glargine; �, [HisA4, HisA8]insulin;B/Bo denotes ratio of bound tracer to maximally bound tracer. F, streptozotocin-induced diabetic male rats were injected subcutaneously with wild-type insulin (X,green line), insulin glargine (f, blue line), [HisA4, HisA8]insulin (�, red line), or buffer control (Lilly diluent;F, brown line). Doses at time 0 were 3.44 nmol of wild-type insulin(20 �g in 100-�l injection volume), 12 nmol of insulin glargine (corresponding to 2.0 units of Lantus), 13.7 nmol of [HisA4, HisA8]insulin, and 100 �l of protein-free buffer(Lilly diluent). Blood glucose concentration was measured from the tip of the tail at indicated times. Each analog was tested in five rats (mean � S.E.); the experimentwas performed twice with similar results. Rats were fed 6–8 h following injections.
REPORT: Zinc-based Protein Design
11758 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 16 • APRIL 16, 2010
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

modifications at other titratable sites (such as the �-amino groupofGlyA1 and imidazole ringofHisB5).Although thepresent crystalstructure provides a model for zinc-bridged self-association sur-faces, thenumber anddistributionofboundzinc ionsmaydiffer ina non-crystalline aggregate formed on injection. In the future, thisissuemay be addressed by biochemical characterization of the ini-tial subcutaneous depot.
CONCLUDING REMARKS
The present study demonstrates the potential of interfacialzinc-binding sites, introduced by design (17, 29), to modify thepharmacokinetics of a protein in a subcutaneous depot. Such bot-tom-up control of assembly illustrates general principles ofsupramolecular chemistry and their application to nanobiotech-nology (1, 2).Optimal treatment of diabetes mellitus often requires com-
bined use of fast and slow acting insulin analogs (7, 30). Theprinciple of isoelectric precipitation underlies the prolongedaction of insulin glargine, an analog in broad clinical use (Lan-tus) (11, 12, 30). Although slow release of zinc-free monomersenables basal metabolic regulation, concerns have been raisedregarding IGF-1R-mediated mitogenicity (14) and possibledose-dependent cancer risk (15, 16). [HisA4, HisA8]insulinachieves slow release by formation of an insoluble hexamer sta-pled by interfacial zinc ions. Because the zinc-free monomerdiscriminates between IR and IGF-1R more stringently thanwild-type insulin or existing analogs, this or related “secondgeneration” analogs may enhance the safety of insulin therapy.Zinc stapling of insulin exemplifies a general strategy tomodifythe pharmacokinetic and biological properties of a subcutane-ous protein depot. The engineering of novel lattice contacts inprotein crystals can thus enable control of supramolecularassembly as a therapeutic protein nanotechnology.
Acknowledgments—We thank S. Wang and W. Jia for technicalassistance.
REFERENCES1. Whitesides, G. M. (2003) Nat. Biotechnol. 21, 1161–11652. Albrecht, M. (2007) Naturwissenschaften 94, 951–9663. Asuri, P., Bale, S. S., Karajanagi, S. S., and Kane, R. S. (2006) Curr. Opin.
Biotechnol. 17, 562–5684. Baker, E. N., Blundell, T. L., Cutfield, J. F., Cutfield, S. M., Dodson, E. J.,
Dodson, G. G., Hodgkin, D. M., Hubbard, R. E., Isaacs, N. W., and Reyn-olds, C. D. (1988) Philos. Trans. R. Soc. Lond. B Biol. Sci. 319, 369–456
5. Dodson, G., and Steiner, D. (1998) Curr. Opin. Struct. Biol. 8, 189–1946. Brange, J., and Langkjaer, L. (1997) Pharm. Biotechnol. 10, 343–4097. Hirsch, I. B. (2005) N. Engl. J. Med. 352, 174–1838. Brange, J., Ribel, U., Hansen, J. F., Dodson,G., Hansen,M. T., Havelund, S.,
Melberg, S. G., Norris, F., Norris, K., and Snel, L. (1988) Nature 333,679–682
9. DeFelippis, M. R., Chance, R. E., and Frank, B. H. (2001) Crit. Rev. Ther.Drug Carrier Syst. 18, 201–264
10. Dodson, G. G., Dodson, E. J., Turkenburg, J. P., and Bing, X. (1993) Bio-chem. Soc. Trans. 21, 609–614
11. Campbell, R. K., White, J. R., Levien, T., and Baker, D. (2001) Clin. Ther.23, 1938–1957
12. Le Roith, D. (2007) Diabetes Metab. Res. Rev. 23, 593–59913. Brange, J. (1987) Galenics of Insulin: The Physico-chemical and Pharma-
ceutical Aspects of Insulin and Insulin Preparations, pp. 18–29, Springer-Verlag, Berlin
14. Mayer, D., Shukla, A., and Enzmann, H. (2008) Arch. Physiol. Biochem.114, 38–44
15. Smith, U., and Gale, E. A. (2009) Diabetologia 52, 1699–170816. Hemkens, L. G., Grouven, U., Bender, R., Gunster, C., Gutschmidt, S.,
Selke, G. W., and Sawicki, P. T. (2009) Diabetologia 52, 1732–174417. Regan, L. (1993) Annu. Rev. Biophys. Biomol. Struct. 22, 257–28718. Kristensen, C., Kjeldsen, T., Wiberg, F. C., Schaffer, L., Hach, M., Have-
lund, S., Bass, J., Steiner, D. F., and Andersen, A. S. (1997) J. Biol. Chem.272, 12978–12983
19. Wan, Z., Xu, B., Huang, K., Chu, Y. C., Li, B., Nakagawa, S. H., Qu, Y., Hu,S. Q., Katsoyannis, P. G., and Weiss, M. A. (2004) Biochemistry 43,16119–16133
20. Kohn,W.D.,Micanovic, R.,Myers, S. L., Vick, A.M., Kahl, S. D., Zhang, L.,Strifler, B. A., Li, S., Shang, J., Beals, J. M., Mayer, J. P., and DiMarchi, R. D.(2007) Peptides 28, 935–948
21. Zhao, M., Wan, Z. L., Whittaker, L., Xu, B., Phillips, N. B., Katsoyannis,P. G., Ismail-Beigi, F., Whittaker, J., andWeiss, M. A. (2009) J. Biol. Chem.284, 32178–32187
22. Wan, Z. L., Huang, K., Hu, S. Q., Whittaker, J., and Weiss, M. A. (2008)J. Biol. Chem. 283, 21198–21210
23. Hu, S. Q., Burke, G. T., Schwartz, G. P., Ferderigos, N., Ross, J. B., andKatsoyannis, P. G. (1993) Biochemistry 32, 2631–2635
24. Weiss, M. A., Hua, Q. X., Lynch, C. S., Frank, B. H., and Shoelson, S. E.(1991) Biochemistry 30, 7373–7389
25. Wan, Z. L., Xu, B., Chu, Y. C., Katsoyannis, P. G., andWeiss, M. A. (2003)Biochemistry 42, 12770–12783
26. Saker, F., Ybarra, J., Leahy, P., Hanson, R. W., Kalhan, S. C., and Ismail-Beigi, F. (1998) Am. J. Physiol. 274, E984–E991
27. Ciszak, E., and Smith, G. D. (1994) Biochemistry 33, 1512–151728. Schneider, S., Weber, R., and Luippold, G. (2004) Arzneimittelforschung
54, 842–84629. Lu, Y., Berry, S. M., and Pfister, T. D. (2001) Chem. Rev. 101, 3047–308030. Baxter, M. A. (2008) Acta Diabetol. 45, 253–26831. Kuerzel, G. U., Shukla, U., Scholtz, H. E., Pretorius, S. G., Wessels, D. H.,
Venter, C., Potgieter, M. A., Lang, A. M., Koose, T., and Bernhardt, E.(2003) Curr. Med. Res. Opin. 19, 34–40
32. Raskin, P. (2007) Endocrinol. Metab. Clin. North Am. 36, 21–3233. Pocock, S. J., and Smeeth, L. (2009) Lancet 374, 511–513
TABLE 1Data collection and refinement statistics
�HisA4, HisA8insulin
Data collectionSpace group R3Cell dimensionsa, b, c (Å) 78.09, 78.09, 36.40�, �, � (°) 90.00, 90.00, 120.00
Resolution (Å) 32.05-1.90Rsym or Rmerge 0.057 (0.422)aI/�I 14.1 (3.0)aCompleteness (%) 99.5 (100.0)aRedundancy 5.49 (5.43)a
RefinementResolution (Å) 32.05-1.90No. of reflections 6475 (955)Rwork/Rfree 0.199/0.257No. of atomsProtein 818Ligand/ion 6Water 82
B-factorsProtein 42.28Ligand/ion 29.03Water 53.94
r.m.s.b deviationsBond lengths (Å) 0.008Bond angles (°) 1.2
a Highest resolution shell is shown in parentheses.b r.m.s., root mean square deviation.
REPORT: Zinc-based Protein Design
APRIL 16, 2010 • VOLUME 285 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 11759
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

