
Photodynamic Therapy Using a Protoporphyrinogen Oxidase Inhibitor1

1 23
Applied Biochemistry andBiotechnologyPart A: Enzyme Engineering andBiotechnology ISSN 0273-2289Volume 171Number 7 Appl Biochem Biotechnol (2013)171:1739-1749DOI 10.1007/s12010-013-0312-1
Specificity of Glucose Oxidase fromPenicillium funiculosum 46.1 TowardsSome Redox Mediators
Tatiana Semashko, Raisa Mikhailova,Almira Ramanaviciene & ArunasRamanavicius

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media New York. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Specificity of Glucose Oxidase from Penicilliumfuniculosum 46.1 Towards Some Redox Mediators
Tatiana Semashko & Raisa Mikhailova &
Almira Ramanaviciene & Arunas Ramanavicius
Received: 8 October 2012 /Accepted: 27 May 2013 /Published online: 31 August 2013# Springer Science+Business Media New York 2013
Abstract Glucose oxidase (GOx) from Penicillium funiculosum 46.1 was purified usingstep-by-step ultrafiltration and it was characterized by spectrophotometric and spectrofluoromet-ric methods. It was shown that spectra of GOx produced by P. funiculosum are typical forflavoproteins. Absorption spectrum has distinct peaks at 380 and 457 nm, excitation spectrum at373 and 447 nm, and emission spectrum at 530 and 562 nm. The pH correlation of enzymeactivity and catalytic characteristics in various buffer systems (phosphate (pH 5.0–9.0), citrate(pH 3.0–5.0), citrate-phosphate (pH 3.0–9.0), and universal (pH 3.0–9.0)) were registered. It wasdetermined that the GOx is the most efficiently interacting with substrate (glucose) in phosphatebuffer at pH 7.0 with kcat/Km=21,825 M−1 s−1. Interaction of several different redox mediators(9,10-phenantroline-5,6-dione, 9,10-phenanthrenequinone, N-methylphenazonium methyl sul-fate, ferrocene, ferrocenecarboxylic acid,α-methylferrocenemethanol, ferrocenecarboxaldehyde)with GOx from P. funiculosum was investigated by evaluation of the difference in fluorescenceemission intensity of FAD(oxidized) and FADH2(reduced) forms. It was found that 9,10-phenantroline-5,6-dione and 9,10-phenanthrenequinone are the best redox mediators for thistype of GOx.
Appl Biochem Biotechnol (2013) 171:1739–1749DOI 10.1007/s12010-013-0312-1
T. Semashko : R. MikhailovaInstitute of Microbiology of Belarus National Academy of Sciences,Kuprevich str., 2, Minsk 220141, Belarus
A. RamanavicieneNanoTechnas—Center of Nanotechnology and Materials Science at Department of Analyticaland Environmental Chemistry, Faculty of Chemistry, Vilnius University, Naugarduko 24,03225 Vilnius, Lithuania
A. RamanaviciusDepartment of Physical Chemistry, Faculty of Chemistry, Vilnius University,Naugarduko 24, 03225 Vilnius, Lithuania
A. Ramanavicius (*)Laboratory of BioNanoTechnology, Department of Material Science and Electrical Engineering,Semiconductor Physics Institute, State Research Institute Center for Physical and Technological Sciences,A. Gostauto 11, 01108 Vilnius, Lithuaniae-mail: [email protected]
Author's personal copy
Keywords Glucose oxidase . Penicillium funiculosum . Redoxmediators . Spectrofluorimetry.
Biosensors . Biofuel cells
Introduction
Glucose oxidase (GOx) (β-D-glucose/oxygen 1-oxidoreductase, EC 1.1.3.4) is FAD-containing enzyme, which is belonging to the class of oxidoreductases. The GOx catalyzesoxidation of β-D-glucose to D-glucono-1,5-lactone and hydrogen peroxide. In the reactioncatalyzed by this enzyme, the β-D-glucose serves as a donor of electrons and hydrogen ions,on other side of this complex reaction, oxygen dissolved in water-based reaction media is asan acceptor. Coenzyme FAD acts as electron shuttle during catalytic action of this enzyme[1] (Fig. 1).
