
Phenotype of atopic dermatitis subjects with a history of eczema herpeticum
Upload
independentCategory
view
0download
0
Clinical and E.xperimeiita} Allergy. 1997. Volume 27. pages 808-815
Specific induction of interleukin-4-producing cells in responseto in vitro allergen stimulation in atopie individuals
S. GABRIELSSON. S. PAULIE. S. RAK*, E. LAGGINGt.M. VAN HAGE-HAMSTENt, B. HAREASTt and M. TROYE-BLOMBERG
Department of Immunology. Stockholm University, ^Allergy Unit. Sahlgrenska Ho.spital and ^Clinical Immunology,Karolin.ska Ho.spital, Stockholm, Sweden
Summary
Background and Objective CD4''' T cells can be divided into two major subsets. T helper(TH)| andTHi cells. Interleukin-4 (IL-4) is produced by TH2 cells and induces switching ofimmunoglobulin (Ig) M/lgG to IgE. Interferon-7 (IFN7) produced by TH, cells counteractsthe IgE-pronioling effects of IL-4. In this study we wanted to investigate whether thenumber of IL 4-producing cells could be a direct measurement of allergen exposure in vitro,and whether this was correlated to the elevated serum IgE-levels seen in atopie persons.Methods We compared the number of IL-4- and IFN7-producing cells using an enzyme-linked immunospot assay (ELISPOT) in response to allergens from birch and cat inperipheral mononuclear cells from atopie and hcaltby individuals.
Results In the two sen.sitized groups there was an increase in the number of lL-4 producingceils in response to the specific allergen which was not seen in the healthy group (1/20000ceils and 1/200000 cells, respectively, P < 0.001 for birch). In criss-cross experimentswhere birch-sensitized individuals were stimulated with cat allergen, no lL-4-producingcells were seen, indicating a high degree of specificity. In individual subjects, the elevatednumbers of IL-4-producing cells were significantly correlated with their allergen-specificserum IgE levels. Wben allergen was combined with a suboptimal dose of PHA, there was asynergistic increa.se in the number of allergen-induced IL-4-producing cells (i/4000 cells)in the atopie donors, which was not seen with the number of IFN7-producing cells.Conclusions Allergen-specific IL-4 producing cells in a peripheral blood mononuclearcell (PBMC) culture can be detected by ELISPOT and the response can synergistically beenhanced by suboptimal concentrations of PHA.
Keywords: allergen-specific, atopy, ELISPOT., iFN7. IL-4
Clinical and Experimental Allergy, Vol 27. pp. 808-815. Submitted 22 September 1996;revised 15 March 1996; accepted 25 August 1996.
Introduction
The acute cause of most symptoms in Type 1 allergies is theIgE-induced release of histamine. prostaglandins and othermediators by mast cells. In allergic individuals, there is anelevated level of IgE in serum [1]. The cytokine IL-4 hasbeen shown to initiate switching to IgE in resting [2.31 andLPS-activated B-cells [4.51 ^nd is al.so needed for main-taining IgE-production [6]. lL-13 is another cytokine that,although less potently, promotes IgE-production 17].
Correspondence: S. Gabrielsson. Departmeni of Immunology. StockholmUniversity. S-106 91 Stockholm. Sweden.
IL-4 is produced by a certain subset of 004"*^ T-helper(TH) cells. Mosmann and Coffman first propo.sed the divi-sion of murine TH cells into TH| and TH2 cells based on thecytokines that they secrete upon antigen stimulation [8].TH, cells produce IFN7, TNFa, TNE/3 and IL-2. while TH.cells produce IL-4, IL-5 and IL-6. There is also an inter-mediate or non-differentiated TH cell called THQ. producinglL-2 and IFN7, as well as IL-4 |9| . Although not as clear-cut, recent data suggest that human CD4''̂ T cells also areheterogenous with regard to cytokine production [101. TH,cell clones provide help for cell-mediated responses andplay an important role in delayed type hypersensitivity.while THi cells are most potent in delivering stimuli to B
© 1997 Blackwell Science Ltd

IL-4-producing cells in response to allergen 809
cells, leading to antibody production [111. Tbe cytokinesof THi and THT cells reciprocally counteract each other,e.g. IL-4 inhibits IFN7 production and IL-4-induced IgE-synthesis is blocked by addition of IFN7 II2|. In murineexperimental infections there is now good evidence that thebalance between the two response patterns determines thetype and efficiency ofthe Jmtnune response.
In this study we wanted to investigate whether theelevated levels of allergen-specific IgE in serum seen inallergic patients were reflected by an increased number ofIL-4-producing cells after activation with allergen. Topermit a minimum of ('/( vitro manipulation we used thehighly sensitive enzyme linked iminunospot (ELISPOT)assay which allows analysis at the single-cell level of invitro activated bulk cultured mononuclear cells.
