
Simulating the effects of reforestation on a large catastrophic fire burned landscape in...
12
Simulating the effects of reforestation on a large catastrophic fire burned landscape in Northeastern China Xugao Wang a,c, * , Hong S. He b , Xiuzhen Li a , Yu Chang a , Yuanman Hu a , Chonggang Xu a,c , Rencang Bu a , Fuju Xie a,c a Institute of Applied Ecology, Chinese Academy of Science, P.O. Box 417, Shenyang 110016, China b School of Natural Resources, University of Missouri, Columbia c Graduate School of Chinese Academy of Science, Beijing 100039, China Received 20 May 2005; received in revised form 2 November 2005; accepted 16 December 2005 Abstract We use the LANDIS model to study the effects of planting intensity and spatial pattern of plantation on the abundance of three main species (larch (Larix gmelini), Mongolian Scotch pine (Pinus sylvestris var. Mongolica), and white birch (Betula platyphylla)) in the Tuqiang Forest Bureau on the northern slopes of Great Hing’an Mountains after a catastrophic fire in 1987. Four levels of planting intensity (covering 10%, 30%, 50%, and 70% of the severely burned area) and two spatial patterns of plantation (dispersed planting and aggregated planting) were compared in a 4 2 factorial design over a 300-year period. The results showed that increasing planting intensity positively influenced larch and Mongolian Scotch pine abundance, but negatively influenced white birch abundance. However, the increased degree of larch abundance with increasing planting intensity was significantly different between intensities. The difference in larch abundance between the 10% planting intensity scenario and the 30% planting intensity scenario was greater than that between the 50% planting intensity scenario and the 70% planting intensity scenario. However, the difference between 30% and 50% planting intensity scenarios was significantly low. Hence, given considerable labor input and economic costs, 30% planting intensity would be effective for forest recovery. In addition, dispersed planting showed more promising results on forest recovery than aggregated planting. However, the difference of larch abundance between dispersed planting and aggregated planting under intermediate planting intensity scenarios (30% and 50% planting intensity) was greater than that under a low planting intensity scenario and a high planting intensity scenario. Therefore, it is necessary to incorporate spatial pattern of plantation into planting practice, especially under an intermediate planting intensity scenario. These results have important implications for forest managers to design sound forest restoration projects for landscapes affected by large infrequent disturbances. In particular, the results suggest that the current planting strategy (50% planting intensity with aggregated planting) employed after the catastrophic fire in 1987 could not be optimum, and the dispersed planting strategy covering about 30% of the severely burned area would better stimulate forest recovery. # 2005 Elsevier B.V. All rights reserved. Keywords: Great Hing’an Mountains; LANDIS; Planting intensity; Spatial pattern of plantation; Forest recovery 1. Introduction The deciduous and coniferous forests of the Great Hing’an Mountains in northeastern China provide the most timber of any forested area in the country; simultaneously, this area encompasses rather unique ecological and environmental systems in the region (Zhou, 1991; Xu, 1998). Human activities, particularly timber harvesting, have substantially altered the spatial pattern and ecological functions of these systems. Decades of fire suppression have reduced fire size, prolonged the fire return interval (i.e., the number of years between two successive fire events for a specific area), and indirectly influenced forest composition and dynamics (Shu et al., 1996). The success of fire suppression, coupled with a warmer, drier climate due to global warming (Xu, 1998), have led to a fuel buildup and resulted in fires of greater intensity and extent than those that occurred previously in the region. Catastrophic fires can have disastrous effects on forest composition and structure, ecosystem processes, and landscape pattern (Romme, 1982; Turner et al., 1997, 1999). On May 6, 1987, a catastrophic fire occurred on the northern slopes of Great Hing’an Mountains, burning a total area of 1.3 10 6 ha. www.elsevier.com/locate/foreco Forest Ecology and Management 225 (2006) 82–93 * Corresponding author. Tel.: +86 24 83970350; fax: +86 24 83970351. E-mail addresses: [email protected], [email protected] (X. Wang). 0378-1127/$ – see front matter # 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.foreco.2005.12.029
Transcript of Simulating the effects of reforestation on a large catastrophic fire burned landscape in...

