
‘St Martin's Summer’: the case of an autumn phytoplankton bloom in the Gulf of Naples...
19
Journal of Plankton Research Vol.17 no.3 pp.575-593. 1995 'St Martin's Summer': the case of an autumn phytoplankton bloom in the Gulf of Naples (Mediterranean Sea) Adriana Zingone, Raffaella Casotti, Maurizio Ribera d'Alcala, Michele Scardi and Donato Marino Stazione Zoologica 'A. Dohrn', Villa Comunale, 80121 Naples, Italy Abstract. Phytoplankton abundance, species composition and primary production of the Gulf of Naples were investigated during an autumn bloom in November 1985. Hydrographic data and surface phytoplankton samples were collected during a 3 day cruise, whereas investigations on in situ primary production and phytoplankton vertical distribution were conducted from a second boat on three different dates. A coast-offshore gradient was recorded for most of the chemical and biological parameters analysed. At stations closer to the coast, which were affected by land run-off, phytoplankton populations attained concentrations of 2.5 x lO* cells P'. with a marked dominance of diatoms belonging to the genera Thalassiosira and Chaetoceros. The most striking character of the system was a remarkably high carbon assimilation rate (up to 1.04 g C m' 2 day" 1 ) at stations closer to the coast. The causative mechanism for this bloom appeared to be land-derived nutrient enrichment, possibly enhanced by autumn rains, followed by a period of favourable meteorological conditions, which occurs recurrently in the region for a brief period around November, known locally as "St Martin's Summer'. We hypothesize that a similar mechanism can stimulate phytoplankton growth more than once every year, since high-stability periods following meteoro- logical perturbations can occur several times in temperate regions of the northern hemisphere in autumn. Introduction The periodicity of phytoplankton in the oceans is driven by seasonal variations in temperature and day length which, besides affecting the physiological response in phytoplankton cells, produce marked changes in the availability of nutrient and light resources by determining the vertical structure of the water column. In coastal waters, phytoplankton periodicity is also affected by the different sources of land-derived nutrients and by their dilution patterns, which in turn are related to the topography and hydrography of the basins. Although each coastal zone has a peculiar ecological physiognomy, which implies that general predictive schemes cannot be applied to these systems, it is now obvious that the contribution of these areas to the global productivity is much higher than expected from their spatial extension (Walsh, 1988; Mantoura et al., 1991). In temperate coastal waters, seasonality results in a major phytoplankton bloom, generally in spring-summer, and eventually in a second bloom in autumn. Besides these two main phases, other phytoplankton blooms and sustained primary production may be observed throughout the summer in some coastal embayments (Takahashi et al., 1977; Ignatiades, 1984), estuaries (Smayda, 1983) and shelf areas (O'Reilly and Bush, 1984). So far, interest has focused on spring-summer blooms and the mechanisms underlying their onset, intensity, duration and fate, whereas autumn blooms have received much less attention. Among the mechanisms postulated for autumn blooms are coastal upwelling (Colebrook, 1979; Estrada, 1984), enrichment from sediments (Smetacek, 1985; Noji et al., 1986), diffusion of nutrients from below the © Oxford University Press 575 at The Open University on December 19, 2010 plankt.oxfordjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of ‘St Martin's Summer’: the case of an autumn phytoplankton bloom in the Gulf of Naples...

Journal of Plankton Research Vol.17 no.3 pp.575-593. 1995
'St Martin's Summer': the case of an autumn phytoplankton bloomin the Gulf of Naples (Mediterranean Sea)
Adriana Zingone, Raffaella Casotti, Maurizio Ribera d'Alcala, Michele Scardiand Donato Marino
Stazione Zoologica 'A. Dohrn', Villa Comunale, 80121 Naples, Italy
Abstract. Phytoplankton abundance, species composition and primary production of the Gulf ofNaples were investigated during an autumn bloom in November 1985. Hydrographic data andsurface phytoplankton samples were collected during a 3 day cruise, whereas investigations on in situprimary production and phytoplankton vertical distribution were conducted from a second boat onthree different dates. A coast-offshore gradient was recorded for most of the chemical and biologicalparameters analysed. At stations closer to the coast, which were affected by land run-off,phytoplankton populations attained concentrations of 2.5 x lO* cells P ' . with a marked dominanceof diatoms belonging to the genera Thalassiosira and Chaetoceros. The most striking character of thesystem was a remarkably high carbon assimilation rate (up to 1.04 g C m'2 day"1) at stations closerto the coast. The causative mechanism for this bloom appeared to be land-derived nutrientenrichment, possibly enhanced by autumn rains, followed by a period of favourable meteorologicalconditions, which occurs recurrently in the region for a brief period around November, knownlocally as "St Martin's Summer'. We hypothesize that a similar mechanism can stimulatephytoplankton growth more than once every year, since high-stability periods following meteoro-logical perturbations can occur several times in temperate regions of the northern hemisphere inautumn.
Introduction
The periodicity of phytoplankton in the oceans is driven by seasonal variations intemperature and day length which, besides affecting the physiological responsein phytoplankton cells, produce marked changes in the availability of nutrientand light resources by determining the vertical structure of the water column. Incoastal waters, phytoplankton periodicity is also affected by the different sourcesof land-derived nutrients and by their dilution patterns, which in turn are relatedto the topography and hydrography of the basins. Although each coastal zonehas a peculiar ecological physiognomy, which implies that general predictiveschemes cannot be applied to these systems, it is now obvious that thecontribution of these areas to the global productivity is much higher thanexpected from their spatial extension (Walsh, 1988; Mantoura et al., 1991).
In temperate coastal waters, seasonality results in a major phytoplanktonbloom, generally in spring-summer, and eventually in a second bloom inautumn. Besides these two main phases, other phytoplankton blooms andsustained primary production may be observed throughout the summer in somecoastal embayments (Takahashi et al., 1977; Ignatiades, 1984), estuaries(Smayda, 1983) and shelf areas (O'Reilly and Bush, 1984). So far, interest hasfocused on spring-summer blooms and the mechanisms underlying their onset,intensity, duration and fate, whereas autumn blooms have received much lessattention. Among the mechanisms postulated for autumn blooms are coastalupwelling (Colebrook, 1979; Estrada, 1984), enrichment from sediments(Smetacek, 1985; Noji et al., 1986), diffusion of nutrients from below the
© Oxford University Press 575
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

\.7Angoneet al.
thermocline (Margalef, 1969) and freshwater run-off (Roden, 1984), all of whichconverge towards episodic enrichment of the photic layer. Light availability inautumn is still high enough for the bloom development. Reduced grazingpressure has also been invoked to explain the development of a high standingcrop(Smayda, 1957; Cushing, 1975).