It should be noted that apart from the molecular oxygen, the GOx may engage verydiverse chemical agents as electron acceptors that are called “redox mediators.” Theapplication of redox mediators ensures more effective electron transport from the active siteof the enzyme [2–5]. High specificity of the GOx towards glucose enables to detectconcentration of this substrate in very complicated samples including blood, saliva, urine,and other body fluids, which contain many other sugars and various reducing non-carbohydrate-based agents. Therefore, the glucose oxidase has been widely applied inbiosensors suitable for express monitoring of glucose and analytical control of manybiological processes. Such biosensors usually are suitable for food processing, medicine,and eco-friendly technologies [6–9]. A promising trend of glucose oxidase application is thedesign of biofuel cells as alternative energy sources [10–16]. It should be noted that recentlyspecial emphasis is focused on the development of enzyme-based biofuel cells, which
Fig. 1 Principle scheme of glucose oxidation reaction catalyzed by GOx
1740 Appl Biochem Biotechnol (2013) 171:1739–1749
Author's personal copy

are used as batteries for the powering of implanted biomedical devices suitable forcontinuous monitoring of glucose. However, currents achieved by these devices stillremain relatively low. Therefore, further progress in the performance of biosensorsand biofuel cells could be achieved by the application of most efficient redoxmediators. There are many substances suitable for electrochemical “wiring” of GOxwith electrodes [17, 18]. Nevertheless, the significance of studies devoted to interac-tion of redox mediators with enzymes remains extremely relevant [13, 14, 16]. Theefficiency of GOx applied in biosensor technologies and biofuel cells is also largelydependent on strains producers and catalytic properties of purified enzymes. Variousglucose oxidases are found in different microorganisms and even in multicellularorganisms [8, 19–26], but the main GOx sources are fungi of genera Aspergillusand Penicillium [27–37].
The screening for strain producer of GOx at the Laboratory of Enzymes, Instituteof Microbiology, which belongs to Belarusian Academy of Sciences, was initiated in1996 [24]. Advanced interest to fungi such as Penicillium genus is based on the factthat the GOx produced by fungi of Penicillium genus show higher affinity to β-D-glucose than many other similar enzymes isolated from different strains of Aspergillusfungi. For example, the value of Michaelis constant (Km) of Aspergillus GOx variesfrom 23.7 to 37.0 mM [27, 30, 32, 34], whereas for GOx from Penicillium, it is inthe range of 3.3–19.0 mM [29, 31, 33, 34, 36, 37]. We constructed highly activestrain producer of extracellular GOx Penicillium funiculosum 46.1 by induced muta-genesis technique [38]. Mutant strain P. funiculosum 46.1 exceeded the parent P.funiculosum BIM F-15 in specific growth rate and glucose oxidase production, andlevel of synthesis of extracellular glucose oxidase and invertase. The GOx from P.funiculosum 46.1 differs from GOx purified from parent culture by Michaelis constant,efficiency of glucose oxidation, pH dependence, and thermal stability, but it has similarthermal optimum. It was shown that enzyme-encoding gene has no special mutation.Homology of amino acid and nucleotide sequences of mutant and parent strains wasin the range of 100 %. The composition of nutrient medium and fermentation conditionsof P. funiculosum 46.1 were optimized; advanced methods of enzyme purification wereelaborated [39–44].
The aim of this study was the evaluation of P. funiculosum 46.1 GOx activity in thepresence of various redox mediators.
Materials and Methods
Chemicals
Major object of studies is extracellular GOx produced by mycelial fungus P. funiculosum46.1. D-(+)-Glucose was obtained from Carl Roth GmbH&Co (Karlsruhe, Germany). The1.0 M solution of glucose was prepared in distilled water at least 24 h before use in order toallow glucose to mutarotate and to reach the steady-state equilibrium between α- and β-forms.Solution (0.1%) of p-benzoquinone (Fluka) for the determination of GOx activity was preparedin distilled water. For experiments, 0.01M phosphate (pH 5.0–9.0), citrate (pH 3.0–5.0), citrate-phosphate (pH 3.0–9.0), and universal (pH 3.0–9.0) buffer systems were used. 9,10-Phenantroline-5,6-dione (PD) (Sigma-Aldrich), 9,10-phenanthrenequinone (PQ) (Sigma-Al-drich), N-methylphenazonium methyl sulfate (PMS) (Sigma-Aldrich), ferrocene (Fer) (Minsk,
Appl Biochem Biotechnol (2013) 171:1739–1749 1741
Author's personal copy

Belarus), ferrocenecarboxylic acid (FCA) (Fluka), α-methylferrocenemethanol (MFM)(Aldrich), and ferrocenecarboxaldehyde (FCL) (Fluka) were used as mediators.
Production and Purification of GOx
The production of GOx was performed by ultrafiltration-based concentration of culturalliquid resulting from submerged fermentation of P. funiculosum 46.1. The fungus was grownin modified liquid nutrient media, pH 5.0, containing 6 % of sucrose as carbon source [36].The fungus was cultured in 250-ml Erlenmeyer flasks, each containing 50 ml of the medium,at 180 rpm and 24–26 °C for 96 h. The density of given seeding was in the range of 2×105–3×105 spores/ml. When the cultivation was completed, the biomass was separated byfiltering, and obtained culture filtrate was used for further isolation of GOx.
