
Squamata) from the Early Cretaceous of Las Hoyas, Spain
31
An unusual lizard (Reptilia: Squamata) fkom the Early Cretaceous of Las Hoyas, Spain SUSAN E. EVANS FLS* Department of Anatomy and Developmental Biology, Universig College London, Goixer Street, London WClE 6BT LUIS JAVIER BARBADILLO Unidad de Puleontologia, Departamen to de Biologia, Facultad de Ciencias, Uniuersidad Autdnoma de Mudrid, Campus de Cantoblanco, 28049, Madrid, Spain Received h.lay 1 9 9 7 ; accepted for publication November 19 97 The Early Cretaceous (Barremian)lepidosaurian assemblage of Las Hoyas (Cuenca Provincr, Castilla-La Mancha), Spain, resembles that of roughly contemporaneous localities at Montsec (Catalonia), Ufia (Castilla-La Mancha) and Galve (Aragon) in being dominated by the scincomorph lizard Mqasuurus (hdaesaums). It differs in the presence of two previously unknown taxa, one of which, described here, shows a strikingly unusual morphology in its ribs and distal limb skeleton suggestive of climbing specialization. This form is sufficiently distinct from other known taxa to warrant the erection of a new genus and species, Scandensiu ciervensis gen. et sp. nov. Cladistic analysis of Scandensiu suggests that it forms a sister taxon to living squamates (Iguania + Scleroglossa and all descendants of their most recent common ancestor). 0 1'398 'l'lir Linriraii Socirt) of Lmd~in ADDITIONAL KEY WORDS:-morphology - cladistics ~ climbing ~ Lepidosauria Diapsida - hyoid. CONTENTS Introduction . . . . . . . . . . . . . . . . Material . . . . . . . . . . . . . . . . . Abbreviations . . . . . . . . . . . . . . . Systematic palaeontology . . . . . . . . . . . Scandensia gen.nov. . . . . . . . . . . . . Description . . . . . . . . . . . . . . . . Phylogenetic position . . . . . . . . . . . . . Function and lifestyle . . . . . . . . . . . . . Scandensia ciervensis sp.nov. Acknowkdgements . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . Appendix 1: Characters used in formulation of data matrix Appendix 2: Data matrix used in construction of cladograin . . . . . . . 236 . . . . . . . 236 . . . . . . . 236 . . . . . . . 237 . . . . . . . 237 . . . . . . . 237 . . . . . . . 238 . . . . . . . 247 . . . . . . . 249 . . . . . . . 250 . . . . . . . 251 . . . . . . . 252 . . . . . . . 259 * Corresponding author. ucgasue@,ucl.ac.uk 0 1998 lhc Linncan S(iciety of 1,ondon 235 0024-4082/98/110235+31 $30.00/0 Downloaded from https://academic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 March 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Squamata) from the Early Cretaceous of Las Hoyas, Spain

An unusual lizard (Reptilia: Squamata) fkom the Early Cretaceous of Las Hoyas, Spain
SUSAN E. EVANS FLS*
Department of Anatomy and Developmental Biology, Universig College London, Goixer Street, London WClE 6BT
LUIS JAVIER BARBADILLO
Unidad de Puleontologia, Departamen to de Biologia, Facultad de Ciencias, Uniuersidad Autdnoma de Mudrid, Campus de Cantoblanco, 28049, Madrid, Spain
Received h.lay 1997; accepted f o r publication November 1997
The Early Cretaceous (Barremian) lepidosaurian assemblage of Las Hoyas (Cuenca Provincr, Castilla-La Mancha), Spain, resembles that of roughly contemporaneous localities at Montsec (Catalonia), Ufia (Castilla-La Mancha) and Galve (Aragon) in being dominated by the scincomorph lizard Mqasuurus (hdaesaums). It differs in the presence of two previously unknown taxa, one of which, described here, shows a strikingly unusual morphology in its ribs and distal limb skeleton suggestive of climbing specialization. This form is sufficiently distinct from other known taxa to warrant the erection of a new genus and species, Scandensiu ciervensis gen. et sp. nov. Cladistic analysis of Scandensiu suggests that it forms a sister taxon to living squamates (Iguania + Scleroglossa and all descendants of their most recent common ancestor). 0 1'398 'l'lir Linriraii Socirt) of Lmd~in
ADDITIONAL KEY WORDS:-morphology - cladistics ~ climbing ~ Lepidosauria Diapsida - hyoid.
CONTENTS
Introduction . . . . . . . . . . . . . . . . Material . . . . . . . . . . . . . . . . . Abbreviations . . . . . . . . . . . . . . . Systematic palaeontology . . . . . . . . . . .
Scandensia gen.nov. . . . . . . . . . . . .
Description . . . . . . . . . . . . . . . . Phylogenetic position . . . . . . . . . . . . . Function and lifestyle . . . . . . . . . . . . .
Scandensia ciervensis sp.nov.
Acknowkdgements . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . Appendix 1: Characters used in formulation of data matrix Appendix 2: Data matrix used in construction of cladograin
. . . . . . . 236
. . . . . . . 236
. . . . . . . 236
. . . . . . . 237
. . . . . . . 237
. . . . . . . 237
. . . . . . . 238
. . . . . . . 247
. . . . . . . 249
. . . . . . . 250
. . . . . . . 251
. . . . . . . 252
. . . . . . . 259
* Corresponding author. ucgasue@,ucl.ac.uk
0 1998 l h c Linncan S(iciety of 1,ondon 235
0024-4082/98/110235+31 $30.00/0
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

236 S. E. EVANS AND L. J. BARBADILLO
INTRODUCTION
Four Spanish fossil localities have produced Early Cretaceous lepidosaurs. The lizard Mqasaurus was originally described from Montsec, Catalonia (Vidal, 19 15), as was Hoffstetter’s (1966) Ilerdaesaurus, now known to be a junior synonym of Mqasaurus (Evans & Barbadillo, 1996, 1997). The lizards of Ufia (Cuenca Province, Castilla-La Mancha) have been described by Richter (199 1, 1994a,b), and consist of four taxa: Mqasaurus (Ilerdaesaurus); two paramacellodid scincomorphs, Becklesius and Paramacellodus; and an indeterminate new genus, Cuencasaurus Richter 1 99410, based on a fragment of dentition. In addition, Richter (1 994b) reported the presence of lizard fragments, referable to both Mqasaurus and paramacellodids, at Galve (Teruel Province, Aragon).
The Early Cretaceous (Barremian) lithographic limestone locality of las Hoyas (Cierva Township, Cuenca, Castilla-L!4 Mancha) has yielded a rich flora and fauna including angiosperms, insects, spiders, birds, crocodiles, turtles, an oviraptorid dinosaur and an assortment of amphibians (Sanz et al., 1988; Sanz & Buscalioni, 1992; Ptrez-Moreno et al., 1994; Fregenal-Martintz & Meltndez, 1995; McGowan & Evans, 1995; Evans & Milner, 1996). The lepidosaurian fauna is dominated by the lizard genus Mqasaurus Vidal 1915 (Barbadillo & Evans, 1995; Evans & Barbadillo, 1996, 1997), but two specimens (LH 1 1000 and LH 1 100 1) represent distinct and novel taxa. LHllOOO is a small short-limbed lizard which will be described elsewhere; LHllOOl forms the basis of this article.
MATERIAL.
Las Hoyas (LH) specimens are generally preserved in a fine lithographic limestone matrix which splits into thin laminae. LH 1 100 1 consists of a mainblock (LH 1 100 1 b) bearing most of the skull and skeleton, and a partial counterpart with parts of the jaw, hyoid skeleton and postcranial skeleton.
ABBREVIATIONS
AC astragalocalcaneum acf anterior coracoid fenestra An angular Ar articular Bh basihyal Br braincase Cb 1 , Cb2 ceratobranchials ce centrale Ch ceratohyal c1 clavicle C/mt5 part of calcaneum or metatarsal 5 Co coronoid D dentary dt distal tarsal
Hb iC1 il Is J mt Mx P Pm Pr Pu R ra Ra
hyobranchial interclavicle ilium ischium
metatarsal maxilla parietal premaxilla prearticular pubis right radiale radius
jugal
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 237
e epiph ysis Eb epibranchial ep entoglossal process Fe femur Fi fibula Fr frontal
scf supracoracoid foramen Sp splenial Su surangular Ti tibia U ulna ul ulnare
SYSTEMATIC PALAEONTOLOGY
Lepidosauria Haechl 1866 Squamata Oppel 1811 Scandensia gen. nov.
Derivation ufname. From the Latin scando, to climb.
Gke species. Scandensia ciervensis sp. nov.
Range. Early Cretaceous (Barremian) of Spain.
Diagnosis. As for species.
Scandensia ciervensis sp.nov.
Derivation ufname. From the township of Cierva, close to the type locality.
Holoppe. LH 1 100 1, Museo de Cuenca, Cuenca, Spain.
Gpe localip. Las Hoyas, Cierva Township, Cuenca, Castilla-La Mancha, Spain.
Gpe horizon. Calizas de la HuCrguina Formation (Limestone Unit 111), Early Cre- taceous, Barremian (Fregenal-MartinCz and MelCndez, 1995).
Diagnosis. A small reptile characterized by simple homodont dentition; paired premaxillae with long slender dorsal processes; dentary shallow, with oblique but shallow subdental shelf and ventrally positioned Meckelian fossa; skull bones either unsculptured or with very light sculpturing; frontal probably fused; complement of 12-1 4 imbricating scleral ossicles; well-developed hyoid apparatus with short entoglossal process; amphicoelous vertebral centra, lacking intercentra except in neck and tail; preserved hypapophysis with strong keel and mid-central position; caudal vertebrae autotomous; ribs relatively short, expanded proximally and tapering distally; interclavicle rhomboid, but with strong anterior and posterior processes; angulated clavicles, medially expanded and perforate; scapulocoracoid fenestrate with scapulocoracoid, anterior coracoid but apparently not posterior coracoid fenestrae; in pelvis, pubis moderately elongated with prominent proximal pectineal process, ilium with elongate, tapering blade; forelimb and hindlimb elements of similar length, long and gracile; ulna tapers gradually, but shows distal twist and strong distal epiphysis; radius slender with styloid process; carpal intermedium apparently absent; radiale and ulnare of roughly equal size, triangular centrale, and five distal carpals of which the fourth is the largest; third and fourth metacarpals of roughly equal length, and fifth short; phalangeal formula 2:3:4:5:3; femur long and
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

