
Studie. and ..... rcb in partial fulfl1l1ent of tb. •
192
os os .. ," " . . " '''' .. - ) 1 ( \ FLORAL DEVELOPMENT :m THE by Ulher Poaluazny .1 Studie. and ..... rcb in partial fulfl1l1ent of tb. l'aquir..-t. fOI: the ... rae of Doctor of PhilOl'ophy. \. . " , .,' '\ • • . ..
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Studie. and ..... rcb in partial fulfl1l1ent of tb. •

os os
.. ,"
" . . "
'''' ..
-
---~ )
1
( \
FLORAL DEVELOPMENT :m THE N~ADALES
by
Ulher Poaluazny
.1
Studie. and ..... rcb in partial fulfl1l1ent of tb. ~
l'aquir..-t. fOI: the ... rae of Doctor of PhilOl'ophy. \.
. "
,
.,' '\
•
•
. ..

"
:
1
\{
1 nom DEVE'LOPMENT t~ THE NAJADALES
•
\
,
•
,."
, , .,
. ..

j
--
'.
, \
The lyabol of the University ta the Iron atatue
outlide the Rathakeller of a barefoot loolegirl that
every etudent kil,ea ~t graduation. The university ia a
Mece. ta which atudenta come with aaa.thing le8. th~n
perfe~aith. lt i. important that .tudent~ bring a
certain ragaauffin, barefoot Irreverence to th.ir etudiee; .
they are not here to vorehlp vb*t i. knovn but to que. tian
it.
.... ,. , , Jacob Iroa.owski
(1973)
, "
\
\ \ \
\ ~ .. "J<.
\ , , , 1
, .. ~ ~. . ,f.
t
..

, '
'{f'l/f'
\" ~{.t c'.
1 L
t , ,
'i \
Ph.D. Biology
U.her Polluuny p
{ Floral development in the Najadalea
ABSTRACT
l'The floral developaent (organogeneaia and hiatogefi.sis) of
PO;&mOleton rlchard.onii, Poc-.o,acon den.us, Ruppia mariti .. var.
mariti .. , Zannichellia palu. tri. and Naja. flexili. ia de.cribed and
~oapar.d. The new and Tel.vant data thu. obtained la applied to (
eontroverlial morphologieal and taxonomie problems. Homologies of
floral appendagel are eatabli.hed, elpeeially the homology of the
t~pal in Pot";reton and the outsrQwth of the atame6 connective in
Ruppl.. a. vell as that of the membranou& envelope about the pistl11ate
flover in Zannichellia and the two about the staminate flower in Najas.
The flover-inflorescence controversy la dilcu.sed and ne. evidence ls
introduead .upportina the concept of partial hoaology with ragard to
bath flover and inflora.canee. A morphological leriel betyeen Potaaogeton
and B!1!! il propo'ad, ba •• d on .iailaritie. in develop.ental pattern. of
fertile brancha. and floral appandas •• , a.pecially of th. androecium and
aynoaciua. The .arphololical aeri •• , in turn, .ugg.ltl relationahip.
withiD the Naja4ala.. Thara i. luppert for ke.pinl PotaDO,aton and
_ppl. within one feaily t whil. Zatm1ch.1l1. and Najal are lufficlently
di.tinet .0 " to Vlrrant th.ir pOlitionlng within I.par.te feailie ••
",':-'
~J

"
o
7
Ph.D. Biologie
Usher Posluszny
Le dévelopPêment floral chez les Najadales
RESUME
• Le développement floral (organogénèse et histogénèse) de Potamogeton
richards9nil, Potamogeton densus, BU2pia maritima var. maritima,
Z~nnichelli8 palustris et NaJas flexilis est décrit et comparé. Les
résultats originaux et pertinents ainsi obtenus sont appllqu~s à des ---~"../'./""
---problèmes morphologiques et taxonomiques controversés. Cert~és _/
homologies. des pièces florales sont établies, partlcu~e~t l'homologie
/ du tépale chez Potamogeton et de l'excroissa~u connectif de l'étamine
chez Ruppià, de même que l'homologie de l~~~ve~euse autour
des deux pistils chez Zannichellia e,t -des deux enveloppes memb
Butour de l'étamine chez Na las //La controverse fleur-inflorescence t
discutée et les renSeigne~ts nOuveaux présentés appuient le concept de
l'homologie partielle~~t à 18 fleur et l'inflorescence. Une série
morphologique entr~~mogeton et Naias est proposée, basée sur les
similitudèS de~des de développements des branches fertiles et des
pièces flO~S. particulièrement de l'androcée et du gynécée. La série
morpholo.glque ainsi êtabl1e sugg~re des liens li l' int6rieur des Najada les. /
Il est proposé de laisser Potamogeton et Ruppia dans la même famille,
tandis que Zan~ichellia et Najas sont suffisamment distinctes pour
justifier le;f reclasaifiC8tion l l'intérieur de familles séparées.
Traduit par
Marc-André Paré

, , 1
•
PREFACE
This thesis was produc~d in the'form of original pepers suitable
for submission ta journals in accordance with section 4.2.7 paragraRh ----- -
(h) of the Faculty of Gradua~t~e~S~uU~~nnr..1«e~sÎEe~alrr~C~h~A~n~n~o~u~n~c~e~m:e:n~t~of " rul.,..."l--1:Irl1u for sub~itting a ihesis. Each chapter ls in-
cor~orated directly forro suitable for publication. Refêrences are
assembled in the sections in which they are cited. Chapters
1-3 have been
(Chapter 1,
4 and.
as papers by Pos1uszny, U. and R. Sattler
J. Bot. 61(2): 209-216. 1974; Chapter 2, Cano J. Bot.
1973; Chapter 3, Cano J. Bot. ~: 1607-1612. 1974). Chapters
be submltted for publicAtion to the Canadian Journal of Botany.
As well, an "INTRODUCTION AND REVIEW OF LITERATURE" and a "SUMMARY AND
LUS IONS" vere added ln order to connect and bring together the topies \
discussed in the five cbapters. Supp1ementary figures were also added to .~ 1
Chapters 1 (Figs. 31, 32) and 2 (Figs. 3l, 33), which did not appear in
the publications.
Voucher specimens for all the mateTial worked on, except Potamogeton
densus, were deposited at the Macdonald Herbarium, Ste. Anne de Bellevue,
Quebec. j ,
1
'" f
l

-,. ~,' r
>' i ',-',' .l
o
,~,~-----
il
CONTRIBUTIONS TO ORIGIN~L KNOWLEDGE
ln the author's opinion. the following items contained in thls
thesie contrlbuted to otiginal knowledge:
a} The tecently refined technique for the study qf floral
organogene,ie vas combinéd vith histogenetic analysis to provide the
tirst ~omprehen81ve developmental study of the flowers of Potamogeton
richard8onii. ~. densus, Ruppla maritima var. maritime. Zannichellia
paly.tria and Najas fIexi!is. Besides 8 genera! description, the fo!low-
ing
\
r--'--.... _____ ,; 1 points wer.rb~Ught to light for the firat time:
1. Th initiation of the lower median stamen and tepal of
î. den.ua beforè the floral apex is èlearly demarcated.
2. The initiation of each of the lateral atamens of P. densus
as two prtmordia.
3. The orlgin of the membranous sbeath about ~he upper
floral bud of !. maritime as a prophyll of the renewal growth
apex.
4. The initiàtion of the outgrowth o.f the stamen connective
in!. maritima after the staminal thecae begin to differentiate.
5. The origin and develop~nt of outgrovths on the upper
abaxial carpel wall in Potamogeton and luppia.
6. A ~learer ca.prehenalon of the developaent of fertile
branche. in~. palUft!i.. Both·.taa1nate and pietillate
branche. tend laward. a .yapod1al pattern.
7. The initiation and d.v.lo~t of th •• a.branou. envelope
about th. platl1late flawer ln Z. p!lUltri., at about , - 1
th.
at ... C).f O'ftl. 1ncaptiOG. Parttally ho.oloaoua Vith a
"( ,.,
.\

(
1
10 !
1 1 , 1
,'" , ... ~,j ! It·~ __ _
in
8. The initiation of the membranous sheath 'pout the node
in Z. p.lu.tri. a. a prophyll of the main axis renewal
growth apex.
9. The detal1s of the development and variation of lobes, at
or just after the inception of the girdling prlmordium about
the floral apex, in!. flexilis.
10. Th@ acropetal development of t~e 1nteguments ln the
.pistillate flower of N. flexilis.
b) This study also represents the first comparative approaeh to the
floral development of a representative sample within the Najadales. It
has provided new ins1ghts ioto old morphologieai and taxonomie problems.
These include the fo~lowing findings:
1. No total homology can be drawn between the tepal of Potamogeton
and the 'outgrowth of the stamen connective of Ruppia. The former
develops acropetally l1ke typ1cal perianth members, while the
latter initiate. after the theeae a.e already forming.
2. Camp.red ta oth.r species of Potamogeton, the development of ~
the flower of !. densus 18 suffiei.ntly different to support the
elas8ifieation of this speeies as a separa te genus (Groenlandla).
3. New floral developaental evidenee for Potaœogeton and Ruppia
indicate. that intem.di,tes between a "flower" and an "lnflores-
cene." .. y occur.
4. Floral and fertile branch development supports a morphologieal
.eri •• lro. Pota.oseton to Naja.(or conver8~ly from t.jas ta
Potaaoutcm).
5. Th. pattern and p.culla~ities of the morphologieal .erlel, ln
particular the di8contlnuity o.tve.n Juppla aad Zanni~helli •• lupport •
the propolal of lad.pendent orilln of fr.'h &ad .~ltwat.r Naj.dal.8.
"

•
Iv
1 ACKNOWLEDGEMENTS
.. First and foremost, l wish ta thank œy supervisor Dr. R. Sattler
for guidance and support above and beyond the calI of duty. His
encouragement and pàtience œade this thesis possible.
1 am very grateful ta the members of my superviaory committee
Dr. W. BoIl, Dr. M. Goldstein and Dr. K. Maier for their thoughtful
consultation and advice, and to my colleagues Dr. Alastair Macdonald,
\ Dr. A.F. Muhammad, Vera Block. Dr. V. Sing~, Tim Dickinson, Dr. ,Uta
Maier. and Eœil Daniel for unselflshly aharing with me, not only an
office and laboratory, but also their support and friendship. As weIl,
many thanks to aIl my fellow graduate students in the deparement who
helped me, in particular Paul Arnison and Seth Mantè who 80 closely
paralleled Many of my own experiences.
l would like to thank Rob_rt Lamarche and Paul Gauthier of the
Photographie Center for their advice and assiatance with the preparat-
ions of the plates, Mona Bissada for her help with some of the micro-
teehniques~ and the .. ny ae.bers of the Biology Deparement who kindly
helped me, vith .p~ci.l th.nks to Miss ~uby Mayhew and Mrs. Shirley
Cahn.
To tho •• who typed vith patience and diligence, Suzann~Anderson
and Bvelyn Fung-A-Ling 1 .. .ast Irateful. To Margaret Lieu wh~ proof-
read the final .. nuacript and to Marc-Andr' Par' who prepared the fr.nch
ab. tract •• arc! beaucoup.
", 0 ..
.. ' . • j

0-'"
-
1 am indebted to the National Reaearch Coune!l of Canada for
the direct support they gave me through a Poatgraduate Scholarship ' ...
for three years (1970-1973) and for the indirect support tQrough •
operating and equipment grants to Dr. R. Sattler. ,
l must alao thank some of my friends, Includ1ng those i~ the
department ••••.• Lynn Boshkov, Matilda Cheung, Claire Cooney, Ian
'airlie, Marg Harris, Stella Humphries, Nathan Maltz, Beesh Per1in,
v
MichAle Sammar1tano, Ravlnder Slngh, Steven Spiegel and Linda Thatcher,
• and thos. out. ide ••••• Francis Davis, Mavis Engel, Ellen Schnieder
and Simon Wasserberler, for their concern and support.
To my mother and stster, thank you for your constant faith and
encouragement duriug the put yeera. lt 1s to you both that 1 dedicate\ ,
thil theds.
) \ \
\
\ \,
\ ,
()
-
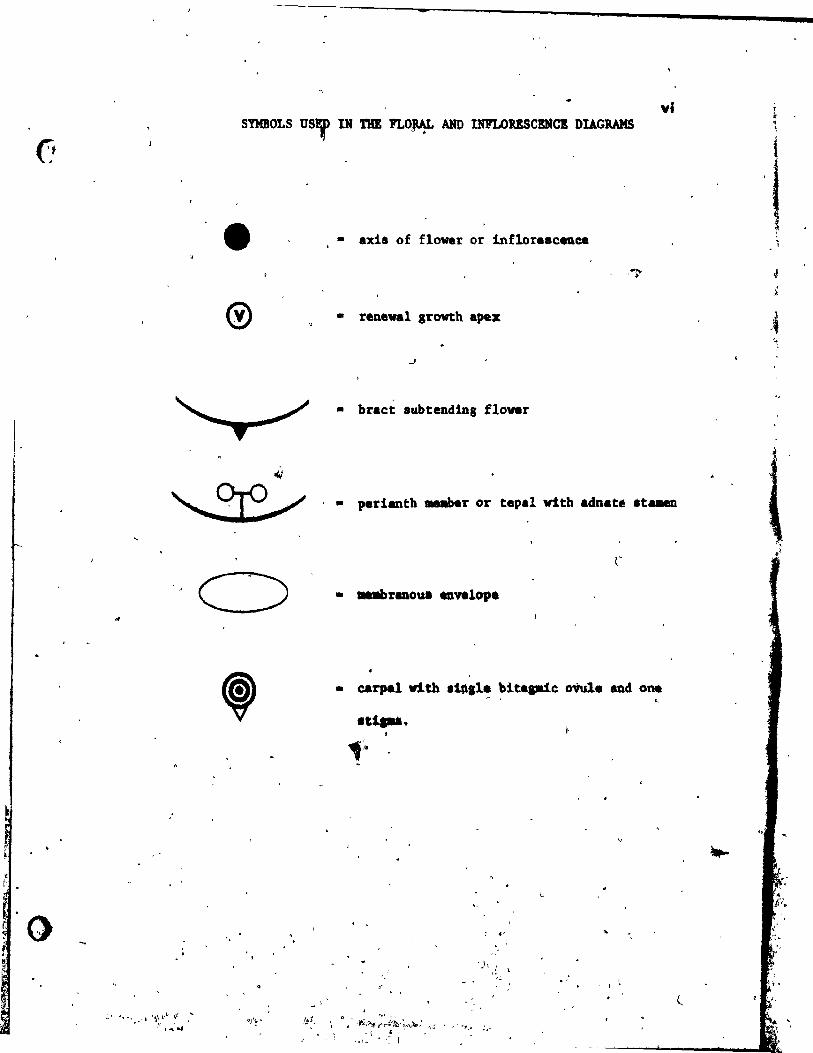
vi
• • axis of flower or inflore.c~ee
, ,~
• reneval growth apex
• br.ct 8ubtend1Dg flovar
<.t
~ . • pedanth 1IIIIaber or tepa! vith adnate te ...
:: 1 ~'~' ... _ t '*i~(j}"" :iJ ...
, r .... ~
, .
• carpe! vith 1i~ll~ ~it.pie oVule and. one ~
.u .....
.' . '
, .
J • ~ ,
'l l
1
f ~
-4 '1

~
t f
1 o
..
TABLE OF CONTENTS
• . ,
Page"
PRIFACE . ........ " ........ " ................................. . i
CONTRiBUTIONS 10 OIlGlNAL KNOWLBDG! .•.•••••••••• , •••••••••••• ~il
AC1CNOWl.ED~S • • • • • • • • • • • • • • • • • • • • • • • • • • • •••••••••••••••• if". 1 v
SYMBOLS USED IN FLORAL AND INFLORESCENCE DIAGRAMS ............. vi
INTRODUCTION AND REVIEW OF LITERATURE . , .................... .. 1
OBSEiVATIONS AND DISCUSSIONS .•...•.•.•.•.••. , .••••....•• " • .• 10
'(' ClIAPTER 1. Floral deve10plllent of l:"'bt!!oseton
\.. dcbardsonH .......................... 11,
J Ab.tract .••.......•. fi. • • • • • • • • • • • • • •• 12
CIIAPTD. 2.
.. p
•
~ Introduction ........................ 13
Katerial and Methode ••.• ; •••••••• \ ••• 13
Observations ........................ • Diacussi(m .......................... 18
Lit.rature Citad ..................... 27
Ploral dev.lopa.nt of Pota.Ds.ton densus. 29 <
Ab. tract , 1
................... ,l', •••••••• 30
Introduction ~ ...................... . '31
.... , ........... . 31
Ob •• rvationa •.••••••••••••.•..•.•.•.. 31
D1acu •• ion ..••••••••••••••••••....•.• 39 1
LiteratUft CiteeS •••••••• : •••••••• ; •••. S5 , "'JI
.. ' ..
..
vil
'f
A

TABLE OF CONTENTS (cont'd) viii
Page
o • CHAPTER 3. Floral development of Ruppia maritime
;
var. maritima. - , ••• II ................... . 58
Abstract .................. 1 •••••••••••• 59
IntroductiOn · ........................ . 60
Material and Methode .................. 60
Obaèrvations · ........................... . 61
Diecussion ..... " ..................... . 65
Literature Cited ...................... 78
CHAPTER 4. Floral development of Zannichellia
palus tris ...................... " ...... . 80
Ab. traC! t .•••••••••••••••••••••.••.••••• 81
I:ntroduction · ......................... . 82
Haterial and Method. .................... 82
Oblervationa · ........................ . 83
DilcUlaion •••• -II ••••••••••••••••••••••• 87
Literature Citad ••••••••••••••••••••• 100
CBAPTlR 5. Plo raI developaent of Naja. flexilie •••• 102
Ab.tract .••. ~ ••••••••• It ••••••••••• 1.1 • ~ lG3
Introductlon ••••••••••••••••••••••••. 104
Matarial &ad Methode .••.••••••••••••• 105 •
Ob.enation. • ........................... lOS
~1.cu •• 1OD ••••••••••••••••••••••••••. 109
Lit.rature Clted ••••••••••••••••••••• 124
o ~ AID COICLUIJOI8 •••••••••••••••••••••••••••••••••••••• 127
LlftlAtuII ClDD XI' SUIIIArr AlI) OCIfCLVII0l8 • •••••••••••••••••• 138
. " ' .
.'

. - •
1
'é)
.,
,
IlI'rBODUCTI0N
and
DVln 'or LITBllATURE
, "
't
•
l'
t" j,
't i .,' "
i j -,
, .-
' .... l
" -• .. '" "
,"
, ~ '.-,

t
2
INTRODUCTION
Evolution of the monocotyledons has been a eontroversial subjeet
for many years (see reviews by Takhtajan, 1969; Moore and Uhl, 1973).
One group that has figured prominently in this controversy iB the
Najada1es, Bome members of vhich have long been of interest aB pOBsibly
primitive monocotyledons. The belief was that monocoty1edons evolved
from primitive aquatic di~otyledons (i.e. Nymphaeales) through secondary
108s of vessels and simplification of flowets (Takhtajan, 1969). Thus,
tpe Najadales vith their aquatie habitat, apoearpous gynoecia (like those l found in so-called primitive dicotyledons) vere proposed a8 possible
intermediate8 between diootyledons and monocotyledons (Schaffner, 1904;
Hallier, 1905; Parkin, 1923). This crucial position brought abou~ a
great deal of interest in thia heterogeneou8 group of planta. In order
to clarify relatioDships vithin this group and to establish phylogenetic
lineage, the Nsjada1ea were studied from almost every coneeivable point
of vlew; vegetative (Chrysler, 1907; Arber, 1921; Singh, 1964; Sculthorpe,
1967), floral morphology and anatomy (Chry.ler, 1907; Uh1, 1947; 8ingh,
1965a, 1965b, 1966; Seu1thorpe, 1967), cyto1ogy (Chase. 1947; Harada,
'1955; Sharma and Chatterjee, ~966; Mlera, 1972), embryology (Swamy and
Lak.ha8nan, 1962; Svamy and Par .. e .. aran, 1963; y .... hita, 1972), pollen
d.velopme~t and morphology (Meier, 1966, Schvanltf' 1967) and blochemlstry
( •• e Kubitski, 1972). But one vital ar ... floral develo,.en~. ha. been
almo.t ca.pletely lsnored for cloae to one buDdred ,.ar ••
Th. flover. of the Najadale" thoulh, ha •• been the focus of auch
.tudy and .. ny .arpholol1cal controver.ie. have lurrounded th .. , for
exa8P1e. whether Pota.ol~tOD haa a tetr ... roUl flower or an inflore.cence
. •

,
- ..
3
composed of four ataminate flowers about one four-carpellate pistillate
flower; whether Potamogeton and Ruppia have perianth mèmbers or out-
growtha on the stamen connectives; what 18 the nature of the envelope
about the pistil1ate flower in Zannichel1ia and those about the stamen
in Najas. Theae and other controverslal questions, along with their
subsequent taxonomie and evolutlonary implications, have been thoroughly
8ummed up in the worka of Uhl (1947), Singh (1965), Sattler (1965) and
Scu1thorpe (1967). Most of the past worka, though, have been based
solely on mature floral structure, which as we know,represnts only one
slice of the space-time.continuum in which an orgauiam exists.
The earliest work on the Najadales was primarily descriptiv~ (Gay.
l8~~). One of the firat investigations of the floral development. though,
vas done by Hegelmaler (1870). He studled the ontogeny of the f10wer of
PotllQleton crlapu8 and c1early showed that the 8teri~~ appendages devel-
oped acropetally llke typical perianth .members and not like outgrowth8 of
the stamen connectives. Simllar re.ults were shawn hy Goebel (1911),
working on ~he floral developaent of !. natane and!. densua. Unfortunately.
th'.e isolated warka contained little comparative data and were virtua11y
ignored for many year. Whilè others (1.e. Rend1e, 1930; Markgraf, 1936. ,
1974) contlnued to argue that this appendage was part 'of the Btamen
coanective. More reeently, Sattler (1965), 1n a floral developmental
study of lotlIOIeton rl~hard.onilJ conf1raed Begelaaier ' • and Go.bel's
original vorka. A.ide fro. th1_, other tecent .tudi •• on Pota.oseton have
eapluada.ct taxonoay (Rayne. f 1974). .. the ouly knowD floTal d."").0PMlltal Itu4y of l.uppia urltiu .e. done
by Orave. (1908). 111. iIlvut1aatioll ... incOliplete aDcl relled only ob.

4
sectioned material. There were camparisons made with Potamogeton • .
Zanniche1l1a and Najas but they involved pt'i1llàrily mictosporogenesfs,
megasporogenesis and embryogenesis.
Twb geners within the Najadsles that have attracted much attention
from developmental floral morphologists are Zannichellia and Najas. This
may be due in part ta the simple nature of the flowers and the compressed
fertile shoots vhich bring many d~fferent stages of development into
close proximity. Magnus (1870, 1894) in his monagraphé, was one of the'
firet ta look at ~loral development in the genus Najas. Schumann (1892)
followed vith his own ontogenetic study of the flowera of Najas and
Zanpiche1lia in which he diaagreed with Magnus about the nature of the
"carpel" in Najas. Magnus homologized the structure about the ovule with
a bract or rudimentsry perianth, comparing it vith the envelope about the
pi.tillate flover in Zannichell1a; while Schumann si.ply considered ft 8
carpel. The finest work on floral development of the above tvo genera,
though, vas done by Campbell (1897). He summarized the works of Magnus
and Schumann and continued where they 1eft off, uBtng primarily histologieal
and some organographic data. He conc1uded in his atudy that Zannichellia
and Na'a. are primitive monocotyledons that are d~rived directly from the
hetaro.porou8 Filie.lea. Hovever, his phylogenetic speculation has 'been
queationed (8ee, e.g., UbI, 1947~, and hi. reliance on mainly hiaUbgenes1s'
has 81ven a,.lanted and et tt.e. incorrect vtev of the floral developaent.
So.. of tba more reeent work that has beau don. on the antoseny of the
flover of lI1u. 1nc1ud. that by Venute.h (1956) 'who .tudied the develop-
-.nt of tha .t .. tnate flover in!. palu.ttis and by Sattler and Cifford
(1967) who inve.ti.ated onto,enetie ~d hiatolo,ieal chant •• in th •• hoot
ti, of !. Ig!dalup!9li'.
~ ........ -------------------- - ~'
•

lt il evident fram the above, that the vork done on floral
dev.lapaent in the Najadalel is very spar.e. As a technique for the
.tudy of floral developaent vaa recently refined by Sattler (1968) and
co.vorkers. an examlnation of certain me.bers of the Najadales was
thought u.eful in approaching the many taxonomie. phylogenetie and
morphologieal probleml lnherent ta th!a group and higher planta in 1
generd.
•
, J ,r
< \
• ..
,; , . :;'i\ . ,-1.,''''
i
--~j

/
•
r.
r , ,
•
LlTERATURE ClTED 6
Arber, A. 1920. Water ~ntl. University Press, Cambridge.
Ca.pbell, D. H. 1897. A morpho1ogical study of Najas and Zanniche1l1a.
Proc. Ca1if. Acad. Sci. ser. 3, 1: 1-61.
Chase, S.S. 1947. Polyploidy in an immersed aquatie angiosperme
Am. J. Bot. 34: 581-582.
Chrys1er, M.A. 1907. 'The structure and relationship of the
Potamogetonaceae and a11ied families. Bot. Gaz. 44: 161-188.
Gay, M.J. 1854. Etudes organographiques sur la famille des Potamêe.
Premier ~moire: Sur les genre Potamogeton, Spiril1us e~
Groenlandia. C.R. Acad. Sci. (Paris) 38: 702-705.
Goebe1, K. 1911. Morphologishe und bio1ogische Bemerkungen. 19. Cher
"gepaarte" Blattanbaen. Flora 103: 248-256.
Grave., A.H. 1908. The morphology of Ruepla maritima. Trans. Connecticut
Acad. Art. and Sei. 14: 59-170.
Hallier, H. 1905. Provilional .ch_e of the natura1. (phylogenetic)
\ ayat_ of floverina pIanu. New Phytol. 4: lSl-l62. . -Barada, 1. 1956. Cyto1oaieal .tudi •• in ae1obiae, 1. Chromosome idiograms
and a liat of chro.o.a.e nuab.ra in'.evln fsai1ies. Cytologia
n(): 306-328.
Bayne •• 1.1. 1974. A revi.ioD of North "'~lcan Pota.ogeton 8ubsection •
Pua1111 (Potaogetonacea.). Rhodora 1!: 564-649 .
JIe •• laal.r, r. 1870. fIbt.r di. Ene,,1ck1una der Illlthenthaile von
las-utop. Bot. Zeit ... 11: 283-320.

r , r. ,
•
"
•
-
7
Kubltzki, K. 1972. Prob1eme der Gro8systematik der BlUtenpflanzen.
Ber. Deutsch. Bot. Gee. 85: 259-277.
Kagnus, P. 1870. Be1trHge zur Kenntnlss der Catung Naja. L. Berlin.
1894. Über die Gat~ung Najas. Ber. d. deutsch. bot. ~----
Gese1lsch. 12: 214-224.
Markgraf, F. 1936. B1Utenbau und Verwandtschaft bel den einfa~8ten
Helobiae. Ber. Deu~sch. Bot. Gesell. 54: 191-229.
------ 1974. Morpho1oglsche K1elngkeiten mit grHsseren Folgen.
Phyton (Austrta) 16t 105-116.
Meh~h N. R. 1966. Development of pol~en gralns of Helobiae and on their
relation to the Nymphaeaceae. Bot. Zh. 51(12): 1736-1740
(in Ru88ian).
Klara, M.P. 1972. Cytologieal studies in .o.e Indlan Potamoseton and
Aponoseton species. Bull. Botan. Soc. Bengal 26(1): 47-51.
Moore, H.!., Jr. and N.W. Uhl. 1973. The monocotyledons: the1r evolut10n
and cotlparaUve biol0tNt. VI. Palma and the!r origin and
evo1utlon of .auocotyledons. Q. Rev. Biol. 48(3): 414-416.
'Parkin. J. 1923. The atrablliathaory of anaiOlpet'1llOUII desc.nt. Proc.
Lin. Soe. Lond., lot. ID: 51-64.
&endl •• A.B. 1930. Th. c1 ... ifièation of flov.ring planta. Vol. 1. '
Univ. ·Pr .... Cubridge •
Sattler t 8.. 1965. Periantll d6valopaent of Potao,atoD -richard.onii.
AIl. J. lot. H(1): 3,..41 •

.... -
Sattler, R. 1967. Ontogenetic ~istochemical changes in the
8
shoot tip of Na las guada1upensis (Sprengel) Morong.
11: 419-428.
'968. A technique for the study of flora 1 deve lc;>pment. ,
Cano J. Bot. ~: 720-722.
. ' __________ , and E.M. Giffard, Jr. 1967. On~ogenetic and histochemical
changes in the shoot tip of Nalas guadalupensis (Sprenge1)
'. Marang. Phytomorphology 12: 419-428.
Schaffner, J.H. 1904. Sorne morphologieal ~eeuliarities of the
Nymphaeaceae and Helobiae. Ohio Nat., ~: 83-92.
Schumann, K. 1892. Morphologisehe Studien. Leipzig. . ,
Schwanitz, G. 1967. Untersuehungen ~ur Postmeiotisehen Mierosporogenese:
1. Morphogenese des Ruppia - Pollen. Pollen Spores 2(1): 9·48.
Scu1thorpe, C.D. 1967. The biology of aquatie vBseu1ar plants. Edward
Arnold (Publishers) Ltd. London.
" S1ngh, V. 1964. Morphologiesl and anatomiea1 studies in Helobiae. 1. \
\ Vegetative anatomy of sorne members of Potamogetonaceae.
Proc. Indian Acad. Sei., ser. B, 60:'214-231.
1965a. Morpho1ogica1 and anatomiea1 studies in He1obiae.
II. Vaseu1ar anatomy of the flower of Potamogetonaceae.
1965b. Morphologiesl and anatomiesl studles ln Helobiae.
III. Vasculsr anatomy of the node and flower of Najadaceae. ,
Proc. Indien Aead. Sei":', ser. B, .§!: 98-108.

• 9
S1ngh, V. 1966. Morphological and anatomiea1 atudies in ae1oblae.
X. Trends of speeialization in placentation in He1obiae.
Curr. Sei. 35(1): 250-251. ,
Sharma, A.K. and T. Chatterjee. 1967. Cytotaxonomy of helob1ae with
\ .,eeial reference to mode of evolue(on. Cytolog1a 32:
286-307.
Swamy, B.G.L. and K.K. Lakshmanan. 1962. Contributlons to the embryology
of the Najadacéae. J. Indian bot. Soc. 41: 247-267.
_________ and N. Parameswaran. 1963. The heIobial endospérm. Biol.
Rev. 38: 1-50.
Takhtajan, A. 1969. Flowering plants; origin and dispersal. OliVer
Boyd: Edinburgh (trans1ated from Ru.sian by C. Jeffrey).
UhI, N.W. 1947. Studiea ln the floral morpho1ogy and anatomy of certain
membera of the Helob1ae. Ph.D. Thesis, Cornell University,
Ithaca, N.Y.
Venkateah, C.S. 1956. Structure and dehiscence of the anther in Rajas.
Bot. Not. !Q!: 75-82.
Y .... hit •• T. 1972. liaenartl,_ Wurae1an1ale 4e. Eabryoa bei 10ppta ~
"rlttaA L. Baltr. Blol. Pflanzan~: 157-170 •
. ,

\ 10
•• ...,
,-
OBsnVATIONS
and
DISCUSSIONS
1
• - - -• 1~' t

-,"" .
11 ..
•
J.
\
CHAPTER l
FLœAL ŒWLOPKBNT OF POTIt.MQÇETON RICHARDSONII
'.~ , ... ,- ~ .-.' . , , , , - "
" ,

'. ",
12
• 1"
.. • AB STRAC T
li)
The inception and development of the sterile floral append-aes
of Potamoseton ricbard.onil have been re-lnvesttgated wlth a refined
dissection technique (Sattler, 1968) and improved microtechntcal
Bethods (F(der and O'Brien, 1968). The result8 obtaine4 ,oy Sattler
(1965) are conflrmed. 1.e., the sterile appendages are initlated at • fi ,
the flanks of the floral apex before the atamen primordia are formed.
Conlequently, they may be bomologlzed with tepals or perianth mem-.. ' bers, although in the .. ~ure flower they are lnsefted at the stamen
".
connective, due to growth between and at the base of each developing
tepal and stamen. Eac~arpel arises as a radi.l primordl~ whlch
beca.es peltate lmmediately a~tër lts inception •. One ovule prlmor-,-
dium 11 lnitiated at the crosi-zone. The stigma becomes bilobed.
A sltght out8rowth de~ops at t~e abaxial side of the style. fbe floral apex bal a two-l.yered tuniéa. The prtœordia of the tepals,
carpel., and ovule. art.e by perieltnal divi.ions.in the second tunica
,layer, wber.a. the at..-n primordia are initiaced by'periclinal dlvi-
lion. in the corpu. and.'lecond tuntca l"yer. Variation in floral r
Pattern" .. peci.Uy with reaard to the nusaber of .ppend..... ha. been , l '
ob •• rYftd in flower. nea:r tbe t1p of the .1nf1ore.cenc~ ail. " \
.:.
~ .1
1
) • 1
~. ,
"

.... e~ .. ~----~----------------------------~=.---------------------------~--~--- -
.t
•
"
13
INTROOOCTION .;Jo'
The Interpretation of the flower of Potamogeton 'has been contro--, t
versial for over a hundred ,ars. With regard, to the sterile appendages
(which are inserted at the- stamen connectives), Sattl!!r (1965) showed
that they are initiated 1ike perianth membera. Markgraf (personal com-
munication) expressed doubts about these findings, still giving pref
erence to his previous interpretation accordlng ta which the sterile
appendages are only ou~growth. of the connective, i.e., they are not
homologous to perianth members (Markgraf, 1936). 1
Since the publication of Sattler 'a (1965) article,- tèchniques have
been ~ch improved. With a refined dissection technique (Satt1er,
we can now show detaila of primordial inception mueh more a;urat y. - \JJ
Even patterns of protoderma1 cells can be demonstrated in t surface
viewa. Further~re, the Methode for preservation and s~i~ng of the ,. , 1 ..
material have been greatly imp~oved by Feder and OtBri~n (1968). With
.his considerable technical progre8s and ~n the light of the still-pre-
>vailing di.agreement on the interpretation of thé sterile appendages, it
app.ared de.irable to r.-lnves~i8ate initiation and development of theae
appendagea with the lmproved technique.. ln addition, ineeptton and
development of the androeeium and gynoeeium have been studied, and vari
atlon Of~al atrUttur. ha. be •• tak •• i.to eon.ideratlon,
MATOUL AND METSons
Tb. _t.rtal of ·Pot.., •• ton richardsonii (Benn.) Rydb. '1" coll
acte4 by che 'author in Lac Rertel at MOnt St. Hilaire near Montreal in
" 1ate June and .ar1y JUly, 1971 and 1972. 'art, of the .. terial wal fixed
.'
'.