Ultrafiltration was conducted using laboratory device based on hollow fiber membraneelement MPVE PS-10М-0.2 (MIFIL, Minsk, Belarus) and UFYA-47 cell (Biotest Company,Kirishi, Russia). Hollow fiber membrane element contained membrane, which wascomposed of aromatic polysulfone with protected surface PS-10М (nominal molecularweight retention limit 10 kDa). For cell ultrafiltration membrane, UPM-10 forming aporous polymeric film based on aromatic polysulforamide Sulfon-IT or polypropylenesupport (retention limit 10 kDa) (Vladipore, Vladimir, Russia) was applied. SDS elec-trophoresis based on Laemmli method [45] was performed for evaluation of enzymepurity.
GOx Activity Assay
The method for the determination of GOx activity was based on enzymatic conversion ofp-benzoquinone into hydroquinone and the measurement of hydroquinone formation rate at290 nm. For this assay as a unit of enzymatic activity (U), the amount of enzyme able tocatalyze transformation of 1 mmol×l−1 p-benzoquinone into hydroquinone during 1 min at25 °C was defined. Enzyme activity is expressed in unit per milliliter (U=mmol×l−1×min−1),unit per milligram protein, or mole per liter per second [46, 47].
Spectrofluorometric and Spectrophotometric Analysis of GOx Activity
Analysis of glucose oxidase was performed using spectrofluorimeter Solar CM2203(Belarus) and spectrophotometer Ultraspec 2100 (Amersham Pharmacia Biotech, Sweden).FAD absorption spectrum was analyzed in the range of 300–500 nm, FAD excitationspectrum at 300–500 nm (at l emission 530 nm), and FAD emission spectrum at 500–600 nm (at l excitation 373, 457 nm). The spectra were measured in 10 mm optical pathlength closed quartz cells. The absorption, excitation, and emission slits were set at 1 nmeach. The measurements were done at ambient room temperature.
Determination of Kinetic Parameters
The apparent Michaelis constant (Km) and maximal rate of glucose oxidation (Vm) both werecalculated in order to evaluate the biological activity of enzyme. The Lineweaver–Burk plot(1/V0; 1/Ms−1) vs. (1/S0; 1/M) was applied for the graphical evaluation of results. Catalyticconstant (kcаt) was calculated using the formula: kcаt=Vm/[GOx]. Molar concentration ofGOx was calculated by the evaluation of FAD spectra using molar extinction coefficient of2.12·104 М−1сm−1 at 457 nm.
1742 Appl Biochem Biotechnol (2013) 171:1739–1749
Author's personal copy

Determination of Reaction Rate of GOx Catalyzed Reaction in the Presenceof Different Redox Mediators
In order to select the optimal buffer composition, which is the most suitable for maximalenzymatic activity, the following 0.01 M buffering systems were tested: phosphate buffer(pH 5.0–9.0), citrate buffer (pH 3.0–5.0), citrate-phosphate buffer (pH 3.0–9.0), and univer-sal buffer, which consist of mixture of H3BO3, H3PO4, and CH3COOH, adjusted to selectedpH in the range of pH 3.0–9.0. One millimolar solutions of PD, PQ, and PMS were preparedin acetonitrile, Fer, FCA, MFM, and FCL in 95 % alcohol. Interaction of GOx (5·10−6 M)with mediators (10−5 M) was studied in 0.01 M phosphate buffer (pH 7.0) at molarenzyme/mediator ratio 1:2, and in order to perform the enzymatic reaction, excessiveconcentration (0.2 M) of glucose was added. In order to perform reliable control, it wasnecessary to study enzyme fluorescence at the same ethanol and acetonitrile concentrationwithout mediators. Final concentrations of acetonitrile and alcohol in analyzed enzymesolution were 1 %.
Protein Determination
Protein content was measured spectrophotometrically using spectrophotometer Ultraspec2100 (Amersham Pharmacia Biotech, Sweden) and it was estimated by the formula:[Protein]=183.0×A230–75.8×·A260 [48].