Materials and methods
(a)
32
Reagents
The monoclonal antibodies used in tbe ELISPOT were:MoAb 1-DIK and 7B6-1 to human IFN7 and MoAb 82-4(IL-4-i) and 12-1 (IL 4 2) to human IL-4 (all MoAb wereobtained from MABTECH, Stockholm, Sweden). The spe-cific allergens used were Aquagen-SQ birch and cat. 1 x 10**SQ-U/tiiL (ALK, Denmark). The mitogen was PHA (MurexDiagnostics Ltd, Dartford, UK).
Subjects
The sensitized groups comprised asthma patients positive inradio-allergosorbent test (RAST. > 4kU/L or > class 2) forbirch (« = 17. aged 27-60, mean 40.1) and/or cat (n = 12,aged 22-34. mean 29.1). All birch patients were studied > 3months after the pollen season. The controls were 17 age-matched volunteers negative in RAST for birch and cat.
Preparation and stimulation of PMNC
Motionuclear cells from heparinized peripheral blood wereisolated by Ficoll-Paque (Pharmacia. Stockholm, Sweden)gradient centrifugation. The cells were then diluted to 2million cells/mL in tissue-culture media (TCM) consistingof RPMI-HEPES with Gentamicin (25^g/mL), 2tnM L-glutamine and 10% heat-inactivated fetal calf serum. Thecells were processed within 24 h of bleeding, or frozen andused later. Eor freezing, PBMC were suspended (10^ to 10̂cells/mL) in TCM containing 10% Dimethyl sulphoxide(DMSO) and frozen gradually TC/min to -70°C in afreezing container (Nalgene Cryo I°C, Nalge Company.USA). Thereafter, tubes were stored in liqtiid nitrogen.Before use. the cells were thawn quickly in a 37°C waterbathand diluted and wa.shed in TCM. Half or I mL cell suspension
20
B 10
(bt
2 8 32
Allergen concentration, xiO^ SQ-U/mL
Fig. 1. The number of IL 4 producing cells in (a) 11 birch sensitiveand (b) nine healthy individuals in response to birch allergen.Peripheral mononuclear cells were cultured for 48 h in mediumcontaining different concentrations of birch allergen. Resultsrepresent the mean values of duplicate determinations.
was transferred to 5 mL-culture tubes (Falcon, Becton-Dick-inson. New Jersey. USA) and allergens and/or PHA wereadded at different concentrations. The tubes were incubatedat 37°C in a humid atmosphere containing 5% CO2 for 4h.After the incubation, the cells were resuspended, and l(X)/iLof the cell suspension were added to each coated ELISPOT-well (200000 cells/well) in duplicates and transferred toELISPOT-plates. In the case of IFN7. cells were first dilutedten times (20000 cells/well).
1997 Blatkwell Science Lid. Clinical and E.\'i>erimental Allergy. 27.

810 S. Gabrielsson et al.
Table 1. The number of IL-4-producing cells/200 0(X) cells from in vitro activated PBMC measured byELISPOT*
StimuliBirch sensitive (17)
Mean ± SDBirch negative (9)
Mean ± SDr-test, unpaired
Pi-test. paired
P
w/oAgPHAPHA + Ag
1.3 ±2.37.9 ±5.6
101.2 ±58,4149.5 ±98.9
1.10.9
46.539.3
±0.8± 1.0±30.2±26.9
NSi0.0010.0200.034
0.0035
0.0013
* PBMC were cultured for 48 h in medium or medium containing 0,25 ^g/mL PHA and/or 32 000 SQ-U birchallergen. Results from each individual represent the mean values of duplicate determinations.t not signiticanl.
ELISPOT
The reverse ELISPOT assay is based upon tbe ELISA-plaque assay for detection of antibodies [13J and wasperformed mainly as described elsewhere 114.16]. Briefly,the anti-human-IFN7 monoclonal antibody 1-DIK or anti-human IL-4 monoclonal antibody 82-4 were diluted to aconcentration of 15/ig/niL in sterile, filtered (0.45 ^m)phosphate buffered saline (PBS), and lOO^L per wellwere added onto nitrocellulose plates (Millititer, MilliporeCorp., Bedford. Massachusetts. USA). Plates were incu-bated overnight at 4°C and thereafter unbound antibodies
s
32
Concentration of allergen, xiO' SQ-U/mL
Fig. 2. Test of crossreactivity to cal allergen in six birch-sensitiveindividuals. • . birch (specific allergen). O. cat (unrelatedallergen). Peripheral mononuclear cells were cultured for 48 h inmedium containing different concentrations of birch or catallergen. Results represent the mean values of duplicatedeterminations.