An autumn peak of phytoplankton biomass and primary production, whichgenerally occurs in October-November, has also often been recorded in theMediterranean Sea, in addition to the main spring bloom [see Estrada et al.(1985) for a review]. In agreement with the most widely accepted explanation,this peak has been related to the autumnal breakdown of the thermocline, whichforces nutrients from deeper waters into the photic zones and stimulates a newpulse in phytoplantkon population growth (Margalef, 1969).
In this study, we present the case of an autumn phytoplankton bloom in theGulf of Naples and suggest that episodic conditions of weather stability followingnutrient enrichment from land run-off can significantly enhance autumnphytoplankton production in this area. Although this phenomenon may belimited in time, it can be quite significant if we consider that periods offavourable weather conditions are recurrent and widespread in mid-latitudeautumn (Kalnicky, 1987).
Study area
The Gulf of Naples is a rather deep (mean depth 170 m) coastal embaymentopening onto the Tyrrhenian Sea. The isobath of 100 m runs more or lessparallel to the coast, at a distance of ~ l - 5 miles, all along its shoreline (Figure1). The water column structure is characterized by a thermal stratification fromApril to early December. Within the gulf, there is a coastal and an open-watersubsystem characterized by distinct physical, chemical and biological properties(Carrada et al., 1980).
In the coastal subsystem, the main source of nutrients is constituted byterrestrial inputs. These include the Sarno River, which has a limited flow, butdrains an area of intense human activities, and untreated urban wastes which arescattered along the whole coast. These inputs, so far not systematicallyquantified, result in nutrient concentrations which can fluctuate widely, but mostfrequently range between 1 and 2 u.mol I"1 of inorganic N, 2-4 of Si and 0.1-0.4 P, and are always detectable. Phytoplankton populations generally attaintheir maximum concentrations and productivity in late spring-early summer, incorrespondence with maximum daylength and thermal stratification of the watercolumn, which limits most of the terrestrial inputs to surface layers (Marino etal., 1984; Scotto di Carlo et al., 1985; Ribera d'Alcala et al., 1989). In the innerpart of the gulf, the most intense summer blooms, mainly formed by small-sizeddiatoms, such as Chaetoceros cf. simplex f. calcitrans. Thalassiosira spp.,Leptocylindrus sp., Skeletonema costatum and small-sized flagellates, succeedand overlap each other, and are confined to surface waters (Modigh et al., 1985;Zingone et al., 1985, 1990a). Monthly averages for primary production,measured biweekly over a 5 year period at the fixed Station MC (marked by anasterisk in Figure 1), show a prolonged period of production values >1 g C m"2
576
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton bkwm in Gulf of Naples
N A P L E S40*50'
-40*40'
h 40*30'13*5" 14*0
Fig. I . Gu l f of Naples with sampling sites. Circles: hydrography stations; full circles: hydrographyand phytoplankton stations; triangles: productivity stations; asterisk: Station MC.
I 1 1 \ 1 1
Fig. 2. Monthly averages, minima and maxima of primary production (mg C m 2 day" ' ) at StationMC in 1984-1988.
577
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

A.Zingoneef al.
day"1 from April to July (Figure 2). A second production pulse constituted byone or more peaks, which are generally shorter in time but occasionally evenmore intense, occurs in autumn [Scotto di Carlo et al. (1985) and unpublisheddata].
By contrast, the open-water subsystem is characterized by a markedoligotrophy. The Levantine Intermediate Water (LIW), the only water mass richin nutrients, is found well below the photic zone all the year round. After amodest spring bloom, nutrients are almost completely depleted in the photiczone (Carrada el al., 1980). The transitional zone between coastal and openwaters is narrow and close to the coast in summer, and wide and far from thecoast in the other seasons (Marino et al., 1984).
Method
To obtain a synoptic hydrographic survey of the area, 23 stations (Figure 1) weresampled by the m/b 'Loran' from 8 to 10 November 1985; this will be referred tohenceforth as the 'hydrographic cruise'. At each station, CTD profiles (IdronautModel 101 sound) were obtained down to a maximum depth of 120 m. From 12selected stations, surface samples were collected with a bucket to determinesalinity, nutrients, chlorophyll a and phytoplankton concentrations.
To measure in situ primary production, six stations were sampled on 9, 14 and15 November by the m/b 'R.Dohrn'; this will be referred to as the 'productivitycruise'. This sampling was planned to be concurrent with the hydrographiccruise, but it was delayed by several days because of technical problems. Four ofthese stations (3, 10, 18 and 19) had been sampled by the m/b 'Loran' a few daysearlier, whereas Station 12 was sampled simultaneously by the two boats on 9November. Station 24 was sampled only during the productivity cruise. Watersamples were collected with Niskin bottles fitted with reversing thermometers atthe surface and at depths corresponding to 50, 25, 10 and 1% of the totalincident photosynthetically active radiation (PAR), which was measured using asubmersible Li-Cor quantum sensor. These samples were used for nutrient,salinity, chlorophyll a and primary production measurements, and thosecollected from the surface and at depths corresponding to 25 and 10% of theincident PAR were also analysed for phytoplankton abundance andcomposition.
Salinity samples were analysed with an Autosal laboratory salinometer(Model 8400). Phosphate, nitrite, nitrate, ammonia and silicate concentrationswere determined with an Autoanalyzer Technicon AAII (Grasshoff etal., 1983).
For chlorophyll a analysis, a volume of water varying from 1.5 to 5.5 1 wasfiltered on 0.5 jim fibreglass GFF filters. Chlorophyll a was extracted with 90%neutralized acetone and its concentration was determined spectrophoto-metrically using the equation indicated by SCOR-UNESCO (1966). Primaryproduction measurements were performed by incubating samples with I4C in situ(Strickland and Parsons, 1965) for 4 h around noon.
