
Spatial distribution of introduced Norway spruce growth in lowland Poland: The influence of changing...
8
Spatial distribution of introduced Norway spruce growth in lowland Poland: The influence of changing climate and extreme weather events Marcin Koprowski Nicolaus Copernicus University, Institute of Ecology and Environment Protection, Laboratory of Dendrochronology, Gagarina 9, 87-100 Torun, Poland article info Article history: Available online 20 April 2012 abstract Norway spruce trees grown outside their natural range were intensively studied in terms of their adaptability to increased late winter temperature. A total of 708 cores of Norway spruce were taken from 35 sites throughout northwestern and western Poland at sites outside or at the margin of its natural range. Clustering reveals that research sites are grouped geographically. The mean ring widths, however, are similar among the three regions. The relationships between tree growth and temperature together with precipitation are comparable to those in lower elevations in Central and Eastern Europe and southern latitudes in Scandinavia. In total, 18% of the single correlations for all months were significant at p < 0.05. The generally positive precipitation effect was found for all months except January. Temper- ature had mixed effects on ring width. Higher temperatures in late winter and early spring increased ring width (13% of all significant correlations). High summer temperatures in both the previous and the current year, on the other hand, generally had a negative impact on tree-ring width during the growing season. On most of the sites, the observed long-term trend for increasing temperatures in February and March had no significant negative influence on tree growth. Also the effect of extreme weather events was taken into account. In 1994 and 1995 the wider rings were mostly observed in humid sites from southwestern Poland. In selected time spans, growth recovery following extreme weather events is particularly strong. Ó 2012 Elsevier Ltd and INQUA. All rights reserved. 1. Introduction Norway spruce (Picea abies [L.] Karst.) occurs naturally in the northeastern and southern parts of Poland while its natural occurrence in the central part of eastern Poland remains contro- versial (Fig. 1)(Lata1owa and van der Knaap, 2006; Dering and Lewandowski, 2009). During the last 200 years spruce has been planted, particularly in mountain areas of southwestern Poland and in the lake-lands of northwestern region. As a result, the occurrence of spruce has been extended to large areas outside its natural range (Boraty nski, 1998) and the importance of this tree in forest management has increased. In the case of spruce, the major influences on geographical distribution other than climate are the evolutionary-genetic tree tolerance, extreme weather events (Podbielkowski, 1991), and negative impact of management activity together with unfavourable soil conditions ( Srodo n, 1967). The effects of climatic change have been observed in Poland (Ko _ zuchowski and Degirmend zi c, 2005), and its impact on tree growth may differ within selected time spans. Further, tree-ring increment patterns might differ across larger regions due to meso-scale differences in climate. In the last 50 years Central Europe has seen increasing average temperatures and higher minimum temperatures in February and March, and thus an earlier onset of spring by more than two weeks (Ko _ zuchowski and Degirmend zi c, 2005). The impact of these higher temperatures on the growth and health status of Norway spruce is still not fully understood. Because spruce is the second most important tree species after pine in Polish forests (Boraty nski and Buga1a, 1998), improved understanding is clearly needed on the impact of climate change on the ecology and growth of spruce (Koprowski and Zielski, 2006). The present work therefore has three major aims: 1. To identify the climate parameters that affect the growth of Norway spruce outside its natural range and to compare these patterns to those found for spruce within its natural range. The latter is already sufficiently known for many European natural stands (Mäkinen et al., 2003; Andreassen et al., 2006; Koprowski and Vitas, 2010). The study of trees outside the natural range might help to determine how spruce reacts after E-mail address: [email protected]. Contents lists available at SciVerse ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint 1040-6182/$ e see front matter Ó 2012 Elsevier Ltd and INQUA. All rights reserved. doi:10.1016/j.quaint.2012.04.020 Quaternary International 283 (2013) 139e146
Transcript of Spatial distribution of introduced Norway spruce growth in lowland Poland: The influence of changing...

at SciVerse ScienceDirect
Quaternary International 283 (2013) 139e146
Contents lists available
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Spatial distribution of introduced Norway spruce growth in lowland Poland: Theinfluence of changing climate and extreme weather events
Marcin KoprowskiNicolaus Copernicus University, Institute of Ecology and Environment Protection, Laboratory of Dendrochronology, Gagarina 9, 87-100 Torun, Poland
a r t i c l e i n f o
Article history:Available online 20 April 2012
E-mail address: [email protected].
1040-6182/$ e see front matter � 2012 Elsevier Ltd adoi:10.1016/j.quaint.2012.04.020
a b s t r a c t
Norway spruce trees grown outside their natural range were intensively studied in terms of theiradaptability to increased late winter temperature. A total of 708 cores of Norway spruce were taken from35 sites throughout northwestern and western Poland at sites outside or at the margin of its naturalrange. Clustering reveals that research sites are grouped geographically. The mean ring widths, however,are similar among the three regions. The relationships between tree growth and temperature togetherwith precipitation are comparable to those in lower elevations in Central and Eastern Europe andsouthern latitudes in Scandinavia. In total, 18% of the single correlations for all months were significant atp < 0.05. The generally positive precipitation effect was found for all months except January. Temper-ature had mixed effects on ring width. Higher temperatures in late winter and early spring increased ringwidth (13% of all significant correlations). High summer temperatures in both the previous and thecurrent year, on the other hand, generally had a negative impact on tree-ring width during the growingseason. On most of the sites, the observed long-term trend for increasing temperatures in February andMarch had no significant negative influence on tree growth. Also the effect of extreme weather eventswas taken into account. In 1994 and 1995 the wider rings were mostly observed in humid sites fromsouthwestern Poland. In selected time spans, growth recovery following extreme weather events isparticularly strong.
� 2012 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
Norway spruce (Picea abies [L.] Karst.) occurs naturally in thenortheastern and southern parts of Poland while its naturaloccurrence in the central part of eastern Poland remains contro-versial (Fig. 1) (Lata1owa and van der Knaap, 2006; Dering andLewandowski, 2009). During the last 200 years spruce has beenplanted, particularly in mountain areas of southwestern Poland andin the lake-lands of northwestern region. As a result, the occurrenceof spruce has been extended to large areas outside its natural range(Boraty�nski, 1998) and the importance of this tree in forestmanagement has increased.
