Sprouting and abnormal contacts of nonmedullated axons, and deposition of extracellular material...
12
BRAIN RESEARCH ELSEVIER Brain Research 718 (1996) 13-24 Research report Sprouting and abnormal contacts of nonmedullated axons, and deposition of extracellular material induced by the amyloid precursor protein (APP) and other protease inhibitors Ricardo D. Moreno a Nibaldo C. Inestrosa a Alan R. Culwell b Jaime Alvarez a, * a Departamento de Biologla Celular y Molecular, Facultad de Ciencias Biolrgicas, Pontificia Universidad Catdlica, Casilla 114-D Santiago, Chile b Athena Neurosczence, South San Francisco, CA, USA Accepted 5 December 1995 Abstract We have reported that the local administration of serine protease inhibitors (amyloid precursor protein with the Kunitz insert (APP K + ), aprotinin, and leupeptin) to the rat sciatic nerve determines a sprouting response of myelinated axons, proliferation of Schwann cells, and demyelination, 5 to 7 days later. Further study of these nerves with the electron microscope revealed (i) a sprouting response of nonmedullated axons, (ii) the appearance of fine axons with a few turns of compact myelin, (iii) abnormal contacts of axons with basal laminae, with fibroblast-like cells, and between them, (iv) the occurrence of hemidesmosome- and desmosome-like junctions between Schwann cell processes, and between Schwann cells and axons, and (v) the appearance of amorphous and fibrillary extracellular deposits alongside the axolemma. The adjacent proximal and distal segments were normal, i.e., axons remained continuous, and the alterations were confined to the segment exposed to the protease inhibitors. Heated APP K +, APP without the Kunitz insert (APP K - ), bovine serum albumin, and saline, did not elicit cytological alterations. Our results suggest that these inhibitors of serine proteases (i) set free a sprouting drive of axons by disrupting an ongoing repressive mechanism; (ii) modify the adhesive properties of axons and Schwann cells, and (iii) alter the natural history of an extracellular material. The imbalance of an extracellular protease system may participate in the pathogenesis of Alzheimer's disease. Keywords: Peripheral nerve; Leupeptin; Aprotinin; APP; Serpin; Deposit; Schwann; Anchoring junction; Desmosome; Alzheimer 1. Introduction The extracellular matrix proteins, proteases and their inhibitors play an important role in the control of the phenotype of a number of cells. Proteases and antipro- teases have been implicated, in the growth of neurites, e.g., collagenase-like proteases, urokinase, tissue plasminogen activator [33], and nexin-I [30], to mention a few. We have shown that local destruction of Schwann cells with actino- mycin-D induces intact axons to sprout [13] which sug- gests that axons have a sprouting programme repressed by Schwann cells. Inhibitors of serine proteases administered to a nerve trunk also induce intact myelinated axons to sprout [2] hence a physiological protease is likely to participate in the repressiwe mechanism. In addition, pro- tease inhibitors induce Schwann cells to proliferate and * Corresponding author. Fax: (56) (2) 222 55 15. Email: j alvarez @genes.bio.puc.cl 0006-8993/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 0006-8993(95)01555-8 demyelinate [3]. Therefore, the salient events that follow a nerve section, namely, axonal sprouting, proliferation of Schwann cells, and removal of myelin, are also triggered locally by protease inhibitors despite the intact condition of axons. Taken together this evidence suggests that a trophic mechanism is specifying the mature condition of the peripheral nerve fibre, and that the breakdown of this mechanism by interruption of axons or by inhibition of proteases releases the processes leading to the early event of nerve degeneration-regeneration. APP has several isoforms, some of which embody an antiprotease domain of the Kunitz type [21]. The natural history of APP is altered in Alzheimer's disease resulting in the deposition of a proteolytic fragment, the /3-amyloid, which forms the core of the senile plaques. In the crown of these plaques, distrophic neurites are a common occur- rence [17,25,26]. Thus the study of the regulatory proper- ties of proteases and their inhibitors, APP in particular, may shed light in the pathogenesis of the disease.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Sprouting and abnormal contacts of nonmedullated axons, and deposition of extracellular material...

BRAIN RESEARCH
E L S E V I E R Brain Research 718 (1996) 13-24
Research report
Sprouting and abnormal contacts of nonmedullated axons, and deposition of extracellular material induced by the amyloid precursor protein (APP) and
other protease inhibitors
Ricardo D. Moreno a Nibaldo C. Inestrosa a Alan R. Culwell b Jaime Alvarez a, *
a Departamento de Biologla Celular y Molecular, Facultad de Ciencias Biolrgicas, Pontificia Universidad Catdlica, Casilla 114-D Santiago, Chile b Athena Neurosczence, South San Francisco, CA, USA
Accepted 5 December 1995
Abstract
We have reported that the local administration of serine protease inhibitors (amyloid precursor protein with the Kunitz insert (APP K + ), aprotinin, and leupeptin) to the rat sciatic nerve determines a sprouting response of myelinated axons, proliferation of Schwann cells, and demyelination, 5 to 7 days later. Further study of these nerves with the electron microscope revealed (i) a sprouting response of nonmedullated axons, (ii) the appearance of fine axons with a few turns of compact myelin, (iii) abnormal contacts of axons with basal laminae, with fibroblast-like cells, and between them, (iv) the occurrence of hemidesmosome- and desmosome-like junctions between Schwann cell processes, and between Schwann cells and axons, and (v) the appearance of amorphous and fibrillary extracellular deposits alongside the axolemma. The adjacent proximal and distal segments were normal, i.e., axons remained continuous, and the alterations were confined to the segment exposed to the protease inhibitors. Heated APP K + , APP without the Kunitz insert (APP K - ), bovine serum albumin, and saline, did not elicit cytological alterations. Our results suggest that these inhibitors of serine proteases (i) set free a sprouting drive of axons by disrupting an ongoing repressive mechanism; (ii) modify the adhesive properties of axons and Schwann cells, and (iii) alter the natural history of an extracellular material. The imbalance of an extracellular protease system may participate in the pathogenesis of Alzheimer's disease.
Keywords: Peripheral nerve; Leupeptin; Aprotinin; APP; Serpin; Deposit; Schwann; Anchoring junction; Desmosome; Alzheimer
1. Introduction
The extracellular matrix proteins, proteases and their inhibitors play an important role in the control of the phenotype of a number of cells. Proteases and antipro- teases have been implicated, in the growth of neurites, e.g., collagenase-like proteases, urokinase, tissue plasminogen activator [33], and nexin-I [30], to mention a few. We have shown that local destruction of Schwann cells with actino- mycin-D induces intact axons to sprout [13] which sug- gests that axons have a sprouting programme repressed by Schwann cells. Inhibitors of serine proteases administered to a nerve trunk also induce intact myelinated axons to sprout [2] hence a physiological protease is likely to participate in the repressiwe mechanism. In addition, pro- tease inhibitors induce Schwann cells to proliferate and
* Corresponding author. Fax: (56) (2) 222 55 15. Email: j alvarez @ genes.bio.puc.cl
0006-8993/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 0 0 0 6 - 8 9 9 3 ( 9 5 ) 0 1 5 5 5 - 8
demyelinate [3]. Therefore, the salient events that follow a nerve section, namely, axonal sprouting, proliferation of Schwann cells, and removal of myelin, are also triggered locally by protease inhibitors despite the intact condition of axons. Taken together this evidence suggests that a trophic mechanism is specifying the mature condition of the peripheral nerve fibre, and that the breakdown of this mechanism by interruption of axons or by inhibition of proteases releases the processes leading to the early event of nerve degeneration-regeneration.
APP has several isoforms, some of which embody an antiprotease domain of the Kunitz type [21]. The natural history of APP is altered in Alzheimer's disease resulting in the deposition of a proteolytic fragment, the /3-amyloid, which forms the core of the senile plaques. In the crown of these plaques, distrophic neurites are a common occur- rence [17,25,26]. Thus the study of the regulatory proper- ties of proteases and their inhibitors, APP in particular, may shed light in the pathogenesis of the disease.

14 R.D. Moreno et al./ Brain Research 718 (1996) 13-24
We report here that, under the influence of protease inhibitors, nonmedullated axons sprout, the bundles ac- quire an organization that reminds that of immature nerves, anchoring junctions develop, extracellular deposits appear alongside the axolemma, and we further characterize the sprouting of myelinated axons.