were washed away by six successive washings with filteredPBS: 100/xLof cell suspension were added to each well andthe plates were incubated 40-42 h at 37°C whereafter thecells were washed away and 100/xL of the biotinylatedIFN7 MoAb 7B6-1 or IL-4 MoAb 12-1 (1/xg/mL) wereadded and incubated for 2h at room temperature (RT).Thereafter, the plates were washed and incubated for90 min at RT with 100 fiL of streptavidin-alkaline-phosphata.se (MABTECH). Stockholm. Sweden) diluted 1/1000. Unbound conjugate was removed by another series ofwashings and finally 100 ^L of BCIP/NBT substrate solu-tion (Bio-Rad, Ricbmond, USA) were added and incubatedat RT until dark spots emerged (l/2h for IFN7 and 1 h forIL-4). The colour development was stopped by three wash-ings with 200/xL of tap-water/well. After drying, the spotswere counted in a dissection microscope (x40).
Statistical analysis
The statistical analyses perfonned were paired student's t-test between w/o and allergen stimulated cultures andunpaired student's /-test between the sensitive and healthygroup. Linear regression analysis on logarithmic values wasalso perfonned.
Results
Estimation of the number of IL-4'producing cells in birchatopic persons and in controls
The ELISPOT assay was used to determine the number ofIL-4 producing cells induced in response to specific aller-gen. For this purpose mononuclear cells from birch allergicpatients or healthy non-allergic individuals were stimulatedwith three different doses of birch allergen. Sera from allallergic patients were classified as being RAST class 3 ormore. As can be seen in Fig. la, there was a response to
1997 Blackwcll Science Lid, Clinical and Experimental Allergy. 27.

IL-4-producing cells in response to allergen 81
200 r
100 •
32
Concentration of allergen, xiO' SO-U/mL
Fig. 3. IL-4-producing cells in two representative birch sensitiveindividuals out of 11. A suboptimal dose of PHA in combinationwith specific allergen induces a synergistic effect on the number ofIL-4-producing cells. The number of spots produced withoutallergen stimuli has been subtracted. Peripheral mononuclearcells were cultured for 48 h in medium containing PHA (0.25 jug/mL) and different concentrations of birch allergen. The number ofPHA induced spots was for donor 1 and 2. 65 and 170 spot.s/20(10(K) cells respectively. Results repre.sent the mean values ofduplicate determinations. • . Patieni 1 w. PHA D. Patient 1 w/oPHA. • . Patient 2 w. 2PHA, O. Patieni I w/o PHA.
birch allergen in all except one birch allergic patient, withthe highest dose (32000 SQ-Units/mL) inducing the highestnumber of IL-4-producing cells. Therefore, the last six indi-viduals tested were only stimulated with the highest allergendose. The response was significantly higher than to mediumonly (P — 0.0035, paired /-test). This response was specific
and not seen in the non-allergic group, where no or very fewIL-4-pr(Kiucing cells were seen (Fig. Ib). The difference ofspots in response to allergen between the two groups wasstatistically highly .significant iP < O.OOL Table 1).
Specificity was further shown by stimulating cells fromsix birch allergic patients with birch allergen and for thesepatients, an unrelated cat allergen. As can be seen in Fig. 2. aresponse was only seen to the specific allergen {P = 0.0017,paired /-test for the highest allergen dose),
Suboptimal polyclonal activation synergistically increa.sesthe frequency of allergen-specific lL-4-producing cells inatopie patients
In this study and in agreement with previously reported [ 17],polyclonal activation by PHA induced more IL 4 producingcells in the atopie group than in the group of healthyindividuals. In individual donors the dose of PHA requiredto induce optimal number of IL 4-producing cells variedbetween I and lO^g/mL (not shown). Also when stimulat-ing with lower concentrations (0.25 ̂ g/mL). the atopiesresponded with more IL-4-producing cells (Table 1).
Although there was an increase in the sensitized patientsin IL-4-producing cells after stimulation with the allergen,the total number of lL-4-producing cells was generally low.However, when combining suboptimal doses of PHA(0.25 fig/mL) with the allergen, there was a synergistieinduction of IL-4-producing cells in the birch sensitiveindividuals (Fig. 3, Table I). In birch sensitive, the differ-ence between PHA -I- allergen and PHA alone was 48 IL-4-spots/2000()() cells. The difference between allergen andmedium only (without the PHA synergy) was 7 lL-4-spots/200000 cells. No increase was seen in healthy individuals.