,/
•
14....--
and preserved in FM (formalin-aeetie-acid"a.leohol) ~e,fore -being stained
in alcoholic acid fuchsin, dissected, and photographed entirely immersed
in 1007. ethyl alcohol following the technique of Sattler (~968). Material
used for sectioning was fixed in 3% glutaraldehyde at 0 C. dehydrated,
and embedded in glycol methacrylate by the general procedures of Feder and
O'Brien (1968). Sections were eut on an ultramicrotome and stained with
0.057. toluidine blue solution in benzoate buffer, ~H 4.4 (Sidman. Mottla,
and Feder, 1961). Cleared preparations were made by the technique of
Rodin and Davis (1967).
OBSERVATIONS
Or~anography
The flower8 are borne on apike8 and are arranged in-alternating
whorls of three. Each flower has four broad.bract-like tepals which are
affixed in the région of the stamen connective. The four nearly sessile
stamens are direct l, opposite the tepals and have large, extrorse anther8~
u.ually a single wborl of four 8eparate carpels alternates wlth the
8tamens. The ovary of each carpe! i. unilocular and contains a single
anatropous ovule attached ta the adaxial portion of the pi8ttl. The style
ia short and bifureates to fora a two-lobed stigma.
Oraanoaenelll
The inflorelcence d.velopl either fram a vegetative apex (Plgl. 1,2)
or gravI out ln the uU of a hat and aa older Inflorucenee (Fig. 3),
The floral bud. are firat Initiat'd on the lower portion of the lnflores-
cenee ub and are lubttmded by youtlf bract pd,lIOt'clia, (P1g. 2), The
Youa& floral bud iner ..... in Ih •• becOIDina doae-.h&ped w:1thout &11y
,k
'. ,

•
l~
external indications at first of the developing floral appendages
(Fig. ), AlI primordia of the floral appendages arise acropetally,
with the lateral tepale being the first ta appear. The two lateral
tepal primordia are inltiated almost simultaneously, growing out as
broad elongate protuberances (Figs. 4,5), The two median tepal pri-
mordia f~rm 800n after, with a slight developmental lag between the
upper median tepal (initiated first) and the lover median tepal (Fige,
5,6). With the development of the tepals, the floral bud becomes
flattened and almost square in the horizontal plane (Fig. 5).
The two laterai stamens appear almost simultaneously opposite the
lateral tepal prlmordta and grow out as elongate prlmordia (Fig. 6).
Soon after i~itiation, more rapid upgrowth occurs at the lateral por-
tions of the primordia, thus beginning the differentiation of the anther'
thecae. There i6 a very short lag between the initiation of the Iateral
stamen8 and median stamens (Fig, 7), The two median Btamen primordia
have a very short (if Any) plastochron, the upper possibly preceding the
lover, and they develop opposite"the median tepals in a manner similar
to the 1ateral atamens (Figa. 6-9).
After the inceptton and early development of the tepal and stamen
primordla, the floral apex, whlch had remained relatlvely inactive
(appearlng fIat ln the harl~ontal plane), becomes slightly dome-ahaped
again '(Fig. 10jr ~e four synoecia1 primordia (for definltion see Sattler,
1973, p. 21) ~e tuttiated at about the a.me time through differential up
,rovth in the re,lon. of.the ~loral apex, alternatlng with the tepal. and
atamen. (Fli. 10). A. the gynoecial primordlua expands, it. peripheral
re,ton crawl more tapidly, thua foraing a peltate primordium (Fi, •. 11~13).
[
____ ~ __ · __ ~ ______ ~t

•
The resulting carpellary tube develops at an equal rate for a short
period of time before initiating the ovule at the adaxial portion of
the carpel wall (Figs. 14, 18). After ovule initiation the abaxial •
18
region of the carpe~ wall grows more râpidly. enveloping the young ovule
prlmordiuœ (Fig. 15). Even at this early stage in gynoecial development,
the two-lobed s~igma can be seen forming by differential upgrowth of the
distal portion of the carpel wall. Shortly after the ovule is enclosed
(Fig. 16), it inHiates two integumen'ts, the inner one forming first and
developing the micropyle (Fig. 19). The ovule then begins to invert so
that the micropyle points downward. As the short style develops~ it
narrowa towards ita distal end (Fig. 21) and forms a slight outgrowth
.• on its abaxial aide Just below the bilobed stigma (Figs. 21, 22).
A number df unu8ual variations were n~particularlY ln floral
bude close to the inflorescence apex. Most common was variation in the
number of carpels, including trtmerou8 (Fig. 12) and even dimerouB ....
(Fig. 13) 'ynoecia, but with tetramerou8 androeeium and perianthe Some
~natanees of eompletely tri.erous flovera were also observed (Fig. i6);
and in one floral bud, h1gh upon the 'infloreacence axis, upsrowth between
two adjacent tepals resu1ted ln the formation on an involueral bract-like
appendage (Fig. 8).
H1etoleneah
As de.crib,d ptevioualy (Sattler, 1965). the floral apex has a two\
layered tunica, tbe second layer of wbicb i. prt.arily involved in pri-
.ordlua initiation. Tbe br.ct i8 Initiated at the junction of floral
apex and tn~lor •• cenc. exl. by per1clinal d191.1oo. ln the •• cond cell
layer, doaa vith cOI"t •• pcmdina ..... 1101\ bd aUe11nal divialonl of tbe
"

•
, 17
surface cells (Fig. 24). At the subsurface level it never involves
more than one or two rows of eells and soon degenerates at about the
stage of gyno~eial inception (Fig. 28), The tepals are initiated through
perielinal divisions in the second tunica layer of the floral apex in
close association with expansion and increased anticlinal divisions of
the surface cells in the region of the tepal primordia (Fig. 24). At
the early stage of tepal initiation there are no signa of stamen incep-
tion, aside from slight cellular expansion in the regions of the pros-
peetive stamen primordia (Fig. 25).
The stamen arises not only as a result of perielinal divisions in /
the second tunica layer but also due to corresponding, simultaneous
periclinal divisions in the corpus (Fig. 26). Soon after stamen incep-
tion, cellular expansion can be observed in the second tunica layer
between the tepal and stamen primordi~. indicating the initiation of
interprimordial upgrowth (Fig. 27). As the floral bùd develops, upgrowth
betwe'n and below both tepal and Itemen primordia becomes more extensive
(Fla', 28, 29).
Perlelinal divisions in the second tunlea layer 18 also Involved in
the initiation of the gynoecial prl~rdi. (Fig. 28). The gynoeclal prl
mordia arow outvard and are distinct from the floral apex before beeomlng !
peltate (Pl,. 4~).
A dqle proc_lal .trand. vhlch dw~lop. from one of the _ln
Itrand. in the lnlloreaceace .. 1 •• differeatiat •• in the central portion
of the 10Una floral bud. POùr proc-.bial .trand. differentiete acropetally
toward the ffr.t re,ioa. of arowtb. tb. tepal prt.ordl~ A. the at ... n
..

;
•
/ 18
primordia develop, at about the stage of gynoecial primordia initia-
tion, procambial strands are Induced towards them, branching trom the
tepal procambial 8trand8 (Fig. 28). The fork of tepal and stamen strands
remains below the region of upgrowth and, even in the near mature
flower, i8 found very close to the central strand in the pedicel (Fig. 30).
The four gynoecial proeambial strands differentiate from the central
strand in the young pedicel towards the gynoecial prlmordia at the stage
when they are becoming peltate (Fig. 29). At a later stage, a ventral
procambial strand differentiates towards the developing ovule, whereas
the original gynoecia1 strand develops towards the prospective stigma
(Fig, 30),
DISCUSSION
Concerning the sterile appendages, the observations of Satt1er (1965)
are confirmed: the primordia of the$e appendages are initiated at the
flanks of the floral apex slightly before thoae of the stamens. Since
they are phyllomic in other respects', they are best interpreted as phyl-
lames and not aB enations or outgrowths of a phyllome. More specifieal1y,
they may be referred to .s tepals or periant~ members if the flover is
interpreted a. a manuial organ sy.stem; If, however, thia flover 18 homo-
loglzed to a conden.ed inflore.cence (i.e., a polyaxial system, see Uhl, .-1947), then the sterile phyllomes would be referred to aS,bracts. A. has
been pointed out before (aattler, 1965; Croizat, 1966), the Potamoge~on
flover is a structure aomevhat Inter.ediate betveen a flover and an
lnflore.cence, although .uch clo.er to a f~.r. Consequently, the f
.terite phyllo.e. are partially homololou, to tepala and bracts (for the
1 nation of "partial tu.olol7" •• e Sattler, 1966), althouah IlUch more bolDo-....

•
19
Iogous to tepals than to braets. For praetieal purposes they are there-
fore referred to as tepals.
The position of these tepals in the mature flower is due to inter-
primordial and interealary growth at the base of the young tepals and
stamens. Interealary growth occurs in many taxa and may involve aIl
sorts of structures of the vegetative region (Bugnon, 1959) as weIl as
the reproductive region (Kaplan, 1967), The tepal-stamen connection in
Potamogeton is just a special case of this widespread phenomenon. It iB
important to keep in mind that the interprimordial ànd intercalary growth
occurs in a region which cannot be clearly asslgned to either the tepal
or stamen primordia, nor the young receptacle. Consequently, the tissues
produced fro~ the growing region at the base of the prlmordia cannot be #
classified as belortging to the axis, or the stamen, or the tepal alone,
• This means saying that the tepsl is insetted at the stamen connective is
not quite correct. First of aIl, the insertion basE of the tepal cannot
be clearly delimited. It might reach down to the receptacle. But it ls -meaningless to search for a delimitation where none exists (or, at least,
where no criteria are available for determining a natural limit).
Secondly, the whole interprimordial and intercalary upgrowth ~annot be
strictly aSligned ta one category, although one would tend to look at
most of it as .tamen.
In Potamoa.ton d.DIU. (Polluazoy and Sattler, 1973) each of the laterai
ata.ena i. initiatad a. two distinct primordia, whereas the median stamens
ari ...... in81e, elonaate primordia. Here in potaœoaeton richardaonil aIl
.t-..n. are ibltiatad a. aiUSI., alona_ted prt.Ordla like tbe median stamans
ln 1L dea., ••

•
20
The development of the gynoecium resembles that of Potamogeton
densus tposluszny and Sattler, 1973) with two major exceptions. (1) In
~ densus the cross-zone arises on the floral apex, whereas in ~
richardsonii even the cross-zone forms on the gynoecial primordium.
(2) In ~ densus the outgrowth on the abaxial side of the style is much
more extensive than in ~ richardsonii.
As in Potamogeton densus (Posluszny and Sattler, 1973), variations
were noted in the patterns of floral buds. In particular, variation
occurred with regard to the number of primordia (see above u!'lder "Organo
genesis"). Structural deviations such as the formation of an involucral
bract-like appendage were a1so observed. In contrast to ~ densus, the
percentage of deviations from the normal pattern was BmBll. This may be
explained by the rule that variations tend to occur malnly in f10wers
near the tip of the inflorescence axis. In ~ richardsonii, which has
many flowers (Figs. 9-27) on one inflorescence, only the few near the tip
may deviate. In ~ densus, with only two flowers per inflorescence, aIl
flowers are near the tip of the inflorescence axls# and consequently, the
percentage of variation\ is much higher. lt ia i.tere~ting to note that in
both ~ denaua and lL richardlonti the upper portion of flowers, i.e.,
the portion nearer to the tip of the lnflorescence, tends to deviate much
more tban tbe lower portion of the flower~ For example, if one carpel
il abaent, it i. usual~y one of the two in the upper portion of the flower l\A.
(Fia. 12); or if an unulual atructure like the'involueral bract-like
'"' appenda&e (rl,. 8) occurl, it i8 a1ao formed in tbe upper pOlition of the
flover •

21
. ACDOWLE OOEMENT
This lnvestlgation vas supported by a Postgraduate Scho1ar-
8h~p 1560 (to U.P.} and by an operating grant A2594 (to R.S.) from
the National Research Couneil of Canada. We wish to thank ~. Mar-
garet McCully pf Carleton University, Ottawa for her kind assistance
in the preparations of the sect~on8 in this chapter.
..
..
•
..
• , ,. ~ l
l .~.. /~, ,.
, .. ", • r ~ 'f .. .r." ~

• •
-
LIst OF ABBREVlATIONS
Al - lateral stamen or 1ta pr1mordlum.
Am - median st8men or lts prlmordium.
Ac • stamen ~onnective.
As - stamen procamblal strand.
B • bract or its primordlum. l'
F • floral apex.
G - carpel or its prlmordlum.
Il - lnner integument or its prlmordium.
10 - outer integument or it8 prlmordium.
L - vegetative appendage or its primordium.
N - nucellus
o - ovule or lts prlmordlum.
" P - tepal or its prlmordlum.
Pl • lateral tepal or ita prlmordlum.
Pm - medlan tepal or lta- prlmordlum.
P. - tepal proc .. blal strand.
R - inflorescence apex (re~oductlve apex).
r - removed, e.g. rt - th.ca removed.
Si • eti..- or developing .t! ....
5, • .t,le or developiaa Ityle. ~
8'0 • outarovth trOll It,te.
T
•
••
'" . . '-

23
,\ •
1
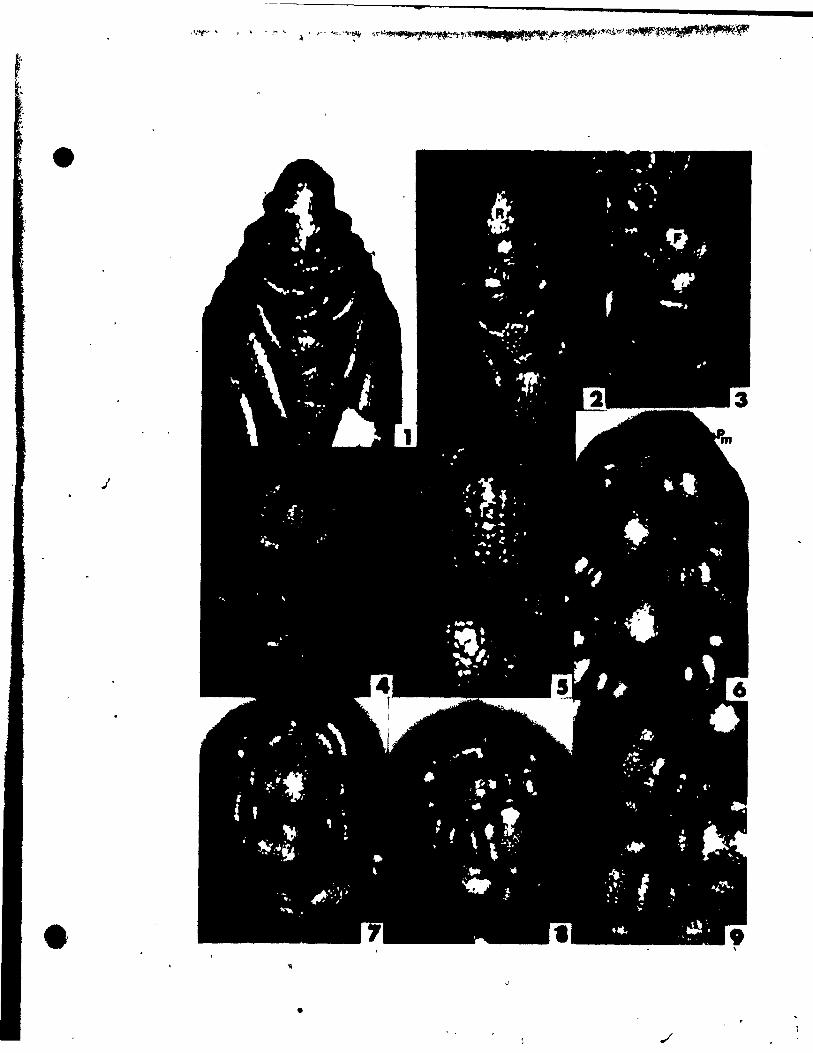
PLATE t Figs. 1-9
Fig. 1. Side view of shoot tip. x 146,
Figs. 2-4. Side view of inflorescences (R-inflorescence apex),
Plg. 2. Simultaneous initiation of floral bud (F) and subtendfng bract
(B). x 146.
Fig. 3. Preceding tepal initiation, the floral bud grows out. becoming
dome-shaped. The inflorescence axis is aubtended by a
sheathing vegetati~e appendage (L). x 146.
Fig. 4. Center top view of a floral bud, showing lateral,tepal (Pl)
initiation. x 146.
Fig. 5. Top view of floral bud at the st.ge of Median tepal (Pm) initi
ation. x 246.
Pig, 6. Young inflorescence with floral buds, showlng lateral stamen
(Al) initiation. Some upgrowth between the lateral tepsl and
stamen can already be aeen. x'l46.
FIg. 7. Top view of floral bud on wb~ch the lateral (Al) and median
(Am)- atamens are developlng an~ beglnning to overgrow the adja-
Pil. 8.
cent tepals. x 146.
Top vi .. of a floral bud. elose to tQe tlp of the inflorescence
apex, exhibltlng unueua1 up.rowth between two tepale, creating
an Inwlucral-Ute bract (1'). Je 146.
'11. 9. Oblique vi_ of • latet'al at ... n (Al) and t.pa~ (Pl)' x 146.
/ "
1
1
•
-
j
..
•

, 1
1 J 1
24
(
PLATE II Figs. 10-17
Flgs. 10-17. Top views of floral buda at the stages of
gynoecial development.
Flg. 10. Two floTal buds Initiating gynoeclal prlmordia (G). The
upper bud 18 a~ a somewhat younger stage. x 146 •.
Fig. Il. Floral bud with gynoecial prlmordia (G) becoming pe1tate.
~, x 146.
Figs. 12 anu 13. IWo buda, at a slight1y aIder stage than Fig. Il,
showing variations in the number of ~ynoecia1 primordia.
The bud in Flg. 12 has a trimerous gynoeci4m, whi1e the one
in Ftg. 13 ha. only a dtœerous gynoecium. x 146.
Fig. 14. OVules initiated at the adaxial portion of ~he young carpel
wall. x 146.
Fig. 15. C4rpel wall beginning to grow up and a~t the developing
ovule. x 146.
Fig. 16. TrimeroU8 floral bud, in which the ovules have been C~
pletely encl~.ed by the carpel wall. x 146.
Fig. 17. Bllobed It~gma (Si) beg!nnlng to dlfferentiate.together with
slight outgrowth at the abaxial portion of the style (~yo).
x 146.
, ,..
•
. ,
1
• . ~
;;.t ••• ,If!,

-
•
L ,
'"

--~~~-------------
"" , ......... ~.~ '"- .... "'t .. ~ «' ....... '.\
t
PlATE III Figs. 18-23
Figs. 18'~nd 19. Side vlew8 of two pistils sectloned
lonaitudinally to reveal developinl ovules.
"g. 18. Ovule (0) Just beginnlng to srow out. x 146.
Fig. 19. Older ovule with both inner (Ii) and outer (10
) tntelumenta
groWing about the nucellua (N). x 146.
Fig. 20. Side view of two bilobed stl~s (S1) and stylar outlrowtha
(Syo). x 146.
Fige. 21 ând 22. Side vlewa of nearly mature flovera •
... 'ig. 21. Portions of the flover and one theca (rT) removed in arder to
r
• .. show tepal (P) inserted high upon thé stamen connective (Ac).
" x 30. "
Pil. 22. Same .tage as Pig. 21. but undisaect~d. x 20.
Pig. 23. Top vlew of nearly mature flower. x 20.
'1
,"
, , ,
, 0
. . \ "J.,
"
/
ri' ,
1
1
/ .

1
•
.,
\

28
PLATE IV Figs. 24-30
Flgs. 24-29. Me4ian longitudinal sections through floral buda,
parallel to the inflorescence axis.
Fig, 24. Early stage, just initiating median tepals (Pm); the upper
one, away trom the bract (8), preceding the one adjacent to the
bract, x 480.
Fig. 25. A slightly o~der atale than in Fig. 24, showing prlmordla of both
median tepals. No clear indication of stamen prlmordla can yet
be seen. x 480.
Flgs. 26 and 27. Stages of stamen inception.
Fig. 26. Cell elongation in the second tunica layer and periclinal divi-
sions in the corpus region of the incipient median stamen pri-
mordium (A ). x 480. m
Flg. 27. Approximately the same stage as in Fig. 26. but showing a
periclinal division in the second tunica layer in the area of
the median stamen Inception (A). Celi expansion between the m
median tepai and stamen prlmo1'dia.can aiready be seen. x 480.
Flg. 28. A floral bud At the stage of inéeption of gynoecia~ primordia
(G). \rocambial strands are already developing in the tepals
Flg. '29.
Fig. JO.
and .tame~ •• x 192.
Developlng gynoeclal prtao1'dia CG) be~a.ing peltate. x 192.
SUe vi..., of, clearilll of .... ture flower t .bovlng forlt of tepai • rl
(PI) and It-.n (b) va.cular .trand. The tepal "a. removed
(1'P). x 30. 0
.. f,'"

'ffl',; """'.ft,' '."lII; ,ft':" Y<je '<;'i.~ ""l:",)lI>I""t.Jil._~ ~"/~"""'. ..... ,..t'"f>il!f,': i!J~'-" .' .' "-7' ,."", , -
•
Si ~ ,
• p
30

" . , ..... .,.",.,....".,
268
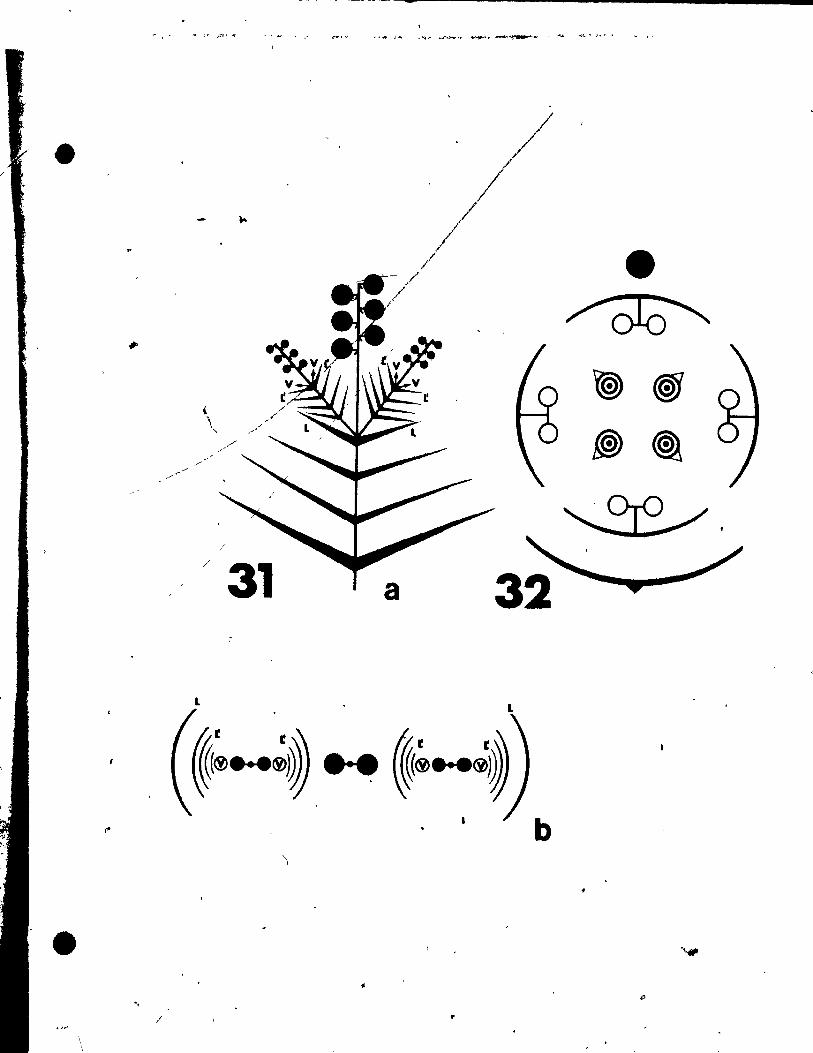
rigs. 31 and 32
Fig. 31. Qiagrammatic representation of the pattern of inflorescence, li t)
from side (a) and top (b) view.
Fig. 32. Floral diagram.
, ,
• ~ f
.. /
o
, 1
, "
1 j
/
\ \ \
/Ii>
"
. .-r .- _ -. 1 T .!'r~' "1' ' , .. , •
{
- ...
, \
"-
~ ~
/
... / /
/
/31
L
1
, ~... • ... ,.. _'Q"'- ~'t .,.,...,.,.0' _~~ "_....... ~., J.~ • ..."
/ • ~
t@ ~
~ ~
~.
a
L
b
Il

• $
•
u
--
27
LlTERATURE CITED
Bugnon, F. 1959. E1~mente d'un chapitre complexe de morphologie
v61~ta1e: Le~ d~formation8 nodales de la pousse par croissance
,intercalaire longitudinale chez les plantes 1 fleurs. Dul1 Sei.
Bourgogne 1i: 29-69.
Croizat. L. 1966. Observations on the ovary of the Juglandaceae.
Southwee t. Nat. 11: 72-117.
Feder. N., and T.P. O'Brien. 1968. Plant mierotechnique: some
princlp1es and new methods. Amer. J. Bot. 55: 123-143.
tAplan, D.R. 1967. Floral' morphology, organogene.l. and Interpretation
of the infedor ovary in Downingi. 1:f"c1ga1upi1. Amer. J. Bot • .2i'
1214-1290.
Markgraf. F. 1936. BIUtenbau und Verwandachaft bei den einfachsten
ge1obiae. Ber. Dtsch. Bot. Ges. 1!: 191-229.
POl1uazny, U., and R. Sattler. 1973. Floral development of Potamogeton
d!Q.ya. Cao. J. Bot. 51: 647-656.
Rodin, R.J. and a.B. Davie. 1967. The ua_ of papain in clearing plant
tb.ue. for vho1aouDta.. Stain techuol. 42: 203-206.
8att1_r, 1. 1965. P.riantb dev,lopaent of Pota.oseton rlchard~onli.
AMr. J. lot. 52: 35-41.
1966 •. Towarda a -.ore adequate approach to cOII'pàratlve
.,rpho1olY. l'hyta.rphololY 1!: 417-429.

.~
Sattl.r,. R. 1968. A techt'lique for the atudy of floral development.
Cano J. Bot. 46: nO-722 • .
-----• 1973. Ofganogeneais of flowers. A photographie text-
atlas. Univeraity of Toronto Press, Toronto.
Sidman, R.L., P.A. Mottla, and N. Feder. 1961. Improved polyester , "
wax embedding for hiatology. Stain. Tec.hnol. ]!: 279-284.
Uhl, N.W. 1947. Studie. in the floral morphololY and anatomy of
certain .eabers of the Heloblae. Ph.D. Thesla, Cornell Unlv.,
lth.ca, N. Y •
....
) .
•
28
'\ .
" i
','

89
••
. '
CHAPTElt 2
FLCltAL ŒVELOPMENT OF .!'OTAMOGITON IQSUS
',.
'. , j
, . . , .' . ït'

•
30
ABSTRACT
The floral appendages of Potamogeton densu8 are inltlated in an acro-
petaI sequence. The first primordla to be seen externally are thoae of the
lateral tepals, though' sectioning young Horal huds (l'ongi tudinally.
paraI leI to the inflorescence axis) reveals initial activity in the règion
of the lower median (abaxial) tepal and stamen at a time when the floral -J
mer!stem la nat yet clearly demarcated. The lateral (transversal) stamens
are initlated simultaneously and unlike the median stamens each arises as
two separate primordia. The upper median (adaxi~l) tepai and stamen develop
late in relation to the other floral appendages, and in sorne specimens are
completely absent. Rates of growth of the primordia vary greatly. Though
the lower median tepal and stamen are initiated firat, they grow slowly
up to gynoecial inception, while the upper median tepal appears Iate in the
fil developmenta1 sequence but grows'rapidly, soon overtakin~ the other tepal
primordla. The four gynoecial prlmordia arise almost slmultaneously,
although vari~tion ln thelr sequence of i~ception occurs. Thé two-1ayered
tunica of the floral apices gives rise to aIl floral appendages through
perielinal divisions ln the second layer. The third layer (corpus) ls
involved a8 weIl in the initiation of the stamen prlmordla. Procambial
stranda develop aeropetally. lagg!ng behind primordial initiation. The
lateral stamen. though initiating aB two prim6rdia eaeh form a single,
central proeambial etrand, which differentiates after growth between the
two primordla of the theeae ha, oeeurred. A Irea~ amount of deviation from
the normal tettamerous'flower'is, found, including completely trimerous
flovera, tri.eroua gynoecia vith tetramerOUI perianth and androecium, and
orlana differentlating partially al tepale and partially as stamen'.
-------~- -----~~--~-~-----~~~\ ~-~-----------

31
INTRODUCTION
This study ls intend~d as a continuation of the w6rk on potamogeton.
rich.rdsonii (Satt1er 1965). It appeared desirab1e to investigate a
species very distantly related to f. richardsonii to find maximal vari
ation wlthin the genus. Potamogeton densus fu1fl11s this requlrement.
Some authors. such as Fourreau (1869). Buteher (1961). and Aalto (1970)
have even separàted this speeies as a monotypic genùs, Groenlandla.
Accordingl'y, a nUlllber of striking differenees in floral development' were
observed. Furthermore, a considerable range 01 variation was noted in
material of the s&me colony. These find!ngs are relevant not only to
the interpretation of the Potamogeto~ flower but generally to floral and
theoretlcal morphology.
MATERIAL AND METHODS
Fertile mater!al of Pot~geton densus L. waa çolleeted by Dr. P.
Leins in Ju1y, 1966 and 1970, at the Botanie Garden in Munlch, West
Germany. It was fixed and preserved in FAA (formalin-acetic aeid-alcohol)
before be~g sta1ned in alcoh6lic acid fuchsin, dissected. and photographed
entirely immersed in 1001. ethyl alcohol following the technique of Sattler
(1968).
The very same floral buds that vere photographed who1e vere dehydrated
in a tartiary buty1 a1eoho1 series and embedded ln tissuemat ~slng stan
dard techniques, Section. vere cut at 6 alcrometers (~), and were stalned
vith orange G. tannic Acld, and lron .lua (ShArman 1943),
OBSERVATIONS
Qrlanoçaphv
The lnflore.caDce of Pot_atOll deow con.bt. of two nover ••

o
32
opposite each other, ln the same plane as the d18tichously arranged
foliage leaves (Fig. 1). In most cases one of the flowers is situated
somewhatehlgher up on the axis (to be referred to here as the upper
floral bud, as opposed to the lower floral bud). ... Usually the mature flower has four stamens with almost sessile
extrorse anthers, eaeh of whlch bears a tepal on the connective, and
four unlovular carpels alternatlng with the stamens and tepals (Fig. 2).
The shoot apex 18 long and narrow, forming leaves in a dlstichous
arrangement, i.e., 1/2 phyllotaxie (Fig. 3), The firet external indl-
cations of floral evoeation are seen as the vegetative apex begins to
enlarge and beeomes bilateral (Fig. 4). As enlargement continues, each
of the two fIat sides starte init~ting a floral apex (Fig. 5). At the
S8mè tlme, in the axile of the last two vegetative appendages for~ed,
axillary buds u8ually develop, growlng vegetatively for a short time
before becoming reproductive and themselves developing axillary buds
(Figs. 6, 7). Thus, a cymose-like pattern of inflorescences res~lte
(Fig. 2).
QnanOlen,es 18
Even before the floral apex can be clearly dlstlngulshed on the )l
inflore.cenee axis. activity·in the region,of'~he lower median tepal and
stamen can already be noted. In a median section through a young bud at
this stage (longitudinally through the inflorescence axis). there are
alré_dy periellnal divisions in the sec~d layer of the two-layered
tunica, ln areas corres'fndlna to the laver median tepal (Pm) and add.l
tian_l peri~linal divi.i~n. in the third layer where the .~amen <Am> 1
appear. (FiS,. 46 1). In al.l1ghtly older bud. eut tran.vene1y to the
\