2. Materials and methods
2.1. Compounds
In this research we used the soluble isoforms of APP75 l and APP695. In Fig. 1, the various domains of these APPs, namely fl-amyloid, transmembrane domain, Kunitz insert, metalloproteinase inhibitor sequence, fibroblast growth stimulating sequence, and the cleavage site within the f l-amyloid sequence are indicated. The Kunitz insert makes this isoform an inhibitor of serine proteases. Two trans- fected human cell lines overexpress the APP751 and APP695 isoforms, respectively [14]. The extracellular domain of this membrane protein is cleaved through the amyloid sequence and released onto the culture medium. The solu- ble isoforms share, except for the Kunitz insert, all the amino acid sequence, including amino acids 1-15 of the fl-amyloid. We denote these soluble isoforms as sAPP K + and sAPP K - , respectively. The purification of these sAPPs, carried out at Athena Neuroscience, South San Francisco, CA, has been reported elsewhere [14]. sAPPs were stored at - 8 0 ° until use. Leupeptin, aprotinin and serum albumin were obtained from Sigma (St. Louis, NO) .
2.2. Administration of compounds
Aseptic solutions were prepared with isotonic sodium chloride (Table 1). sAPP K + was used native or heated
Ext c,.0 c~0 v
/1 . t . : -~
N/.'"[ T cp0 I ( ; ,c.o
M Int
APP695
C APP751
Fig. 1. Schema of APP695 and APP75 r Figures indicate the number of amino acids of the protein. APPs are membrane proteins with a single transmembrane domain. M, cell membrane; Ext, and Int, extracellular and intracellular spaces, respectively; N, and C, amino and carboxi termini of the protein, respectively; CHO, sites of glycosylation; /3 (solid box), /3-amyloid sequence; PP, pentapeptide with fibroblast growth stimulating activity; K (empty box), Kunitz antiprotease sequence, which is the only difference between APP75 ~ and APP695. Dotted lines indicate the site where K is inserted. Horizontal arrow, metalloproteinase inhibitor se- quence. Arrowhead, a protease cleaves APP through the fl-amyloid sequence solubilizing the extracellular domain. The truncated fl-amyloid sequence becomes the carboxi terminal domain of soluble APPs. For sources, see Inestrosa and Soto [21], Miyasaki et al. [29], and Ninomiya et al. [31].
Table 1 Administration of APPs and other compounds
Compound Dev. [ ] Days n
Native sAPP K + inj 520 2 2 inj 520 5 4 pump 52 7 1
Heated sAPP + inj 520 5 3 sAPP K - inj 790 2 2
inj 790 5 4 pump 79 7 1
Leupeptin inj 5 5 2 pump 0.5 7 1 pump 1.0 7 3 inj 1.4 10 1
Aprotinin inj 0.5 5 2 pump 0.4 7 3
Serum albumin inj 0.5 5 4 Saline inj - 5 3
pump - 7 1
Dev, device, injection or osmotic pump. [ ], concentration in nM for APPs, and mg/ml for others. Days, of survival, n, size of group.
for 10 min in a boiling water bath. Rats (Sprague-Dawley , 200 g) were used; surgery was carried out under pentobar- bitone anesthesia (30 m g / k g , i.p.). In most rats, 4 - 5 mm of the peroneal nerve were infiltrated in the distal third of the thigh with 2 /xl of the solution; 2 days later, a second injection was made. In a few rats, an Alzet osmotic pump (rate: 1 / x l / h for 7 days) was implanted; one end of a silicone tubing was connected to the pump and the other end, fitted with a 4-mm-long cuff, was secured to the sciatic nerve with hair stitches. Survival times are indi- cated in Table 1.
2.3. Histological processing
The nerves were fixed by vascular perfusion or infiltra- tion (4% paraformaldehyde, 0.3% glutaralhehyde, 0.01% picric acid, buffer phosphate 0.1 M, pH 7.4). The nerves were removed and left overnight in the same fixative. After a short rinse, the tissue was immersed in buffered 1% osmium tetroxide for 2 h followed by 2% uranyl acetate for 2 h; finally, the tissue was dehydrated with graded acetones and infiltrated with Epon (for details, see [13]). For light microscopy, sections were stained with Toluidine blue. For electron microscopy, thin sections were double stained with uranyl acetate and lead citrate.
2.4. Morphometry of nonmedullated fibres
To estimate the relative the population of nonmedul- lated fibres, we used a modification of the method of Jenq and Coggeshall [22]. The estimate of the nonmedullated population is given by the formula
Nf = M f × ( N / B ) × ( B / M )
where Nf stands for total nonmedullated fibres; Mf, for total myelinated fibres; ( N / B ) , for nonmedullated axons
R.D. Moreno et al. / Brain Research 718 (1996) 13-24 15
per bundle; and ( B / M ) , for bundles per myelinated fibre. Mf and ratios should be determined experimentally. In our experiments we could not count Mf. Instead, two reason- able assumptions were made, namely, that Mf is constant for the peroneal nerve (coefficient of variation is 5% in the tributaries of the sciatic nerve [22]), and that the treatment did not change measurably the original population of Mf. This method permit comparison between the relative sizes of two nonmedullated populations. In practice, in each nerve surveyed, we counted more than 100 myelinated and demyelinated fibres in consecutive squares of the grid, the corresponding nonmedullated bundles, and the axonal pro- files within each bundle.
3. Results
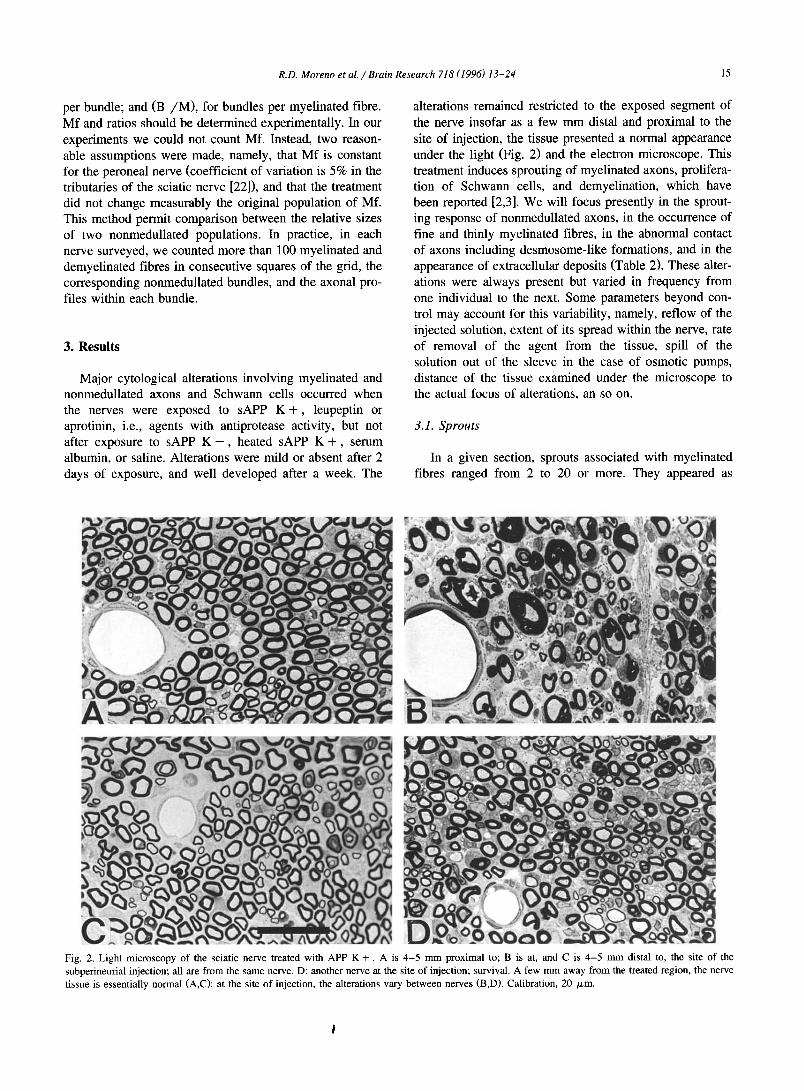
Major cytological alterations involving myelinated and nonmedullated axons and Schwann cells occurred when the nerves were exposed to sAPP K + , leupeptin or aprotinin, i.e., agents with antiprotease activity, but not after exposure to sAPP K - , heated sAPP K + , serum albumin, or saline. Alterations were mild or absent after 2 days of exposure, and well developed after a week. The
alterations remained restricted to the exposed segment of the nerve insofar as a few mm distal and proximal to the site of injection, the tissue presented a normal appearance under the light (Fig. 2) and the electron microscope. This treatment induces sprouting of myelinated axons, prolifera- tion of Schwann cells, and demyelination, which have been reported [2,3]. We will focus presently in the sprout- ing response of nonmedullated axons, in the occurrence of fine and thinly myelinated fibres, in the abnormal contact of axons including desmosome-like formations, and in the appearance of extracellular deposits (Table 2). These alter- ations were always present but varied in frequency from one individual to the next. Some parameters beyond con- trol may account for this variability, namely, reflow of the injected solution, extent of its spread within the nerve, rate of removal of the agent from the tissue, spill of the solution out of the sleeve in the case of osmotic pumps, distance of the tissue examined under the microscope to the actual focus of alterations, an so on.