In the case of spruce, the major influences on geographicaldistribution other than climate are the evolutionary-genetic treetolerance, extreme weather events (Podbielkowski, 1991), andnegative impact of management activity together with unfavourablesoil conditions (�Srodo�n, 1967). The effects of climatic change havebeen observed in Poland (Ko _zuchowski and Degirmend�zi�c, 2005),
nd INQUA. All rights reserved.
and its impact on tree growth may differ within selected time spans.Further, tree-ring increment patterns might differ across largerregions due to meso-scale differences in climate. In the last 50 yearsCentral Europe has seen increasing average temperatures and higherminimum temperatures in February and March, and thus an earlieronset of spring by more than two weeks (Ko _zuchowski andDegirmend�zi�c, 2005). The impact of these higher temperatures onthe growth and health status of Norway spruce is still not fullyunderstood. Because spruce is the second most important treespecies after pine in Polish forests (Boraty�nski and Buga1a, 1998),improved understanding is clearly needed on the impact of climatechange on the ecology and growth of spruce (Koprowski and Zielski,2006). The present work therefore has three major aims:
1. To identify the climate parameters that affect the growth ofNorway spruce outside its natural range and to compare thesepatterns to those found for spruce within its natural range. Thelatter is already sufficiently known for many European naturalstands (Mäkinen et al., 2003; Andreassen et al., 2006;Koprowski and Vitas, 2010). The study of trees outside thenatural range might help to determine how spruce reacts after
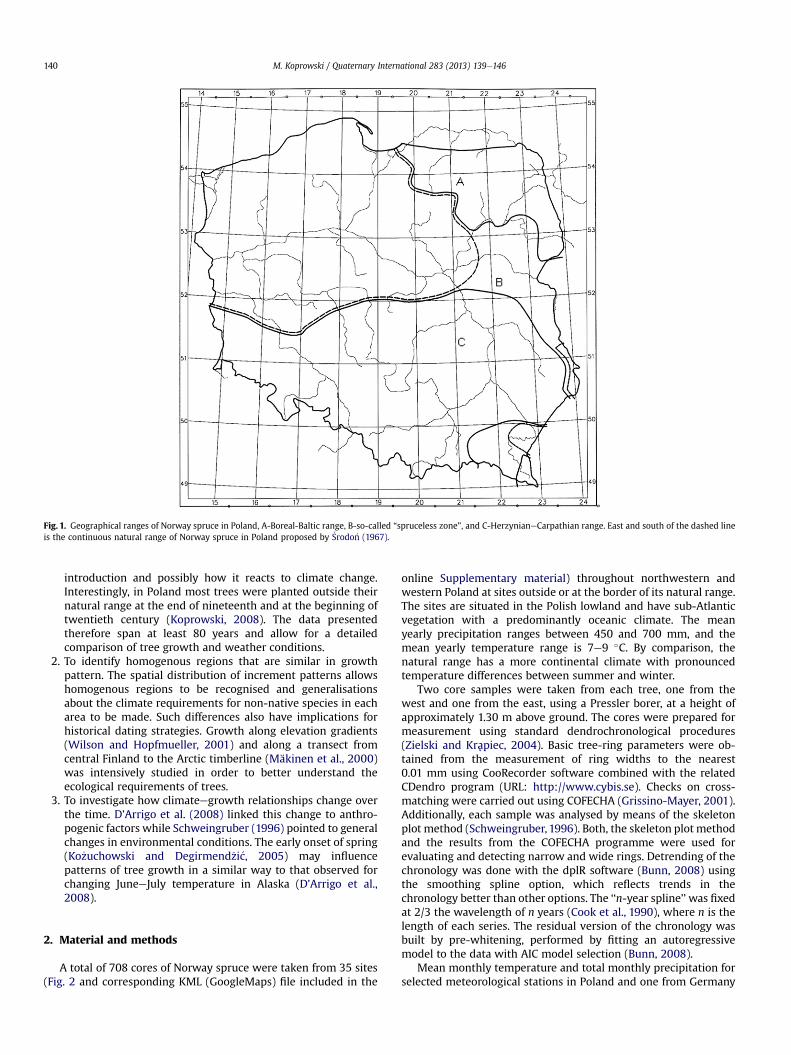
Fig. 1. Geographical ranges of Norway spruce in Poland, A-Boreal-Baltic range, B-so-called “spruceless zone”, and C-HerzynianeCarpathian range. East and south of the dashed lineis the continuous natural range of Norway spruce in Poland proposed by �Srodo�n (1967).
M. Koprowski / Quaternary International 283 (2013) 139e146140
introduction and possibly how it reacts to climate change.Interestingly, in Poland most trees were planted outside theirnatural range at the end of nineteenth and at the beginning oftwentieth century (Koprowski, 2008). The data presentedtherefore span at least 80 years and allow for a detailedcomparison of tree growth and weather conditions.
2. To identify homogenous regions that are similar in growthpattern. The spatial distribution of increment patterns allowshomogenous regions to be recognised and generalisationsabout the climate requirements for non-native species in eacharea to be made. Such differences also have implications forhistorical dating strategies. Growth along elevation gradients(Wilson and Hopfmueller, 2001) and along a transect fromcentral Finland to the Arctic timberline (Mäkinen et al., 2000)was intensively studied in order to better understand theecological requirements of trees.
3. To investigate how climateegrowth relationships change overthe time. D’Arrigo et al. (2008) linked this change to anthro-pogenic factors while Schweingruber (1996) pointed to generalchanges in environmental conditions. The early onset of spring(Ko _zuchowski and Degirmend�zi�c, 2005) may influencepatterns of tree growth in a similar way to that observed forchanging JuneeJuly temperature in Alaska (D’Arrigo et al.,2008).
2. Material and methods
A total of 708 cores of Norway spruce were taken from 35 sites(Fig. 2 and corresponding KML (GoogleMaps) file included in the
online Supplementary material) throughout northwestern andwestern Poland at sites outside or at the border of its natural range.The sites are situated in the Polish lowland and have sub-Atlanticvegetation with a predominantly oceanic climate. The meanyearly precipitation ranges between 450 and 700 mm, and themean yearly temperature range is 7e9 �C. By comparison, thenatural range has a more continental climate with pronouncedtemperature differences between summer and winter.