238 S. E. EVANS AXD L. J. BARBADILLO
Figure 1. Scandensis ciwvensis gen. et sp. nov., holotype-LH11001, Las Hoyas, Spain. A, skeleton on mainblock; B, outline figure of skeleton on mainblock. Scale bars: A = 10 nim; B = 1 mni.
slender; tibia somewhat shorter and quite robust; fibula slender but with triangular medial expansion roughly one third of distance from proximal head; foot somewhat longer than hand; metatarsals 3 and 4 roughly equal in length, fifth unknown; penultimate phalanges slender and elongate, equal or exceed the metapodials in length.
DESCRIPTION
LH 1 1001 comprises a small skeleton (snout-pelvis length 25 mm, total length roughly 50 mm) in which the skull is poorly preserved, but the postcranial skeleton is fully articulated and shows fairly good, if variable, preservation (Fig. 1A,B). The rather weak ossification and detached epiphyses suggest immaturity, but the carpus is fully formed, with geometrical rather than circular elements, and the animal was probably subadult.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 239
A
Fi'gure 2 . Scandensia ciervensis gen. et sp. nov., holotype-LH11001, Las Hoyas, Spain, skull. A, view of skull as preserved on mainblock; B, superimposition of bones from main and counterpart blocks, with left side bones shown in black, and midline elements dotted; right side bones have been omitted. Scale bar = 1 mm.
Skull The skull is preserved in left dorsolateral view but the roofing and temporal bones
are very damaged (Fig. 2A,B). The premaxillae are paired, with long slender ascending processes. There were at least four tooth positions on each side. The maxilla has a slender anterior process and a deep facial process but its alveolar margin is broken away. In the skull roof, the nasals are paired and border moderate- sized terminal nares. Posteriorly, they appear to have broadened and may have met the prefrontals. The frontals were probably fused and roughly parallel-sided. The cristae cranii (olfactory or descending flanges) are deep but do not approach one another in the midline. The parietal has been split between the two blocks and is therefore difficult to reconstruct. It seems to have been a broad, thin bone, without strong lateral borders and with at best light sculpture. The adductor muscles were probably confined to the ventral surface and the upper temporal fenestrae may have been reduced.
The circumorbital bones are fragmentary but clearly formed a complete and relatively robust series. They surround a series of imbricating scleral ossicles (1 2-14 elements). The prefrontals are large but the presence or absence of a lacrimal cannot be confirmed. The post- and preorbital bones did not meet. Parts of both jugals are preserved, the postorbital portion is broad and strong, but there is a long narrow suborbital process which would have extended into the anteroventral orbital rim- contacting either the prefrontal or lacrimal (if present) (Fig. 2B).
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

240 S. E, EVANS AND L. J. BARBADILLO
The braincase has been split open and shows the channels and grooves of the bony labyrinth; a large mass on the mainblock appears to correspond to a broad basioccipital. The impression is of a relatively large structure with short paroccipital processes, overlain to a large degree by the parietal.
Palatal elements are preserved as fragmented sheets, but show no detail of structure. None, however, bears any trace of teeth.
Lower jaw The mandibles of LH 1 100 1 are relatively poorly preserved but their basic features
can be reconstructed from elements on the part and counterpart blocks. The right dentary is visible below and deep to the left maxilla, but the implantation
and lingual detail are obscured. There is a wide but shallow subdental shelf which restricts the Meckelian fossa to the ventral margin. The left dentary has separated from the skull and is visible in lateral view (Fig. 2A). It is shallow and bears a line of small sensory foramina but no obvious ventral muscle scar. There are teeth in eight positions but the spacing suggests rapid replacement (and therefore a higher number of actual, if unfilled, tooth positions). These teeth are simple and homodont, with some labiolingual compression, in contrast to the teeth of the contemporary Meyasaurus which are strongly bicuspid at the rear of the jaw, even in very young animals. The teeth of Scandensia bear some resemblance to those of Cuencasaurus from Uiia (Richter, 1994b), but the status of this taxon is problematical. The single fragmentary holotype specimen consists of a portion of a tooth row, described as a dentary but equally possibly a maxilla. The simple teeth show a general similarity to those of Scandensia, but the fragmentary nature of the material renders detailed comparison impossible.
The counterpart block preserves the rear of the left mandibular ramus in medial view (Figs 3B, 4A). An anterior impression shows the rear of the dentary to have been bifurcate, providing a space for a bluntly tapering surangular. The upper dentary process is more slender than the lower but they are of similar length. The surangular extends the full length of the postdentary mandible. It is overlain anteriorly by an element which tapers posteroventrally. Seen in cross-section (Fig. 4B), this element is thin and is separated from the surangular and dentary by a small space. It must be the posterior end of the splenial. Behind this splenial and below the surangular is a long slender prearticular. It can be followed towards the distal end of the bone but the articular is not preserved. The adductor fossa is small. Overlying the junction of the surangular and prearticular is a thicker triangular fragment which is probably part of the coronoid. Finally, between the splenial and surangular anteriorly, there is a further thin slip of bone with little medial exposure. It is presumably a small angular.
On the right side, the articular is preserved, its retroarticular process appears parallel-sided and relatively untwisted.
Hyoid shbton The lepidosaurian hyoid apparatus has been described in many taxa (Tanner &
Avery, 1982). As seen in Sphenodon and in the primitive lizard condition (e.g. Fig. 3C,D), it consists of a central unpaired basihyal associated with three pairs of rod- like attachments (reducing to two or one pair in more derived forms). The basihyal
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022
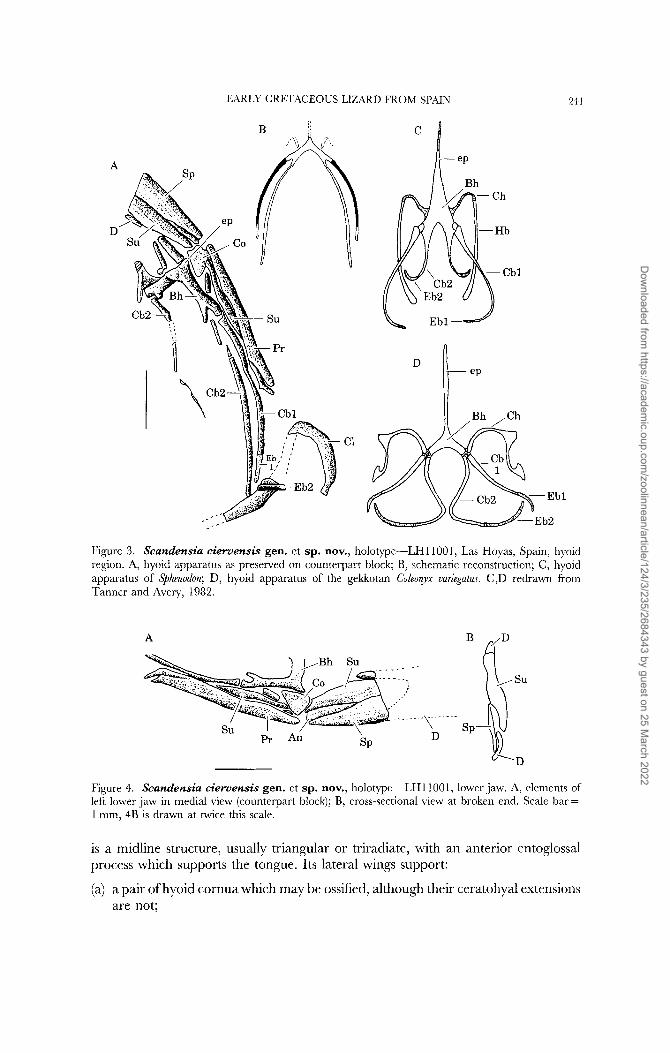
EARLY CRETACEOUS LIZARD FROM SPAIN 2-1 1
A
D’
Bh
Figure 3. Scandensia cieruensis gen. et sp. nov., holotpe-LH11001, Las Hoyas, Spain, hyoid region. A, hyoid apparatus as preserved on counterpart block; B, schematic reconstruction; C, hyoid apparatus of Sphenodon; D, hyoid apparatus of the gekkotan Coleorpx uuriegatus. C,D redrawn from Tanner and Avery, 1982.
A dD
Figure 4. Scandensia cieruensis gem et sp. nov., holotpe-LH11001, lower jaw. A, elements of left lower jaw in medial view (counterpart block); B, cross-sectional view at broken end. Scale bar = 1 mm, 4B is drawn at twice this scale.
is a midline structure, usually triangular or triradiate, with an anterior entoglossal process which supports the tongue. Its lateral wings support:
(a) a pair of hyoid cornua which may be ossified, although their ceratohyal extensions are not;
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