3.1. Sprou t s
In a given section, sprouts associated with myelinated fibres ranged from 2 to 20 or more. They appeared as
Fig. 2. Light microscopy of the sciatic nerve treated with APP K +. A is 4-5 mm proximal to; B is at, and C is 4-5 mm distal to, the site of the subperineurial injection; all are from the same nerve. D: another nerve at the site of injection; survival. A few mm away from the treated region, the nerve tissue is essentially normal (A,C)I at the site of injection, the alterations vary between nerves (B,D). Calibration, 20 /xm.
16 R.D. Moreno et a l . / Brain Research 718 (1996) 13-24
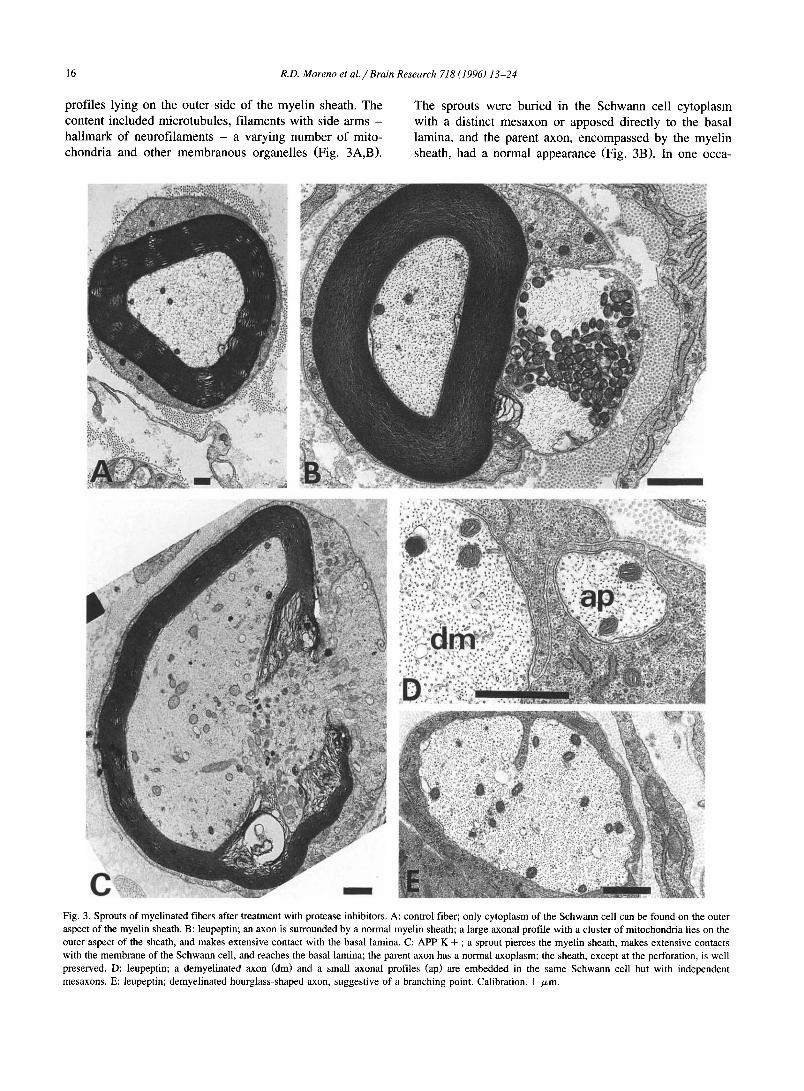
prof i les lying on the outer s ide o f the mye l in sheath. The
content inc luded micro tubules , f i laments wi th side arms -
ha l lmark o f neurof i l aments - a varying n u m b e r o f mito-
chondr ia and other m e m b r a n o u s organel les (Fig. 3A,B).
The sprouts were bur ied in the S c h w a n n cell cy top la sm
with a dis t inct m e s a x o n or apposed direct ly to the basal
lamina, and the parent axon, e n c o m p a s s e d by the mye l in
sheath, had a normal appearance (Fig. 3B). In one occa-
Fig. 3. Sprouts of myelinated fibers after treatment with protease inhibitors. A: control fiber; only cytoplasm of the Schwann cell can be found on the outer aspect of the myelin sheath. B: leupeptin; an axon is surrounded by a normal myelin sheath; a large axonal profile with a cluster of mitochondria lies on the outer aspect of the sheath, and makes extensive contact with the basal lamina. C: APP K + ; a sprout pierces the myelin sheath, makes extensive contacts with the membrane of the Schwann cell, and reaches the basal lamina; the parent axon has a normal axoplasm; the sheath, except at the perforation, is well preserved. D: leupeptin; a demyelinated axon (dm) and a small axonal profiles (ap) are embedded in the same Schwann cell but with independent mesaxons. E: leupeptin; demyelinated hourglass-shaped axon, suggestive of a branching point. Calibration, 1 /~m.
R.D. Moreno et al. / Brain Research 718 (1996) 13-24 17
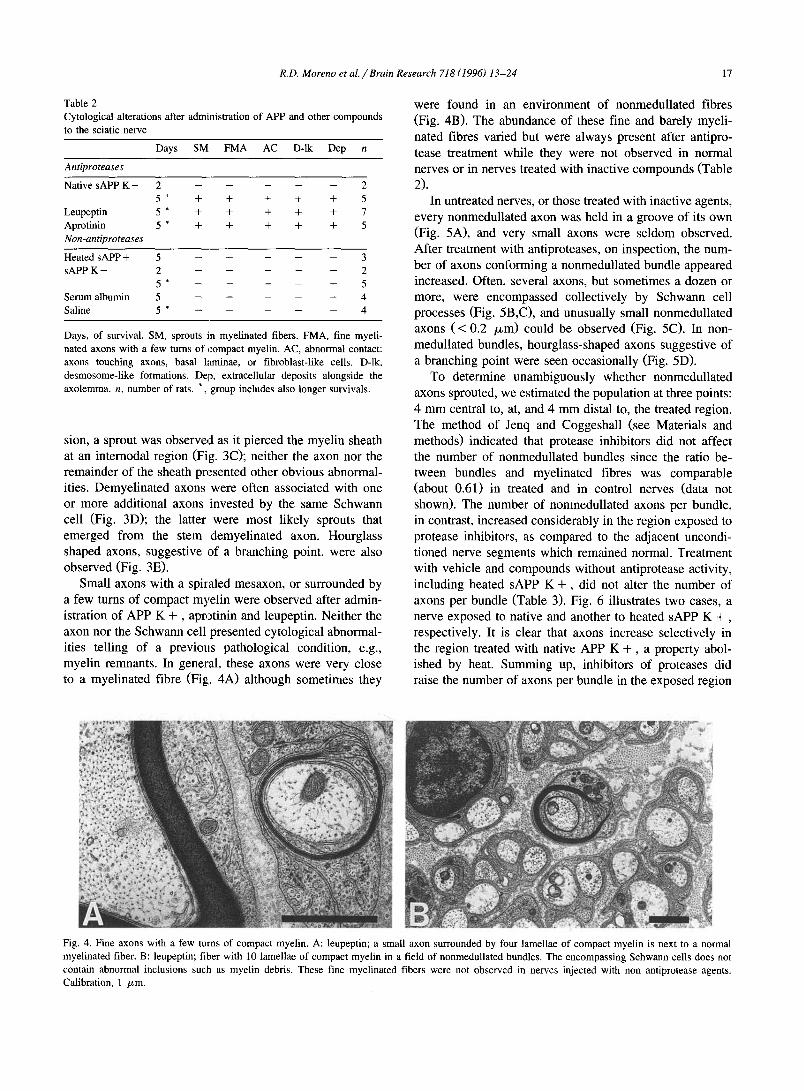
Table 2
Cytological alterations after admi~aistration of APP and other compounds
to the sciatic nerve
Days SM FMA AC D-lk Dep n
Antiproteases
Native sAPP K + 2 . . . . . 2
5 * + + + + + 5 Leupeptin 5 * + + + + + 7
Aprotinin 5 * + + + + + 5
Non-antiproteases
Heated sAPP + 5 . . . . . 3 sAPP K - 2 . . . . . 2
5 * - . . . . 5 Serum albumin 5 . . . . . 4
Saline 5 * - . . . . 4
Days, of survival. SM, sprouts in myel inated fibers. FMA, fine myeli-
nated axons with a few turns of compact myelin. AC, abnormal contact:
axons touching axons, basal laminae, or fibroblast-like cells. D-lk,
desmosome-l ike formations. Dep, extracellular deposits alongside the axolemma, n, number of rats. *, group includes also longer survivals.