Phytoplankton samples were fixed with neutralized formaldehyde to a finalconcentration of 1.6% and observed with a Zeiss IM35 inverted microscope.
578
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton btoom in Gulf of Naples
Cells were counted along two diametric transepts of the bottom plate, aftersedimentation for 1-3 days of subsamples varying from 5 to 100 ml, dependingon cell concentration. The whole bottom plate was also examined for scarcelyrepresented species. Algae which could not be identified to the species or genuslevel were assigned to suprageneric groups such as centric diatoms, coccolitho-phorids, euglenophyceans, cryptophyceans, prasinophyceans, dinoflagellatesand other phytoflagellates. The latter two groups were further subdivided intosmall (<15 .̂m) and large (>15 ^m) naked and thecate dinoflagellates, andsmall (<10 u.m) and large (>10 |xm) phytoflagellates. The principal taxonomictexts used for species identification were those proposed by Zingone et al.(1990b).
Correspondence analysis was performed to define the main coenoclinestructure. In order to define spatial partitions on the basis of phytoplanktonpopulation structure, constrained clustering (Legendre, 1987) was performedusing a complete linkage algorithm on a percentage similarity matrix. Stationcontiguity was established by means of a Gabriel net (Gabriel and Sokal, 1969)and provided the spatial constraint for the clustering algorithm. For both theseanalyses, the whole data set was reduced to 67 phytoplankton taxa by mergingpoorly represented species or congeneric species which supposedly have similarecophysiological characteristics.
Results
Hydrographic data, chlorophyll a and primary production
The water column structure was mainly determined by temperature, which washomogeneous down to —30-40 m, values being in the range of 19.16-19.67°C.The spatial distribution of terrestrial inputs was clearly reflected in surfacesalinity values (Figure 3a), and in salinity values averaged on the first 30 m(Figure 3b). Freshwater content within the mixed layer decreased with thedistance from the coast; however, the diluting effect of freshwater was stilldetected at the most offshore stations. Minimum salinity values (down to 37.54PSU) were recorded in surface layers at stations nearest the mouth of the RiverSarno (Stations 11 and 12), where they determined a weak stratification in the
Fig. 3. Hydrographic cruise, (a) Surface salinity contours (PSU). (b) Contours of salinity values(PSU) averaged on the first 30 m depth.
579
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from
\.Z.\ngone et al.
first metres of the water column. At other inshore stations, values <38 PSUwere recorded throughout the whole mixed layer.
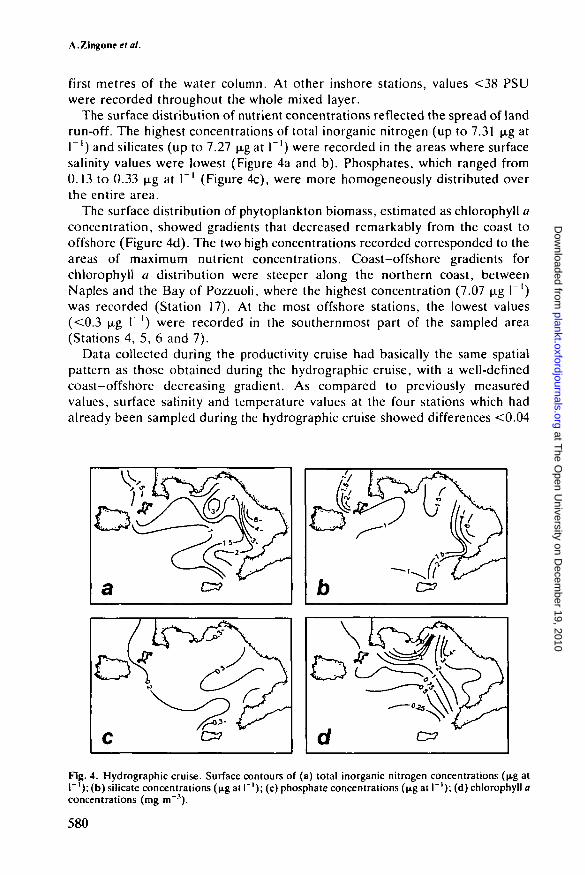
The surface distribution of nutrient concentrations reflected the spread of landrun-off. The highest concentrations of total inorganic nitrogen (up to 7.31 .̂g atP') and silicates (up to 7.27 u,g at I"1) were recorded in the areas where surfacesalinity values were lowest (Figure 4a and b). Phosphates, which ranged from0.13 to 0.33 \i.g at I"1 (Figure 4c), were more homogeneously distributed overthe entire area.
The surface distribution of phytoplankton biomass, estimated as chlorophyll aconcentration, showed gradients that decreased remarkably from the coast tooffshore (Figure 4d). The two high concentrations recorded corresponded to theareas of maximum nutrient concentrations. Coast-offshore gradients forchlorophyll a distribution were steeper along the northern coast, betweenNaples and the Bay of Pozzuoli, where the highest concentration (7.07 (ig 1~')was recorded (Station 17). At the most offshore stations, the lowest values(<0.3 u.g I"1) were recorded in the southernmost part of the sampled area(Stations 4, 5, 6 and 7).
Data collected during the productivity cruise had basically the same spatialpattern as those obtained during the hydrographic cruise, with a well-definedcoast—offshore decreasing gradient. As compared to previously measuredvalues, surface salinity and temperature values at the four stations which hadalready been sampled during the hydrographic cruise showed differences <0.04
Fig. 4. Hydrographic cruise. Surface contours of (a) total inorganic nitrogen concentrations (\i% atr ' ) ; (b) silicate concentrations (u,g at I"1); (c) phosphate concentrations (ugat I"'); (d) chlorophyll aconcentrations (mg m"3).
580
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton bloom in Gulf of Naples
PSU and 0.4°C, respectively, thus indicating that basic hydrographic features inthe area had not changed substantially between the two sampling cruises.
