
Gene set enrichment analysis of the NF-κB/Snail/YY1/RKIP circuitry in multiple myeloma

alaeoecology 254 (2007) 550–560www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Stable isotope composition of Late Glacial land snail shells fromGrotta del Romito (Southern Italy): Palaeoclimatic implications
Andre Carlo Colonese a, Giovanni Zanchetta b,⁎, Anthony E. Fallick c,Fabio Martini d, Giuseppe Manganelli e, Domenico Lo Vetro d
a Dipartimento di Archeologia e Storia delle Arti, Sezione di Preistoria, University of Siena, via delle Cerchia, 5, Italyb Dipartimento di Scienze della Terra, University of Pisa, via S. Maria, 53, Italy
c Scottish Universities Environmental Research Centre, East Kilbride G75 0QF, UKd Dipartimento di Scienze dell'Antichità “G. Pasquali”, University of Firenze, Piazza Brunelleschi 4, 50121 Firenze, Italy
e Dipartimento di Scienze Ambientali, Università di Siena, via P.A. Mattioli 4, 53100 Siena, Italy
Received 3 November 2006; received in revised form 29 June 2007; accepted 4 July 2007
Abstract
Stable isotope composition of living and fossil land snail shells was determined at Grotta del Romito (Southern Italy) with theaim to reconstruct environmental and climatic variation in the area during Late Upper Palaeolithic. The investigated successioncomprised 15 different excavated layers spanning between ca 13,000 and 14,500 yr cal BP. The oxygen isotope composition ofsnail shells indicates a marked decrease at the layer D8 suggesting a climatic deterioration consistent with the GI 1d climatic event(Older Dryas). This climate deterioration may have been related to a substantial decrease of mean annual temperature withassociated changes in the regional atmospheric circulation. However, the environmental conditions at the time of shell's growth inthe other intervals sampled suggest condition comparable to the present day. The carbon isotope composition of fossil snail shells isin agreement with other records, which indicate a general increase of the δ13C values of organic matter during Pleniglacial to LateGlacial caused by substantially lower atmospheric CO2 concentration at that time.© 2007 Elsevier B.V. All rights reserved.
Keywords: Land snail shells; Stable isotopes; Late Glacial; Palaeoclimate; Southern Italy
1. Introduction
The 13C/12C and 18O/16O ratios of land snail shells aregoverned by a wide number of physiological and envi-ronmental factors which make it difficult to obtain asimple quantitative reconstruction of the past climateparameters by fossil shells (e.g. Lécolle, 1985; Goodfriendet al., 1989; Goodfriend and Ellis, 2002; Balakrishnan andYapp, 2004; Zanchetta et al., 2005). Specifically, the
⁎ Corresponding author.E-mail address: [email protected] (G. Zanchetta).
0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2007.07.005
18O/16O ratio is related to the isotopic composition ofenvironmental water ingested by snails (e.g. water vapour,dew, local meteoric precipitations, e.g. Goodfriend et al.,1989; Lécolle, 1985; Zanchetta et al., 2005), an isotopiceffect linked to the exchange of fluid between the externalenvironment (through the body of the snails) and internalfluid (Balakrishnan and Yapp, 2004), relative humidity(Yapp, 1979; Balakrishnan and Yapp, 2004) and thetemperature of carbonate precipitation. Different directcorrelations between oxygen isotope composition ofmeteoric water and oxygen isotope composition of landsnail shells have been found within living populations

551A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
(Lécolle, 1985; Goodfriend and Ellis, 2002; Zanchettaet al., 2005), but in very arid lands this correlation is oftennot apparent (Goodfriend et al., 1989). Probably the mostcomplete model to interpret oxygen isotope compositionof land snail shells is that proposed by Balakrishnan andYapp (2004). However, quantitative prediction from thismodel involves several assumptions which complicate itsapplicability to past samples (Balakrishnan et al., 2005a).An additional issue is related to the large oxygen isotopevariability usually measured both on living and fossilpopulations (Goodfriend et al., 1989; Leone et al., 2000;Goodfriend and Ellis, 2002; Balakrishnan et al., 2005a,b).If, for fossil remains, this variability can partially beexplained by accepting that fossils assemblages accumu-lated during a certain interval of time (and so shells canactually record variability in the climatic parametersduring time), for living populations this spread is probablydue to very localised variability of local environmentalconditions (e.g. temperature, moisture) during the life ofthe snails. Furthermore, different species collected in thesame locality can have different 18O/16O ratios (e.g.Lécolle, 1985; Balakrishnan et al., 2005b) but the wideisotopic variability found in modern populations can hidesmall vital offsets (Zanchetta et al., 2005).
The carbon isotope composition has mainly beenrelated to the isotopic composition of the ingested food(e.g. Stott, 2002; Goodfriend and Ellis, 2002;Metref et al.,2003; Balakrishnan and Yapp, 2004). This depends on thefact that land snails incorporate more CO2 throughrespiration than CO2 from the surrounding environment(McConnaughey et al., 1997). However, ingestion ofcarbonates (Goodfriend and Hood, 1983; Yates et al.,2002), exchange with atmospheric or soil CO2, andmetabolic rate (Balakrishnan and Yapp, 2004) maycomplicate the picture. Scrutiny of the data available inliterature suggests that the carbon isotope composition ofsnail shells is more species-dependent than for 18O/16O(e.g. Yapp, 1979; Lécolle, 1983, 1984; Yates et al., 2002;Zanchetta et al., 2004, 2005).
