
Смирнова О.В., Чистякова А.А., Истомина И.И....
10
T. XLV, № 2 ЖУРНАЛ ОБЩЕ Й'БИОЛОГИИ 1984 УДК 581.524 : 575.3 КВАЗИСЕНИЛЬНОСТЬ КАК ОДНО ИЗ ПРОЯВЛЕНИЙ ФИТОЦЕНОТИЧЕСКОЙ ТОЛЕРАНТНОСТИ РАСТЕНИЙ О. В. СМИРНОВА, А. А. ЧИСТЯКОВА, И. И. ИСТОМИНА В статье рассмотрено одно из проявлений фитоценотической толерант-' ности видов — квазисенильность. Описаны конкретные проявления квазисе- нильности у растений разных жизненных форм. Показано, что переход в квазисенильное состояние часто представляет собой единственный способ сохранения ценопопуляции в фитоценотически неблагоприятных условиях. Явление толерантности широко известно в экологии (Schelford, 1913), Под толерантностью обычно понимаются экологические пределы суще- ствования растений и животных (Одум, 1975; Пианка, 1981). Эти эколо- гические пределы могут быть обусловлены абиотическими или биотиче- скими факторами. Одним из вариантов толерантности, прямо или кос- венно обусловленной взаимодействием растений в ценозе, мы считаем фитоценотическую толерантность. Ранее были показаны основные про- явления, характеризующие этот вариант толерантности у растений (Смирнова, 1980; Смирнова, Чистякова, 1980). К их числу относится ква- зисенильность. ■ Впервые представление о том, что растения, имеющие сенильный об- лик, при изменении условий существования могут давать генеративные побеги, было высказано Т. А. Работновым (1975). Ему же принадлежит этот термин. Квазисенильность можно определить как явление морфологической имитации сенильности, возникающее у растений в фитоценотически не- благоприятных условиях. Квазисенильные особи— это растения, спо- собные длительно находиться на низшем уровне жизненности и омола- живаться при улучшении условий существования. Проявления квазисенилы-юсти несколько различаются у растений разных жизненных форм. Е1иже это состояние описано на модельных ви- дах деревьев, кустарников, трав. Среди деревьев выбраны: дуб черешча- тый (Quercus robur L.), клены остролистный (Acer platanoides L.) и по- левой (A. campestre L.), ясень обыкновенный (Fraxinus excelsior L.), лип^ сердцевидная (Tilia cordata Mill.) и вяз шершавый (Ulmus scabra Mill.). Перечисленные виды деревьев переходят'в квазисенильное со- стояние из имматурного или виргинильного состояний в условиях край- не низкой освещенности. iyi деревьев выделено два варианта формы роста квазисенильных ра- стений: компактная и распростертая (Чистякова, 1978). Распростертые формы имеют значительные по протяженности (десятки сантиметров) плагиотропные подземные побеги — ксилоризомы. В подземной части у растений обеих форм роста хорошо развит ксилоподий (термин Lind- man, 1914). Это вертикальный утолщенный подземный орган, состоящий из разросшихся эпикотиля, гипокотиля и базальной части главного кор- ня. В надземной части квазисенильные растения имеют одну или не- сколько скелетных осей и «пеньки», образовавшиеся после отмирания осей. Эти структуры могут быть расположены непосредственно на кси- лоподии или ксилоризомах. Базальные участки, отмерших, а также ныне живущих скелетных осей и ксилоризомы несут на себе запас спящих по- чек. Последние дают начало периодически сменяющимся надземным ске- летным осям и новым ксилоризомам (рис. 1). Надземная часть обычно календарно моложе подземной, так как в условиях затенения каждая скелетная ось живет не более 15—20, иногз да 30 лет. Наиболее длительно она существует у дуба, граба, вяза, липы., Квазисенильные древесные растения обладают своеобразным изменени- 216
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Смирнова О.В., Чистякова А.А., Истомина И.И....

T. XLV, № 2 Ж У Р Н А Л О Б Щ Е Й ' Б И О Л О Г И И 1984
УДК 581.524 : 575.3
КВАЗИСЕНИЛЬНОСТЬ КАК ОДНО ИЗ ПРОЯВЛЕНИЙ ФИТОЦЕНОТИЧЕСКОЙ ТОЛЕРАНТНОСТИ РАСТЕНИЙ
О. В. СМИРНОВА, А. А. ЧИСТЯКОВА, И. И. ИСТОМИНА
В статье рассмотрено одно из проявлений фитоценотической толерант-' ности видов — квазисенильность. Описаны конкретные проявления квазисе- нильности у растений разных жизненных форм. Показано, что переход в квазисенильное состояние часто представляет собой единственный способ сохранения ценопопуляции в фитоценотически неблагоприятных условиях.