Nutrients were distributed rather homogeneously within the photic zone, withthe exception of Station 12, where total inorganic nitrogen and silicates rapidlydecreased from 5.67 to 1 and from 7.27 to 1.3 p.g at I"1, respectively, within adepth of 10 m.
Surface values for chlorophyll a concentration (Figure 5) were >1.5 mg m~3
at the four most inshore stations and reached 3.7 mg m~3 at Station 12, in thewestern part of the gulf. The vertical distribution for these values showed amarked decrease below 10 m at Station 12 and below 20 m at the other inshorestations. At the two stations located in the central area of the gulf, chlorophyll avalues were lower (0.31-0.45 mg m~3) and more homogeneously distributedthroughout the photic zone. All the four stations that had been sampledpreviously during the hydrographic cruise, including Station 3 which had beensampled only 1 day before, had surface chlorophyll a concentrations notablylower than those previously measured.
Production was measured in situ at six stations on 3 days during which lightranged from 10.22 to 15.43 E m"2 day"' (Table I). Primary production valuesfollowed the same spatial pattern as chlorophyll a values (Figure 5). At stationscloser to the coast, values ranging from 77 to 85% of the total integratedproduction were obtained between the surface and a depth of 10% of theincident PAR (10-20 m). Integrated values over the euphotic zone ranged from441 mg C m"2 day"' at Station 18 to >1000 mg C m"2 day"' at the two stations
100 150 2002 3 4
Chi
p
c0
•*-V 10'\\ 20
ft 30j*4*** An.
111
501
1002
/ -
/I
1503
- — -
2004
2505
—I12
Fig. 5. Productivity cruise. Vertical distribution of chlorophyll a (mg m~'; full line) and primaryproduction values (mg C m~* day"1; dashed line). The dotted line corresponds to the lowest limit ofthe water layer within which 90% of the total integrated production occurs.
581
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

N I
Tab
le I
. P
rim
ary
prod
ucti
on a
nd s
ome
rela
ted
para
met
ers
at t
he s
ix p
rodu
ctiv
e st
atio
ns
Sta
tion
3 12 19 24 10 18
Dat
e
9.11
9.11
14.1
114
.11
15.1
115
.11
PA
R(E
m-:d
ay
"')
10.2
210
.22
11.0
711
.07
15.4
315
.43
Chl
a 0
m(m
g m
"3)
0.32
3.70
0.45
2.37
2.23
1.64
Pro
duct
ion
0 m
(mg
C m
"1 h"1
)
1.04
30.1
11.
2611
.37
12.8
77.
43
P/B
()m
(mg
C(m
gc
hlo
)"1 h
"1)
3.24
8.14
2.81
4.80
5.77
4.53
Inte
grat
ed p
rodu
ctio
n(m
g C
m"2
day
"1)
50.3
510
33.1
614
8.47
798.
0210
40.4
744
1.33
at The Open University on December 19, 2010plankt.oxfordjournals.orgDownloaded from

Autumn phytoplankton bloom in Gulf of Naples
in the eastern part of the gulf (Stations 10 and 12). In the central area of the gulf,markedly lower production values along the whole water column were recorded,with surface values of 8.87 and 7.6 mg C m"3 day"1, and integrated values of 148and 50 mg C m"2 day"1 at Stations 19 and 3, respectively. It should be noted thatthe six stations where production was measured did not include the stationswhich had been found to be the richest (Station 17) and the poorest (Stations 4,5, 6 and 7) in terms of phytoplankton biomass during the hydrographic cruise.
Phytoplankton
A total of 194 microalgae—80 diatoms, 69 dinoflagellates, 35 coccolithophorids,three silicoflagellates and seven flagellates belonging to different taxonomicgroups—were identified to species level.
During the hydrographic cruise, phytoplankton abundance decreased fromcoastal to offshore stations, with a maximum of 2.5 and a minimum of 0.09 x 106
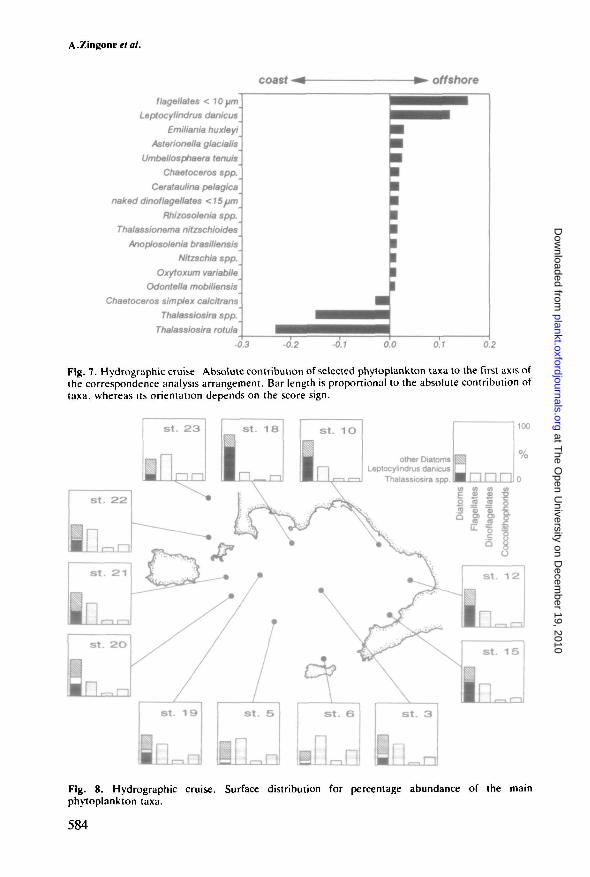
cells 1~' at Stations 18 and 6, respectively (Figure 6a). This spatial pattern alsoemerged from the correspondence analysis, thus demonstrating a similargradient for community structure (Figure 6b). As shown in Figure 7, thisgradient was determined by the asymmetric distribution of a small number oftaxa, which nonetheless determined marked differences in species dominanceand percentage composition of the communities (Figure 8). At stations closer tothe coast (Stations 10, 12, 15 and 18), diatoms including Thalassiosira rotula (upto 49% of diatom populations), T.allenii and several small-sized species of thisgenus, as well as some species of Chaetoceros (C. cf. simplex f. calcitrans),dominated surface phytoplankton populations, attaining percentages from 58 to75%. In the central area of the gulf (Stations 3 and 19) and in the northwesternpart of the Bocca Grande Channel (Stations 20 and 21), phytoplanktonpopulations were also dominated by diatoms (from 49 to 59%), although with amore diversified population and a lower contribution of T. rotula (from 12 to25% of the total number of diatoms). Similar phytoplankton populations werefound in the Gulf of Gaeta, but Station 23 had higher percentages ofphytoflagellates as compared to diatoms. In contrast, at the open-water stations
Fig. 6. Hydrographic cruise, (a) Surface contours of total phytoplankton cell numbers (cells x 10h
I" ). (b) Surface contours of the first axis station scores obtained by correspondence analysis basedon phytoplankton data. The first axis accounted for 33.6% of the total variance.