sion, a sprout was observed as it pierced the myelin sheath at an internodal region (Fig. 3C); neither the axon nor the remainder of the sheath presented other obvious abnormal- ities. Demyelinated axons were often associated with one or more additional axons invested by the same Schwann cell (Fig. 3D); the latter were most likely sprouts that emerged from the stem demyelinated axon. Hourglass shaped axons, suggestive of a branching point, were also observed (Fig. 3E).
Small axons with a spiraled mesaxon, or surrounded by a few turns of compact myelin were observed after admin- istration of APP K + , aprotinin and leupeptin. Neither the axon nor the Schwann cell presented cytological abnormal- ities telling of a previous pathological condition, e.g., myelin remnants. In general, these axons were very close to a myelinated fibre (Fig;. 4A) although sometimes they
were found in an environment of nonmedullated fibres (Fig. 4B). The abundance of these fine and barely myeli- nated fibres varied but were always present after antipro- tease treatment while they were not observed in normal nerves or in nerves treated with inactive compounds (Table 2).
In untreated nerves, or those treated with inactive agents, every nonmedullated axon was held in a groove of its own (Fig. 5A), and very small axons were seldom observed. After treatment with antiproteases, on inspection, the num- ber of axons conforming a nonmedullated bundle appeared increased. Often, several axons, but sometimes a dozen or more, were encompassed collectively by Schwann cell processes (Fig. 5B,C), and unusually small nonmedullated axons ( < 0.2 /xm) could be observed (Fig. 5C). In non- medullated bundles, hourglass-shaped axons suggestive of a branching point were seen occasionally (Fig. 5D).
To determine unambiguously whether nonmedullated axons sprouted, we estimated the population at three points: 4 mm central to, at, and 4 mm distal to, the treated region. The method of Jenq and Coggeshall (see Materials and methods) indicated that protease inhibitors did not affect the number of nonmedullated bundles since the ratio be- tween bundles and myelinated fibres was comparable (about 0.61) in treated and in control nerves (data not shown). The number of nonmedullated axons per bundle, in contrast, increased considerably in the region exposed to protease inhibitors, as compared to the adjacent uncondi- tioned nerve segments which remained normal. Treatment with vehicle and compounds without antiprotease activity, including heated sAPP K + , did not alter the number of axons per bundle (Table 3). Fig. 6 illustrates two cases, a nerve exposed to native and another to heated sAPP K + , respectively. It is clear that axons increase selectively in the region treated with native APP K + , a property abol- ished by heat. Summing up, inhibitors of proteases did raise the number of axons per bundle in the exposed region
Fig. 4. Fine axons with a few turns of compact myelin. A: leupeptin; a small axon surrounded by four lamellae of compact myel in is next to a normal myelinated fiber. B: leupeptin; fiber with 10 lamellae of compact myel in in a field of nonmedullated bundles. The encompassing Schwann cells does not contain abnormal inclusions such as myel in debris. These fine myelinated fibers were not observed in nerves injected with non antiprotease agents. Calibration, 1 /xm.

18 R.D. Moreno et al. /Brain Research 718 (1996) 13-24
bu t no beyond , wh i l e the p o p u l a t i o n o f bund l e s r e m a i n e d
u n c h a n g e d . H e n c e axons were c o n t i n u o u s fo l l owing inhibi -
t ion o f ex t race l lu la r p ro teases bu t the p o p u l a t i o n inc reased
cons ide rab ly ( 4 0 - 9 0 % ) in the t rea ted segment , i.e., non-
m e d u l l a t e d axons d e v e l o p e d a spou t ing response .
3.2. A b n o r m a l contac ts be t ween axons a n d S c h w a n n cel ls
In a n o r m a l nerve , the a x o l e m m a of m y e l i n a t e d and
n o n m e d u l l a t e d axons is a lways c lose ly apposed to a
S c h w a n n cell m e m b r a n e , excep t at d iscre te regions , e.g.,
node o f Ranvie r , t e rminals . A d i f fe ren t p ic ture was seen
af ter admin i s t r a t i on o f s A P P K + , ap ro t in in and leupept in .
As m e n t i o n e d a l ready, a fu r row in the S c h w a n n cell
a c c o m m o d a t e d m a n y n o n m e d u l l a t e d axons , w h e r e b y axons
apposed ex tens ive ly to each o ther (Fig. 5B,C; see also Fig.
8A). The in te rce l lu la r c lef t was f r equen t ly w ide r than usual
(Fig. 8B). N o n m e d u l l a t e d axons and sprouts o f m y e l i n a t e d
axons s o m e t i m e s touched a long span o f basa l l a m i n a (Fig.
Fig. 5. Abnormal axonal contacts. A: control bundle; APP K + (B,E,F); leupeptin (C,D). In a normal bundle, Schwann cell processes separate axons from each other and from the basal lamina (A). After treatment with protease inhibitors, axons touch each other (B,C), and the basal lamina (C). Axonal profiles are usually of a regular size (compare A and B) but sometimes exceedingly small profiles can be seen after treatment (C, arrows), suggestive of sprouts, as well as hourglass-shaped nonmedullated axons, suggestive of a branching point (D). Desmosome-like densities are frequent after treatment; single densities in a Schwann cell process face an axon (D, fine arrows); double densities may involve two Schwann cell processes (D, coarse arrows) or a Schwann cell process and an axon (C and D, arrowheads). Strands bridge the gap between densities of opposing membranes, and bundles of fibrils may be recognized in the cytoplasmic aspect (D, inset that corresponds to the upper coarse arrow). A large axon, probably a sprout of a myelinated fiber, is partially invested by layers of a fibroblast-like cell (E, arrowheads). At a higher magnification, the basal lamina of the Schwann cell is clearly seen (F, arrow) whereas the fibroblast-like cells does not have it (F, arrowheads). Calibration, 0.5 /zm.

R.D. Moreno et al. / Brain Research 718 (1996) 13-24 19
3B,C). We even observed axons, probably sprouts of myelinated axons, partially encompassed by thin processes of fibroblast-like cells (Fig. 5E,F). In normal nerves or those treated with non anfiprotease compounds, such ab- normal associations were not seen.