Явление толерантности широко известно в экологии (Schelford, 1913), Под толерантностью обычно понимаются экологические пределы существования растений и животных (Одум, 1975; Пианка, 1981). Эти экологические пределы могут быть обусловлены абиотическими или биотическими факторами. Одним из вариантов толерантности, прямо или косвенно обусловленной взаимодействием растений в ценозе, мы считаем фитоценотическую толерантность. Ранее были показаны основные проявления, характеризующие этот вариант толерантности у растений (Смирнова, 1980; Смирнова, Чистякова, 1980). К их числу относится квазисенильность. ■
Впервые представление о том, что растения, имеющие сенильный облик, при изменении условий существования могут давать генеративные побеги, было высказано Т. А. Работновым (1975). Ему же принадлежит этот термин.
Квазисенильность можно определить как явление морфологической имитации сенильности, возникающее у растений в фитоценотически неблагоприятных условиях. Квазисенильные особи— это растения, способные длительно находиться на низшем уровне жизненности и омолаживаться при улучшении условий существования.
Проявления квазисенилы-юсти несколько различаются у растений разных жизненных форм. Е1иже это состояние описано на модельных видах деревьев, кустарников, трав. Среди деревьев выбраны: дуб черешча- тый (Quercus robur L.), клены остролистный (Acer platanoides L.) и полевой (A. campestre L.), ясень обыкновенный (Fraxinus excelsior L.), лип^ сердцевидная (Tilia cordata Mill.) и вяз шершавый (Ulmus scabra Mill.). Перечисленные виды деревьев переходят'в квазисенильное состояние из имматурного или виргинильного состояний в условиях крайне низкой освещенности.
iyi деревьев выделено два варианта формы роста квазисенильных растений: компактная и распростертая (Чистякова, 1978). Распростертые формы имеют значительные по протяженности (десятки сантиметров) плагиотропные подземные побеги — ксилоризомы. В подземной части у растений обеих форм роста хорошо развит ксилоподий (термин Lind- man, 1914). Это вертикальный утолщенный подземный орган, состоящий из разросшихся эпикотиля, гипокотиля и базальной части главного корня. В надземной части квазисенильные растения имеют одну или несколько скелетных осей и «пеньки», образовавшиеся после отмирания осей. Эти структуры могут быть расположены непосредственно на кси- лоподии или ксилоризомах. Базальные участки, отмерших, а также ныне живущих скелетных осей и ксилоризомы несут на себе запас спящих почек. Последние дают начало периодически сменяющимся надземным скелетным осям и новым ксилоризомам (рис. 1).
Надземная часть обычно календарно моложе подземной, так как в условиях затенения каждая скелетная ось живет не более 15—20, иногз да 30 лет. Наиболее длительно она существует у дуба, граба, вяза, липы., Квазисенильные древесные растения обладают своеобразным изменени-
216

ем длин годичных приростов по главной оси (рис. 2). Максимальные размеры приростов характерны для первых лет после пробуждения спящей почки. Со временем они уменьшаются до минимальных (0,5— 2,0 см), и надземная ось отмирает либо целиком, либо частично. В последнем случае (рис. 2, б) нарастание носит волновой характер, так как при каждом пробуждении спящей почки первоначально формируется наибольший прирост, который в дальнейшем сменяется убывающими по величине. Таких «волн» роста за время существования скелетной оси может быть до шести у дуба, до четырех у граба, а у остальных видов> два-три.
Рис. 1. Компактное квазисенильное растение дуба («торчок»). 1 — скелетные оси, 2 — ксилоризом, 3 — придаточные корни, 4 — ксило-
подий, 5 — главный корень, х — отмирающие верхушки осей
217

В лесоводственной литературе квазисенильные растения компакт? ной формы известны под названием «торчков». Литературные данные о длительности жизни торчков имеются для дуба и клена остролистной (Михайлов, 1909; Гузовский, 1913; Морозов, 1918; Шеф, 1925; Пятницкий, 1949; Денисов, 1954; Мушинская, 1977). По этим данным максимальная длительность жизни торчков дуба— 15—25, клена остролистного — 25—30 лет.
Определение абсолютного возраста торчков на срезах через среднюю •часть ксилоподия (путем подсчета под микроскопом годичных колец з
древесине) показало, что продолжи* тельность их жизни у всех исследованных видов может достигать 50 и более лет. У некоторых деревьев, в частности у дуба и клена полевого, отмирание скелетной оси и развитие новой из спящей почки может быть отделено промежутком в несколько лет. В это время квазисенильное растение не имеет надземных побегов и находится в состоянии вторичного покоя (термин Т. А. Работнова, 1950). Подобное состояние многократно отмечалось исследователями у травянистых растений (Прозоровский, 1940; Работнов, 1950; Куркин, 1976; Мусина, 1-976)- Учитывая возможность перехода в состояние вторичного покоя, можно предположить, что абсолютный возраст отдельных квазисенильных особей больше числа годичных колец на срезе. Такое допущение вполне вероятно, так к'ак у угнетенных деревьев мы неоднократно наблюдали отсутствие вторичного утолщения в древесине в наиболее неблагоприятные годы.