583
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from
A.Zingone«a/.
flagellates < Wfim
Leptocylindrus danicus
Emitiania huxleyi
Asterionella glacialis
Umbellosphaera tenuis
Chaetoceros spp.
Cerataulina pelagica
naked dinoflagellates <1Sfuv
Rhizosolenia spp.
Thalasslonema nitzschloides
Anoplosolenia brasiliensis
Nitzschia spp.
Oxytoxum variabile
Odontella mobiliensis
Chaetoceros simplex calcitrans
Thalassiosira spp.
Thalassiosira rotula
coast • - • offshore
•0.3 -0.2 -0.1
Fig. 7. Hydrogruphic cruise Absolute contribution of selected phytoplankton taxa to the first axis ofthe correspondence analysis arrangement. Bar length is proportional to the absolute contribution oftaxa, whereas its orientation depends on the score sign.
100
other DiatomsLeptocylindrus danicus
Thalassiosira spp-
Fig. 8. Hydrographic cruise. Surface distribution for percentage abundance of the mainphytoplankton taxa.
584
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton bloom in Gulf of Naples
(Stations 5 and 6) the phytoplankton populations were dominated by smallflagellates (up to 47%). Coccolithophorids, mainly represented by Emilianiahuxleyi, also reached high percentages at these two stations (up to 24%). Amongdiatoms, Leptocylindrus danicus was dominant, together with several Chaeto-ceros species.
The main spatial partitions in phytoplankton community distribution, as theyresult from constrained clustering analysis, are hierarchically ranked in Figure 9.Note the similarity among populations from coastal stations, especially betweenthose from the eastern part of the gulf, and the marked change in populationstructure in the central part of the gulf.
Phytoplankton populations from the production sites (Figure 10) hadpractically the same composition as those of the hydrographic cruise, withdiatoms dominating at coastal stations as well as at the two stations in the centralpart of the gulf (Stations 3 and 19). As compared to stations closer to the coast,Stations 3 and 19 had a lower number of cells, and a more diversified diatompopulation. Species composition and population structure, which were analysedwithin the most productive layer, down to a depth corresponding to 10% of theincident PAR, were generally similar throughout the water column (Figure 11).This occurred both at the two stations in the central part of the gulf, which werealso quantitatively homogeneous throughout the most productive part of thewater column, and at stations closer to the coast, which had surface orsubsurface maxima for cell numbers.
Analogously to data on chlorophyll a, all the four stations previously sampledshowed a notable reduction in cell number, as well as differences in the
Smin>0.7 ox
Smin>0.6 ox
Fig. 9. Hydrographic cruise. Gabriel net links between stations with similarity >0.8. 0.7 and 0.6.
585
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

A.Zingonee/a/.
St. 2-4
In „
St. 18 St. 1O
Innn
St. 12
Fig. 10. Productivity cruise Surface distribution for percentage abundance of the main phyto-plankton taxa.
Table I I . Phytoplankton abundance and similarity index between samplings at the same stations ondifferent sampling dates
Station 1st samplingdate
Cells I"1
on 1st date2nd samplingdate
Cells I"1
on 2nd dateSimilarityindex
3101819
08.0910.10.
11111111
.85
.85
.85
.85
0.382.062.480.42
X
X
X
X
10"If/1
10"10*
09.15.15.14.
11.8511.8511.8511.85
0100
.24
.11
.99
.34
X
X
X
X
10*10"10"10"
0.670.680.700.68
percentage composition of the population compared with data obtained 1-5days earlier (Table II).
Discussion
In the Gulf of Naples, oligotrophic waters of the Southern Tyrrhenian Seaborder on eutrophic coastal waters all year round, although the boundary regiondiffers in extension and distance from the shore, depending on the season(Marino et al., 1984). In autumn 1985, all chemical and biological parametersmeasured were distributed along a coast—offshore gradient, which was steeper inthe area closer to the shoreline, within the isopleth of an averaged salinity of 38PSU, which in this study can be considered the limit of the coastal subsystem(Figure 3a). A smooth gradient for the same parameters was evident even
586
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

cells
l"1x1
06
1 2
3
Fig.
11.
Pro
duct
ivity
cru
ise.
Phy
topl
ankt
on v
ertic
al d
istr
ibut
ion,
> | 5" O o Z
00
at The Open University on December 19, 2010plankt.oxfordjournals.orgDownloaded from

A.7Jngone el al.
between the most offshore stations (Stations 5, 6 and 7). Hence, the boundaryregion between coastal and open waters is wider and farther from the shore inautumn than in other periods of the year. This may be attributed to the fastervertical and advective water movements, typical of autumn and winter, thatfavour the mixing and dilution of terrestrial inputs over larger water volumes(Carrada et al., 1981).
The spatial distribution of phytoplankton populations also reflected the effectthat water mixing and dilution processes had on the fate of terrestrial inputs. Infact, our data demonstrated a great similarity among coastal stations in differentsites of the gulf, which were presumably under the influence of heterogeneousterrestrial inputs. Moreover, differences among coastal and offshore stationswere due to the asymmetric distribution of only a few dominant taxa rather thanto the substitution of the majority of species. Finally, there was a scarcedifferentiation in the vertical distribution of the phytoplankton populationwithin the most productive water layers. On the contrary, a remarkable spatialheterogeneity in phytoplankton communities characterized the innner part ofthe gulf during summer; indeed, populations dominated by different species co-existed in a very restricted area in this season (Zingone et al., 1990a).