Results and Discussion
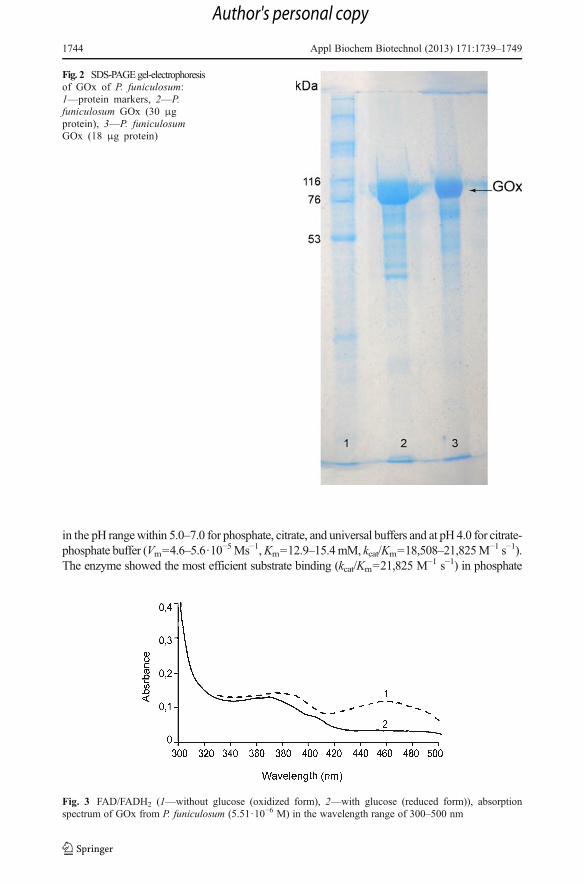
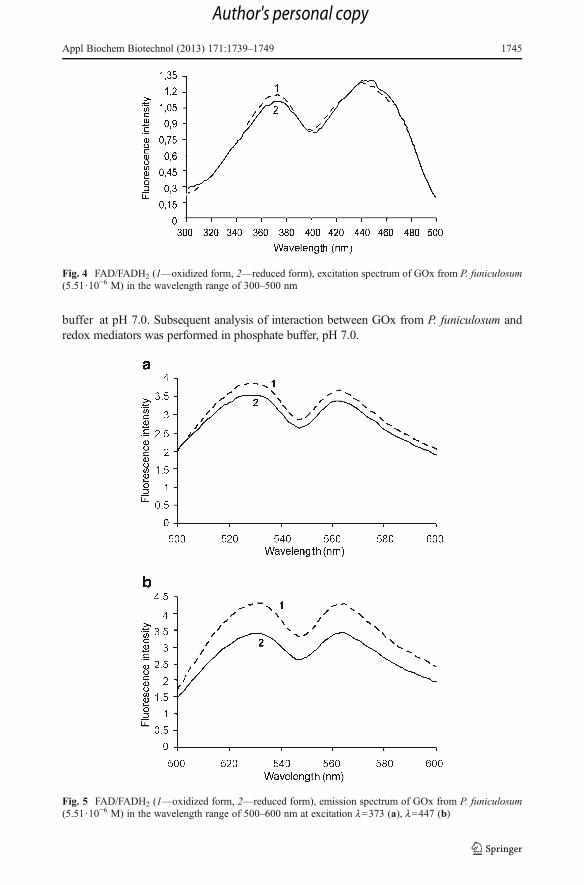
The harvesting of extracellular GOx was carried out by sequential ultrafiltration. As aresult, concentrated GOx solution of 2,900 U total activity and specific activity of85.4 U/mg protein was obtained. Concentration of impurity proteins in enzyme prepara-tion was not significant (Fig. 2). According to FAD extinction, calculated enzymeconcentration was 8·10−5 M. High specificity of this enzyme to glucose was shown inprevious research [39]. Both spectrophotometric and spectrofluorometric analysis haveshown that spectra of GOx, which was produced by P. funiculosum, are typical forflavoproteins. Absorption spectrum has distinct peaks at 380 and 457 nm (Fig. 3),excitation spectrum at 373 and 447 nm (Fig. 4), and emission spectrum at 530 and562 nm (Fig. 5). It should be noted that glucose addition to GOx solution changesabsorption spectrum: the peak at 457 nm decreases because FAD is converted to FADH2
(Fig. 3), glucose addition does not affect excitation spectrum, while intensity of emissionspectrum declines (Figs. 4 and 5).