Распростертые квазисенильные растения могут жить гораздо дольше компактных: их календарный возраст измеряется сотнями лет, если учитывать жизнь отдельных партикул, на которые способен разделяться ранее единый организм. Длительное пребывание в квазисенильном состоянии обеспечивает выживание особей деревьев семенного происхождения вплоть до момента вывала старых генеративных экземпляров, которые составляют верхние ярусы древостоя. Переход в квазисенильное состояние возможен у особей деревьев не только семенного, но и вегетативного происхождения (Чистякова, 1978).
Судьба квазисенильных особей различна: при улучшении световой обстановки отдельные скелетные оси начинают усиленно развиваться и растение переходит в генеративное состояние. Если же условия освещения остаются прежними, растение может отмереть целиком или же происходит отмирание отдельных скелетных осей и замена их новыми за счет развития из спящих почек.
Роль квазисенильных особей в ценопопуляциях всех видов широколиственных деревьев была исследована в дубово-грабовых лесах УССР, Изменение доли квазисенильных особей в пределах ареала изучалось у липы сердцевидной.
Доля квазисенильных особей в молодых частях ценопопуляций (проростки— виргинильные растения) всех видов широколиственных деревьев в дубово-грабовых лесах очень невелика (табл. 1). Однако, если рассчитать процентное участие квазисенильных особей в стабильной, уже прижившейся части потомства, которая включает имматурные и виргинильные растения, оно окажется существенным (1,2—18,8%).
Рис. 2. Динамика роста главных скелетных осей ясеня (а) и дуба (б) в квазисенильном состоянии (статистически достоверные усредненные данные). На, оси ординат — годичный
прирост, см
.218
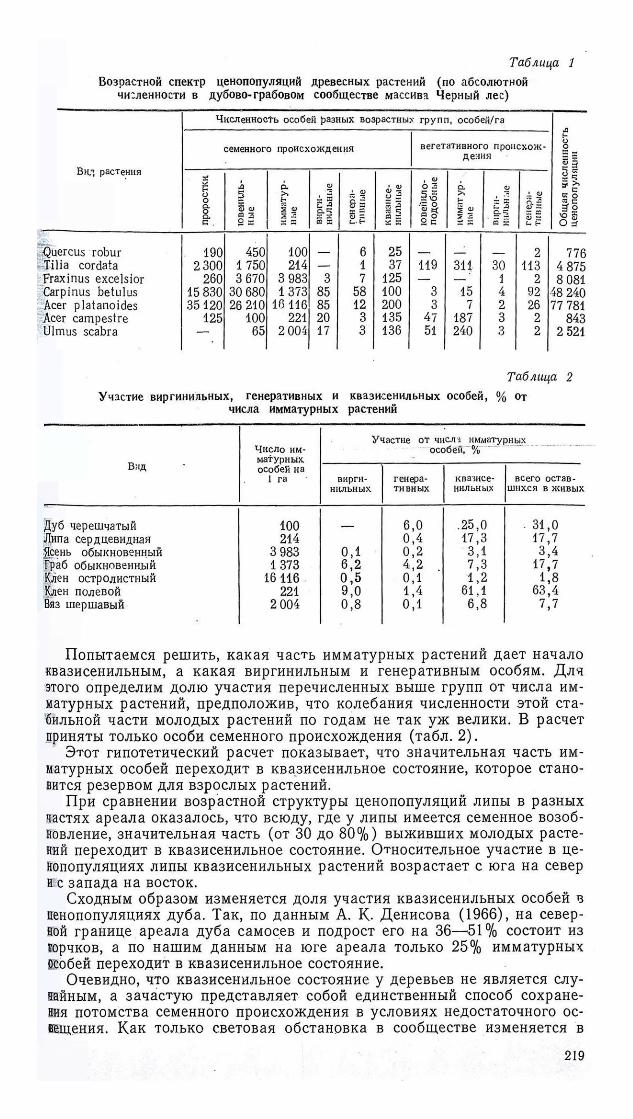
Таблица 1Возрастной спектр ценопопуляций древесных растений (по абсолютной
численности в дубово-грабовом сообществе массива Черный лес)
Вил растения
Численность особей разных возрастных групп, особей/га
Общ
ая ч
исле
ннос
ть
цено
попу
ляци
йсеменного происхождения вегетативного происхождения
прор
остк
и.