Two core samples were taken from each tree, one from thewest and one from the east, using a Pressler borer, at a height ofapproximately 1.30 m above ground. The cores were prepared formeasurement using standard dendrochronological procedures(Zielski and Krapiec, 2004). Basic tree-ring parameters were ob-tained from the measurement of ring widths to the nearest0.01 mm using CooRecorder software combined with the relatedCDendro program (URL: http://www.cybis.se). Checks on cross-matching were carried out using COFECHA (Grissino-Mayer, 2001).Additionally, each sample was analysed by means of the skeletonplot method (Schweingruber, 1996). Both, the skeleton plot methodand the results from the COFECHA programme were used forevaluating and detecting narrow and wide rings. Detrending of thechronology was done with the dplR software (Bunn, 2008) usingthe smoothing spline option, which reflects trends in thechronology better than other options. The ‘‘n-year spline’’was fixedat 2/3 the wavelength of n years (Cook et al., 1990), where n is thelength of each series. The residual version of the chronology wasbuilt by pre-whitening, performed by fitting an autoregressivemodel to the data with AIC model selection (Bunn, 2008).
Mean monthly temperature and total monthly precipitation forselected meteorological stations in Poland and one from Germany
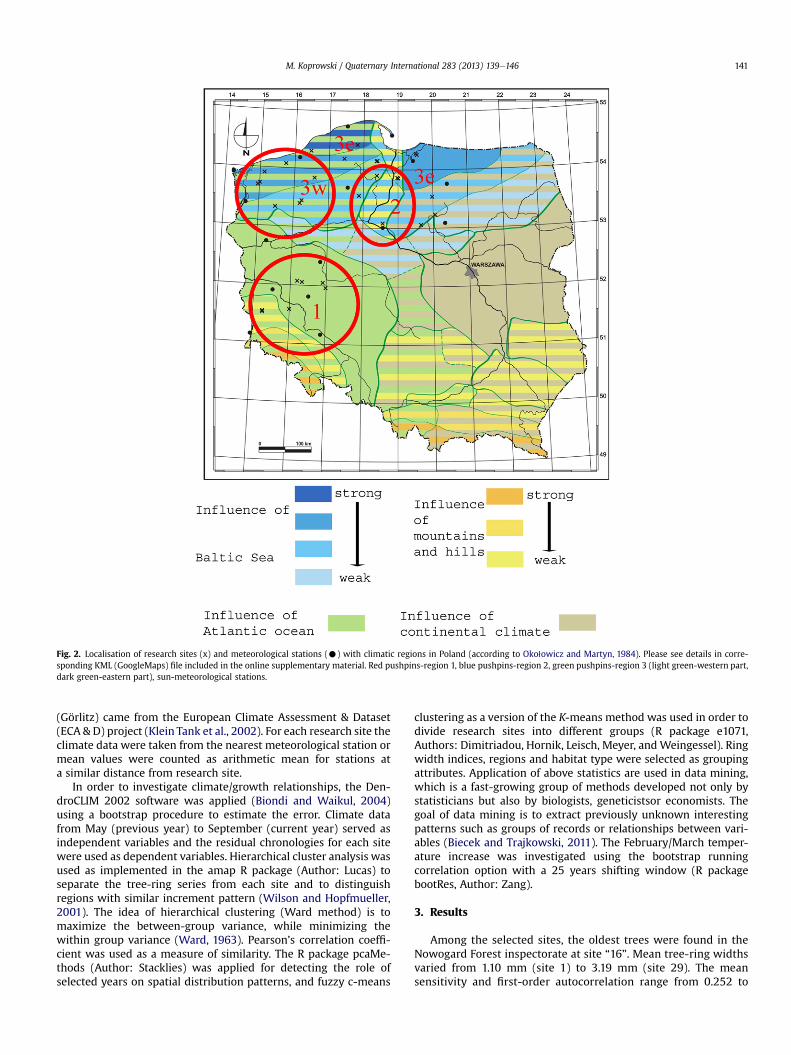
Fig. 2. Localisation of research sites (x) and meteorological stations (C) with climatic regions in Poland (according to Oko1owicz and Martyn, 1984). Please see details in corre-sponding KML (GoogleMaps) file included in the online supplementary material. Red pushpins-region 1, blue pushpins-region 2, green pushpins-region 3 (light green-western part,dark green-eastern part), sun-meteorological stations.
M. Koprowski / Quaternary International 283 (2013) 139e146 141
(Görlitz) came from the European Climate Assessment & Dataset(ECA & D) project (Klein Tank et al., 2002). For each research site theclimate data were taken from the nearest meteorological station ormean values were counted as arithmetic mean for stations ata similar distance from research site.
In order to investigate climate/growth relationships, the Den-droCLIM 2002 software was applied (Biondi and Waikul, 2004)using a bootstrap procedure to estimate the error. Climate datafrom May (previous year) to September (current year) served asindependent variables and the residual chronologies for each sitewere used as dependent variables. Hierarchical cluster analysis wasused as implemented in the amap R package (Author: Lucas) toseparate the tree-ring series from each site and to distinguishregions with similar increment pattern (Wilson and Hopfmueller,2001). The idea of hierarchical clustering (Ward method) is tomaximize the between-group variance, while minimizing thewithin group variance (Ward, 1963). Pearson’s correlation coeffi-cient was used as a measure of similarity. The R package pcaMe-thods (Author: Stacklies) was applied for detecting the role ofselected years on spatial distribution patterns, and fuzzy c-means
clustering as a version of the K-means method was used in order todivide research sites into different groups (R package e1071,Authors: Dimitriadou, Hornik, Leisch, Meyer, andWeingessel). Ringwidth indices, regions and habitat type were selected as groupingattributes. Application of above statistics are used in data mining,which is a fast-growing group of methods developed not only bystatisticians but also by biologists, geneticistsor economists. Thegoal of data mining is to extract previously unknown interestingpatterns such as groups of records or relationships between vari-ables (Biecek and Trajkowski, 2011). The February/March temper-ature increase was investigated using the bootstrap runningcorrelation option with a 25 years shifting window (R packagebootRes, Author: Zang).