Moreover, the land snails adjust the time of shellgrowth during the most favourable condition avoidingboth too high and too low temperature, and in function onthe local humidity (Balakrishnan and Yapp, 2004), so thatthe isotopic composition of their shells is not strictlygoverned by the “average” local environmental conditionsand other factors like seasonality can have an importantrole (e.g. Leng et al., 1998). In conclusion one could arguethat the use of isotopic shell composition to infer pastclimatic change can be partially hidden by the largeisotopic variability we can find on past assemblages orthrough the effect of smoothing of the climatic signalregulated by physiological requirement. Despite these
limitations, land snail shells are commonly preserved incontinental deposit of many regions and their isotopicstudy can represent an additional important source ofinformation for the understanding of the past environ-ments (e.g. Goodfriend and Ellis, 2000; Leone et al.,2000).Moreover, most species of land snails present in theQuaternary fossil record are extant, so it is possible toestablish the relations between isotopic composition offossil and living samples (Goodfriend and Ellis, 2000).
In particular, land snail shells are common fossilremains of archaeological excavations and, the study oftheir isotopic composition can aid palaeoenvironmentalreconstructions at time of human occupation (Abell andPlug, 2000; Goodfriend and Ellis, 2000; Balakrishnanet al., 2005a). The aim of this paper is the palaeoenviron-mental reconstruction, through stable isotope recordobtained from land snail shells, of the Late Glacial depositpreserved at the Grotta del Romito (Southern Italy, Fig. 1)archaeological excavation. This archaeological excava-tion supplies a high resolution, well dated, stratigraphiccontinental succession between ca 13,000 and 14,500 yrcal BP, and represents the opportunity to test if the isotopiccomposition of land snail shells is a suitable tool toreconstruct rapid climatic fluctuations.
2. Geological and climate setting
The Grotta del Romito (39° 54′N, 15° 55′E) is a smallhorizontal cave open in the western side ofMonte Ciagolalimestone hill at 275 m a.s.l., about 25 km from theTyrrhenian coast (Fig. 1). This region is characterised bymountains rising to more than 2000 m, the Apennineridge, which descend steeply toward the MediterraneanSea. The study area is dominated by winds blowing fromwest supplying air masses that are partially dammed by theApennine ridge resulting in an abundant annual precip-itation (from 904 mm to 1502 mm according to localelevation and slope aspect). Mean monthly temperaturesat the site vary from a mean minimum of ∼8.5±4 °C,during January to February, to amaximumof∼27±3.1 °Cin August (Fig. 2). The mean annual oxygen isotopecomposition of meteoric water at the site is ca −6‰(Longinelli and Selmo, 2003; Leone and Mussi, 2004).
The deposits of Grotta del Romito contain one of mostimportant sequences of the Upper Palaeolithic of theItalian peninsula (from Gravettian to Late Epigravettian),recording a detailed prehistoric occupation of ca 13,000 yrin the immediate entrance of the cave (e.g. Graziosi, 1962,1971; Martini et al., 2003; Cattani et al., 2004; 2004a,b).Since the first seasons of excavation Grotta del Romitohad revealed a detailed prehistoric settlement, severalimportant manifestations of prehistoric art and an intense

Fig. 1. Location map of the studied area.
552 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
funerary use, thus assuming an important position in theframe of the Late Upper Palaeolithic of theMediterraneanbasin.
Fig. 3a shows the general stratigraphy of thearchaeological excavation within the cave, while Fig. 3bshows the location of the excavated area. Area A concernsthe old excavations performed at the beginning of 1960,whereas areas B and B′ are the recent excavations thatstarted in 2000.
3. Materials and methods