242 S. E. EVANS AND L. J. BARBADILLO
(b) a pair of ossified first ceratobranchials extending at their tips into short car- tilaginous epibranchials;
(c) a pair of cartilaginous second ceratobranchials which may be continuous with the basihyal (as posterior extensions) or may be jointed with it running back parallel to the first ceratobranchials. The second ceratobranchial may also be tipped with an epibranchial (Fig. 3 C,D).
Variation exists both within and between groups as to the retention, form and size of the second ceratobranchial element, the presence or absence of the second epibranchial and its continuity with the ceratobranchial, and the length and shape of the hyobranchial elements (Estes, Gauthier & de Queiroz, 1988; Tanner & Avery, 1982).
The hyoid apparatus lies in the floor of the mouth and has no bony connection with the skull. It is consequently rarely preserved in fossils, but LHllOOI is an exception. The hyoid lies to one side of the braincase on the counterpart block (Figs 2B, 3) and was exposed by the removal of a thin layer of matrix.
The basihyal is crescent-shaped, extending anteriorly into a narrow entoglossal process. Distally, each arm of the basihyal bifurcates into two short processes, of which the anterior is the thicker. The structure is clearest on the right side (as viewed) where two long rods articulate with these processes (fragments of the contralateral pair can also be seen, Fig. 3A). The more lateral element is ossified except at its tip where there is clearly a separate cartilaginous epibranchial element. The medial rod is of similar length and shape. It appears to be cartilaginous and its connection with the basihyal is more slender. It ends close to the clavicle but a second shorter element extends across the clavicular stem and may be another epibranchial.
Since only two pairs of elements are visible they could be alternatively interpreted as the ceratohyal and first ceratobranchial, or the first and second ceratobranchials. The latter seems more probable since the more lateral element of the pair is ossified and has a short epibranchial associated with it. The ceratohyal does not take this form. The hyoid arch is therefore missing but its proximal end may be represented by the short bar of bone associated with the basihyal.
As preserved, the hyoid apparatus of Scandensia shows a primitive structure in retaining both ceratobranchial elements and their epibranchials, and in the simple trifurcate shape of the basihyal. As reconstructed (Fig. 3B), the ceratobranchials were well-separated in the midline.
Axial skeleton The preservation of the skeleton on two blocks makes an exact vertebral count
difficult, but there were an estimated 25 or 26 presacral vertebrae, of which eight are cervicals. The vertebral centra lack condyles and are notochordal and amphicoelous. Neural spines appear to have been broad based but low.
The cervical centra are short, with short arch pedicels (Fig. 5A). Hypapophyses are not well preserved, but one is visible below cervical vertebra 5, in a roughly midcentral position, and has a well-developed keel. Dorsal vertebrae are slightly longer (Fig. 5B), with square amphicoelous centra; there is no trace of persistent intercentra in this region. Two sacral vertebrae are followed by a series of 3 or 4 anterior caudals bearing strong broad transverse processes, which curve forward and provided a strong base to the tail (Fig. 5C). The remainder of the tail (16 +
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 243
A
.-.-
B
C
Figure 5. Scundensiu cimensis gen. et sp. nov., holotpe-LH11001, vertebrae. A, dorsomedial view of anterior cervical vertebrae showing amphicoelous condition of the centrum; B, outline view of middorsal vertebrae to show relationship in size between ribs and vertebral centrum; C, anterior caudal vertebrae, ventral view. All scale bars = 1 mm.
vertebrae) is narrow with fully autotomous vertebrae (the fracture plane dividing the centrum into roughly equal parts). Assuming most of the tail to be preserved (it tapers rapidly from its base), it is relatively short (roughly equal in length to the presacral column).
The first clear rib is on the eighth vertebrae, although there must have been short ribs in more anterior positions. The dorsal ribs are characterized by their relatively wide proximal parts @resumably for muscle attachment) which taper to a narrow distal end (Figs 1, 5B). Such ribs continue to the end of the presacral series and remain robust. It is striking that while the vertebrae themselves are thin-walled and delicately built, the ribs that articulate with them are thick and heavily ossified. A somewhat similar pattern (in the vertebrae at least) is seen in some crevice-living modern lizards (e.g. the gekkotans Tarentola and Hemidacplus; lacertids Podarcis hispanica, AlQroides marchi, LJB pers. observ.), often in conjunction with low neural spines and a dorsoventrally compressed body. It is possible that the unusual rib shape reflects strong axial muscle attachments (longissimus, iliocostalis) used to flatten the body out.
Pectoral girdle and forelimb Elements of the pectoral girdle are best seen on the left side of the main block
(Fig. 6), although the counterpart provides some information. As with other specimens from Las Hoyas, both bony and cartilage skeletal parts are preserved.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

241 S. E. EVANS AND L. J. BARBADILLO
U
Figure 6. Scandensiu cierumsis gen. et sp. nov., holotype-LH11001, pectoral girdle. A, lrft scapulocorocoid, ventral region, and interclavicle as seen in lateral view; B, interclavicle, as preserved, with block reorientated; C, reconstruction of interclavicular shape. Scale bar = 1 mm.
The scapulocoracoid is clearly visible, at least in its ventral part. It is fenestrated with scapulocoracoid and anterior coracoid fenestrae separated by a strong spur of bone. Below the anterior coracoid fenestra, and separated from it by another thickened spur, is a region of extremely thin bone in the position of the posterior coracoid fenestra. The scapular blade is not visible. From the right side, there is evidence of a large suprascapula. The scapulocoracoid is bordered anteriorly by a cartilaginous body passing from central sternal cartilage along the ventral and anterior border of the scapulocoracoid. This is the mesosternum and extends at least to the level of the bar separating the scapulocoracoid and anterior coracoid fenestrae.
The interclavicle is partially preserved and has an unusual shape, being rhomboid rather than cruciform, with lateral arms, a very thin tapering posterior process and a well-developed anterior process (Fig. 6B,C). The anterior process is an indication of large medially expanded clavicles, and one of these is preserved on the counterpart block. It is perforate and tapers laterally into a narrow portion which angles sharply away from the midline, apparently meeting the scapula rather than the suprascapula dorsally.
The right humerus is partially obscured, but shows the elbow clearly. The humeral and olecranon epiphyses are incompletely ossified and the latter is detached. The bones of the hand are disrupted but the phalangeal arrangement is clear. The left humerus is partially preserved on the counterpart block, while the forearm and hand are perfectly preserved and in articulation, although the digits (fully articulated) have been separated from their metacarpals (except in digit five) (Fig. 7A), presumably as a result of postburial compression of the matrix.
All the forelimb elements are gracile and elongated, and they are nearly equal in length to their hindlimb counterparts (9l0/o H/F and R,U/T) (Fig. 9C,D). The humerus retains a small ectepicondylar foramen. The radius is thinner than the ulna and 70% of the humeral length; it ends distally with a small epiphysis. The ulna is broadest proximally, tapering gradually distally, but then narrowing abruptly
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022
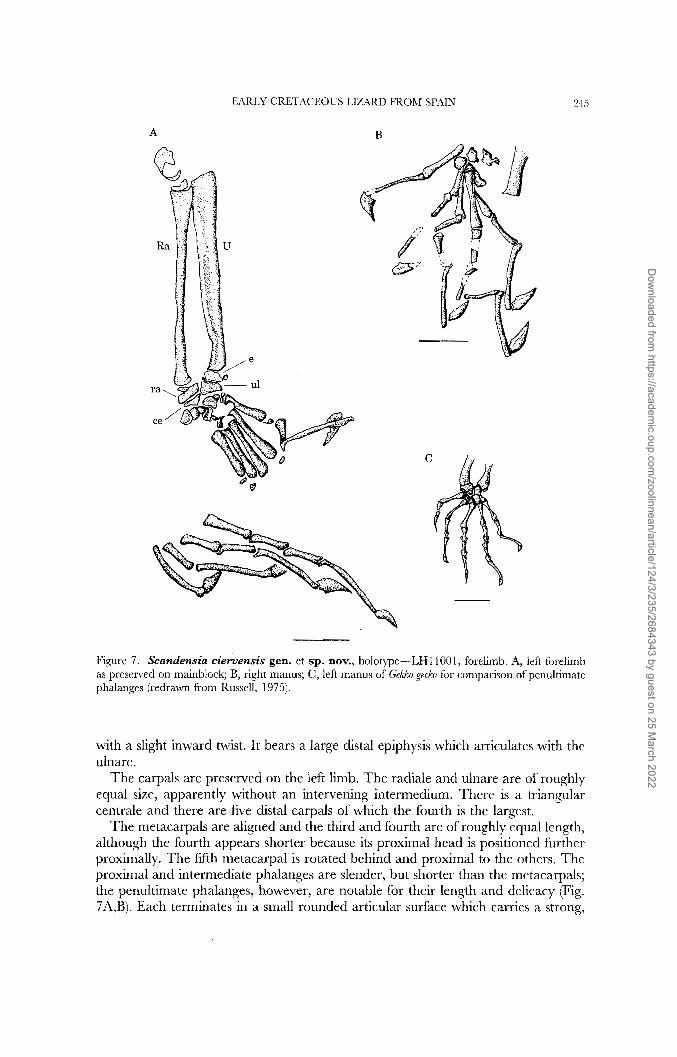
A
EARLY CRETACEOUS LIZARD FROM SPAIN
B
2-15
C
Figure 7. Scandensia ciervensis gen. et sp. nov., holotype-LHl1001, foreiimb. A, left forelimb as presened on mainblock; B, right manus; C, left manus of GeMo gecko for comparison of penultimatc phalanges (redrawn from Russell, 1975).
with a slight inward twist. It bears a large distal epiphysis which articulates with the ulnare.
The carpals are preserved on the left limb. The radiale and ulnare are of roughly equal size, apparently without an intervening intermedium. There is a triangular centrale and there are five distal carpals of which the fourth is the largest.
The metacarpals are aligned and the third and fourth are of roughly equal length, although the fourth appears shorter because its proximal head is positioned further proximally. The fifth metacarpal is rotated behind and proximal to the others. The proximal and intermediate phalanges are slender, but shorter than the metacarpals; the penultimate phalanges, however, are notable for their length and delicacy (Fig. 7A,B). Each terminates in a small rounded articular surface which carries a strong,
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