Cat atopies
The response to the cat allergen in cat allergic patients was
Table 2. The number of IL-4-producing cells/200000 cells from in vitro activated PBMC measured byELiSPOT*
Stimuli
w/o
AgPHAPHA + Ag
Cat atopies (12)Mean
0.7 ^10.0^98.8 ^
133.0 i
± SD
:0.7: 12.1: 100.9: 147.1
Cat negative (8)Mean ± SD
0.5 ± 0.62.0+ 1.8
47.9 ± 28.656.4 ± 30.1
f-test, unpairedP
NStNSNSNS
r-test. pairedF
0.016
0.017
*PBMC were cultured for 48 h in medium or medium containing 0.25/ig/mL PHA and/or 32 000 SQ-U catallergen. Results from each individual represent the mean values of duplicate determinations.t not significant.
IW7 Blackwell Science Ltd. Clinical and Experimental Allersy. 27. 808-815
812 S. Gabrielsson et al,
o
s
32
30
20
sooo
10
(b)
32
Allergen concentration. SQ-U/mL
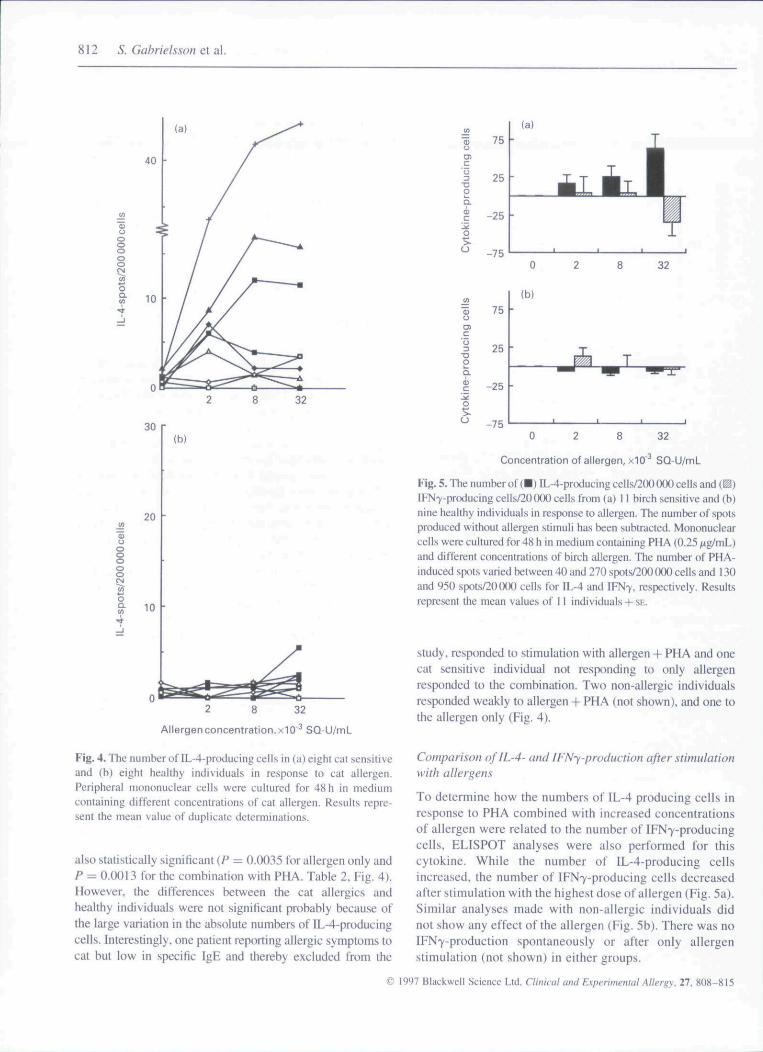
Fig. 4. The number of IL-4-producing cells in (a) eight cat sensitiveand (b) eight healthy individuals in response to cat allergen.Peripheral mononuclear cells were cultured for 48 h in mediumcontaining different concentrations of cat allergen. Results repre-sent the mean value of duplicate determinations.
also statistically significant (F — 0.0035 for allergen only andP = 0.0013 for the combination witb PHA. Table 2, Fig. 4).However, the differences between the cat allergies andhealthy individuals were not significant probably because ofthe large variation in the absolute numbers of IL-4-producingcells, interestingly, one patient reporting allergic symptoms tocat but low in specific IgE and thereby excluded from the
•5 75
25
-25
CJ
-75
75
25
-25
-75
32
(b)
32
Concentration of allergen, x lO SQ-U/mL
Fig, 5. The numher of (•) IL-4-producing cells/2(M) (XX) cells and (^)[FN7-producing cells/20(X)0 ceils from (a) 11 birch sensitive and (b)nine healthy individuals in response to allergen. The number of spotsproduced without allergen stimuli has been subtracted. Mononuclearcells were cultured for 48 h in medium containing PHA (0.25 ;xg/mL)and different concentrations of birch aliergen. The numher of PHA-induced spots varied between 40 and 270 spots/200 (XX) cells and 130and 950 spots/20000 cells for IL-4 and IFN7, respectively. Resultsrepresent the mean values of II individuals + SE.
study, responded to stimulation with allergen-I- PHA and oneeat sensitive individual not responding to only allergenresponded to the combination. Two non-allergic individualsresponded weakly to allergen -I- PHA (not shown), and one tothe allergen only (Fig. 4).