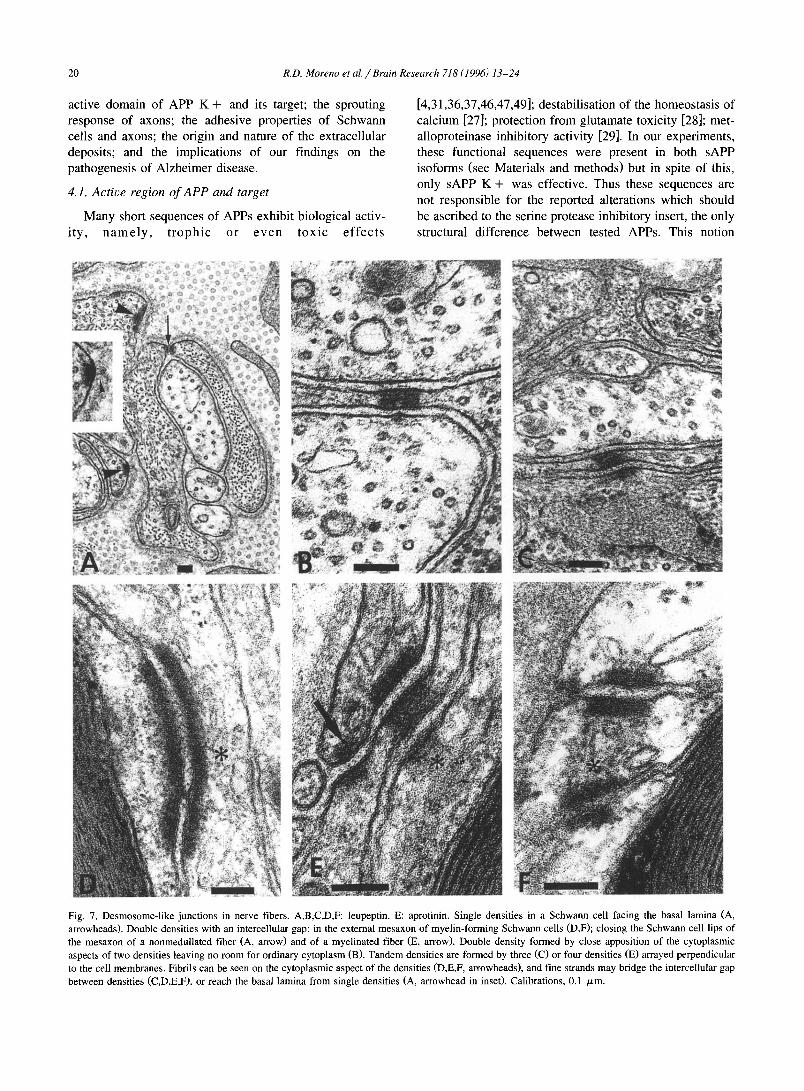
After antiprotease treatment, subplasmalemmal densi- ties were regularly observed in Schwann ceils irrespective of whether they formed myelin or not, and less frequently, in axons (Fig. 5C,D; Fig. 7). Prevailing lengths were ~ 100 nm, and widths, .-~ 30 nm. Fibrils could be ob- served on the cytoplasmic aspect of the densities; in some cases the diameter was close to the width of the plas- malemma, and in others, considerably larger, 10-12 nm (Fig. 7B,D,F). These sizes are reminiscent of micro- and intermediate filaments. Densities were observed in three associations: single, pair, and tandem. Single densities were abundant in Schwann cells; they faced the basal lamina (Fig. 7A), another Schwann cell process, or a nonmedullated axon (Fig. 5D). In front of the density, a fluffy material was sometimes recognized between the Schwann cell membrane and the basal lamina (Fig. 7A). In nonmedullated axons, single densities were seen occasion- ally, and never in myelinated axons.
Paired densities occured in myelin- and nonmyelin-for- ruing Schwann cells. Pairs. were observed in the last turns of the myelin that had became loose, in the external mesaxon, close to the basal lamina (Fig. 7D,E,F), and occasionally in the lips of the external mesaxon. In Schwann cells investing nonmedullated bundles, paired densities could involve two adjacent processes, one density facing the other across a cleft of intercellular space (Fig. 5D). Density pairs also developed between a Schwann cell and a nonmedullated axon (Fig. 5C,D) but these were less frequent. In nonmedullated fibres, density pairs were also found between the lips of the mesaxon (Fig. 7A). The cleft between densities sometimes was bridged by strands of an electron dense material (Fig, 5C,D; Fig. 7). The intercellu- lar gap at the density pair was most of the time similar to the distance between membranes away from the gap but occasionally it was conspicuously widened (Fig. 7E). Fi- nally, a density pair could arise also by apposition of the cytoplasmic aspects of two densities held in a thin Schwann cell process, leaving no room for ordinary cytoplasm (Fig. 7B). We think that these are tandem configurations (see below) in which the density of the cell across the intercel- lular cleft was smaller and did not fall in the plane of the section.
We denote tandem configuration arrays of three or four densities organized in a cascade involving at the same time plasmalemma and cytoplasmic faces. These tandems were found in Schwann cells associated with myelinated and with nonmedullated fibres (Fig. 7C,E). Ocassionally, a nonmedullated axon contributed to a tandem. In normal nerves or those treated with non-antiproteases, the densi- ties described above were exceptional in Schwann cells and never recognized in axons.
12,
10'
g 8
0
~ 2
"[~
I l
native heated
Fig. 6. Nonmedullated fibers per bundle. Two nerves, one treated with native APP K + , and the other, with heated APP K + . Axonal profiles (mean + S.E.M.) were assessed 4 -5 mm proximal to ( - ), at (0), and 4 - 5 mm distal to ( + ) , the site of injection. More than 50 bundles were assessed at each point. *, P < 0.05 (ANOVA, and Student /Newman- Keuls tes0. Native APP K + raises locally the number of nonmedullated axons at the site of injection, property that is abolished by heat (for complete survey, see Table 2).
Anchoring junctions are seen as subplasmalemmal den- sities similar to those reported here. These junctions bind a cell to another cell or to the extracellular matrix, and involve cytoskeletal, transmembrane, and extracellular ma- trix proteins. Single densities reveal focal contacts and hemidesmosomes while paired densities reveal adhesion belts and desmosomes. On the cytoplasmic aspect, anchor- ing junctions associate characteristically with intermediate filaments or with microfilaments. The densities that we found in treated nerves share some of these features whereby we cannot identify them with certitude. Their general structure notwithstanding grants that they belong to the class of anchoring junctions.
3.3. Extracellular deposits
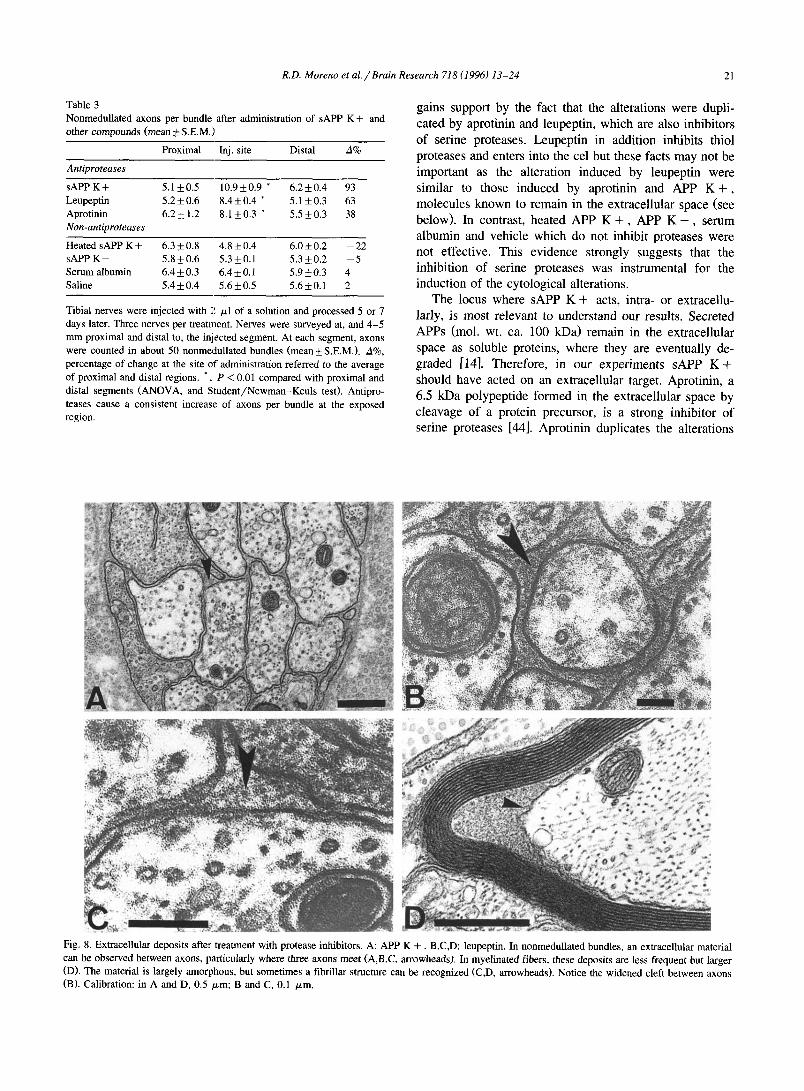
In normal nonmedullated bundles, the cleft between axon and Schwann cell is narrow and electron lucent. After treatment with sAPP K + , leupeptin or aprotinin, the cleft was often widened and extracellular material was observed at some points, particularly where three nonmedullated fibres met (compare Fig. 5A with Fig. 7A and Fig. 8A,B,C). A material of the same kind was also observed between axon and myelin sheath; in these cases, the myelin was thin, probably corresponding to remyelination (Fig. 8D). These deposits were found more often associated with nonmedullated axons but the size was greater in myeli- nated fibres. The material was largely amorphous, but fibrils could be clearly discerned in some instances. The diameter of these fibrils was about 7 nm (Fig. 8C,D).