246
A
S. E. EVANS AND L. J. BARBADILLO
Figure 8. block, left pubis and ischium; B, outline reconstruction of pelvic elements. Scale bar = 1 mm.
Scandensia ciewensis gen. et sp. nov., holotype-LH11001, pelvis. A, counterpart
deep (but mediolaterally thin) claw. The same arrangement is seen in the foot, and most closely resembles that of some derived scansorial geckos (Russell, 1975; Russell & Bauer, 1990). The phalangeal formula is 2:3:4:5:3. In total, the hand slighty exceeds the length of the humerus and is a little over 77% of the length of the pes.
Pelvic gzrdle and hind limb The pelvic girdle is best preserved on the right side. The ilium has been rotated
so that its apex (still cartilaginous) faces forward. The blade is long and tapering but provides little detail. The pubis differs from that of Meyusuum in being generally somewhat longer and more gracile (Fig. 8A). It has a well-developed, proximally- placed pectineal process, directed posterodorsally (i.e. without a ventral inflexion) and a proximal obturator foramen. The ischium is much shorter with no obvious processes, tubercles or expansions (Fig. 8).
The femur is long and sigmoid, with a slender shaft and an incompletely ossified epiphysial expansion distally. The tibia is shorter and somewhat more robust, with a strong, broad head and a shaft that remains sturdy. The distal head is not clearly preserved, but appears to lack the notch for the astragalocalcaneum typically found in scleroglossan lizards (Estes et ul., 1988). The fibula is much thinner with a narrower proximal head, a shaft of almost constant width and a slight expansion at the distal end. However, at the junction of the proximal one-third of the shaft and the remainder, there is a pronounced triangular expansion which overlaps the tibia and probably corresponds to the position of insertion of the iliofibularis muscle (Fig. 9A,
The preservation of the ankle and foot is, unfortunately, rather poor. The ankle is partially preserved on the left in what is apparently a dorsal view, so that the medial metatarsals overlap the lateral ones. A medially placed element appears to be the astragalus, with tibia1 and partial fibular facets. This articulates laterally with a broken element, presumably the calcaneum, and distally with a large fourth distal tarsal. At least one additional distal tarsal is preserved but interpretation is complicated by the presence of several detached metatarsal epiphyses.
C).
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 247
,-- .._ < . C D n
B AC
Figure 9. Scandensiu cimensis gen. et sp. nov., holowe-LH11001, hindlimb. A, hindlimb elements as preserved on mainblock; B, enlargement of left ankle region; C and D, hind (C) and forelimb (D) in schematic reconstruction to compare lengths of individual components.
In the same left foot, the heads of metatarsals two, three and four are in articulation with the tarsus, but metatarsal five is missing (its proximal part may be amongst the broken material proximal and lateral to metatarsal four). The metatarsals are relatively longer than the metacarpals but follow the pattern of the hands, with the third and fourth of similar length. The similarity also extends to the short proximal phalanges and the long slender penultimate phalanges bearing deep claws.
In total, the foot is somewhat longer than the hand, with the long penultimate phalanges being roughly equal in length to metatarsals. The phalangeal formula is 2:3:4:5:3/4.
PHYLOGENETIC POSITION
Scandensia shows a unique combination of character states which distinguishes it from other lepidosaurian reptiles, both extant and extinct. Unfortunately, the poor preservation of the skull hampers discussion of relationships. On the basis of foot
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

218
Cordyliformes
- Scincidae
S. E. EVANS AND L. J. BAKBADILLO
-
-! Xantusiidae
I '
IGUANIA
GEKKOTA
SNAKES
AMPHISBAENIA
Dibamidae
Helodermatidae
Varanus
Lanthanotus
Anguidae
Xenosauridae
Bavarisaurus
OUTGROUP
Figure 10. Cladogram showing hypothesis of relationships for Scandensia.
and vertebral structure, placement in or close to Gekkota might seem the best solution, but assumptions of gekkotan relationship have been wrong in the past (e.g. Bauarisaurus, Ardeosaurus, Estes, 1983, Evans, 1993). For that reason, we ran a cladistic analysis using a matrix of 254 characters (taken mostly from Estes et al., 1988; Gauthier, Estes & de Queiroz, 1988) and a total of 26 taxa, including those constituting the outgroup (Saurosternon, Marmoretta, kuehneosaurs and rhynchocephalians). Data for living taxa was taken mostly from Estes et al. (1988) and Gauthier et al. (1988), with data on the fossils from published and unpublished work by one (SE) or both of the current authors (Evans, 1991, 1993, 1994, work in progress; Evans & Barbadillo, 1997; Evans & Chure, in press a,b; Waldman & Evans, 1994). The matrix was run using the heuristic search mode of PAUP version 3.1.1 (Swofford, 1993) with multiple states treated as polymorphisms. This resulted in six equally parsimonious trees of length 840 steps (CI, 0.73; RI, 0.58; RC, 0.42). A strict consensus tree is given in Figure 10. Most of the variation between the trees was in the relative positions of the outgroup taxa. Three trees placed Eichstaettisaurus and Scandensia as sister taxa, the remaining three placed Eichstaettisaurus and then Scandensia as successive sister taxa to Ardeosaurus + the crown group. The position of Bavarisaurus remained unchanged.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SP.4IN 2.1.9
The observation that some Jurassic and early Cretaceous taxa (e.g. Mqasaurus, Paramacellodus) consistently nest within the crown group, while others (e.g. Bavarisaurus, Ardeosaurus, Scandensia) equally consistently fall outside it, was first made by Reynoso (1 996a,b). The crown group share a suite of character states (subsequently reversed in some modern families) including: jugal forms ventral margin of orbit (25); dorsal process of the squamosal lost (28); reduction/loss of pterygoid tooth rows (73); nasals reduced in width (130); reduction of palatal flange of pterygoid (137); shortened angular (1 46); coronoid eminence formed by coronoid alone (1 47); anterior cervical ribs single-headed (1 48); gastralia absent (1 64); loss of quadratojugal(l98); reduction of quadrate/pterygoid overlap (203).
A further suite places Ardeosaurus, Eichstaettisaurus and Scandensia above Bavarisaurus including: forked postfrontal (1 3); loss of trunk intercentra (86); anterior coracoid emargination (94); a perforate clavicle (97); and a cruciform interclavicle (1 0 l), although some of these states may subsequently be found to be more inclusive. In this analysis, two characters place Ardeosaurus closer to the crown-group, a fused parietal (1 7) and procoelous vertebrae (82), although the state of the latter is uncertain in Eichstaettisaurus. It now seems likely that the primitive condition of the vertebral centra in stem squamates is amphicoely, with procoely characterizing the crown- group, and subsequent reversal within Gekkota.
The clustering of gekkotans, dibamids, amphisbaenians and snakes was a consistent feature in the data runs performed for this analysis, and was unaffected by the inclusion or deletion of Scandensia. However, while the position of Scandensia and the other stem taxa is unchanged by the addition of further crown group taxa (e.g. mosasaurs, dolichosaurs), these taxa do affect the stability of the gekkotan-snake- amphisbaenian clade, raising alternative arrangements within the crown group-most notably the grouping of snakes with varanoid lizards (as recently proposed by Lee, 1997), or gekkotans with scincomorphs (as suggested by Presch, 1988). A number of research groups, including ourselves, are currently addressing these questions but they are beyond the scope of the present paper.
FUNCTION AND LIFESTYLE
The most common members of the Las Hoyas assemblage are aquatic animals (fish, crocodiles, salamanders, frogs) and those that may have lived and foraged close to the edges of the lake (e.g. the lizard Meyasaurus, Evans & Barbadillo, 1997). More terrestrial elements, like the dinosaur Pelecanimimus (Perez-Moreno et al., 1994) and the lizard described here, are much rarer and would have occupied the more arid hinterland, living amongst the rocks and vegetation. They were presumably carried into the lake by small surface streams. Several aspects of the postcranial skeleton of Scandensia are peculiar (the lightly built vertebrae, thick ribs, long hands and feet) and suggest a specialized lifestyle, but it has proved impossible to match them exactly in any living lizard. As observed above (Description), some features of the vertebrae and ribs resemble those of crevice dwelling recent lizards which can flatten the body dorso-ventrally. In addition, the relatively long gracile limbs, the long penultimate phalanges, and the similarity in length between the fore and hind limbs, are traits found in some climbing lizards. In order to consider the significance of these limb parameters more objectively, we ran a principal components analysis combining
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