Comparison of IL-4- and IFNy-production after stimulationwith allergens
To determine bow the numbers of IL-4 producing cells inresponse to PHA combined with increased concentrationsof aliergen were related to the number of IFN7-producingcells, ELISPOT analyses were also performed for tbiscytokine. While the number of IL-4-producing cellsincreased, the number of IFN7-producing cells decreasedafter stimulation with the highest dose of allergen (Fig. 5a).Similar analyses made with non-allergic individuals didnot show any effect of the allergen (Fig. 5b). There was noIFN7-production spontaneously or after only allergenstimulation (not shown) in either groups.
1997 Blackwell Science Ltd. Clinical and Experimental Allergy. 11. 808-815

IL-4-producing cells in response to allergen 813
1.2
log(iL-4 spots/200 000 cells)
Fig. 6. Linear regression analysis between the logarithm of specificserum IgE (measured by RAST) and the logarithm ofthe numberof IL-4-producing cells in response to specific allergen <p = 0.80.P = 0.001) in 13 birch- or cat-allergic individuals. Peripheralmononuclear cells were cultured for 48 h in medium containingPHA only or PHA together with birch or cal allergen. The numberof spots produced without allergen stimuli ha.s been subtracted.PHA alone gave 40-270 lL-4-spots/200 000 cells. Results representihe mean value of duplicate determinations.
Specific IL-4 response vs specific IgE
To investigate if the IL-4-response wus correlated to aller-gen-specific serum IgE-levels, the tiumbers of allergen-specific IL-4-producing cells were cotnpared with the itidi-viduai allergen-specific IgE levels (Fig. 6). Linear regres-sion analysis revealed a statistically significant relationshipbetween the two tneasures (p = 0.80. P = 0.001). This wasin contrast to what was seen between IL-4-response to PHAand total IgE in serum (not shown).
Discussiun
Resting T cells are poor producers of IL-4, but whenstimulated in vitro with mitogens or anti-CD3 antibodiesin the presence of IL-2 and IL-4, they can develop into IL-4-producing cells. Still, very low frequencies of IL-4producing cells are seen after stimulation compared toseveral other cytokines. e.g. IL-2 [ 18|. Here, we bave usedthe ELISPOT-method, since this method has been shownto be more sensitive tban conventional ELISAs 119] andthe frequency of cytokine producing cells can be mea-sured. In ELISPOT the cytokine is measured close to tbecell where it is produced and not in the supernatant, whereit will be diluted or ean bind to cell-bound or solublereceptors. As few as one eytokine producing eell in200000 cells ean be detected by this assay. In ELISPOTthe true frequency of cytokine-producing cells might be
underestimated if more IL-4 is produced later than 44 h orif tbe MoAbs or cytokine molecules dissociate during tbetime ofthe incubation. An alternative sensitive method todetect cyokine producing cells is RT-PCR. However,since tbe IL-4 production is transient and RNA isdegraded faster than protein, the time-points are morecritical [20].
Type 1 allergic responses are THi-dependent [10,21].Tbus, cells from atopic patients produce more IL-4 afterpolyclonal stimulation tban healthy people [17.22]. Tbe IL-4 production is also higher in polysensitized patients com-pared with monosensitized patients [17]. In agreement withthis, in the present study many more IL-4-produeing cells inresponse to PHA were found in the atopic group comparedto tbe healthy donor group.
Allergen-specific IL-4 can be induced in atopic indivi-duals. However, in these instances even smaller amounts oflL-4 are produced than after polyclonal stimulation. Severalauthors have reported that in order to he able to detectantigen-specific IL-4-production, repeated antigen stimula-tion [23-25] or in vitro pulsing with polyclonal stimuli [26]are needed. Other reports on allergen-specific IL-4-produc-tion have been performed using specific T-cell clones[27,28]. Here, we demonstrate that by using the ELISPOTassay it is possible to detect IL-4-producing cells directlyafter a short incubation with specific allergen. Thus, whenPBMC obtained from bircb allergic patients were in vitroactivated with the allergen, tbe numbers of IL-4-producingcells significantly increased over non-stimulated cultures.As analyses were performed out of season for bireh pollenallergies these in vitro tests seem to refiect a secondaryresponse. In similar cultures obtained from healthy Indivi-duals, bircb allergen did not induce any IL-4-secretion.Moreover, in the atopic individuals tbe number of IL-4spots was significally correlated to tbeir specific serum IgElevels.