3. Results
Among the selected sites, the oldest trees were found in theNowogard Forest inspectorate at site “16”. Mean tree-ring widthsvaried from 1.10 mm (site 1) to 3.19 mm (site 29). The meansensitivity and first-order autocorrelation range from 0.252 to
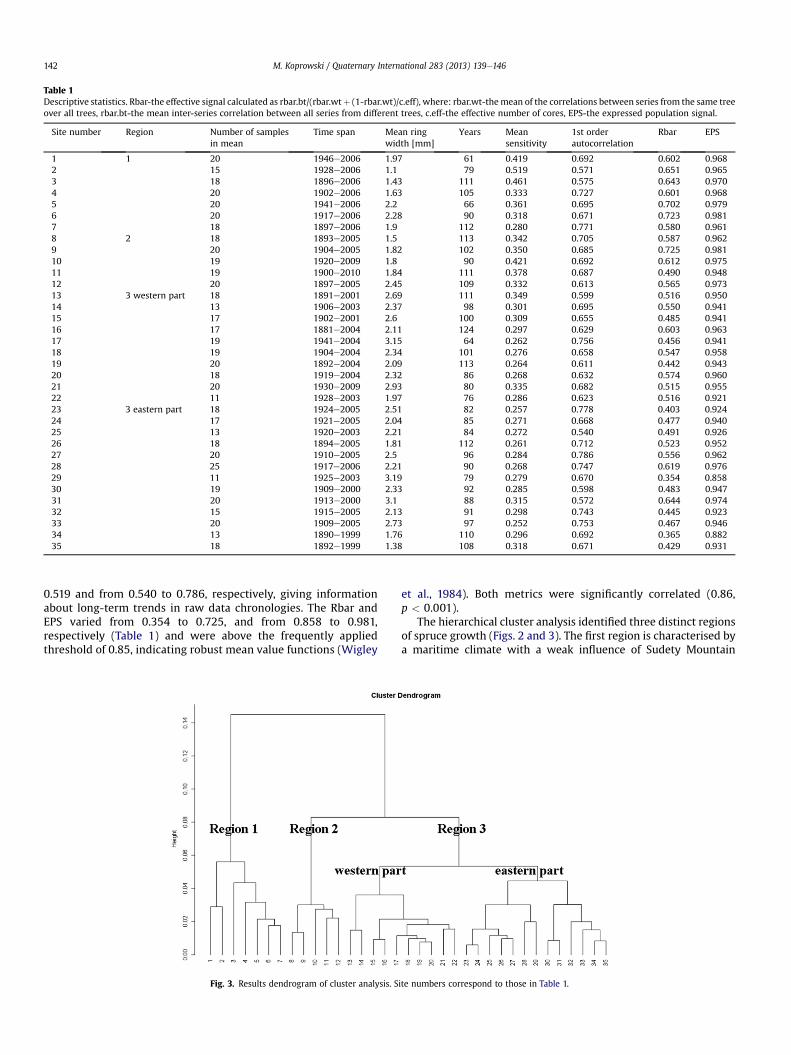
Table 1Descriptive statistics. Rbar-the effective signal calculated as rbar.bt/(rbar.wtþ (1-rbar.wt)/c.eff), where: rbar.wt-themean of the correlations between series from the same treeover all trees, rbar.bt-the mean inter-series correlation between all series from different trees, c.eff-the effective number of cores, EPS-the expressed population signal.
Site number Region Number of samplesin mean
Time span Mean ringwidth [mm]
Years Meansensitivity
1st orderautocorrelation
Rbar EPS
1 1 20 1946e2006 1.97 61 0.419 0.692 0.602 0.9682 15 1928e2006 1.1 79 0.519 0.571 0.651 0.9653 18 1896e2006 1.43 111 0.461 0.575 0.643 0.9704 20 1902e2006 1.63 105 0.333 0.727 0.601 0.9685 20 1941e2006 2.2 66 0.361 0.695 0.702 0.9796 20 1917e2006 2.28 90 0.318 0.671 0.723 0.9817 18 1897e2006 1.9 112 0.280 0.771 0.580 0.9618 2 18 1893e2005 1.5 113 0.342 0.705 0.587 0.9629 20 1904e2005 1.82 102 0.350 0.685 0.725 0.98110 19 1920e2009 1.8 90 0.421 0.692 0.612 0.97511 19 1900e2010 1.84 111 0.378 0.687 0.490 0.94812 20 1897e2005 2.45 109 0.332 0.613 0.565 0.97313 3 western part 18 1891e2001 2.69 111 0.349 0.599 0.516 0.95014 13 1906e2003 2.37 98 0.301 0.695 0.550 0.94115 17 1902e2001 2.6 100 0.309 0.655 0.485 0.94116 17 1881e2004 2.11 124 0.297 0.629 0.603 0.96317 19 1941e2004 3.15 64 0.262 0.756 0.456 0.94118 19 1904e2004 2.34 101 0.276 0.658 0.547 0.95819 20 1892e2004 2.09 113 0.264 0.611 0.442 0.94320 18 1919e2004 2.32 86 0.268 0.632 0.574 0.96021 20 1930e2009 2.93 80 0.335 0.682 0.515 0.95522 11 1928e2003 1.97 76 0.286 0.623 0.516 0.92123 3 eastern part 18 1924e2005 2.51 82 0.257 0.778 0.403 0.92424 17 1921e2005 2.04 85 0.271 0.668 0.477 0.94025 13 1920e2003 2.21 84 0.272 0.540 0.491 0.92626 18 1894e2005 1.81 112 0.261 0.712 0.523 0.95227 20 1910e2005 2.5 96 0.284 0.786 0.556 0.96228 25 1917e2006 2.21 90 0.268 0.747 0.619 0.97629 11 1925e2003 3.19 79 0.279 0.670 0.354 0.85830 19 1909e2000 2.33 92 0.285 0.598 0.483 0.94731 20 1913e2000 3.1 88 0.315 0.572 0.644 0.97432 15 1915e2005 2.13 91 0.298 0.743 0.445 0.92333 20 1909e2005 2.73 97 0.252 0.753 0.467 0.94634 13 1890e1999 1.76 110 0.296 0.692 0.365 0.88235 18 1892e1999 1.38 108 0.318 0.671 0.429 0.931
M. Koprowski / Quaternary International 283 (2013) 139e146142
0.519 and from 0.540 to 0.786, respectively, giving informationabout long-term trends in raw data chronologies. The Rbar andEPS varied from 0.354 to 0.725, and from 0.858 to 0.981,respectively (Table 1) and were above the frequently appliedthreshold of 0.85, indicating robust mean value functions (Wigley
Fig. 3. Results dendrogram of cluster analysis. S
et al., 1984). Both metrics were significantly correlated (0.86,p < 0.001).
The hierarchical cluster analysis identified three distinct regionsof spruce growth (Figs. 2 and 3). The first region is characterised bya maritime climate with a weak influence of Sudety Mountain
ite numbers correspond to those in Table 1.