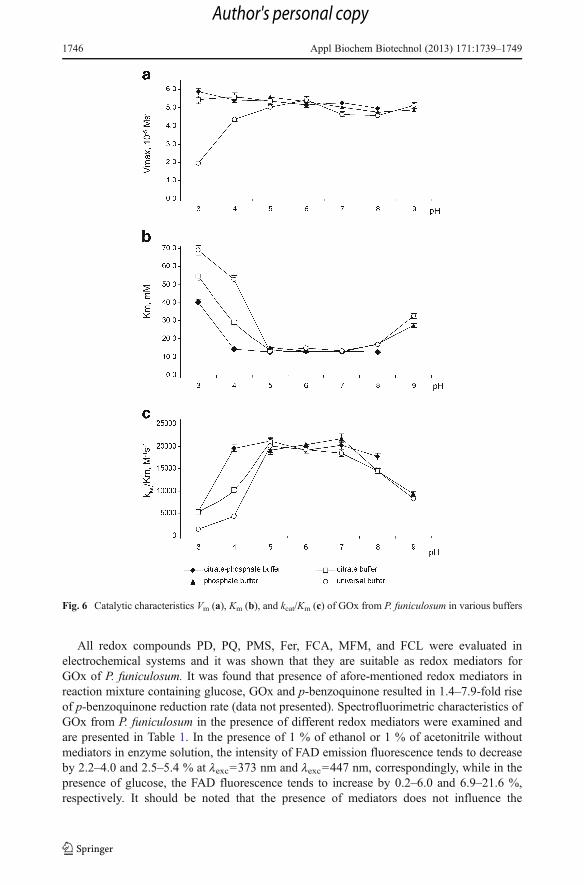
An essential stage during the development of biocatalytic system is the choice of optimalbuffer system where the enzyme displays its maximal catalytic activity. Investigations wereconducted in the following solutions: phosphate buffer (pH 5.0–9.0), citrate buffer (pH 3.0–5.0), citrate-phosphate buffer (pH 3.0–9.0), and universal buffer (pH 3.0–9.0). Enzymecatalytic activity was assessed by the estimation of: (a) maximal rate of 1,4-benzoquinonereduction to hydroquinone (Vm), (b) Michaelis constant (Km), which is an important indica-tor of substrate affinity, and (c) the efficiency of glucose oxidation reaction (kcat/Km).
It was found that for GOx from P. funiculosum the highest Km values, which were in therange of 27.4–69.0 mM, were observed at pH 3.0, pH 4.0 and at pH 9.0 (Fig. 6). High Km
values are indicating much lower affinity of GOx to glucose at these pH values, with lowestactivity at pH 3.0. Optimal values of catalytic parameters of P. funiculosumGOxwere observed
Appl Biochem Biotechnol (2013) 171:1739–1749 1743
Author's personal copy
in the pH rangewithin 5.0–7.0 for phosphate, citrate, and universal buffers and at pH 4.0 for citrate-phosphate buffer (Vm=4.6–5.6·10
−5Ms−1,Km=12.9–15.4mM, kcat/Km=18,508–21,825M−1 s−1).
The enzyme showed the most efficient substrate binding (kcat/Km=21,825 M−1 s−1) in phosphate
Fig. 3 FAD/FADH2 (1—without glucose (oxidized form), 2—with glucose (reduced form)), absorptionspectrum of GOx from P. funiculosum (5.51·10−6 M) in the wavelength range of 300–500 nm
Fig. 2 SDS-PAGEgel-electrophoresisof GOx of P. funiculosum:1—protein markers, 2—P.funiculosum GOx (30 μgprotein), 3—P. funiculosumGOx (18 μg protein)
1744 Appl Biochem Biotechnol (2013) 171:1739–1749
Author's personal copy
buffer at pH 7.0. Subsequent analysis of interaction between GOx from P. funiculosum andredox mediators was performed in phosphate buffer, pH 7.0.