This study investigated 15 archaeological layers rela-tive to the Late Epigravettian succession (ca 13,000 and14,500 yr cal BP; Table 1) coming only from excavatedarea B (Fig. 3b) to avoid any problem of stratigraphiccorrelations. 14C ages were obtained on charcoals and arein stratigraphic order, indicating no reworking fromdifferent layers (Table 1). Land snail shell remains arerelatively abundant, but to reduce the possibility ofanalysing reworked specimens and to control thepreservation state, complete well preserved shells wereselected. Two herbivorous species were chosen for stableisotope analyses: Discus rotundatus (Müller, 1774)(Punctoidea, Patulidae), and Helix cf. ligata (Müller,1774) (Helicoidea, Helicidae). D. rotundatus is a
semelparous species (Cameron, 1982) living in habitatscharacterised by sufficient moisture such as dampsheltered places, in woods, maquis, under stones, andfallen trunks in damp herbaceous vegetations (Giusti et al.,1995; Kerney and Cameron, 1999). H. cf. ligata is apoorly known species endemic to Central–Southern Italy.According to Settepassi and Verdel (1965) its optimumseems to occur between 400 and 1000 m a.s.l. preferringwoody areas and avoiding long periods of drought. H. cf.ligata is probably an iteroparous species as other largespecies of subgenus Helicinae. For each archaeologicallayer 2 to 6 shells were analysed for the appropriatespecies. Living land snail shells of D. rotundatus, and H.cf. ligata were also collected at the cave entrance during2004–2005. Random samples of archaeological andmodern shells were analysed using X-ray diffraction(XRD) and scanning electron microscopy (SEM) to lookfor evidence of recrystallization. The results indicate thatthe internal aragonite structures were preserved. D.rotundatus occurs more abundantly from layer C toD8, whereas in lower levels it is less abundant andfragmented. By contrast H. cf. ligata has well preservedand complete specimens only below layer D8. Completeshells were cleaned in an ultrasonic bath, rinsed severaltimes in deionised water, powdered and ashed in a lowtemperature oxygen plasma to remove organic
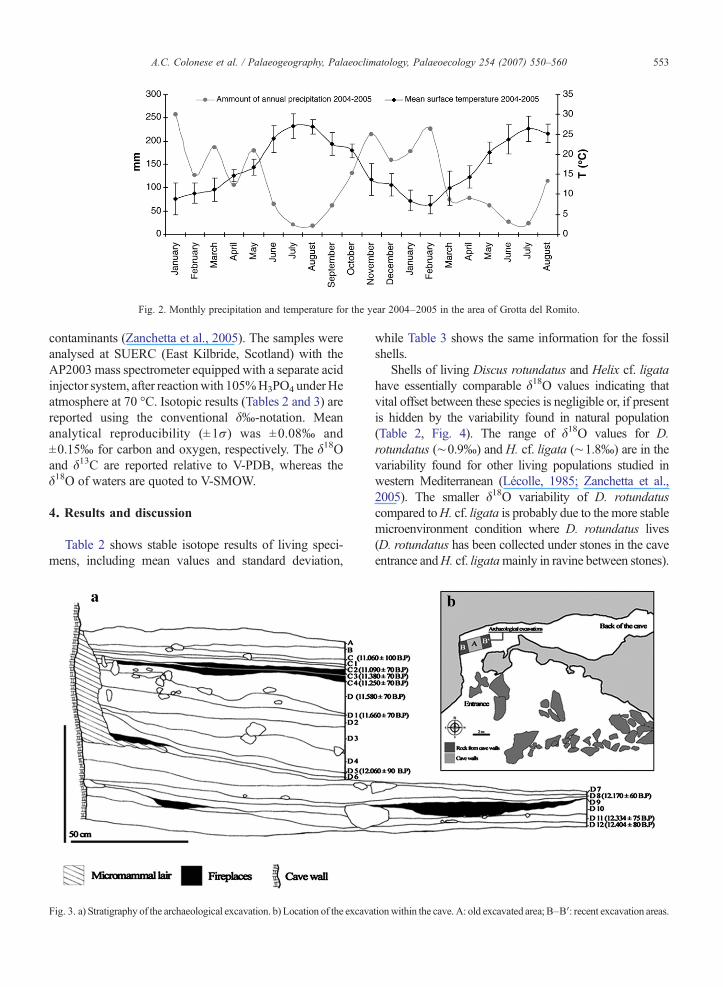
Fig. 2. Monthly precipitation and temperature for the year 2004–2005 in the area of Grotta del Romito.
553A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
contaminants (Zanchetta et al., 2005). The samples wereanalysed at SUERC (East Kilbride, Scotland) with theAP2003 mass spectrometer equipped with a separate acidinjector system, after reactionwith 105%H3PO4 underHeatmosphere at 70 °C. Isotopic results (Tables 2 and 3) arereported using the conventional δ‰-notation. Meananalytical reproducibility (±1σ) was ±0.08‰ and±0.15‰ for carbon and oxygen, respectively. The δ18Oand δ13C are reported relative to V-PDB, whereas theδ18O of waters are quoted to V-SMOW.
4. Results and discussion
Table 2 shows stable isotope results of living speci-mens, including mean values and standard deviation,
Fig. 3. a) Stratigraphy of the archaeological excavation. b) Location of the excava
while Table 3 shows the same information for the fossilshells.
Shells of living Discus rotundatus and Helix cf. ligatahave essentially comparable δ18O values indicating thatvital offset between these species is negligible or, if presentis hidden by the variability found in natural population(Table 2, Fig. 4). The range of δ18O values for D.rotundatus (∼0.9‰) and H. cf. ligata (∼1.8‰) are in thevariability found for other living populations studied inwestern Mediterranean (Lécolle, 1985; Zanchetta et al.,2005). The smaller δ18O variability of D. rotundatuscompared toH. cf. ligata is probably due to the more stablemicroenvironment condition where D. rotundatus lives(D. rotundatus has been collected under stones in the caveentrance andH. cf. ligatamainly in ravine between stones).
tionwithin the cave. A: old excavated area; B–B′: recent excavation areas.