www.elsevier.com/locate/foreco
Forest Ecology and Management 225 (2006) 82–93
Simulating the effects of reforestation on a large catastrophic fire
burned landscape in Northeastern China
Xugao Wang a,c,*, Hong S. He b, Xiuzhen Li a, Yu Chang a, Yuanman Hu a,Chonggang Xu a,c, Rencang Bu a, Fuju Xie a,c
a Institute of Applied Ecology, Chinese Academy of Science, P.O. Box 417, Shenyang 110016, Chinab School of Natural Resources, University of Missouri, Columbia
c Graduate School of Chinese Academy of Science, Beijing 100039, China
Received 20 May 2005; received in revised form 2 November 2005; accepted 16 December 2005
Abstract
We use the LANDIS model to study the effects of planting intensity and spatial pattern of plantation on the abundance of three main species
(larch (Larix gmelini), Mongolian Scotch pine (Pinus sylvestris var. Mongolica), and white birch (Betula platyphylla)) in the Tuqiang Forest
Bureau on the northern slopes of Great Hing’an Mountains after a catastrophic fire in 1987. Four levels of planting intensity (covering 10%, 30%,
50%, and 70% of the severely burned area) and two spatial patterns of plantation (dispersed planting and aggregated planting) were compared in a
4 � 2 factorial design over a 300-year period. The results showed that increasing planting intensity positively influenced larch and Mongolian
Scotch pine abundance, but negatively influenced white birch abundance. However, the increased degree of larch abundance with increasing
planting intensity was significantly different between intensities. The difference in larch abundance between the 10% planting intensity scenario
and the 30% planting intensity scenario was greater than that between the 50% planting intensity scenario and the 70% planting intensity scenario.
However, the difference between 30% and 50% planting intensity scenarios was significantly low. Hence, given considerable labor input and
economic costs, 30% planting intensity would be effective for forest recovery. In addition, dispersed planting showed more promising results on
forest recovery than aggregated planting. However, the difference of larch abundance between dispersed planting and aggregated planting under
intermediate planting intensity scenarios (30% and 50% planting intensity) was greater than that under a low planting intensity scenario and a high
planting intensity scenario. Therefore, it is necessary to incorporate spatial pattern of plantation into planting practice, especially under an
intermediate planting intensity scenario. These results have important implications for forest managers to design sound forest restoration projects
for landscapes affected by large infrequent disturbances. In particular, the results suggest that the current planting strategy (50% planting intensity
with aggregated planting) employed after the catastrophic fire in 1987 could not be optimum, and the dispersed planting strategy covering about
30% of the severely burned area would better stimulate forest recovery.
# 2005 Elsevier B.V. All rights reserved.
Keywords: Great Hing’an Mountains; LANDIS; Planting intensity; Spatial pattern of plantation; Forest recovery
1. Introduction
The deciduous and coniferous forests of the Great Hing’an
Mountains in northeastern China provide the most timber of
any forested area in the country; simultaneously, this area
encompasses rather unique ecological and environmental
systems in the region (Zhou, 1991; Xu, 1998). Human
activities, particularly timber harvesting, have substantially
altered the spatial pattern and ecological functions of these
* Corresponding author. Tel.: +86 24 83970350; fax: +86 24 83970351.
E-mail addresses: [email protected], [email protected]
(X. Wang).
0378-1127/$ – see front matter # 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.foreco.2005.12.029
systems. Decades of fire suppression have reduced fire size,
prolonged the fire return interval (i.e., the number of years
between two successive fire events for a specific area), and
indirectly influenced forest composition and dynamics (Shu
et al., 1996). The success of fire suppression, coupled with a
warmer, drier climate due to global warming (Xu, 1998), have
led to a fuel buildup and resulted in fires of greater intensity and
extent than those that occurred previously in the region.
Catastrophic fires can have disastrous effects on forest
composition and structure, ecosystem processes, and landscape
pattern (Romme, 1982; Turner et al., 1997, 1999). On May 6,
1987, a catastrophic fire occurred on the northern slopes of
Great Hing’an Mountains, burning a total area of 1.3 � 106 ha.

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–93 83
Fig. 1. The location of Tuqiang Forest Bureau and burn severities after the
catastrophic fire in 1987 (1, unburned area (MA 1); 2, severely burned area
where conifers were dominant before the fire (MA 2); 3, severely burned area
where conifers were not dominant before the fire (MA 3); 4, other burned area
(MA 4)).