Fig. 4 FAD/FADH2 (1—oxidized form, 2—reduced form), excitation spectrum of GOx from P. funiculosum(5.51·10−6 M) in the wavelength range of 300–500 nm
Fig. 5 FAD/FADH2 (1—oxidized form, 2—reduced form), emission spectrum of GOx from P. funiculosum(5.51·10−6 M) in the wavelength range of 500–600 nm at excitation l=373 (a), l=447 (b)
Appl Biochem Biotechnol (2013) 171:1739–1749 1745
Author's personal copy
All redox compounds PD, PQ, PMS, Fer, FCA, MFM, and FCL were evaluated inelectrochemical systems and it was shown that they are suitable as redox mediators forGOx of P. funiculosum. It was found that presence of afore-mentioned redox mediators inreaction mixture containing glucose, GOx and p-benzoquinone resulted in 1.4–7.9-fold riseof p-benzoquinone reduction rate (data not presented). Spectrofluorimetric characteristics ofGOx from P. funiculosum in the presence of different redox mediators were examined andare presented in Table 1. In the presence of 1 % of ethanol or 1 % of acetonitrile withoutmediators in enzyme solution, the intensity of FAD emission fluorescence tends to decreaseby 2.2–4.0 and 2.5–5.4 % at lexc=373 nm and lexc=447 nm, correspondingly, while in thepresence of glucose, the FAD fluorescence tends to increase by 0.2–6.0 and 6.9–21.6 %,respectively. It should be noted that the presence of mediators does not influence the
Fig. 6 Catalytic characteristics Vm (a), Km (b), and kcat/Km (c) of GOx from P. funiculosum in various buffers
1746 Appl Biochem Biotechnol (2013) 171:1739–1749
Author's personal copy
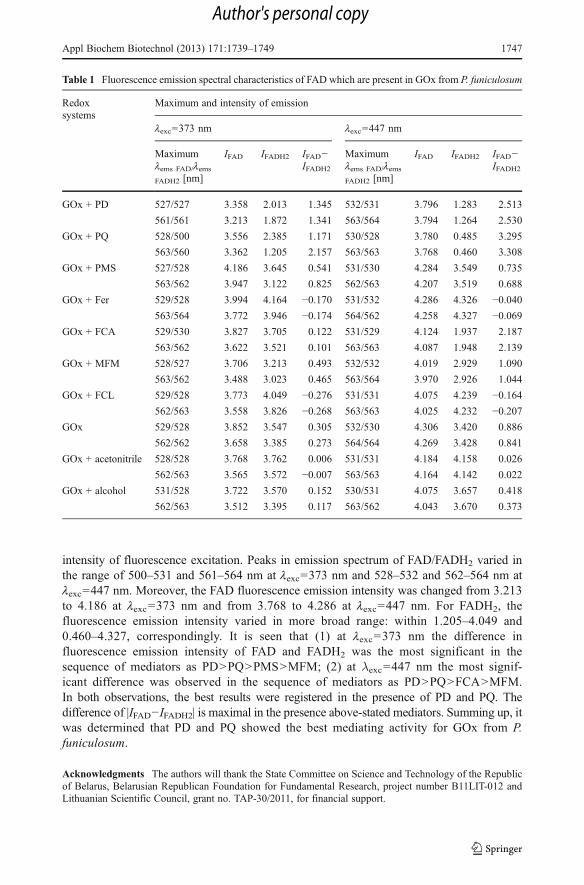
intensity of fluorescence excitation. Peaks in emission spectrum of FAD/FADH2 varied inthe range of 500–531 and 561–564 nm at lexc=373 nm and 528–532 and 562–564 nm atlexc=447 nm. Moreover, the FAD fluorescence emission intensity was changed from 3.213to 4.186 at lexc=373 nm and from 3.768 to 4.286 at lexc=447 nm. For FADH2, thefluorescence emission intensity varied in more broad range: within 1.205–4.049 and0.460–4.327, correspondingly. It is seen that (1) at lexc=373 nm the difference influorescence emission intensity of FAD and FADH2 was the most significant in thesequence of mediators as PD>PQ>PMS>MFM; (2) at λexc=447 nm the most signif-icant difference was observed in the sequence of mediators as PD>PQ>FCA>MFM.In both observations, the best results were registered in the presence of PD and PQ. Thedifference of |IFAD−IFADH2| is maximal in the presence above-stated mediators. Summing up, itwas determined that PD and PQ showed the best mediating activity for GOx from P.funiculosum.
Acknowledgments The authors will thank the State Committee on Science and Technology of the Republicof Belarus, Belarusian Republican Foundation for Fundamental Research, project number B11LIT-012 andLithuanian Scientific Council, grant no. TAP-30/2011, for financial support.