юве
ниль
ны
е
имма
тур-
иые
вирг
и-ни
льны
е
2_ ОIIЕ. ь кв
азпс
е-ни
льны
е
юве
нило
-по
добн
ые
имма
т ур -
ны
е
О
U 3С. 5 С2 Н
- S ic l £ о ~о £ и н
-Quercus robur 190 450 100 6 25 2 776Tilia cordata 2 300 1 750 214 — 1 37 119 311 30 ИЗ 4 875Fraxinus excelsior 260 3 670 3 983 3 7 125 — — ' 1 2 8 081Carpinus betulus 15 830 30 680 1373 85 58 100 3 15 4 92 48 240Acer platanoides 35 120 26 210 16 116 85 12 200 3 7 2 26 77 781Acer cam pest re 125 100 221 20 3 135 47 187 3 2 843Ulmus scabra — 65 2 004 17 3 136 51 240 ОО 2 2 521
Таблица 2Участие виргинильных, генеративных и квазисенильных особей, % от
числа имматурных растений
ВидЧисло имматурных особей на
1 га
Участие от числа имматурных особей,"%
виргинильных
генеративных
квазисенильных
всего оставшихся в живых
Дуб черешчатый 100 _ 6,0 .25,0 • 31,0Липа сердцевидная 214 0,4 17,3 17,7Ясень обыкновенный 3 983 0,1 0,2 3,1 3,4Граб обыкновенный 1373 6,2 4,2 . 7,3 17,7Клен остролистный 16 116 0,5 0,1 1,2 1,8Клен полевой 221 9,0 1,4 61.1 63,4Вяз шершавый 2 004 0,8 0,1 6,8 7,7
Попытаемся решить, какая часть имматурных растений дает начало квазисенильным, а какая виргинильным и генеративным особям. Для этого определим долю участия перечисленных выше групп от числа имматурных растений, предположив, что колебания численности этой стабильной части молодых растений по годам не так уж велики. В расчет приняты только особи семенного происхождения (табл. 2).
Этот гипотетический расчет показывает, что значительная часть имматурных особей переходит в квазисенильное состояние, которое становится резервом для взрослых растений.
При сравнении возрастной структуры ценопопуляций липы в разных пастях ареала оказалось, что всюду, где у липы имеется семенное возобновление, значительная часть (от 30 до 80%) выживших молодых растений переходит в квазисенильное состояние. Относительное участие в це- нопопуляциях липы квазисенильных растений возрастает с юга на север й с запада на восток.
Сходным образом изменяется доля участия квазисенильных особей в ненопопуляциях дуба. Так, по данным А. К. Денисова (1966), на северной границе ареала дуба самосев и подрост его на 36—61% состоит из юрчков, а по нашим данным на юге ареала только 25% имматурных Есобей переходит в квазисенильное состояние.
Очевидно, что квазисенильное состояние у деревьев не является случайным, а зачастую представляет собой единственный способ сохранения потомства семенного происхождения в условиях недостаточного освещения. Как только световая обстановка в сообществе изменяется в
219

лучшую сторону (вывал, вырубка), квазисенильные растения переходят в генеративное состояние, образуя одно- или многоствольные деревья.
Среди кустарников в качестве модельного вида выбрана лещина обыкновенная (Corylus avellana L.). Это обычный в широколиственных лесах геоксильный кустарник (Серебряков, 1962), достигающий максимального развития и нормально плодоносящий в условиях полного освещения. У лещины можно выделить два варианта жизненной формы. К первому относятся растения, у которых взрослые особи образуют компактные кусты. Надземная часть кустов состоит из большого числа многолетних (25—30 лет) скелетных осей, а подземная сформирована короткими (до 5 см) базальными участками живых и отмерших осей, несущих почки возобновления. Эти базальные участки в совокупности составляют короткое симподиальное корневище — ксилоризом. Корневая
система куста образована придаточными и главным корнем. Последний отмирает в начале генеративного периода. К концу онтогенеза единый куст распадается на партикулы вследствие разрушения ксилоризома.
Второй вариант жизненной формы представлен особями, у которых помимо короткого симподиального ксилоризома, начиная с имматурно- го возрастного состояния, образуются длинные гипогеогенные ксилоризо- мы (до 100 см). Из их верхушечных почек формируются надземные оси, а на их основе новые компактные кусты с несколько меньшим, чем у кустов семенного происхождения, числом скелетных осей и придаточной корневой системой. Дальнейшее развитие таких кустов происходит так же, как и у кустов семенного происхождения, т. е. онтогенез кустов вегетативного происхождения соответствует развитию особей, возникших из семян.