Phytoplankton populations at stations that were sampled twice within a fewdays showed a change in total abundance and in the relative abundance for thedifferent species. The short interval (just 1 day in the case of Station 3) and thevariations observed suggest that the changes occurred mainly by way oftransport rather than by in situ evolution of phytoplankton communities. Thestations in the central part of the gulf, where the major changes were found,were influenced by inshore waters. It can be argued that, in this period, theopen-sea current moved in a northwesterly direction, which is the prevailingcondition for Southern Tyrrhenian waters (De Maio et al., 1985). This is alsoconfirmed by the relatively high salinity values in the northwestern part of thesampled area. In contrast, when the open-sea current moves towards thesoutheast, the northwestern and central parts of the gulf are affected by coastalwaters of the adjacent Gulf of Gaeta, which are fertilized by the outfall of theCuma sewage treatment plant and by the Volturno river plume (Carrada et al.,1981).
Phytoplankton populations at coastal stations were characterized by thedominance of a few diatom species, mainly belonging to the genera Thalassiosiraand Chaetoceros. A similar community has already been recorded in autumn incoastal waters of the Gulf of Naples (Carrada et al., 1979). Also late-spring andsummer blooms in coastal waters of the gulf are most often dominated bydiatoms, although in the latter case they comprise smaller sized species, reachingnumbers up to 1.15 x 10* cells 1"' (Zingone et al., 1990a).
Like other diatom-dominated communities frequently found in associationwith autumn blooms in such temperate regions as the western Mediterranean(Estrada, 1982; Estrada etal., 1985) and the eastern coast of the USA (Marshall,1984; Marshal! and Lacouture, 1986; Durbin etal., 1975), these communities areclassifiable in the first stages of the phytoplankton succession (Margalef, 1958)which is restarted whenever new nutrients are supplied to the photic zone.
588
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton bloom in Gulf of Naples
Regardless of the mechanism producing this enrichment, a similar ecologicalresponse is seemingly evoked. In contrast, in Irish coastal waters (Roden, 1984),the west coast of Sweden (Lindahl and Hernroth, 1983; Olsson and Edler, 1991)and the Kiel Bight (Smetacek, 1985; Noji et al., 1986), the autumnal nutrientenrichment determines the development of a late-summer community thatrepresents an advanced stage of the phytoplankton succession, consisting mainlyof dinoflagellates belonging to the genera Ceratium and Gyrodinium. In the KielBight, both types of bloom occur: a dinoflagellate-dominated bloom, then adiatom-dominated bloom.
Phytoplankton abundances at offshore stations were two orders of magnitudelower than in coastal waters, with a higher percentage of coccolithophorids andphytoflagellates. Even lower cell numbers (3 x 104 cells l~'), as well as a moremarked dominance of coccolithophorids and small flagellates, were recorded inautumn 1981 in the open waters of the neighbouring Gulf of Salerno (D.Marinoand A.Zingone, unpublished data).
The most interesting aspect of this study are the values for primary productionobtained at the stations closer to the coast, especially in the western part of thegulf. Although very high (up to 1 g C m~2 day"'), these values are probably notthe highest reached in this area in autumn. In fact, biomass values twice as highhad been measured a few days before, during the hydrographic cruise, at someof the stations. Moreover, a 5 year investigation at the MC station, 2 miles offthe city of Naples, demonstrates that, besides an annual maximum in spring-summer, one or more production peaks occur in October-mid-November thatare often of the same order of magnitude as the main annual peak (Figure 2).These peaks are always shorter in time than the spring-summer peak, especiallywhen they occur in November, because of the high meteorological variabilitythat characterizes this season in the Mediterranean Sea. Thus, the biweeklysampling at the MC station may have failed to detect these short-lasting autumnblooms in some years. For example, in the autumn of this investigation primaryproduction values recorded in November (5 and 22) at this station were ratherlow (219 and 77 mg C m~2 day"1, respectively).
An autumn peak has been described in the annual cycle at various sites in theMediterranean [see Estrada et al. (1985) for a review] and it has generally beenattributed to the recirculation of nutrients from subpycnocline layers into thephotic zones caused by the breakdown of the thermocline (Margalef, 1969).However, this explanation does not apply to our area, because in autumn theonly nutrient-rich water mass (LIW) which could sustain the enrichment wemeasured is confined to —100 m below the lower limit of the photic zone(Carrada et al., 1980), and the presence of a rather sharp thermocline around40 m does not allow for nutrient replenishment from lower layers of the watercolumn. Moreover, such an explanation would not account for the lowproduction values that we recorded in the central part of the gulf.
Neither do other mechanisms proposed to explain autumn blooms in otherareas seem to fit our observations. Water column stratification, which in someshallow, light-limited estuaries reduces turbidity and hence promotes autumnblooms (Pennock, 1985), probably does not exert the same influence in our
589
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

A. Zingone el al.
rather deep coastal waters. Indeed, integrated primary production was very highat both Stations 10 and 12, which had an isopycnal and stratified water column,respectively, within the photic layer. Although enrichment from sediments,induced by the deepening of the thermocline down to the bottom (Smetacek,1985; Noji et al., 1986), cannot be excluded as a fertilizing mechanism, in theGulf of Naples the isobath of 30-40 m, within which this should occur, runs veryclose to the coast. Finally, data obtained at the fixed station MC show that thereis no decrease in planktonic herbivores in autumn that would account for areduced grazing pressure (Mazzocchi and Ribera d'Alcala, 1995), as hypo-thesized for other areas (Samyda, 1957; Cushing, 1975).
The inshore-offshore gradient for productivity, as well as the salinity andnutrient distribution, clearly indicate that the main factors driving productivityin the Gulf of Naples in this study are nutrient inputs of terrestrial origin,possibly enhanced by autumn rains typical of the Mediterranean area in autumn.These inputs are associated with waters that are always less dense than thewarmest surface waters, and their dilution in the surface mixed waters alwaystakes place following wind events. Thus, the autumnal meteorological per-turbations and deepening of the thermocline promote both vertical andhorizontal dilution of nutrients.