• •
33
inflorescenc~ aXiA, perielin.1 divi.ionR ln the 8~cond tunica lay~r arp
also seen to be involved in the 8imultaneou8 initiation of the lateral
tepala (Fig. 47). ,-Even,though there is a very small plastochron between the inItiation
of the lower median tepal and stamen on one hand, and the lateral tepals
on the oth, et', the rate of growth of these floral appendages varies ~reat ly. , . ... Thu8, the first primordia of floral appendages to be clearlY di8tingulshed
externally are those of the lateral tepals (Ffgs. 8, 48). The primordia
of the lower median tepal and stamen grow at a very 8l~ rate and are
rather difficult to make out ln the e.rlY stages (Fig. 9). ,
The lateral stamens appear next in the deveiopmental sequence. Up-
growth proceeda more rapidly in the central portion of the flotal bud
leaving a deprea'ion between the lateral tepal primordia and the floral
apex (Figs. 10, 12), This pattern of growth seems closely related to
the formation, subsequently, of the lateral stamens, each of'which arises
1 as two separate primordia, adjacent to the prlmordium of the lateral tepsl
(Flgs. 15. 16). The latetal atamens, in fact. not only °develop as (wo
separate primordia each, but the lower of theae prlmordia i8 initlated
before the upper (Fig8. Il, 13). Sections through the young androecial
p~lmordia at th!8 stage show the mode of initiation to be bath through,
periel!nal divisions in the second tunica layer and in the third layer ...../. ..... ,
corpuê regton, and confirm the observation that,\he lateral stamen arises
a. two separate primordia (Fig8, 49 ~ 50).
Tbe d •• elopment of the hpper median t , and stamen is not e.~y to
under.tand. In one tnflorelc~nce observe t the upper median tepal of thé
o l~.r floral bud II inltiated directly r
the prlmordia of the lateral
...
-

-t
•
34
, tepala and stamens, leavlng very litt le room for t~e upper median stamen
, J • J
'(Fig. 15). The upper floral bud on the s&me i?fl~~e8cence, althougb
only slightly aider, has an already well-developed upper median tep.l
and is jùst initiating the upper median stamen (Fig. 14). Observations
of la'ter stages suggest that two developmental patterns are possible.
(1) There may be a r.pid expansion in the upper regton of the bud, pro-
viding the needed space for upper stamen formation. (2) There may be
a failure of one or more organs ta develop in the upper portion of the -hud, resulting in a trlmerous whorl of stamens or completely trimerous
flovers. These deviations, whieh were very frequent in the material
Itudied, will be delcribed rh greater deta!l beloW.
lt should he noted here that the initiation and development of the
upper and lover median stamens. varies quite signifieantly from that of
the lateral ones. The median st~en usually does not arise as tvo
markedly sep.rate primordia but as one elongate primordium (Figa. 18.
19). Concurrent with the i?cep.t;lon of the upper median stmen" are the
firat signs of interprlmordlal upgrowth between the double prlmbrdia of
the lateral stamenw (Flg. 17). ...
Unlike the lateral tepals and Itmens whieh a~e inittated in rapid
o
sucee .. ion and grov at about the"s&me rate, the median tepals and stamens
differ,ln their t~e of inceptlon and grovth rate. Although the lower
medlan tepal and .tamen are inltiated long before the upper pne., their
rate of, growth in th •• arly sta.,s of de~lopment t, very slow'and they .
are hardly visibl. unt!l about the sta.e of Iynoèclal inception (Fig_ 20).
The upp.r medlan tepal, on the other hand. once lnltiated (deflnit.Iy
Mach later than rhe low.r me dl an t.pal) B~ow. rapldly and

•
-,
not only the lower median tepal but the already lar~ lateral tepals
• (Fig. 14).
36
Upgrowth between tepal and stamen occurs early in the development of
the lateral appendages, at about the same time interprimordial upgr~wth
.~ ls noted betwéen the two primordia of each lateral stamen (Fig. 14). In
the case of the median tepals an~ stamens, upgrowth between them le not
seen until the stage of gynoecial initiation (Fig. 20). In any case,
upgrowth between tepai and stamen in f. densus does not proceed very far
and even in the mature flover is not very pronounced (Fig. 44).
The development of the hypogynou8, apocarpou8 gynoecium, wlth four
carpele, each containing a single bitegmlc, anatropous ovule begins at ,
about the same time the staminal thecae start to differentlate (rigs. 19,
20). Four (three in the case of tr!merous bude, or trimerou8 gynoecia)
nearly radial gynoecial primordia are Initiated on the floral apex,
alter~ating with the stamens. A lon~itudinal section through a young
gynoeci&l primordium reveal. that initiation Is due primar!ly to perl-
clinal divisions in the seeond tunica layer accompanied by cellular expan·
aion and anticlinal divisionl ~ the firlt tunlca layer -(Fig. 51). The
lequence of inception varie.. In seme floral buda the gynoecial primordia
. aril. at about the 1 ... t~ (Pig. 20), while in otherl the lower two are
forw.d firlt (Plg, 22). In one particularly peculiàr bud one can see the
gyno.elal prt.ordia At thr •• diff.rent .t .... of development; the lover
~o are already at 'the It ... of ovule inception, the upper left primer-
dtum ia ju.t becoaina .. tt.te, and the upper rtght pr~rdium il Itill
radial (Ptg. 29). A pett.te prt.ordtua ia formed- by uP8rowth proce.ding ~
1 7
/

l"
36
from the abaxial portion of the gynoecial primordium, spreading around
the periphery to a portion of the floral apex (PtS. 25). Just after
the Iynoecial primordia have become completely pelt.te, the ovules are f
initlated at the adaxial portion of the young inner wal~ of the carpe1 ~
(Querzon~) (Figa. 26, 27). A longitudinal section through the ovule
shows that periclina1 divisions in the second.tunica layer are primarily
responsible for prÛDordial initiation (Fig. 52). The ovule develops
twq lnteguments through cellular expansion and division in the p~oto-
dermal layer, wi~h the inner integument forming first (Fig. 40). From
the beginning, the abaxial portion of the carpel wall grows At a more
rapld rate, partially enclosln~he young ovule (Fi~, 28). The ovule,
though, 18 not compleiely covered unt!l a relatlvèly late stage (Fig. 37),
and may still be seen at the stage wher.e the tepals and stamens are
reaching maturity (Fig. 34), Just after the carpellary tube hal enclosed
the ovule and begins to form the style, a primordium is initlated at the
upper abaxial portion of the carpel wall, just be10w the region. that later
becomes the Itigma (Figs. 38, 39). In the mature carpel th1s outgrowth ~
can in ~~e cases be very promin~nt "Fig• 41).'
The much diseussed .t~en connective develops in some cases to rather
outltandinl proportions, sometimes towering above the pollen sacs (Fig.
44). BefDre'anthe.i8 the tepall undergo a rapid expanaion, completely
encloslua the other floral appendages. lt is therefore nece.sary to ,
remove the tepals to ob.erva the ftnal stases of 8~lgma and style develop-
.. nt (rll" 41, 42, 43). At anth~.i. the tepals open and bend back
revealinl the tertil. flof~l orlans (Fig, 45) •
. ID addition to the Dor.al tetramerOU8 flow.ra many unu8ual variants
, ,

/ , .
1
t . . are al.110 observed. Lower floral buds tend in many case8 to be cam-
•
pletely trimerous, with no indications what.oever of Any rudi~entary
upper median tepal or stamen (Figs. 21, 25, 26). In other buds inter-
mediate outgrowth8 at the p08ition of the upper median tepal and sta-
'~en are seen. The lower bud of one partlcular inflorescence has at
this position only an elongate primordium (Fig •. 22, 23, 24). The
lateral 8tamens of this bud als6 exhibit strange growth patterns,
growing much mor~ rapidly in their lower (abaxial) regions. In another
floral budt the appendage (PmG) in the upper tepal - stamen region
could be interpreted either as a fifth carpel or as a deformed tepal
Àdjacent to what see~ a rudiment~ry stamen (U) (Fig8. 29, 30, 31).
Finally, in an instance of variation at the upper tepal and stamen posi-
tion the appendage formed 8eem8 80mewhat intermediate between a tepal
and 8tamen (Fig. 28).
Flowers vith tetramerOU8 perianth and androecium but only trimerouB
gynoecia a~e also observed (Fig. ~). Here the t~o,lower carpels appear
quite normal, alternating with the laterai and lover median stamens, but
in place of the two carpele in the upper portion only a single, larg~,
elongate car pel ie formed directly opp08ite the upper median stamen. Two 1
ovule8 develop within the enlarged carpeL. Failure of the upper left-hand
portion of one floral bud ta differentlate fully re.ults in a rudimentary
car pel and upgrovth betveen the theea of the left lateral and upper median
.t_ens, (Fig. 35).
fIocaœbl.1 Qevelop!!!t
In a cro ••• ection tbrOugh t~e inflore.cence axis a number of collateral

•
-
38
bundles are found, two being rather prominent. Procambial strands
differentiate from the.e two towards the lower (abaxtal) portion of
the young floral buds, a single strand entering each. After tepal
primordia are initiated on the floral bÙd, four procambial strands
d~velop acropetally towards the tepals from the single strand entering
the flower (Fig. 53). Stamen strands fOTm acropetally as weIl, dif
ferentiating from the tepal atrands towards\fhe stamen primordia, at ~ v
a stage after interprimordial upgroWth has begun between t~p.l and
st&men (Fig. 54). In the case of the léterai st*mens, each of which
arises as two separate primordia, a procambial strand differentiatè8
after interprimordial upgrowth between them ~8 occurred, and ln a posi-
tion between the original two primordla.
The gynoeci8l primordia begin to form procambial tissue late in
their development, at about the time of ovule inception. Here again
the strands develop acropetally, from the central strand in the pedicel
of the flover towards the young carpels. From the main carpellary strand
a branch differentiates ventrally into the ovule. The main strand itself
continues to develop within the dorsal portion of the growing carpel. ,
reaching up to the stigmatic region in the mature carpel. No procambial
strand differentiatel ln the outgrowth of the ~pper abaxial porttpn of ,
the carpel previou.ly de.cribed.
Flnally and rather UDulually. a aingle procamb~al strand differentiates
from the central procambial a.semblage in the pedlcel lnto the domed up·
growth of th. ceâtral floral bud reglon. (Pig. 55) •
}

t
3&
DISCUSSION
Although this' investigation of f. densu8 was intended to focus
prlmarily on the stamen connective - tepal ~ue8tion (see Croizat 1966),
the unusual inlti.~lon of the stamens proved an interesting and impor-
tant diveréion. The medlan stAmens arise 8S elongate primordia in a
fashlon slmilar to that of the stamens in Potamog~ton richardsonii.
However, each of the lateral stamens is initi_ted as two primordia
which ooly later become connected by tnterprimordia1 growth to form
• stamen of the .~e shape and structure as the median stameos, This
ob.ervation demon.trate. two phenomena: (1) equi-finality, which in _
de,criptive and comparative sense means that two identica1 strùctures
are formed by different developmental proces.es, in this case, from two
different starting points; (2) one organ which normal1y i8 formed from
one primordium arise., in the case of the lateral stamens, from two
primordla. The lnterpretation of this diprimordlal stamen inception ls
difflcult. Miki (1937) .peculated th.t,"the tvo-celled anther (of L
Potamoseton) may be considered &s two sessile atamens which indicated
probably a reduced flower on the stalk of the bract, " Perhaps the dipri-
mordia1 stamen inc.ption could support this view or rather the conclusion
that a .tamen of 1. den.us i8 partially homologous to two 8tamen8. In
other yord., the Itructure that 1. termed a stemen ha8 characteriatic.
of a .ln81e st..-n and a pair of stamenl. The properties that it shaTes
vith • pair of .t ... n ... y be demon.trated by a ea.parl.on with flovers
of Ali ... tale.. For .~le, ln Butoau. or All,.. one flnds a pair of
.t",n ~iaordi. luperpo •• d to tbe prt.ordiua oÏ • tepal or petaI, re.
pectivel, (Sattler 1973;'Slnah and Sattler ~; Lein. and St~ler 1972).
-
,

-, '101%' J r -~ .. "
40
The position of ~he pair of stamen primordia corresponds to the posl-
tion of the two prlmordia of the single stamen ln g. denaus. Hence,
there 18 homology wlth respect to position (homotopy). To transform the c: r'
pattern of Butomus or Al4sma into the pattern of Potamogeton densus one
would have to assùme interprlm6rdial growth, a reduction in the number
of pollen sacs, and changes in the vaecularization, which may be a neces-
sary consequence of the very early inception,of interprimordia1 growth.
Regarding the controversy concerning the outer floral appendages,
each of whlch has variously been Interpreted as "outgrowth of the stAmen
connective" (Irmisch 1851; Elch1er 1875j Aacherson 1889; Rendle 1930j
Markgraf 1936), &8 "perlanth segment" (Graves 1908; S1ngh 1965; and
others), as "perigon segment" (Hegelmaier 1870; and others), etc., there
was no evidence in the ontogeny of the flower of f. densus of Any out-
growth on the young stamen primordium. Development was for the most
part acropetal, in that the tepa1 prlmordium was initiated be~e any
indication of the stamen opposite it could be seen, The lower (abaxial)
, median tepal and stamen primordia, though, ~re initiated simultaneously
(Fig. 46)~ In Any case, tepal and stamen inceptlon ~a8 spatially sepa-
rated. Sectioning the materia1 (1n many cases the SAme buds that were
dislected and photographed whole) "supported thele findinss. This vas in
accord vith the floral organogenetlc atudie. done by Hegelmaier (1870)
on 1. crheu., Goebel (1911) on-f •. natans and 1. den.ua, and Sattler.(1965)
on!. richardlonii.
The orsan .y.tem vhich (for convenience) i8 termed a '!lower' in th!s
paper. ha. been hoao1og1ced vith an lnf1ore.cence (Kunth 1841; MIki 1937; '.
Ubl 1947) or a flover (H .. e181er 1870; Markaraf 1936; Sinlh 1965; and
, ---------------------~~-------

•
•
•
41
,1'
the majority of the botanists dealing with this genus). In this respect
the inception of the lower median (abaxial) tepal and its superposed
atamen ls interesting: it occurs concomitantly wlth, or possibly even
before, the Inception of the floral apex; hence, the incipient primqrdia
'of the abaxial tepal and stamen resemhle strikingly those of a bract and
it. axillary branch (Fig. 46). As the floral apex develop., the primordia
,of the abaxial tepal and stamen become part of the young floral bud and
the other floral appendages are initiated on this bud. One might inter-
pret these events as the spatial and temporal superimposition of a bract-
&xillary bud configuration and the development of a flower (which in turn
May he partially homologous to an inflorescence). In other words, organs
that are Inl~lated like bract and axillary reproductive bud become during
their development part of a flover and assume aIl the properties of tepals
and stameoa, respectively. Thls Interpretation seems to be in agreement
vith Emberger's (1950) thesis that recognized intermediates between
"inUorescence" and "~lower" and emphasized the occurrenoe of prefloral
organizations that have not as yet fully contracted into the organization
of a flower. However, vith regard to the contraction of inflorescences
into flovers, one might also con.ider the opposite trend, namely, an
expansion from a flover ta an inflorescence. <
It ha. been previausly mentioned that variation occurred vith greater
frequeney and in a greater variety than ean be termed normal. Variation
in floral .tructure ia nothing new, and in vater plants it seems e.peeially
high. Arber (1920) ha. noted that, "a eharaeter whieh increasei the difft-
eul~y of identifyins them (Potamogetonace •• ) i. the.capacity for variation
in form .hovn by one and th • .... individusl. If But in l. den.U8 at lust
•

l.
•
•
'. 507.. of the flowera observed ahoved some fom of deviation from the nor-
mal tetramerous pattern. A partlcu1ar1y lnterelting variation obaerved
val. blending or.merglng of two appendages, 1.e. tepai and stamen (Fig.
28) and tepa1 and carpe1 (Fig. 29).
A comparison of the development of g. densus and that of thé distantly
related species 1. richardsonii and f. crispus (Table 1) aptly demonstrates
the remoteness of f. densus, and supports to some extent the reasoning of
those who separated th18 spec!es as a monotypic genus, Groenland!a
(Fourreau 1869; Butcher 1961; A~lto 1970). However, tt should be noted
that 1. crlspu. 1. intermediate between 1. densus and 1. richardsonii.
The initiation of lateral stamens, amount of upgrowth between tepaI and
.tamen, and the lack of a bract subtending the floral bud. closely resemb1es
1. densus; yet, the sequence of tepal initiation and growth of!. crispus ;
i. identieal with 1. richardsonii. This distribution of attributes is in '~
agreement w1:th the separation of these three spectes into different sec·
tionl by Aleherlon and Gr.ebner (1907),

·,
TABLE 1
Developmental differences between P. de.sus, P. crispus, al
P. densus
Iwo floral buds initiated on the inflorescence axis, in opposite arrangement
No bracts subtending the floral buds
Early, almost simultaneous inceptlon of lower median (abaxial) tepal and stamen beIore or concomitant with initiation of floral apèx
• Eacn of the two lateral stamens initiated as two separate prllIIOrdia
Th~ upper median (adaxial) tepai primordium ls initiated late, after the lateral stamens, but growa tapidly
Very little upgrowth between tepaI8 and 8tamens
Extensive ter.1nal upgrowth of , It ... n connective
,. Extensive outarovth on upper abu1al portiOn of carpel vaU
P. crispus (Hegelmaier,1870)
S~x floral buds initiated .on the inflorescence axis, in spiral arrangement
Same" as P. densus
Lateral tepala initiated first on promil
Origin of la,teral stamen not as clearly diprimordial as in P. densus
The two median tepal primordia are init after the lateral tepal primordia and . lateral 8tamen primordia
)
Some upgrowth between t~pals and 8t.~.n. but not significantly more than in P. densus e
None
None
A lar,e, l1nale unbranched v •• cular Itra04 develop. iD the .t ... ~ eonnaè t ive'
, . .
/

-..
TABLE 1
, . ta1 differences betwee~.!:. densu,s, P. crispus, and p. dchardsonii
. P. crispus (Hegelmaier,1870)
Six floral buds initiated on the inflorescence axis, in spiral arrangement
Same as P. densus
P. richardsonii (Sattler,1965)
Floral buds (9-27) ~nitiated on the inflorescence axis, in whorls of
cthree
Bracts subte~ding floral buds .1'
Lateral tepals initiated firat on prominent floral apex
Origin of 1ateral stamen not as clearly diptimordial as in ~. deoBus -
AlI four .tamens initiated as sinSle, elongate primordia
The two median tepal primordia are initiated almo8t simultaneou.ly juat after the Iateral tepal primordia and about the same time as the lateral 8tamen primordia
.', Some upsrowth between tepals and
stamens but not lignifieantly more than in P. denlus
None
None
[rAll4 developi in ·th. ltan
"'. ,.
Tepals carrled ~igh up on stamen connectives
None
Sl1,bt outarowth on uppér abax~l' carpel wall
~ Tb. ItaaiD6l buadl* birfureatel th. lOVer portiOn of tb. CODQIcUve
. .. .&
•
43

• •
-
, ,~.
ACtc:NM.E OOEMENT
... r ,
This investigation va. lupported by a Postgraduate Scholarship
~ 1560 '(to U. P.) and by an operating grant A2594 (to R. S.) from the
Natiànal Research Couneil of Canada.
, 1
•
• •
44
•
:, ~'
~ ,.~
.h~ .~
r ,,' • '" .;,
,' .
t·;