У лещины в квазисенильное состояние могут переходить как молодые, так и взрослые растения. Квазисенильные особи, возникшие на ранних этапах онтогенеза, могут быть как семенного, так и вегетативного происхождения. Первые из них полностью соответствуют «торчкам» у деревьев, а вторые представляют собой молодые по абсолютному возрасту кусты вегетативного происхождения с крайне пониженной жизненностью. Надземная часть последних состоит из одной-двух слабо ветвяг щихся малолетних осей с небольшими приростами, а в подземной части сохраняются остатки длинного ксилоризома и базальных частей отмерших малолетних осей (рис. 3). Квазисенильные особи, возникшие из
Рис. 3. Квазисенильное растение лещины до (а) и после (б) омоложения, вызванного улучшением световюй обстановки
220

Таблица 3Возрастной состав денопопуляции лещины обыкновенной в различных
условиях освещения
Численность особей разных возрастных групп на 1 га
<D335£оео
<03Яа>>КЗгг
О3 генеративные _• 1 Освещен-Условия
освещенияЕдиница
измеренияКVРооо.оо.с
XЛ5£Uо»£
оRоо2 ср
едне
воз
раст
ные
<1)3C LсаНо се
ниль
ные
\
кваз
исен
ши
ные
ность, % от
полной
Под ПОЛОГОМ особей/га . 3 3 39 3 18 144 375 0,9—1,5грабового % 0,513 0,513 5,667 0,513 — 3,077 24,615 64,102леса
В условиях час- особей/га — — 100 340 200 20 — 200 10— 20тичного освет- % — — 11,628 39,535 23,256 2,325 — 23,255ления
На вырубке особей/га%
— 202,041
202,041
10010,204
62063,256
22022,449 — — 100
взрослых растений, обычно невозможно отличить от истинно сенильных без эксперимента (природного или проводимого человеком). Как и сенильные, они имеют одну — три слаборазвитых скелетных оси в надземной части и полуразрушенный крупный разветвленный ксилоризом в подземной части. Однако при осветлении эти особи способны омолаживаться до ювенильного-виргинильного состояния, образуя хорошо развитые надземные скелетные оси с мощной системой придаточных корней. В течение нескольких лет после омоложения сохраняются остатки старого ксилоризома, что дает возможность определить происхождение таких особей.
Для того чтобы выяснить роль квазисенильных особей в ценопопу- лядиях лещины, был проведен анализ возрастного состава ценопопуля- ций на участках с разным световым режимом в пределах одного типа леса: дубо-грабинника снытево-осокового.
Под пологом дубово-грабового леса, где освещенность составляет Ю,9—1,5% от полной, в возрастном спектре денопопуляции лещины имеется один максимум на сенильных и квазисенильных особях (табл. 3). Это свидетельствует о том, что условия существования денопопуляции 'близки к пессимальным. При улучшении освещенности в полосе леса, которая граничит с девятилетней вырубкой и освещается за счет косых лучей солнца (средняя освещенность 10—20% от полной), максимум численности в денопопуляции лещины приходится на виргинильные и молодые генеративные растения. Однако детальный анализ подземных частей этих растений показывает, что все они возникли из квазисенильных особей. В их подземной части еще сохранились остатки крупных старых ксилоризомов и базальные участки надземных осей сенильного типа. Одновременно определение по анатомическим признакам возраста новых надземных осей и придаточных корней показывает, что возникли они через 1—2 года после рубки соседнего участка, которая привела к значительному улучшению световой обстановки под пологом леса.
На 10-летней вырубке в условиях 100%-ной освещенности максимум в возрастном спектре денопопуляции лещины приходится на молодые генеративные растения, а сенильные особи отсутствуют полностью. Таким образом, рассмотренный ряд демонстрирует особенности развития ценопопуляций лещины в лесных ценозах. Определение возрастного состава денопопулядий этого вида в ценозах с различным режимом освещенности показывает, что пополнение взрослой части денопопуляции при улучшении освещенности происходит почти исключительно за счет квазисенильных особей. Семенное возобновление при этом не играет заметной роли.
Среди травянистых растений выбрано три модельных вида: чистяк весенний (Ficaria verna Huds.), медвежий лук (Allium ursinum L.) и
221

сныть обыкновенная (Aegopodium podagraria L.), различающихся по-, особенностям пространственной структуры взрослых особей и времени возникновения в онтогенезе квазисенильных растений.
Чистяк весенний — многолетнее травянистое олигокарпическое кистекорневое растение. Часть придаточных корней выполняет запасающую- функцию. Структурным элементом побеговой части взрослой особи является полурозеточный монокарпический, моноциклический побег. Сенильные растения имеют небольшой розеточный побег с одним-двумя ассимилирующими листьями ювенильного типа и разрушающимся старым корневищем, на котором еще сохранились крупные запасающие корни прошлых лет и следы от генеративных побегов.