250 S. E. EVANS AND L. J. BARBADILLO
A
G G SA
X
PC4
B
X G G I I S
G G
c S S I A
€I1 :I I
I A G G A] c I s C I I G G I I l l 1 I I
I
I1 I 1 1
PC5
Figure 1 1. Plot of results from principal components analysis of body and limb parameters performed using 55 taxa of recent lizards and LHIIOOI. The plots shown are those of principal component 2 (relating mostly to head length and snout-vent length) against (A) PC 4 and (B) PC 5, both of which relate to foot parameters, especially those of the penultimate digit. Both plots highlight the distinctive form of the hand and foot in LHl 1001 in comparison with modern lizards. Key: A, anguimorphs; G, gekkotans; I, iguanians; S, scincomorphs; X, LH11001.
data on Scandensia with an existing data set based on 22 scaled skeletal measurements taken from a range of modern lizard taxa (n = 55). Two of the resulting plots are shown in Figure 1 1. They show the second principal component (relating mostly to snout-vent length and head length) plotted against the fourth (Fig. 11A) and fifth (Fig. 11B) principal components, both of which are related to digital proportions. On a general level, the grouping of Scandensia with predominantly climbing lizards such as gekkotans and chamaeleons, lends support to our interpretation of its lifestyle. The fact that Scandensia stands somewhat apart is evidence of its phalangeal specialization. However, its closest neighbours are derived scansorial gekkotans such as Gekko and Homopholis and it is possible that Scandensia had independently acquired digital climbing pads of some form.
ACKNOWLEDGEMENTS
Our thanks are due to Professor J.-L. Sanz and his group at the Autbnoma Universidad de Madrid for their support throughout this work; to, Sefior Armando Diaz-Romeral who found the specimen described here and made it available for study; and to Dr Victor-Hugo Reynoso, Mexico, for discussions on early lizard
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 25 1
relationships. This work was supported under an EC Capital and Mobility Pro- gramme grant, and by funding from the Generalitat of Castilla-La Mancha. Dr Paul O'Higgins (UCL) assisted us with the Principal Components Analysis; Aysha Raza (UCL) prepared the ink figures and collected most of the skeletal measurements of living taxa.
REFERENCES
Barbadillo J, Evans SE. 1995. Lizards (Reptilia: Squamata) from the Early Cretaceous of Las Hoyas, Spain. Second International Symposium on Lithographic Limestones, Lleida-Cuenca, 1995. Elctended Abstracts pp. 27--28. Universidad Autonoma de Madrid.
Estes R. 1983. Sauria terrestria, Amphisbaenia. In: Wellnhofer P, ed. Handbuch der Palaoherpetologze, 1 OA. Stuttgart: Gustav Fischer Verlag.
Estes R, Gauthier J, de Queiroz K. 1988. Phylogenetic relationships within Squamata. In: Estes R, Pregill G, eds. Phlogenetic Relationships of the Lizard Families. Stanford: Stanford University Press,
Evans SE. 1991. A new lizard-like reptile (Diapsida, Lepidosauromorpha) from the Middle Jurassic of Oxfordshire. zoological Journal ofthe Linnean SocieQ 103: 39 1-41 2.
Evans SE. 1993. Jurassic lizard assemblages. Second Ge0Ige.e.r Cuvier Symposium. Revue Paliobiologie, Volume Sppciale 7: 55-65.
Evans SE. 1994. The Solnhofen (Jurassic: Tithonian) lizard genus Bavarisaurus: new skull material and a reinterpretation. Neues Jahrbuch f u r Geologie und Palaontologie Abhandlungen 192: 37-52.
Evans SE, Barbadillo J. 1996. The early Cretaceous lizards of Montsec (Catalonia: Spain). Treballs del Museu de Geologia de Barcelona 5: 5-1 3.
Evans SE, Barbadillo J. 1997. Early Cretaceous lizards from Las Hoyas, Spain. zoologiralJourna1 OJ' the Linnean Socieo 119: 23-49.
Evans SE, Chure DC. 1998a. Morrison lizards: structure, relationships and biogeography. Modern Geology 23: 35-48.
Evans SE, Chure DC. 1998b. Paramacellodid lizard skulls from the Jurassic Morrison Formation at Dinosaur National Monument, Utah. Journal of Ertebrate Paleontology 18:99-114.
Evans SE, Milner AR. 1996. A metamorphosed salamander from the early Cretaceous of Las Hoyas, Spain. Philosophical 7ransactions of the Royal SocieQ o f london, B 351: 627-646.
Fregenal-MartinCz MA, Melendez MN. 1995. I. 1. Geological Setting. In: Meltndez MN, ed. Lm Hoyas. A lacustrine konseruatlagerstatte, Cuenca, Spain. II International Symposium on Lithographic Limest0ne.r. Field trip guide book-Juh 16, 1995. Madrid: Ediciones de la Universidad Autonoma de Madrid, 1-10,
Gauthier J, Estes R, de Queiroz K. 1988. A phylogenetic analysis of Lepidosauromorpha. In: Estes R, Pregill G, eds. Pblogenetic Relationships of the Lizard Families. Stanford: Stanford University Press, 15-98.
Hoffstetter R. 1966. Les Sauria ( = Lacertilia) du Jurassique supkrieur du Montsech (Espagne). Bulletin de la Sociite giologique de France 7: 549-557.
Lee MSY. 1997. The phylogeny of varanoid lizards and the affinities of snakes. Philosophical Transactions ofthe Royal SocieQ, B, 352: 53-97.
McGowan G, Evans SE. 1995. Albanerpetontid amphibians from the Cretaceous of Spain. Nature, London 373: 143-145.
Perez-Moreno BP, Sanz JL, Buscalioni AD, Moratalla JJ, Ortega F, Rasskin-Gutman D. 1994. A unique multitoothed ornithomimosaur dinosaur from the Lower Cretaceous of Spain. Nature, London 370: 363-367.
Presch W. 1988. Phylogenetic relationships of the Scincomorpha. In: Estes R, Pregill G, eds. Phlogenetic Relationships ofthe Lizard Families. Stanford: Stanford University Press, 47 1-492.
Reynoso VH. 1996a. ,4 primitive lizard from the early Cretaceous of Mexico and the phylogenetic position of early lizards. Journal of Ertebrate PaleontoloQ 16: 60A.
Reynoso VH. 199613. Early Cretaceous lepidosaurs (Reptilia: Diapsida) from Central Mexico and the phylogeny of lepidosauromorphs. Ph.D Thesis, McGill University, Montreal, Canada.
Richter A. 1991. A new species of Ilerdaesaurus and other lizards from Ufia, Spain (Lower Cretaceous). In: Kielan-Jaworowska Z, Heinz N, Nakrem HA, eds. F$h Symposium on Mesozoic Tierestrial Ecosystems and Biota, Extended AbJtracts. Contributions o f the Palaeontological Museum, Uniuersip o f Oslo 364: 55-56.
119-281.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

252 S. E. EVANS AND L. J. BARBADILLO
Richter A. 1994a. Der problematische Lacertilier Zlerdaesaums (Reptilia: Squamata) aus der Unter- Kreide von Ufia und Calve. Berliner geowissenschaftliche Abhandlungen 13: 135-161.
Richter A. 199413. Lacertilia aus der Unteren Kreide von Uiia und Galve (Spanien) und Anoual (Marokko). Berliner geowissenschaftliche Abhandlungen 14: 1-147.
Russell AP. 1975. A contribution to the functional analysis of the foot of the Tokay, Gekko gecko (Reptilia: Gekkonidae). Journal of<oology 176: 437-476.
Russell AP, Bauer A. 1990. Digits in pad-bearing gekkonine geckos: alternate designs and the potential constraints of phalangeal number. Memoirs o f the Queensland Museum 29: 453-472.
Sanz JL, Wenz S, Yebenes A, Estes R, Martinez-Declos X, Jimenez-Fuentes E, Dieguez C, Buscalioni A, Barbadillo LJ, Via L. 1988. An early Cretaceous faunal and floral assemblage: Las Hoyas fossil-site (Cuenca, Spain). Geobios 21: 61 1-638.
Sanz JL, Buscalioni AD. 1992. A new bird from the Early Cretaceous of Las Hoyas, Spain and the early radiation of birds. Palaeontology 35: 829-845.
Swofford DL. 1993. PAUP: Phylagenetic Analysis Using Parsimory. Version 3.1. Computer program, distributed by Illinois Natural History Survey.
Tanner W, Avery D. 1982. The buccal floor of reptiles, a summary. Great Basin Naturalist 42: 273-349. Vidal LM. 1915. Nota geologica y paleontologica sobre el Jurasico superior de la provincia de Ltrida.
Waldman M, Evans SE. 1994. Lepidosauromorph reptiles from the Middle Jurassic of Skye. <oological Boletin del Instituto Geolbgico de Espaiia 36: 17-55.
Journal o f the Linnean Socieg 112: 135-150.
APPENDIX 1: CHARACTERS USED IN FORMULATION OF DATA MATRIX
Character coded 0,1 etc but polarity not fixed; character states unordered; multistate characters considered as polymorphisms.
Combines/extends characters from Estes et al. (1988) and Cauthier et al. (1988), with one original character (253). The principal changes have been to combine two or more 2 state characters into multi-state characters, or to extend characters so that they apply also to more primitive taxa.
Characters shown in italics were designated uninformative (PAUP 3.1.1) with respect to the taxa included in the data runs and were therefore ignored in the tree searches. 1. Ontogenetic fusion of premaxillae: (0) paired well into postembryonic ontogeny; ( I ) fused in embryo. 2. Bony external nares extent: (0) opening not extended posteriorly, does not reach frontal; ( I ) opening extended posteriorly reaching or nearly reaching frontal. 3. Ontogenetic fusion of nasals: (0) paired well into postembryonic ontogeny; (1) fused in embryo. 4. Nasal-prefrontal contact: (0) present; (1) anterior process of frontal separates nasal and prefrontal. 5. Prefintal contact with posterior orbital bones: (0) does not contact posterior orbital bones; (1) does contact above orbit. 6 . Ontogenetic fusion of frontals: (0) paired well into postembryonic ontogeny; (1) fused in embryo or early posthatchling. 7. Lateral borders of frontals: (0) more or less parallel; (1) strongly constricted between orbits. 8. Frontal shelf: (0) lacking broad shelf below nasals; ( I ) broad shelf with frontals often exposed dorsolaterally as wedges or spikes. 9. Descending processes of frontals: (0) weakly developed and prefrontals broadly participate in orbitonasal fenestra; (1) strongly developed and replace prefrontals. 10. Median contact of descending processes of frontals: (0) none; (1) in contact below olfactory tracts. 11. Frontal tabs: (0) no tabs; (1) frontal tabs project posteriorly over parietal. 12. Postfrontal: (0) present, large, separate at some stage; ( I ) postfrontal reduced to a small nubbin; (2) absent (could be lost or fused, but this might be determined by the topology of the resulting tree). 13. Postfrontal forking: (0) subtriangular, not forked medially; (1) semilunate, forked medially . 14. Postorbital contribution to posterior border of orbit: (0) forms about one half of orbital border and is mainly an orbital bone; ( I ) forms less than one half and is mainly a temporal bone; (2) no postorbital. 15. Jugal-squamosal contact: (0) no contact, bones widely separated or ( I ) contact or approach closely. 16. Supratemporal fenestra: (0) open; (1) restricted mainly by postorbital; (2) restricted mainly by postfrontal. 17. Ontogenetic fusion of parietals: (0) paired well into postembryonic ontogeny or (1) fused early. 18. Parietal tabs: (0) absent or (1) present as thin triangular structures extending into fossae below frontal.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 253
19. Parietal downgrowths: (0) absent; (1) pointed ventral downgrowths; (2) deep ventral flanges. 20. Parietal table: (0) extensive posterior table covers most of braincase, postparietal processes short; (1) table short, braincase exposed, postparietal processes long. 2 1. Parietal foramen: (0) within parietal; (1) at frontoparietal suture; (2) in frontal; (3) absent . 22. Posterior extent of maxilla: (0) extends well back under orbits; (1) only just beyond anterior edge of orbit. 23. Lacrimal: (0) separate, large with extensive exposure on cheek; (1) separate small, confined to orbital rim; (2) fused to prefrontal; (3) absent. 24. Lacrimal foramen number: (0) single; (1) double. 25. Anteroventral border of orbit: (0) formed by maxilla with jugal confined to medial surfacc; (1) formed by jugal. 26. Jugal/postorbital bar: (0) jugal large, bar complete; (1) jugd reduced or absent. 27. Squamosal: (0) present; (1) absent. 28. Dorsal process of squamosal: (0) present; (1) absent. 29. Supratemporal: (0) present; (1) absent. 30. Palpebral ossifications: (0) present; (1) absent. 31. Pterygoid lappet of quadrate: (0) present; (1) absent. 32. Vomer fusion: (0) separate well into postemblyonic ontogeny; (I)&sed in embryo or early in postembryonic ontogeny. 33. Vomer size: (0) relatively small, extends posteriorly less than half the length of the maxillary tooth row; (1) elongate posteriorly, extends one half or more the length of the maxillary tooth row and usually restricts internal naris. 34. Median contact of septomaxillae: (0) separated by a gap filled by the cartilaginous internarial septum; (1) meet or nearly meet on midline in a raised crest. 35. Dorsal expansion of septomaxilla: (0) flat or concave, vomeronasal organ small; (1) expanded and convex, vomeronasal organ large. 36. Posterior border of opening for vomeronasal organ: (0) not closed by bone (palaeochoanate); (1) closed by contact or overlap of maxilla and vomer (neochoanate). 37. Medial extensions of palatine: (0) no medial extensions; (1) medial extension from ventrolateral edge of palatine forms air passages for bony secondary palate. 38. Choanal fossa of palatines: (0) small in relation to palatine size; (1) relatively prominent in relation to palatine size. 39. Ectopterygoid contact with palatine: (0) fails to contact palatine anterolaterally, maxilla forms part of the lateral border of the suborbital fenestra; (1) contacts palatine anterolaterally, excluding maxilla from border of suborbital fenestra. 40. Ectopterygoid: (0) relatively slender, fenestra widely open; (1) ectopterygoid enlarged medially, restricting suborbital fenestra. 41. Pyriform recess: (0) narrow throughout; (1) broad. 42. Alar process of prootic: (0) relatively short and primarily directed dorsally; (1) elongated and anterodorsally directed. 43. Supratrigeminal process of prootic: (0) undeveloped or feebly developed; ( I ) Jinger-like projection above trigeminal notch. 44. Enclosure o f lateral head vein in bony canal formed lp crista prootica: (0) crista prootica does not extend forward to enclose the vein; (1) extendsfornard and encloses vein in a bony canal. 45. Posterior opening o f vidian canal: (0) within basisphenoid; ( I ) at bansphenoid-prootic suture. 46. Jaw adductor musculature: (0) extends onto dorsal surface o f parietal; (1) attaches on& onto ventral su face. 47. Meckels- canal: (0) open; (1) clospd with or (2) without Juture. 48 . Intramandibular septum o f Meckel's canal: (0) subdivision o f Meckels- canal occurs well anterior to the posterior end o f the tooth row, with intramandibular septum poor& or moderate4 developed; ( I ) subdivision occurs near postm'or end o f dentaly tooth row with intramandibular septum well developed. 49. Meckelian canal: (0) medial& for whole length; ( I ) ventral anterior to anterior inferior alveolar foramen. 50. Subdental she@ (0) weakly developed; ( I ) large; (2) absent. 51. Dorsal extension o f coronoid process o f dentaly: (0) small or absent; ( I ) large extending onto coronoid. 52. Suraqular in lateral view: (0) tapers anteriorly, pointed d?stal&; ( I ) expanded anterodorsally and near& vertical at anterior margin; (2) as in (1) but anterior end terminates relatively closer to level of coronow' eminence on surangulal: 53. Medial view o f prearticular with splenial and dentary removed: (0) prearticular extends near& to anterior end o f surangular, well anterior to coronoid bone; (1) reduced, not extending well anterior to coronoid bone. 54 . Postemlateral dentary shape: (0) no surangular or comnoid notches; (1) notches pre.ant; (2) notches reduced. 55. Dentary postdentaly: (0) much overlap; ( I ) overlap reduced. 56. Splenial reduction: (0) splenial extends anterior to mid tooth row; (1) shorter; (2) absent.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