Table 3Stable isotope data of the archaeological excavation. Mean andstandard deviation for each layer are also reported
Layers δ18OV-PDB‰ Mean SD δ13CV-PDB‰ Mean SD
Discus rotundatusC −0.20 −9.34
−0.29 −10.72−0.53 −10.52−0.19 −0.30 0.16 −10.72 −10.33 0.66
C1 −0.27 −10.14−0.49 −8.83−0.09 −8.50−0.24 −0.27 0.17 −9.22 −9.17 0.71
C2 0.46 −8.200.28 −12.110.53 −10.561.00 0.57 0.31 −9.26 −10.03 1.69
C3 0.70 −9.890.35 −10.820.70 −10.590.57 0.58 0.17 −10.03 −10.33 0.44
C4 0.75 −8.300.39 −7.511.12 0.75 0.37 −9.10 −8.30 0.80
D −0.20 −10.43−0.30 −11.800.13 −11.40
−0.12 −9.210.50 −10.08
−0.48 −0.08 0.35 −9.61 −10.42 1.01D1 0.08 −10.48
−0.15 −9.59−0.53 −8.410.46 −0.04 0.41 −9.60 −9.52 0.85
D2 −0.76 −10.42−0.52 −10.75−0.19 −0.49 0.29 −10.09 −10.42 0.33
D3 −0.48 −10.620.54 −9.820.54 −9.58
−0.73 −0.03 0.67 −11.39 −10.35 0.82D4 −0.16 −10.93
0.70 −10.440.49 −10.70
Table 114C dates available for the Grotta del Romito archaeologicalexcavation. Calibration was performed using the program Calib Rev.5.0.1 (Reimer et al., 2004)
Layer 14C yr BP 14C cal yr BP(±2σ)
δ13CV-PDB‰ Lab. code
C 11.060±100 13.170–12.890 – Beta-160295C1C2 11.090±70 13.190–12.890 – Beta-160296C3 11.380±70 13.500–13.140 – Beta-160297C4 11.250±70 13.460–13.010 – Beta-160298D 11.580±70 13.240–13.190 – Beta-160299D1 11.660±70 13.606–13.419 – Beta-160300D2D3D4D5 12.060±90 13.970–13.830 – Beta-160302D8 12.170±60 13.857–14.171 −26.8 CeDaD-LTL234AD9D11 12.334±75 14.013–14.748 −24.7 CeDaD-LtL238AD12 12.404±80 14.107–14.876 −25.6 CeDaD-LtL235A
554 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
However, Helix cf. ligata has, on average, lower δ13Cvalues compared toDiscus rotundatus probably related tothe different physiology and diet, whereas δ13C variability(∼2.5‰ and ∼3‰, for Helix and Discus respectively) iscomparable. Other studies have already shown that Helixspp. have the lowest δ13C values of shells when differentspecies are compared (Lécolle, 1983, 1984; Yates et al.,2002). This may be due to the efficiency of the oxygentransport system of medium–large land snail, such asthose of the Helicids, compared to small land snail, whichregulates the amount of CO2 adsorbed from air (McCon-naughey et al., 1997). This is in agreement with the Stott's(2002) results which show that 13C/12C of Helix aspersashell (Cornu aspersum according to recent literature) islittle influenced by diffusion of atmospheric CO2 intothe hemolymph. However, it is also possible thatD. rotundatus includes in its diet more carbonate than
−0.29 0.19 0.48 −10.50 −10.64 0.22D5 −0.07 −9.05
0.21 −10.12−0.66 −0.17 0.45 −11.81 −10.33 1.39
D8 −3.43 −10.33−2.15 −9.96−4.16 −3.25 1.02 −11.96 −10.75 1.06
Helix cf. ligataD9 −0.18 −8.96
−0.50 −9.05−1.39 −0.69 0.62 −9.61 −9.20 0.35
D11 −0.22 −9.550.44 0.11 0.47 −9.00 −9.27 0.39
D12 −0.28 −8.66−0.20 −8.67
Table 2Stable isotope data of living shells collected at the entrance of the cave
δ18OV-PDB‰ Mean SD δ13CV-PDB‰ Mean SD
Discus rotundatus0.13 −10.850.54 −9.78
−0.08 −12.78−0.35 0.06 0.38 −12.64 −11.51 1.45
Helix cf. ligata−0.39 −12.030.88 −13.290.10 −13.24
−0.93 −14.430.87 0.10 0.79 −12.98 −13.20 0.90

Fig. 4. δ18O–δ13C diagram of the whole set of isotopic data from Grotta del Romito.
555A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
H. cf. ligata (e.g. Yates et al., 2002). According to theexperiments Stott (2002) and Metref et al. (2003)performed on H. aspersa raised on controlled food, theδ13C of the living shells of Romito is compatible with adiet mainly composed of C3 plant material. A minor effectcan be due to annual/seasonal variability of 13C/12C ratiodue to different sources of food, soil CO2 production oronset of reproductive activity (e.g. Magaritz and Heller,1983; Leng et al., 1998; Baldini et al., 2007).
For each layer fossil populations show δ13C and δ18Oranges of values within that found for living counterpart,supporting the view of the absence of reworkedmaterial inthe set of shells analysed. The only exception is the δ13Cvalues for layer C2 (range: ∼3.9‰) but the δ18O has arange within of that of the living population (∼0.7‰). Asfor living populations the δ13C shows larger rangescompared to δ18O. The larger δ18O range of values wasfound at layer D8 (∼2‰). Zanchetta et al. (2005) havesuggested that the increasing δ18O spread of values foundin living populations may indicate an increase ofenvironmental stress. Comparison of the δ18O and δ13Cof the living shells with those of the fossil populations(Figs. 4 and 5, Tables 2 and 3) shows some interestingdifferences. The Discus rotundatus and Helix cf. ligataδ18O values are roughly similar in both fossil and livingpopulations.Only the three samples coming from layerD8appear strongly 18O-depleted compared to thewhole set ofdata. The SEM analyses of a specimen from this layer rule
out a possible diagenetic effect. The δ13C values ofH. cf.ligata fossil populations are basically 13C-enriched,whereas for the δ13C values of D. rotundatus a partialoverlap is apparent (Fig. 3). According to Goodfriend andEllis (2000) snail populations which lived after theindustrial revolution (ca after the AD 1860) may havebeen affected by the progressive 13C-depletion ofatmospheric CO2 due to the burning of fossil fuel (Friedliet al., 1986) superimposed on the natural δ13C variabilityusually found in living populations. The δ13C reduction ofthe atmospheric CO2 is today of ca 1.5‰ with respect tothe 19th century. If this reduction is completely propagat-ed, via ingested food, to the shell of living land snails itmay account for most of the observed δ13C variationbetween fossil and living populations ofD. rotundatus butonly a part of the difference found in theH. cf. ligata. It isimportant to note that δ13C values of plants depend onwhich part of the plant is considered, but more importantthey are species-dependent and vary according to manyenvironmental parameters, including the isotopic compo-sition of atmospheric CO2, atmospheric CO2 concentra-tion and water stress (e.g. White et al., 1994). Indeed, thedecrease of the δ13C of the atmospheric CO2 in the lastcentury is not clearly observed in the δ13C recordsobtained by tree-rings (Friedli et al., 1986). Therefore, thismay not be the ultimate reason for the observed δ13Cvalues in the fossils of Romito. Rather, it is interesting tonote that many δ13C records of “continental organic