Один из способов вегетативного размножения этого вида состоит в- образовании ювенильноподобных особей из придаточных почек на запасающих корнях. Такой способ вегетативного размножения приводит к образованию компактных клонов. Помимо молодых растений в клонах чистяка можно найти растения небольшого условного возраста, у которых, в отличие от молодых растений, не происходит ежегодного увеличения числа и размеров ассимилирующих листьев, запасающих и всасывающих корней. В отличие же от старых растений у них нет следов генеративных побегов и остатков корневища. Кроме того, у них появляются новые структуры — столоны «убегания» длиной 1—2 см. Такие особи мы относим к квазисенильным. Судьба их различна. Если длина столона оказывается недостаточной для того, чтобы верхушечная почка растения и новые придаточные корни оказались вне компактного клона, растение умирает. Если верхушка побега укореняется вне клона, _р_ас-- тение продолжает развиваться и переходит к цветению. Таким образом,, у чистяка квазисенильные особи возникают на ранних этапах онтогенеза и в этом отношении могут быть сопоставлены с «торчками» древесных растений.
Медвежий лук — многолетнее травянистое поликарпическое луковичное растение с ежегодно сменяющимися симподиальными двуосными: луковицами. Вегетативный малолетник (Старостенкова, 1978). Вегетативное размножение у этого вида начинается в конце виргинильиого — в молодом генеративном состояниях и не сопровождается вегетативным разрастанием, что приводит к образованию сверхкомпактных клонов. В этих клонах образуются, как и у чистяка, квазисенильные особи со столонами «убегания», но формируются они не из молодых, а из взрослых особей. Образование квазисенильных особей со столонами «убегания» обнаружено также у Allium victorialis, Ranunculus cassubicus, Des- champsia flexuosa. Причем, в отличие от чистяка, у перечисленных видов в квазисенильное состояние переходят генеративные особи. Как показывают определения, доля квазисенильных особей в возрастных спект: рах ценопопуляций Ficaria verna, Allium ursinum невелика (3—7%), однако именно их наличие определяет возможность циклического развития компактных клонов этих видов по типу: молодость — зрелость — ста^рость — омоложение. Омоложение в них происходит за счет большого резерва (в некоторых клонах до 60—70%) квазисенильных особей, которые после отмирания части старых и взрослых особей претерпевают омоложение.
Таким образом, описанная выше квазисенильность определяется особенностями пространственной структуры ценопопуляций, т. е. эндогенным по отношению к ценопопуляций фактором. В данном случае она является следствием угнетающего воздействия разрушающихся частей материнских особей на дочерние.
Сныть обыкновенная — многолетнее травянистое длиннокорневищное поликарпическое растение. Взрослые генеративные растения состоят из нескольких парциальных монокарпических полициклических полурозе- точных побегов с придаточной корневой системой, соединенных гипогео- генными корневищами (Смирнова, 1067). Сенильные особи состоят из одного парциального побега с разрушающимся корневищем и розеточ-
222

ным побегом ювенильного типа. Н овообразование придаточных корней Практически не происходит.
Помимо описанных выше сенильных особей в ценопопулядиях можно обнаружить растения, у которых из спящих пазушных почек парциальных побегов сенильного облика развиваются гипогеогенные корневища, дающие начала системам парциальных побегов взрослого типа (рис. 4). Такие растения рассматриваются нами как квазисенильные. Решить, что представляет собой каждое конкретное растение сныти, имеющее сенильный облик: собственно сенильное растение или квази- сенильное,— в настоящее время не представляется возможным без эксперимента (природного или антропогенного).
Рис. 4. Выход сныти обыкновенной из квазисенильного состояния: qs — квазисенильный,v — виргинильный, парциальные побеги
Рис. 5. Схема циклического развития ценопопуляций сныти обыкновенной в возрастном ряду древостоев. Возраст насаждения и освещенность: 1 — 20 лет, 1—2%; 2 — 40 лет, 3—5%; 3 — 90 лет, 4—6%; 4 — 180—220 лет, 8 — 12(20)%. Численность парциальных побегов: а — от проростков до средневозрастных генеративных; б — от старых генера
тивных до сенильных и квазисенильных.
Наиболее четко роль квазисенильных растений в ценопопулядиях сныти проявляется в возрастных рядах древостоев, формирующихся после сплошных рубок. Определяющим экологическим фактором здесь является существенное изменение режима освещенности. Описание изменений возрастного состава и численности ценопопуляций сныти в возрастном ряду широколиственных ценозов (Смирнова, 1977) показывает, что под пологом старовозрастного естественного насаждения, где освещенность достигает 7—10%, общая численность парциальных побегов — 56,64 шт/м2, а доля «субсенильных» и «сенильных» парциальных побегов (часть из которых является квазисенильными) составляет 12,7%. На вырубках 1—3 лет при полной освещенности суммарная численность побегов сныти увеличивается в 6 раз, а численность «сенильных» падает до 2,1%. После смыкания полога нового поколения леса освещенность на уровне травяного покрова снижается до 1—2%, общая яисленность ценопопуляций сныти резко уменьшается, а господство переходит к «сенильным» парциальным побегам (рис. 5). В дальнейшем по мере взросления нового поколения леса освещенность увеличивается, параллельно возрастает численность взрослых побегов, достигая максимума к 180—250 годам, а численность «сенильных» падает. Полное отсутствие в рассматриваемых ценопопулядиях молодых растений семенного происхождения приводит к заключению, что переход господствующего положения от «сенильных» парциальных побегов к взрослым гене- Еативным объясняется тем, что большая часть «сенильных» побегов яв-
223

.ляются квазисенильными. Определяющее влияние освещенности на переход квазисенильных побегов в генеративные удалось пронаблюдать в 40-летнем насаждении дуба, где проводились опыты по прореживанию древостоя. На контроле, где сомкнутость крон дуба была 0,8—0,9, в возрастном спектре ценопопуляции сныти господствовали сенильные побеги, на опытных участках с сомкнутостью крон 0,5—0,6 — генеративные.