In this scenario, a crucial role can be played by the establishment of a periodof meteorological stability, during which calm weather and clear skies lead toirradiance values that can enhance phytoplankton growth, notwithstanding thereduced daylength, which was the case in the present investigation. Our cruisewas conducted during the so-called 'St Martin's Summer', i.e. a shortinterruption of the shift from autumn to winter characterized by mild dry highpressure, which frequently occurs in the first half of November in our region.Unfortunately, besides popular tradition, no systematic study is available for therecurrence of 'St Martin's Summer' which can, however, be likened to a late'Indian Summer' episode. These meteorological singularities have been demon-strated to occur most frequently in three different periods in the northernhemisphere south of 50°N, i.e. 25 September-1 October, 6-22 October and 15-24 November, with a frequency of 38, 66 and 61%, respectively, over a 70 yearperiod (Kalnicky, 1987). Therefore, at least one or more periods of meteoro-logical stability recur every year in autumn between October and mid-Novemberin the northern temperate regions. The high productivity recorded in this study,although not representative of the whole season, could at least be as recurrent asthe meteorological singularities that trigger it. The impact of these singularitieson different coastal and offshore waters will probably vary depending onnutrient availability and local hydrographic features.
As expected from the nutrient distribution pattern, primary production valuesfell dramatically in the central stations of the gulf. We cannot exclude thepresence of a deep chlorophyll maximum below our sampling depth (Estrada,1982; Lohrenz et al., 1988; Estrada et al., 1993). However, this would probablynot contribute significantly to the total water column production because of lightlimitation (Estrada, 1981).
Within the framework of the debate on the contribution of eutrophic coastal
590
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton bloom in Gulf of Naples
areas to the primary production of oligotrophic seas, the case under studydemonstrates that the effects of the enrichment produced by intense municipaland industrial outfalls, coupled with appropriate meteorological conditions,although remarkable at coastal sites, fade rapidly within a few miles from thecoast. However, should high productivity in coastal waters related to climaticsingularities be shown to be a recurrent and widespread phenomenon, the rolethat autumn peaks could play in the annual production cycle is far from beingnegligible, especially if they are phased with the outburst of other compartmentsof the pelagic system, so amplifying the transfer of energy through the trophicnet.
Acknowledgements
We are particularly grateful to Colin S.Reynolds for reading and commentingupon the manuscript. Anonymous referees are sincerely acknowledged for theircritical advice and suggestions.
References
Carrada.G.C. Marino,D.. Modigh.M. and Ribera d'Alcala.M. (1979) On the distribution ofUtermohl phytoplankton in a coastal sub-area of the Gulf of Naples. Rapp. Comm. Int. MerMtdit., 25. 73-74.
Carrada.G C . Hopkins.T.S.. Bonaduce.G.. Ianora.A.. Marino,D.. Modigh.M.. Riberad'Alcala.M. and Scotto di Carlo,B. (1980) Variability in the hydrographic and biological featuresof the Gulf of Naples. P.S.Z.N I Mar. Ecol., 1. 105-120.
Carrada.G.C. Fresi.E.. Marino,D., Modigh.M. and Ribera d'Alcala.M. (1981) Structural analysisof winter phytoplankton in the Gulf of Naples. J. Plankton Res.. 3, 291-314.
Colebrook.J.M. (1979) Continuous plankton records: seasonal cycles of phytoplankton andcopepods in the North Atlantic Ocean and the North Sea Mar. Biol.. 51, 23-32.
Cushing.D.H. (1975) Marine Ecology and Fisheries. Cambridge University Press, Cambridge, pp. 1-278.
De Maio.A.. Moretti.M., Sansone.E.. Spezie.G. and Vultaggio.M. (1985) Outline of marinecurrents in the Bay of Naples and some considerations on pollutant transport. Nuovo Cimento,8C, 955-969.
Durbin.E.G.. Krawiec.R.W. and Smayda.T.J. (1975) Seasonal studies on the relative importance ofdifferent size fractions of phytoplankton in NarTagansett Bay (USA). Mar. Biol., 32, 271-287.
Estrada.M (1981) Biomasa fitoplanctonica y producci6n primaria en el Mediterraneo occidental, apnncipios de otono. Invest. Pesq.. 45. 211-230.
Estrada.M. (1982) Phytoplankton of the western Mediterranean at the beginning of autumn. Int.Rev. Ces. Hydrobiol.. 67, 517-532.
Estrada.M. (1984) Phytoplankton distribution and composition off the coast of Galicia (northwest ofSpain). J. Plankton Res., 6, 417-434.
Estrada.M., Vives,F. and Alcaraz.M. (1985) Life and productivity of the open sea. In Margalef.R.(ed.). Key Environments: Western Mediterranean. Pergamon Press, Oxford, pp. 148-197.
Estrada,M., Marrase\C, Latasa.M., Berdalet.E., Delgado.M. and Riera.T. (1993) Variability ofdeep chlorophyll maximum characteristics in the Northwestern Mediterranean. Mar. Ecol. Prog.Ser., 92, 289-300.
Gabriel,K.R. and Sokal.R.R. (1969) A new statistical approach to geographical variation analysis.Syst. Zool.,%, 259—278.
Grasshoff.K... Ehrhardt,M. and Kremling.K. (1983) Methods of Seawater Analysis. Verlag Chemie,Weinheim, pp. 1-419.
Ignatiades.L. (1984) Coarse-scale horizontal distribution of phytoplankton in a semi-enclosedcoastal area. P.S.Z.N. I Mar. Ecol., 5, 217-227.
Kalnicky.R.A. (1987) Seasons, singularities, and climatic changes over the midlatitudes of thenorthern hemisphere during 1899-1969. J. Climate Appl. Meteorol., 26, 1496-1510.
591
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

A.Zlngpne el al.
Legendre.P. (1987) Constrained clustering. In Legendre.P. and Legendre.L. (cds). Developments inNumerical Ecology NATO AS1 Series Vol. GI4. Springer-Verlag. Berlin, pp. 289-307
Lindahl.O. and Hernroth.L. (1983) Phyto-zooplankton community in coastal waters of westernSweden—an ecosystem off balance? Mar. Ecol. Prog. Ser.. 10. 119-126
Lohrenz.S.E.. Wiesenburg.D.A.. De Palma.I.P.. Johnson.K.S. and Gustafson.D.E. (1988)Interrelationship among primary production, chlorophyll, and environmental conditions in frontalregions of the western Mediterranean Sea. Deep-Sea Res.. 35. 793-810.