Table 1 Fluorescence emission spectral characteristics of FAD which are present in GOx from P. funiculosum
Redoxsystems
Maximum and intensity of emission
lexc=373 nm lexc=447 nm
Maximumlems FAD/lems
FADH2 [nm]
IFAD IFADH2 IFAD−IFADH2
Maximumlems FAD/lems
FADH2 [nm]
IFAD IFADH2 IFAD−IFADH2
GOx + PD 527/527 3.358 2.013 1.345 532/531 3.796 1.283 2.513
561/561 3.213 1.872 1.341 563/564 3.794 1.264 2.530
GOx + PQ 528/500 3.556 2.385 1.171 530/528 3.780 0.485 3.295
563/560 3.362 1.205 2.157 563/563 3.768 0.460 3.308
GOx + PMS 527/528 4.186 3.645 0.541 531/530 4.284 3.549 0.735
563/562 3.947 3.122 0.825 562/563 4.207 3.519 0.688
GOx + Fer 529/528 3.994 4.164 −0.170 531/532 4.286 4.326 −0.040563/564 3.772 3.946 −0.174 564/562 4.258 4.327 −0.069
GOx + FCA 529/530 3.827 3.705 0.122 531/529 4.124 1.937 2.187
563/562 3.622 3.521 0.101 563/563 4.087 1.948 2.139
GOx + MFM 528/527 3.706 3.213 0.493 532/532 4.019 2.929 1.090
563/562 3.488 3.023 0.465 563/564 3.970 2.926 1.044
GOx + FCL 529/528 3.773 4.049 −0.276 531/531 4.075 4.239 −0.164562/563 3.558 3.826 −0.268 563/563 4.025 4.232 −0.207
GOx 529/528 3.852 3.547 0.305 532/530 4.306 3.420 0.886
562/562 3.658 3.385 0.273 564/564 4.269 3.428 0.841
GOx + acetonitrile 528/528 3.768 3.762 0.006 531/531 4.184 4.158 0.026
562/563 3.565 3.572 −0.007 563/563 4.164 4.142 0.022
GOx + alcohol 531/528 3.722 3.570 0.152 530/531 4.075 3.657 0.418
562/563 3.512 3.395 0.117 563/562 4.043 3.670 0.373
Appl Biochem Biotechnol (2013) 171:1739–1749 1747
Author's personal copy

References
1. Roth, J. P., & Klinman, J. P. (2003). Proceedings of the National Academy of Science USA, 100, 62–67.2. Katz, E., Shipway, A. N., & Willner, I. (2002). In G. S. Wilson (Ed.), Encyclopedia of electrochemistry.
vol. 9 (pp. 559–626). Weinheim: Wiley.3. Leskovac, V., Trivic, S., Wohlfahrt, G., Kandrac, J., & Pericin, D. (2005). The International Journal of
Biochemistry & Cell Biology, 37, 731–750.4. Löffler, U., Wiemhöfer, H.-D., & Göpel, W. (1991). Biosensors and Bioelectronics, 6, 343–352.5. Savitri, D., & Mitra, C. K. (1999). Journal of Biosciences, 24, 43–48.6. Reshetilov, A. N. (1996). Applied Biochemistry and Microbiology, 32, 72–86.7. Wilkins, E., & Atanasov, P. (1996). Medical Engineering & Physics, 18, 273–288.8. Wilson, R., & Turner, A. P. F. (1992). Biosensors and Bioelectronics, 7, 165–187.9. Wong, C. M., Wong, K. H., & Chen, X. D. (2008). Applied Microbiology and Biotechnology, 78, 927–
938.10. Ramanavicius, A., & Ramanaviciene, A. (2009). Fuel cells, 9, 25–36.11. Ramanavicius, A., Kausaite, A., & Ramanaviciene, A. (2008). Biosensors and Bioelectronics, 24, 761–
766.12. Ramanavicius, A., Kausaite, A., & Ramanaviciene, A. (2005). Biosensors and Bioelectronics, 20, 1962–
1967.13. Davis, F., Higson, S., & S’eamus, P. J. (2007). Biosensors and Bioelectronics, 22, 1224–1235.14. Ivnitski, D., Branch, B., Atanassov, P., & Apblett, C. (2006). Electrochemistry Communications, 8, 1204–
1210.15. Kannan, M., Renugopalakrishnan, V., Filipek, S., Li, P., Audette, G. F., & Munukutla, L. (2008). Journal
of Nanoscience Nanotechnol, 8, 1–13.16. Neto, S. A., Forti, J. C., & Andrade, A. R. D. (2010). Electrocatal, 1, 87–94.17. Oztekin, Y., Yazicigil, Z., Solak, A. O., Ustundag, Z., Okumus, A., Kilic, Z., Ramanaviciene, A., &
Ramanavicius, A. (2012). Sensors and Actuators B-Cemical, 166–167, 117–127.18. German, N., Ramanavicius, A., Voronovic, J., Oztekin, Y., & Ramanaviciene, A. (2011). Microchimica
Acta, 172, 185–191.19. Bean, R. C., & Hassid, W. Z. (1956). Journal of Biological Chemistry, 218, 425–436.20. Bi, J., Felton, G. W., Murphy, J. B., Mathews, M. C. Patent US6303326 (2001). pp. 27.21. Cavener, D. R. (1980). Biochemical Genetics, 18, 929–937.22. Crueger, A., & Crueger, W. (1990). In W. M. Fogarty & C. T. Kelly (Eds.), Microbial enzymes and
biotechnology (pp. 177–226). London: Elsevier.23. Kupletskaya, M. B., & Kurakov, A. V. (1998). Applied Biochemistry and Microbiology, 35, 140–144.24. Mikhailova, R. V., Lobanok, A. G., Shishko, Z. F., & Yasenko, M. I. (1998). Mycological
Phytopathology, 1, 60–64.25. Ohashi, K., Natori, S., & Takeo, K. (1999). European Journal of Biochemistry, 265, 127–133.26. Smotrova, N. G., & Kremenchuzki, G. N. (2002). Mikrobiologichny zhurnal, 64, 28–34 (in Ukrainian).27. Bachmatova, I., Kurtinaitiene, B., & Marcinkeviciene, L. (1992). Biotechnol. in Estonia. Latvia and
Lithuania, 11, 47.28. Eriksson, K. O., Kourteva, I., Yao, K. Q., Liao, J. L., Kilár, F., Hjertén, S., & Chaga, G. (1987). Journal of
Chromatography, 397, 239–249.29. Garzillo, A. M., Paolo, S. D., Petruccioli, M., Fenice, M., Buonocore, V., & Federici, F. (1995).