\
~ LIST OF ABBREVlATIONS
o
Al - lateral stamen or tts primordia.
~ • median stamen or lts primordlum.
Ac - stcmen connective.
F • floral apex.
FI - lower floral apex or bud.
Fu • upper floral apex or bud.
G • carpel or lts prlmordium.
Ii • inner tntegument or ttl primordium.
10 • outer int.sument or ltl prlmordlum.
L • vegetative appendage or tts primordium.
N • nucelluB.
o • ovule or lt. prtmordium.
Pl • lateral tepal or itl primordlum.
Pm • median tepal or lt, prlmordlum.
r • remov.d, e.g, rT • theca removed.
SI • atlgma or developins Iti ....
8y • Ityle or developing .tyle.
Syo • outgrowth fram Ityle.
T • thec. or it, pri.or41um.
U • undiffèrentiated .'pend .....
v • .... t.t! •• apex (ren..al arowth).
-

~~~.--~~------~--------~~-.--------~--~--................ ...
46,
-Figs, 1 and 2
"
\ 'Fig. 1. Diagrammatic representat1Qn of the pattern of Infloréscences,
Fig. 2.
from s1dè (a) and top (b) view.
Floral diagram.
•
" --,
,
•
•
..
o
-• 1:' ., ""J -, , 11-,
,
\ ." , .
, , ' .
• ~
1 1 t
f ' 1 . , a , .
"
L L \
,
~(® ... ~0 ~ ~~ ... ®j~ l
6
b
, 1 :
'"

47 1
PLATE l Figs. '3-11
Fig. 3. Side'view of vegetative apical meristem (V) producing leaf
primordia in a distichoua arrangement. x 146.
Fig. 4. ,Top vi~ of floral bud (F) Just being initiated. L ·'leaf
primordium. x 146. i
Fig. 5. Side vlew of inflorescence axis showing upper floral bud ('u)
and lower floral bud (FI) a. we~l ~a the lateral vegetative
apex (V). being initiated in the axil of the last vegetative
appendage (L). x 146.
1
Figs. 6 and 7" Side and top views at a s1ightly older stage. Roth are
views of a aingle inflorescence. x 146. 1
Fig8. 8 and 9f Top views •• hovins the lover medlan (abaxial) tepal
(Pm> and both lateral tepals (Pl) just at inception on the ,.
upper floral bu~ in Flg. 8, while the lower floral bud in Fig. 9
on the .ame inflorescence axis shows the fir.t indications of
lateral stamen primordia (Al). x 146.
,F1g •• 10 and 11. 'ToP view and oblique view of two ,floral buds on the
same infloreacence axis at 81mllar .tages aa thoae of Flga. 8
and 9 but at • gre.t~r __ Inifieatlon. x 246 •
. ,
, \
t

-
•
•
li
•
e. ,

48
-
PLATE II Figl. 12-20
, Fig. 12. Top view of the lover floral bud of an inflorescence, shoving
the lower median (abaxial)"tepal (Pm> and stamen (Am> just
becoming distinguishable at this stage. x 246.
Fig. 13. Side view of the upper floral bud of the samè inflorescence as (
in Fig. 12, showing the loWer primordium of the lateral stsmen
'(Al). x 246.
Figs.14-l7. A single inflorescence from different views
f and different magnifieations.
Fi8s. 14 and 15. Top view8 in whtch the upper floral bud in Fig. 14 has
already initiated the upper median (adaxial) tepal (Pm> and
stamen (Am>, while the lower floral bud in Fig. 15 has only
ihitiated the upper median tepal. The two separat~ primordia
of each of the lateral stamens are clearly visible (Al)' x 146.
Figl. 16 and 17. Side views (slightly oblique) shoving higher magniflca-
tlons of the lame budl, foeusing on the lataral stamens. x 246.
Flg. 17. Interpr,imordla1 upgrovth juat beginning. x 246.
FiSI. 18-20. Top vievi of flora~ buds.
Flg. 18. Floral bud .hoving elonsate form of the meaian stamen pr~rdia.
x 246.
Flg. 19." Floral bud .hovlns lnterprlmordial growth betveen the ~wo'prl
mordia of each lataral .t ... n. x.246.
Flg. -20. Floral bud .howinl ineeptlon of STAoeclal prlmordla (G).
.ote: dark .pot. on the, bud are due to exce.' uptake of .tain
by .OIN caU •. x lit6.
.1
f .. ' i ,1
~
0 --.;:: ~ 1 ,

-
,)
" ,
,
.e

49
0
, ~ ,
-,.F" .... '~,
PLATE III Figs. 21-28
Fig. 21. Top view of a trimerous floral bud. x 146. /
Figs. 22-24. Buds of one inflorescence fram diff~rent s.
Fig; 22. Top vi.. of lower fl~ra:,;:: showing an unusua1 append '1
at the uppe~ median (ad al) tepa1, atamen region. ;146.
Fig. 23: Side view of .... e but Fig. 22. x 146. / -!uda. Fig. 24. Top vlew of lnfloresc nce8 tip (1\) with two floral x l~6.
, Figa. 25-28. Top viewa of 'floral buda. ,
-Fig. 25. Trimerous floral bud in which the gynoeclal primordia (G)
have become péltate. x 146,
Fig. 26. A trimerous floral bud at a slightly ,older stage. initiatlng
ovules at adaxla1 portlon of carpel valle x 146.
Fig. 27. Tetrameroua floral bud at a simi1ar stage ae that of Fig. 26.
x 146.
Fig. 28. A floral ~ud vith a trimerous gynoeclum but tetramerous perlanth
and androecium. The upper medlan appendage. though. 8e8m8
partia11y tepa1-l1ke, partially 8tamen-1ike (Pm Am). x 146 • •
... . \,
, . ....
...

" ' .
• ~f.:ç,~'fk' -è""f!l\t •• ;"$d_I •• ~ _
....
L . a
1
"
"
_ h

&0
PLATE IV Figl. 29- 33
ngs. 29-32.
" 1
A single inflorescence ,viewed in different ways.
Fig. 29. Top view of the lower floral bud in which the gynoecial prl-
mordia are at different stages of development. The upper
median (adaxial) appendage (Pm G) seems partia11y, carpel and
partially tepal. tt aubtènds an undifferentiated primordium
(U). x 146.
Fig. 30. Top view (sHghly oblique) looking down on inflorescence tip,
• showing the unusual upper m~dlan appendage: x 146.
Fig. 31. A median longitudinal section through the saUle inflore8cene'li!.
~ 233.
Fig. 32. Top view of the tetramerOU8 upper floral bud on this tnfldres-
cence. x 146.
Fig. 33. Top vi'ew of a Horal bud Ihowlng upgrowth of the carpe1 wall
and exten,ive growth of the stamen connective (Ac). x 146.
4
. , ,
,\

-
t
"
< . .. ":"
, 1
"

-.. y , ~
&1
PLATE V Flg.. 34~38
~ Flgs. 34"38. Top vins of floral bud •.
,~g. 34. carpel wall beglnnlng to grow up and about the deve10ping
ovule. x 146 •.
Ptg. 35. Fatlure of complete dtfferentt.tton in the upper 1eft-hand \
rtg. 36.
portion of thls-bud resu1ttng in a rudi~ntary earpe1 and
interprtmordta1 upgrowth between tvo prtmord~ of adjacent
(latera1 and Median) ItaDen •• x 146. \
Elongate upper (adaxial) 'carpe1 eontainlng tvo ovule prt-
mordt.. ,146.
Ptg. 37. 11 ene10.ing ovule. x 146.
F~a. 38. Style bel~nning to differentiate ~d an outgrowth at the
abaxi.1 p~rtlon of the Ityle (Syo) 1e dev.loping. x 85. 1
, i'
1 1
'. ,
."
, .'
, :
/
/
'.
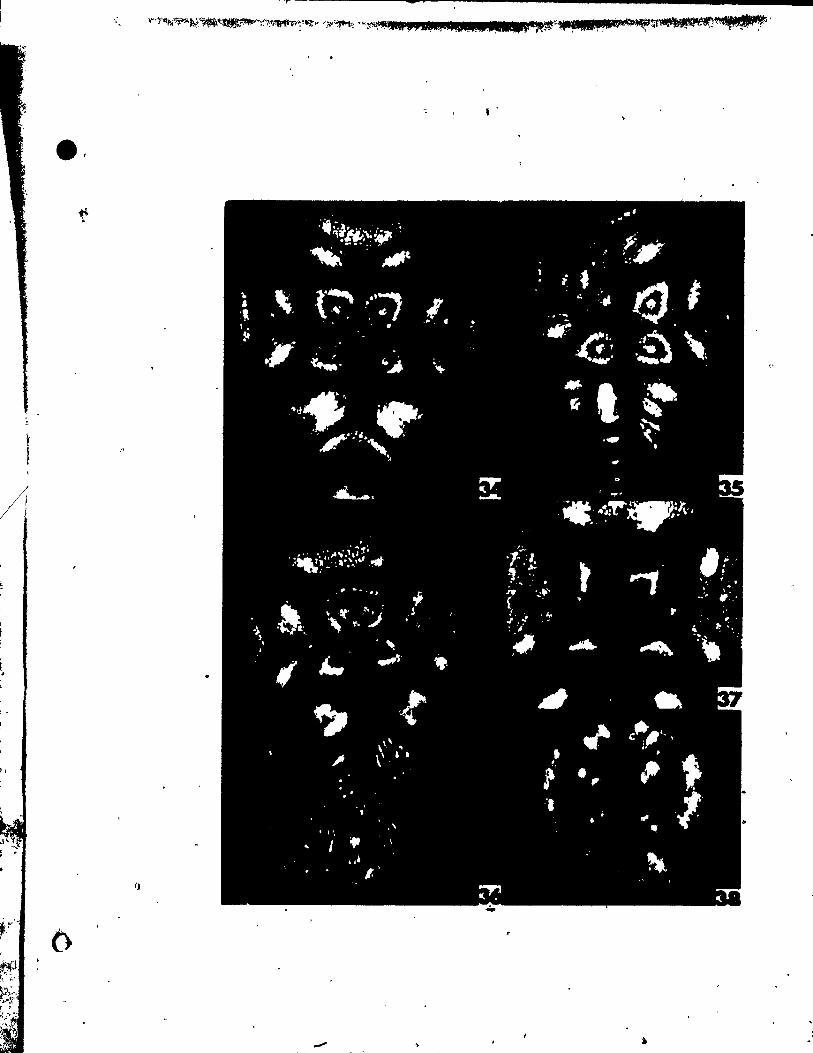
...-'
t .
i.
'1
IJ
..- ·1

PLATE VI Figl. 39-45
Fll. 39. Slda view of .t1g'111& (Si) beglnnlng ta devel'op on tbe upper
portion' of the 8tyle (Sy) iDote stylar outgrowth (Syo>. x 146.
Fil. 40: Top vi~ of dll1ected carpel show4ng lnner (Il) and outer
(la) integuments growlng about the nucellu8 (N) of the
young ovule. x 146.
Fil. 41. Sida view of carpel .hoving extensive stylar outgrowth (Syo)
and well developed .ti..- (S1). x 85.
Fig. 42. Top view of two Itigmaa. x 146.
Fig. 43. Adaxial vlew of carpel •• howlng upgrowth tram baie ta stigma.
x 146.
Fig. 44. Nearly mature stamen in which o~e of the thecae (rT) and the
tepal Crp) were removed to show the extensive Irowth of the
.taminal connective (Ac). x 85.
Fil. 45. Open mature flower. x 16.
\
o " 1

~ r.
.'
(
45. -

-l • Jo.. _ 'C. '
Figs. 46-50
Flg. 46. Longitudinal .ection through young inflorescence axil at
about the same stage shawn in Fig.~. Note the 'f1rlt
indications ofmedian tepal (Pm> and median Itamen (~)
iniUat.ion. x 480.
Fig. 47. The same inflorescence .. in Fig. S, eut tranlverlely to
the inflorelcence axil, showing the first .ign~ of laterai
tepai (Pl) formation. x 516.
Flg. 48. ,-The same bud as that of Fig. 9, eut transversely to the
inflorelcence axi., .hoving lateral 'tepal primordia. x 480.
Pia. 49. Slightly oblique crol. section of floral.bud shoving
lateral stamen (Al) ln~ti&tion. Axillary vegetative axis
(V), Just below floral bud, vas eut obliquely. x 480 • ..
li'ig .• 50. The same floral bud as shawn whole in Figs. 22-24, sectloned
transversely to the inflorelcenee axil, .howing the devel-
oping lateral Itamen prt.ordia (Al). ~ 480. \
.! J ·t~
.'
• . .
r .....
. "
. ~ ,

i ~ . • f
( .~
•
-
..
. .
\
1

Fig •. 51-55 ~ Fig. 51. Longitudinal section through floral bud ahowing initiation
of gynoeciaJ primordium (G). x 800.
Fig. 52. Median longitudinal section of the floral bud of Fig. 28,
showing the developing ovule (0) in the single upper median . ,. .. 1
carpel and the Appendage with partial .tamen ~d partial
tepal characteristlc8 (Pm Am). x 480. \
\ \
Figs. 53-55. A series of cross sections through ~learly
mature buds, revealing the pattern of vas
eular strands. Within t~ne8 d'eUmttiing
th~ vascular Itrands, concentrations of 'li'
xylem ve.aels have been outlined.
Fig. 53. Vascular strands entering the tepals, and the .tamen strand.
ju~t beginning to branch off. x 32.
Flg. 54. V .. cular .trand. ln the atamen connective r.,ion and the
beglnning. of brancblng of the central buDdle,tovardl the
car.,.lI. x 32. Ji '
Plg. 55. Strand. ent.rl~ th. carpel. and a 1~11 central strand ln the'
ranat of tbe f1~al :.x'- x 32.
......
, '
• 1.
·t
. '
, .'
fr.; , ,
,~,
') ~
~
•

•
f ,/
.'
, "
.,''1. .. ' .1,. .. .,. '1' " . "
53
i'

t
l'
o
LITERATURE CtTED
Aalto, M. 1970. Potamogetonaceae. Fruits 1. Recent and subfo8ei~
endocarpe of fenn08candian species. Act Bot. Fenn. ~:1-25.
Arber. A. 1920. Water plants. University Pre~l, Cambridge.
Âscherson, P. 1889. Pbtamogeton.ceae. ln A. Eng1er and K. Prantl:
Ole natUrlichen Pf1anzenfami1ian. Il. Teil. W. Engelmann Verlag,
Leipdg.
A.cher'lon, P. t and P. Graebner. 1907. Potamogetonaceae. ln Das
pflanzenreich. IV. Wilhelm Engelmann. Leipzig. pp. 1-184.
Butcher, R. W. 1961. Â new i11uatrated British f10ra. Part Il.
Leonard Hill L1mited, 'England.
Croisat, \L. 1966. Observations on the ovary of the Jug1andac.a •.
Southw. Nat. 1!:72-117.
E~~ Â. W. 1815. BluthendlqT_e. 1". TeU. W. Eng'elmann VeTiag t
Leipda. "
Z.b.r,al', L. 1950. La val.~ ~pbolos1qu. et l'origine de la 'fleur.
(A propo. d'une th'ol'ie nouvelle). Ann6e Bl01. (Paria), ~:279-296.
Poutt.au, P. J. 1869. Catalolue des plante. du cour. du Rhane. Ann.
Soc. Linn. Lyon, Sel'. 2, !1:l69.
COebel.~. 1911. Morpholo81scbe und blolOlllche Bael'mngeft. 19. t'ber,
",.paute" Bl.~u.nl .. eQ. Flora,' 103:248-256.

•
, Graves, A. H. 190a. The mor~hology of RUppla marltlma. Trana. Conn.
i
Ac.d. Arta S~l. l!:59-170. "l,l'
Hegelmaler, F. 1870. Ober die ~ntwlcklung der BIUthentheile von Pota
mogeton. Bot. Ztg. (Berlin), ]1:283-320.
Irmlsch. T. 1~51. "Ueber die Inflore.cenzen der deutschen Potameen.
Flora, !=81-93.
Kun th , C. S. 1841. Enumeratio plant.rum. Vol. 3. J. G. Cotta~ Stuttgart •
and Tubingen.
Lein., P., and P. Stadler. 1972. Entwlcklun8Ig •• ehi~htllche Unter.uehungen ~
am AndrQeceum der Aliimatal.e. ~.terr. Bot. Z. ln pre ••. #
Markgraf. F.. 1936. B1Utenbau un~ Verwand.chaft bel den einfach.ten
ReIob!ae. Ber. Dtlch. B9t. Ge •. ~:19I-229.
Miki. S. 1937. 'l1le crilin of Najas and po ..... ton·.
.li: 4 72-180 ~
Bot. MaS. (Tokyo), 1
aendle, A. B. 1930. Th. clal.iticatlon of flow.ring plantl. Vol. 1.
U~iv. Pr •• ~, C.-brid.e.
Sattler" l. 1965. Per1antb development of PO~leton rlcb!rd,onll.
. J. Bot. 3,(1):35-41.
1968. A ~.chDlque for the It"dy ot flO1.'al deveto_nt.. Cara. .. .
J. Bot. !!§.17Zo-722. ' .
"
, ,',
, . , "
.. . ,

'\ , '
17
Sattlu, R. 1973, Organogenesi8 of f1ower.. A photographie text-atlas. ,
University of Toronto Pre" t Toronto.
Shàrman, ,lB. C. 1943. Tannic acid and iron a1U11l with .afranln and
orange G in .tudies of the shoot apex. Stain T.chnol. ~~105-lll.
S1ngh, V. 1965. Morphologieal and ahat~ieal .tudies in He1o~ ~
Va.eular an.tomy of the flower of Potamogetonaeeae. Bot; Gaz.
ll,!(2) :137-144.
81ngb, V'., and R. 8att1er. 1972. Floral development of 'AUIU triviale:
• Cano J. Bot. 50:619-627.
Uhl, N. W. \947', Studies in the floral morphology and anatomy of eer-
tain members of the Helobiae, Ph.D. Theli., Corne11 University,
Ithac;a, N.Y.
Il'
o
• •
. .'

• '.
/ /. ,
.... '
."
CHAPTER :3
FLœA'L ŒVELOPMENT OF RUPPIA MARITIKA VAR. KAllITlMA
'.
,.
",
.. <"
r ~ 'tir ,: ~,l ' .' ,
, .
/ /
"
,
.e",
. . . -' ,~'.
' .
1 j
~ 1

'\
•
=.
ABSTRACT
A hyaline. unvasculari.ed sheath envelops a portion of the inflores-• cence near maturity. Though resembling an appehdage of the main axis. in
early ontogeny it develop. as a prophyll of the renewal growth apex below
1 Two flowera ~evelop on the inflorescence axi8, sub-
opposite e.ch other. Fertile appendages are initiated in an acropetal
sequence on each floral bud. '" The firat to form, in the meoian position.
are th~ two stamen. t the lower preeed,ing the upper. Rach stamen deveto'ps
" two bisporangiate thecae separated by a broad connective. A dorsiventral
j outgrowth ia initiated slightly abaxially near the tip of the connective
at the stage ,of theca differentiation. This outgrowth appears to be homo-
IOIOU. vith a aimilar outgrovth in Potamogeton den.u., but not with the
aterile appendage. of the Pot8mOseton flover which, by some authors. have
incorrectly been interpreted a~ connective outgrowths. Rach carpel arise.
a. a radial primordium which become. peltate aft*r it. i~ception. One
o~le ie Initiated at the adaxlal ~ortlon (Querzone). The,.tigma become.
broad and fIat, lobina at it. margina. A ~118ht outgrovth develop. at the
.' -abaxial wall of the carpel. The floral apex ha •• tvo-layerad tun~. The
primordia of the .tamen., carpel., an~ ovul •• a~iae by p.riclinal divi.ions
ln the .econd layer.
prt.ordial inc.ption.
Proc.ablal development ,_ acropetal followlng clo.ely
Bach appendage 1 inclJina the ow1e. recelve. O'(le (1
1
proca.bial atrand. Tbe out~owth. of th. ponnective and the carpel lack
proca.bium • l'
( l ,
/ ,/
/

t 1
'.
-
60
INTRO meTION
There has been much discussion ln the past on the nature of the
flower of ~ppia and the small dorsiventral outgrowth on lts stamen
connective (see, for example, Markgraf 1936; Uh1 1947; Singh 1965; and
G~erro 1968), It has been generally concluded that the flower of
Ruepia is equivalent to the flower of Potamogeton and, specifically,
that the smali outgrowths of the connectives in Ruppia are homologous
vith the sterile appendages of the Potamogeton flower that have been
shawn to be sterile phyllom~8 (Sattler 1965: Posluszny and Sattler 1973.
1974). These conclusions have been based on mature or well-deyeloped
flowers, 8ince no developmental data have been àvailable for the flover
of Rupp!a. The purpose of this ~ork 1s ta prov1de a detailed account of
the floral development of Ruppia and to use this nev information for the
evaluation of homologies and relationships with Potamoaeton. i
MATEIHALS AN D METHOœ ~
The .arly to near-mature floral stages of RHPeia maritim& L. var.
m&f1ttea vere collected in June, 1972, at a brackish water inlet near
Buctouche, New Brunlwick. Tbe fruitins .tages vere collected near the
8 ... locality in'$ept .. ber, 1973, by Dr. Philippe Forest. The material
va. t.bed and prelerveln PM (forMUn-aceÙc acld-alcohol) before being
stained ln alcoholic &cId tucblln, 41 •• ected, and photogra~h.d entirely
1IIiMr.ect ln 100\ et..,l alcobol follow1", the teclm1qua of Sattlu (196.8).
Alter belai pbototrAPbed -'ole. ~tatD floral ~dl v.~. then dehy-,
drated lD a t~t1er1 '-tJ1 a1~1 ..ri •• .ad ••••• "d, la tl •• uemat u.la,
.t ........ teclma~.. IMC~ ... ne _ • IIler_CIrI (~) ad vere
, ,
" ,

•
'" \
61
stalned with DelafIeld t.a hematoxyl1n. Cleared specimens vere prepared
uaiA8 the technique of Rodin and Davis (1967).
OBSERVATIONS • . . 1
Organography
Once Ruppia marltima becomes reproductive, the inflorescence terminates
the main axis and vegetative growth continues for a short tiœe in the ax~
of the foliage leaves beneath the inflorescence. These renewal shoots in
turn terminate aS,inflorescences, resultln& in a sympodial pattern of
growth (Fig. 32 a,b).
The inflorescence consists of two flowers subopposite each other. To
d1stinguish between the two, ,one will be referred to as the upper floral
bud (Fu) and the other a8 the lower floral bud (FI)' Each flower has two
median 8eaal1e stemena. The two blsporangiate thecae of each stamen are
separaud by a broad connectbe with a dorsiventral outgrowt:h. The gynoe
ciua conaiats of three ta five (u8ually four) paltât. carpe1s each of which
bears a single, pendulou8, bitegmle ovule (Fig. 33). The fruit i8 an asyur
metric drupe on a long, 8lender .talk or podoJYnA which develops after
fertUhation.
,~
Otaanovnut.
A. the vegetative apex b.~. Teproductive, lt enlargea and initiates
bath the upp.r (Pu) and 1~ (Pl) floral buda .ubop,..lte each other on the
inllor •• canc. ail (rta. If:" At about the ... u. .. , vegetaUve renaval
growth .,le •• are tDittated iD th. axll. of 'foll... leave. immediately below
tu Inllore'ceDee (ri, •. 1,2).' The ran..,al arowth apex belov tbe UP9&r
floral bud ,. of ,articuler inter •• t. Adaxlal1y, lt ,ive. ri •• ta a thin, , ~ "\

-
« hyaline, sheathing prophyll (Figs. 4- 8)). The base of this prophy11
1 1
extend. to some extent anto the main .xia (Figs. 6, Il) and thus ~ 1
gives the impression of be10nging to"' the main axis, especially in IJ
later de.~lopmental stages and at maturity (Fig. 25). The tip of
the prophyll becomes lobed (Figs. 7, 25).
The firat floral organ to form ls the lower atamen. It arises
as an e.longate prlmordium on the 'lover periphery bf the sornewhat
fIat floral apex (Figa. 3. 4), thus producing a allghtly two-lobed
atarnen primordium. The upper atarnen is initlated next on the floral
apex, ln a very simi~ar manner as the lower stamen (Fig. 7). It
growa rapid1y and at the stage of theca differentiation ia already
1arger and more developed than' the loWer stamen primordium (Fig. 9),
The young stamens form a broad, slightly curved, connective between
the developing thecae and a1most encircle the floral apex (Figa. 10,
82
Il). It la toward the eb~ial portion of thia large atamen connective
1 very claie ta its tip thet an outgrowth ia initiated at the atage
ahortly after gynoecial inceptlon (Figs, 12, 13). This outgrowth aril"
firlt on the more rap!dly developing upper stàmen of bath t10ral huda
on the infloretcence axia (Fig. 13); and growa rapidly, at fint as a
radial primordlum (Fig. 14) and then becoming doraiventral (Figa, 16,
20, 21), It doe. not develop to any.grest extent, and remain. in con-
spicuoua betveen the tvo thecae of the Itamen even at a atage nearing ...
maturlty (Pig. 27). The tvo thecae of each Itamen become bilporangiate
at abaut the .taae of ovule inception (Figa, 15, 16), Brenching of
therconn'ctive re.ult. in a very short atalk beneeth eech theca
(PiS. 20). /
1
As the stamen. ara formins thalr thec&a, two carpel primordia are
•

•
• •
83
initiated simultaneously on the transversal flanks of the floral apex
(Fig. 9)~ After a short pla8tochron, two median carpel primordia are
initiated at right angles to the initial two carpel primordia (Fig. 10).
Each carpel primordium i8 nearly radial at firat, and then through more
rapid upgrowth about its periphery becomes peltate (Figs. 10, 15).
One ovule is i~itiated at the adaxial w$ll (Querzone) of the peltate
primordium, first in the two transversal carpels, then in the two
median ones (Figs. 15, 16). !Wo integuments are formed on the ovule
(Figs. 24, 30). The more rapidly growing abaxial carpel wall envelops
the developing ovule primord!um, leaving only a small slit on top
(Fig. 19). About this 8l1t the stigma differentiates as a fIat pel-
tate structure (Figs. 20, 22, 23). lt becomes very broad with lobed
marg{ns near maturity (Figs. 26, 27). Each carpel forms an outgrowth
on its upper abaxial (dorsal) side at about the éame time as the stigma
begtns to differentiate (Figs. 20, 22, 23). After ferti1ization the
.hort stalk below each carpe! extends to form a tlender but str.ight
podogyn& (Fig. 31).
Some variation va. noted in the development of the flower of
Ruppla maritlma. . ,r
Although .o.t flowera formed tetramerou. gynoecia, , ,,'----
.ame vith trlmerou. (Fig. 23) and p~ntameroU8 (Flg, 21) gynoecla vere
obl.rved. The number of stamens va. a1ways tvo. Abortion of a .tamen
theca occurred ln one flover (Fig. 23), vhlle ln another, the stamen
connective had aeemingly bifurcated_produclng tvo connective outgrowths
(Flg. 20).
llittosenel1s '.'
The floral apex ha •• two--layered tun1cA. Th- .tamen pri1llOt'dt'a are
a 'I.~"'" 1t".
il t
'0: ~( .{
, , =

~
•
-
84
initiated through periclinal divisions in the second tuniea layer
accompanied by cellular expansion and increased anticlinal divisions of
the outer layer (Figs. 5, 8). The outgrowth of the éonnective re8ults
from an increase of cellular activity abaxially near the tip of the con· ,
nective. This involves. periclina1 divisions and expansion of 8ubpro-
todermal cells Along with increased anticlinal divisions of tne p~to-
dermal cel1s in that region (Fig. 14). The ovUles are formed through
periclinal divisions of the subprotodermal cells of the adaxial wall of
the developing carpe18 (Fig. 17).
Conc~7rent w\th stamen inception i9 the initiation of the sheathing ,/
prophyll by perlclinal divisions ln the first tunica layer of the
ren~al growth apex (Fig. 5). The prophyll consists of only two cel1
layers with cells that rapidly become large and vacuolated '(Figs. 14,
17, 18).
fTocambial Development
Soon after the inceptlon of stamen primordia, proc~bial development
besins in the infloreacence axis in the form of one strand (Fig. 8). As
thia strand entera the ba.e of the upper floral bUd, two staminal strande
diverge from it (Flg. 8). Tke atrand leading to the upper stamen pri- 1
mo~dlum il almo.t a .tr~lght continuation of the atrand entering the
floral bud (Figs. 8. 17, 28). In the case of t~ lover floral bud, pro
c.abial strand. develop toward. bath the upper and lover stamen prl~dla
fram the Inflpr •• cenc •• trand (Pl' •. 8. 18), ~ the atamen. mature. ~
tb •• e .trand. biturc.t. in the upper portion of tbe connective (Fig. 29),
t~U' 8upplytna tb. tvo It-.1nal thecae • N
At tbe .t .. _ ~ the carpel prt.ordla beca.e ,elt.te, one proc .. bi.l

a
(\
s
J ,
strand beglns to dlfferentlate towards each of them froro the point of
divergence of the staminal strands (Figs. 18, 28). The deve10ping
ovule ptimordium receives a procambial strand trom the main strand in
the dorsal portion of the carpe1 (Figs. 28, 30r. .
No procambial development could be detected in the sheathlng
66
prophyll, the outgrowth of the stamen connective, and the outgrowth of
the carpel.
DISCUSSION .
The development of the sheathing prophyl1 provides an excellent ,
~ example of the advantages on ontogenetic studies. At matu~ity lt consists
di only two cell layers without v&scularization and appears to belong to
the inflorescence axis, enclosing its upper flover. This has led sorne
workers to describe it as a tspathe t (Graves 1908). However, the early J'
stagés ob8erved fn this investigation indicate that it 18 ~nitiated as a
prophyll on the adaxial f1ank df the renewa1 growth apex immediately
below the upper floral bud ... · ouring Hs ~evelopment 1t appears to be
"captured" by the rapidly developihg inflorescence axis, and eventually ,
envelops the upper flover.
Most workers today cons!der Ruppia and Potamogeton to be closely
related ln many respects, with the most strlkins difference being in the
number of stamen.. potamoleton has four stamens per floral unit, while
Rupp1a has only two. .Here again r ~~~lopmental ob.ervations provlde
~ew indghta. A compariaon betweenAuppia marltlma and two apecies of 1
fOtamoseton, who.e floral development was lnv.,tlgated (l. dén.U8 and
1. r~ch.rd'fonii, po.lu'ImY and Sattler 1973, 1974), reveala that at fhe . . . , ._+ .tage of .tamen inc.ptlon, Ruppla maritlme ha. a ~loral apex on1y about
i
, .. '
, , -<
'II

•
1 66
half as large as potamogeton even though the stamen prlmordia are the
aame size. Thus, no space ls availaole for the ~nception of two
transversal stamens as in Potamogeton.
Similarities in stamen initiation and development between Ruppi&
and Potamogeton ar~ evident. As in Potamoseton richardaonii (aee
Sattler 1965, Fig. 12; Posluazny and Sattler 1974), inrRuepia maritima