251 S. E. EVANS AND L. J. RAKBADILLO
57. Splenial posterior&: (0) posterior to apex $ coronoid process; ( I ) not posterior to coronoid p ~ c ~ s s ; (2) limited overlap posterior&. 58. Splenial-dentav suture: (0) extensive bone to bone contact; (1) zduced. 59. Coronoid with lateral lappet: (0) absent; (1) present; (2) covered b~ posterior process of dentay. 60. Coronoid process anterior&: (0) curves smooth& onto dentay; ( I ) anterior border levelh out bpjore reaching dentag) producing long low extension. 61. Anterior end of coronoid: (0) clasps dentay overlapping medial& and lateral&; ( I ) abuts against den f a y . 62. Angular: (0) present; ( I ) absent. 63. Prearticular crest: (0) absent or (1) present; (2) prominent with embedded angular process. 64. Retroarticular process: (0) dorsal surface with sulcus or pit; (1 j none. 65. Retroarticular process direction: (0) posterior; ( I ) inflected medially. 66. Retroarticular process medial margin: (0) smooth; (1) tubercle or flange. 67. Retroarticular process offset: (0) no ofset; (1) offset with lateral notch. 68. Retroarticular process: (0) tapered or parallel-sided; (1) broader posteriorly . 69. Retroarticular process torsion: (0) no posterior torsion; (1 j posterior torsion. 70. Finger-like angular process: (0) absent; (1) present. 71. Adductor fossa: (0) small or moderate; (1) expanded and inflated (linked to adductor expansion). 72. Palatine teeth: (0) present, multiple rows; (1) present, single middle row; (2) absent. 73. Pterygoid teeth: (0) present, multiple rows; ( I ) present reduced row or patch; (2) absent. 74. Marginal teeth: (0) subpleurodont (lingual and labial walls roughly equal); (1) weakly pleurodont (labial wall twice lingual wall); (2 ) fully pleurodont, labial wall more than twice lingual wall; (3) acrodont. 75. Replacement: (0) iguanid type; ( I ) posterolingual. 76. Plicidentine: (0) absent; (1) present. 77. Step or offset in tooth margin of maxilla: (0) absent; (1) present. 78. Scleral ossicle number: (0) more than 14 ossicles; (1) 14 ossicles or fewer. 79. Second epibranchia 80. Second ceratobranc als: (0) present; (1) absent. 81. Vertcbral condyle: (0) not oblique; ( I ) oblique. 82. Vertebral centrum: (0) amphicoelous, notochordal (1) amphiplatyan (2) procoelous. 83. Centrum constricted before condyles: (0) not; (1) constricted. 84. Zygosphene and zygantrum development: (0) absent (1) weakly developed; (2) strongly developed. 85. Cervical intercentra attachment 1: (0) intervertebral; (1) sutured under anterior part of following centrum; (2) fused under anterior part of following centrum; (3) sutured to posterior part of preceding centrum; (4) fused to posterior part of preceding centrum. 86. Trunk intercentra: (0) present; ( I ) absent. 87. Transverse processes on caudals: (0) single pair of transverse processes; (1) two pairs of converging transverse processes; (2) two pairs of diverging transverse processes; (3) anterior pair of transvcrse processes absent. 88. Autotomy septa in caudal vertebrae: (0) a.s. within a single pair of transverse processes, or between two pairs of transverse processes, or anterior to a single pair of transverse processes; (1) a.s. located posterior to a single pair of transverse processes; (2) as . absent. 89. Number of presacrals: (0) 25; (1) fewer than 25; (2) 26; (3) 27 or more. 90. Number of cervicals: (0) 8 or more; (1) fewer than 8. 91. Number of rib attachments on sternum: (0) 5; (1) 4; (2) 3; (3) 2 or fewer. 92. Postx$histernal inscriptional ribs: (0) none continue midventralb; ( I ) at least some do. 93. Scapular emargination: (0) absent; (1) present. 94. Anterior primary coracoid emarghation: (0) absent; (1) present. 95. Posterior secondary emargination: (0) absent (1) present. 96. Epicoracoid cartilage extent: (0) contacts mesoscapula and usually suprascapular; ( I ) fails to contact either suprascapula or mesoscapula. 97. Clavicle: (0) present, simple; (1) present, expanded and perforated; (2) absent. 98, Clavicle angulation: (0) simple curved rods or (1) angle away from scapulocoracoid. 99. Clavicle meets: (0) scapula or suprascapula (1). 100. Interclavicle; (0) present, robust; (1) present, gracile; (2) absent. 101. Interclavicle lateral process: (0) present, t-shaped; (1) present, cruciform; (2) absent. 102. Sternal fontanelle: (0) absent; (1) present. 103. Ectepicondylar foramen: (0) present only as groove; ( I ) foramen formed in postembryonic ontogeny; (2) formed in embryo; (3) groove and foramen absent. 104. Ventral view of pubis: (0) pubis short or only moderately elongated with symphysial process ventrally directed; ( I ) pubis very long with symphysial process elongate and anteriorly directed.
(0) present; (1) absent.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 255
I05. Postcloacal bones: (0) absent; ( I ) present. 106. Ventral body osteoderms: (0) absent; (1) present. 107. Dorsal body osteoderms: (0) absent; (1) present. 108. Cephalic osteoderms: (0) absent; (1) present. 109. Dermal rugosities: (0) absent; ( I ) present not vermiculate; (2) present, vermiculate. 110. Epiphysis fusion: (0) at same time or after fusion of braincase elements; (1) prior to fusion of braincase elements. 1 1 1. Extension of m.adductor posterior: (0) not extending into Meckel’s canal; (1) extends far into Meckel’s canal. 1 12. Origin of m.pseudotemporalis superficialis: (0) not extended posteriorly along mesial margin of temporal fossa; ( I ) extended posteriorly. 113. Anterior head of m.pseudoprofundus: (0) absent; (1) present. 1 14. M.rectus abdominis lateralis: (0) absent; (1) present. I 15. M.extracolume1laris: (0) absent; ( I ) present. 116. Foretongue retractability: (0) absent; (1) retracts within posterior (hind) tongue at zone of invagination. 1 17. Notching of free part of tongue: (0) no notching present; (1) notched less than 10%; (2) notched 10-20%; (3) notched 20-40%; (4) notched 40-50%; (5) notched more than 50%. 1 18. Anterior tongue cross-section and tongue keratinization: (0) rounded, glandular papillae present throughout tongue; ( I ) much wider than tall, non-glandular, posterior (hind) tongue keratinized; (2) mushroom shaped in cross-section, entire tongue keratinized and non-glandular. 1 19. Tongue plication: (0) entire tongue scaly or papillose; (1) posterior (hind) tongue plicate; (2) entire tongue plicate. 120. Ciliary restraint system for hair cells: (0) tectorial system (restraint imposed by tectorial membrane); ( I ) combined tectorial and sallet system; (2) sallet system with restraint imposed by inertial bodies (sallet or culmen). 12 I . Internal (quadrate) process of stapes: (0) present; ( I ) absent. 122. Ulnar nerve position in forelimb: (0) superlicial (‘lacertid’) condition; ( I ) deep (‘varanoid’) condition. 123. Innervation of dorsal muscles of lower leg: (0) peroneal nerve; (1) interosseus . nerve. 124. Fcmoral or pre-anal pores: (0) absent; (1) present. 125. Course of stapedial artery: (0) passes anterior to stapes; (1) perforate stapes; (2) passes posterior to stapes. 126. Modged middorsal scale mw: (0) present; (1) absent. 127. Cephalic scales: (0) relatively small; (1) enlarged. 128. Cycloid scales: (0) absent; (1) present. 129. Pubis tubercle position: (0) tubercle posterodorsally placed; (1) tubercle anteroventrally placed. 130. Nasal width: (0) nasals broad, equal or greater in width than width of conjoined nares; (1) nasals reduced, width less than that of conjoined nares. 131. Fronto-parietal suture: (0) u or w shaped in dorsal view; (1) more or less transverse. 132. Frontoparietal suture: (0) equal or narrower in width than naso-frontal suture; (1) greater in width than naso-frontal suture. 133. Supratemporal, where present: (0) superlicial in position on squamosal/parietal junction; (l), deeply placed, between parietal and squamosal. 134. Squamosal ventral ramus: (0) squamosal bears ventral ramus extending down anterior margin of quadrate; (1) ventral ramus lost. 135. Qnadrate dorsal head: (0) solid; ( I ) notched or perforated for ventral peg o f squamosal (or f o r chorda gmpani nerve). 136. Kmerine teeth: (0) present, numerous; ( I ) absent (or with at most one or two remnant teeth). 137. Pterygoid, anterior process: (0) long anterior process of pterygoid reaches vomer, and usually opposite pterygoid; ( I ) pterygoid process reduced, pterygoids separated from one another and from vomers by palatines. 138. Palatine/pterygoid contact: (0) palatine posteromedially extended, restricts entry or pterygoid into margin of suborbital fenestra; (1) palatine reduced posteromedially, pterygoid with broad entry into suborbital fenestra. 139. Septomaxilla: (0) simple outline, no posteroventral process; ( I ) posteroventral projection extending towards dorsal surface of uomer to form posterior margin o f duct ofJacobson’s organ. 140. S@tomaxilla: (0) simple, limited to posteroventral edge o f external nares to form side and part o f j o o r o f nasal capsule; ( I ) expanded bone covering enlarged uomeronasal organ dorsal& andjooring nasal passage ventral&. 141. Paroccipital process: (0) short, narrow, does not contact quadrate; ( I ) narrow, contacts quadrate; (2) meets quadrate and is expanded distally to support quadrate dorsally.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