Biotechnol. Applied Biochemistry, 22, 169–178.30. Hatzinikolaou, D. G., Hansen, O. C., Macris, B. J., Tingey, A., Kekos, D., Goodenough, P., & Stougaard,
P. (1996). Applied Microbiology and Biotechnology, 46, 371–381.31. Hayashi, S., & Nakamura, S. (1976). Biochimica et Biophysica Acta, 438, 37–48.32. Kalisz, H. M., Hecht, H. J., Schomburg, D., & Schmid, R. D. (1991). Biochimica et Biophysica Acta,
1080, 138–142.33. Kalisz, H. M., Hendle, J., & Schmid, R. D. (1997). Appl. Microbiology and Biotechnology, 47, 502–507.34. Rando, D., Kohring, G. W., & Giffhorn, F. (1997). Applied Microbiology and Biotechnology, 48, 34–40.35. Takegawa, K., Fujiwara, K., Iwahara, S., Yamamoto, K., & Tochikura, T. (1989). Biochemistry and Cell
Biology, 67, 460–464.36. Sukhacheva, M. V., Davydova, M. E., & Netrusov, A. I. (2004). Applied Biochemistry and Microbiology,
40, 25–30.37. Davydova, M. E., Kurova, V. S., Sukhacheva, M. V., Kupletskaya, M. B., Ryabov, A. D., & Netrusov, A.
I. (2002). MSU Vestnik. Series 2. Chemistry, 43, 366–370 (in Russian).38. Mikhailova, R. V., Semashko, T. V., & Lobanok, A. G. (2002). Applied Biochemistry and Microbiology,
38, 236–239.
1748 Appl Biochem Biotechnol (2013) 171:1739–1749
Author's personal copy

39. Semashko, T. V., Mikhailova, R. V., & Eremin, A. N. (2003). Applied Biochemistry and Microbiology, 39,368–374.
40. Eryomin, A. N., Semashko, T. V., & Mikhailova, R. V. (2006). Applied Biochemistry and Microbiology,42, 138–144.
41. Semashko, T. V., Mikhailova, R. V., & Lobanok, A. G. (2004). Microbiology, 73, 286–292.42. Semashko T. V., Mikhailova, R. V., Demeshko, O. D. (2011). Abstracts in Scientific Conference
“BIOLOGICALLY ACTIVE SUBSTANCES: fundamental and applied problems” Novy Svet. ARCrimea. Ukraine. 312.
43. Mikhailova, R. V., Semashko, T. V., Zhukovskaya, L. A., & Lobanok, A. G. (2010). Proceedings of theNational Academy of Sciences of Belarus. Series of Biological sciences, 1, 58–61 (in Belarusian).
44. Mikhailova, R. V., Semashko, T. V., Lobanok, A. G. Patent of the Republic of Belarus no. 14007 (2011)Official bulletin of National Centre of Intellectual Property. 1. 108.
45. Laemmli, U. K. (1970). Nature, 227, 680–685.46. Ciucu, A., & Patroescu, C. (1984). Analytical Letter, 17, 1417–1427.47. Markwell, J., Frakes, L. G., Brott, E. C., Osterman, J., & Wagner, F. W. (1989). Applied Microbiology and
Biotechnology, 30, 166–169.48. Poloznikov, A. A., Savitsky, P. A., Khushpulyan, D. M., Chubar, T. A., Gazaryan, I. G., & Tishkov, V. I.
(2006). MSU Vestnik. Series 2. Chemistry, 47(1), 20–24 (in Russian).
Appl Biochem Biotechnol (2013) 171:1739–1749 1749
Author's personal copy
Copyright © 2022 FDOKUMEN




