M. Koprowski / Quaternary International 283 (2013) 139e146 143
microclimate at its southern boundary. The second region isa mixture of Baltic climate aspects and is modulated by the pres-ence of the hills. The third region in northern Poland is predomi-nantly under the influence of the Baltic Sea.
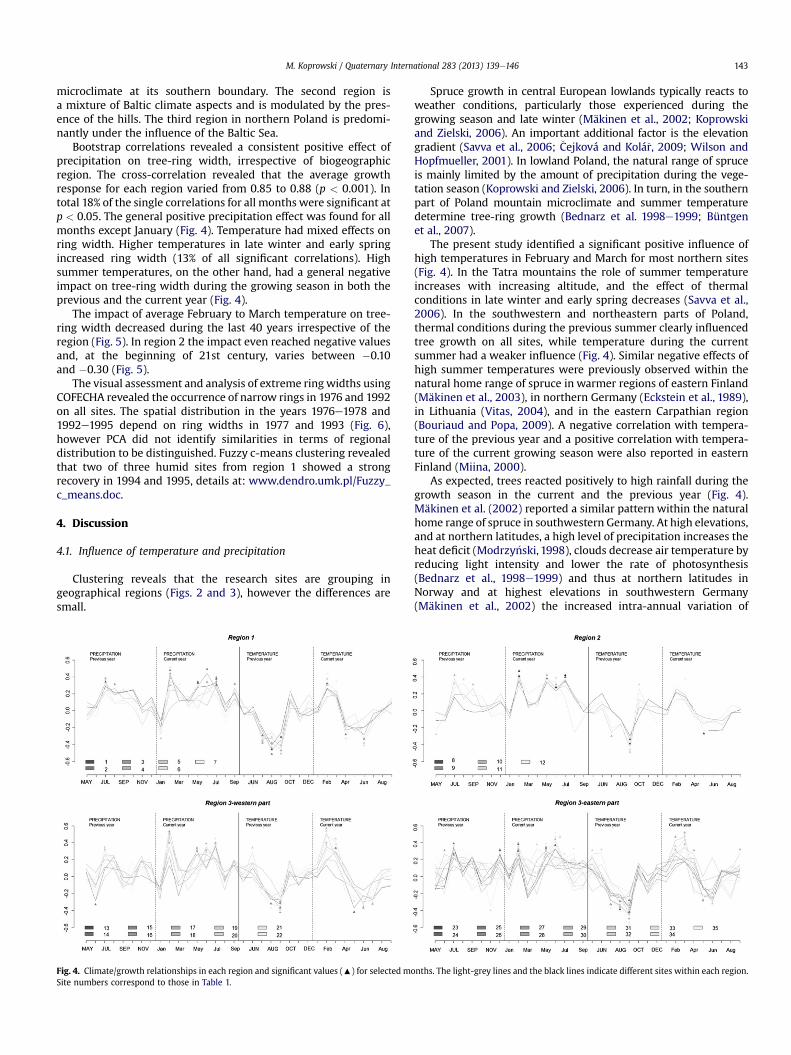
Bootstrap correlations revealed a consistent positive effect ofprecipitation on tree-ring width, irrespective of biogeographicregion. The cross-correlation revealed that the average growthresponse for each region varied from 0.85 to 0.88 (p < 0.001). Intotal 18% of the single correlations for all months were significant atp < 0.05. The general positive precipitation effect was found for allmonths except January (Fig. 4). Temperature had mixed effects onring width. Higher temperatures in late winter and early springincreased ring width (13% of all significant correlations). Highsummer temperatures, on the other hand, had a general negativeimpact on tree-ring width during the growing season in both theprevious and the current year (Fig. 4).
The impact of average February to March temperature on tree-ring width decreased during the last 40 years irrespective of theregion (Fig. 5). In region 2 the impact even reached negative valuesand, at the beginning of 21st century, varies between �0.10and �0.30 (Fig. 5).
The visual assessment and analysis of extreme ring widths usingCOFECHA revealed the occurrence of narrow rings in 1976 and 1992on all sites. The spatial distribution in the years 1976e1978 and1992e1995 depend on ring widths in 1977 and 1993 (Fig. 6),however PCA did not identify similarities in terms of regionaldistribution to be distinguished. Fuzzy c-means clustering revealedthat two of three humid sites from region 1 showed a strongrecovery in 1994 and 1995, details at: www.dendro.umk.pl/Fuzzy_c_means.doc.
4. Discussion
4.1. Influence of temperature and precipitation
Clustering reveals that the research sites are grouping ingeographical regions (Figs. 2 and 3), however the differences aresmall.
Fig. 4. Climate/growth relationships in each region and significant values (:) for selected moSite numbers correspond to those in Table 1.
Spruce growth in central European lowlands typically reacts toweather conditions, particularly those experienced during thegrowing season and late winter (Mäkinen et al., 2002; Koprowskiand Zielski, 2006). An important additional factor is the elevationgradient (Savva et al., 2006; �Cejková and Kolá�r, 2009; Wilson andHopfmueller, 2001). In lowland Poland, the natural range of spruceis mainly limited by the amount of precipitation during the vege-tation season (Koprowski and Zielski, 2006). In turn, in the southernpart of Poland mountain microclimate and summer temperaturedetermine tree-ring growth (Bednarz et al. 1998e1999; Büntgenet al., 2007).
The present study identified a significant positive influence ofhigh temperatures in February and March for most northern sites(Fig. 4). In the Tatra mountains the role of summer temperatureincreases with increasing altitude, and the effect of thermalconditions in late winter and early spring decreases (Savva et al.,2006). In the southwestern and northeastern parts of Poland,thermal conditions during the previous summer clearly influencedtree growth on all sites, while temperature during the currentsummer had a weaker influence (Fig. 4). Similar negative effects ofhigh summer temperatures were previously observed within thenatural home range of spruce in warmer regions of eastern Finland(Mäkinen et al., 2003), in northern Germany (Eckstein et al., 1989),in Lithuania (Vitas, 2004), and in the eastern Carpathian region(Bouriaud and Popa, 2009). A negative correlation with tempera-ture of the previous year and a positive correlation with tempera-ture of the current growing season were also reported in easternFinland (Miina, 2000).