In patients witb atopic dermatitis, an inllammatory skindisease associated with high IgE-levels in serum, adecreased I EN 7-production as compared with tbat innormal individuals in response to non-specific stimulationhas been reported, suggesting a switcb to TH^ in the atopiepatients [29.30]. This was also tbe case in our study, wherean increased number of IL-4-producing cells and a reducednumber of IFNy-producing cells were found. Tbe fact thatthere was no reduction of IEN7-producing cells in tbehealthy individuals after allergen stimulation suggests thatthe TH I-cells are suppressed by the IL-4 induced in theatopic individuals, leading to a reduced number of IFN7-producing cells.
Wben PBMC were obtained from cat allergic patients notall of them responded to cat allergen. Since the number ofIL-4-producing cells in some cat sensitized individuals wasas bigh as for the birch sensitized group, tbe lack of response
6 1997 Blackweil Science Lid. Clinical and Experimental Allergy. 11. 808-815

814 S. Gabriels.wn et al.
is not likely due to fewer T-cell epitopes in the cat allergenpreparation. Rather the reason for the low response in someindividuals is probably due to that cat allergen remainsairbome for a long while, and thus the patients have beenexposed without actually being in contact with the animal.Tbis might lead to migration of tbe specific T cells into thetarget organ, and the cells remaining in the periphery wouldbe difficult to stimulate further. The migration of activatedcells from the periphery has been demonstrated in malaria[31]. The reason why two non-allergic individualsresponded to the cat allergen is unknown. The response isvery low and only to the highest dose of allergen, and couldbe due to a subcHnical allergy or the crude nature of theallergen.
When PHA and allergen were u.sed together, a synergistieincrease in the number of IL-4-producing cells was seen inthe atopic group. The reason for tbe synergy is most likelythat PHA stimulates T cells, hereby leading them to aggre-gate and to gain cell-cell contact, facilitating interactionbetween cell-surface molecules. Several cytokines will alsobe produced, e.g. IL-2. which is known to be a costimulatoryfactor necessary for IL-4-production. since anti-iL-2 com-pletely inhibits IL-4-production |32]. The frequency ofallergen stimulated IL-4-producing cells with the PHAprobably reflects better the in vivo situation in the lung,where the cells more easily reach contact with other celtsthan in a culture tube in vitro. Furthermore, additionalcytokines produced in vitro by the PHA could in vivo beproduced by other cells not present in the in vitro PBMCculture. The synergistie increase was not seen in the numberof IFN7-producing cells. Thus, tbis suggests an increasedactivation of TH2 cells, or alternatively of other cellsproducing a TH^-like cytokine pattern in atopic individuals.However, our data does not exclude the possibility ofexisting allergen specific TH[ or THo type of cells. Cellsproducing TH^-eytokines bave been shown to includeCD8^ T ceils [33], 76 T cells [34]. CD4 CD8-a,3 T cells[35] as well as non-B non-T cells [36], However, recent datasuggest that the IgE initiating factor most likely is CD4"'̂derived IL-4 [37]. The synergistie effect observed with PHAis not specific for TH2 cells since in psoriasis, a TH|-associated disease, the synergistie effect of PHA is seen asan increased number of IFN7-produeing cells |38|.
In conclusion, the results demonstrate an increased numberof allergen-induced IL-4-pr(Klucing cells in the atopic indi-viduals which was not seen in the beaithy individuals. Thenumber of IL-4-producing cells was significantly correlatedwith presence of allergen-specific serum igE-levels. At pre-sent we do not know whether this IL-4 is produced by T ornon-T cells. Despite this, the ELISPOT assay was shown tobe a useful tool for detection of allergen-specific IL-4-produeing eells in a PBMC-cuiture.
Aci^nowiedgements
We thank Professor Peter Pcrlmann for critically readingthis manuscript and Mrs Agneta Breitholz for technicalassistance. This work was supported by The King Gustafthe 5th 80-years Jubilee Foundation, The Swedish Asthmaand Allergy Association. The Swedish National Board forLaboratory Animals and the Vardal Foundation.
References
1 Johansson SGO. Bennich H, Berg T. In vitro diagnosis ofatopic allergy. 3. Quantitative estimation of circulating IgEantibodies hy radioallergosorbent test, lnt Arch Allergy ApplImmunol 1971; 41:443-51.
2 Del Prete G, Maggi E, Parronchi P, Chretien I, Tiri A, MacchiaD. Ricci M. Banchereau J, De Vries JE. Romagnani S. IL-4 isan essential factor for the IgE synthesis induced in vitro byhuman T cell clones and their supematants. J Immunol 1988;140:4193-98.