This immense, high-intensity fire consumed vegetation cover
and precipitated the exposition of mineral soils – as well as the
subsequent erosion and runoff – during post-fire rain events
(Xiao et al., 1988; Shu et al., 1996). Forest recovery in such
vastly burned areas is challenging because the long-term
landscape-level vegetation dynamic in a forest landscape is
complicated by spatial and temporal interactions among
multiple ecological and anthropogenic processes.
In many cases, natural succession can eventually lead to
post-fire recovery. This is especially true for cases in which
there are sufficient residual forests remaining nearby to act as
seed sources (Turner et al., 1999; Borchert et al., 2003). In cases
such as the 1987 fire in the Great Hing’an Mountains, natural
recovery is difficult because the severely burned area is
extensive, the burn severities are high, and the seed sources are
far removed (Xiao et al., 1988). In these situations, the process
of vegetation recovery is slow, increasing the risk of soil erosion
and environmental degradation. Thus, ecological restoration
through human mediation is necessary. After the 1987 fire,
forest management in this region shifted from timber harvesting
to reforestation – particularly in the severely burned area – in
order to accelerate forest restoration.
Various approaches have been developed to restore forest
vegetations for degraded systems where natural recovery is
unlikely. Grass seeding provides quick, temporary vegetation
ground cover; these are typically annuals or short-lived
perennials that can hold soil (Beyers, 2004). However, such
a treatment does not facilitate long-term ecosystem restoration.
Long-term ecosystem restorations are accomplished either
through the planting of a small number of early successional
nursery trees or shrubs to create habitats for seed-dispersing
birds (e.g., Lamb, 1998), or through high-density plantation of
tree species once present in the disturbed areas (e.g., Smale
et al., 2001; Nagashima et al., 2002).
Although these approaches have shown promising results for
development of forest structure, increase of species richness,
and recovery of natural successional processes, they are often
limited to relatively small areas (Lamb, 1998). In the Great
Hing’an Mountains, it is not sufficient to restore only a few
burned areas, and it is highly impractical to plant trees in the
vast severely burned area due to the considerable labor and
economic resources required. It is therefore necessary to assess
the planting intensity (the proportion of the burned area for
reforestation) and spatial patterns of plantation (the spatial
allocation of reforestation in the field) for forest recovery. In the
Great Hing’an Mountains, the planting strategy after the
catastrophic fire in 1987 was to plant coniferous seedlings in an
aggregated fashion over about 50% of the severely burned area
where coniferous forests used to grow. Others such as dispersed
planting, an alternative to the aggregated planting, have not
been evaluated and compared. A dispersed planting would
entail planting trees in a random fashion across the entire
severely burned area.
Descriptive studies and field experiments are often
inadequate sources of input for managers developing and
implementing reforestation plans. Comparing ecological
restoration strategies on a large-scale landscape is often
beyond the limits of traditional or experimental studies. Models
have therefore become an important tool for predicting the
effects of alternative management options. Landscape models
are particularly important in this study because other types of
models, such as gap models and ecosystem process models, are
often limited in spatial extents (He et al., 2002b; Mladenoff,
2004). While other anthropogenic disturbances – such as fire
suppression and forest harvesting – have been studied using
landscape models (Gustafson et al., 2000; Franklin et al., 2001;
Sturatevant et al., 2004; Wimberly, 2004), the effects of
reforestation on long-term forest dynamics has not been
explicitly simulated.
The purpose of this research is to study the effects of
different reforestation scenarios on vegetation dynamics in the
severely burned area of the Great Hing’an Mountains. We will
examine the effects of planting intensities and spatial patterns
of plantation (dispersed vs. aggregated) on forest landscape
recovery. We will apply a factorial design of planting intensity
and spatial pattern of plantation on the realistically para-
meterized forest composition maps of 1987 to identify the
combinations of planting intensity and method that could best
accelerate forest restoration. Understanding the probable post-
fire dynamics of the region under different reforestation
scenarios will not only provide insights into landscape scale
processes, but also will provide baseline information on forest
landscape restoration in northeast China after catastrophic fire
disturbances.
