
Study of Oral Tolerance and Its Indirect Effects in Adoptive Cell Transfer Experiments
7
321 Ann. N.Y. Acad. Sci. 1029: 321–327 (2004). © 2004 New York Academy of Sciences. doi: 10.1196/annals.1309.013 Study of Oral Tolerance and Its Indirect Effects in Adoptive Cell Transfer Experiments ANDRÉ PIRES DA CUNHA, a NELSON MONTEIRO VAZ, a AND CLÁUDIA ROCHA CARVALHO b a Departmento de Bioquímica-Imunologia ICB-UFMG, Brazil b Departmento de Morfologia, ICB-UFMG, Brazil ABSTRACT: Parenteral exposure to antigens to which oral tolerance had been previously induced results in the inhibition of immune responses to other un- related antigens. Herein we tested whether indirect effects of oral tolerance could be adoptively transferred. Anti-Ova– and antihemoglobin-specific responsiveness as well as oral tolernace to Ova were transferred to irradiated, but not to normal, nonirradiated recepients. Irradiation, thus, facilitated adop- tive transfer of oral tolerance. However, the inhibitory (indirect) effects upon the unrelated immunogen were not adoptively transferred, even to irradiated recepients. In addition, we studied adoptively transferred CFSE-labeled spleen cells by flow cytometry in recipient spleen, inguinal lymph nodes, and bone marrow, both in irradiated and nonirradiated recipients, 1, 3, or 5 days after cell transfer. Comparing the percent and absolute number of CFSE-labeled cells in each organ displayed significant differences in the dynamics of decay of adoptively transferred cells from tolerant or immune donors. KEYWORDS: oral tolerance; syngeneic barrier; indirect effects; adoptive transference INTRODUCTION Oral tolerance was first described by Wells in 1911. 1 In recent years, there has been renewed interest in this phenomenon as a possible therapy for autoimmunity. 2 Although many aspects of oral tolerance have been resolved, its fundamental nature remains elusive. Several results from our laboratories 3,4 and the laboratories of others 5 have shown that in addition to blocking immune responsiveness to the specific antigen, parenteral exposure to an antigen against which oral tolerance has been previously induced can induce blockage of the initiation of immune responses to unrelated antigens. Many laboratories have used the adoptive transfer of cells between syngeneic animals as a tool for seeking a better understanding of the mechanisms involved in Address for correspondence: Nelson Monteiro Vaz, Laboratório de Imunobiologia, Departa- mento de Bioquímica e Imunologia, ICB-UFMG, Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte, MG, Brazil, CEP: 31270-901 Caixa Postal 486. Voice: +5531 3499-2662; fax: +5531 3499-2640. [email protected]; [email protected]
Transcript of Study of Oral Tolerance and Its Indirect Effects in Adoptive Cell Transfer Experiments

321
Ann. N.Y. Acad. Sci. 1029: 321–327 (2004). © 2004 New York Academy of Sciences.doi: 10.1196/annals.1309.013
Study of Oral Tolerance and Its Indirect Effects in Adoptive Cell Transfer Experiments
ANDRÉ PIRES DA CUNHA,a NELSON MONTEIRO VAZ,a AND CLÁUDIA ROCHA CARVALHOb
aDepartmento de Bioquímica-Imunologia ICB-UFMG, BrazilbDepartmento de Morfologia, ICB-UFMG, Brazil
ABSTRACT: Parenteral exposure to antigens to which oral tolerance had beenpreviously induced results in the inhibition of immune responses to other un-related antigens. Herein we tested whether indirect effects of oral tolerancecould be adoptively transferred. Anti-Ova– and antihemoglobin-specificresponsiveness as well as oral tolernace to Ova were transferred to irradiated,but not to normal, nonirradiated recepients. Irradiation, thus, facilitated adop-tive transfer of oral tolerance. However, the inhibitory (indirect) effects uponthe unrelated immunogen were not adoptively transferred, even to irradiatedrecepients. In addition, we studied adoptively transferred CFSE-labeled spleencells by flow cytometry in recipient spleen, inguinal lymph nodes, and bonemarrow, both in irradiated and nonirradiated recipients, 1, 3, or 5 days aftercell transfer. Comparing the percent and absolute number of CFSE-labeledcells in each organ displayed significant differences in the dynamics of decay ofadoptively transferred cells from tolerant or immune donors.
KEYWORDS: oral tolerance; syngeneic barrier; indirect effects; adoptivetransference
INTRODUCTION
Oral tolerance was first described by Wells in 1911.1 In recent years, there hasbeen renewed interest in this phenomenon as a possible therapy for autoimmunity.2
Although many aspects of oral tolerance have been resolved, its fundamental natureremains elusive.