556 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
matter” (e.g. bone collagen, leaf, organic lake sediment)show a significant decrease in δ13C between Late Glacialand Holocene transition, with δ13C values substantiallyhigher during the Last Glacial and Late Glacial (Stevensand Hedges, 2004 and references therein) despite of anegligible δ13C decrease of the atmospheric CO2 (ca0.3‰, Leuenberger et al., 1992). The higher δ13C valuesof many proxies during Late Glacial have been mainlyinterpreted as due to lower CO2 concentration (Krishana-murthy and Epstein, 1990; Stevens and Hedges, 2004).Therefore, the higher δ13C values of snail shells duringLate Glacial at Romito compared to living populations arein agreement with this very general tendency of theorganicmatter of plants and herbivorous tissues and bones
Fig. 5. Grotta del Romito δ18O and δ13C records. For compariso
to have higher δ13C values during the Last and LateGlacial compared to Holocene. However, the difference inδ13C between living and fossil snails, especially evidentfor H. cf. ligata, can be due to other factors. For instance,this difference could be easily produced by a diet of thelate Pleistocene snail populations containing basically C3
vegetation with different proportions of different plants(e.g. shrubs and tree ratio and/or evergreen and deciduousspecies) and/or withmixedC3 andC4 diet. It is also knownthat environmental stress like lower meteoric precipitationand humidity can produce increases in plant δ13C values(Stuiver and Broziunas, 1987).
Fig. 5 shows the detailed trend of the stable isotopecomposition of fossil specimens ofDiscus rotundatus and
n the isotopic values of living specimens are also reported.

557A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
Helix cf. ligata. As already observed the most consistentvariation is recorded by δ18O in layer D8 (13,857 to14,171 yr cal BP) with a strong 18O-depletion. Apart fromthis layer the trend does not show significant differencescompared to δ18O values of living population. The D8layer δ18O excursion is in very good agreement with theGI 1d climatic event (or Older Dryas, Björck et al., 1998;Fig. 6) in the GISP 2 ice core record (Stuiver et al., 1995).In Fig. 6 the ages of layers C1, D2 to D4 and D9 werelinearly extrapolated from ages of the bracketing layers.Although, this is not strictly correct because it assumes acontinuous sedimentation within the cave succession, itdoes not introduce particular bias concerning layer D8 agebeing directly dated.
As discussed in the Introduction a general model forinterpreting the oxygen isotope composition of snail shellsin termof past climatic changes is not yet available. For theItalian peninsula, living populations of land snail shellsshow a good relationship between shell δ18O and localδ18O of meteoric precipitation (δ18Os and δ18Op respec-tively) given by the equation (Zanchetta et al., 2005):
d18Op ¼ 0:65 d18Os� 5:44 ð1Þ
According to this equation layer D8may have recordeda 18O-depletion of ca 2‰ in the local mean precipitationcompared with the average. If the calculated variation ofthe δ18Op values was mainly due to changes in averagetemperature the layer D8 may have recorded a decrease intemperature of ca 5 to 10 °C taking into account that todaythe relation between the mean δ18Op and surfacetemperature in the Mediterranean ranges from 0.2 to0.4‰/°C (Hauser et al., 1980; Bard et al., 2002; Argiriouand Lykoudis, 2006). Obviously, these estimates rely onthe assumption that the δ18Op/°C ratio was valid also forpre-Holocene periods, an assumption supported byclimate simulations on very different climatic modesperformed by Bard et al., 2002 for the westernMediterranean basin. Despite this, a so drastic decreasein mean annual temperature, as suggested by δ18O valuesof shells, is probably unrealistic and the low δ18Op valuescould have been associated both to a change in tem-perature and atmospheric circulation. A substantialdecrease in δ18Op could have been produced by the so-called “amount effect” (Dansgaard, 1964), in which adecrease in δ18Op is associated to an increase in meteoricprecipitation. This mechanism has been invoked as one ofthe most important in driving many δ18O proxy records inthe Mediterranean basin (e.g. Bar-Matthews et al., 2000;Bard et al., 2002; Drysdale et al., 2004; Zanchetta et al.,2007). However, pollen (e.g. Sadori and Narcisi, 2001;Follieri et al., 1989) and δ18O records on speleothems (e.g.
Bar-Matthews et al., 2000; Drysdale et al., 2004, 2007)and lake carbonates (Baroni et al., 2006; Zanchetta et al.,1999, 2007) have shown that in the Mediterranean basinan increase in precipitation is related mainly to warmcondition whereas cold phases are usually associated todry conditions on land.
Therefore, the striking correlation of the event recordedin the layer D8 with GISP 2 and the fact that precipitationat basin scale seems to increase mainly during warmingphases, suggest that the δ18O of shell at layer D8 wasmainly influenced by a substantial decrease in the δ18O ofthe precipitation due to decrease in temperature but notassociated to the “amount effect”. It is worthy of mentionthat the other layers show isotopic composition essentiallysimilar to the living shells, which suggest conditions attime of shell growth similar to the present day. This mayalso support the interpretation that δ13C values of fossilshells are lower compared to living shells mainly becauseof lower CO2 concentration rather than different climateconditions.
The δ13C record of Discus rotundatus does not showmarked variation in layer D8 and also general trends arenot apparent (Fig. 4). This probably may indicate that theinferred climatic changes at layer D8 did not producesignificant isotopic impact on the snail diet and it is also inagreement with the fact that a consistent part of the δ18Opvariation inferred by shells is not probably related totemperature alone.