2. Study area
The Tuqiang Forest Bureau (Fig. 1), encompassing
approximately 4 � 105 ha on the northern slopes of Great
Hing’an Mountains, is in the Mohe County of Heilongjiang
province in northeast China (from 5281505500 to 5383304000N,
and 12281800500 to 12382900000E). It borders Russia to the north
(separated by Heilongjiang River), the Xilinji Forest Bureau to
the west, the Amur Forest Bureau to the east, and the Inner
Mongolia Autonomous Region to the south. The area has a
cold, continental climate, with an average annual temperature

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–9384
at �5 8C. Monthly mean temperature ranges from �47.2 8C in
January to 31.4 8C in July. The average annual precipitation is
432 mm, with great inter-annual variations. Seventy-five
percent of the rainfall occurs between June and August.
Uplands and small hills characterize this region, though it
possesses a relatively smooth topography. Slopes are generally
less than 158; the maximum slope is less than 458. Hills
undulate throughout this area, and the mountain ranges mostly
run in north and south directions. Elevations range from 270 m
to 1210 m; the mean elevation is 500 m. Brown coniferous
forest soil is representative in the Bureau. Vegetation is
dominated by larch (L. gmelini) forests. White birch (Betula
platyphylla) is the major broad-leaved species in the region. In
addition to larch and white birch, the tree species include
Mongolian Scotch pine (P. sylvestris var. mongolica), spruce
(Picea koraiensis), aspen-D (Populus davidiana), black birch
(Betula. davurica), aspen-S (Populus suaveolens), and willow
(Chosenia arbutifolia).
On May 6, 1987, a catastrophic forest fire of 1.33 � 106 ha
took place on the northern slopes of the Great Hing’an
Mountains. The burned area covered 2.31 � 105 ha in the
Bureau, and the severely burned area covered roughly
9 � 104 ha (Fig. 2). The conflagration caused incredible
damage to Tuqiang Forest Bureau, and this has led to great
difficulty in restoring the forest ecosystems.
Fig. 2. Landtype maps of Tuqiang Forest Bureau.
3. Methods
3.1. Description of LANDIS
LANDIS is a landscape disturbance and succession model
that facilitates the study of the effects of natural and
anthropogenic disturbances on forest landscapes, and has been
described extensively elsewhere (Mladenoff and He, 1999; He
and Mladenoff, 1999; He et al., 1999; Gustafson et al., 2000).
Here we provide a general description of the model. In
LANDIS, a landscape is organized as grid of cells, with
vegetation information stored as attributes for each cell. Cell
size can be varied from 10 m to 500 m depending on the
research scale. At each cell, the model tracks a matrix
containing a list of species by rows and the 10-year age cohorts
by columns. The model does not track individual trees. This
differs from most from stand simulation models than track
individual trees (Grimm, 1999). Additionally, computational
loads are greatly reduced, because actual species abundance,
biomass, or density is not calculated. A species presence/
absence approach allows LANDIS to simulate large landscapes
and avoids any false precision of predicting species abundance
measures with inadequate input data or parameter information.
LANDIS stratifies a heterogeneous landscape into land
types, which are generated from GIS layers of climate, soil, or
terrain attributes (slope, aspect, and landscape position). It is
assumed that a single land type contains a somewhat uniform
suite of ecological conditions, resulting in similar species
establishment patterns and fire disturbances characteristic,
including ignition frequency, mean fire return interval, and fuel
decomposition rated (He and Mladenoff, 1999). These
assumptions have been supported by many empirical and
experimental studies (e.g., Brown and See, 1981; Kauffman
et al., 1988). Furthermore, land types can be redefined by users
to partition the landscape into strata that are most relevant for a
particular application.
Four spatial processes and numerous non-spatial processes
are simulated by LANDIS. The spatial processes are fire,
windthrow, harvesting, and seed dispersal. Fire is stochastic
process based on the probability distributions of fire cycle and
mean fire sizes for various land types (He and Mladenoff,
1999). LANDIS simulates fire levels of fire intensity from
surface fire to crown fires. Fire intensity is determined by the
amount of fuel on a site. Tree species are also grouped into fire
fire-tolerance classes based on their fire-tolerance attributes and
five age-based fire susceptibility classes from young to old,
with young trees being more susceptible to damage than older
trees. Thus, fire severity is the interaction of susceptibility
based on species age classes, species fire tolerance and fire
intensity. A new LANDIS fire module (Yang et al., 2004)
employs hierarchical probability theory to allow even more
explicit simulation of different fire regimes across landscapes.