1
Supplemental Information
for
Supramolecular Protein Engineering DESIGN OF ZINC-STAPLED HEXAMERS AS A LONG-ACTING DEPOT
Nelson B. Phillips, Zhu-li Wan, Linda Whittaker, Shi-Quan Hu, Qing-xin Hua, Jonathan Whittaker, Faramarz Ismail-Beigi, and Michael A. Weiss*
Table of Contents
Purpose of Supplement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 Supplemental Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 Supplemental References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5 Supplemental Figure S1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Supplemental Figure S2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8 Supplemental Figure S3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 Supplemental Figure S4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 Supplemental Figure S5 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 Supplemental Figure S6 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 Supplemental Table S1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 Supplemental Table S2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17 Supplemental Table S3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18 Supplemental Table S4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19

2
Purpose of Supplement
This Supplement contains methodological details concerning the synthesis of [HisA4, HisA8]-insulin by chain combination; isoelectric focusing gel electrophoresis; protein solubility assay; receptor-binding assays including expression and purification of the insulin receptor (IR isoform B) and the Type 1 IGF receptor (IGF-1R); and the crystallization and crystallographic analysis of the insulin analog. Six supplemental figures (Supplemental Figure S1-S6) and four supplemental tables (Supplemental Tables S1-S4) are also provided.
Supplemental Figure S1 provides a ribbon model of the classical zinc finger to highlight its HX3H α-helical element. In Supplemental Figure S2 are shown 1H-NMR spectra of an engineered insulin monomer with substitutions HisA4 and HisA8, demonstrating absence of zinc-ion binding. Stereo views of the salient hexamer-hexamer interface in the wild-type and variant crystal lattice are shown in Supplemental Figure S3. Supplemental Figure S4 provides an alignment of the T- and R-state protomers of [HisA4, HisA8]-insulin with a collection of wild-type crystallographic protomers, demonstrating retention of native-like main-chain conformation. In Supplemental Figure S5 are shown additional views of the crystal structure, highlighting the TRf dimer in the asymmetric unit and surface of the T-state protomer. Electron-density maps are shown in Supplemental Figure S6 comparing the three types of Zn-binding sites in the crystallographic hexamer of [HisA4, HisA8]-insulin (the axial Zn-binding site of the T3 trimer, the axial Zn-binding site of the Rf
3 trimer, and the novel A4-A8-related peripheral Zn-binding site).
Supplemental Table S1-A-D provides RMSD values for alignment of the respective T- and R-state protomers of [HisA4, HisA8]-insulin relative to a collection of crystal structures of wild-type insulin and unrelated insulin analogs obtained from the Protein Data Bank; the similarity between the analog RSMD values and control RMSD values indicates that the new structure falls within the envelope of prior structures as visually apparent in Supplemental Figure S4. Supplemental Table S2 provides zinc-binding geometries. In Supplemental Table S3 the side-chain conformations of the histidines are defined. In Supplemental Table S4 are given the absolute and relative hormone-receptor dissociation constants.
Supplemental Methods
Synthesis of Insulin Analogs. Chain combination (1) was effected by interaction of the S-sulfonated derivative of the A chain (41 mg) and B-chain analog (21 mg) in 0.1 M glycine buffer (pH 10.6, 10 ml) in the presence of dithiothreitol (7 mg) (2,3). CM-52 cellulose chromatography of each combination mixture enabled partial isolation of the hydrochloride form of the protein contaminated by free B-chain. Final purification was accomplished by reverse-phase HPLC. The predicted molecular mass of [HisA4, HisA8]-insulin was verified by MALDI mass spectrometry. The final yield (6.1 mg) were similar to those obtained in a control synthesis of wild-type insulin (4). The corresponding yield of [HisA4, HisA8]-DKP-insulin was 8.8 mg.
Isoelectric Focusing Electrophoresis. The pI values of insulin and insulin analogs in their native states were measured by IEF gel electrophoresis using pre-cast pH 3-10 IEF gels, (125 x 125 mm, 300 µ m, SERVALYT® Precotes® from SERVA Electrophoresis GmbH, Heidelberg; obtained from Crescent Chemical Co. Hauppauge, NY). The Precotes® were set up in a horizontal IEF apparatus, Multiphor II (Pharmacia Biotech) according to the manufacturer’s protocol. The unit was pre-cooled to 4° C using a circulating water bath (Brinkman), before