As expected, trees reacted positively to high rainfall during thegrowth season in the current and the previous year (Fig. 4).Mäkinen et al. (2002) reported a similar pattern within the naturalhome range of spruce in southwestern Germany. At high elevations,and at northern latitudes, a high level of precipitation increases theheat deficit (Modrzy�nski, 1998), clouds decrease air temperature byreducing light intensity and lower the rate of photosynthesis(Bednarz et al., 1998e1999) and thus at northern latitudes inNorway and at highest elevations in southwestern Germany(Mäkinen et al., 2002) the increased intra-annual variation of
nths. The light-grey lines and the black lines indicate different sites within each region.
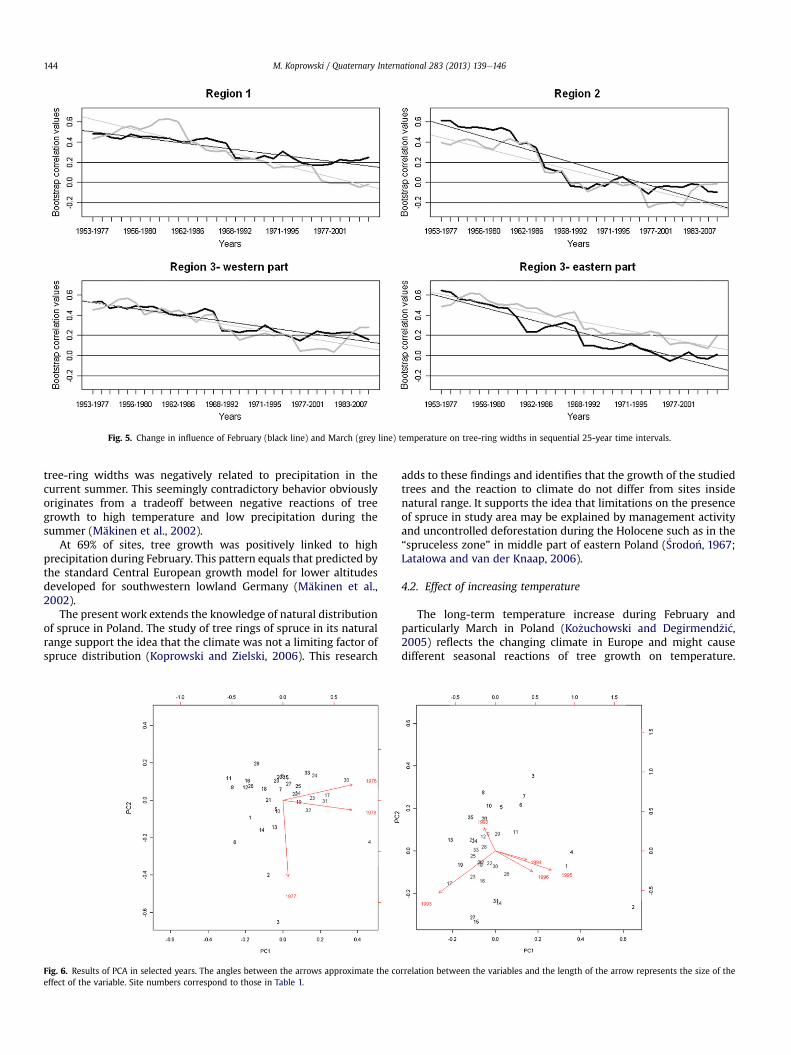
Fig. 5. Change in influence of February (black line) and March (grey line) temperature on tree-ring widths in sequential 25-year time intervals.
M. Koprowski / Quaternary International 283 (2013) 139e146144
tree-ring widths was negatively related to precipitation in thecurrent summer. This seemingly contradictory behavior obviouslyoriginates from a tradeoff between negative reactions of treegrowth to high temperature and low precipitation during thesummer (Mäkinen et al., 2002).
At 69% of sites, tree growth was positively linked to highprecipitation during February. This pattern equals that predicted bythe standard Central European growth model for lower altitudesdeveloped for southwestern lowland Germany (Mäkinen et al.,2002).
The present work extends the knowledge of natural distributionof spruce in Poland. The study of tree rings of spruce in its naturalrange support the idea that the climate was not a limiting factor ofspruce distribution (Koprowski and Zielski, 2006). This research
Fig. 6. Results of PCA in selected years. The angles between the arrows approximate the coeffect of the variable. Site numbers correspond to those in Table 1.
adds to these findings and identifies that the growth of the studiedtrees and the reaction to climate do not differ from sites insidenatural range. It supports the idea that limitations on the presenceof spruce in study area may be explained by management activityand uncontrolled deforestation during the Holocene such as in the“spruceless zone” in middle part of eastern Poland (�Srodo�n, 1967;Lata1owa and van der Knaap, 2006).
4.2. Effect of increasing temperature
The long-term temperature increase during February andparticularly March in Poland (Ko _zuchowski and Degirmend�zi�c,2005) reflects the changing climate in Europe and might causedifferent seasonal reactions of tree growth on temperature.
rrelation between the variables and the length of the arrow represents the size of the

M. Koprowski / Quaternary International 283 (2013) 139e146 145
D’Arrigo et al. (2008) called this phenomenon the “divergenceproblem”. It had been observed in northern Alaskan forests andassociated with JuneeJuly mean temperatures (D’Arrigo et al.,2008). The correlation observed here, using a 25-year movingwindow, revealed a decreasing impact of late winter temperatures(Fig. 5). The strongest impact was observed from the 1940s to the1970s. In region 2 the impact even reached negative values,meaning that higher temperatures negatively influenced cambiumactivity during the growth season. This may lead in the long run tounfavourable tendencies. For instance, a temperature of �10 �Cinduces tree resistance to frost from �20 �C to �30 �C(Mitscherlich, 1971). The increase of mean FebruaryeMarch airtemperatures and its fluctuations can disturb this hardening oftrees. The highest level of tree adaptation to low temperatures isreached when temperatures are sustained between �5 �Cand �15 �C, and is connected with the phenomenon of super-cooling, allowing water in the plants to remain liquid (Larcher,2003). The temperature variations in late winter and early springcan lead to a decrease of the trees’ resistance to low air tempera-ture. However, at this stage of the research, the negative effect oftemperature increase is visible in March only in region 2 at the endof the twentieth century. In other regions, on most of the sites, thetemperature increase in February and March had no significantinfluence on tree growth (Fig. 5). The question of whether the treesgrow better or worse under projected climate change than do thetrees in the natural distribution is crucial for forest management.Because the relationships between climate and tree rings arecomparable in the whole of lowland Poland, the temperatureincrease will have similar effect on tree growth, both outside andinside natural range. Interestingly, a pollen study revealed thatduring the Preboreal (period of the Holocene), spruce re-expandedas the climate warmed (Lata1owa and van der Knaap, 2006).