256 S. E. EVANS i\ND L . J . BARBADILL0
142. Stapes: (0) thick, perforated; ( I ) relatively thinner, imperforate though footplate significantly largcr than shaft; (2) very slender, columelliform, small footplate. 143. Epipterygoid: (0) with broad base, facetted to pterygoid and contacting quadrate; ( I ) with columella base, in fossa columellae, and not contacting quadrate; (2) epipterygoid absent. 144. Metotic fissure: (0) no anteroventral extension, exoccipital meets opisthotic both above and belo\\ small metotic fissure; (1) exoccipital does not meet opisthotic ventrally, metotic fissure extends anteroventrally to reach basal plate; (2) metotic fissure divided into separate openings for the vagus nerve dorsally, and the lateral opening of the recessus scala tympani ventrally; usually with expansion of crista tuberalis to form enlarged frame for fenestra rotunda.
This extends an EstesIGauthier character. In primitive lepidosauromorphs (Mamoretta, kuehn~osanrus) as well as Eungina, the exoccipital meets the braincase both above and below a small undivided mctotic fissure which can have had little or no function as a compensatory opening. In rhynchocephalians (including the basal Gep/y~xaurzls), the metotic fissure is extended anteroventrally and includes the basal plate in its boundaries. The anteroventral extension may have accommodated a small perilymphatic duct. Further expansion and subdivision of the metotic fissure occurs in squamates. 145. Vidian canal: (0) open, runs in groove between lateral edge of parasphenoid and basisphenoid; ( I ) closed by lateral expansion of the parasphenoid. 146. Angular: (0) long, reaches mandibular condyle; (1) shortened, fails to reach condyle. 147. Coronoid eminence: (0) little or no eminence; ( I ) eminence present but mostly coronoid process of dentary and coronoid; (2) eminence formed by coronoid alone. 148. Anterior cervical ribs: (0) double headed; (1) single headed. 149. Cervical intercentra: (0) simple bars; (I) prominent hypapophyses. 150. Sacral and caudal ribs: (0) fuse to respective ribs posthatching; (1) j u s e during a b y o n i c development. 151. Neural arches: (0) j k s e to their respective vertebra posthatching; (1) j k s e during emblyonic development. 152. Entepicondylar foramen of humerus: (0) present; (1) lost. 153. Ulna distal epiphysis: (0) weakly developed; (1) enlarged distal epiphysis of ulna that is nrarly hemispherical in profile and fits into a corresponding depression in the ulnare. 154. Radial distal epiphysis: (0) weakly developed, simple; ( I ) develops ‘styloid’ process which fits into a concave depression on the proximal end of the radiale. 155. Carpal intermedium: (0) lage, present; ( I ) small or absent. 156. Centrale o f hand: (0) medial centrale contacts third distal caqal; (1) lateral centrale c0ntact.r Jecond distal carpal to .separate medial centra1e)om dc3. 157. First metatarsal: (0) contacts only dcl; (1) is expanded to contact medial centrale and dc2. 158. Pubic symphysis: (0) strong contact in front of relatively small thyroid fenestra; ( I ) reduced contact in front of wide open thyroid fenestra. 159. Tibio-astragalar joint: (0) fixed, with tibial ridge fitting into groove on astragalus; ( I ) loss of ridge and groove structure, more or less plane surface; (2) notched tibial epiphysis fits over single ridgr on astragalus. 160. Fibuloastragalar joint: (0) involves only a small part of distal fibular head; ( I ) involves most of distal head. 16 1. Fourth distal tarsal: (0) simple proximal surface; ( I ) complex tongue in goove ,joint between fourth distal tarsal and distal surface of astragalocalcaneum. 162. Fifth metatarsal: (0) simple, straight, with some expansion of distal head; (1) hooked, but lacking prominent medial and lateral plantar tubercles; (2) hooked with prominent medial plantar tubercle and laterally displaced lateral plantar tubercle. 163. Distal tarsal 2: (0) present; ( I ) absent. 164. Gastralia: (0) present; ( I ) absent. 165. hf. bursalis: (0) no discrete m.bursalis; (1) discrete m.bursalis develops f i m part o f m.retractor bulbi. 166. Anterior braincase and interorbital septum: (0) laze4 cartilaginous, small amounts o f membrane and f imtrae; (1) reduced cartilaginous component, large fenestrae and s&$icant amounts of membrane. 167. Toque pallets: (0) smooth surface; (1) pallets on ventral surface o f tongue. 168. Penis: (0) absent; (1) hemipenes developed. 169. Lacrimal duct: (0) associated with posterior end of choanal ,pmoue; ( I ) extends antarorb to become assoc.iatpd with vomwnasal duct. 170. Vomeronasal organ: (0) diverticulum of nasal capsule; (1) separate from nasal capsule and with fungiform body. 171. Sensov epithelium o j vomeronasal ogan: (0) weak& developed; (1) extensive& developed. 1 72. l6mmnasal ogan and paraseptal cartilage: (0) paraseptal cartilage supports j o o r of vomeronacal organ, lateral4 placed duct opens into medial wall ofchoana; ( I ) vomeronasal ogan rotated nine0 degees about its long ax&, paraseptal cartilage lies media& and vomeronasal duct opens ventral& into oral c a v q
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 25 7
173. Lateral nasal gland: (0) small, outside cavum conchale; (1) enlaTed, within cauum conchale. 174. Caruncle: (0) caruncle and egg tooth present at hatchling; ( I ) on& egg tooth, caruncle lost. 175. M m intermandibularis and mandibulohyoideus: (0) non-interdgtating; (1) multiple interdigitation. 176. M.depressor mandibulae: (0) partb coguent with m.episternocleidomastoideus; (1) completeb separate. 177. Innervation o f m.intermandibula&: (0) involves facial nerve; ( I ) no involvement o f facial nerve. 178. M.clavodeltoideus: (0) extends on5 to dorsal surface o f clavicle; ( I ) extends to ventral suface o f clavicles. 179. Meniscus o f knee: (0) subdivided into medial and lateral parts b~ cruciate ligament; (1) single plate pierced by cruciate ligament. 180. Pm'bmphatic sac: (0) small; (1) prominent filling recessus scala gmpani (this is linked to form o f metoticjssure). 181. Cochlear duct: (0) faces ventralb; (1) faces lateralb(varies bw embyos in squamates [O] and adults [ l ] , except [O] in some snake.r as welq. 182. Pars tuberalis o f adenohypophysis: (0) present; (1) absent. 183. Interhyal: (0) cartilaginous; (1) membranous or absent. 184. Ovaries: (0) solid; (1) saccular. 185. Chemoreception: (0) co$ned to oral cavig; (1) tongue mediated extraoral chemoreception (i.e. tongue protrusion in licking and tasting unique to squamates. 186. Sublingual glands: (0) simple epithelial invaginations; (1) compound. 187. M.genioglossus: (0) does not insert into buccaljloor; (1) inserts into buccaljoor (Schwenk, 1988). 188. ALgenioglossus: (0) undivided; (1) divided into medial and lateral parts. 189. Mtransversalis fibres: (0) curl ventralb around hyoglossus and insert on ventral transverse septum; (1) forms uninlerrufited loop. 190. M.verticalis m. of tongue: (0) has ventral transverse sqtum; (1) lacks median septum and showsfibre crossova 191. Parasphenoid and basisphenoid: (0) separate in hatchling; (1)J;sed in embTo. 192. Pmatlas: (0) present; (1) absent. 193. Atlantal neural arches: (0) separate; (1) broadly in contact to cover neural canal. 194. Atlantal neural arches: (0) remain free; (1) fuse to intercentrum at cessation of growth. 195. Postparietals: (0) present; (1) absent. 196. libulars: (0) present; (1) absent. 197. (0) Postorbital overlain by pos@ntal; (1) pos$?ontal overlain by postorbital. 198. Quadratojugal: (0) present, anterior process; (1) present, reduced, no anterior process; (2) absent. 199. Posterior process ofjugal: (0) extends about halfway back (or less) below lower temporal fenestra; (1) more than half way; (2) no posterior process, rounded or angled margin. 200. Adductor chamber: (0) small, quadrate does not extend far below level of occipital condyle; (1) enlarged, quadrate extends well below occipital condyle. 201. Nares: (0) paired; ( I ) coy¶uent. 202. Quadrate width: (0) narrow in posterior view; (1) wide in posterior view with development of a conch. 203. Quadrate/pterygoid relationship: (0) quadrate medial wing overlaps pterygoid flange to form a firm contact; (1) quadrate and pterygoid wings reduced, no strong overlap. 204. Quadrate: (0) with quadratojugal foramen; (1) without quadratojugal foramen. 205. Palatine teeth: (0) no enlarged lateral tooth row; (1) lateral tooth row converging laterad, as one of several rows; (2) anterior four or five teeth enlarged in relation to others; (3) lateral palatine row parallels maxillary row. 206. Pterygoid flange: (0) teeth present; (1) teeth absent. 207. Abducens canal: (0) bony canal incomplete or absent; (1) present and complete. 208. Quadrate shape in lateral view: (0) straight; (1) bowed. 209. Postorbital/parietal: (0) postorbital contacts parietal; (1) does not contact parietal. 210. Relation of palatine to suborbital fenestra: (0) suborbital fenestra large and suboval; (1) palatine broadened laterally, constricting suborbital fenestra anteriorly; (2) palatine further broadened lateral, constricts all suborbital fenestra; (3) lateral margin of palatine parallel to maxilla, suborbital fenestra almost closed. 211. Nasal process o f premaxilla: (0) narrow; (1) broad. 2 12. Exoccipitals: (0) separate; (1) fused to basioccipital; (2) fused to opisthotic in mature animals; (3) fused to opisthotic prior to hatching. 213. Posterior extension of dentary: (0) extends posteriorly to level of coronoid eminence or slightly beyond; (1) extends posteriorly more than halfway between coronoid eminence and articular condyle. 214. Posterior extension of angular: (0) extends posteriorly to articular condyle; (1) does not extend to articular condyle. 215. Lateral extent ofangular: (0) angular extends more than one third up lateral surface ofmandible; (1) extends less than one third.
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