Анализ литературы показывает, что не только недостаток света определяет переход части побегов сныти в квазисенильное состояние. Так, по данным В. Г. Карпова (1969), это происходит в южной тайге и при острой нехватке элементов минерального питания, в первую очередь азота. Тот же вывод можно сделать из работы В. Н. Сукачева (1941). Описанные Н. А. Борисовой (1966) изменения ценопопуляций ландыша в сукцессионных рядах ельников заставляют предполагать наличие у этого вида квазисенильных особей.
Таким образом, сравнение видов разных жизненных форм заставляет предполагать существенную роль явления квазисенильности в жизни растений. Квазисенильность вызывается эндо- или экзогенными по отношению к ценопопуляции факторами. Среди эндогенных факторов ведущая роль принадлежит первичной пространственной структуре ценопопуляции, определяемой биологией вида. Среди экзогенных факторов, определяющих переход растений в квазисенильное состояние, к настоящему времени удалось наиболее четко выяснить роль низкого освещения и, в меньшей степени, конкуренции за элементы минерального питания.
Для видов, имеющих компактные клоны, переход в квазисенильное состояние определяется как эндогенными, так и экзогенными по отношению к ценопопуляции причинами; для видов, образующих диффузные клоны — только экзогенными. Эндогенная квазисенильность в компактных клонах — это следствие угнетающего воздействия разрушающихся частей материнских особей на дочерние. В ценопопуляциях, первичная агрегированность которых определяется наличием компактных клонов, появление квазисенильных особей отмечается довольно часто. Переход части особей в квазисенильное состояние в компактных клонах дает возможность им выжить, а после гибели большинства особей омолодиться и вновь занять территорию вегетативным или семенным путем. Таким образом, переход в квазисенильное состояние является одной из причин циклического развития компактных клонов по типу: молодость—'Зрелость — старость — омоложение и дает возможность неопределенно долгого удержания ценопопуляцией занятого места в ценозе.
В диффузных клонах эта сторона квазисенильности практически не проявляется, так как в них происходит постоянное убегание молодых особей или побегов (у полицентрических особей) от взрослых и старых.
Вне зависимости от времени возникновения в онтогенезе общим для квазисенильных особей является все усиливающееся преобладание процессов старения и отмирания над новообразованием. Это выражается в уменьшении длины приростов скелетных осей у древесных растений, в уменьшении размеров и числа листьев и корней у трав.
Биологический смысл квазисенильности — в способности растений длительно существовать на предельно возможном низком уровне жизненности, с отрицательным балансом органического вещества и минимальными затратами накопленных ранее пластических веществ. В этом отношении переход в квазисенильное состояние подобен переходу в состояние вторичного покоя. Оба эти состояния представляют собой проявления фитоценотической толерантности видов, позволяющей им неопределенно долго сохраняться в фитоценозах на низком уровне численности и жизненности и резко увеличивать их при нарушениях ценотиче- ской замкнутости.
Полученные результаты дают основание заново пересмотреть данные многих ценопопуляционных исследований, где показана существенная роль старых особей в ценопопуляциях. Вероятно, необходимо отличать истинную онтогенетическую старость, от «старости» ценотической, которая представляет собой не что иное, как квазисенильность.
224

ЛИТЕРАТУРАБорисова Н. А. Некоторые механизмы равновесия в лесных биогеоценозах.— Тр. Ле-
нингр. лесотехн. акад. им. С. М. Кирова, 1966, вып. 104, с. 29.Гузовский Б. И. Казанские нагорные дубравы.— Лесной ж., 1913, вып. 1—2, с. 5. Денисов А. К. Пойменные дубравы лесной зоны. М.— Л.: Гослесбумиздат, 1954. 211 с. Денисов А. К. Дубовые леса Севера. Автореф. дис. на соискание уч. ст. докт. биол.