Windthrow is also stochastically simulated. In the LANDIS
wind module the probability of windthrow mortality increases
with tree age and size. Windthrow events interact with fire
disturbance such that windthrow increases the potential fire
intensity class at a site due to increased fuel load.

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–93 85
The LANDIS harvest module simulates forest harvesting
activities based on management area and stand boundaries
(Gustafson et al., 2000). These maps are predefined and are only
used by the LANDIS harvest and fuel modules. Harvest activities
are specified through rules relative to spatial, temporal, and
species age-cohort information tracked in LANDIS. The spatial
component determines where harvest activities occur and may be
used to enforce stand boundary and adjacency constraints. The
temporal component determines the timing (rotations) and
manner (single versus multiple-entry treatments) of harvest
activities. The species age-cohort component allows specifica-
tion of the species and age cohorts removed by the harvest
activities. For example, a clearcut removes all species and all
ages, whereas a selection harvest typically removes only a few
species and age cohorts. The ability to use a combination of
spatial, temporal, species, and age information to specify harvest
action independently allows a great variety of harvest prescrip-
tions to be simulated (Gustafson et al., 2000).
LANDIS simulates seed dispersal based upon species’
effective and maximum seeding distance (He and Mladenoff,
1999). Seed dispersal probability is modeled for each species
using an exponential distribution that defines the effective and
maximum seed dispersal distances.
Non-spatial processes of succession and seedling establish-
ment are simulated independently at each site. They also
interact with spatial processes such as seed dispersal, harvest-
ing, and disturbances. Succession is a competitive process
driven by species life history parameters in LANDIS. It is
comprised of a set of logical rules primarily using the
combination of shade tolerance, seeding ability, longevity,
vegetative reproduction capability, and the suitability of the
land type (Mladenoff and He, 1999). These rules are used to
simulate species birth, growth and death at 10-year intervals.
For example, shade intolerant species cannot establish on a site
where species with greater shade tolerance are present. On the
other hand, the most shade tolerant species are unable to occupy
an open site. Without disturbance, shade tolerant species will
dominate the landscape given that other attributes (e.g.,
dispersal distances) are not highly limiting and the environ-
mental conditions are otherwise suitable. Species establishment
is regulated by a species-specific establishment coefficient
(ranging from 0 to 1.00), which quantifies how different land
types favor or inhibit the establishment of a particular species
(Mladenoff and He, 1999). These coefficients, which are
Table 1
Species’ life attributes for Tuqiang Forest Bureau in northeastern China
Species LONG M
Larch (Larix gmelini) 300 20
Mongolian Scotch pine (Pinus sylvestris var. mongolica) 250 40
Spruce (Picea koraiensis) 300 30
White birch (Betula platyphylla) 150 15
Aspen-D (Populus davidiana) 180 20
Black birch (Betula davurica) 150 15
Aspen-S (Populus suaveolens) 150 25
Willow (Chosenia arbutifolia) 200 30
LONG: longevity (years); MTR: age of maturity (years); ST: shade tolerance (1–5
seeding distance (m); VP: vegetative reproduction probability; MVP: minimum ag
provided as input to LANDIS, are derived either from the
simulation results of a gap model (e.g., He et al., 1999) or from
estimates based on existing experimental or empirical studies
(Shifley et al., 2000).
Due to the stochastic nature of processes such as fire,
windthrow, and seedling dispersal simulated in the model,
LANDIS is not designed to predict the specific time or place that
individual disturbance events will occur. Rather, it is a cause–
response type of scenario model that simulates landscape
patterns over time in response to the combined and interactive
outcomes of succession and disturbance. It can provide managers
with guidance about management practices that can mitigate
current or anticipated problems on the forest landscape, and
provide a better understanding of long-term, cumulative effects
that may result from the combination of natural disturbances and
management practices. LANDIS has been applied and tested
with different species and environmental settings (Gustafson
et al., 2000; Shifley et al., 2000; Franklin et al., 2001; He et al.,
2002a,b; Mehta et al., 2004; Wimberly, 2004). In addition, the
current version, LANDIS 4.0, added new capabilities that
simulate explicit fuel dynamics, fuel-fire interactions, and
biological disturbances (He et al., 2004). Parameterization of
LANDIS 4.0 for the Tuqiang Forest Bureau involved several
aspects: species’ vital attributes, a forest composition map that
contains individual species presence/absence and age classes at
each cell, a land type map, establishment probabilities for each
landtype, fire disturbance regimes for each landtype, and forest
management scenarios.