258
2 16. Retroarticular process size: (0) relatively small; (1) large; (2) absent. 2 17. Surangular participation in articular condyle: (0) outer third to one half of condyle formed by surangular; (1) condyle formed entirely by articular, surangular forms only outer rim. 21 8. Condylar surface: (0) articular condyle wider than long, anteroposterior ridge small or absent; ( I ) condyle as long as wide, strong antemposterior articular ridge present; (2) condyle longer than wide, antemposteror ridge present. 21 9. Transverse processes in trunk: (0) short or absent; ( I ) elongated. 220. Posterior process on second sacral rib: (0) absent; (1) present. 221. A l f o m outgrowths on ribs: (0) absent; ( I ) present. 222. Xz$histernum: (0) relativeb hge; (1) small; (2) absent. 223. Rib bearing portion of sternum: (0) simple rod; (1) enlarged and formed of two plates remaining separate until nearb maximum size; (2) paired sternal platespse in emblyo. 224. Cleithrum: (0) present; ( I ) absent. 225. First distal carpal: (0) separate bony element; (1) fused to mcl or absent. 226. Mc thickness: (0) first and fifth metacarpals relatively thin compared to 2 4 ; (1) 1 & 5 relatively thick, 2-4 gracile. 227. Mc4: (0) longer than 3; (1) 3 & 4 subequal in length; (2) mc4 shorter than 3. 228. Medial and lateral centralia: (0) mc and lc subequal in size; (1) mc as much as twice size of lc. 229. Olecranon: (0) well developed in all except juveniles; (1) poorly developed in full grown animals (polarity uncertain). 230. Shajh ofradius and ulna: (0) subequal in length; ( I ) radius longer than ulna. 231. Humeral entepicondyle: (0) not strongb developed in adult; (1) strongb developed. 232. Humerusfemur ratio: (0) in adult, humerus shorter than f m u r ; ( I ) as long or longer than femur. 233. Acetabulum: (0) ilium includes 80-85% acetabulum; (1) 60-65% acetabulum. 234. Pubic jlange of ilium: (0) absent; ( I ) present. 235. Dorsal edge of ilium: (0) horizontal; (1) steeply inclined. 236. Thyroid fenestra: (0) absent, pelvis a solid plate; (1) present, small; (2) present, large, pubic symphysis narrow. 237. Fusion of pelvic elements: (0) separate in full grown adult; (1) fused. 238. Femur/fibula: (0) fibula and femur articulate end to end, distal femoral condyles symmetrical; (1) fibula sits in recess on lateral margin of distal end of femur, femoral condyles markedly assymetrical. 239. Astragalus/calcaneum: (0) separate throughout ontogeny; (1) fused prior to fusion of sca- pulocoracoid. 240. Lateral pes centrale: (0) present as separate element throughout ontogeny; ( l ) f ; . d to astragalus in emblyo. 241. First distal tarsal: (0) present; (1) absent. 242. Dt5: (0) present as discrete element at some point in ontogerp; ( I ) absent or f u e d to d t 4 ~ r AC during ontogerp. 243. 3rd branchial arch: (0) complete; (1) interrupted. 244. Bony epiphyes: (0) absent; ( I ) present. 245. Extra ossifications: (0) few connective tissue ossifications; (1) calcification of many tendons, sheets of connective tissue, cartilage etc occurs prior to full maturity. 246. Cloacal slit orientation: (0) antempastenor; ( I ) transverse. 247. Kidney: (0) without sexual segment; ( I ) sexual segment present. 248. %ngue use: (0) not used to secure small pry; ( I ) used. 249. Cilialy process: (0) present; ( I ) absent. 250. Mict i tans: (0) tendon gm.nictitans attached to m$yrimidalir, the latter taking origin on surface ofgeball; (1) tendon attaches to n.retractor bulbi and interorbital septum. 251. Skin shedding: (0) skin not noma& shed in its entirep; ( I ) repularh shed in its entirep. 252. Cartilaginous disc in eyeball: (0) absent; (1) present. 253. Symphysis: (0) large plane terminal symphysial surface, round or ovoid, surrounding tip of Meckelian canal; (1) reduced symphysial surface limited to dorsal edge of Meckelian canal; (2) little or no symphysial surface-small rugosity or raised edge on dorsal surface of Meckelian canal. (Primitive lepidosauromorphs have a strong ovoid symphysial surface which is substantially reduced in squamates.) 254. Perforating foramen in ankle: (0) present; (1) absent.
S. E. EVANS AND L. J. BARBADIILO
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 239
- 4 5 3 c c 5 c - c 5 3 3 3 n. c c 3 c s 5 5 c 5 -
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

S. E. EVANS AND Id. J. BARBADILLO
- - 3 go 0 c 0 - a 3 0 0 c 0 C . 0 3 c 0 . A . 0. n . n . 0 - 3
- - - 8 - - g 3 - - g - - - - - n. - - - C . - ”. 0 c 0 3
- c 3 c - - 0 c - 0 3 c c 0 0. c 0 c 5 5 = c -. =
- c 0 0 - - pT: 3 - c 5 c c c= c c 0 3 0. * . I _ 0 c c -
3 0 - c 5 - 3 3 0 5 c c c c c 5 c n . ~ = c = =
3 0 c - Nc - N O 5 5 0 0 0 . c 0 c c. n. *. c= - e - - - - - - id a c c 0 a a c c c 0 0 3 c 3 a- c 0 n. = a = 3 0 5 0 - 5
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 26 1
- - = c e 3 3 d 0 3 g o 0 3 c 0 - 0 - 0 s I. n . 0 c
g 2 - - - - - - - - - - - n. n. - n. n. ... -
- - n. n. a
- - - j 3 &5 c c ;a s c 6 3 3 0 c 5 n. - - - - 0 7.0 c
3 3 c
3 1 - - - 0
N
3 ." NN- c 3 0 0 3 0 0 n N 0 0 C. 0 *.;a me d
- r . ~ o - 3 5 0 0 0 3 - - . . ; a ~ c n , n . . r . 0 3 "
0
N
d - - &5 N N N N N N N N N N N 0 N N N 5 n. N N C1.a
c 3 c c = 3 c c c - - - n. c 3 3 n. n. n. 0 0 n.
- - : $ - 3 3 - 0 3 - - - g- 3 3 0 3 3 3 0 *. 3 3 -
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

S. E. EVA1L’S AND L. J. RARBADILLO
- N d N N N N N N N N N N N “. 0 N C 0 . C . 0. n . 0. c
0
- N 3 .N N N c.4 N ?I N c4 N N N N N N - N 0 . I
N N g -
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022
EARLY CRETACEOUS LIZARD FROM SPAIN 263
n. _L. n.
n. n. 0.
n. n. n.
0. n . n .
n. -. _i.
-. *. -.
0. ̂ . n.
n. 0. 0.
C. 0. 0.
-. c
*. a
n. 5
n . 5
n. c
0. c
P . c
r.. c
C . c
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

0 N N
2 N
m N
h
N -
s Y - - N
1:
n 3 - N
N -
- - Y
- .. - N
m w
m N
r. w
‘a w
10 w
* 3 N
m N
Y
N
- 0 N
S. E. EVANS AND L. J. BARBADILLO
Tc;
3 0 c c 0 3 3 c 0 0 3 3 0 n.O m N c 0 C . 3 3 .a 0
3 a x
.? u
n
4
m
* N
m m N
N cc N
- * Y
3
Y
3 ’I
0
’I
r. N
a N N
Lo
N
d N N
m N N
N N
- N N
3 3 0 0 0 c 3 5 3 3 c c 0 c 0 0 .. 0 5 3 n. c 0
= 0 0 0 3 3 0 0 5 0 3 = 0 0 n . o r . 0 c 3 -. 0 c
- d ~ . ” . n . * . n . r . r . r . r . r . r . - . r . n . r . O r . r . - r , r . =
0 0 c 0 5 3 3 5 0 3 0 0 0 0 3 = 0 0 - 0 1.5 .t“ -
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022

EARLY CRETACEOUS LIZARD FROM SPAIN 265
2
s N
N 0 N
- N
0 I.-, N
fl d
CD
N
* N
c %
+
* u
I *
u ?d
+
Dow
nloaded from https://academ
ic.oup.com/zoolinnean/article/124/3/235/2684343 by guest on 25 M
arch 2022




