4.3. Effect of extreme weather conditions
In different regions of northern and central Europe, Mäkinenet al. (2003) reported negative effects on spruce growth of theextreme drought in 1976. A growth decrease in 1977 observed inthe humid Belgium Ardennes must have been the consequence ofthe drought in 1976 (Misson et al., 2003). Vitas (2002) identified theyear 1992 as a pointer year in Lithuania and Koprowski and Zielski(2008) confirmed this for the Bia1owie _za National Park. The presentdata reveal the effects of these droughts for spruce both within andoutside its natural range (Fig. 6). A visual assessment of ring widthsand the PCA also point to differences in the subsequent recovery(Fig. 6). The strongest growth recovery in 1977 was observed at 2sites from region 1, one site from region 2 and one from region 3.Trees from southwestern Poland (region 1) show a more markeddecline in growth in 1993. PCA revealed that the abnormal growthin 1993 determined the differences between the growth models inyears 1992e1996 (Fig. 6). Each humid site (except site 28, region 2)and each site from region 1 had narrow rings in 1993 and recoveredin 1994 and 1995 (www.dendro.umk.pl/Fuzzy_c_means.doc).
In conclusion, the relationships between tree growth andtemperature together with precipitation are comparable to those inlower elevations in Central and Eastern Europe and southern lati-tudes in Scandinavia. It supports the idea that presence of spruce instudy area may be limited by the management activity anduncontrolled deforestation. The spatial modelling clearly groupsthe research sites on geographical and climatological factors. Thenegative effect of increased temperature in February and Marchwas identified at sites from region 2. For most of the research siteslocated in southwestern and northern Poland there was no rela-tionship between late winter temperature and tree-ring growth. Atpresent, climate warming has no negative, significant influence on
tree rings. However, the divergent influence of air temperature indifferent time intervals can lead to problems in terms of climatereconstructions. This phenomenon may disturb the process ofcalibration for modelling climate and tree rings of Norway spruce inCentral and Eastern Europe. Relationships between growthrecovery in 1977 and forest habitat have not been demonstrated. In1994 and 1995 wider rings were mostly observed in humid sitesfrom southwestern Poland. The most widespread strong influenceon growth was regeneration after extreme weather events.
Acknowledgements
This work was supported by the National Science Centre in theyears 2011e2013, habilitation grant number N N309 7091410. Ithank Martin Bridge for language correction and Werner Ulrich fordiscussions on the manuscript. I thank also Jerzy Mia1dun fordrawing maps. I owe special thanks to the staff of The State Forestsfor their hospitality and help. I would like also to thank familymembers and friends who helped in the fieldwork. This paper isa part of the habilitation thesis.
Appendix A. Supplementary material
Supplementary material associated with this article can befound, in the online version, at doi:10.1016/j.quaint.2012.04.020.
References
Andreassen, K., Solberg, S., Tveito, O.E., Lystad, S.L., 2006. Regional differences inclimatic response of Norway spruce (Picea abies (L.) Karst.) growth in Norway.Forest Ecology and Management 222, 211e221.
Bednarz, Z., Jaroszewicz, B., Ptak, J., Szwagrzyk, J., 1998-1999. Dendrochronology ofNorway spruce (Picea abies (L.) Karst.) in the Babia Góra National Park, Poland.Dendrochronologia 16e17, 45e55.
Biecek, P., Trajkowski, K., 2011. Na prze1aj przez Data Mining. URL. http://www.biecek.pl/NaPrzelajPrzezDataMining.
Biondi, F., Waikul, K., 2004. DENDROCLIM 2002: a Cþþ program for statisticalcalibration of climate signals in tree-ring chronologies. Computers &Geosciences 30, 303e311.
Boraty�nski, A., Buga1a, W., 1998. Biologia �swierka pospolitego. Bogucki Wydaw-nictwo Naukowe, Pozna�n.
Boraty�nski, A., 1998. Disjunctions in the range of the spruce. In: Boraty�nski, A.,Buga1a, W. (Eds.), Biologia �swierka pospolitego. Bogucki WydawnictwoNaukowe, Pozna�n, pp. 79e90.
Bouriaud, O., Popa, I., 2009. Comparative dendroclimatic study of Scots pine,Norway spruce and silver fir in the Vrancea range, Eastern Carpathian Moun-tains. Trees-Structure and Function 23, 95e106.
Bunn, A.G., 2008. A dendrochronology program library in R (dplR).Dendrochronologia 26, 115e124.
Büntgen, U., Frank, D.C., Kaczka, R.J., Verstege, A., Zwijacz-Kozica, T., Esper, J., 2007.Growth responses to climate in a multi-species tree-ring network in theWestern Carpathian Tatra Mountains, Poland and Slovakia. Tree Physiology 27,689e702.
�Cejková, A., Kolá�r, T., 2009. Extreme radial growth reaction of Norway spruce alongan altitudinal gradient in the �Sumava Mountains. Geochronometria 33, 41e47.
Cook, E.R., Briffa, K., Shiyatov, S., Mazepa, A., Jones, P.D., 1990. Data analysis. In:Cook, E.R., Kairiukstis, L.A. (Eds.), Methods of Dendrochronology: Applicationsin the Environmental Sciences. International Institute for Applied SystemsAnalysis, Kluwer Academic Publishers, Boston, pp. 97e162.
Dering, M., Lewandowski, A., 2009. Finding the meeting zone: where have thenorthern and southern ranges of Norway spruce overlapped? Forest Ecologyand Management 259, 229e235.
D’Arrigo, R., Wilson, R., Leipert, B., Cherubini, P., 2008. On the ‘divergence problem’
in northern forests. A review of the tree-ring evidence and possible causes.Global and Planetary Change 60, 289e305.
Eckstein, D., Krause, C., Bauch, J., 1989. Dendroecological investigation of sprucetrees (Picea abies (L.) karst.) of different damage and canopy classes. Holzfor-schung 43, 411e417.
Grissino-Mayer, H.D., 2001. Evaluating crossdating accuracy: a manual and tutorialfor the computer program COFECHA. Tree-Ring Research 57, 205e221.