each stamen h lnitlated as a remarkably elongate primordiUID whose
lateral portions (that will form the thecae) grow more rapidly, thua
ptoducing a two-Iobed atamen primordium. ln the transversal stam~n8
of Potamogeton den sus the two later.l portions (that will fOTm the
thecae) are initiated as separate primordia. Then, interprimordial
groYth leada to the formation of one stamen pri~ordium that has two
lobes as in Potamogeton richardaonii. What appear to be fundamental
• differences in the mode of stamen inceptlon can be understood aa ver,
alight differences ln the timing of the onset of growth ib varioua
regions of the stamen primordiumj or possibly the differences are ,only
due to different growth rates in various regions of the stamen p~imor-
dium. Although Ruee!a marltlma appears to folloy the pattern of ;
"
Potamogeton riehard.onli, lt tends to appro.ch the unuaual pattern of
pqtamo.eton den.u.. Sine. differences in amount of growth ar~ 10
.inute, ve eanDot exelude vlt~ eertalnty th. posalbility tha! at least
th. lov.r at ... u of lu"!. !!littpa approxtmat~. the patte1 of the
tran.Vft.al .t_n. of Pot_a.ton den." •.
'l'b. interpr.tatloD of the outarowth of tJte .t .. n cJnecth'. ha. 1
b •• n very alch debat.d. An .xe.n,nt .~ of tb. e rov.r.1e. can
be found in GMerro '(1968). _nt, no
.. eUr whather th. outll'owth ... cOtl.llderad _ "e.U "p.rf.anth
\
1

•
' ..
- 67
segment." or & "bract, It it was taken for granted that it h totaUy
homologous with the appendage on the connective of potCD08etôn. , At J
~turity, one may indeed reach this conclusion quite ea.tly. However.
developmental studies reveal • mOlt remarkable difference between .
Poumogeton and Ruppia. In Potamogeton the appendages are inUiated
acropetaUy on the floral apex and develop like typical phyllomes •
(Poaluszny and Sattler 1973. 1974). In contTast, the Telults of this
work show clearly that in Ruppia maritima the connective outgrowth
develops, unlike elther a tepal or a bTact, after stamen inception.
in fact after the beginning of theca differentiation. It results
fTom growth veTy.sllghtly abaxial to the tip of the stamen connective.
Althoulh it bec~e. dorliventra1, it differentiates no pTocambia1
ti •• ue and consequent1y lackl Any vasculaTizatlon at maturity. In .
geneTal, it ia ai~ilar to a vertical extension of the stamen connec-
tive in Potamogeton denlul (Pol1uszny and Sattler 1973). and may be • conlidered homologous vith uhe latter.
~ ,
CaTpel development 1. limilaT in Ruppia and Potameseton. In both
g.neTa. the cArpel primoTdlum il Initiated a. a protubeTance tbat
approach.1 radial aymmetry. It loon becomes p~ltate and forme one
ovule at the crOl1 aone. In later stagea an outsrowth arise. at the
upper abaxlal portion of the carpel wall.
Many who have .. de cOllP&~ilon. of vesetatl VI appendagel. mature
flover., _d ••• d. (Marltaraf 1936: tJhl 1947; Sln8h 1965 j Gamerro 1968)
c1alll tbat 1»,,1a il elo •• t, l'elated to" Pot.,..tOD. Doe. floral
cle",CopMnt .uPport luch a poatulat.? It do •• not ctmtradict it. but
it .c:ert&inly do .. DOt brlDa th. tvo •• nera cloler ~olether. 'Th.ra are
..

1
•
88
.. many similaritt.s, for example, stamen and carpel development; but
there are also considerable differences such as a lack of homologues
of .terile phyllomes in Rupp!a and a difference in the number of
stamens. The floral develop..nt of !. mari~~. thoulh. doe. fit into a . . ...
trend of simplification and reduction between Potamogeton and
Zannichell1 •• as nb~ed by Singh (1965~. According,to this trend, reduc-
tion OCCUTS in the number of· ,ppendA8e~ per flower, flower. per inflores-
cenee, and vegetative appendages on the renewal shoots before floral
induction. Whether the trend goe. indeed from complex ta reduced 4T
vice verIa cannat be a.certained without clear chronological data. More
detàl1ed pal.obotanieal evidence 18 needed to elaborate on the prom~slng
relearch of Doyle (1973) and other paleobotanists. / ,
..
'}
.)
1
1

"
•
• .-
AClCNOWLE DQoŒNTS
Thi. inve.tigation wal 'upported by a POltgraduate Scholarlhip "1
1560 (to U.P.) and by an operatins grant Aa594 (to ~.S.) from the,
National Relearch Council df Canada. Ve ~i.h to thank Dr. Philippe
Forelt for co11ectinl lame of the mat.rtal uled in thil .tudy and
Dr. P. B. Toml1nlon for valuable commenta on the final manu.cript .
-, ,-, • +
, *~ ........ -- 1 .,
f. -J
•
.. 1
.' .
'. l,',
•
q'
'89,
j
1 /
, ""
< ,~; 4"1
" • i
. ~
~
" .. i " dO,
, lJJ

. ,
• -
, • ..
'- LIST OF ABBkEVIATIONS
A - stamen or its primord~
Ac - stamen connective.
Aco - connectiJe outgrowth.
As -procembial strand of stepen.
F - floral apex. , ~ Ji : ~ Fi • 10000er floral apex or bud.
Fu - upper , '" flor .. 1 apex or bud.
G • carpel or iu primordium.
Go - carpellary outgrowth.
G. - pro cambial strand of carpe1.
11 - inner integument or its primordium.
10 - outer intekument or its primordium.
L - veget.rive appendase or its primordium.
L. - aheathing prophyll or its pri~rdium.
N - nuce1lua.
-R • inflore.cence apex (reproductive apex).
al • proc .. bial .trand of Inflor •• cenee &xii •.
r • removed ••• 1. rL • ~Ietative app.ndaae reaoved. - '
8i • Itia-a or developlaa .t! ....
T -. theca or it. prt.o~diua.
Ta • aborted t~._.
v • ft •• taU~ apu (1'0"'1 ar.CNtb apu) •
.• t',
" o
1
70
.. l "

-
71
PLATE l Figs, 1-8
), Figa. 1 and 2. Side vtewa of very young inflorescences (R). \
Flg. 1. Both upper (Fu) and lover (FI) floral aplces being'lnltlated
1 at the Rame time al the vegetative rènewal apex (V). x 146.
Ftg. 2. Sllghtly oider stage than that of Fig. 1. The renewal growth
apex tV) la developing in the axil of a leaf (L) on the main
\
Fl~ • .J.
\ '
Fig. 4.
axis. x 146. \
Top vie~ ~f upper floral bud (Fu) beginning to Inltiate the
lover stamen primordium (A). Two renewal growth apiees (V) .. can be seen, A leaf (rL) of the main axis was removed. x 146.
Top view of an upper floTal bud (Fu) and two renewal growth
'" apice. (V}. The uppermoat renewal growth apex 18 Just initia-. \,
ting it. prophyll (LI). x 146. ,
~ig, 5. A median longitudinal section, through the inflorescence of 1
Fig. 4 •• hoving initiation of Itamenl (A) &Rd the prophyll (La):
x 237.
Fig" 6. Side view of yOung inflorescence. The prophyll (L8 ) te begin-
,\ .
ning to extend on to the inflorescence axis, below the up~er
flbral bud (Fu). x 146 • •
7, Top view of an upper floral bud (Fu), Ihoving both Itamen pri-
mordia (A). The mArgin of the prophyll primordium (L.) 18
becomina lobed:'x 146.
Median longitudinal I.ction~hrou~h th. ~nflorelcence of Fig. 7.
Procambial atrandl are beginning to d1ffarentl&te towardl the ,
upper and lowcr floral bud.. x 370 • j
. '
------'------ -~-- -- .
1
•

< '
If
• "-
f -• . ' ,
$

72
'PLATE II Figs. 9-14
Figs. 9 and 10. Top vievs of two floral buds.
F1g. 9. A lover floral bud (FI)' in which the stamens (A) are already "
developi~g the thecae. Two carpel primordia (G) are just
~ forming on the transversal flanks of the floral apex. x 146.
Fig. 10. An upper floral bud shoving four gynoecial primordia (G).
The tvo transverlal gynoecial primordia are becoming peltate.
The prophyll (Ls) has overgrown the lower stamen primordium.
Note the broad connective (Ac). x 146. ~"...
Flg • .1l. Oblique view o~ a young inflorescence showing the 8light ly 1
curved upper staDen'connective (Ac). x 146.
Flg. 12. Top view of young inflorescence looking down on the upper
stamens (A) of the upper (Fu> and lover (FI) floral b~ds,
.hoving the initiation of connective outgrovths (Aco). x 146.
Flg. 13. Top view of lover floral bud (FI), ehowing the connective
outgr~th (Aco) on its upper lUmen primordium and also on
the upper .tamen primordium of the upper floral bud. x 146.
Fig. 14. Median longitudlnal •• etion through the young inflorescence,
.bowing formation of connective outgrowth (Aco) and of carpel
pri.ordia (G). Tb. developing prophyll (L.) ls made up of
on1y two ceU 1ayer'l. x 370.
41
•
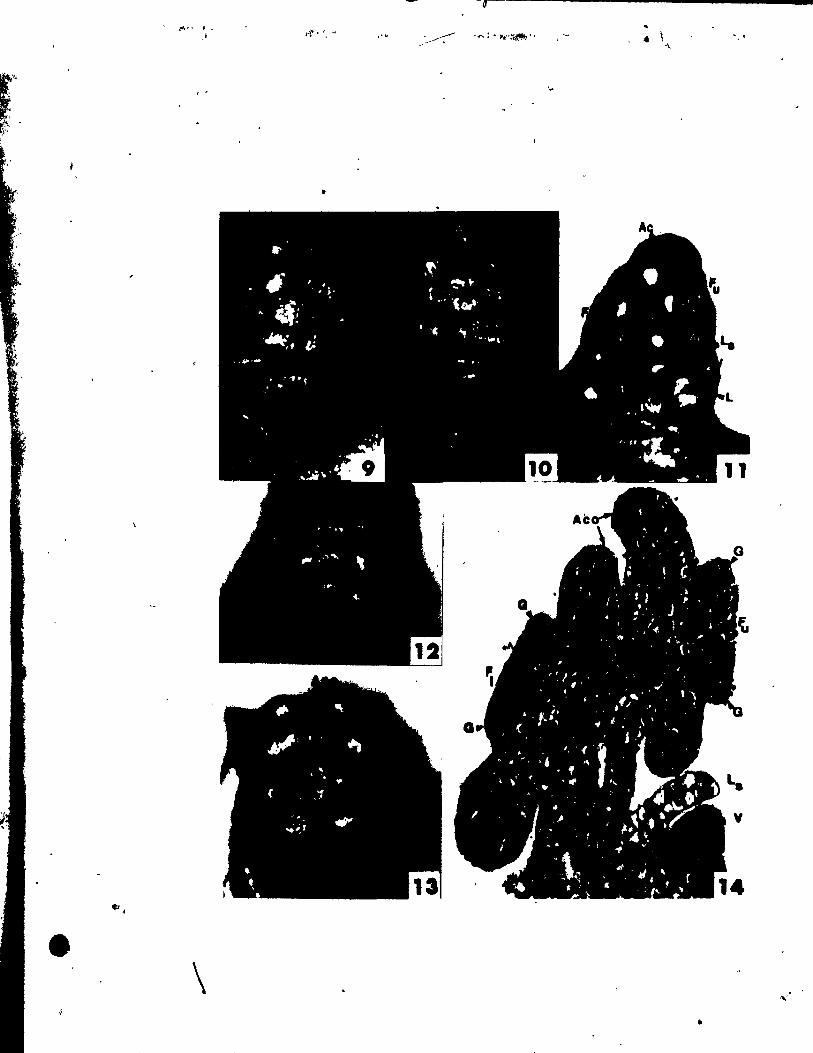
'" f-, .. ~ .
ô"
\
t4'." ~ . ,
•
,'b
.. .. \ ....
..

73
-
PLATE III Figsf\ 15-18.
Figs. 15 and 16. Top viewl of two lower floral buds.
Fig. 15. The connective outgrowth (Aco) can be seen developing between
the bisporangiate thecae of the upper stamens. AlI four car~
pel primordia (G) have become peltate. x 146.
Fig. 16. The connective outgrowth ~f the' upper stamens is becoming
doreiventral. The tranaversal carpel primordia are forming
t ovules (0). x 146.
Figs. 17 and 18. Median longitudinal sections through the
young inf/~cence 0f1fig. 16. /J~~
The upper floral bud (Fu) .howi the connective outgrowth (Aco)
and ovule (0). initiation; The prophyll (L.) already envelopl
a portion of the upper floral bud. x 370.
Fig. 18. The lover floral bud (FI) .howi the ataminal procambial Itrande
(AI) already well-differentiated. Procambi~l development tovarda
the carpel primordla (G) il jUlt beglnning. x 370.
J, _,-~
•
o ---

, .
,

.. "
74
PLATE IV Figl. 19-22
" Figs. 19-22. Top vie.Ja of various florallbùda.
r ...
Fig. 19. An upper floral bud, showing developing carpel. (G)
which already enclole the ovules. x 146.
Fig. 20. Viev from behind the upper floral bud showing connective
" o~tgrowth (Aco). The upper stamen of the lover floral bud
has a branched connective outgrowth. The developing car-
pela of the lower floral bud are forming atigmas (Si) and
outgrowths on their upper abaxial,aides (Go). x 146.
Fig. 21. View from behind the upper floral bud (Fu) .hoving the con-
nective outgrowth (Aco). The outgrowth ~ thil .tage i.
dor8iventral. The laver floral bud (F1) ha. a pentamerOu8
gynoecium. x 85.
Fig. 22. The developing atlgmaa (Si) are becoming flat~ened. The
•
earpellary outgrowth (Go) i. quite prominent. x 146.
• r-' 1
•
-
1
-, !
1
' •
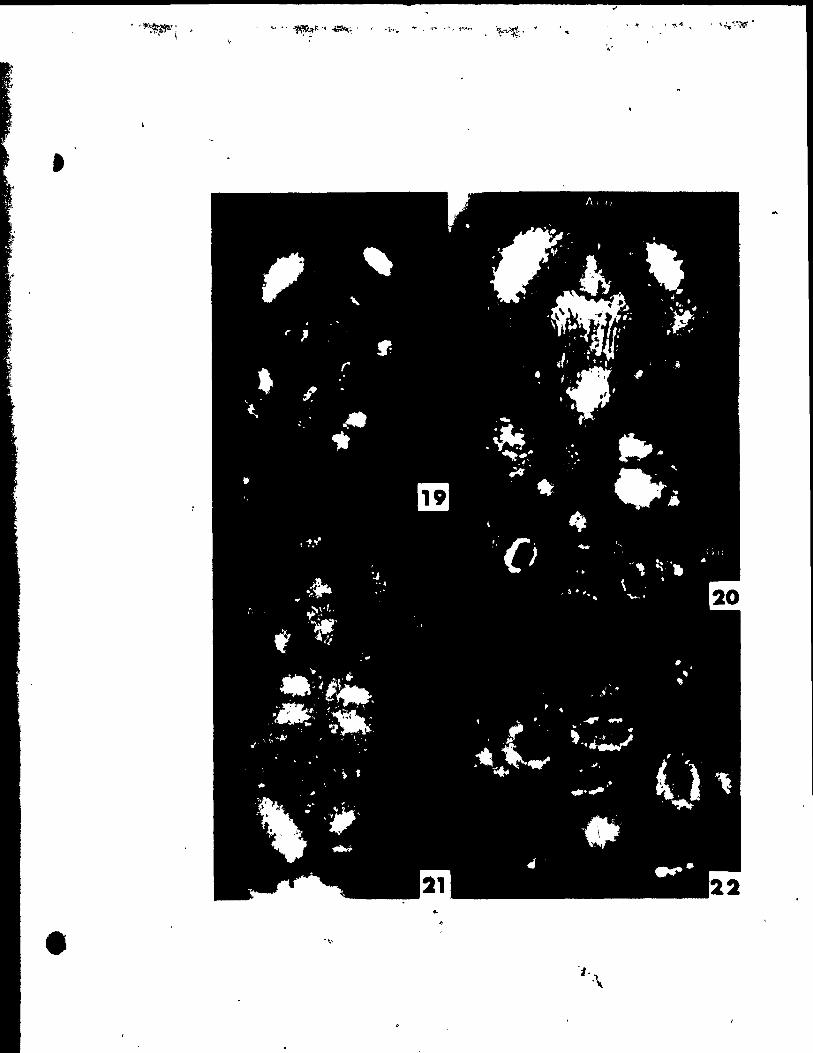
.'
•
..

"
o •
PLATE V Flge. 23-27
Fig. 23. Slde vlew of 'a trlmeroui gynbectum, showlng the peltate
stlsma (S1), and the carpellary outgrowth (Go). One of ,. the 8tamen thecae (Ta) has aborted. x 146 •
. ~ Fig. 24. The carpel wall has been d1ssected avay to reveal the
developlng ovule (0). The ou~er ~Io) and 1nner (Il)
tntegUDentl have almast enveloped the nucellu8 (N). x 146.
'118s. 2S and 26. S1de vievs of almalt mature inflorescence.
F18. 25. The sheathl" prophyll (La) almost covera the uppf!lr flord
bud. x 37.
Fig. 26. Inflor,.acence vith two nearly mature flowera. x 48.
Fi8. 27. Top vlev of the Im.er flover and rear vin of the at_en of
the upper flower. x 48.
4'
..
,~ ~.
," ..... " .. : ;"J" i~. ....-k,:;J~. '''; .. ' ... ' ~ ., ~~, ..
_ ,~ \l J.Jt,i,..', ~~~ r ;:~ ~,f ..,' ~ '"
l, ":, . -, . i, • ~
• -_ ... ',,! 1"4
! t ,: :,:;.t;:~r
, '
• .,~
~
! t
.-1 '" •
" ,. " } (
'\
• ,. 1 j

•
•
.. "

1
1>It,.r l "l
78
, PLATE VI Fig ••. 28- 31
~ Flg. 28. Side view of cleared tnflor •• cence ,hoving valcu1ar .tranda
,,,,"
of stamena (As) and carpéh (Ga). x 48.
FiS. 29. Top vi~ of the lower flover (Fl ) of the same cleared
inflorescence a. Fig, 28, The branchlng of the staminal
8trand (AI) in the lover portion of the connective can be
seen. x 48.
Pig. 30. Median longitudinal section through a carpel, .boving proc~
bi.l strand developing towards the bitesmic ovule, The basal 1
portion of al'lother carpel (G) can alto be .een. x 148,.
Fig. 31. Side vi .. of mature fruitl. x 4.
" , " i'
;:~, • v,, , .... \"1' ..,r
~'
',,'.~~~ .. " , 1 t ~ r
... ., _...!. r ... ~. 1. ,' •..
-,
.. ~.. "'"
•
, ,

, , •
,#
-',
28 , ,
,>
\ o 1

77
c ,
, '
FiaI, 32 and 33
Pla. 32.
Pla. 33.
1 MAlr&1lD&t1c repruentaU, 6f
/ from'aide (a) and top (b) ~lew.
, 'loral diaaram.
/
,
/ /
/ /
..
1
/ .
•
/
/ ;'
the pattern of
/
/
1 1
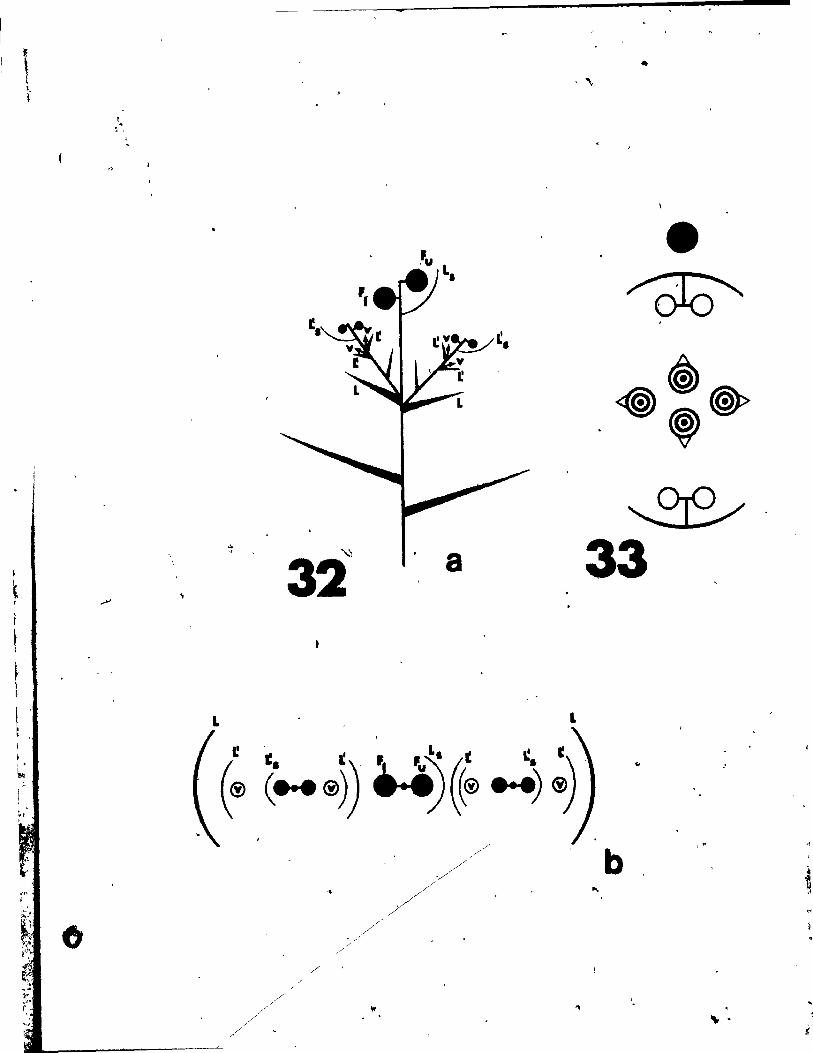
.,
32'" a
L L
I! .
(® ê~·. ®)) •. i)«~ •. ~ ;) . ..
/'
• ~
~ <@) @» , ~
33
..
b ,.
,
'" . .. 1
t

~
F1~ r~ 'j
~,
• :
-.
""
LITERA TURE CITE D
DO e,' J. A. 1973. The monocoty1edons: thelr evo1ution and cOm"-
parative biology. V. Fossi1 evldence on eaF1y evolution of the
monocotyledons. A. Rev. 8101. 48(3):399-413:
, Gam rro, J. C. 1968. Observaciones sobre la bioldgiA floral y
morfoligia de 'la PotamogetonAcea Ruppia cirrhosa (Petag.) Grande
(-!. spirille L. ex Oum.). Darwiniana (8. Aires), ~:575-608.
s, A. H. 1908. The morphology of Rupp!a maritlma. Tran~.
Conn. Acad. Arts Sei. 14:59-170.
raf, F. 1936. BIOtenbau und Vervandschaft bei den einfachsten
He1obiae. Ber. ntsch. Bot. Ges. 54:191-229.
78
Pos1iulllny. U., and R. Sattler. ,1973. Floral developlDent of PotAmogeton 1
densus. Cano J. Bot. 51(3):647-656.
1974. Floral develo~nt of Pot8DOgeton richard.onii.
Am. J. Bot. !!(2):209-216.,
Rodin, R. J., and R. E. Oavia. 1967. The us. of papa in in clearing
plant tissu.s for vho1emount •• Stain Technol. 42(4):203-206. . -,
Sattl.r. R. 1965. Perianth development of Potamogeton richard.oni!.
I.a. J. Bot. ll(l) :35-41.
1968. A t.ehnique for tbe .tu"," of floral d.velopaent. (
Cano J. Bot. .!l: 720-722 •
(
-
•
i 1
i

r
•
lr' .. " ...-' '
79
Singh, V. 19~5. Morphologieal and anatomieal studies in Helobiae.
Il. Vascular anatomy of the flover of potamogetonaceae. Bot.
Gaz. j1!(2):137-144.
Uhl. N. W. 1947. Studies in'the floral morphology and anatomy of cer
tain membera oi':~e Ilelobaie. Ph. D. Thesis, Cornell University, •
Ithaca. N. Y •
. ,
: .... ' 1 ". "
,,!,",

1 .. '
• '" ,.
-
r;t,
CHAP'l'ER 4
FLCl.AL DlV!LOPMENT OF ZANNICHELLIA pAJ,USmts
..
, , ',.. t(.( ~ < t,,) , .
80
,.' '"

•
-
81
ABSTRACT
What, at maturity, appears to be a bisexual flower in the axil of
one of two subopposite leaves, is revealed as a fertile nodal complex
with quite different organization. Three appendages develop at each nodal
complex. The first girdles the stem and becomes at maturity a membranous
sheath about the entire node. The second subtends the axillary meristem
which term1natè9 as the staminate flower, continuing laterally as a
renewal growth in the axil of a sterile appendage Just below the stamen.
The third appendage is subopposite the terminal meristem which gives rise
to the pistillate floral bud towarda the staminate flower, and a renewal
growth apex towards the appendage. This renewal growth apex repeats the
cnUre pattern at 'lllOlt 8 900 .hlft to the right or left, depending on the
shoot. The single stamen of the staminate flower develops as those studied
ln Potamogeton and Ruppia. The pistillate flower develops two carpel pri-
mordia, which become peltate before initiating a single ovule primordium on
the ad axial portion (Querzone). The membranou8 envelopé whi~h eovera the
carpel. at maturity ia initiated àt ovule inception, below one of the car-
pela. A peltate .tigma differentiates ,on a short style and at maturity .,;
beca.e. broad and lobed. The renewal growth apex hàs a one-layered tuniea.
The -..branoua .heath. of the node and of the pistillâte flower are pri-
marl1y epidermal in or1gi~, While the remainder of tbe ~terile and reproduc
tive appendq •• ub. throuah aetinty in aubprotodermal' cells. Procamhial
developMnt 1. acr~p.tal follovina cl0.al~prt.ordlal inception. Rach organ , Il. "" ,
(.terlh or fertile) racelve. one procamblal .trend, axcept for th~~m ...
branous .heath about the Dode and th. one about tbe.pl.tl11ate flover.
t
'. , ,

•
4
82
INTRODUCTION
Since Campbe11's (1897) comparative study of Najas and Zanniehe11ia
no further critical study of the floral development of Zannichellia has
been publiahed. Nonetheless, many intérpretations regarding its floral
appendagea have bean .ttempted. For examp1e, it was proposed that each
car pel represents a 8ing1e pistillate flower (Campbell, 1897; Uhl, 1947),
'. that the sing~e stamen of the staminate f10wer may be a fusion product of
two stamens (Eames, 1961; Sinah, 1965) and that the envelope about the
pi.tillate flower ia homologou8 to a perianth (Magnuè, 1869; Graebner and
F1ahau1t, 1908; Wettstéin, 1935), and 80 on. These assumptiona had taxo-
nomic repereu88ion8 8S to the positioning of Zannichellia within the
N~jad.le8. Sinee the Interpretation of the flowers of Zannichellia ia so
crltlcal and controveralal, the uae of up-to-date technique8 for the study
of floral development was thought us.ful to further eluc~date the work
be,un by C .. pbell. 1
MATEIlIAL AND METHODS
The material of Zannlch.~lla palustris L. wa. collected by the
author in July, 1971 in the St. Lawrence River between the Mercier. Bridge
and the Lachine lapida. It va. flxed and preserved in FAA (formalln-acetic
acid-alcoho1) before bain, .talned ln alcohollc acid fUéhain, di.sected, and
photolraphed .ntirel, t..&r.ed in 100% ethyl alcohol folloving the technique
of Satt1er (1,-68).
"ter1a1 ... prapar.d lo~ .ectlODlna hi 4ehydratina lt in a tertiary
butyl alcoho~ •• ri •• CDd .abed41na iD tl'.ue.at ua1Aa .taud.rd technique ••
• 1

•
-
83
OBSERVATIONS
At maturity both the staminate and pistillate flowers are found in the
axil of the lower (L2) of two subopposite leaves (Figs, 1, 17, 24), An /
&Xil1ary branch (V), positioned at the base of the staminate f1ower, can
a180 be seen in the/axil of this leaf (Figs, 1, 17, 21), The entire node
i8 enclosed by a membranous sheath (LI) originating below the two subopposlte
le.ves (Fig, 1), As weIl, a smatl sca1e-like prophyll (La) sepBrate8 the
pistil1ate and staminate flowers (Fig, 1),
The staminate flower consists of a,9ingle atamen with two bisporangiate
thecae (some with unisporangiate thecae were a1so seen) separated by a broad
connective, which 18 al1ghtly extenaed at the tip (Figs, 17, 18, 22, 24),
The pistillate f10wer consists of two to three earpels (usually two) encloséd
by a membranous envelope (Figs. 1, 19, 24), The style is short, the stigma
i8 fIat and peltate, alightly lobed st the margin (Figs. 23, 24). The fruit
ia an Achene on a short stalk, with a characteristically lobed dorsal keel,
The organlzation c~ be grasped and understood only on a developmental
basis. Figure 1 18 • scheme Vhich ineludes developmental evidence up to
maturity.
Qrl&nosenesia
In lta flowèring pba.e the .hoot of Zannicbellia p*luatris forms thr~e '" " . , , .
sterile app.ndase. (L l' L 2' L 3) at each nodal camplex, The firlt (L 1),
vhieh at maturity bec~~1 the megbranous aheath, girdles the axis 800n after
initiation (F1S" 2, 3, 6, 7). . ,
The second appendage (L 2) la.inltiated oppo-
.ite the flrlt, but 11i&htly h1sher up on the axis (Fl'8. 1, 3, 6, 7). An

•
84
• axillary meristem (am), that will terminate as a staminate flower, i8 ioi-
tiated in the axil of this second appendage (Figs. 2-7). At about the same , ,
time, the third sterile appendage (L 3) is initiated above L 1 and opposite
the axillary meristem (Figs. l, 2, 6), The tip of the main apex or terminal
meristem (tm) continues to enlarge, becoming dome-shaped before giving rise
, to a renewal growth apex (V) in the Bxil of the third appendage and the
rernaindér forming the pistillate (G) floral bud (Figs. 3-5). This sequence
of development la ~ted by the renewal growth apex in the axil of the
third appendage but with an Almost 90° shift to the lett (Figs. 2-6) or .~
to the right (Flgs. 7,9) dependlng on the shoot.
The staminate (Iower (A) deve10ps from the apical portion of the axillary
meristem in the axil of the second appendage of the nodal complex (Figs. 6, 7).
The axillary meristem first produces two sterile appendages (La); one toward
the plstillate floral bud (G) which remalns small and scale-like, and the
other towards the outside which subtends a renewal growth apex (V) (Figs. 6,
7). This renewal growth apex develops at a very slow raté, and even as the
stamen reaches maturity it remains smali and insignifieant (Figs. 17, 21).
At the same time as this renewal growth ts being inltiated, the apex of the
axil1ary merlstem terminates 8S the staminal floral bud (A), which rapid1y
de~eIops into an e10ngate primordium (Figs. 8, 9). At the distal portions
of thi8 primordium the staminal thecae are initiated (Figs. 10. Il, 14, 15).
The 81ight extenlion of the tip of the 8tamen connect'ive (Ac) occors 1ate in
development, after thecal dlff.rentiatlon (Flg8. 17, 18, 22). The atamen
remaina lea811. unt11 Just before anthe.ls, wben rapl~ upgrowth of the fila-
ment carrie. the anther up and out of the aheath (LI) enclol1ng the nodal
reglon ~'ig •• 21, 22, 24).
'ï~" ~!:.l , }
,,1 . , , 1
1
.1
f

" /
1
"
, \
•
·80
The piatillate flower termina tes the main axis. The floral bud is
radial at first, before initiating, the two carpel primordia distally
(Fig. 2,6). The carpel primordia ~re more or less radial (Fig. 2). before
. becoming peltate through a more ~apid,u~growth at the ~eriphery than the
central portion (Figs. 3, 8, 9). At about the time the stamen la differen-
tiating ita thecae. the single ovule primordium is in1tiated at the adaxial
wall (Querzone) of the carpel (F1gs. 10, Il). The abax1al portion of the
carpel wall, through rapid upgrowth, soon encloses the ovul~ (Figs. 13, 15) , 1
and begins to d1fferentiate a short style and peltate stigma (Figs. lG-LÜ).
The stigma, as it nears maturity, becomes very broad,and lobed at its
margin (Figs. 23, 24). At the stage of ovule inception, the sheathing
envelope (E) is initiated below one of the c~pels (Figs. 9, 12, 15). It
soon girdlea the peduncle of the pistiliate flower (Figs. 13, 14, 16) and
a,t ma turity forms a membranoua cave ring enclosing the carpels (Figs. 19, 24).
Towards maturity upgrowth occura at the internode be10w the sheathing
'leaf (LI) •• carryi,ng the uppermoat nodal comp1ex up and, away from the oider
one below {Fig. 1). A slight distortion occura as the renewal shoot, which
is positioned laterally at firat, straightens .out through more rapid up-
Irowth on one aide, bringina the staminate and pistillat.e flowers together ,
(Fia. 24), thu. cr •• ting the falae iapression of a bisexual flower in the
A .liaht ..,unt of variation wae notad in the flovera of Zannichellia
palu.tria. loae piatillate flowera vith three carpels were seen, but only
at lata .tasea of developaent. Staaens vith uni.poranaiate thecae w.re noted , \
quit. fraquentIy (Fia', 22, 24) •

f r
r.
, a
•
86
•
Hiatogenesis
.The meristem of the renewal growth apex (V) has a one-layered tunica. ,
The .first sterile appendage (L 1) la initiated primarily by divisions in
the tunica (Figs. 25-27). The second (L'2) and third (L'3) appendages
arise through periclinal divisions and cellular expansion in the subproto
~ dermal cells alone,accompanied by anticlinal divisions in the tunica
(Figs. 25. 27). The axillary merlstem (am) which terminatea à8 the stami-;
nate flower, initiate8 primar1ly through periclinal divisions in the corpus -
vith concomitant anticlinal divisions in the protodermal cells (Fig. 25).
The termin.l merlatem (tm) gives rise t~ the pistillate floral bud (F)
and the continuation renewal growth (V) throUlh periclinal divisions in
the .ubprotoderm and anticlina~ divisions in the tunlca (Fig. 26).
The pistillate floral b~d also has a one-Iayered tunica. lt initiates
two carpel primordia (G) through pericllnal division in the subprotodermal ~
cell (Fig. 28). Tbe carpel primordia become pe~tate (Fia. 30). befor~
glvina rise to ovule primordia through the actlvlty of aubprotodermal cells
at the adaxial p~rtion of the carpel wall (Fla. 31): The ovule develops
twô 1ntesu.ent ••. Both thè inner (Ii) and outer (la> integument. are
~
protoderaal in or1ain (Fig. 33). The sheathina envelope (E) initiates
belov one ~arpel thtouah periclinal divisiona of th. protodermal ~d Bub
protodermal cella (Fiai. 30, 32). ~e envelope only dev.lopa tvo cell
layer.f •• it alrdle' the peduncle of the pi.tillate flover and eventuall1.
,enclo.es lt (P1, •• 32, 33).
~ --The .inale .taaen of the ataa1nate flower bea1n. to dlfferentiate the
~ell. taat vill fora it. aicro'PlCe. At the/.taae the carpel. of the piatll-