3
placing the PRECOTE IEF gel on electrophoresis bed coated with light mineral oil for efficient heat exchange. The gels were connected to electrodes using filter paper wicks wetted with Anode Fluid pH 3 and Cathode Fluid pH 10 (both from SERVA) at the two ends of the gel. Prior to loading the samples, the gel was pre-focused at an initial voltage setting of 200 volts and a final setting of 500 volts for 30 min using a high voltage power supply (LKB model 2197). After loading the samples and the IEF standards (5-10 µL, at a loading protein concentration of 5-10 µg), isoelectric focusing was performed at 500 – 2000 volts for 2 hrs or until the final voltage of 2000 volts was reached, after which focusing was continued for an additional 15 min. After IEF, the gel was fixed with 200 ml of 20% trichloroacetic acid for 20 min, rinsed for 1 min with 200 ml of deionized water and stained with Serva Violet 17 solution and destained with 86% phosphoric acid according to the SERVA manual protocols. The IEF standard proteins (from SERVA) used are as follows, with their respective pI’s in parentheses: horse heart cytochrome C, (10.7), bovine pancreas ribonuclease A (9.5), lens culinaris Lectin (8.3, 8.0, 7.8), horse muscle Myoglobin (7.4, 6.9), bovine erythrocytes Carbonic anhydrase (6.0), bovine milk β -lactoglobin (5.3, 5.2), soybean trypsin inhibitor (4.5), Aspergillus niger glucose oxidase (4.2), Aspergillus niger amyloglucosidase (3.5). The pI’s of the protein samples were determined by comparison to a linear regression plot of migration distance versus pH gradient of the IEF standards.
Solubility Assay. The pH-dependent solubility of the insulin analogs was evaluated by a modification of the method of DiMarchi and coworkers (Kohn, W. D., Micanovic, R., Myers, S. L., Vick, A. M., Kahl, S. D., Zhang, L., Strifler, B. A., Li, S., Shang, J., Beals, J. M., Mayer, J. P., and DiMarchi, R. D. Peptides 28, 935-48 (2007); Ref. 20 in main text). In brief, wild-type human insulin, insulin glargine or [HisA4, HisA8]-insulin were made 0.60 mM in an unbuffered solution containing dilute HCl at pH 4.0; the composition of the solution, similar to that employed in the pharmaceutical formulation Lantus (Sanofi-Aventis), contained 0.52 mM ZnCl2, 20 mg/ml of an 85% vol/vol glycerol solution (to a final concentration of 185 mM), and 2.7 mg/ml meta-cresol (25 mM) as antimicrobial preservative. Each of the three proteins exhibits a solubility in this pH 4.0 solution exceeding 0.60 mM. A series of identical aliquots (10 ml) was removed and diluted 50-fold into buffers at various pH values (in the range 5.0 – 9.0) to a final volume of 500 ml; respective pH values were then re-adjusted to be 5.0, 6.0, 7.4, 8.0, 8.5, and 9.0. The diluent was composed of 10 mM Tris-HCl and 140 mM NaCl with pH values adjusted by dilute HCl or NaOH. The multiple samples were then mixed 20 times by inversion and centrifuged for 5 min at 14,000 rpm in a micro-centrifuge. 200 µl of supernatant was then removed in duplicate from each tube and injected onto an analytical reverse-phase HPLC (C4 column; 25 cm x 0.46 cm) with an elution gradient of acetonitrile containing 0.1% trifluoroacetic acid. In each case a single elution peak was observed, and its area quantified by integration using vendor software (Waters, Inc.). The wild-type insulin values at pH 7.4-9.0 provided a control for losses unrelated to solubility; percent recoveries were typically in the range 85-90%. The solubility of insulin glargine at pH 7.4 was found to be between 1 and 2 µM in accord with the results of DiMarchi and coworkers. This limited solubility was similar at molar ratios of zinc-to-analog of 5.2:6 (i.e., 5.2 zinc ions per hexamer) and 2.2:6 (2.2 zinc ions per hexamer), consistent with an axial role of zinc ion in the glargine hexamer. The solubility of [HisA4, HisA8]-insulin at pH 7.4 was also found to be 1 and 2 µM at a molar ratio of 5.2 zinc ions per hexamer.

4
Plasmids of Receptor Expression. For expression of epitope-tagged IR and IGFR, the mammalian expression vector pcDNA3.1Zeo+ was obtained from InVitrogen and was modified for C-terminal epitope tagging by subcloning an in-frame oligonucleotide cassette encoding in-frame triple repeats of the FLAG M2 epitope (Asp-Tyr-Lys-Asp-Asp-Asp-Asp-Lys) between the BamHI and XbaI restriction sites. Respective cDNAs encoding IGFR and the B-isoform of IR were as previously described (5,6). They were modified for subcloning into the modified expression vector by introduction of a BamHI site encoding an in-frame C-terminal Gly-Ser linker at their 3’ ends just prior to the stop codon by site-directed mutagenesis.
Expression of Receptor cDNAs. DNA for transfection was prepared as previously described (7). The receptor cDNAs were expressed transiently in PEAK rapid cells using polyethyleneimine (8). Cells were harvested three days post-transfection when receptor expression was maximal. Lysis was accomplished in a buffer consisting of 0.15 M NaCl and 0.1M Tris-Cl (pH 8.0), containing 1% (v/v) Triton X-100 and a protease inhibitor cocktail (Roche). Lysates were stored at -80o C until required for assay.
Receptor Binding Assays. Respective IGFR and IR-B binding assays were performed by a modification of the microtiter plate antibody-capture assay that Whittaker and colleagues have described previously (6,7,9). Microtiter strip plates (Nunc Maxisorb) were incubated overnight at 4 oC with anti-FLAG IgG (100 µl/well of a 40 µg/ml solution in phosphate-buffered saline). Washing and blocking were performed as previously described (6,7,9). Detergent lysates of 293 PEAK cells transiently transfected with cDNAs encoding full-length IR-B or IGFR with C-terminal FLAG-tags were partially purified by wheat germ agglutinin (WGA) chromatography (10) to deplete lysates of receptor pre-cursors. Wheat-germ eluates were then incubated in the antibody-coated plates for 1 hour at room temperature to immobilize receptors. After extensive washing to remove unbound proteins, competitive binding assays with labeled insulin tracer (125I-[TyrA14]-insulin) or labeled IGF-I tracer (125I- Tyr31-IGF-I) and unlabeled insulin analogs were carried out as described (7,9). All insulin analogs were assayed with either insulin or IGF-I receptor as control ligands in the same set of assays. Binding data from homologous competition assays were analyzed by non-linear regression analysis using a 2-site sequential model to obtain dissociation constants for insulin and IGF-I (11,12). Binding data for heterologous competition experiments were analyzed by the method of Wang (13); this method uses an exact mathematical expression to describe the competitive binding of two different ligands to a receptor.
X-Ray Crystallography. Crystals were grown by hanging-drop vapor diffusion in the presence of a 1:1.7 ratio of Zn2+ to protein monomer and a 3.5:1 ratio of phenol to protein monomer in Tris-HCl buffer as described (14). Drops consisted of 1 µl of protein solution (8 mg/ml in 0.02 M HCl) mixed with 1 µl of reservoir solution (0.38 M Tris-HCl, 0.1 M sodium citrate, 9% acetone, 4.83 mM phenol, and 0.8mM zinc acetate at pH 8.4). Each drop was suspended over 1 ml of reservoir solution. Crystals were obtained at room temperature after two weeks. Data were collected from single crystals mounted in a rayon loop and flash frozen to 100º K. Reflections from 32.05-1.90 Å were measured with a CCD detector system on synchrotron radiation in Berkeley National Laboratory. Data were processed with the program DTREK. The crystal belongs to space group R3 with unit cell parameters: a=b=78.09 Å, c=36.40 Å, α=β=90º, γ=120º. The structure was determined by molecular replacement using CNS (15). Accordingly, a model was obtained using the native TR dimer (Protein Databank (PDB) identifier 1RWE following removal of all water molecules, zinc- and chloride ions). A translation-function search was performed using coordinates from the best solution for the