4. Discussion
sAPP K + but not sAPP K - affects the cytology of axons and Schwann cells, and induces the accumulation of an extracellular material. These observations bear on the
20 R.D. Moreno et al./ Brain Research 718 (1996) 13-24
active domain of APP K + and its target; the sprouting response of axons; the adhesive properties of Schwann cells and axons; the origin and nature of the extracellular deposits; and the implicat ions of our f indings on the pathogenesis of Alzhe imer disease.
4.1. Active region o f APP and target
Many short sequences of APPs exhibit biological activ- i ty , n a m e l y , t r o p h i c or e v e n t o x i c e f f e c t s
[4,31,36,37,46,47,49]; destabil isat ion of the homeostasis of ca lc ium [27]; protection from glutamate toxicity [28]; met- al loproteinase inhibi tory activity [29]. In our experiments, these funct ional sequences were present in both sAPP isoforms (see Materials and methods) but in spite of this, only sAPP K + was effective. Thus these sequences are not responsible for the reported alterations which should
be ascribed to the serine protease inhibi tory insert, the only structural difference be tween tested APPs. This not ion
Fig. 7. Desmosome-like junctions in nerve fibers. A,B,C,D,F: leupeptin. E: aprotinin. Single densities in a Schwann cell facing the basal lamina (A, arrowheads). Double densities with an intercellular gap: in the external mesaxon of myelin-forming Schwann cells (D,F); closing the Schwann cell lips of the mesaxon of a nonmedullated fiber (A, arrow) and of a myelinated fiber (E, arrow). Double density formed by close apposition of the cytoplasmic aspects of two densities leaving no room for ordinary cytoplasm (B). Tandem densities are formed by three (C) or four densities (E) arrayed perpendicular to the cell membranes. Fibrils can be seen on the cytoplasmic aspect of the densities (D,E,F, arrowheads), and fine strands may bridge the intercellular gap between densities (C,D,E,F), or reach the basal lamina from single densities (A, arrowhead in inset). Calibrations, 0.1 /zm.
R.D. Moreno et al. / Brain Research 718 (1996) 13-24 21
Table 3 Nonmedullated axons per bundle after administration of sAPP other compounds (mean _+ S.E.M.)
K + and
Proximal Inj. site Distal A%
Antiproteases
sAPP K + 5.1_+0.5 10.9_+0.9 * 6.2_+0.4 93 Leupeptin 5.2 _+ 0.6 8.4 _+ 0.4 * 5.1 _+ 0.3 63 Aprotinin 6 .2+1.2 8.1_+0.3 * 5.5_+0.3 38 Non-antiproteases
Heated sAPP K + 6.3_+0.8 4.8_+0.4 6.0_+0.2 - 2 2 sAPP K - 5 .8+0.6 5.3_+0.1 5.3_+0.2 - 5 Serum albumin 6.4 ___ 0.3 6.4 -+ 0.1 5.9 _+ 0.3 4 Saline 5.4_+0.4 5.6_+0.5 5.6-+0.1 2
Tibial nerves were injected with 2 /zl of a solution and processed 5 or 7 days later. Three nerves per treatment. Nerves were surveyed at, and 4 -5 mm proximal and distal to, the injected segment. At each segment, axons were counted in about 50 nonmedullated bundles (mean + S.E.M.). A%, percentage of change at the site of administration referred to the average of proximal and distal regions. *, P < 0.01 compared with proximal and distal segments (ANOVA, and Student /Newman-Keuls test). Antipro- teases cause a consistent increase of axons per bundle at the exposed region.
gains support by the fact that the alterations were dupli- cated by aprotinin and leupeptin, which are also inhibitors of serine proteases. Leupeptin in addition inhibits thiol proteases and enters into the cel but these facts may not be important as the alteration induced by leupeptin were similar to those induced by aprotinin and APP K + , molecules known to remain in the extracellular space (see below). In contrast, heated APP K + , APP K - , serum albumin and vehicle which do not inhibit proteases were not effective. This evidence strongly suggests that the inhibition of serine proteases was instrumental for the induction of the cytological alterations.
The locus where sAPP K + acts, intra- or extracellu- lady, is most relevant to understand our results. Secreted APPs (mol. wt. ca. 100 kDa) remain in the extracellular space as soluble proteins, where they are eventually de- graded [14]. Therefore, in our experiments sAPP K + should have acted on an extracellular target. Aprotinin, a 6.5 kDa polypeptide formed in the extracellular space by cleavage of a protein precursor, is a strong inhibitor of serine proteases [44]. Aprotinin duplicates the alterations
Fig. 8. Extracellular deposits afte:c treatment with protease inhibitors. A: APP K + B,C,D: leupeptin. In nonmedullated bundles, an extracellular material can be observed between axons, particularly where three axons meet (A,B,C, arrowheads). In myelinated fibers, these deposits are less frequent but larger (D). The material is largely amorphous, but sometimes a fibrillar structure can be recognized (C,D, arrowheads). Notice the widened cleft between axons (B). Calibration: in A and D, 0.5 /xm; B and C, 0.1 /zm.

22 R.D. Moreno et a l . / Brain Research 718 (1996) 13-24
observed after APP K + treatment, as also does leupeptin, a tripeptide antiprotease (mw: 476). From these observa- tions we propose that all three agents operate through a common target, an extracellular protease.
4.2. Sprouting
In present experiments, axons were not severed since the nerve was normal distal to the treated segment. After treatment the nonmedullated bundles per myelinated axon remained constant, and the number of myelinated (plus demyelinated) fibres is unlikely to have changed measur- ably, hence the number of bundles also remained constant (for mathematical analysis, see Materials and methods section, Morphometry). The increased number of non- medullated axons per bundle in the conditioned segment (40-90%) indicates thus a local and sizeable sprouting response of uninterrupted nonmedullated axons. This result extends the ability of antiprotease to induce sprouts, al- ready reported for myelinated axons [2], to nonmedullated axons.
In myelinated fibres, the sprouts may originate in the internodal region, as observed. They may also emerge at a node of Ranvier, but such event was not identified. Addi- tional axonal profiles of varying sizes were seen embedded in a Schwann cell that encompassed a demyelinated axon, indicating that demyelinated axons also sprouted. Whether the sprouting response precedes or follows demyelination cannot be answered presently. Fine axons with a few turns of compact myelin appeared after treatment with protease inhibitors; they were seen neither away from the treated region nor in nerves injected with non antiproteases. These fine and thinly myelinated axons are sprouts that left the parent myelinated fibre and are already undergoing myeli- nation; alternatively, former nonmedullated axons acquire the ability to induce myelination under the influence of the treatment.
4.3. Repression of axonal growth
That a normal peripheral nerve represses the growth of axons is an old notion. At the turn of the century, Langley and Anderson [23] showed that axons of a sectioned nerve do not invade a normal nerve but do regrow along a degenerated nerve. Recently, this behaviour has been du- plicated in vivo and in vitro. In the Wld s mouse mutant, both degeneration and regeneration following a nerve crush are delayed by several weeks [8] but if the distal segment is mechanically damaged, axons do regrow until they are halted upon reaching the undamaged nerve tissue [12]. In culture, degenerated nerves or proliferating Schwann cells support the growth of neurites but normal nerves or quies- cent cells do not [5,6]. Descriptively, normal nerves and stabilized Schwann cells are repressive for axonal growth while degenerated nerves and proliferating cells are per- missive. Another line of evidence indicates that in vitro,
proteases affect the development of neurites. In a culture medium supplemented with serum, neurones do not extend processes but they do so after inhibition of thrombin, a serine protease [30], i.e., an extracellular protease makes the environment repressive for the extension of neurites, and its inhibition makes the environment permissive. Our results extend this in vitro pattern to the in vivo condition; in fact, stem axons do not sprout in a normal nerve, but protease inhibitors induce a local sprouting response, i.e., they remove a repression. Moreover, the hypothesis of an ongoing repression predicts that, following a nerve injury, the distal stump will take some time to become permissive, thus explaining the observed 1-2 day delay before the onset of regeneration [7,9,15,38 ]. Contrariwise, a reduction of the delay of regeneration is predicted if the repression is removed. This was observed when the nerve stump distal to the crush was frozen to kill the Schwann cells or conditioned with protease inhibitors [41]. Taken together, the evidence presented strongly support the notions of a sprouting drive of axons and an ongoing repression, in which Schwann cells and a physiological protease partici- pate.