Several results from our laboratories3,4 and the laboratories of others5 haveshown that in addition to blocking immune responsiveness to the specific antigen,parenteral exposure to an antigen against which oral tolerance has been previouslyinduced can induce blockage of the initiation of immune responses to unrelatedantigens.
Many laboratories have used the adoptive transfer of cells between syngeneicanimals as a tool for seeking a better understanding of the mechanisms involved in
Address for correspondence: Nelson Monteiro Vaz, Laboratório de Imunobiologia, Departa-mento de Bioquímica e Imunologia, ICB-UFMG, Av. Antônio Carlos, 6627, Pampulha, BeloHorizonte, MG, Brazil, CEP: 31270-901 Caixa Postal 486. Voice: +5531 3499-2662; fax: +55313499-2640.

322 ANNALS NEW YORK ACADEMY OF SCIENCES
immunological phenomena. A dominant consideration in these studies is based onthe fact that the main determinant in cell transplantation is MHC dependent, suchthat histocompatibility determines whether transferred cells will be incorporated inthe immunological activities of the recipient or not. However, a more fundamentalrole of the recipient’s cells in the survival of transferred cells and the establishmentof these immunological activities, which prevails even in syngeneic transplants, hasalso been demonstrated: the so-called “syngeneic barrier.”6 Some form of depletionof the recipient’s lymphocytes is believed to be necessary to provide “space” fortransferred cells to operate. Experimentally, this is usually achieved by whole-bodyirradiation of recipient animals,6 or other immunosuppressive treatments. Notwith-standing, adoptive transference of oral tolerance has been demonstrated usingsyngeneic animals with an intact immune system.7 The subject, however, is still amatter of debate, with most authors proposing that depletion of the recipient’s cellsis necessary for the persistence of isogeneic transferred cells and, beyond this, topromote homeostatic expansion of transferred cells.8,9
With these questions in mind, the present experiments were undertaken to assessthe possibility of adoptively transferring oral tolerance and its indirect effects tonormal or previously irradiated syngeneic animals, and to evaluate which factorsinterfere in these events and by what means.
MATERIALS AND METHODS
Young adult C57BL/6 mice (7 to 10 weeks of age) were made orally tolerant toovalbumin (Ova) by drinking a solution of 4 mg/mL Ova (Ova, Sigma Grade III;Sigma Chemical Co., St Louis, MO, USA) for 3 days. Seven days later, they receivedintraperitoneally 2 µg of hemoglobin from Biomphalaria glabrata (Hb, kindly pro-vided by Prof. Marcelo Santoro from our department) mixed with 10 µg of Ova plus3 mg aluminum hydroxide (Al(OH)3). Fourteen days thereafter, 5 × 107 spleen cellsfrom these mice or from nontolerant controls were transferred intravenously intonormal or previously irradiated mice (600 rad). All recipients were boosted intrap-eritoneally with a mixture of 2 µg Hb and 10 µg Ova without adjuvant immediatelyafter cell transfer and 7 d later. Serum was collected 7 d thereafter and analyzed byELISA; results are represented as Elisa∗ as previously described.4 Results are ex-pressed as means plus one standard deviation (1 SD). To test significance, Student’st test was performed. CFSE-labeled cells were adoptively transferred as previouslydescribed.10 Pooled donor spleen cells were traced by flow cytometry in irradiatedand nonirradiated recipient spleen, inguinal lymph nodes, and bone marrow 1, 3, or5 d after cell transfer, using the same protocol described above.