5
rotation function following analysis of data between 15.0 and 4.0 Å resolutions. Rigid-body refinement using CNS, employing overall anisotropic temperature factors and bulk-solvent correction, yielded values of 0.325 and 0.344 for R and Rfree, respectively, for data between 19.2 and 3.0 Å resolution. Between refinement cycles, 2Fo-Fc and Fo-Fc maps were calculated using data to 3.0 Å resolution; zinc and chloride ions and phenol molecules were built into the structure using the program O (16). Water molecules were calculated and checked using DDQ program (Focco Van Akker and Wim Hol, Acta Cryst. 1999, D55, 206-218). The geometry was continually monitored with PROCHECK (17); zinc ions and water molecules were built into the difference map as the refinement proceeded. Calculation of omit maps (especially in the first eight residues of B chain N terminus of each monomer) and further refinement were carried out using CNS (15), which implement maximum-likelihood torsion-angle dynamics and conjugate-gradient refinement. Statistics data are provided in Table 1 in main text.
Supplemental References 1. Chance, R. E., Hoffman, J. A., Kroeff, E. P., Johnson, M. G., Schirmer, W. E., and
Bormer, W. W. (1981) The production of human insulin using recombinant DNA technology and a new chain recombination procedure. In: Rich, D. H., and Gross, E. (eds). Peptides: Synthesis, Structure and Function; Proceedings of the Seventh American Peptide Symposium, Pierce Chemical Co., Rockford, IL
2. Hu, S. Q., Burke, G. T., Schwartz, G. P., Ferderigos, N., Ross, J. B., and Katsoyannis, P. G. (1993) Biochemistry 32, 2631-2635
3. Hua, Q. X., Hu, S. Q., Frank, B. H., Jia, W., Chu, Y. C., Wang, S. H., Burke, G. T., Katsoyannis, P. G., and Weiss, M. A. (1996) J. Mol. Biol. 264, 390-403
4. Weiss, M. A., Nakagawa, S. H., Jia, W., Xu, B., Hua, Q. X., Chu, Y. C., Wang, R. Y., and Katsoyannis, P. G. (2002) Biochemistry 41, 809-819
5. Whittaker, J., Okamoto, A. K., Thys, R., Bell, G. I., Steiner, D. F., and Hofmann, C. A. (1987) Proc. Natl. Acad. Sci. USA 84, 5237-5241
6. Mynarcik, D. C., Williams, P. F., Schäffer, L., Yu, G. Q., and Whittaker, J. (1997) J. Biol. Chem. 272, 18650-18655
7. Whittaker, J., and Whittaker, L. (2005) J. Biol. Chem. 280, 20932-20936 8. Pham, P. L., Kamen, A., and Durocher, Y. (2006) Mol. Biotechnol. 34, 225-237 9. Whittaker, J., Groth, A. V., Mynarcik, D. C., Pluzek, L., Gadsboll, V. L., and Whittaker,
L. J. (2001) J. Biol. Chem. 276, 43980-43986 10. Hedo, J. A., Harrison, L. C., and Roth, J. (1981) Biochemistry 20, 3385-3393 11. Christoffersen, C. T., Bornfeldt, K. E., Rotella, C. M., Gonzales, N., Vissing, H.,
Shymko, R. M., ten Hoeve, J., Groffen, J., Heisterkamp, N., and De Meyts, P. (1994) Endocrinology 135, 472-475
12. Levitzki, A. (1978) Quantitative aspects of allosteric mechanisms, Springer-Verlag, Berlin, New York
13. Wang, Z. X. (1995) FEBS Lett. 360, 111-114 14. Wan, Z., Xu, B., Chu, Y. C., Katsoyannis, P. G., and Weiss, M. A. (2003) Biochemistry
42, 12770-12783 15. Brunger, A. T., Adams, P. D., Clore, G. M., DeLano, W. L., Gros, P., Grosse-Kunstleve,
R. W., Jiang, J. S., Kuszewski, J., Nilges, M., Pannu, N. S., Read, R. J., Rice, L. M.,

6
Simonson, T., and Warren, G. L. (1998) Acta Crystallogr. D Biol. Crystallogr. 54, 905-921
16. Jones, T. A., Zou, J. Y., Cowan, S. W., and Kjeldgaard, M. (1991) Acta Crystallogr. A 47, 110-119
17. Laskowski, R. A., Macarthur, M. W., Moss, D. S., and Thornton, J. M. (1993) J. Appl. Crystallogr. 26, 283-291
18. Zhao, M., Wan, Z. L., Whittaker, L., Xu, B., Phillips, N. B., Katsoyannis, P. G., Ismail-Beigi, F., Whittaker, J., and Weiss, M. A. (2009) J. Biol. Chem. 284, 32178-32187

7
Supplemental Figure S1. Structure of a classical Zn finger. The sequence motif (XArXCX2,4CX4FX3LX3HX3HX; ligands in bold) encodes a ββα structure in which paired (i, i+3) histidines (red) on one edge of an α-helix (blue) contribute to tetrahedral coordination of an engaged zinc ion (magenta sphere). Paired substitutions GluA4→His and ThrA8 →His in insulin (bottom of Fig. 1A in main text) recapitulates this portion of the Zn finger (Fig. 1B of main text).