Mantoura.R.F.C MartinJ.-M. and Wollast.R. (eds) (1991) Ocean Margin Processes in GlobalChange: Report of the Dahlem Workshop on Ocean Margin Processes in Global Change. JohnWiley & Sons. Chichester. pp. 1-469.
Margalef.R. (1958) Temporal succession and spatial heterogeneity in phytoplankton. In Buzzati-Traverso.A.A. (ed.). Perspectives in Marine Biology. University of California Press. Berkeley.CA. pp 323-349.
Margalef.R. (1969) Composici6n especifica del fitoplancton de la costa catalano-levantina(Meditei-raneo occidental) en 1962-1967. Invest. Pesq., 33. 345-380.
Marino.D.. Modigh.M. and Zingone.A. (1984) General features of phytoplankton communities andprimary production in the Gulf of Naples and adjacent waters. In Holm-Hansen.O., Bolis.L. andGilles.R. (eds). Marine Phytoplankton Productivity. Lecture Notes on Coastal and EstuarineStudies. 8. pp. 89-100
Marshall.H.G. (1984) Phytoplankton distribution along the eastern coast of the USA. Part V.Seasonal density and cell volume patterns for the north-eastern continental shelf. J. Plankton Res .6. 169-193.
Marshall.H.G. and Lacouture.R. (1986) Seasonal patterns of growth and composition ofphytoplankton in the lower Chesapeake Bay and vicinity. Estuarine Coastal Shelf Sci , 23,115-130.
Mazzocchi.M G. and Ribera d'Alcala.M. (1995) Recurrent patterns in zooplankton structure andsuccession in a variable coastal environment. ICES J. Mar. Sci., in press.
Modigh.M.. Ribera d'Alacala.M., Saggiomo.V.. Forlani.G. and Tosti.E. (1985) Time relationshipbetween physico-chemical parameters and biological properties of phytoplankton blooms in theinner part of the Gulf of Naples, summer 1983. Rapp Comm. Int. Mer Mtdit., 29, 109-110.
Noji.T., Passow.U. and Smetacek.V. (1986) Interaction between pelagial and benthal duringautumn in Kiel Bight. I. Development and sedimentation of phytoplankton blooms. Ophelia, 26,333-349.
Olsson.P. and Edler.L. (1991) Dinoflagellate distribution in the southern Kattegat during an autumnbloom. Sarsia, 76, 23-28.
O'ReillyJ.E. and Bush.D.A. (1984) Phytoplankton primary production in the northwesternAtlantic shelf Rapp. P.-V. Rtun. Cons. Int. Explor. Mer. 183. 255-268
Pennock.J.R. (1985) Chlorophyll distributions in the Delaware estuary: regulation by light-limitation. Estuarine Coastal Shelf Sci . 21. 711-725.
Ribera d'Alcala.M., Modigh.M., Moretti.M., Saggiomo.V., Scardi.M., Spezie.G. and Zingone.A.(1989) Una storia infinita. Eutrofizzazione nella Baia di Napoli. Oebalia. 15. 491-501.
Roden.C.M. (1984) The 1980/1981 phytoplankton cycle in the coastal waters off Connemara.Ireland. Estuarine Coastal Shelf Sci.. 18, 485-497.
SCOR-UNESCO (1966) Determination of Photosynthetic Pigments in Sea Water. Monographs onOceanographic Methodology. UNESCO. Paris, pp. 1-64.
Scotto di Carlo,B., Tomas.C.R., Ianora.A., Marino,D., Mazzocchi.M.G., Modigh.M.,Montresor.M., Petrillo.L., Ribera d'Alcala.M., Saggiomo.V. and Zingone.A. (1985) Uno studiointegrato dell"ecosistema costiero del Golfo di Napoli. Nova Thalassia, 7(Suppl. 3), 99-128.
Smayda.T.J. (1957) Phytoplankton studies in lower Narragansett Bay. Limnol. Oceanogr., 2,342-359.
Smayda.T.J. (1983) The phytoplankton of estuaries. In Ketchum.B.H. (ed.). Estuaries and EnclosedSeas. Elsevier, Amsterdam, pp. 65-102.
Smetacek.V. (1985) The annual cycle of Kiel Bight plankton: a long-term analysis. Estuaries. 8.145-157.
Strickland J.D.H. and Parsons.T.R. (1965) A manual of sea water analysis. Bull. Fish. Res. BoardCan.. 125. 1-203.
Takahashi.M., Seibert.D.L. and Thomas,W.H. (1977) Occasional blooms of phytoplankton duringsummer in Saanich Inlet, B.C., Canada. Deep-Sea Res., 24, 775-780.
Walsh J.J. (1988) On the Nature of the Continental Shelves. Academic Press. San Diego, CA, pp.1-520.
Zingone.A.. Montresor.M. and Marino,D. (1985) Temporal and spatial variations of phytoplankton
592
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from

Autumn phytoplankton bloom in Gulf of Naples
communities from the inner part of the Gulf of Naples, summer 1983. Rapp. Comm. Int. MerMtdit., 29, 103-105.
Zingone.A.. Montresor.M. and Marino.D. (1990a) Summer phytoplankton physiognomy in coastalwaters of the Gulf of Naples. P.S.Z.N. I Mar. Ecol.. 11. 157-172.
Zingone.A.. Honsell.G.. Marino.D.. Montresor.M. and Socal.G. (1990b) Fitoplancton. InInnamorati.M., Ferrari.I.. Marino.D. and Ribera d'Alcala.M. (eds). Metodi nell'Ecologia delPlanaon Marino. Nova Thalassia, 11, 183-198.
Received on April 13, 1994; accepted on October 19, 1994
593
at The O
pen University on D
ecember 19, 2010
plankt.oxfordjournals.orgD
ownloaded from




