The δ13C values of Helix cf. ligata (layers D9, D11and D12) appear particularly high if compared to livingHelix and, once normalized to Discus rotundatus adding1.7‰ as mean differences observed in the livingpopulation (Table 2), these are the highest δ13C valuesof the record. The high δ13C values ofHelix at layers D9,D11 and D12 can be tentatively explained by the loweratmospheric CO2 concentration during the early Bølling(Monnin et al., 2001).
5. Conclusion
Despite the origin and environmental significance ofthe isotopic composition of land snail shells beingparticularly complex, this study demonstrates that stableisotope composition (in particular 18O/16O ratio) of landsnail shells can retain a signal of short term climaticchanges. Owing to the large isotopic variability of livingand fossil populations only stratigraphic successionssampled with adequate resolution and detail will usuallydeliver significant results; this can often be achieved atarchaeological excavations. However, archaeologicallayers can suffer different processes of reworking andstratigraphic disturbances produced by human occupation.

Fig. 6. Comparison between the δ18O records of Grotta del Romito and GISP 2. For Grotta del Romito the δ18O values are the mean values for eachlayer.
558 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
The data discussed indicate that this was not the case forGrotta del Romito.
The isotopicmeasurements on two species of land snailnot previously analysed show that there is no substantialoffset in the oxygen isotope composition between thesetwo species, but carbon isotopes appear to be species-dependent. This is in agreement with previous findings onother species (e.g. Zanchetta et al., 2005).
The oxygen isotope composition of land snail shellsshows a good agreement with the general trend observedin the GISP 2 δ18O ice core record. In particular, the δ18Orecord indicates a clear brief climatic deterioration chrono-logically consistent with the GI 1d climatic event.
The carbon isotope composition of living land snailshells appears 13C-depleted compared to fossil populations(in particular for fossilHelix cf. aspersa). The higher δ13Cvalues of fossil snail shells are in agreement with otherrecords suggesting a general increase of δ13C of
“continental organic matter” during Last to Late Glacialas effect of substantial lower CO2 concentration atthat time. However, other causes can have produced the13C-enriched values of fossil populations likemixedC3/C4
plant or different proportions of C3 species in snails diet.
Acknowledgements
The authorswish to thank T. Donnelly, J. Dougans, andA. Tait for their support in stable isotopes analyses. Thiswork is partially funded by the University of Pisa (ex 60%G. Zanchetta). SUERC is funded by a consortium ofScottish universities and NERC. This paper is part of thePh.D. thesis of A.C. in “Preistoria - Ambienti e Culture” atthe University of Siena, Sezione di Preistoria, XVIII ciclo.We also thank two referees for the comments andsuggestions which greatly improved the quality of thepaper.

559A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
References
Abell, P.I., Plug, I., 2000. The Pleistocene/Holocene transition in SouthAfrica: evidence for the Younger Dryas event. Glob. Planet. Change26, 173–179.
Argiriou, A.A., Lykoudis, S., 2006. Isotopic composition of precipitationin Greece — past and recent data. International Workshop onIsotopic Effects in Evaporation. Pisa 3–5 May 2006, pp. 14–17.
Balakrishnan, M., Yapp, C.J., 2004. The flux balance model for theoxygen and carbon isotope compositions of land snail shells.Geochim. Cosmochim. Acta 68, 2007–2024.
Balakrishnan, M., Yapp, C.J., Meltzer, D.J., Theler, J.L., 2005a.Paleoenvironment of the Folsom archeological site, New Mexico,USA, approximately 10,500 14C yr B.P. as inferred from stableisotope composition of fossil land snail shells. Quat. Res. 63, 31–44.
Balakrishnan, M., Yapp, C.J., Theler, J.L., Carter, B.J., Wyckoff, D.G.,2005b. Environmental significance of 13C/12C and 18O/16O ratios ofmodern land-snail shells from southern great plains of NorthAmerica. Quat. Res. 63, 15–30.
Baldini, M.L., Walzer, S.E., Railsback, L.B., Baldini, J.U.L., Crowe,D.E., 2007. Isotopic ecology of the modern land snail Cerion, SanSalvador, Bahamas: preliminary advances toward establishing alow-latitude island paleoenvironmental proxy. Palaios 22, 174–187.
Bard, E., Delaygue, G., Rostek, F., Antonioli, F., Silenzi, S., Schrag, D.,2002. Hydrological conditions over the western Mediterranean basinduring the deposition of the cold Sapropel 6 (ca. 175 kyr BP). EarthPlanet. Sci. Lett. 202, 481–494.
Bar-Matthews, M., Aylon, A., Kaufman, A., 2000. Timing andhydrological conditions of Sapropel events in the Eastern Mediter-ranean, as evident from speleothems, Soreq cave, Israel. Chem. Geol.169, 145–156.
Baroni, C., Zanchetta, G., Fallick, A.E., Longinelli, A., 2006. Molluscstable isotope record of a core from Lake Frassino, northern Italy:hydrological and climatic changes during the last 14 ka. Holocene16, 827–837.
Björck, S., Walker, M.J.C., Cwynar, L.C., Johnsen, S., Knudsen, K.-L.,Lowe, J.J., Wohlfarth, B., Intimate members, 1998. An eventstratigraphy for the Last Termination in the North Atlantic regionbased on the Greenland ice-core record: a proposal by theINTIMATE group. J. Quat. Sci. 13, 283–292.