8
Supplemental Figure S2. 1H-NMR studies. Aromatic region of 700 MHz 1H-NMR spectra of engineered monomer [HisA4, HisA8]-DKP-insulin in 10 mM deuterated Tris-HCl (pD 7.6) in D2O in the absence (A) or presence (B) of equimolar ZnCl2. No changes are observed in His imidizole ring resonances. 2DNOESY and 2D-TOCYS spectra (data not shown) likewise indicate no significant changes in aromatic or aliphatic spin systems. The DKP template contains two substitutions in the classical dimer interface (ProB28→Lys and LysB29→Pro as in Humalog; Eli Lilly and Co.) and one substitution in the trimer interface (HisB10→Asp). The latter substitution precludes conventional zinc-mediated assembly (ref. 21 of main text).

9
Supplemental Figure S3. Stereo view of interfacial zinc ions in lattice assembly. This figure corresponds to Fig. 2a,b in main text. (A) Hexamer-hexamer packing in crystal lattice: in each hexamer the upper trimer has T3 conformation, and lower trimer Rf
3. Axial zinc ions are shown as purple spheres, and A4-A8-coordinated zinc ions as magneta spheres. The A-chains are shown in green, and B-chains in black. (B) Expansion of boxed region in panel A. Three novel zinc ions (magenta) are observed at hexamer-hexamer interface. Brown arrows indicate Rf-state side chain HisA4’ (from bottom trimer of top hexamer), which complete the zinc-binding sites. Purple sphere toward bottom is axial zinc ion of T3 trimer of bottom hexamer.
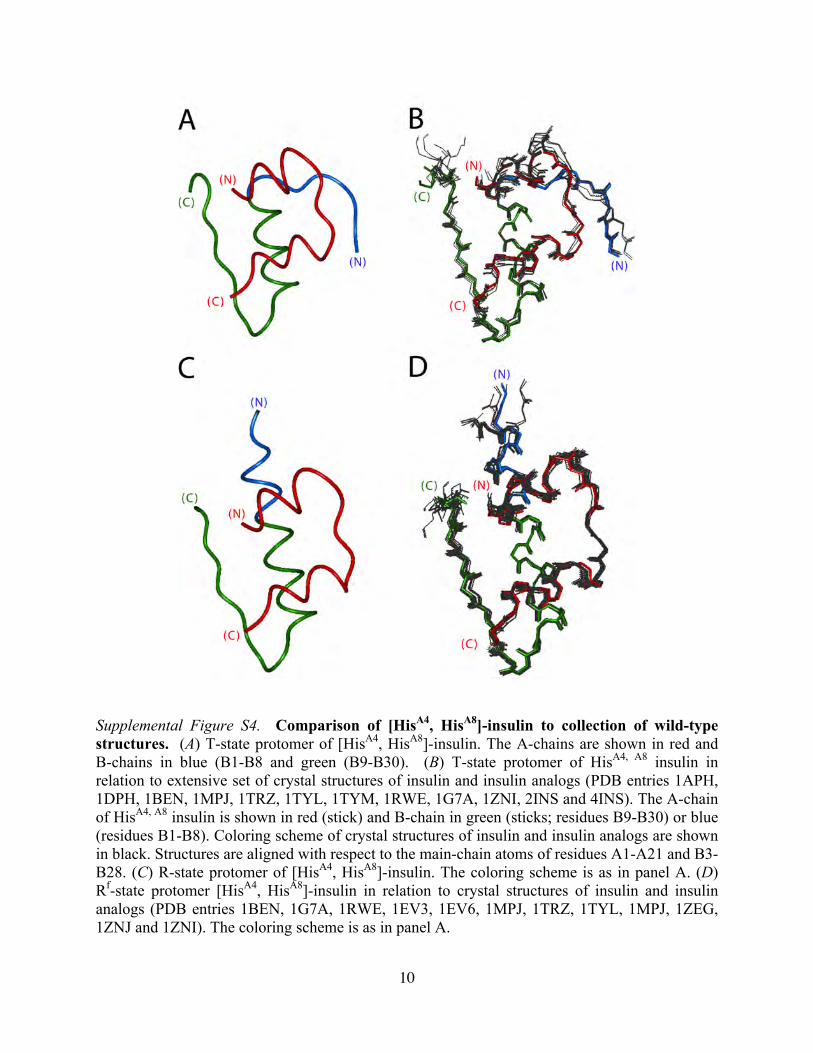
10
Supplemental Figure S4. Comparison of [HisA4, HisA8]-insulin to collection of wild-type structures. (A) T-state protomer of [HisA4, HisA8]-insulin. The A-chains are shown in red and B-chains in blue (B1-B8 and green (B9-B30). (B) T-state protomer of HisA4, A8 insulin in relation to extensive set of crystal structures of insulin and insulin analogs (PDB entries 1APH, 1DPH, 1BEN, 1MPJ, 1TRZ, 1TYL, 1TYM, 1RWE, 1G7A, 1ZNI, 2INS and 4INS). The A-chain of HisA4, A8 insulin is shown in red (stick) and B-chain in green (sticks; residues B9-B30) or blue (residues B1-B8). Coloring scheme of crystal structures of insulin and insulin analogs are shown in black. Structures are aligned with respect to the main-chain atoms of residues A1-A21 and B3-B28. (C) R-state protomer of [HisA4, HisA8]-insulin. The coloring scheme is as in panel A. (D) Rf-state protomer [HisA4, HisA8]-insulin in relation to crystal structures of insulin and insulin analogs (PDB entries 1BEN, 1G7A, 1RWE, 1EV3, 1EV6, 1MPJ, 1TRZ, 1TYL, 1MPJ, 1ZEG, 1ZNJ and 1ZNI). The coloring scheme is as in panel A.

11
Supplemental Figure S5. Additional views of crystal structure of [HisA4, HisA8]-insulin. (A) Stereo view of variant TRf insulin dimer (ribbon model). The A-chains of molecule 1 and 2 are shown in blue (A1-A8 α-helix) and black (A9-A21); B-chains in green. The HisA4- and HisA8 side chains are depicted as red sticks whereas the novel Zn2+ ion is magenta. (B) Space-filling representation of T-state promoter showing A4 and A8 side chains (red sticks) in relation to the protein surface, novel zinc ion (magenta), and HisA4’ from adjoining hexamer. White cross (x) indicates position of chloride anion. The chain color code is as in panel A.