late flowe~ are for.iaa-the1r ovule prt.ord1a <'ia. 31). The .llaht .. ten
.ion of the .e .... cODDecti.e occura late ta d ... lo,..at (Pla. 34).

•
-
87
Pro cambial Development
A 8ingle procambial strand difierentiates acropetally towards the f
renewal growth complex (Fig. 25). As the primordia of the second (L'2) •
and third (L'3) sterile appendases develop, s ptocambial strand differen-
tiates towards them from the main strand (Fils. 26, 29). No procambial
strand forms in the aembranous sheath (L'I) that girdles the node. The
mÀin procambial strand continues to develop towatds the terminal and axil-
lary merieteml (Fig. 26). After the formation of the ren~wal growth apex
(V) it branche., differenti.ting a strand towards the renewal growth apex
(where the entire pa~tern il rep.ated) And one towards the pistillate
floral bud (Fig. 29). 1
AI each of two carpels initiates it8 ovule, a single procambial strand
differentiates towar~s each one (Fig. 31). In the carpél the strand bifur-
cates,differentiating ventrally into the ovule, and dorsally towards the
relion that will form the style and sti ... (Figs. 32,~33). The st.-en dif-
ferentiates a .in,l~pr6c .. b1al strand in the connective relion (FilS. 30,
31). Tovard ... turity tWe valcular bundle in the stamen connective doea not
axt_nd into the exten.ion at the tlp (Fi,'. 34).
DISCUSSION
Althouah Campbell (1897) in hie develo~ntal study of Zannichellia 1
att.-ptad to clarify the probl ... ae.ociited vith the fl~er., hie u.e of
.. ialy .actlaned astertal'led to e.veral .teinterpretatlonl. For exa-ple.
~e cla{Md that th. u:lllary Urilt .. (aa). avbtended by the .econd ItarUel ,
appeada.e (L 2), .ave riae ta, the 1,taa1nate flover and a reneva1 arowth by
dichotOllY. Our ~baervationl clearly ahow that the uillary .rbt_ at
fir.t 1n1tiatee twa .'erile appen4aa.. (La) vith a renewal .rowth apex i~ J
. '

,. o
88
, the axil of the outer one (Figs. 6, 7), before terminating ~ a staminate
flower. Furthermore, in describing carpe! development and ovule Inception
Campbell claimed that, "the ovule is really an axial structure, being the
termination of a branch, and the carpellary leaf merely subtends it." We
found j.nstead that the carpel firat develops as a pel,tate primordium and r
gives riSè ta the ovule secondarily on the adaxial portion of the carpel
wall lFlgs. 9, 10). This supports the observations of Schumann (1892) and
Magnus (1894). .. ...
Probably one of the most contentlous issues in dea1ing with
Zannicheilla ls the nature of the envelope about the pistillate flower.
Magnus (1870), Graebner and F1ahau1t (1908). and Wettstein (1935) described , ~
It as a perianth surroundlng the pistillate flower. Campbell (1897) and
Uhl (1947) hOlllOloglzed the envelope to the tubular ''bract'' below each node
(LI) and further claimed that each carpel represents a sIngle flower. Our
developmantal work does not nec,esurily support these, interpretations. The
envalope la ptimarily protodermal in origin. much lik~ the Integuments of
an ovule, and dlfferentlates nO vaacular tis~ue (Fi88. 30-34). lt may-be , of • status inter&ediate between a phyllome and an emergence.
Much of the discueaion of whether Zannichellia has a tetra-carpe11ate
pi.tillate flower or an inflorescence in which each carpel represents a
.lnal. flover (an ln~.rpr.t.tion followed by Many flora., Ruch as Marie-
Vlctor1n, 1964, or Ma.o~. 1957). has focused on the inceptlon and posltlonlng
of the earpel'a. Sinah (1965) elalaed tl'lat the clrpeb &1:'e only part o.f a
pl.till.te flover aince they aril. in Ipiral luceesllon, whi1e'Campbell '. ..
(1891) atated t~t eaeh carpe! repre •• nta a piltillate flover .ince the two
earpele O'dl.1D.ated .1 a 'dlehotOllOUI branchlq .yu .. 11lte that of the teralnal
and axl11ary _rtn_. We éannot support e1ther atat ... nt. Firatl,.. our

o
89
obàervations show that the two carpel primordia are initiated almost
simUltaneously on the flanks of the pistillate floral apex, the central
portion of which i8 still visible at early stages (Figs. 3, Il), and
not through either spiral succession or dichotomy of the, apex; and
secondly, neither the axillary Cas discussed earlier) nor the terminal
meristeme clearly branch dichotomously. Though it is impossible to
establish beyond doubt whether the terminal meristem (tm) gives rise
to the renewal growth apex and the pi.tillste floral bud through dichoto~y
(as Campbell claims) or whether the pisti1late floral bud terminates the
shoot vith the main renewal growth continuing laterally (Schumann, 1892),
developmental evidence tends to support the latter view. In fact, it ia
only at young atages that the branching pattern can be clearly seen. for
as the internode. extend the initial branching pattern i8 distorted, 80
tha~ the pistillate f10wer is brought into a seemingly axillary position
next to the staminate f10wer (Fig. la, lower nodal comp1ex). "
The deve10pment of the single stamen of the staminate f10ver does
not 1end support to thoae (lames. 19~1; Singh, 1965) who believe that it
may repreaefit the connation of two anthers. As in Potamogeton richardsonii
and Rueela maritima the atamen initiates as an e10ngate primordium which
different~te& it. thecae at the distal portion.. Cases of tetrathecal
at ... na vith eight micro.porangia were cited by Singh (1965) as possible
~pl •• of connation. In our materlal,primarily bitbecal 8tamens with
four .tero.poranai. were found.,and when there was variation it vas in the
other'direction, that ia, uniaporanaiate the ca. occurred. This would seem
to in4icate that ~.rl.tlon in the nuaber of th.cae, or .lero.porangia per
thec.a, '1. 'luite ca.on in thia tamn.
Tha cl ... 1ficat1on of Zanaichal11a bal .raflected the conflict1ng

•
•
-
90
morphologieal interpretat1ons. 51ngh- (1965). for example, using floral
and vegeta'tive data, has argued for keep1ng Zannichellia together with
Potamogeton and Ruppia in one family, Potamagetonaceae (see Wettstein,
• 1935). Others. like Hutchinson (1959), have p1aced Zanniche11ia in
a separate family. '" Comparing the floral development of ,Zannichellia
Ralustris,with that of Potamogeton richardsoni1 (Posluszny and Sattle~,
1974&), !. densus (Poa1uszny'and Satt1er. 1973) and Ruppia maritima il
(Posluszny and Sattler, 1974b) some similarities as weIL as Bome differences
are revealed. Carpel inception and development in the four species
studieâ are similar; initiated as a more or less radial primordium. the
carpel becomes peltate and forms a single ovule at the adaxial portion
(Querzone) of the carpel wall. Zanniqhellia palus tris, though, does not
fom an outgrowth on the upper abaxial carpel wall as in Potamoseton and
Ruppia. The development of thé stamens i;"tt the specie9 i9 alike,
however, in Zanniehellia the stamen origlnates in a terminal position and
the number of microspotangia per st~en varies. As ln Potamogeton the stamen
connective in Zannichellia palustris forms a slight extension st tbe tip.
Comparing the branchlng pattern of the floral shoots, a morphologieal ...
series trom Potamogetonrto Zannieheilla (as proposed by Singh, 1965) was
noted. In both Zannichellla palu8trls and Ruppia maritima. the prophyll of
the tenewal growth apex beeomea a membranous sheath; but in Ruppia it sheaths
one of the flowers on th~ inflorescence whereas in Zannichellia lt sheaths
the'entire node of thè renewal growth. '1
The MOst atriking differenee.between Ruppia and Potamogeton on one hand il!'
and Zanniehelli. on the othar 1a the separation of sexes in Zanniebellia •
Althoush at aaturity the stamen and tvo carpel. of Zannichellia palus tris
are clo •• toaether, aeeminaly in the &xi1 of one 1e.f, they arise indepen-
$ il t a 'i i .L ~

1
•
-
91
dently of one another. the pisti11ate flower being s product of the main
shoot, while the staminate flower termlnatt!s the axillary shoot. ThIs
does not necesssrily negate a link between Ruppia and ZBnnichellia, for
separation of the sexe'8 18 seen as a common phenomenon in other groups
(e.g., Hydrocharitaceae) where trends cu1minate in hyarophily (Sculthorpe,
1967); but it still leaves a Véry large gap between them. On the basis of
our comparative observations we tould, therefore, $ee some evidence for
8ssigning Zannichell'ia to a sepsrate family. However, before ma1dng 8
suggestion in this regard, we would prefer to study other sPecies of
Zannichelli; and related genera that might bridge the gap between Zanniche11ia
and other Potamogetonaceae. \
\,

• ' iIf
1 LIST OF ABBREVIATIONS
A .. etamlnate flower or its primordium.
Ac • tlp of stamen connective.
am • axlllary merlstem (in the axil of appendage L2' develops into
staminate flower and renewal growth). , E· membranou8 envelope around plstillate flower.
" F· floral apex of pietillate flower.
G· arpel or its primordlum.
li .. 1 lntelument or its pfimordium. ,
1 • 0 Int_loment or lt. primordium.
..
, L !' L' 3" aterile appendagee of the uppermost renewa! growth
or
LI' L2 •
enewa! gtowth.
the ~aaedi.~elY ,
.. sterile appendage of below upper-
La e appendases of the staminal branch.
N • \
o • prilDOrdium.,
r • • 8.1. t:L· v ••• taUv8 appandale re.aoved.
S • intravallnalil.or ita prlmerdlum. 1
r de~lop1D1 It~.
Sy • developina .tyla."
ta • .. riat.. (developa into piatillate flower and renewal
p-awth)
V· r ..... 1 ITOWth .,. •
,l
"
92

-
93
Fig. 1, a e.nd b r
(
, .
Fig. 1. D.Lagr .... tic repr~8entation of sterile and ,reproductive appendages
of ~wo node. at different .tage. of deve10pment •• ide <a> afid top
(b) vie" •.
". ~, \
.)
...
• ,
" ~ ... • ,
\\, ... .
\
"',
.,
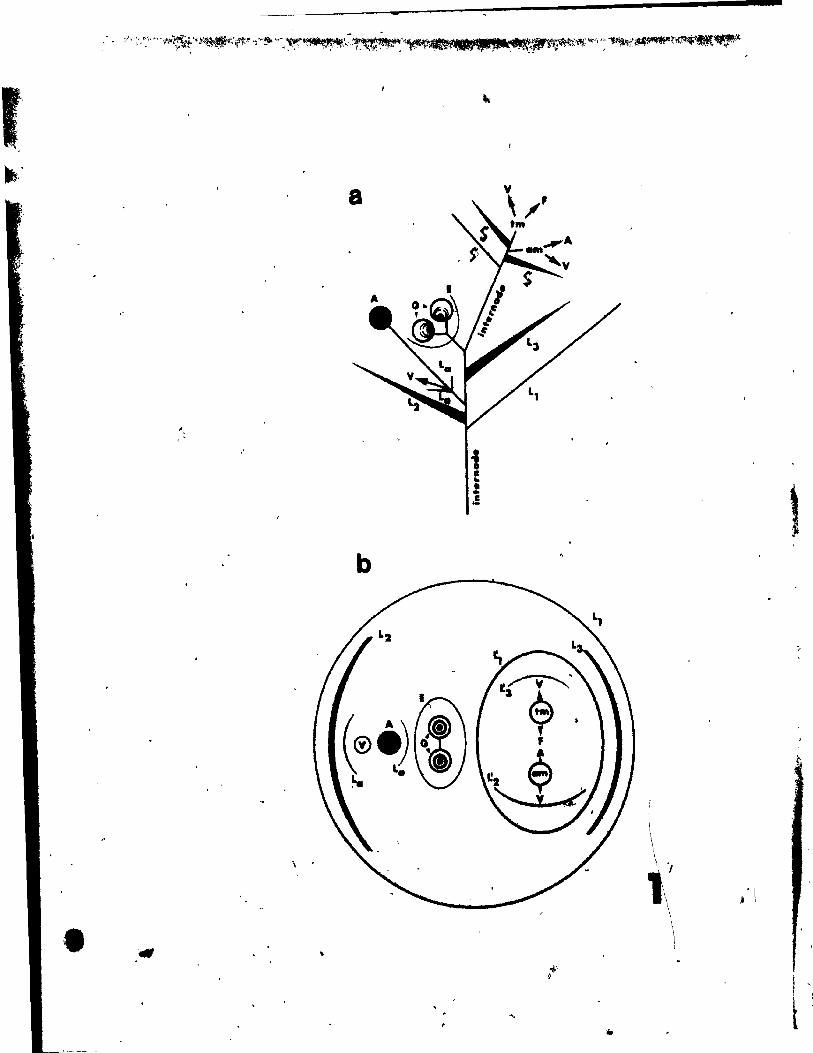
-
,"
b
i e ! .!
~.
f \ 1
1 .t '
l " t j

PLATE l F18S. 2-11
Figs. 2 and 3. Top views of apical renewal growth complex
and young floral stages.
Fig. 2. A young renewal growth having already initiated sterile appep-
dases (L'l' L'2' L'3) with terminal (tm) and axillary (am)
lIleristems. Sterile a:ppendages and floral' buds of underlying
node are also seen including the third (L3), a portion of the
second appendage (rL2), the staminate branch which termina tes
ln a stamen (A), continuing in a renewal growth apex (V)
which ia subtended by a sterile appendage (La)' and the floral
apex of the piatillate flower (F) wh1ch 18 Just initia tin .. !ts •
two carpels (G). x 146.
Fia. 3. A stage allghtly older than Fig. 2. The uppermost renewal
arovth apex has alr4 weIl developed ste'rUe appendales (L' 1
bellnnlng ta girdle the node, L'2_ L'3). The terminal meristem
(tm) la Just forming the apex of the pi8tiIIate flover (F) and
the renewal growth apex (V). The axillary meristem (am) i8 in
the &XlI of the second 4Ppendage (L' 2)' The sterile appendages
of the ,underlylng node have been rem~ed, ~xcePt for a portion
of the aecond one (L2)' The slngle atamen prtmordium (A) of
th. staa1nate flover has elongated prior to thecal lnit~ation.
The lateral renewal arawth apex (V) of the _tainsl branch 18 /
(Q) of
.xll of a atarile appenda,e (La)' The carpel primordla
.Ile piotilla.e fl_ aro j",,' bO~" paltot •• x 1~6\ in the
\
, \ ,
\ \
, •
o

•
e
Figa. 4 and 5. Side view8 of uppermost renewal growth complex .
• Fig. 4. The te~inal meristem (tm) has ndw clearly developed into the floral
Apex of the pistillate flover (F) and a renewal growth apex (V).
x 146.
Fig. 5. A stage similar but slightIy aIder than that in Fig. 4. x 146.
Figs. 6 and 7. Side views showing young staminal branches.
1
Ftg. 6. The young staminal branch has already initiated two sterile appen-. dag.s (La)' a~lateral renewal growth (V) and a termInal stamlnate
flover (A). x 246.
Fig. 7. A staminal branch slightly older than that in FIg. 6. x 246. , Flgs. 8 - 11. Top vievs'of young carpels.
Fig. 8. The two carpel primordia of the pistillate flower are Just
becomlng peltate. The young staminate flover to the right (A) i8
elongating prior to thecal initiation; the older one to the left
has already developed a sliaht extension of the connactive (Ac).
x 146.
Fig. 9. A s1ml1ar ata.e aa that of Fia. 8, but'fram a alightly different
anale. Envelope (E) about pi.cillate flover i8 Just inltiating
balow one of the carpels. x 146.
Fi •• la. Bach carpel (G) i. juat initiating its ovule (0). The 8tamen to
the rl&ht 1. diff.rantiatins lt. thaeaa. x 146.
FlI. Il. ~ carpel vall. (G) are be,ianins ta overlrov the youna ovule.
(0). x 146. •
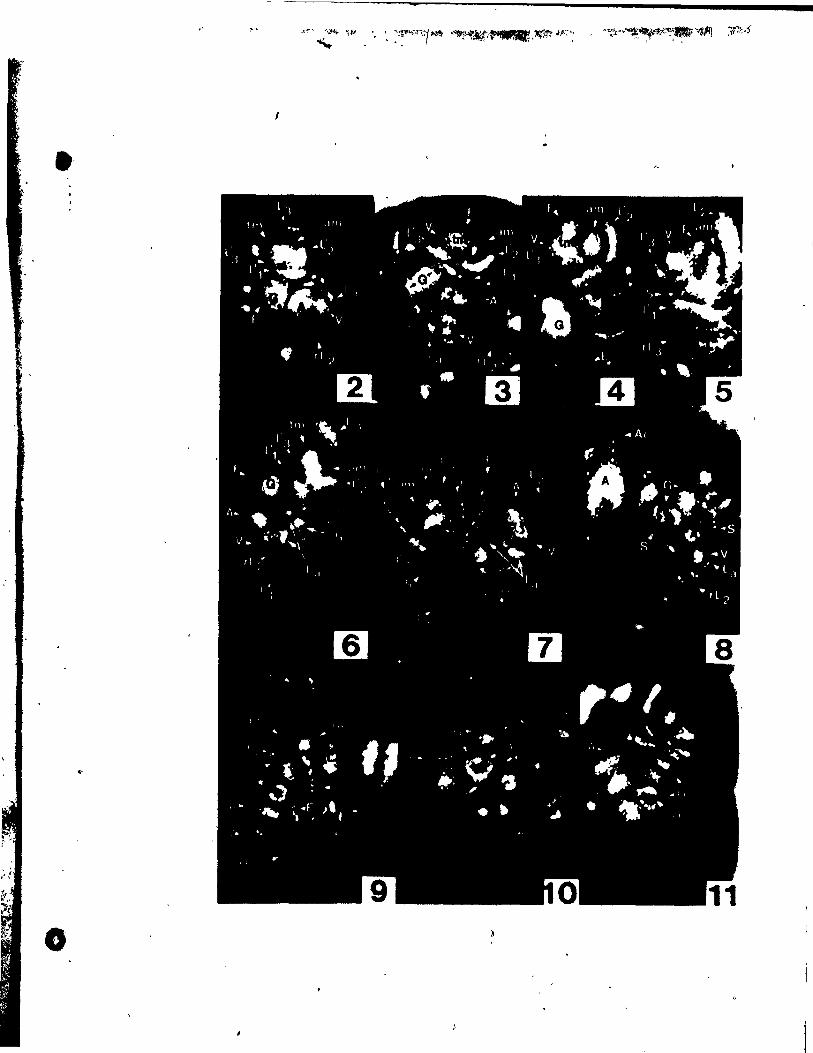
.'
1
.'
o

-
96
PLATE II Figs. 12-20
Flga. 12-14. Side viewi of developing plsti1late flowerl
and thei~ sheathing envelopes.
, Fig. 12. The aheathing envelope (E) can be seen juat at initiation
below one of the carpels. x 146. .,
Fig. 13. Carpels (G) just prior to total enclosure of the ovule (0).
The sheathing envelope (E) 18 about ta gird1e the peduncle
of the pi8tillate flower. x 146.
Fig. 14. The sheathing envelope (E) completely girdles the peduncle
of the pistillate flower. x 146.
Fig. 15. Top view of carpels prior ta complete enclosure of ovules
(0). x 146.
Figa. 16-20. Side views of carpels approaching maturity.
Ug. 16. Two carpels (G) at a stage jUlIt .fter eneloaing their ovules",
The atamen of the ataminate flower to the right i8 forming a
alight extension of the connectiv~ (Ac). An apical renewal
ca.plax can be aeen to the left of the platillate flower.
x 146.
Pig. 17. The carpel. (G) of the pi.tillate flover have already developed
peltate atia-aa (Si). The thecaa and extenaion of the connec
'tive (Ac) of the .taaen (A) afe nov claarly differentiated. x 85.
A portion of the .heathi.na _velope ~. baen rellOved to show two
~acpe1a (C) just Inlel.tine thelr 'ti .... (81). x 146. - j
\ 1
Plg.' 19 • .11atUlate flow.t Vith the abuthilll 8Ilvelop4t intact. x 85.
Fig. 20. Piatill.te flower differ.atiatiDa at1l-a (Si) and acyle (Sy). "
K 146. •
----------~---- ,
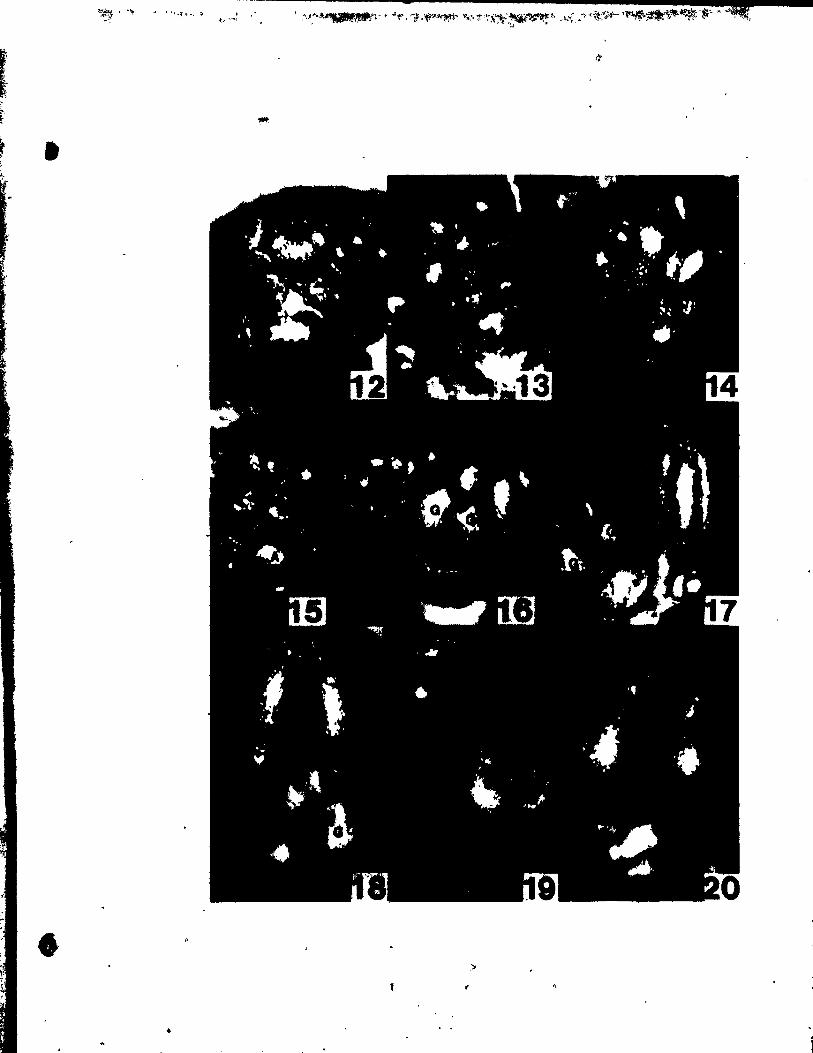
-,_, l .~ '.~, ':)< !
i"," --,
•
)
•

\' \
PLATE III !iga. 21-24
Figs. 21-24. Side view. of nearly mature atadinate and
piatillate flowera,
Fi8. 21. A lateral renewel growth coapl~ at the baae of a ~e.rly
fully devaloped .taaen (A). x 146.
118. 22. A .tameo that ha. dav,eloped uni8potangia~e thecae. x 74.
F.18. 23. Naarly mature carPeli of a pi.till.te flower in which "the
.heathiDa env.lope (rE) haB bean r~ved. The atyles (Sy)
and atiamaa (Si) are a1mo8t fully developed. x 146.
Fi.. 24. A _ture aea.e ahovin. po.iUoual ralat10nBhip between
pi.tillate end .taa1na~e f~ower.. Portions of Bterile
o
. ,
• fil
. . .. '.
•
r
•
,<1 ' .
)
o
1 , ~
J
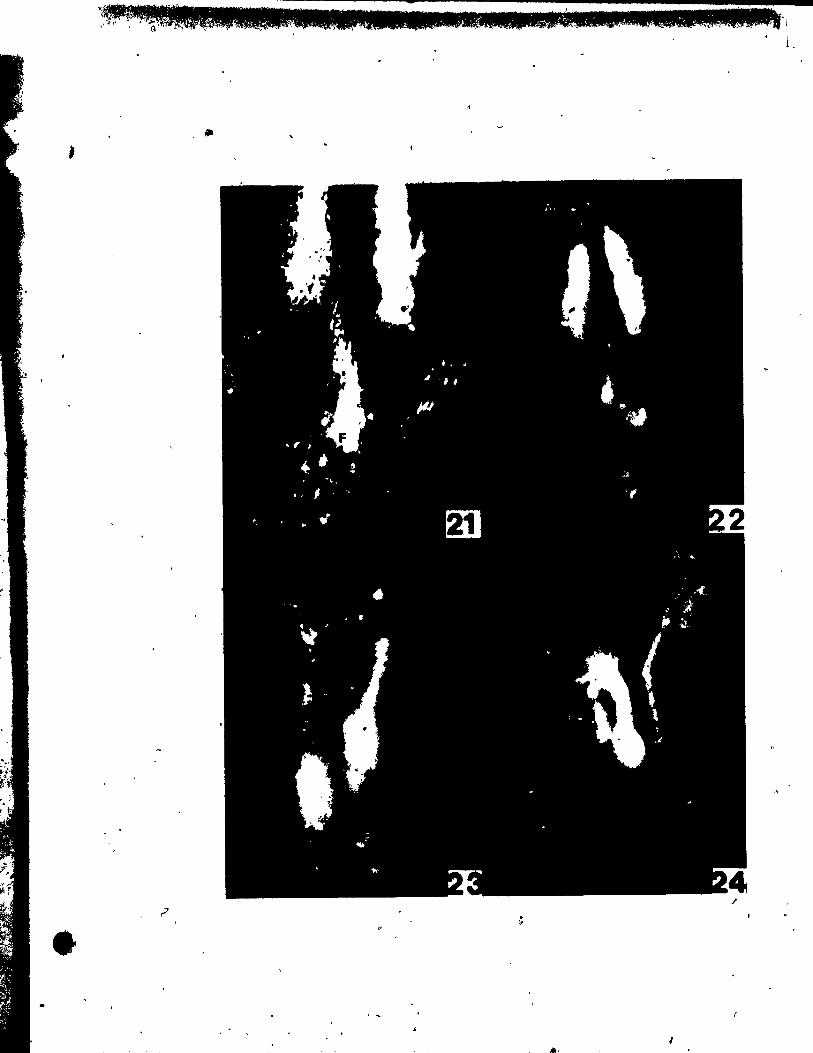
~Pl''IW . , G IE.l0 ..... _,as • .. •.. 11,. , ..... Q. lU· l, .'w.' .: . L
,fil ,

98
PLATE ~V Figs. 25-29
Figs. 25-29. Median longitudinal sections t~rough the uppermost
Fig. 25. A section
rene)rl srowth complexes.
Iho~nitiation of Iterile appendagea (L'l' , ,
L 2' L 3)' and axillary meristem (am) at the periphery ~f
the terminal merlstem (tm). x 370.
Fig. 26. The terminal meristem (tm) i8 jU8t beginnins to develop
Into the floral apex (F) of the pistillate flower 4,nd a
renewal growth apex (V). ~ 3~O.
\
Figs. 27 and 28. Iwo sections through different portions
of a ~newal growth complex.
FiS. 27. The very younS renewal srowth apex (V) 1. jUlt initiating ,
the firat and second ~terile appendage. (L'l' L'2)' A
portion of the floral apex of the piatillate flower (f) can
alao be seen. x 370.
Fia. 28. A tanaenUal aection of the apex of the pisti11ate flower
(F) ahawn in FiS. 27. Initiation of a carpel (G) can be
••• n on the left. The axillary aeriatam (am) of the
.,tadnal brech ia in' the axil of thl 8econd appandaga
(L2)' l[ 370.
Fil. 29. A a.~tioD .i.tlar to tn-t in Pi •• 26. but at a'ali,htly
'older .t •••• l[ 370.
/
, , f

-, -'
:". > ••• ' ·r:~"~i:Y~·;4.~~.!' ••• V,_ijJ" .• l .. ,a;·~ .. ~.p·,,,~"~~·
1 j 1 1
J

! 1
1 1
1 •
99
-,,"jI~ j Jr~I" "'-o\'li;t "".f""'"f' ,W~.r..("I~""""'t,"It*~""";' .. ~l""ift~"'I"~
PLATE v- Figs. 30-34
Figs. 30-3A. Median long1tudinal sections through various
stages of developing pistillate and stamlnate
-flowers.
Fig. 30. Young carpel primordia (G) Just becoming peltate (tangen-
tial section). The sheath1ng envelope (E) 1s being 1nitiated.
The microsporangial tissue of the stamen (A) Is starting to
differentiate. x 237.
Fig. 31. lneeption of ovule primordium. x 237.
Fig. 32. Ovule primordium (0) just prior to integument development.
x 370.
Fig. 33. Carpel with young ovule showing nucellus (N). inner (li)
and outer {Io> 1nteguments. Developlng style (Sy) and
/t/ stigma (Si) can al80 be seen. x 237.
1 1 Fia. 34. Néarly mature ataminate and pistillate flowera. One
-.
ea~el 18 eut ob1ique1y: The extension of the 8tamen ebn
nect~ve (Ac) protrudea ab ove mlcroapor'angia. x 148. \
--
If
•
\ ,
..

r 1
• ..

1;: ~ , • ,
• ,'"
• .,.'t ,' .. l.
LITERATURE CITED
100
Campbell, D.H. 18 7. A morpho1ogical study of Najas and Zanniche11ia.
Eames, A.J. 1961.
ser. 3, 1:1-61. v
of the ansiosperms. McGraw-Hi11, New
1908. II. Reihe He1obiae. Lebengeschichte
der B1Utenpf1anze von n von Kirchner, E. Loew, and
C. Schrater. Band l'tAbt. !:394-584. \
Magnus, P. 1869. der Gattung Najas L. Bot. Zeit. 27:769-
713.
1870. e zur kenntniss der Gattung Najas L. Berlin.
1894. Ober die Gattung Najas. B~r. d. deut8ch. bot. Gese1lach.
12:214-224.
Ma.on, H.L. 1957. A f10ra 0 the mar.hes of Californie. University of
California.
Karie-Victorin, R.C. '1964.
de Ho"nt~'al.
e Laurentlenne. Le,s presses de L'Université
Poslu.zny, U. and R. Sattler. 1973. F~oral dev.lopment of Potamoleton
dIQ8U!. Cano J. Bot. ~:647-656.
1974.. Floral dav.lo~nt
J. lot. il (2) : 20~-216 •
richard.onU. Amer .. ~~------- ,
\

101
, . , ' Posluszny, U. and R. Sattler. 1974b. Floral development of Ruppia
maritima var. maritima. Cano J. Bot. 52: 1607-1612.
Sattler, R. 1968. A technique for the study of Horal development.
Can. J. Bot. 46: 7,20- 722 •
Schumann, K. 1892. Morphologische Studien. Leipzig.
Sculthorpe, C.D. 1967. ,The bio1ogy of aquatie v8seu1ar plants. Edward
Arnold (Publishers) Ltd. London.
8ingh, V. 1965. Morphologieal and anatomica 1 studies in He1obiae. II.
Vascular anatomy of the f10wer of Potamogetonaceae. Bot. Gaz. 126(2):
137-144.
Uh1, N.W. 1947. Studies in the floral morphology and anatomy of certain
members of the Helobiae. Ph.D. Thesis, Corne1l University, Ithaca,
Wet~stein, R. 1935. Handbuch der systematischen Botanik, Vo,!. 2. Franz
Deuticke, Leipzig •
•

102
•
/
-, ,
CBAPTER 5
lL01W. DBVILOPMBIrr OF NAJAS J.anLIS
,
'.
f
, •

o
\
103
ABSTRACT
Tvo subopposite leaves form at a node. The lower one arises almost
simultaneously with the axillary meristem which it subtends. The upper
lesf initistes sfter the lower one and does not 8ubtend any structure.
The axillary meristem gives rise to a rènewal growth apex and a floral bud
almost at its incèption. In some cases the axillary meristem forma only a
floral bud. The floral bud may be either ataminate or pistillate. The
mâin axis and the renewal growth in the axil of the lower leaf repeat this
pattern of development. Stamlnate and piatillste flowers are ~lmost indis-
tinguishsble at inception. They form as dome-like protuberances and both
initiate girdling primordia which become lobed at or immediately after incep-
tion. In the staminate flower the glrdllng primordlum becomes the outer envel
ope, While a .èco~d girdling primordlum formed acropetally becomes the Inner
envelope. Bath envelopes overgrow the one-celled anther whlch Is the trans-
for.ed staminate floral apex. In the pistillate flowér the glrdllng primord-
lua becomes the gynoeelal vall that encloaed the single bitegmt~ ovule which
la the transformed pistillate floral apex. A short style forms and a stigma
vith 2-4 branches. The renewal growth apices have a one-layered tunica. The
two .ubopposite le.ves initiate through eeli division in both tuniea and aub-
p~otodermal cells. The axillary meristem ar18~8 through pericllnal divisions
in the .ubprotodenaal cells. The girdling priaordia of both staminate and
pi.tillate floral bud. are epideraal in o~igin as are the integuments of the
ovula. Proc .. bial developaent i8 acropetal following clo.ely primordia 1
incaption. Eàch leaf, floral bud and renaval growth apex receivea a single
.trand. No va.cularisation 1a eeen-in anvalopes of tha at .. inate flover or
the l1DOe~~~ vall of the pl.t~llate flower, all of vhlch reaain tvo cell
layer. thlck ...n at .. turity.
•

o
-
104
INTROruCTION
The flowers of Najas are amongst the simplest of the angiosperms. Yet,
they have been the source of much eontroversy for over 100 years. In parti-
cu\fr, the controversy has focueed on the two envelopes about the etaminate
flower. Swamy and Lakshmanan (1962) have summarized the many terme used ta
destribe these envelo~~ and the subsequent Interpretations that have
followed. Seulthorpe (1967) in reviewing the morphologiea1 and taxonomie
problems assoeiated w·ith the envelopes of Najas stated that, "until the deve1op-
ment and possible homologies of these struétures are better understood, it ,
might perhaps be poli~c ta avoid morphological eommitment." Furthermore,
there has been much disagreement about the nature of the plstillate flower. Its
terminal ovule has been used as evidence of a derived origin (Singh, 1965),
while a swelling âbout the style was thought ta be the remnant of a perianth, •
thus making it an inferior ovary (Miki, 1937).
Slnce Campbell's (1897) morphologieal study of Najas and Zanniehellia, no
further investigations on the floral development of Najas have been undertaken.
Sattler and Gifford (1967) studied the development of the vegetative phase and
floral induction in Najas 8uadalupen,ls. whereas SwamY and Lakshmanan (1962)
concentrated on embryology. Therefore. It appeared desirable ta fill the gap
befween the induction of flowering and the latest stages of floral development
by the present investigation which focuses on organogenesls and hi8togenesis
of the flower. of Naja. flexilil. Thil would provide ft basis for the inter-
pretive problems and make passible a comparison vith similar developmental
ob •• rvation.in the flowers and inflore.cences of Pot.-oseton. Ruppia and
Zapplche1Ua.

o
-
10~
MATERIAL AND METHODS
The material of Najas flexilis (Willd.) Rostk. aod Schmidt. was
collected by the author in July, 1971 at the Morgan Arboretum, Ste.
Anne de Bellevue, Quebec. It was fixed and preserved in FAA (formalin-
acetie acid-alcohol) before being stained in aleoholic acid fuchsin,
dissected, and photographed entirely immersed in 100% ethyl alcohol fo1low-
lng the technique of Sattler (1968).
Material was prepared for sectioning by dehydration in a tertiary
but yI alcohol series and embedding in tissuemat using standard techniques.
Sections vere cut at 6 micrometers (~m). Most of the sections were stained
vith Delafield'B hematoxylinô but some (Figs. 29, 30) were Btained with
Toluidine Blue 0 while still embedded in paraffin uBing the technique of
Sakai (1973).
OBSERiATIONS
Orgaqography
The flowers on a fertile ahoot are found in the axil of the lower (L'l'
LI) of tWQ suboppoaite leavea (Fig. la). On1y a single flover, either
staminate (A) or pietillate (G) is positioned adjacent to a reneval growth
branch (V). No pattern could be detected in the diatribution of staminate
~.nd pistillate flovers QP the fertile b~anche8.
The ataminate flover con.iata slmply of • uniaporangiate anthet on a
short filament (Figl. la, 24, 26). The ~nther la enclosed by two membranoua
I~ruèturea (Ei , B ); the outer one (E ) being hilhly lobed or toothed at the o 0
.pez (li .. _ 8, 26), the inner one (11) .~h.rinl ,to the anther, and being
bilobed at the apex ('il, 11). The pi.tillate fl~er~conslata of a unilocul- '
ar lYDoeeium, .nelo.ing • b •• al, aaatropoUl, bitegâie ovule. The etyle ia
.bort .ad'eade 1D 2-4 tootb-like atl ... tic projection. ('iBa. 24, 25).