RESULTS
We investigated the possibility of adoptively transferring the “indirect (inhibito-ry) effects” of parenteral exposures to a tolerated antigen. In mice that were orallyexposed to Ova and then immunized with Ova+ Hb mixtures, anti-Ova and anti-Hbtiters were both significantly lower than in mice not orally exposed to Ova, as ex-pected (FIG. 1). These mice were used as donors of spleen cells to normal or previ-

323DA CUNHA et al.: ORAL TOLERANCE AND ITS INDIRECT EFFECTS
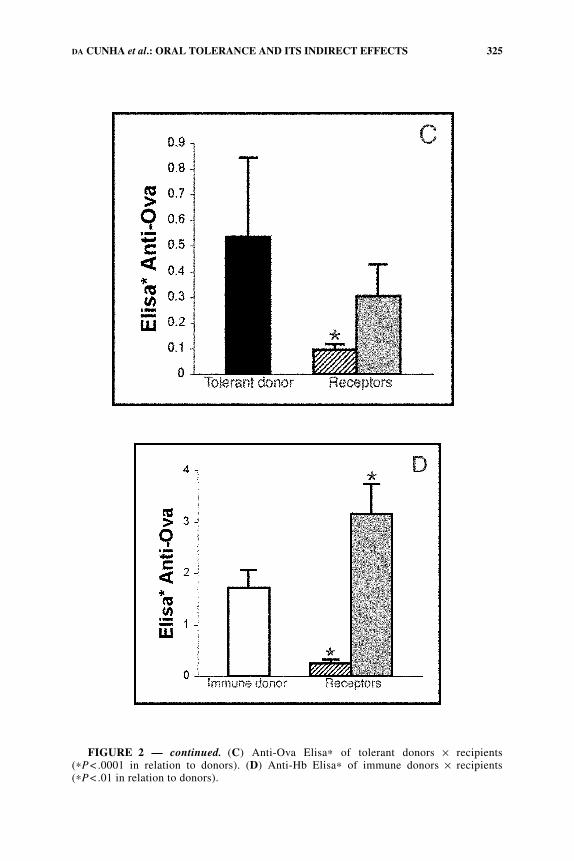
ously irradiated syngeneic recipients. Subsequent bleedings of mice that receivedcells from nontolerant donors showed that antibody responses to Ova and Hb werepresent in irradiated, but not in nonirradiated syngeneic recipients (FIG. 2, B and D).Irradiated recipient mice that received cells from tolerant donors showed low re-sponses to Ova but stronger responses to Hb. This showed that oral tolerance wasadoptively transferred, but its indirect (inhibitory) effects were not (FIG. 2, A and C).In nonirradiated recipients, there was no evidence of adoptive transfer (FIG. 2) ofimmunization, tolerance, or its indirect effects, a phenomenon that may be ascribedto the so-called “syngeneic barrier.”6
Further experiments were performed by adoptively transferring CFSE-labeledcells. Preliminary results indicate that more CFSE-labeled donor cells may be recov-ered from nonirradiated recipients (data not shown). However, owing to the lympho-cyte depletion caused by irradiation, a larger percentage of CFSE cells may berecovered from irradiated recipients, independent of the origin of the transferredcells (tolerant or normal donors).
DISCUSSION
We were unable to transfer the indirect (inhibitory) effects of parenteral expo-sures to a tolerated antigen to either normal or irradiated recipients, whereas immuneresponsiveness (immunization) and tolerance could be successfully transferred toirradiated recipients. This suggests that the indirect (inhibitory) effects of the toler-ated antigen are more demanding of a systemic architecture than oral tolerance.
FIGURE 1. Anti-Ova and anti-Hb antibodies in C57BL/6J donor mice 14 d afterprimary immunization with Ova +Hb+Al(OH)3. Figure indicates immune (open bars) andtolerant (black bars) donors. ∗P< .0001 and ∗∗P< .05.

324 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 2. Anti-Ova and anti-Hb antibodies in C57BL/6J donor and recipient mice.Donors were immunized with Ova+ Hb+ Al(OH)3; 14 days later, sera were collected forantibody testing, and 5 ×107 spleen cells from these mice were transferred intravenously tosyngeneic irradiated (dotted bars) or normal (hatched bars) mice. These recipients werethen immunized intraperitoneally with Ova+Hb immediately after transference and 7 dthereafter. The figure indicates immune (open bars) and tolerant (black bars) donors. (A)Anti-Hb Elisa∗ of tolerant donors × recipients (∗P<.0001 in relation to donors). (B) Anti-Hb Elisa∗ of immune donors × recipients (∗P< .05 and ∗∗P< .001 in relation to donors).
325DA CUNHA et al.: ORAL TOLERANCE AND ITS INDIRECT EFFECTS
FIGURE 2 — continued. (C) Anti-Ova Elisa∗ of tolerant donors × recipients(∗P< .0001 in relation to donors). (D) Anti-Hb Elisa∗ of immune donors × recipients(∗P< .01 in relation to donors).

326 ANNALS NEW YORK ACADEMY OF SCIENCES
Cells transferred to nonirradiated syngeneic recipients have their specific activitiesstrikingly blocked, such that neither immunization nor tolerance could be transferredto nonirradiated recipients. However, a few previous studies have demonstrated thepossibility of transferring oral tolerance to intact recipients.7,11 Nonetheless, thetransference is made at different time points after oral tolerance induction and usu-ally donor mice are fed but not challenged before donation. We are unaware whetherthis could be an explanation for this discrepancy.