12
Supplemental Figure S6. 2Fo-Fc electron-density maps of zinc-binding sites (1σ; stereo views). Additional views of crystal structure of [HisA4, HisA8]-insulin. (A) axial zinc ion of T-state trimer; (B) axial zinc ion of Rf-state trimer; (C) novel A4-A8-coordinated peripheral zinc-binding site (repeated for convenience from Fig. 1E in main text). Whereas chloride ion occupancy is essentially 100% in panels B and C, only weak density is observed in the T-state trimer.
13
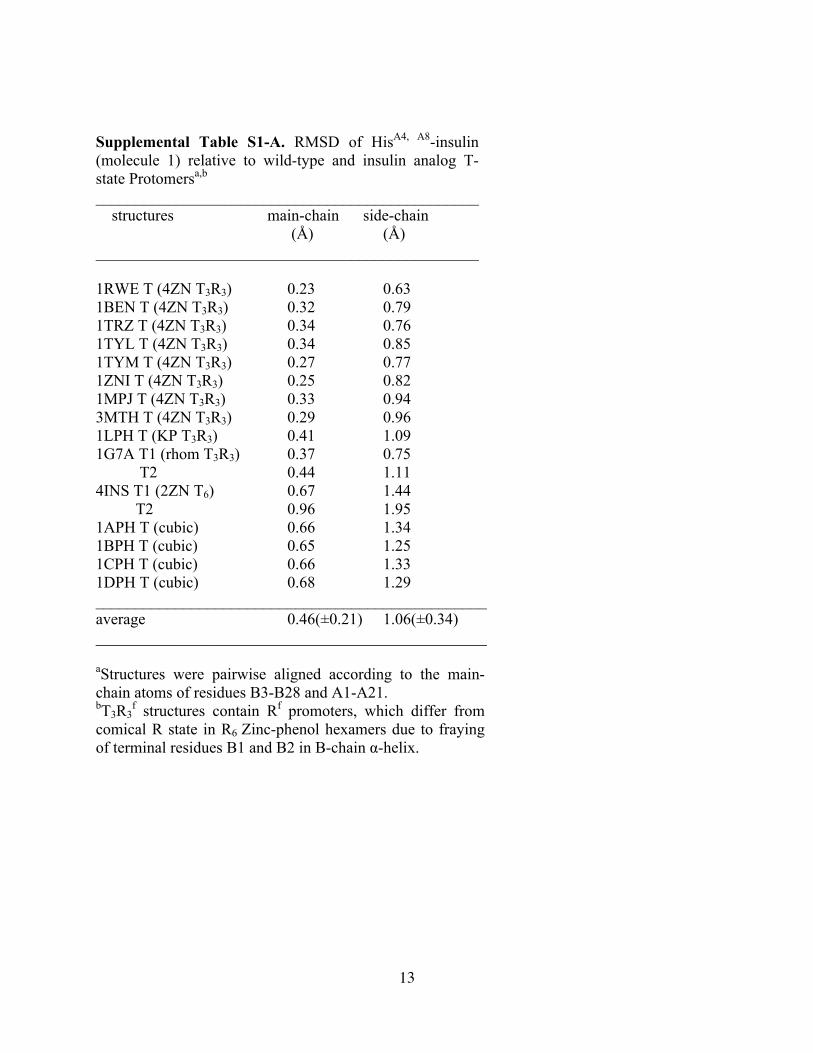
Supplemental Table S1-A. RMSD of HisA4, A8-insulin (molecule 1) relative to wild-type and insulin analog T-state Protomersa,b ________________________________________________ structures main-chain side-chain (Å) (Å) ________________________________________________ 1RWE T (4ZN T3R3) 0.23 0.63 1BEN T (4ZN T3R3) 0.32 0.79 1TRZ T (4ZN T3R3) 0.34 0.76 1TYL T (4ZN T3R3) 0.34 0.85 1TYM T (4ZN T3R3) 0.27 0.77 1ZNI T (4ZN T3R3) 0.25 0.82 1MPJ T (4ZN T3R3) 0.33 0.94 3MTH T (4ZN T3R3) 0.29 0.96 1LPH T (KP T3R3) 0.41 1.09 1G7A T1 (rhom T3R3) 0.37 0.75 T2 0.44 1.11 4INS T1 (2ZN T6) 0.67 1.44 T2 0.96 1.95 1APH T (cubic) 0.66 1.34 1BPH T (cubic) 0.65 1.25 1CPH T (cubic) 0.66 1.33 1DPH T (cubic) 0.68 1.29 _________________________________________________ average 0.46(±0.21) 1.06(±0.34) _________________________________________________ aStructures were pairwise aligned according to the main-chain atoms of residues B3-B28 and A1-A21. bT3R3
f structures contain Rf promoters, which differ from comical R state in R6 Zinc-phenol hexamers due to fraying of terminal residues B1 and B2 in B-chain α-helix.

14
Supplemental Table S1-B. RMSD of HisA4, A8-insulin (molecule 2) relative to wild-type and insulin analog R-state protomersa,b ___________________________________________________ structures main-chain side-chain (Å) (Å) ___________________________________________________ 1RWE R (4ZN T3R3) 0.35 0.76 1BEN R (4ZN T3R3) 0.69 1.04 1TRZ R (4ZN T3R3) 0.73 1.05 1TYL R (4ZN T3R3) 0.42 0.84 1TYM R (4ZN T3R3) 0.44 0.88 1ZNI R (4ZN T3R3) 0.66 1.17 1MPJ R (4ZN T3R3) 0.59 1.02 3MTH R (4ZN T3R3) 0.66 1.34 1G7A R1 (rhom T3R3) 0.70 1.25 R2 0.66 1.11 1EV3 R1 (rhom R2) 0.52 1.22 R2 0.41 0.81 1ZEG R1 (rhom R2) 0.53 1.01 R2 0.53 1.30 1ZEH R1 (rhom R2) 0.82 1.63 R2 0.92 1.86 1ZNJ R1 (monoclinic) 0.57 1.20 R2 0.51 1.35 R3 0.62 1.33 R4 0.54 1.04 R5 0.50 1.02 R6 0.55 0.99 1EV6 R1 (monoclinic) 0.59 1.21 R2 0.46 1.20 R3 0.84 1.53 R4 0.50 1.03 R5 0.42 1.15 R6 0.60 1.15 _____________________________________________________ average 0.58(±0.14) 1.16(±0.24) _____________________________________________________ aStructures were pair-wise aligned according to the main-chain atoms of residues B3-B28 and A1-A21. bT3R3 structures contain Rf promoters, which differ from comical R state in R6 Zinc-phenol hexamers due to fraying of terminal residues B1 and B2 in B-chain α -helix