"r'
o
106
Organogenesis
The fertile ~ranch of Najas flexilie forms two subopposite leaves
at each node (Lill' L"Z)' The Eirst {lower) leaf (Ln 1) initiates almost
concomit~t1y with the axillary meristem (am) which it subtends (Figs. 2,
8, 14). The second leaf (L"2) initiates slightly higher up on the axis
(Figs. 3, 6, 8, 9, 14). It does not subtend any structure. The main
apex (Vm) then continues to grow for a short time before repeating the
o above pattern but with a shift, approachiog 90 , to the right (Figs. 4,
7, 8, 9, 14) or the left (Figs. 2, 3, 5, 6). Relative1y little upgrowth
in the internodal region was found 1n our mater1al.
The ax1llary meristem (am) g1ves rise to t~o apices immediately after
or p08sibly even simultaneous with 1ta incept10n (Figs. 4, 5). One of the
aplces becomes floral (F), either male or fema1e, growlng rapidly and at
times obscur1ng the other apex (V) which develops vegetatively at first
before itself becoming fertile (Figs. 3-10, 13, 14). It shou1d be noted,
that not aIl axillary meristems divide. Cases where they form ooly a
single floral bud were seen (Figs. Z, 9).
The early development of the piatlilate (G) and staminate (A) flowera
are 80 similar as to be almast indistinguishable. Both are dome-like at
Inception (Flgs. 2, 4, 5), and both initiate glrdling pr1mordia about their
• periphery (Figa. 2-4, 6-8). This girdling primord1um becomes bi- or tri-
lobed at or 800n after its inception (Figs. 11, 12, 18). The firat clearly
t di.tingui.hable feature betveen pi8tillate and staminate floral bud Is the
shape of the developlng apex. The pistil1ate.!loral apex becomes narrow
and 'polnted (Fig •• 13, 32), while the staminate floral apex becomes round
and broad (Figa. Il, 14-16, 31).
The girdling prlaordiua in the staminate flower develops into the outer \
envelope (K ), which arowa rapidly vith much lobing at its tip "(Fig. 14). o
)

)
o
-.. -...
107
As the outer envelope begins to overgrow the apex, a second girdling pri-
mordium is initiated (Figs. 14,16). lt deve10ps into the Inner envelope
(Ei) which forms only two lobes and becomes closply adpressed to the
developing anther (Fig. 11). The staminate floral apex, after the incep·
tion of the two envelopes, is transformed into the unisporangiate anther,
which i8 soon enclosed by the ,overgrowing envelopes (Flgs. 6,8.11).
Towards maturity, the distal portion of th~ membranous outer envelope
elongates, becoming style-llke with serrated branching at the tip (Fig. 26).
Th~ inner envelope becomes almost completely postgenitally fused with the
anther and 18 almost indistinguishable at maturity. e'icept for the slight
extension of its bilobed tip (Fig. 24). The staminate flower develops a
very short stalk (Fige. 24, 25),
In the pistil1~te f1ower. the girdling prlmordium develops into the
lynoecia1 wall (G). As ln the outer envelope of the staminate flower, it
too becomes lobed at Its tip (Figs. 14, 17. 18). The f~oral apex 18 trans-
for~d into an ovule primordlum (0) as the gynoecia1 wall overgrows ft
(F'igs. 17-19, 31, 32). l'Wo integuments are initiated on the'ovule pri-
mo,dium, the outer (10) preceding the Inner (Ii) (Fig. 20). The rapld
growth of the. outer Integument may at t fmes obscure the inner one (Fig. 21).
After the Inceptlon of Integuments, the ovule turns downward, eventually
becomins anatropous (Flgs. 35, 36). As the gynoecia1 wall encloses the
ovule it belln. ta de~lop a Ihort style (81) and a lt1gma (Si) vith 3 or
4 branche. (:rigs. 2, 1" 22. 23)'. Towards Mturity,' these stigmatlc branches
become qulte 'long and filamentous (Fiss. 24, 25).
The apex of the reaeW&l Irowth,ot'the .. 1n &xi. (Vm) has a one-1ayered
,'\

r f ,
o
~
-, v "-l'~-
\ 108
" tunica. Roth the first (L"l) and se'cond (L"2) leaves are initiated
through periclinal divisions in the tuniea at their marginal portion
(Fig. 27) and pericllnal divisions ln the subprotodermal cells in their
median portion (Fig. 28);' As a consequence of this initiation, the upper
and marginal portions of the léaves remain oàly two cell layers thick,
while the bases have a one- to four-layered meso4erm.
The axii~ary meristem (am) initiatee through expansion and peri-
clinal divisions of cells in the subprotodermal layer, accompanied by
anticlinal divisions in the tuniea (Fig. 27). The major component of the
'axillary meristem, the rapidly growing floral apex (F) initiates ita girdling
primordium primarily through periclinal divisions in the tunies (Arrow,
Fig. 29). In the case of the staminate flower (A), the initial girdling pri-
mordium becomes the outer envelope (Eo)- The inner envelope (E i ) ariées
almilarty and both envelopes remain two cell layers thtck throughout thelr
dev~lopment (Figs. 30, 31). As well, both envelopes show a signifieant Al
amount of celltl1ar expAnsion at their upper portions towards maturity
(Figs. 33, 34). Stamen differentiation can be seen after the envelopes
enclose the anther (Fig8. 33. 34).
ln the pistillate flower the girdling primordium ~come8 the gYQoecial
~ \ ' wall (G). which encloses the ovule primordlum (0, transformed floral apex)
(Fig. 32). Like the envelopes of the staminate flower the gynoec!al wall
remalns tw,o celi layera thick and exhlbits'extenslve cellular expAnsion in
lts upper portion toward.'maturity (F1gs. 35. 36). Both inner and~outer ~
integumenta are protodermal ln orlgin, lite the gynoeeial wall, and 'they
enclo •• the rapldly d.Jèlop1ng nueel1us Just .a 1t turns downv.rd iuto the
&D4tropoU8 position. In turning downward. a portion of the outer integWll8nt
beca.e. .0 cl0 •• 1, adpre •• ~ ta the funicl. th.t lt 1. barely dt.tingui.hable
(P1&. 36).
"

•
• ' ... '1
109
Procambial oevelopment
From thé 8ing~e procambial strand in tpe main axis (Fig. 27), brancQes • differentiate acropetally towards the two leaf primordia and the axl11.ry
merlstem,followlng closely the sequence of development (Fig. 28). Single
frocambial strands develop towards the pistillate or stamlnate flôral
apices and supply only the dev~loPlng ovule or anther (11gs. 31-36), No
procambial strands differentlate ln either envelope of the staminate 1
flawer or the gynoecial wall of the pistil~ate flower (Figa. 30, 33-36).
DISCUSSION
Thé flowers, af Najas flex!11s are remarkable ln many ways. Probab1y
the molt unusual feature i8 the formation of two envelopes about the single,
stamen of the staminate flower (some like Schumann, 1892; Eber, 1934;
Lawrence, 1~51; and Benson, 1957, reported on1y one envelope). In their
development and farm these envelopes re8amble very much the gyn~ecial wall
and outer integument in the pistillate flover. As the latter two, they
(arise ln an acropetal .equence &8 girdling prl~dia (see page xxi Sattler,
1973 for deflnition) whlch are Initiated in the tuniea and they remain two
eeU -layera thick without. valctllarization even at maturlty. The interpre-
tation of the staminal envelopes has been rather eontroverslal~ For example,
t ~t haa been debated whether or not the inner and/or outer envelopes are r . , '- , homolpsoua ta foliar appendagea, bract. or perianth members. Some have
pr.f~rred to u •• the more neutral term, .he4th, for one or both envelopes.
Th •• e *Dd otber interprétationa have been tborouah1y summarized in the worka
of Uhl (1947)" SVa.y and takahaanan(1962), 81ngh (1965) and Scu1tborpe
(1967),
In our opinion tbe .orphological f.nterpretat,lon of the enve\op.. aSO\lt
l

o
110
the stamen ls be~t approached from tpe point of view of comparative .. morphogenesla (van Heel, 1973). As pointed out, theae envelopes re8~ble
in their whole deve10pment the two outermoBt envelopes of the female /
1
flower (i.e. the gynoecial wall and outer integument). This resemblance ,.'
would aeem to indicate that during the inception and development of the
envelopes around the stamen primordium, the same (or aimilar) morpho-
genetic factors are active as ih the envelopes around the nucellua. This
meana, that marphogenetic factors no~11y testricted ta the Immediate
proximity of the nucellu8, have become expressed at a MOst unusual place,
i.e. about the stamen primordium on the male flower (for further discussion
of aim11ar phenomena, aee Cutter, 1965). Genera11y speaking, we have here
--an example df novel organization occurrini through a spatial (and maybe
al.o temporal) ehift of organ-determining factors (Zimmermann, 1959;
Heyen,.197); Sattler, 1974a, b). In this case, this leads ta a Most
unuaual spatial configuration of organs.
We showed that the ovule primordiu., including the nucellu~ i8 the
tranaformed f'loral apex, i.e. the ovule la initiated in an exactly terminal
(basal) poaition. The ovule cert.inly doea not arise laterally on the
,ynoecial appendage (i.e., the gynoecia1 wall). Therefore, it.i8 not
poesible to apply the car pel concept to this aituation (~e Sattler, 1974.).
The lYDoectUlll of NaJa8 ia clearly acâ"rpellate. Whether thk8 acarpellate
condition i. primitive 'or de~ived from a carpellate aynQecium ie a different
que.tion (S.ttler. 1974a). Sinah (1965) a.aumea ,that the ~ondition in
Najas ie d.rived by .hifttn, the latera1 ovule down into ba.al poaition.
Hov.v.r, on.'could a1.0 .. ka the oppo.1te aesuapt10n (see, a.g. Maanua, 1869~
1871, 1894; campbell, 1891). Such id ... have been expreaead vith resard to
other taxa with ba~al placentatioo br Haeu.e (1966) _ud Croiz4t (1960),
.'

-
111
" '" Macdonald and Sattler (1973) and Macdonald (1974) 'diseussed this iasue .. with regard to the basal ovule ln the Kyrlcaeeae.
In dealing with the deve10pment of the pisti11ate flow~r. other
questionable points ln the lite~ature were clarified. For example, Miki
(1937) claimed that an annular swelling developed st the lower part of
the stlgma. He then used this observation to eBtablish that the flower was
epigynous. No clear indication of a swelling was seen in the material w~ .. studled nor wal lt reported ln the work of Singh (1965). If such a
swelling did occur in some~species of Najas, it could be comparable to the
Ilight outgrowth on the carpel walls ~bserved in Potamogeton and Ruppia
(Po.lu.zny and Sattle%, 1973, 1974a, b), making it very unlikely evidence
of epigyny. Another point that was clarified is the description of integu-
lDent development in Campbell'. (1897) work on Najas flexil1s. He clàimed
that the Inner integument formed first, followed by the outer which was
partia1ly 8uppresaed on the side that the ovule turned towards its funicle. )
We obaerv.4. that the Integuments deve10ped in an acropeta1 sequence,' ~nd
tbat it wa. ouly after the ovule turned down Into It8 anatropous position
that a porticn of the outer Integument became closely adpressed ta the
funlcle, thua making it d1ff1cult to dist1nguish. TheBe observations are
8upported by Kagnus ~1894) in his study of Najas marina.
lt 1. not 8urpri.inl that vith aIl the se contentioue issu$s, the taxo-
naaic po.itionlna of Naja. ha. beau controver.~a1.' Miki (1937), Uhl (19~7)
and Sculthorpe (1967) have suamarized eome of the past taxonoaic interpre-- 1. 1
tationa. Most conaidet Najas as derived (exce~1on. are Campbell, 1897 and
Rand1e, 1930), but there i. 11ttle aareement on what ie ie deri~ed from or
hov it relatee to other .ener~ within the Najada1e.. Sinee .uch of the past
taxoDOaic epeculat10n v .. b ... d on va._tative and mature floral structures,

,.
•
...
112
It ls usefu1 to compare the floral deve10pment of Najas flexllls with that
of other taxa of the Najadale. previous1y .tudled, i.e. Potamogeton
richard.onii, P. denau., Ruppia ~ritlma and ZaQniehe11ia pa1ustris
(Pos1ueznyand Satt1er, 1974a, 1973, 1974b, 1975 in press). Although there
are many differences, the above speeies have certain develapmental charae-o • •
teri.tics in coœaon and a morpho10gieal series can be enviaaged from
Potamogeton to ZannicheIIla (Paslusany and Sattler, I974b, 1975 in press).
For examp1e, these species have slmilar gynaecla1 and andraecial deve1op-
ment and slmllar sympodial patterns of branehing in their fertile shoots.
Najas flexi1ls, however, has • monopodia1 pattern of branChintra10ng the
main axis. But with the compressed nature of the deve10ping node, it would
take on1y a slight spatial and temporal shift to praduce a situation slmilar
to that of Zannichellia pa1ustris. With this in mind Najas could have
certain affinities with the abave morphologica1 series. Even sa, major
differences in floral development exist between Najas and the other species,
primarily the development of the gynoecium and andro~cium and the enve10pes
about the staminate flower. These envelop.s are unique to Najas, and there-
for~, cannot be compared with s1mila~ structures ~n other Najadales, with
the possible exception of the ridae around the staminete flower of
Althen1a. Miki (1931), though, does use this eharacteriatic to derive Naja$
from the submerged Hydrocharitaeeae, where spathes are found about the
staminate flowers. lt would ~e neeessary to compare the development of
these spathes with the envelopes in Naja. to see how va11d thi. homology is. 1
One possible 1ink betwee~ the pisti11ate,flower of Najas and that of
Zannichellia wa. proposed by Magnus (1811), in eoapar1na the membranou •
envalope .round the carpela in Zannichellia vith the tlcarpe1-Hu" wall of II ..
I&Ja •• This id .. ha. fOUQd very litu. acc.ptance (».endle, 1899; Uhl. 1947; o '

I! ,,~~\
~
•
•
,t _
113
Singh, 19~5). Out developmental observations show that the envelope ln
Zanniehellia ia initiated after ovule inception on only one side of the
pedunele, while the gynoeelal wall ln Najas lnltlates as a glrdllna pri-
mordium prior to ovule development.
lt ls elear from our work on thé floral development, that Najas ls
distinct in the Rajadales even with respect to Zannichellia. If Najas la
to fit into a morphologieal serles, as dls~uaaed above, lnter&ediates
would have to be found, sueh as. for exemple, Althenia which is close te
Zannlchellia, yet has a rldge-like outgrowth about the ba.e of the staminate
flower. Unti1 such poseible links can"be more thoroughly studied, it
would see& advieable to keep Najas within a aeparate family, the Najadaceae,
which along wlth the Potamogetonaceae (Potamoleton, Rupgla) and the
Zannichel1iaceae (Zanniehe11ia) would be placed wlthln one order (Rend1e,
1930; Sculthorpe, 1967; Takhtajan, 1969), Furthermore, it eeem. useful to
follow Sculthorpe's (1967) advlce, "to refrain from phylogeneUc speculation
until the floral and vegetative morphol~gy of aIl genera are much bettei
undentood,1I and until additional evidenee from genetics, ecology, geography
and palaobotany 1s _v.il_bls.
';0

l,
f! ' f'i l
,
.".
o
,.- , , , \
LIST OF ABRJEVLATIONS
A '~ staminate flower or its primordium • . '
am • axillary meristrem (in the uU of leaf LI. develoPIii into t1~ .. r
and renewal Irdwth).
Ei • innar membranoua ,envelope around staalnate flover.
lo • outer membranoua envelope around staminate flover.
F • floral apex.
G • gyooeciua or lts primordium.
Ii • iuner intesument or lte prtmord~um.
i • outer intagument or lts primordium. o
114
L"b L"2 • leaves of the uppermost renewal growth or th.ir pd.llordia.
L'l' L'2 • leavaa of the node immediately below uppe~.t ranawal grovth.
LIt L2 - laava. of the third nQde from the top.
N • nucallu ••
o • ovule or it. primordium.
r • removed. e.l. rL • vaa_tative .ppandasa re.aved.
8 • .quaula intravaginalis or ita prillOrdi~.
81 • .ti..- or dev.lopin, .t! ....
5y • .tyle or ~avalopinl st y
V • ~.ter.l ran .. al Irowth
". - ape~ of _in ale. .~,
/ ,); ~"
1
:~ .,.1 '~ .
'~',i :i;~
.} 1
, ..
IJ

-lU~
t.'. 'ria. 1. D1a.r~t1c repr •• entatlon of .terile and reproductive app.oda.e. of chr •• no4 •• et 4ifferent .taaèa of 4avalopaent, .ide (a) and
top (b) vieY ••
• 0 •
1
< ~ "~ ____ ...... _____ ... 1
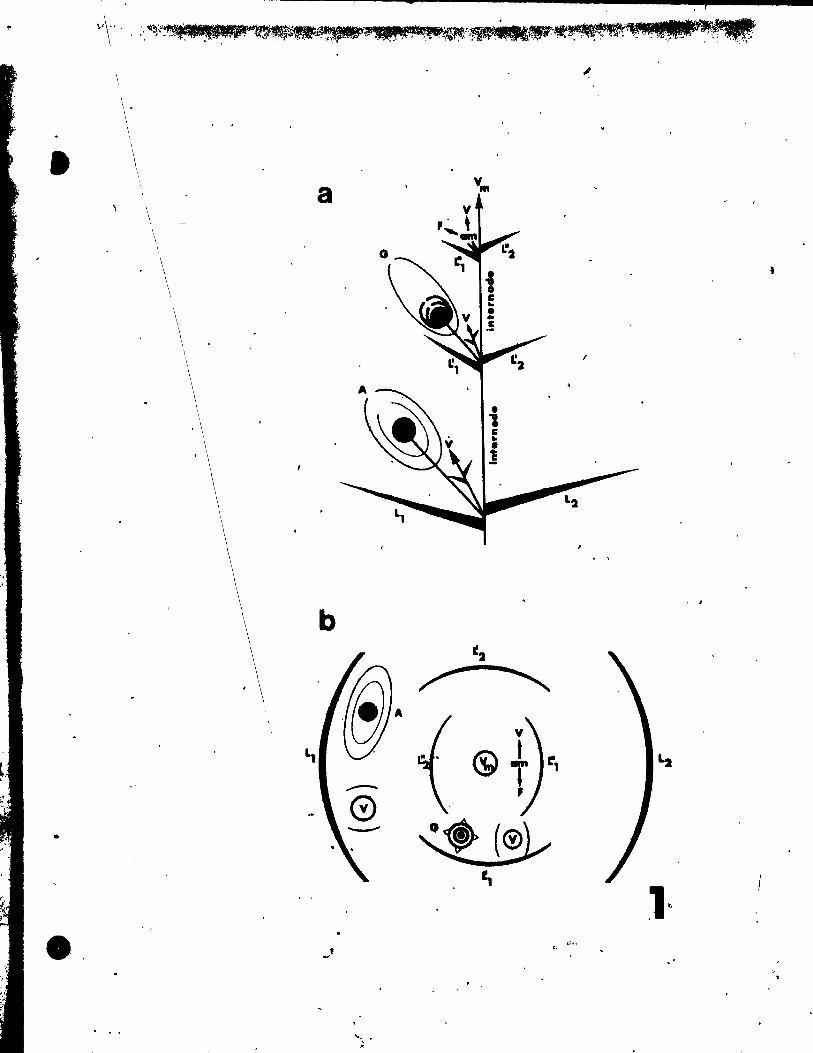
1
•
-" . "
\ \
\
a
b
L1
--0
.J
- , ,
1
, 1
~2
~
V
~t T
1:',
~ ~
1~ b V'~

-116
PLATE 1 Figl. 2-13
Fig. 2. Side view of three nodal coaplexea. The uppermost node ia just
lnltiating lu firet leaf (L\) along with the axl11ary mer18tem
(am) which it subtends. At the tip, the apex of the main axis
(Vm) can be seen. The second nodal complex Is developing Just
below, with Its flrst leaf (L'l) and Its floral apex (F). The
lataral renewal growth apex 1& not visible. The fir8t leaf of
the third nodal eomplex has been removed (rLl) to reveal a young
ataminate flower (A) developing Its inner (Ei ) and outer {Eo}
envelopea. On the opposite .ide of the stem i8 a portion of the
.econd leaf' (L2). A p18tillate flower (G) be10nglng to a still
older nodal complex ia seen in the background. lt i8 already
developing !te stipas' (Si) • .x 146.
Fig. 3. Top view of an uppermost nodal complex whlch Is sllghtly older than
that: in Fig. 1. lta second leaf (L"2) 18 Just Initiating. The .
&xillary meriltem of the second nbdal co~l.x (in the &xil of leaf
L'l) ha. already given ri •• to a floral apex (F) and a lateral
ren.wal apex (V). The outer envelop. (10) of the staminate flower
(A) of tbe thlrd nodal coaplex. 1. overgrowing the young atamen. x 146.
Fig •• 4 and S. Sida viev., lookinl .traight onto ~illary aeriatem
(&a) of upperllOBt nodal complexe ••
Pig. 4. An u1l1ary _dIt .. (aa) about to live ria. to floral and ren.".l . apic... a. tho .. ' Cf and. V) ln th. axil of the fir.t le.f (L' 1) of th.
1laC0Dd DOd.al eoIIPlu: x 146.
/
,/ r
,
o

-/
117
Fig. 5. A stage similar to that of Fig. 4, but enlarged showing that one
portion of the axillary meristem i8 growing faster (F) than the
other that will form the lateral renewal growth (V). x 246.
F~s. 6-9. Side views, each one of three nodal complexes at
different stages of development.
Fig. 6. The uppermost node has just initiâted the firllt (L\) ,and second
(L"2) leaves and the axillary lIleristem (am) in the axi! of the
firat leaf while continuation growth is occurring by means of the
apex of the main axis (Vm). The axillàry meristem of the second
nodal coaplex has already formed floral (F) and lateral renewal
growth (V) apices, while that of the third nodal complex has a
" {oung staminate flover (A) with an oVeTgrowing outer envelope (Eo>
~d a lateral renewal growth (V) with sterile and possible developing
reproductive appendage,S. x 146.
Figa. 7 and 8. Stagea slailar to thoae in Fig_ 6, but on shoota with rlght-
handed rather than left-handed developm8nt. x 146.
Fig. 9. A stag_ vith an axillary meri_te. (am) tbat,un11ke the one_ in Fi~8.
4 and S, 1a not foraing floral and vegetative apicea. ~ 146.
Fig. 10. Side view of a'latera! reneval grovth (older tban the one in Fig. 6)
li,. 12.
that haa 11van rl •• to a floral (P) and a reneval arawth (V) apex.
x 146.
Vl ..... of .t8lÙll&te f1rra at two ata,e. of d.vdopaent. One'
devalop~DI the outer ~nverop. (Bo> and the other vith both outer and
ion.r (11) enyelope. OYerlrow1nl th. younl at ... n. x 146.
Top yi_ of Y~I floral bud (r) 4evelO91nl an uneven gl1'd11nl primor
cil_. x 246.
)
J f f •
!
\ \

118
Fil. 13. Side viev of à youn~ pistillate floral apex (F) tnitiating the
,yoo.cial wall and a lateral reDewal growth apex (V) 'Wh1ch i.
just in1tlatina aterile appendaaea. x 246.
1
, .
, ,
1
"
• \
)
l ,
• •
•
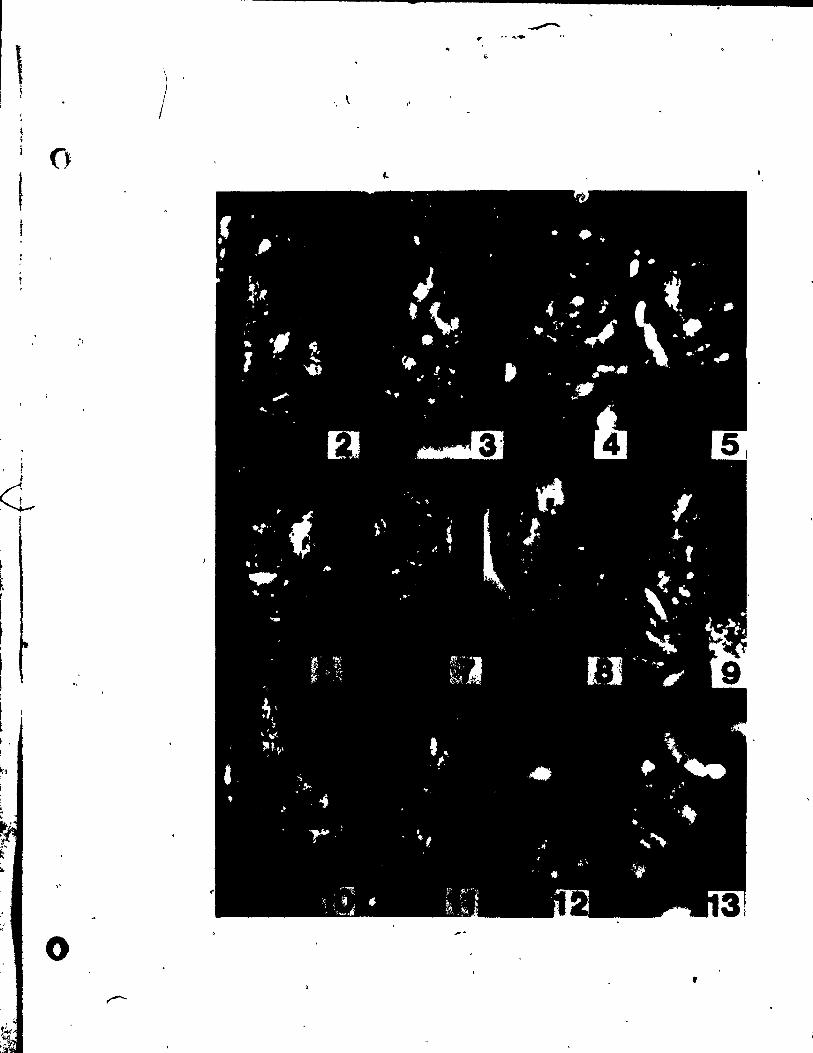
\ t ~ r
t 1 ()
1
, 1
~
" "
.0
-
) " \
,

•
119
PLATE II Figs. 14-22 ..
Fig. 14. Side view of three nodal complexes showing the single stamen (A)
of a' staœinate flower developing its outer envelope (Eo) and thé
single carpel (G) of a pistillate flower in which the ovule (0)
is b.ing enclosed and atigma primordia (Si) are deve1oping. x l4~.
Figs. 15 and 16. Top views of developing staminate f1owers.
Fig. 15. ~e outer envelope (Eo> has three lobes whi1e the inuer envelo~e
(Ei ) is Just initiating abou~' the stamen (A). x 246.
Fig. 16. Sim!lar stage in the development of à stamipate flowèr as that of
r Fig. 14. but showing an outer enve10pe (Eo) w\th lees lob1ng. x 146.
Fig. 11,. Side view of two stages show!ng the development of a pist1l1ate
flower. On the right, a young gynoecium (G) Just enclosing the
ovule (0). On the left, an aIder gynoecium (G) with a developed
'style (Sy) and st1gma (S1). x 146 • •
Fil_. 18-21. Side views of gynoecia, showing the development of
the ovule. f Fig.'18. Young gynoecium (G) at a atagé whe~e its wall ia overgrowing the
incipient ovule (0). Belowand ta the right' 1. a young floral
bud (F). x 146.
Fil_ 19. A portion of the IYDoecial wall ha. ~n removed ta ahow an ovule
primordium (0) juat prior ta the developm8nt of integamenta. x 246.
Fil. 20. The lYDoecial vall .a. entirely removed ta reveàl the young ovule
ahowinl aueellua (H) in1tiatina iuner (li) and ou~er (10) inteau
mente. x 246.
. ..
.J
o

-. ~
120
Fig. 21. Young ovule with nucellus (N) and outer integument (10 ) visible.
r x 246. \ '
~" ' J. -,' Fig. 22. Gynoecium (~) at a stage similar ta that in Fig. 21, shawi~g
stigma (Si) develapm~nt. x 146 .
..
..
, . ,
/
•
a. .~
\l . .', . '
ï. i
ï
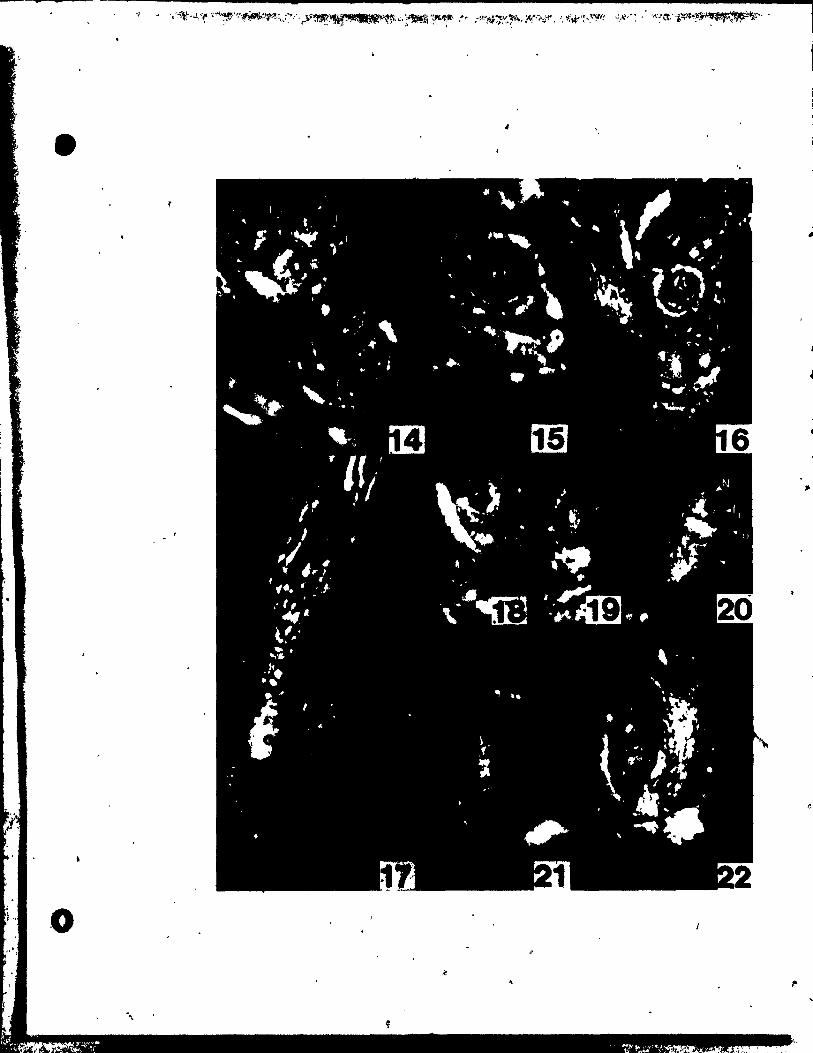
~
1
-,.1
1
"
o ,.
-',

.. • r
, 121
. ,-
PLATE lIt Fia •• 23-26
Fig. 23.
. Fig_. 23-26. Side views of nearl1 mature flowers.
The gynoecium (G) of the pi.tillate flower -to the riaht hal an
already weIl developed style (Sy). The 8tigm4 i8 beyond the
upper portion of the -figure. The yo~nger gyngecium CG) to the
1eft shows both deve10ping style (S1) and atigma (Si). In
between the two piatillate flowera ia a .ingle stamen (A) of a
.taminate flower. The floral and renewal apicas of two nodal
compleXe. (amtin the axi! of Lill and am.in the axU of L'l) are
seen in the &x!l of the second leaf (L2)_of the third nodal
cOlllplex. x 146.
Fil. 2~. A nearly mature staminate flower (A) ta the righttin which the
anther 18 v~81ble through the two transpareat envelopes (Eo and
Ei)' Ta the 1eft i. the gynoecium (G) of • pi.tillate flover
_hoving developed style (S1) and sti ... (Si). x 46.
Fig. 25. Mature pi.tillate flower. x 37. r
F18. ' 26 • Mature nalinate flover. x 46.
-1
, ,
\
•
• •
\
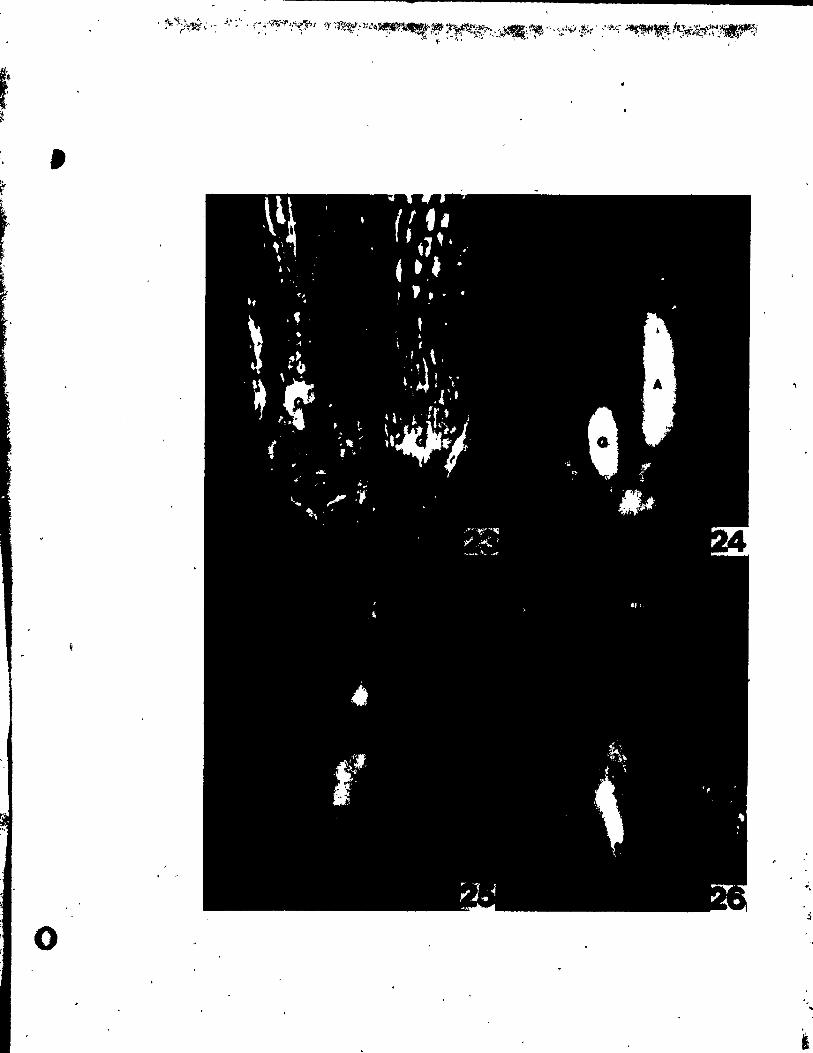
-
,
"
o ..

,
t 1 l ;,
1 /
t 1
f, 1
~.
/
1
182
, ,
PLATE IV . Fig~. 21-3'2
/
1 ./J
/
Fiss. 27-29. He~ian longitudinal sections throush uppermo8t , /
and second nodal complexes. 1
,. .. /
A aection th~fuSh an uppermo8t nodal complex at a staSi/
correapondi~8 to the one in F1&. 2. showing 1nitiati~ of
fint and ,~eond leaves . (LlI~. '\'2)' ax1l1ary meristem (d) and
the rene~~l growth of the main ~iS ~Vm)' ~ portion of t.he
second ~~af (L'2) ~f the second no~al complex can also qe
seen. -ITo the 1eft i8 a tiangential section thiough a young i
gynoe~lU\Jl (G). x 237.
Fig. 28. ~ al~tion simllar to that in Fig. ,1, but at a sHghtly aIder
srge. x 231.
Fig. 29. f mediau section through a shoot tip shovinS a floral apex
/ (F) initiating !ta ~irdlin8 primordiUJll, (arrow) '. x 237.
/
Fill. 30-32. ",dian lonlitud1nal 8ection8 throuJh young
pistillate and ataminate flowera.
Fil. 10. Tva Itamtnate flowera (A) at dlfferertt 8tages of development. ,
Th. one to the riaht i. j\~t initiating the outer envelope
(Bo, labelled only on one 8ide). while the one On the left ia
~ aIre.dy eneloled by the outer eavelope and i8 laitiatina ita
'faner env.lope (Ei • labe\led o~y on one .ide). x 237. \ ' 1
Fi,. 31. YbUDI at ... n (A) of a atamiaate flower dev.lopinS ita outer
" envelope (Bo). x 370.
F1,. 32. GJDOecial vall (G) of a piatillate flower,ju.t be,1nnina to
overlrow cbe tacipient ovule (0). x 370 • . .
/
)
..
o

-
•
o

-
123
PLATE V F1gl. 33-36. •
Figl. 33-36. Median longitudinal·.ec~ions through nearly
mature ataminate and piatill.te flowera.
&
Fig. 33. The inner envelope (Ei) of thi. staminate flower 1a '
beg1nn1ng to enclole the anther (A). x 237.
Fig. 34. The anther (A) nearing a.turity is already covered by both ,
inner (El) and outer (Eo) envelapes. A tangentia! aection