4.4. Extracellular deposits and abnormal contacts
The electron dense material that appeared alongside the axolemma should be proteinaceous. Under the effect of protease inhibitors, axons or Schwann cells may have secreted a novel product; alternatively, the turnover of a normal extracellular component was altered resulting in its accumulation. The identification of this material will help to clarify this issue.
In a normal nerve, Schwann cells are practically the exclusive neighbours of axons. This association was dis- rupted by antiproteases since axons were often contacting extensively other axons, basal laminae and even fibroblast-like cells, and the clefts were wider. These abnormal arrays suggest an alteration of functional compo- nents of the extracellular matrix and/or surface proteins involved in adhesion, e.g., collagen IV, laminin, fi- bronectin, cell adhesion molecules, cadherins, integrins, proteoglycans [10,19,42,43], etc. During development and in the initial stages of regeneration, peripheral axons con- tact each other extensively. The morphological maturation of the nonmedullated bundle includes the encompassing of every axon by Schwann cell processes such that an axon touching the basal lamina or another axon is a rarity in adult peripheral nerves [32]. Thus protease inhibitors in- duce a new organization of the adult nerve tissue that resembles in this respect that of the developing nerve. Other tissues are known to vary cyclically in register with the activity of a protease-antiprotease system that remodels the extracellular matrix, e.g., mammary cells [40]. It is tempting to think that the organization of the nerve as a tissue, among other events, results of the development of a protease-antiprotease system that adjusts continuously the extracellular matrix.

R.D. Moreno et al. / Brain Research 718 (1996) 13-24 23
Anchoring junctions do not belong to the inventory of a normal peripheral nerve [32] although in some nerve fibres of invertebrates desmosome-l ike formations are ubiquitous [20]. In demyelinating diseases of man desmosome-l ike junctions have been noted between astrocytes and oligo- dendrocytes, and between Schwann cell processes [16,34]. In present experiments, junctions were extremely rare in either normal nerves or tho,;e treated with nonantiproteases but after exposure to antiproteases, abundant anchoring junctions occurred between Schwann cell processes or between axons and Schwann cells. Central or distal to the treated segment, axons did not exhibit junctions. In conse- quence, densities occurring in axons should result of a local re-ordering of existing axoplasmic proteins rather than requiring new proteins coming from the perikaryon. Whether re-ordering of existing proteins also holds for Schwann cell densities or the expression of genes is neces- sary, is an open question. In brief, protease inhibitors disrupt locally the mature adhesion between axons and Schwann cells, and exaggerate another type of adhesion that hardly belongs to their inventory.
4.5. B id i rec t iona l regu la tory m e c h a n i s m
When axons degenerate, Schwann cells resorb the myelin sheath, and a burts of mitosis ensues [11], i.e., Schwann cells are regulated by axons. In turn, axons are controlled by Schwann cells, e.g., in vivo, destruction of Schwann cells results in a local sprouting response of axons and a rise of axonal microtubules [13]; in vitro, axons enlarge up to a certain size and further radial growth occurs only upon myelination [48]. Protease inhibitors brake the mutual regulation of axons and Schwann cells, as shown by cell proliferation and myelin resorption [3], sprouting of axons, and disorganization of the nonmedul- lated bundles. This suggests that several programmes of Schwann cells and axons are coherently activated, or deac- tivated, but on a local basi,;.
4.6. A l z h e i m e r d i sease
Finally we want to comment on the pathogenesis of Alzheimer disease in relation to protease inhibitors. The natural history of APP is altered in patients [for a review, see [24]] and, in particular, the isoform with antiprotease activity [45]. Several protease inhibitors are deposited in the plaques, e.g., a 1-antichymotrypsin, a 1-antitrypsin, nexin-I [1,18,35,39]. Sprouts occur around the plaques [17,25,26] which in the light of our results may be induced in vivo by protease inhibitors. Thus the imbalance of an extracellular protease system may participate in the patho- genesis of Alzheimer disease lesions.
In conclusion, in a nerve treated with sAPP K + or other inhibitors of serine preoteases, a sprouting response of axons ensues, the cellular contacts are disrupted, and extracellular deposits appear. We propose that axons are
endowed with a sprouting programme repressed continu- ously; that the repression involves the extracellular matrix and a protease-antiprotease system; and that this system participates in the pathogenesis of Alzheimer ' s disease.
Acknowledgements
R.M. was a recipient of a fellowship of the P. Universi- dad Catt l ica. We thank M. Ptrez, G. Mtndez , and J. Moril las for technical assistance. This project was sup- ported by grants FONDECYT 0 6 6 0 / 9 2 and 1950999, Stiftung Volkswagenwerk, and Deutsche Gesellschaft ftir Technische Zusammenarbei t GmbH.
References
[1] Abraham, C.R., Selkoe, D.J. and Potter, H., Immunochemical identi- fication of the serine protease inhibitor a 1-antichymotrypsin in the brain amyloid deposits of Alzheimer disease, Cell, 52 (1988) 487- 501.
[2] Alvarez, J., Moreno, R.D., Llanos, O., Inestrosa, N.C., Brandan. E., Colby, T. and Esch, F.S., Axonal sprouting induced in the sciatic nerve by the amyloid precursor protein (APP) and other antipro- teases, Neurosci. Lett., 144 (1992) 130-134.
[3] Alvarez, J., Moreno, R.D. and Inestrosa, N.C., Mitosis of Schwann cells and demyelination are induced by the amyloid precursor pro- tein and other protease inhibitors in the rat sciatic nerve, Eur. J. Neurosci.. 7 (1995) 152-159.
[4] Araki, W., Kitaguchi, N., Tokushima, Y., Kazuhiro, I., Aratake, H., Shimohama, S., Nakamura, S. and Kimura, J., Trophic effect of /3-amyloid precursor protein on cerebral cortical neurons in culture, Biochem. Biophys. Res. Comm., 181 (1991) 265-271.
[5] Baehr, M. and Bunge, R.P., Functional status influences the ability of Schwann cells to support adult rat retinal ganglion cell survival and axonal regrowth, Exp. NeuroL, 106 (1989) 27-40.
[6] Bedi, K.S., Winter, J., Berry, M. and Cohen, J., Adult rat dorsal root ganglion neurons extend neurites on predegenerated but not on normal peripheral nerves in vitro, Eur. J. Neurosci., 4 (1992) 193-200.
[7] Beremberg, R.A., Forman, D.S., Wood, D.K., De Silva, A. and Demaree, J., Recovery of peripheral nerve function after axotomy: effect of triiodothyramine, Exp. Neurol., 57 (1977) 349-363.
[8] Bisby, M.A. and Chen, S., Delayed wallerian degeneration in sciatic nerves of C57B1/Ola mice is associated with impaired regeneration of sensory axons, Brain Res., 530 (1990) 117-120.
[9] Bisby, M.A. and Keen, P., The effect of a conditioning lesion on the regeneration rate of peripheral nerve axons containing substance P, Brain Res., 336 (1985) 201-206.
[10] Bixby, J.L., Pratt, R.L., Lilien, J. and Reichardt, L.F., Neurite outgrowth on muscle cell surfaces involves extracellular matrix receptors as well as Ca2+-dependent and -independent cell adhesion molecules, Proc. Nat. Acad. Sci. USA, 84 (1987) 2555-2569.
[11] Bradley, G.W. and Asbury, A.K., Duration of synthesis phase in neurilemma cells in mouse sciatic nerve during degeneration, Exp. Neurol., 26 (1970) 275-282.
[12] Brown, M.C., Perry, V.H., Hunt, S.P. and Laper, S.R., Further studies on motor and sensory nerve regeneration in mice with delayed Wallerian degeneration, Eur. J. Neurosci., 6 (1994) 420- 428.
[13] Bustos, J., Vial, J.D., Fafindez, V. and Alvarez, J., Axons sprout and microtubules increase after local inhibition of RNA synthesis, and

24 R.D. Moreno et al . /Brain Research 718 (1996) 13-24
microtubules decrease after inhibition of protein synthesis: a mor- phometric study in rat sural nerves, Eur. J. Neurosci., 3 (1991) 1123-1233.