Cameron, R.A.D., 1982. Life histories, density and biomass in awoodland snail community. J. Mol. Struct. 4, 159–166.
Cattani, L., Colonese, A.C., Ricciardi, S., 2004. Ricostruzione climaticae ambientale di Grotta del Romito: analisi palinologiche emalacofaunistiche dei livelli epigravettiani C e D. Atti della XXXVIIRiunione Scientifica, Preistoria e Protostoria della Calabria, II,pp. 637–640.
Dansgaard, W., 1964. Stable isotopes in precipitation. Tellus 4, 436–468.Drysdale, R.N., Zanchetta, G., Hellstrom, J.C., Zhao, J., Fallick, A.E.,
Isola, I., Bruschi, G., 2004. Palaeoclimatic implications of the growthhistory and stable isotopes (δ18O and δ13C) geochemistry of a middleto late Pleistocene stalagmite from central–western Italy. EarthPlanet. Sci. Lett. 227, 215–229.
Drysdale, R.N., Zanchetta, G., Hellstrom, J.C., Fallick, A.E., McDonald,J., Cartwrigth, I., 2007. Stalagmite evidence for the precise timing ofNorth Atlantic cooling events during the early Last Glacial. Geology35, 77–80.
Follieri, M., Magri, D., Sadori, L., 1989. Pollen stratigraphical synthesisfrom Valle di Castiglione (Roma). Quat. Int. 3–4, 81–84.
Friedli, H., Lötscher, H., Oeschger, H., Siegenthaler, U., Stauffer, B.,1986. Ice core record of the 13C/12C ratio of atmospheric CO2 in thepast centuries. Nature 324, 237–238.
Giusti, F., Manganelli, G., Schembri, P.J., 1995. The Non-marineMolluscs of Maltese Islands. . Monografie, vol. XV. MuseoRegionale di Scienze Naturali, Torino.
Goodfriend, G.A., Ellis, G.L., 2000. Stable carbon isotope record ofmiddle to late Holocene climate changes from land snail shells atHinds Cave, Texas. Quat. Int. 67, 47–60.
Goodfriend, G.A., Ellis, G.L., 2002. Stable carbon and oxygen isotopicvariations in modern Rabdotus land snail shells in the southern GreatPlains, USA, and their relation to environment. Geochim. Cosmo-chim. Acta 66, 1987–2002.
Goodfriend, G.A., Hood, D.G., 1983. Carbon isotope analyses of landsnail shells: implications for carbon sources and radiocarbon dating.Radiocarbon 25, 810–830.
Goodfriend, G.A., Magaritz, M., Gat, J.R., 1989. Stable isotopecomposition of land snail body water and its relation toenvironmental water and shell carbonate. Geochim. Cosmochim.Acta 53, 3215–3221.
Graziosi, P., 1962. Découvert de gravures rupestres de type paléolithiquedan l'abri del Romito (Italie). L'Anthropologie 66 (3–4), 262–268.
Graziosi, P., 1971. Dernières découverts de gravures paléolithiques dansla grotte du Romito en Calabre. Mélanges André Varagnac, Paris,pp. 355–357.
Hauser, S., Dongarrà, G., Favara, R., Longinelli, A., 1980. Composizionedelle piogge in Sicilia. Riferimenti di base per studi idrogeologici erelazione con altre aree mediterranee. Rend. Soc. Ital. Mineral. Petrol.36, 671–690.
Kerney, M.P., Cameron, R.A.D., 1999. Guide des escargots et limacesd'Europe. Delachaux et Niestlé, Paris.
Krishanamurthy, R.V., Epstein, S., 1990. Glacial–interglacial excursionin the concentration of atmospheric CO2: effect in the
13C/12C ratio inwood cellulose. Tellus 42B, 423–434.
Lécolle, P., 1983. Relation entre les teneurs en 18O and 13C des test deGastéropodes terrestres et le climat océanic et alpin. C. R. Acad. Sci.Paris 297, 863–866.
Lécolle, P., 1984. Influence de l'altitude en climat méditerranéen sur lesteneurs en oxygène-18 et carbon-13 des coquilles de Gastéropodesterrestres. C. R. Acad. Sci. Paris 298, 211–214.
Lécolle, P., 1985. The oxygen isotope composition of landsnail shells asa climatic indicator: applications to hydrogeology and paleoclima-tology. Chem. Geol. 58, 157–181.
Leng, M.J., Heaton, T.H.E., Lamb, H.F., Naggs, F., 1998. Carbon andoxygen isotope variations within the shell of an African land snail(Limicolaria kaumbeul chudeaui Germani): a high-resolution recordof climate seasonality. Holocene 8, 407–412.
Leone, G., Bonadonna, F.P., Zanchetta, G., 2000. Stable isotope record inmollusca and pedogenic carbonate fromLate Pliocene soils of CentralItaly. Palaeogeogr. Palaeoclimatol. Palaeoecol. 163, 115–131.
Leone, G., Mussi, M., 2004. Environmental isotopes and well waters andundersea sulphurous outflows at Capo Palinuro, Salerno. J. Technol.Environ. Geol. 3, 35–48.
Longinelli, A., Selmo, E., 2003. Isotopic composition of precipitation inItaly: a first overall map. J. Hydrol. 270, 75–88.
Leuenberger, M., Siegenthaler, M., Langway, C.C., 1992. Carbonisotope composition of atmospheric CO2 during the last ice age froman Antartic ice core. Nature 357, 488–490.
Magaritz,M.,Heller, J., 1983.Annual cycle of 18O/16O and13C/12C isotoperatios in landsnail shells. Chem. Geol. (Isot. Geosc.) 1, 243–255.