.'
through the uppermost noda~ complex la sean to the left. x 148.
Fia. 35. A young ovule juat pr10r to beeom1ng anatro,ous w1th nueellus
(N) and primordia of both inner (Il) and outer (10 ) integuments.
x 237.
Flg. 36. A section through a pistillate flover nearing maturity. shoving
. the anatropOu8 ovule in which the nucellus (N) has already
formed the embryo sac. x 148.
' .. 1
.",
' •.
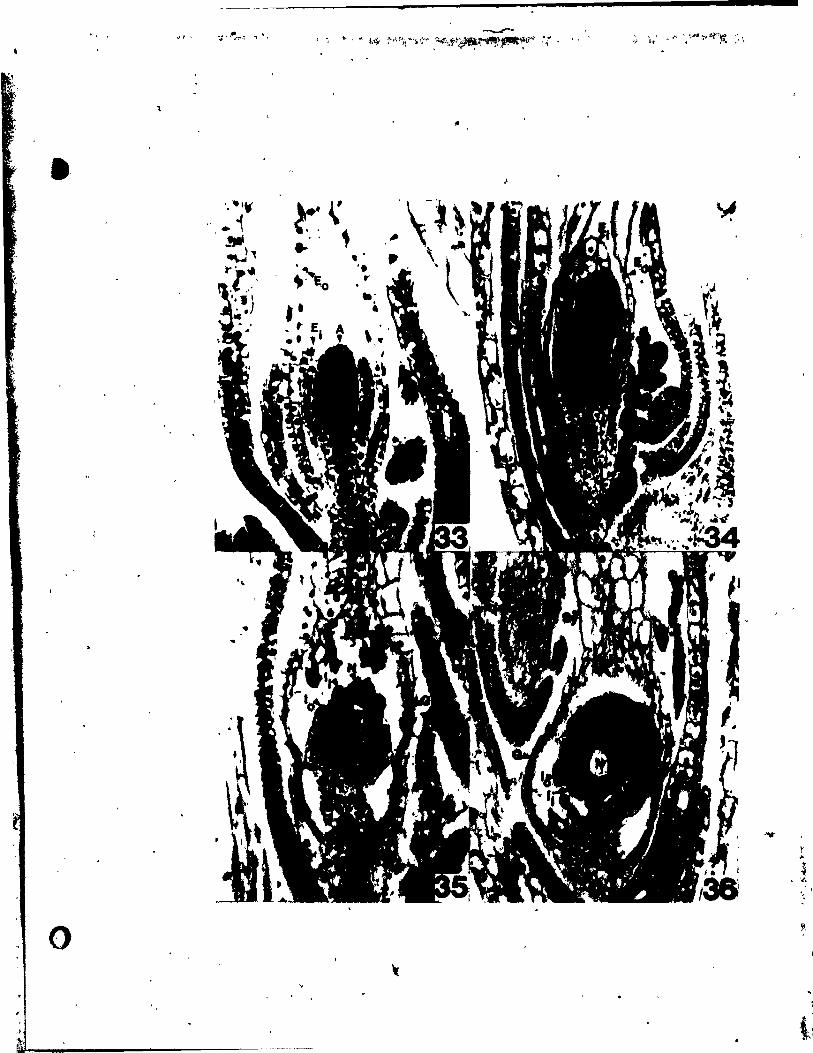
"
0
_f
' •• I~
" . i '. ~, ,.' .. l,.-' . ') , .
"
:~ ~,
, \, ~.
-
~ .. .; ~," , ~~Ei) r
• • _ ,. El .. ,
-'If
~
" $ , '#~
" -,
li,
~ ,
li

124 ~.
LITERATURE CITBD
,.' Banson, L. 1957. Plant cla.sification. Boston.
campbell, p. fi. 1897. A JDOrpho1o.qica1 st-udy of Najas and Zannichellia.
Proe. Calif. Acad. Sci., sere ~, ~:1-6l.
Croizat, L. 1~60. Principia botanica. ·2 Vols. Caracas.
Cutter, E.G. 1965. Recent experimental studies of shoot apex and ahoot
morphoqeneais. -Bot. Rav. ~J7-113.
!ber, E. von. 1934. Karpellbau und Plazentation.verhKltnisse in der
Reihe der Halobiae. Flora 1271273-330.
Heal, W.A. van. 1973. Flowera and fruits in Flaoourtiaceaa I. •
Scaphocalyx spathacea Rid1. B1umeà 31:259-279 .
• Lawrence, G.H.M. 1951. Taxonomy of vascular planta. New York.
Macdonald, A.D. 1974. Floral development of Comptonis peregrina
(Myricaceaa). CaJ1. J. BOt: 52:2165-2169 • ..
_____ ' and R. Sattler. 1973. Floral development .of !'lYrics sale and
the controve:ny over flo~al concepts. Cano J. Bot. 5l,:1965-1915.
Magnu., 'P. 1969. Zur IIOrphologi. der ~tW\q Najas L. Bot. Zeit. 271769-
1773.
1811. Beitrag. au JteMtn1 •• 4Br Gattun9 Naja.. hr1in.
. ; 1894. Uber 41. G&ttunf "jas. 'JIU. d. 4e\JUch. bot.' -1\
O' ... l1.ab. .!:!. 214-224.
" "

•
v 1
1~
Meeuae, A.D.J. 1966. Fund .. entals of phytomorpho1ogy. The Ronald
Press Co. t New York •
., .... Meyen, S.V. 1973. Plant morphology in lts ndmothetlcal aspecta.
Bot. Rev. 39(3):205-260. ,.-
Miki, S. 1937. The origln of Najas and Potamoaeton. Bot. Mag. (Tokyo),
Po.luazny, U. and R. Sattler. 1973. Floral deve10pment of Fot8aOgeton
denlul. Can. J. Bot. 51:647-656.
1974a. Floral developaent of Potamoleton richardsonii. Amer.
J. Bot. 61(2):209-216.
1974b. Floral developaent of Ruppia maritima var. maritima.
Cano J. Bot. 52:1607-1612.
1973.' Floral dev_lopa_nt of Zannlche1lia paluler1a. (In press).
'-landl., A.l. 1899. A ayatematic reviaion of the I_nua Naja.. Trans. ,
\
1 Linn. Soc. Sere 2, Bot. 1~379-444.
1930. The clalsification of flowerinl planta. Vol. 1 • ...
Monocotyledona. 2ad .d. Caabrid,_ Univeraity Pre.a, Cambridge.
1
S.t&i, W.B. 1973. aiaple .. thad for difterential Italnina of paraffin l'
1 .. bldded plant .. tarial ullng To1uidlue Blue O. Stain Technol.
.!!oU) :247-249.
--.. .... -----.....
I.tt~.r. 1. 1968. -~~bnlque for the atudy of floral davelopaent.
CaA. J. lot. 461720-722. -,1
/

126
Sattler, R. 1973. Organogenesia of flowera. A photographie text-atla ••
Univeraity of Toronto Press, Toronto.
, . 1974&. A ne'" conception of the .hoot of hiqher planta.
J. Theor. Biol. ~:367-3B2.
1(.
1974b. A ne." approach to 9YJloecial lDOrphology. PhytOll'Orphology
~:22-34.
_____ " and E.M. Gifford, Jr. 1967. Ontogetletic: and hi.tochaaical
ohanqe8 in the shoot tip ot Najas pdalupensi. (Sprengel) Marang.
Phyto~rphology !l:419·428. •
Schumann, K. 1892. Morphologisch Studien, I. Leipzig.
Sculthorpe, C.D. 1967. The biology of aquatic vascular plants. E4ward
Arnold (Pu~li.her8) Ltd. London.
Singh, V. 1965. Morphologioal and anatoaica1 atudie. in Helobiae. III.
Vascular anatomy of the node and flover of Najadaceae. ~roc. Indian
Acad. sei., SerY. B. ~t98-108.
s._y, I.G.L., and ~.lt. Lak.haanan. 1962. Contributions to the embryology
ot the Najadaceae. J. IncUan Bot. Soc. !!..247-267 •
. ~ 'l'akht;ajan, A. 1969. !'lonring plàJltat oriQin and dispersal. Oliver and
•
, Boyd, Bdinl>urgh. (Tranalated troll Rua.lan by C. Jettrey~)
Ohl, If •• ,' 1947. Studie. in the noral MOrpbOlOfY and anatomy of certain
MllbeZ's of the Halo!)1&.. Ph.D. 'J'IÎes1a, Comell \l\ivenity, Ithaoa, R.Y.
I~,.. 1959. Dle PhrlogenJ.e au .11...... Jena, 't\ltqutl
G. riltClbeZ' V_la9.-
-
(

-
( 1
127
. ;/
OONCLUSIONS
..
• • ,
,1
j-j:,_.~, MI" , ln. j .... , ..
,~L~"l.Îl~ .. j~.,,~ ,_ ,"( ..
. , j " :"'. i
::::~. ~~;, :.l~',;.k,,: '.1

" l t' ." .~
'"
128
The floral developmental observations in this thesls helped clarify
the following morphologieal and taxonomie prob1ems involving the five
spec1es studied. ,f
1. lt was shawn that the deve10pment of the perianth members (tepals)
in Potamoseton richardsonii and f. densus is acropetal, supparting the
previou8 floral developmental work o'f Sattler (1965). and refuting the ides
that these appendages are outgrowths of the stamen connective. The position
of these tepals in the mature flower ls due ta interprimordial and inter- '
calary growth at the base of the young tepals and stamens.
2. The flower of Potamogeton densue, though resembling that of
f. rlchardsonii at maturity, demon8trat~d several unique developmental
features. The lower Median tepal and stamen are In1tiated at a tlme when
the floral meristem 1s nct yet clearly demarcated. This was interpreted 8S
the spatial and temporal superimpositian of a bract-axillary bud conf1gur-
ation and the deve10pment of a flover. With regar~ ta the f1ower-inflores-
cence controversy, this observation 8upported the ideas of Emberger (1950)
Who recognized intermedlates between "inflorescence" and "flower". As
weIl, !n ~n8u8. the lateral st .. ens each initiated 88 tvo separa te
primordia, len4ing support to the conclusion that the stamen i8 partially
homologoua to two atamen8, which Kiki (19\7) elaimed weie redueed flowers
. on the .talk of a bract (tepal).
3. A comparison of the development of P,' den.ua and that of the
diatantly ralated .pecie.!. richard.on!! and!. cri.pua, 8cipported to
.a.e extant the aeparation of !. den. ua a. a aonotyplc ganu., Groenlandia.
4. tt wae de.onstrated that the membranoua abaeth about ~he mature ... é'
upper ,floral bud ln luppia .. titilla :$.nftiated a. a prophyll on the adaia!
,/j l, ~ 1
• '1
..

, l'
"
,1 129
flank of the renewal growth apex tmmediately below th& upper floral bud.
During its detvelopment it appeau to be "captured" by the rapidly
developing inflorescence axis.
5. the development of the controverslal outgrowth of the stamen
connective in !. maritime suggested no homology with the tepai of the
flower of Potamogeton. Unlike.a tepal or a bract it initiates after
" stamen inceptioq, in fact after the beginning of theta differentiation. ~
ln general, lt is similar to a vertical exension of the stamen connective
in Po tamoge ton .
6. Though floral development study does not bring Ruppia and
Potaaoseton closer together. it does fit !. maritime into a morphological
series between Potamoseton and Zannichellia as postulated by'Singh (1965).
Whether this series goes from complex ta simple or vice versa cannot be
ascertained without clear chronological data.
./
7. Regarding the development of the flower of Zannich~llis palustris,
a number of miainterpretations by Campbell (1897) were clarified. Firstly,
it was clearly shawn that the staminate flower terminated an axis with a
reneval gro~th apex continuing in the axil of a sterile sppendage below;
and secondly, it was shown that the carpel first develops as a peltate
primordium (aimilar to Potamoseton and Ruppia) giving rise to the ovule
aecondarily on the adaxial portion of the carpel wall. ,
8. The envelope .ur~ounding the piatillate flover in Z. palustris was
shawn to be ·pr~ .. rl1y p~otod.rmal ln origin, much like the integuments of
an oVule. Yet, Its po.ition and inception were like that of a phYllome • ... lt .. y be of a etatua 1ntermediate between a phylloae and an emergence.
9. The rUlOn. gi ven for tendng each carpel of Zannichellia a8 a
•• parate pi.tillate flower vére not 8upported. They do not arise in spiral
, '
•
"

-t
=
130
succession as Singh (1965) claimed, nor are they part of a dichotomous
branching sys~em as proposed by Campbell (1897). Jhe two carpel
primordia are initiated almost simu1taneously on the fl!nks of the
pistillate floral ape~.
10. Due to the compressed nature of the developing nodal complexer of
k. palustris, it was impossible to establish beyond dou6t whether the
terminal meristem gives rise to the renewal growth apex and the pistillate
floral bud through dichotomy (Campbell, 1897) or whethèr the pistillate
floral bud terminates the shoot with the main renew~l growth continuing
laterally (Schumann, 1892). Developmental evidence, though, tended to
support th~ latter view.
Il. Th~ development of the single stamen of the staminate flower of
k. palus tris did not lend support ta those (Eames, 1961; Singh, 1965)
who believe that it may represent the cannat ion of two anthers. As in
~. ~ichardsonii and R. maritima the stamen first develops as an elo~gate
primordium-wh{ch differentiates its thecae at the distal portions.
12. A comparison of ~. palustris with f. richardsonii, f. densus and
~. maritime, revealed sorne sirnilar1ties (i.e: gynoecial development and
sympodial pattern of fertile branches) but there were also sorne striking /,
differences, most notable being the separation of sexes in Zannichellia . .
This did not necessarily negate a link between Ruppia and Zannichellia,
but on the basis of our comparative observations we could see sorne evidence
for assigning Z'annichellia ta a separate fâmily.
13. In their de~elopment and form, the two envelopes about the staminate
flower in BA1&a fl~x11is resemble very much the gyn8ecial wal~nd outer \
inte~ument 11 th~ pistill~tT f1ower. This resemblance would seem t~ indicate
that during the inception and'development of the envelopeg around the stamen
,

•
primordium, the aame (or aimilar) ~orpho8enetic factors are active as
ln the envelopes around the nucel1us.
14. As ln other cases vhere the ovule la initiated in an exactly
te~inal (basal) position (e.g., Myricaceae), the gynoeclym of Najas
was ~learly shown to be acarpellate (as defined by Battler, 1974). \
131
15\. Some questionable points in the literature were clarified regard-
in8 the pistl11ate fraver of Najas. No annular swe1ling was seen to
develop At the Iower part of the 8tigma as Miki (1937) claimed; and
çontrary ta Campbell's (1897) de8c~lptlon. the integuments deve10ped ln
acropetal sequence wlth a portion of the outer integument becomlng so
clos.ly adpressed to the funiele at rnaturity that it was difficuit to
dilt:l.nguish.
16. In comparing the floral development of li. flexl1is vith those of
~. richardsonii, 1. densul, !. marit!ma and!. palu.tris, numerous
dif~.rences were.noted; ln partlcular. the branehing pattern of the fertile
Ihoot, the development of the androe~lum and synoeclum and t~e two envelopes
about the staminate flower (see, Fig. l, Table 1). Najas, tberefore,
app •• ~ed distinct'w1thin the Najadales even with respect to Zanniehellia • .tI
Untll po.a1ble l1nks'.with other Najadales (e.g. Althen1a) ean be more
thorou,hly .tudied, ·it se .. adviaablè to keep Najas withln a separate
family, the Najadace •• , vhich along w~th the Potamogetonaceae (Pot&mo8!ton,
iUIIP") and~th. Zanniehelliaceae (Zannicbell1a) would be pl.cad within one / ',\,
- order.
17. Vith relard to th. re1.Uon.h:l.p. betw.an the' ·fresh and brackieh
vat.r W.j.dal •• , lit. tho ••• tudi.d in thi. the.i., and the .. rine i.jadale. , . (i ••• Zo.tar.e .... ~ddoui.e ••• t Cyaocloceaeea.) ~J tvo line. of evolution
"
~ • , .

, f
/ J
1
-
132
have been suggested. ArboT (1920), suggeeted that the marine species
derlved from freshwater species with the salt-tolerant and bracklsh
water speciés being Intermedlate. Den Rartog (1970), on the other
hand sU8gested t~ both freshwater and marine specles evolved In-
dep~ndently fram salt-tolerant ~hrub-llke terre. trials with sympodial
rhizomes and cymose inflorescence (see Fig. 2). He, a1so noted some
fossil evidence to back his clalm. The resu1ts in thls thesis tend to
support Den Hartog's proposaI more than Arber's. As weIl, a possible
reason for the gap in the morphological series between Ruppla and
Zappichellia could be explalned by the faet that they originated indep-
énde~tly (Fig. 2).
18. Observations' in thie theeis supportèd propoeed links between the
" Najadales and the Hydrocharitales (Sc:haffner, 1904; Miki, 1937). Many
apacies within the Hydrocharitaceae, for exemple, 'exhibit eimilar
patterns of inflorescence and aXlll,ary bud development (Cutter " 1964;
Charlton and Ahmad; 1972). Also, Kaul (1970) and Sculthorpe (1967)
diacusaed a Biailar aer!es of morphologieal modifications in the
inflorescences of the Hydrocharitaceae,whieh like the one in tqe Najadales,
~lves separation of aexea. modification of a complex aympodial aystem
and reduction ln part ••. The morpholoalca1 leriea deacrib.d.in th1.
th.li., therefore. 1. not an laolated occurrence but part of a pattern
of eventa 1nvolving aquat1c plante goin, from anemophiloua ~o hydroph11ou8
--pollination orv1ce ~r...
,
"

133
-
Fig. 1. Comparison of the diagrammat1c represéntations of
. .
..
.. ~ ..
the inflorescences and flowers of, a) Potamogeton
ri,hardloni!, b) potamogetOn densus, c) Rupp1a
mar!ttœA, d) ZAnn1chellia palustr1s, and e) Najas
flex111 •.
, ~ ..
, .
. "
. . , ,·L'. .,. .',' ~ i'I "
: ~::,.;,,'~~~~ __ .~r"':,.· _~. . ' '.~ :;;} .... ~ ~ .. .. '
" .f;.. J..9fAt/..< ~.
t" .. r dO._'" .>f.: . ,. .' ,"~
..
.. .
• 1
1
.1
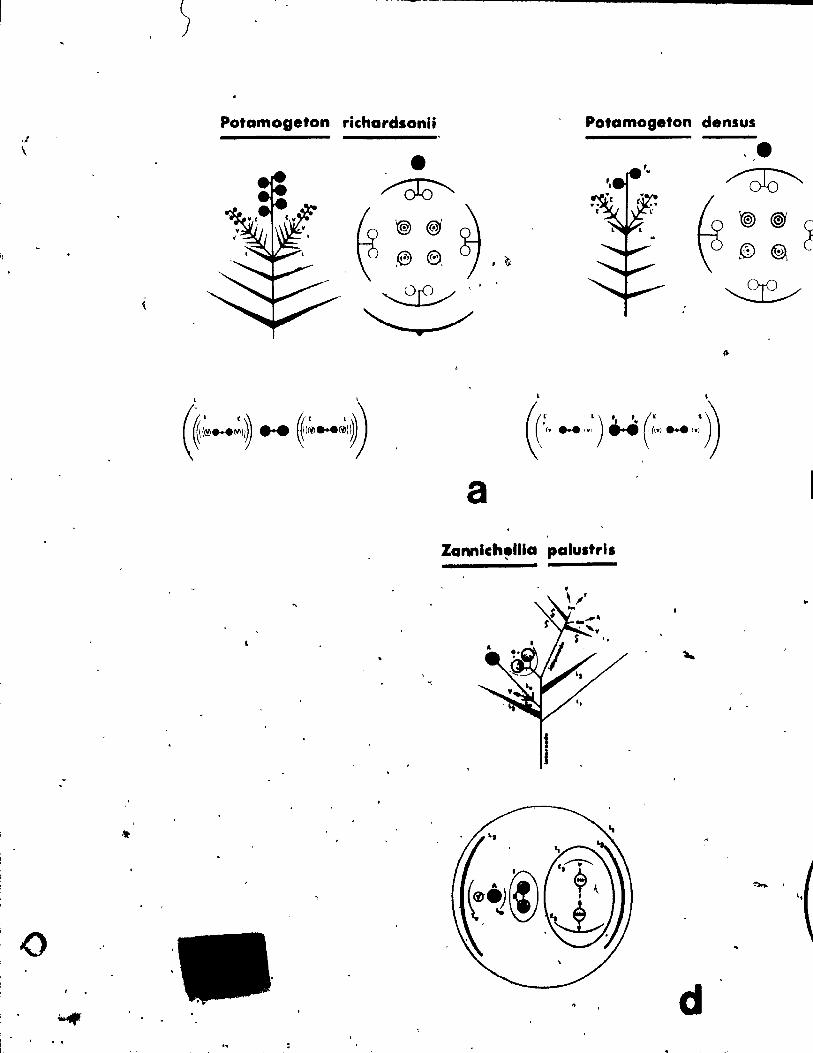
Potamoseton richardsonii Potamogeton densus ·: \
L L
ft(r L)" (r ~)~ ~,'" ... '" "',""''''''' ~
a Zannfch!tllla palultrls
..
1
~ .
, . d . .,
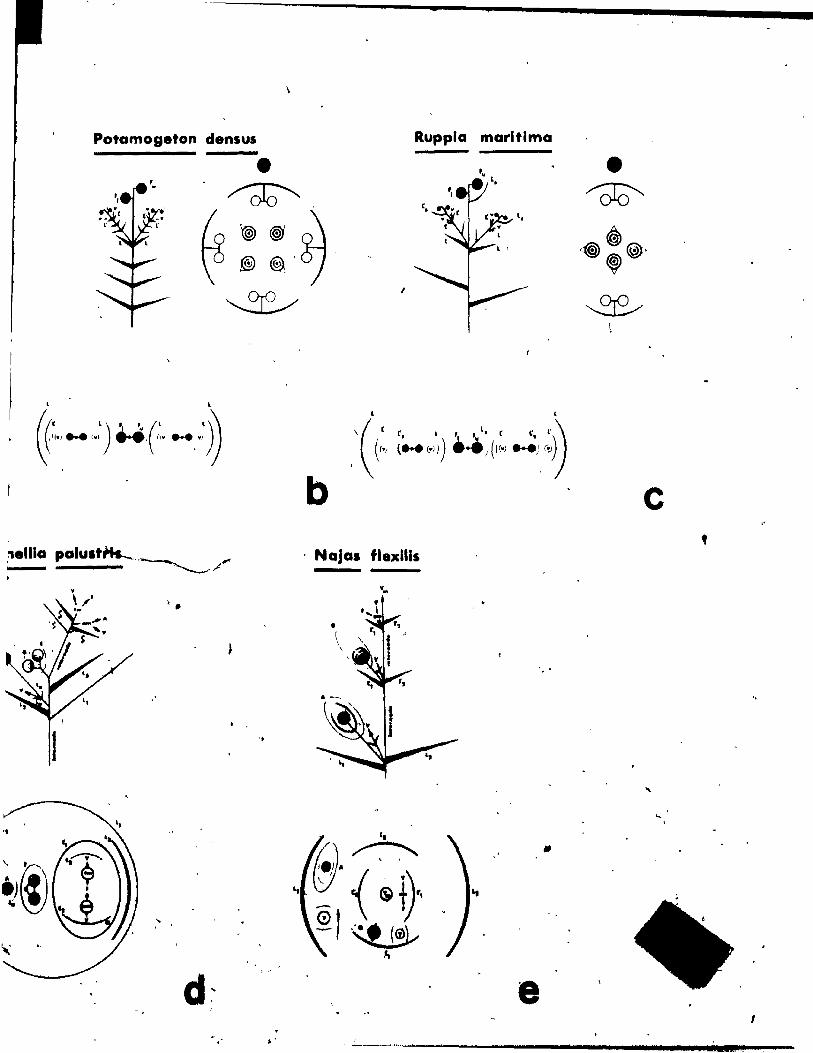
Potamogeton densus
, ,
((fr ')" (' f)~ ~ •• ''' ... '" ..... O· ... :" ')
" .
1 '.
. . '
•
Ruppla marltlma
1
, '
,'( (:, ;;...,:)) .~:(I:'> ";::'»)
b c , Najas fl.xlli,
•
e 1
" ___ ~ ___________________________ d.lE ____ --
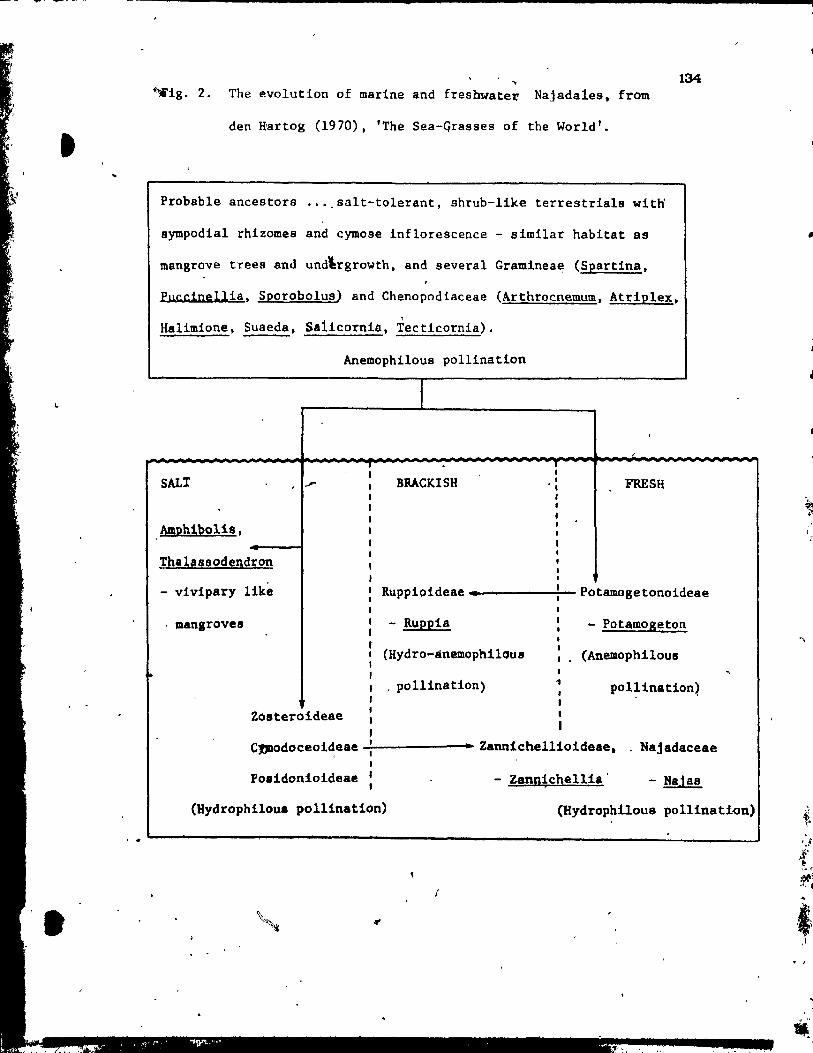
~ig. 2. The evolution of marine and freshwater Najadales, from
den Hartog (1970), 'The Sea-Grasses of the World'.
Probable ancestors .•. _salt-tolerant, shrub-like terres trials witH
sympodial rhizomes and cymose inflorescence - similar habitat as
mangrove trees and und\rgrowth, and several Gramineae (Spartina,
Pucdnellia, Sporobolus) and Chenopodiaceae (Arthrocnemum, Atriplex.
Halimione, Suaeda, Sa1icornia, Tecticornla).
Anemophilous pollination
1
SALI SRACKtSH FRESH
Aml>b1bol1s,
Thahssodendron
134
- vi vipary like Ruppioideae~.~--------~- Potamogetonoideae
mangroves
1 ,
- Ruppia
(Hydro-anemophilous
- Potamogeton
(Anemophilous
, . pollination) , 1 , po 11 ination)
Zosteroideae t , 1
1 1
1
CJJDodoceoi~eae ...:::..------•• Zannlchellioideae •. Najadaceae J
Poaidonioldeae 1 1
(Hydrophilou8 pollinatlon)
- Zannichellia - Najas
(Hydrophilou8 pollination)
. . L-----------------------------------________________________________ ~ ________ _J
...
1
1
l 1
~
, r
. , 1
.1

-'
+
\
TABLE l. Comprehensive comparison of morpho1ogical and developmenta1 features
Potamogeton richardsonii, Potamogeton densus, Ruppia maritima, ~ , .
POLLlNATION
DEVELOPMENT OF FERTILE BRANCHES
\,
P. t'ichardsonii P. densus g,. maritima
Anemophilous Anemo ph ilou s Hydro -anemophi 10u:
Sympodia1 branching pattern. - Inflorescence terminates the shoot renewal growth apices in axils of the last vegetative appendage' grow vegetatively before themselves terminating as Infiorescencl Leavea. and inflorescences arranged è1istichously.
STERILE APPENDAGES 8-20 6-15 4 BETWEEN INFLORESCENCES
FLGlERS PER INFLORESCENCE ARRANGEMENT MiD DEVELOPHENT
SHBATHS OR ENVBLO~
..
"
9-27 flowers inltiated ln whorls,of 3. Each flower is subtended by a bract.
NoM
.,i
2 flowers initiated on the inflorescence axis, in subopposite arrangement
b--
na bract
•
no bract
.,.. l ~heath partla11y
envelops the upp floral bud (Fu)' Develops initial aB a prophy.l~ of renewal growth· a below the hflor' cence.
" .
J.A.I"'~ __ "&!V_~_~ _______ -""" ---------------

•
.. 13~
} of morphological and developmental features of the flowers of J
\ l, Potamogeton densus, ~uppla marltima, Zannlcheilla palustris, and Najas flexills
P. densus ~. maritima
Anemophilous Hydro-anemophilous
pattern. - Inflorescence terminates the shoot; ces in axils of the last ve~etative appendages before themselves terminating as inflorescences. scences arranged distichously.
:ed Ich ~d
6-15 4
2 flowers initiated on the inflorescence axis, in subopposite arrangement
no bract
None
no bract
1 sheath partially envelops the upper floral bud (Fu>,' Developa inltially as a prophyll of the renewal growth apex below the inflorescence.
"
Z. palustris N. flexilis
Hydrophllous , Hydrophilous
Symp~odial (possibly Monopodial intermediate between sympodial and ~ono-podial with regard to pistillate flower)
Each successive nodal complex ia formed at almost 900 from the one below.
3
1 9taminate flower i9 inltiate in the axil of the 2nd appenrlage (L2) of the node.
1 pistillate flower formed by division of terminal merlstem (tmY.
Both staminate and plstillate flowers appeat axl11ary at 1D8: turity.
l envelope about the pistl11ate flower. It ls initiated at ovule inceptLon, below one of the carpel.. Origin lé primarl1y protodermal on one side of the pedunele, whlch It later ,irdles.
f
2
Either l 8ta~inate
or 1 pistillate flower ls formed ln the axl1 of/the Iower (LI) of two subopposite leaves at a -Rode.
2 envelopes about the staminate flower. They .develop acrop'etally a8 girdling primordia, ~r1marily protodermal in ori81n. The outer envelope (Eo) 18 100.e wlth teeth-like projections at it. apex; the Inner envelope ' (11) i. bilobed and elolelY'adpre'led to the an.ther.
...

o
/
TABLE 1. (cont'd)
PERIANTH MEMBERS (TEPALS)
ANDROECIUM
OUTGROWTH OR EXTENSIONS OF THE STAMEN CONNECTIVE
GYNOECIUM
OUTCltCJWTHS OF CARPEL WALL
\
!. richardsonii
4 tepals are initiated l'he 2 later~1 tepals
~ precede the 2 median tepais.
p, densus
in acropetai sequence. The Iower med,ian tepai
initiates first, followed by the 2 la'terai tepals and che upper median tepal.
4 stamens are init1ated opposite the tepals. ALI the stamen The 2 laterai stamens
primordia are init- each initia~e as 2 iated as elongate &eparate primordia, primordia. The 2 . foilowed by the 2 lateral stamens pre- median s""tame~ which cede the 2 median initiate as elongate atamens. primordia.
Slight vertical extension of stamen • connective. Late in development.
Vertical extensions of stamen connective. Late in development. In some cases quite prominent.
4~carpels are init1ated, alternating with the stamens.
R. maritima
None
2 median !ltamcn., are initiated 0
floral' apex, ea elonaate primor
---J
Dorsiventral outg of the- starncn connectiv~ meri aqc origin. Initiates just , s tamen begins t~
dUferentiatc tl
Eacb car pel forms as a radial pr1mordium, whiçh at the crosi-zone (Quétzone).
2 lateral and 2 m. carpel prirnordi. initiated.
becomes Ipeltate l
Sllght outgrowth on upper abaxial carpel 1lall. Late in dévelopment.
rl"
"
In some case.s, extensive~outgrowths on upper abAleial car!fl wall.
. ~t~1n deve1opment.
. ).
...
At maturity fruit elong8us •
SI1ght outgrowth .( upper abaxial C~
wall. Late in development •

P. densus R. maritima
in acropetal sequence. Non~
The lower median tepal initiates first, followed by the 2 lateral tepals and the upper median tepal.
èd opposite the tepaIs. The 2 lateral &~amens
each initiate as 2 separate' primordia, followed by the 2 median stamens which initiate as elongate prlmordia,
Vertical extensions of stamen c~ctive. Late in development. In some cases quite promi.nent.
2 median stamens ~are initiated on the floral apex, each as
. elongate primordia.
Dorsiventral outgrowth of the stamen connective meristematic origin. Initiates Just after stamen begins tb differentiate thècae.
~. paIustris
'II None
138
N. flexilis
None
1 terminal stamen. forms from ~he transformed stamlnate floral apex.
Early stages similar Anther forms only ,1 to Pota~geton and cell . Ruppia, elongate primordium different-iates staminal the~ae at distal portions.
Slight vertical extension of stamen connective~ Late in devèlopment.
None
!d, alternating with the 2 lateral and 2 median 2 carpel primordia form No carpel, gynoecial
1 radial primordium, which !uerzone) .
ln some ca ••• , extensive outgrowths on upper abaxial carpel wall. Late in development.
car pel primordia are on the pistillate wall initiates as a initiated. .. floral apex. girdHng prim"ordium
becomes peltate and initiates a 8ingl~ ovule which encloses the ,
At maturity fruit stalk elongates.
Slight outgrowth on upper abaxial çarpel wall. Late in development.
None
\
-'
pistillate floral apex which later transforms directly into the ovule.
None

, l , Il 1 ~
\
o
TABLE 1. (cont'd)
VARIATIONS •
J {
,
i
, .
."
f. richardsonU
Most variations occur close to inflorescence apex. Most common variations inelude trimerous and dimerous gynoecia wlth tetraMerous stamens and tepals, completely trimerous flowers, upgrowth between adjacent tepals.
,.
f. densus
A grest amount of variation occurred including completely trimerous flowers, trimerous gynoecia with tetramerous androecium and perianth, and organs 'd1fferent1at1~g
. partially 1 as tepals and partially as stamens.
,
.. .
B.. mar!tima
'" Somé variation oc
éd including tr merous and pent ous gynoecia, Il
ed staminal the bifurcated stam connective.
/ .

f. densus 3
A great amount of variation occurred including completely trimerous flowers, trimerous gynoecia with tetramerous androecium and perianth, and organs differentiating partially as tepals and partially as starnens.
R. maritima
Sorne variation occurred including trimerous and pentamerous gynoecia, aborted staminal theca, .. bifureated staminal connective.
.'
.~.
Z. palustris
Very little variation. Some stamens with unisporangiate thecae, gynoecia with t~ree rather than two carpels .
•
. , J '
•
137
!. flexlUs
Very l'ittle variation in development of flowers.
. ,
r
;;;
, .. . ' ,

c
".
..
138
LITERATURE CITED
Arber, A. 1920. Water plants. University Press, Ca~brtdge.
Campbel!, ,D.H. 1~7. A morphologieal study of Najas and Zanniehelli.a. I!" ,
Proe. Calif. Acad. Se1. ser. 3, !.: 1-61.
Charlton, W.A. and A. Ahaad. 1973. Studies in the Al1smataceae.
IV. Developmental morphology o~ Ranalisma huml1e and comparisons
with two membera of the Butomaceae, Hydrocleis nymphoides, and
Butomua umbellatuB. Cano J. Bot. 51: 899-910. r-
Cutter, E.~964. Observations on leaf and bud foraati9n in
Hydroeharls morBus-ranae. Amer. J. ~t. 51(3): 318-324.
Eamel, A.J. 1961. Morphology of the angio.perms. KeGraw-Hi11 Book
Co. r New York. "
EabeJiger, L. 1950. La valeur morphologique et l'origine .de la fleur: '"
CA propol d'une thiorie nouvelle). Ann'e Biol. '(Paria), 26:
279-296.
Bartol, C. den. 1970. Th ••• --Ir ••••• of the world. Verh.ndelingen l' ."
f
d.r ~niDkl1jke .eder1.nd •• Akad_ie van We"tenechap-p-en, Afd. Natu,urkude.
Tweedl Ieek., ne.l 59(1): 1-275. ,-
lau1. a.B. 1970. Evolution And .d.pt.tion of inflor •• cencee in th.
Hydrocharit.c.... Mer. J. Bot • .21.(6): 708-715. /
\ Miki,' 8. 1937. The oriltn of •• j •• and Potaao •• too. Bot. Mal. Tokyo;
. .as 472-480.
\.
1.
. ."
Il

, r
,~
\
Sattler, ~. 1965. Perianth development of Potamoaeton riehard.onii.
Amer. J. Bot. 52(1): 35-4J.
1974. A nev approaeh to gynoeeial morphology. Phytomorphology
24: 22-34.
Sehaffner, J.H. l~04. Soae morphologiea1 peculiaritle. of 'the
Nymphaeaceae and Heloblae. Ohio Nat. ·i= 83-92 •.
~
Schumann, K. 1892. Morphologllche Studlen. Leipzig. 1
Seulthorpe, C.D. 1967. The blology of aquatle vaeeular planta;
Edtfard Arnold (Publiehet.)' Ltd. London.
Singh, V. 1965.~ Morphologiea! etudlel'in Re10biae II.
Va.eula~ anatomy of the ~lower of Pot g.tonac.... Bot. Gaz.
126(2): 137~l44.
)




