3 Pene J. Rousset F, Briere F, Chretien I. Bonnefoy J-Y. Spits H.Yokota T, Arai K-I, Banchereau J, De Vries JE. IgE productionby normal human lymphocytes is induced by interleukin 4 andsuppressed by interferons 7 and a and prostaglandin E .̂ ProcNatI Acad Sci USA 1988: 85:6880-84.
4 Vercelli D. Geha RS. Regulation of isotype switching. CurrOpin Immunol 1992; 4:794-97.
5 Coffman R. Carty J. A T ceil activity that enhances polycionalIgE production and its inhibition hy interteron-7. J Immunol1986; i 36:949-54.
6 Finkelman FD. Katona ID, Urban JF. Holmes J, Ohara J. TungAS, Sample JvG, Paul WE. IL-4 is required to generate andsustain in vivo IgE responses. J Immunol 1988; 141:2335-41.
7 Punnonen J, Aversa G, Cocks BG, McKen:die ANJ, Menon S.Zurawski G, de Waal Malefyt R, de Vries JE. Interleukin 13induces interleukin 4-independent IgG4 and IgE synthesis andCD23 expression by human B cells. Proc Natl Acad Sci USA1993:90:3730-34.
8 Mossmann TR, Cherwinski H. Bond MW. Giedlin MA, CoffmanRL. Two types of murine helper T cell clone. J Immunology1986; 136:2348-57.
9 Firestein GS, Roeder WD, Laxer JA, Townsend KS. WeaverCT. Hom JT. Linton J, Torbett BE, Glasebrook AL. A newmurine CD4"'' T cell subset with an unrestricted cytokineprofile. J Immunol 1989; 143:518-25.
10 Romagnani S. Lymphokine production by human T cells indisease states. Ann Rev Immunol 1994; 12:227-57.
11 Abbas AK. Williams ME, Burstein HJ. Chang TL. Bossu P.Lichtman AH. Activation and functions of CD4"'" T-cel! sub-sets, Imm Rev 1991: 123:5-22.
12 Snapper CM. Paul WE. Interferon-7 and B cell stimulatoryfactor-1 reciprocally regulate Ig isotype production. Science1987; 236:944-47.
13 Sedgwick JD. Holt PG. A solid-phase immunoenzymatietechnique for the cnuineration of specific antibody-sec ret ingcells. J Immunol Melhods 1983; 57:301-9.
14 Versteegen JMT. Logtenberg T. Ballieux RE. Enumeration of
1997 Blackwell Science Lid. Clinical and Experitnenlal Allergy. 27.808-815

IL-4-producing cells in respon.se to allergen 815
IFN-7-prodiicing human lymphocytes by spot-ELISA. J Immu-nol Methods 1988: 111:25-9.
15 Czerkinsky C, Andcrsson G. Fcrrtia B. Nordstrom I. QuidingM, Eriksson K, Lar.sson L. Hellstrand K, Ekre H-P. Detectionof human cytokine-secreting cells in distinct anatomical com-partments. Immunol Rev 1991; 119:5-22.
1 (1 ElGhazall GEB, Paulie S, Andersson G. Hansson Y, HolmquistG. Sun J-B. Olsson T, Ekre HP, Troye-Blomberg M. Numberof interleukin-4- and interferon-7-secreting human T cell.sreactive with tetanu.s toxoid and the mycobacterial antigenPPD or phytohemagglutinin: distinct response profiles depend-ing on the type i)f antigen used lor activation. Eur J Immunol1993; 23:2740-45.
17 Pene J. Rivier A, LagierB, Becker WM, Michel F-B. BousquetJ. Differences in lL-4 release by PBMC are related withheterogeneity of atopy. Immunology 1994: 81:58-64.
IS Powers GD, Abbas AK. Miller RA. Frequencies of IL-2- andIL-4-seereting T cells in naive and antigen-stimulated lympho-cyte populations. J Immunol 1988: l4{):3352-57.
19 Kabilan L. Anders.son G. Lolli F. Ekre H, Troye-Blomberg M.Detection of intracellular expression and secretion of intcr-feron-7 at the single-cell level after activation of human T cellswith tetanus toxoid in vitro. Eur J Immunol 1990: 20:1085-89.
20 Goodier MR. Lundqvist C. Hammarstom ML, Troye-Blomberg M, Langhome J. Cytokine profiles for humanV79^ T cell.s stimulated by Plasmodium faleiparum. ParasiteImmunol 1995; 17:413-23.
21 Del Prete GF. de Carli M. D'Elios MM. Maestrelli P, Ricci M,Fabbri L. Romagnani S. Allergen exposure induces the activa-tion of allergen-specitie Th2 cells in the airway mucosa ofpatients with allergic respiratory disorders. Eur J Immunol1993:23:1445-49.