15
Supplemental Table S1-C. Control RMSD of prototypical wild-type insulin T state (4INS, molecule 1) related to wild-type and insulin analog crystal structuresa ________________________________________________ Structures main-chain side-chain (Å) (Å) ________________________________________________ 1APH (cubic) 0.73 1.54
1BPH (cubic) 0.73 1.54
1CPH (cubic) 0.77 1.69
1DPH (cubic) 0.78 1.62
1BEN T (4ZN T3R3) 0.61 1.16
1MPJ T (4ZN T3R3) 0.63 1.26
1TRZ T (4ZN T3R3) 0.60 1.19
1TYL T (4ZN T3R3) 0.60 1.19
1TYM T (4ZN T3R3) 0.64 1.19
1ZNI T (4ZN T3R3) 0.61 1.26
1LPH T (KP- T3R3) 0.63 1.25
3MTH T (4ZN T3R3) 0.65 1.23
1G7A T1 (rhom- T3R3) 0.69 1.35
T2 (rhom- T3R3) 0.71 1.38
4ZN T2 (2ZN T6) 0.68 1.34
__________________________________________________ Average 0.67(±0.06) 1.35(±0.17) __________________________________________________ aStructures were pair-wise aligned according to the main-chain atoms of residues B3-B28 and A2-A20.
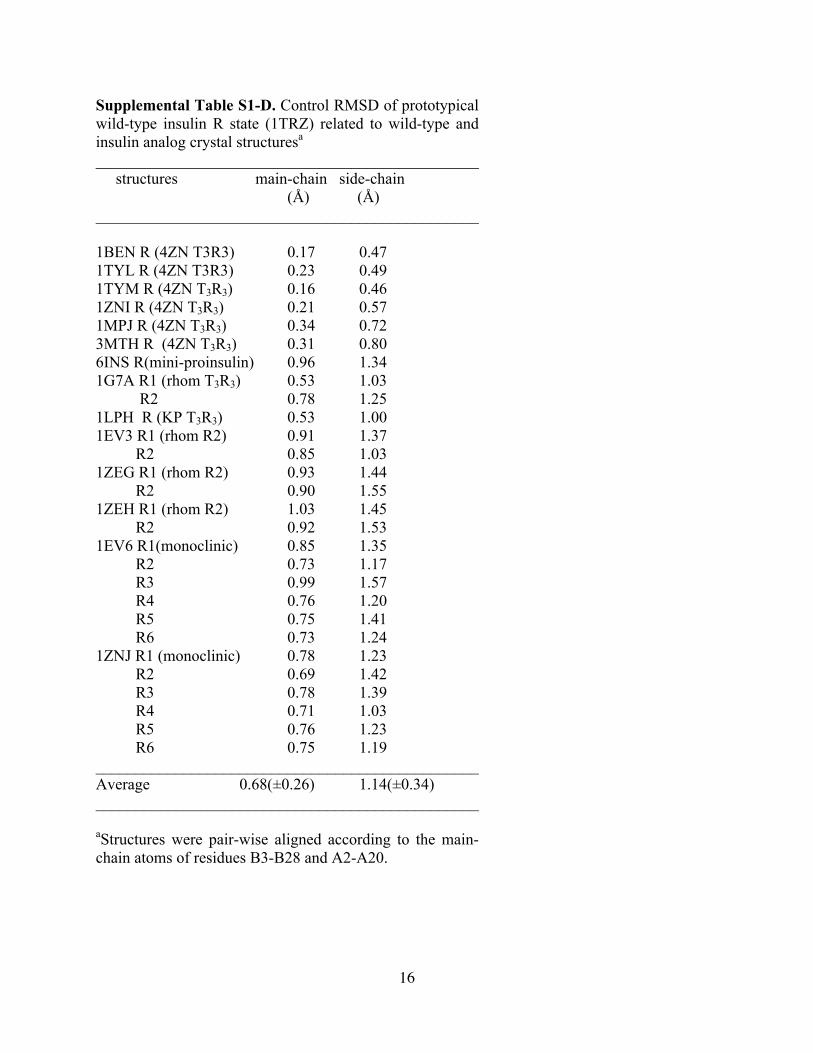
16
Supplemental Table S1-D. Control RMSD of prototypical wild-type insulin R state (1TRZ) related to wild-type and insulin analog crystal structuresa ________________________________________________ structures main-chain side-chain (Å) (Å) ________________________________________________ 1BEN R (4ZN T3R3) 0.17 0.47 1TYL R (4ZN T3R3) 0.23 0.49 1TYM R (4ZN T3R3) 0.16 0.46 1ZNI R (4ZN T3R3) 0.21 0.57 1MPJ R (4ZN T3R3) 0.34 0.72 3MTH R (4ZN T3R3) 0.31 0.80 6INS R(mini-proinsulin) 0.96 1.34 1G7A R1 (rhom T3R3) 0.53 1.03 R2 0.78 1.25 1LPH R (KP T3R3) 0.53 1.00 1EV3 R1 (rhom R2) 0.91 1.37 R2 0.85 1.03 1ZEG R1 (rhom R2) 0.93 1.44 R2 0.90 1.55 1ZEH R1 (rhom R2) 1.03 1.45 R2 0.92 1.53 1EV6 R1(monoclinic) 0.85 1.35 R2 0.73 1.17 R3 0.99 1.57 R4 0.76 1.20 R5 0.75 1.41 R6 0.73 1.24 1ZNJ R1 (monoclinic) 0.78 1.23 R2 0.69 1.42 R3 0.78 1.39 R4 0.71 1.03 R5 0.76 1.23 R6 0.75 1.19 ________________________________________________ Average 0.68(±0.26) 1.14(±0.34) ________________________________________________ aStructures were pair-wise aligned according to the main-chain atoms of residues B3-B28 and A2-A20.