наук. Красноярск: Ин-т леса АН СССР, 1966. 38 с.Карпов В. Г. Экспериментальная фитоценология темнохвойной тайги. Л.: Наука, 1969,
335 с.Куркин К. А. Системные исследования динамики лугов. М.: Наука, 1976. 284 с. Михайлов Н. А. Естественное и искусственное возобновление дубрав лесостепной зо
ны.— Докл. XI съезду лесохозяев в Туле, 1909, с. 17.Морозов Г. Ф. К вопросу о возобновлении дубрав.— Лесной ж., 1918, с. 383.Мусина Л. С. Побегообразование и становление жизненной формы некоторых розет
кообразующих трав.— Бюл. Московск. о-ва испыт. природы, 1976, т. 81, вып. 6, с. 123.
Мушинская Н. И. Экология семенного размножения клена остролистного и липы мелколистной в Башкирии. Автореф. дис. на соискание уч. ст. канд. биол. наук. Свердловск: Ин-т экологии раст. и животн. Уральск, фил. АН СССР, 1977, 34 с.
Одум Ю. Экология растений. М.: Мир, 1975. 740 с.Пианка Э. Эволюционная экология. М.: Мир, 1981. 396 с.Прозоровский Н. А. Наблюдения над осенним и зимним состоянием растений.— Тр.
Центр.-Черноземн. заповеди., 1940, вып. 1, с. 273.Пятницкий С. С. Самосев дуба в качестве посадочного материала.— Лес и степь, 1949,
№ 1, с. 85.Работное Т. А. Жизненный цикл многолетних травянистых растений в луговых цено
зах.— Тр. Ботан. ин-та АН СССР. Сер. 3. Геоботаника, 1950, вып. 6, с. 7. Работное Т. А. Изучение ценотических популяций в целях выяснения стратегии жизни
видов растений.— Бюл. МОИП, 1975, т. 80, вып. 2, с. 5.Серебряков И. Г. Экологическая морфология растений. М.: Сов. школа, 1962. 378 с. Смирнова О. В. Онтогенез и возрастные группы осоки волосистой и сныти обыкновен
ной.— В кн.: Онтогенез и возрастной состав популяций цветковых растений. М.: Наука, 1967, с. 100.
Смирнова О. В. Динамика ценопопуляций на протяжении интервалов времени разного порядка на примере сныти.— В кн.: Ценопопуляции растений. Развитие и взаимоотношения. М.: Наука, 1977, с. 57.
Смирнова О. В. Поведение видов и функциональная организация травяного покрова широколиственных лесов европейской части СССР.— Бюл. Московск. о-ва испыт. природы, 1980, т. 85, вып. 5, с. 53.
Смирнова О. В., Чистякова А. А. Анализ фитоценотических потенций некоторых древесных видов широколиственных лесов европейской части СССР.— Ж. общ. биол., 1980, т. 41, № 3, с. 350.
Старостенкова М. М. Лук медвежий.— Биол. флора Московск. обл., 1978, вып. 4, с. 52. Сукачев В. Н. О влиянии интенсивности борьбы за существование между растениями
на их развитие.— Докл. АН СССР, 1941, т. 30, № 8, с. 752.Чистякова А. А. О жизненной форме и вегетативном разрастании липы сердцевидной.—
Бюл. Московск. о-ва испыт. природы, 1978, т. 83, вып. 2, с. 129.Шеф М. Д. Естественное возобновление главной породы в Казанских дубравах.— Изв.
Казанск. ин-та с. х. и лесоводства, 1925, вып. 2, с. 46.Lindman С. А. М. Nagra bidrag till fragan: buske eller trod.— In: Kungl. Svenska Ve-
tenskapsakademichs Arskok. Stokholm, 1914, S. 231.Shelford V. E. The reaction of certain animals to gradient of evaporation power and air.
A study in experimental ecology.— Biol. Bull., 1913, v. 25, p. 79.Московский государственный Поступила в редакциюпедагогический институт 6.1.1982
QUAS1SENII4ITY AS ONE OF MANIFESTATIONS OF PHYTOCOENOTIC TOLERANCE OF THE PLANTS
о. V. SMIRNOVA, A. A. CHISTYAKOVA,- I . I. ISTOMINA
V. I. Lenin State Pedagogical Institute of Moscow
Summary
Quasisenility is a morphological imitation of senility arising in plants under phyto- coenotically unfavourable conditions. The capacity of transition in a quasisenile state manifests itself in plants belonging to different life forms and at different ontogenetic stages. The biological sense of quasisenility consists in the ability of plants to exist for a long time at the lowest possible level of activity. In this respect the transition into a quasisenile state is similar to the transition in a state of secondary rest. Quasisenility is one of possible (in some cases a single) ways of preservation of a coenopula- tion under phytocoenotically. extremely unfavourable conditions. Quasisenility can be considered as one of the mechanisms of succession processes.
6 Журнал общей биологии, № 2 225




