[14] Esch, F.S., Keim, P.S., Beattie, E.C., Blacher, A.R., Culwell, R.C., Oltersdorf, T., McClure, D. and Ward, J.P., Cleavage of amyloid /3 peptide during constitutive processing of its precursor, Science, 248 (1990) 1122-1124.
[15] Forman, D.S., McQuarrie, I.G., Labare, F.W., Wood, D.K., Stone, L.S., Braddock, C.H. and Fuchs, D.A., Time course of the condition- ing lesion effect on axonal regeneration, Brain Res., 182 (1980) 180-185.
[16] Ghatak, N.R., Occurrence of oligodendrocytes within astrocytes in demyelinating lesions, J. Neuropath. Exp. Neurol., 51 (1992)40-46.
[17] Geddes, J.W., Anderson, K.J. and Cotman, C.W., Senile plaques as aberrant sprout-stimulating structures, Exp. Neurol., 94 (1990) 767- 776.
[18] Gollin, P.A., Kalaria, R.N., Eikelenboom, P., Rozemuller, A. and Perry, G., ~ 1-antitrypsin and a l-antichymotrypsin are in the lesions of Alzheimer's disease, NeuroReport, 3 (1992) 201-203.
[19] Hall, D.E., Neugebauer, K.M. and Reichardt, L.F., Embryonic neu- ral retinal cell response to extracellular matrix proteins: developmen- tal changes and effects of the CSAT antibody, J. Cell Biol., 104 (1987) 623-634.
[20] Heuser, J.E. and Doggenweiler, C.F., The fine structural organiza- tion of nerve fibers, sheaths, and glial cells in the prawn Palaemon- etes vulgaris, J. Cell Biol., 66 (1966) 381-403.
[21] Inestrosa, N.C. and Soto, C., Molecular biology of the amyloid of Alzheimer's disease, Biol. Res., 25 (1992) 63-72.
[22] Jenq, C-B. and Coggeshall, R.E., Long-term patterns of axon regen- eration in the sciatic nerve and its tributaries, Brain Res., 345 (1985) 34-44.
[23] Langley, J.N. and Anderson, H.K., The union of different kinds of nerve fibres, J. Physiol., 31 (1904)365-391.
[24] Marotta, C.A., Majocha, R.E. and Tate, B., Molecular and cellular biology of Alzheimer amyloid, J. Mol. Neurosci., 3 (1992) 111-125.
[25] Masliah, E., Mallory, M., Hansen, L., Alford, M., DeTeresa, R., Terry, R., Baudier, J. and Saitoh, T., Localization of amyloid precursor protein in GAP43-immunoreactive aberrant sprouting neu- rites, Brain Res., 574 (1992) 312-316.
[26] Masliah, E., Mallory, M., Deerinck, T., DeTeresa, R., Lamont, S., Miller, A., Terry, R., Carragher, B. and Ellisman, M., Re-evaluation of the structural organization of neuritic plaques in Alzheimer's disease, J. Neuropathol. Exp. Neurol., 52 (1993) 619-632.
[27] Mattson, M.P., Cheng, B., Davies, D., Bryant, K., Lieberburg, I. and Rydel, R.E., /3-amyloid peptides destabilize calcium homeostasis and render human cortical neurons vulnerable to excitotoxicity, J. Neurosci., 12 (1992) 376-389.
[28] Mattson, M.P., Cheng, B., Culwell, A.R., Esch, F.E., Lieberburg, I. and Rydel, R.E., Evidence for excitoprotective and intraneuronal calcium-regulating roles for secreted froms of the /3-amyloid precur- sor protein, Neuron, 10 (1993) 243-254.
[29] Miyazaki, K., Hasegawa, M., Funahashi, K., Umeda, M.A., Metallo- proteinase inhibitor domain in Alzheimer amyloid protein precursor, Nature, 362 (1993) 839-841.
[30] Monard, D., Cell-derived proteases and protease inhibitors as regula- tors of neurite outgrowth, Trends Neurosci., 11 (1988) 541-544.
[31] Ninomiya, H., Roch, J-M., Sundsmo, M.P., Otero, D.A.C. and Saitoh, T., Amino acid sequence REMS represents the active domain of amyloid /3 /A4 protein precursor that promotes fibroblast growth, J. Cell Biol., 121 (1993) 879-886.
[32] Peters, A., Palay, S.L. and Webster, H.deF., The Fine Structure of the Nervous System, Oxford Univ. Press, Oxford, 1991, pp. 218-264
[33] Pittman, R.N. and Buettner., Degradation of extracellular matrix by neuronal proteases, Dev. Neurosci., 11 (1989) 361-375.
[34] Prineas, J.W., Acute idiopathic polyneuritis. An electron microscope study, Lab. Invest., 26 (1972) 133-147.
[35] Rosenblatt, D.E., Geula, C. and Mesularn, M.M., Protease nexin 1 immunostaining in Alzheimer's disease, Ann. Neurol., 26 (1989) 628-634.
[36] Saitoh, T., Sundsmo, M., Roch, J-M., Kimura, N., Cole, G., Schu- bert, D., Oltersdorf, T. and Schenk, D.B., Secreted form of amyloid protein precursor is involved in the growth regulation of fibroblasts, Cell, 58 (1989) 615-622.
[37] Schubert, D., Jin, L.W., Saitoh, T. and Cole, G., The regulation of amyloid /3 protein precursor secretion and its modulatory role in cell adhesion, Neuron, 3 (1989) 689-694.
[38] Sketelj, J., Bresjanac, M. and Popovic, M., Rapid growth of regener- ating axons across the segments of sciatic nerve devoid of Schwann cells, J. Neurosci. Res., 24 (1989) 153-162.
[39] Takahashi, H., Hirokawa, K. and Tsukagoshi, H., Immunohistologi- cal study on the distribution of a l-antichymotrypsin in Alzheimer's brain, compared to /3-amyloid precursor protein and /3-amyloid protein, J. Neurol. Sci., 99 (1990) 301-310.
[40] Talhouk, R.S., Bissell, M.J. and Werb, Z., Coordinated expression of extracellular matrix-degrading proteinases and their inhibitors regulates mammary epithelial function during involution, J. Cell Biol., 118 (1992) 1271-1282.
[41] Tapia, M., Inestrosa, N.C. and Alvarez, J., Early axonal regenera- tion: repression by Schwann cells and a protease?, Exp. Neurol., 131 (1995) 124-132.
[42] Tomaselli, K.J., Damsky, C.H. and Reichardt, L.F., Interactions of a neuronal cell line (PC 12) with laminin, collagen IV, and fi- bronectin: identification of integrin-related glycoproteins involved in attachment and process outgrowth, J. Cell Biol., 105 (1987) 2347- 2358.
[43] Tomaselli, K.J., Neugebauer, K.N., Bixby, J.L., Lilien, J. and Re- ichardt, L.F., N-cadhherin and integrins: two receptor systems that mediate neuronal process outgrowth on astrocyte surfaces, Neuron, 1 (1988) 33-43.
[44] Trautschold, E., Werle, E. and Zickgraf-Rudel, G., Trasylol, Biochem. Pharmacol., 16 (1967) 59-72.
[45] Van Nostrand, W.E., Farrow, J.S., Wagner, S.L., Bhasin, R., Goldgaber, D., Cotman, C.W. and Cunningham, D.D., The predomi- nant form of the amyloid /3-protein precursor in human brain is protease nexin 2, Proc. Natl. Acad. Sci. USA, 88 (1991) 10302- 10306.
[46] Whitson, J.S., Glabe, C.G., Shintani, E., Abcar, A. and Cotman, C.W., Amyloid protein promotes neuritic branching in hippocampal cultures, Neurosci. Lett., 110 (1990)319-324.
[47] Whitson, J.S., Selkoe, D.J. and Cotman, C.W., Amyloid r-protein enhances the survival of hippocamplal neurons in vitro, Science, 243 (1989) 1488-1490.
[48] Windebank, A.J., Wood, P., Bunge, R.P. and Dyck, P.J., Myelina- tion determines the caliber of dorsal root ganglion neurons in culture, J. Neurosci., 5 (1985) 1563-1569.
[49] Yankner., B.A., Dawes, L.R., Fisher, S., Villa-Komaroff, L., Oster- Granite, M.L. and Neve, R.L., Neurotoxicity of a fragment of the amyloid precursor associated with Alzheimer's disease, Science, 245 (1989) 417-420.