17
Supplemental Table S2. Zinc Coordination Geometries A. Bond Distances novel site T-state trimer (axial) R-state trimer (axial) A4 ND1--Zn 2.09 B10NE2--Zn 2.06 B10 NE2--Zn 2.03 A8 ND1--Zn 2.09 ZN – Cl/H2O 2.29 ZN -- Cl 2.25 Zn -- Cl 2.26 B. Bond Angles novel site A4 ND1--Zn--Cl 93.3 deg A8 ND1--Zn--Cl 101.7 A4 NE2'--Zn--Cl 135.0 A4 ND1--Zn--A8 ND1 104.8 A8 ND1--Zn--A4 NE2' 111.0 A4 NE2'--Zn--A4 ND1 106.7 T-state trimer (axial zinc ion) B10 NE2--Zn-- Cl 120.6 deg B10 NE2--Zn-- B10 NE2' 96.4 R-state trimer (axial zinc ion) B10 NE2--Zn—Cl 109.4 deg B10 NE2--Zn-- B10 NE2' 109.5
18
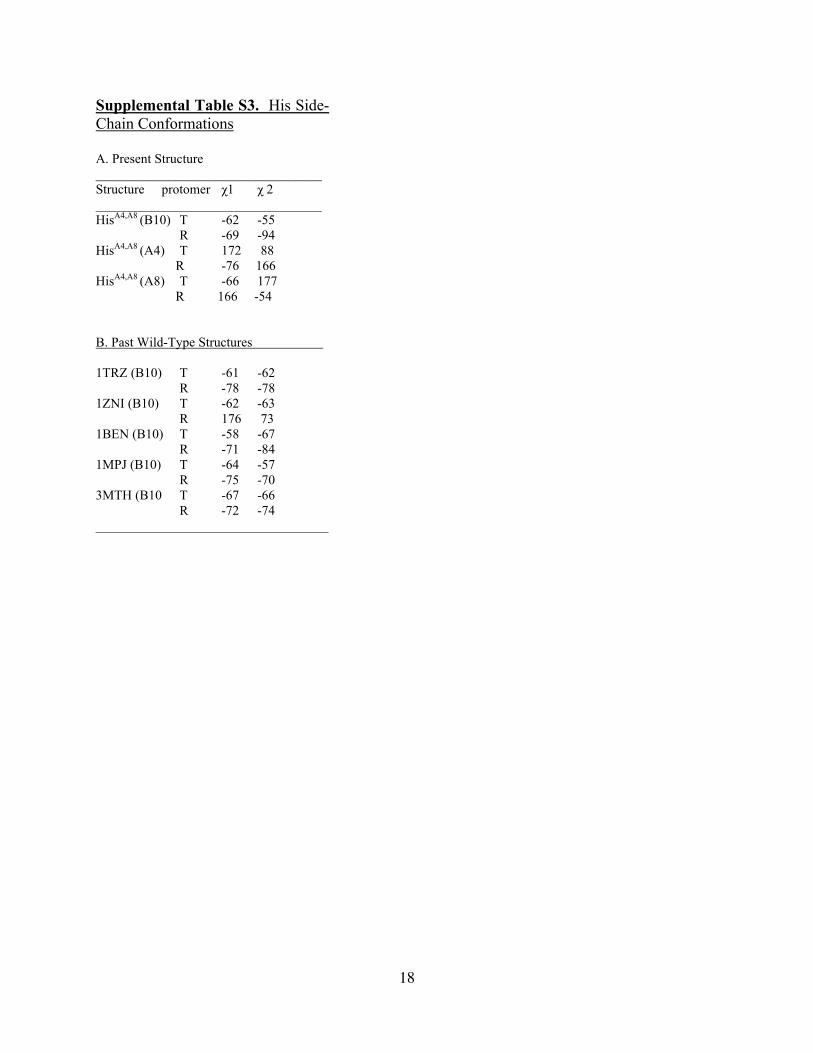
Supplemental Table S3. His Side-Chain Conformations A. Present Structure __________________________________ Structure protomer χ1 χ 2 __________________________________ HisA4,A8 (B10) T -62 -55 R -69 -94 HisA4,A8 (A4) T 172 88 R -76 166 HisA4,A8 (A8) T -66 177 R 166 -54 B. Past Wild-Type Structures 1TRZ (B10) T -61 -62 R -78 -78 1ZNI (B10) T -62 -63 R 176 73 1BEN (B10) T -58 -67 R -71 -84 1MPJ (B10) T -64 -57 R -75 -70 3MTH (B10 T -67 -66 R -72 -74 ___________________________________

19
Supplemental Table S4. Receptor-Binding Studiesa A. Dissociation Constants HIRB IGFR Kd SEM Kd SEM insulin 0.060 0.009 12.2 1.8 Lantus 0.110 0.016 3.1 0.44 [HisA4,A8]-HI 0.045 0.007 71.2 14.7 [HisA4,A8-GlyA21]-HI 0.091 0.012 133.3 33
B. Relative Receptor-Binding Properties protein relative affinitiesb receptor selectivityc IR IGF-1R ratio relative wild-type insulin 100 0.30 ± 0.02 333 ± 36 1
insulin glargine 43 ± 7 0.92 ± 0.12 47 ± 9 0.14 ± 0.02
HisA4, HisA8-insulin 125 ± 18 0.05 ± 0.01 25000 ± 830 7.5 ± 2.5 aEstimates of dissociation constants are based on an exact mathematical expression to describe the competitive binding of two different ligands to a receptor (13,18). Errors derive from standard errors of the mean (SEM). bRelative IR affinity of wild-type insulin is defined as 100%; relative affinity of IGF-I for IGF-1R (not shown in Fig. 2e) is likewise defined at 100%. The affinity of insulin for IR (isoform B) is similar to the affinity of IGF-I for IGF-1R. cReceptor selectivity is calculated either as an absolute value (the ratio of dissociation constants for IR versus IGF-1R) or as a relative value (the ratio of the absolute selectivities of an analog versus wild-type insulin). The relative selectivity of wild-type insulin is 1 by definition.

Hua, Jonathan Whittaker, Faramarz Ismail-Beigi and Michael A. WeissNelson B. Phillips, Zhu-li Wan, Linda Whittaker, Shi-Quan Hu, Kun Huang, Qing-xin
HEXAMERS AS A LONG ACTING DEPOTSupramolecular Protein Engineering: DESIGN OF ZINC-STAPLED INSULIN
doi: 10.1074/jbc.C110.105825 originally published online February 24, 20102010, 285:11755-11759.J. Biol. Chem.
10.1074/jbc.C110.105825Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/02/24/C110.105825.DC1.html
http://www.jbc.org/content/suppl/2010/04/05/285.16.11755.DC1.htmlRead an Author Profile for this article at
http://www.jbc.org/content/285/16/11755.full.html#ref-list-1
This article cites 32 references, 5 of which can be accessed free at
by guest on September 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from




