Klein Tank, A.M.G., Wijngaard, J.B., Können, G.P., Böhm, R., Demaré, E.G.,Gocheva, A., Mileta, M., Pashiardis, S., Hejkrlik, L., Kern-Hansen, C., Heino, R.,Bessemoulin, P., Müller-Westermeier, G., Tzanakou, M., Szalai, S., PáLsdóTtir, T.,Fitzgerald, D., Rubin, S., Capaldo, M., Maugeri, M., Leitass, A., Bukantis, A.,Aberfeld, R., Vanengelen, A.F.V., Forland, E., Mietus, M., Coelho, F., Mares, C.,Razuvaev, V., Nieplova, E., Cegnar, T., AntoniolóPez, J., Dahlström, B., Moberg, A.,

M. Koprowski / Quaternary International 283 (2013) 139e146146
Kirchhofer, W., Ceylan, A., Pachaliuk, O., Alexander, L.V., Petrovic, P., 2002. Dailydataset of 20th-century surface air temperature and precipitation series for theEuropean Climate Assessment. International Journal of Climatology 22,1441e1453. Data and metadata available at URL: http://eca.knmi.nl.
Koprowski, M., 2008. Dendrochronologiczna analiza przyrostów rocznych �swierkapospolitego (Picea abies (L.) Karst.) poza naturalnym zasiegiem a cechy bio-metryczne szyszek. Sylwan 3, 50e57.
Koprowski, M., Vitas, A., 2010. Growth/climate relationships in tree-ring widths ofPicea abies in Lithuania and Poland. In: Przybylak, R., Majorowicz, J., Brázdil, R.,Kejna, M. (Eds.), The Polish Climate in the European Context: An HistoricalOverview, pp. 327e340.
Koprowski, M., Zielski, A., 2006. Dendrochronology of Norway spruce (Picea abies(L.) Karst.) from two range centres in lowland Poland. Trees-Structure andFunction 20, 383e390.
Koprowski, M., Zielski, A., 2008. Extremely narrow and wide tree rings in theNorway spruce (Picea abies (L.) Karst.) of the Bia1owie _za National Park.Ecological Questions 9, 73e78.
Ko _zuchowski, K., Degirmend�zi�c, J., 2005. Contemporary changes of climate inPoland: trends and variation in thermal and solar conditions related to plantvegetation. Polish Journal of Ecology 3, 283e297.
Larcher, W., 2003. Physiological Plant Ecology: Ecophysiology and Stress Physiologyof Functional Groups.
Lata1owa, M., van der Knaap, W.O., 2006. Late Quaternary expansion of Norwayspruce Picea abies (L.) Karst in Europe according to pollen data. QuaternaryScience Reviews 25, 2780e2805.
Mäkinen, H., Nojd, P., Mielikäinen, K., 2000. Climatic signal in annualgrowth variation of Norway spruce (Picea abies) along a transect from centralFinland to the Arctic timberline. Canadian Journal of Forest Research 30,769e777.
Mäkinen, H., Nöjd, P., Kahle, H.P., Neumann, U., Tveite, B., Mielikäinen, K., Röhle, H.,Spiecker, H., 2002. Radial growth variation of Norway spruce (Picea abies (L.)Karst.) across latitudinal and altitudinal gradients in central and northernEurope. Forest Ecology and Management 171, 243e259.
Mäkinen, H., Nöjd, P., Kahle, H.P., Neumann, U., Tveite, B., Mielikäinen, K., Röhle, H.,Spiecker, H., 2003. Large-scale climatic variability and radial increment varia-tion of Picea abies (L.) Karst.in central and northern Europe. Trees-Structure andFunction 17, 173e184.
Miina, J., 2000. Dependence of tree-ring, earlywood and latewood indices of Scotspine and Norway spruce on climatic factors in eastern Finland. EcologicalModelling 132, 259e273.
Misson, L., Nicault, A., Guiot, J., 2003. Effects of different thinning intensities ondrought response in Norway spruce (Picea abies (L.) Karst.). Forest Ecology andManagement 183, 47e60.
Mitscherlich, G., 1971. Wald, Wachstum und Umwelt. In: Waldklima und Wasser-haushalt, Bd. 2. Sauerländer`s Verlag, Frankfurt am Main.
Modrzy�nski, J., 1998. Zarys ekologii �swierka. In: Boraty�nski, A., Buga1a, W. (Eds.), Bio-logia�swierkapospolitego.BoguckiWydawnictwoNaukowe,Pozna�n,pp. 303e359.
Oko1owicz, W., Martyn, D., 1984. Regiony klimatyczne. In: Atlas Geograficzny Polski.PPWK, Warszawa.
Podbielkowski, Z., 1991. Geografia ro�slin. WSiP, Warszawa.Savva, Y., Oleksyn, J., Reich, P.B., Tjoelker, M., Vaganov, E.A., Modrzy�nski, J., 2006.
Interannual growth response of Norway spruce to climate along an altitudalgradient in the Tatra Mountains, Poland. Trees-Structure and Function 20,735e746.
Schweingruber, F.H., 1996. Tree Rings and Environment: Dendroecology. Birmens-dorf, Swiss Federal Institute for Forest, Snow and Landscape Research, Berne,Stuttgart, Vienna, Haupt.
�Srodo�n, A., 1967. Norway spruce in the Quaternary Poland. Acta Paleobotanica VIII.Nr 2. Kraków. Institute of Botany.
Vitas, A., 2002. Drought of 1992 in Lithuania and consequences to Norway spruce.Baltic Forestry 7, 25e30.
Vitas, A., 2004. Tree rings of Norway spruce (Picea abies (L.) Karsten) in Lithuania asdrought indicators: dendroecological approach. Polish Journal of Ecology 2,201e210.
Ward, J.H., 1963. Hierarchical grouping to optimize an objective function. Journal ofthe American Statistical Association 58, 236e244.
Wigley, T.M.L., Briffa, K.R., Jones, P.D., 1984. On the average value of correlated timeseries, with applications in dendroclimatology and hydrometeorology. Journalof Climate and Applied Meteorology 23, 201e213.
Wilson, R.J.S., Hopfmueller, M., 2001. Dendrochronological investigations of Norwayspruce along an elevational transect in the Bavarian Forest, Germany.Dendrochronologia 19, 67e79.
Zielski, A., Krapiec, M., 2004. Dendrochronologia. Wydawnictwo Naukowe PWN,Warszawa.




