Martini, F., Martino, G., Rolle, R., 2003. L'Epigravettiano finale diGrotta del Romito a Papasidero: L'industria litica degli strati C e D.Riv. Sci. Preist. 53, 55–138.
Martini, F., Cattani, L., Colamussi, V., Colonese, A.C., Martino, G.,Mallegni, F., Noto, F., Ricciardi, S., Rickards, O., Rolle, R., 2004a.

560 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 254 (2007) 550–560
Primi risultati delle nuove ricerche nei livelli epigravettiani di Grottadel Romito a Papasidero (scavi 2000–2002). Atti della XXXVIIRiunione Scientifica I.I.P.P. Preistoria e Protostoria della Calabria,vol. 1, pp. 35–58.
Martini, F., Bisconti, M., Casciarri, S., Fabbri, P.F., Leonini, V., Lo Vetro,D., Mallegni, F., Martino, G., Noto, F., Ricci, S., Ricciardi, S.,Rickards, O., 2004b. La nuova sepoltura epigravettiana “Romito 7” aPapasidero. Atti della XXXVII Riunione Scientifica I.I.P.P. Preistoriae Protostoria della Calabria, vol. I, pp. 101–111.
McConnaughey, T.A., Burdett, J., Whelan, J.F., Paul, C.K., 1997. Carbonisotopes in biological carbonates: respiration and photosynthesis.Geochim. Cosmochim. Acta 61, 611–622.
Monnin, E., Indermühle, A., Dällenbach, A., Flückinger, J., Stauffer, B.,Stocker, T.F., Raynaud, D., Barnola, J.-M., 2001. Atmospheric CO2
concentration over the Last Glacial Termination. Science 291,112–114.
Metref, S., Rousseau, D.-D., Bentaleb, I., Labonne, M., Vianey-Liaud,M., 2003. Study of the diet effect on δ13C of shell carbonate of theland snail Helix aspersa in experimental conditions. Earth Planet.Sci. Lett. 211, 381–393.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand,C., Blackwell, P.G., Buck, C.E., Burr, G., Cutler, K.B., Damon, P.E.,Edwards, R.L., Fairbanks, R.G., Friedrich, M., Guilderson, T.P.,Hughen, K.A., Kromer, B., McCormac, F.G., Manning, S., BronkRasmsy, C., Reimer, R.W., Remmele, S., Southon, J.R., Stuiver, M.,Talamo, S., Taylor, F.W., van der Plicht, J., Weyhnmer, C.E., 2004.IntCal04 terrestrial radiocarbon age calibration 0–26 cal kyr BP.Radiocarbon 46, 1029–1050.
Sadori, L., Narcisi, B., 2001. The Postglacial record of environmentalhistory from Lago di Pergusa. Holocene 11, 655–670.
Settepassi, F., Verdel, V., 1965. Continental Quaternary molluscs of theLower Liri Valley (Southern Latium). Geol. Rom. 4, 369–451.
Stevens, R.E., Hedges, R.E.M., 2004. Carbon and nitrogen stable isotopeanalysis of northwest European horse bone and tooth collagen,
40,000 BP–present: palaeoclimatic interpretation. Quat. Sci. Rev. 23,977–991.
Stott, L.D., 2002. The influence of diet on the δ13C of shell carbon in thepulmonate snailHelix aspersa. Earth Planet. Sci. Lett. 195, 249–259.
Stuiver, N., Broziunas, T.F., 1987. Tree cellulose 13C/12C isotope ratiosand climate changes. Nature 328, 59–60.
Stuiver, M., Grootes, P.M., Braziunas, T.F., 1995. The GISP2 18O climaterecord of the past 16,500 years and the role of the sun, ocean andvolcanoes. Quat. Res. 44, 341–354.
Yapp, C.Y., 1979. Oxygen and carbon isotope measurements of landsnail shell carbonate. Geochim. Cosmochim. Acta 43, 629–635.
Yates, T.J.S., Spiro, B.F., Vita-Finzi, C., 2002. Stable isotope variabilityand the selection of terrestrial mollusc shell samples for 14C dating.Quat. Int. 87, 87–100.
White, J.W.C, Ciais, P., Fige, R.A., Kenny, R., Markgraf, V., 1994. Ahigh-resolution record of atmospheric CO2 content from carbonisotopes in peat. Nature 367, 153–156.
Zanchetta, G., Bonadonna, F.P., Leone, G., 1999. A 37-meter record ofpaleoclimatological events from stable isotope data on continentalmolluscs in Valle di Castiglione, near Rome, Italy. Quat. Res. 52,293–299.
Zanchetta, G., Bonadonna, F.P., Ciampalini, A., Fallick, A.E., Leone, G.,Marcolini, F., Michelucci, L., 2004. Intratyrrhenian cooling eventdeduced by non-marine mollusc assemblage at Villa S. Giorgio(Livorno, Italy). Boll. Soc. Paleontol. Ital. 43, 347–359.
Zanchetta, G., Leone, G., Fallick, A.E., Bonadonna, F.P., 2005. Oxygenisotope composition of living land snail shells: data from Italy.Palaeogeogr. Palaeoclimatol. Palaeoecol. 223, 20–33.
Zanchetta, G., Borghini, A., Fallick, A.E., Bonadonna, F.P., Leone, G.,2007. Late Quaternary palaeohydrology of Lake Pergusa (Sicily,southern Italy) as inferred by stable isotopes of lacustrine carbonates.J. Paleolimnol. doi:10.1007/s10933-006-9070-1.
Copyright © 2022 FDOKUMEN




