22 Ishizuka T, Kawagoe M. Suzuki K. Hara M. Harigai M.Kawakami M. Kawaguchi Y. Hidaka T. Matsuki Y, TanakaN, Kitani A, Nakamura H. An ultrasensitive system to detectIL-4: enzyme-linked immunosorbent a.ssay (ELISA) combinedwith an avidin-biotin and enzyme amplification system. JImmunol Methods 1992; 153:213-22.
23 Marshall JD. Wen Y. Abrams JS, Umctsu DT. In vitro synth-esis of IL-4 by human CD4''' T cells requires repeated antigenicstimulation. Ceil Immunol 1993: 152:18-34.
24 Secrist H, Chelen CJ. Wen Y. Marshall JD, Umetsu DT.Allergen immunotherapy decreases interleukin 4 productionin CD4-f T cells from allergic individuals. J Exp Med 1993:178:2123-30.
25 Secrist H. DeKruyff RH. Umetsu DT. Interleukin 4 productionby CD4+ T cells from allergic individuals is modulated byantigen concentration and antigen-presenting cell type. J ExpMed 1995: 181:1081-89.
26 Kurtzhals JAL. Hansen MB. Hey AS. Poulsen LK. Measure-ment of antigen-dependent interleukin-4 production by human
peripheral blood mononuclear cells. J Immunol Methods 1992;156:239-45.
27 Romagnani S. Maggi E. Parronchi P. Maeehia D, Piccinni MP.Ricci M. Increased numbers of Th2-like CD4+ T cells in targetorgans and in the allergen-speeifie repertoire of allergicpatients. Int Arch Allergy AppI Immunol 1991; 94:133-36.
28 Wierenga EA. Snoek MdG C. Chretien 1. Bos JDJHM, Kap-senberg ML. Evidence for compartmentalization of functionalsubsets of €04"^ T lymphocytes in atopic patients. J Immunol1990; 144:4651-56.
29 Romagnani S, Del Prete G. Maggi E, Parronchi P. Tiri A,Macchia D, Giudizi MGAE, Ricci M. Role of interleukins ininduction and regulation of human IgE synthe.sis. Clin ImmunImmunopath 1989; 50:sl3-23.
30 Reinhold U. Wehrmann W. Kukel S. Kreysel HW. Evidencethat defective interferon-gamma production in atopie dermati-tis patients is due to intrinsic abnormalities. Clin Exp Immunol1990; 79:374-9.
31 Hviid L, Theander TG. Abdulhadi NH, Abu-Zeid UA.Bayoumi RA. Jensen JB. Transient depletion of T cells withhigh LFA-1 expression from peripheral circulation duringacute Plasmodium faleiparum malaria. Eur J Immunol 1991;21:1249-53.
32 Seder RA. Le Gros G. Ben-Sasson SZ, Urban J. Finkelman FD.Paul WE, Increased frequency of interleukin 4-producing Tcells as a result of polyclonal priming. Use of a single-cellassay to detect interleukin 4-producing cells, Eur J Immunol1991; 21:1241-47,
33 Le Gros G. Erard F. Non-eytotoxic, IL-4. IL-5, IL-IO produ-cing CD8-I- T celts: their activation and effector functions. CurrOpin Immunol 1994; 6:453-57.
34 Ferrick DA. Schrenzel MD. Mulvania T, Hsieh B. Ferlin WG.Lepper H, Differential production of interferon 7 and inter-leukin-4 in response to Th I- and Th2-stimulating pathogens by76T cells in vivo. Nature 1995; 373:255-57.
35 Zlotnik A. Bean AG. Production of IL4 by non-Th2 T-cellsubsets: possible role of CD4- CDS- alpha beta TCR+ andCD4 subset T cells in T helper subset regulation. Res Immunol1993: 144:606-9,
36 Ben-Sasson S. Le Gros G. Conrad DH. Finkelman FD, PaulWE. Cross-linking Fc receptors stimulate splenic non-B. non-Tcells to secrete interleukin 4 and other lymphokines. Proc NatlAead Sci USA 1990: 87:1421-25.
37 Schmitz J. Thiel A. Kuhn R. Rajewsky K, Muller W, Assen-macher M. Radbrueh A, Induction of interleukin 4 (IL-4)expression in T helper (Th) cells is not dependent on IL-4from non-Th-cells. J Exp Med 1994: 179:1349-53.
38 Siginundsdottir H, Sirgurgerrsson B. Troye-Blomberg M et al.Circulating T-cells of patients with active psiorisis respond tostreptococcal M-peptides sharing sequences with human epi-dermal keratins. Scand J Immunol 1997; in press.
1997 Blackwell Science Ltd, Clinical and E.xperimental Allergy. 17.

Copyright © 2022 FDOKUMEN




