Overall, these results imply that there are barriers other than MHC-mismatch tothe functional implantation of the transferred lymphocytes. As classically demon-strated by Celada,6 antibody-producing cells transferred to normal syngeneic recip-ients of different ages face a “syngeneic barrier” that significantly hinders theiroperation, even in newborn recipients, and grows steadily thereafter. In irradiatedrecipients, who had most of their cycling cells destroyed, transferred cells may befully operant and may respond 50- to 100-fold better to subsequent exposures to thespecific antigen than when transferred to normal recipients.
A further point is worth attention: We could only tell that oral tolerance was trans-ferred to irradiated recipients because we compared this group with another that hadalso been irradiated and received cells from immune donors. However, we could notevaluate whether oral tolerance was transferred to nonirradiated recipients becauseimmunization was not transferred to the immune control group.
We used CFSE cell-labeling to study the distribution of cells derived from toler-ant or immune donors in irradiated or normal syngeneic recipients. Our preliminaryresults suggest that on the first day the pattern of distribution of cells derived fromimmune or tolerant donors did not differ in any of the analyzed organs (data notshown). By the third day, however, recipients of cells from immune but not tolerantdonors showed a decrease in both the relative and absolute number of cells (data notshown). This suggests that there may be differences in the dynamics of decay ofadoptively transferred cells from tolerant or immune donors.
An issue arising from these results is why oral tolerance is transferable only toirradiated recipients if, as mentioned above, nonirradiated recipients maintain ahigher absolute number of donor cells. It may be that although animals with an intactimmune system display a higher number of donor-derived cells, they represent asmaller proportion of total cells, and may be unable to establish the interactions nec-essary to maintain tolerance. An environment where transferred cells predominatemay be necessary to form the connections involved in the production of activating orinhibitory cytokines.
ACKNOWLEDGMENTS
We thank Ms. Ilda Marçal and Frankcinéia Assis for their excellent technicalassistance. CNPq and FAPEMIG, Brazil supported this work.
REFERENCES
1. WELLS, H.G. 1911. Studies on the chemistry of anaphylaxis. III. Experiments with iso-lated proteins, specially those of the hen’s egg. J. Infect. Dis. 8: 147–171.

327DA CUNHA et al.: ORAL TOLERANCE AND ITS INDIRECT EFFECTS
2. FARIA, A.M. & H.L. WEINER. 1999. Oral tolerance: mechanisms and therapeutic appli-cations. Adv. Immunol. 73: 153–264.
3. VAZ, N.M., D.G. HANSON, L.C.S. MAIA, et al. 1981. Cross-suppression of specificimmune responses after oral tolerance. Mem. Inst. Oswaldo Cruz. 76: 83–91.
4. CARVALHO, C.R., B.A. VERDOLIN & N.M. VAZ. 1997. Indirect effects of oral tolerancecannot be ascribed to bystander suppression. Scand. J. Immunol. 45: 276–281.
5. MILLER, A., O. LIDER & H.L. WEINER. 1991. Antigen driven bystander suppressionafter oral administration of antigen. J. Exp. Med. 174: 791–798.
6. CELADA, F. 1966. Quantitative studies of the adoptive immunological memory in mice.I. An age-dependent barrier to syngeneic transplantation. J. Exp. Med. 124: 1–14.
7. RICHMAN, L.K., J.M. CHILLER, W.R. BROWN, et al. 1978. Enterically induced immuno-logic tolerance. I. Induction of suppressor T lymphocytes by intragastric administra-tion of soluble proteins. J. Immunol. 121: 2429–2434.
8. MAINE, G.N. & J.J. MULÉ. 2002. Making room for T cells. J. Clin. Invest. 110: 157–159.9. TANCHOT, C., A. LE CAMPION, B. MARTIN, et al. 2002. Conversion of naive T cells to a
memory-like phenotype in lymphopenic hosts is not related to a homeostatic mecha-nism that fills the peripheral naive T cell pool. J. Immunol. 168: 5042–5046.
10. OEHEN, S., K. BRDUSCHA-RIEM, A. OXENIUS, et al. 1997. A simple method for evaluat-ing the rejection of grafted spleen cells by flow cytometry and tracing adoptivelytransferred cells by light microscopy. J. Immunol. Methods. 207: 33–42.
11. CHEN, Y., J. INOBE, H.L. WEINER. 1995. Induction of oral tolerance to myelin basic pro-tein in CD-8 depleted mice: both CD4+ and CD8+ cells mediate active suppression.J. Immunol. 155: 910– 916.




