3.1.1. Species attributes and forest composition map
A total of eight tree species were incorporated into LANDIS.
Species’ vital attributes (Table 1) were estimated based on
existing studies of the region (Ai et al., 1985; Duan, 1991; Hu
et al., 1991; Xu, 1998; He et al., 2002a; Xu et al., 2004) as well
as consultation with local experts. A forest composition map
was derived from an extant forest stand map of 1987, a stand
attribute database, and one scene of Landsat TM imagery taken
in 1987. The forest stand map recorded boundaries of stands
and compartments. (A compartment is a unit of forest inventory,
generally containing 10–100 stands.) The stand attribute
database provided information on the relative percentage of
canopy species, the average age of dominant canopy species,
timber production, and crown density. To reduce computational
loads during model simulations, the forest composition map
TR ST FT ED MD VP MVP
3 3 150 300 0 0
1 1 100 200 0 0
4 2 10 150 0 0
1 3 200 4000 0.8 40
1 3 �1 �1 1 40
1 4 200 1000 0.8 40
1 4 �1 �1 0.9 40
2 2 �1 �1 0.9 30
); FT: fire tolerance (1–5); ED: effective seeding distance (m); MD: maximum
e of vegetative reproduction (years). �1 represents unlimited seeding range.

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–9386
Fig. 3. Spatial distribution of part landscape occupied by larch at simulation year 200. (I1, I2, I3, and I4 represent simulation scenarios where the planting intensity
covers 10%, 30%, 50%, and 70% of the severely burned area, respectively. M1 and M2 represent dispersal planting and aggregated planting, respectively.)
Table 2
Attributes for each land type of Tuqiang Forest Bureau in China
Land type MFRI TSLF EC1 EC2 EC3 EC4 EC5 EC6 EC7 EC8
SS 300 150 0.3 0.25 0.03 0.3 0.01 0.1 0 0
NS 350 180 0.25 0.2 0.05 0.25 0.01 0.05 0 0
T 1000 500 0.002 0 0 0.004 0.001 0 0.01 0.05
BSS 300 10 0.3 0.25 0.03 0.3 0.01 0.1 0 0
BNS 350 10 0.25 0.2 0.05 0.25 0.01 0.05 0 0
BT 1000 10 0.002 0 0 0.004 0.001 0 0.01 0.05
MFRI: mean fire return intervals in years; TSLF: time since last fire disturbance;
EC1, EC2, EC3, EC4, EC5, EC6, EC7, EC8 are the establishment coefficient
for larch, Mongolian Scotch pine, spruce, white birch, aspen-D, black birch,
aspen-S, and willow. SS: southern slope; NS: northern slope; T: terrace; BSS:
burned southern slope; BNS: burned northern slope; BT: burned terrace.
was processed at 90 m � 90 m resolution, which yielded 1604
rows � 873 columns. Each cell contains the presence/absence
and age cohorts of all eight tree species.
For each cell in a stand, we used a stand-based assignation
(SBA) approach (Xu et al., 2004) to stochastically assign
species age cohorts to each cell based on forest inventory data.
Xu et al. (2004) used uncertainty analysis to evaluate the
approach, and their results demonstrated that uncertainty was
relatively low at the cell level during the beginning of the
simulation. The uncertainty increased with simulation years,
but the uncertainty finally reached an equilibrium state in which
input errors in original species age cohorts had little effect on
the simulation outcomes. At the landscape level, species
abundance and spatial patterns were not substantially affected
by the uncertainties in species age structure at the cell level.
Since the typical application of LANDIS is to predict long-term
landscape pattern changes, SBA can be used to parameterize
species age cohorts for individual cells.
3.1.2. Landtype map
LANDIS stratifies the heterogeneous landscape into relatively
homogeneous units (landtypes or ecoregions). Within each
landtype, similar environments for species establishment are
assumed (Mladenoff and He, 1999). In the study, we derived
eight landtypes, primarily based on terrain attributes, TM images
taken in 1987, and the catastrophic fire of 1987 (Fig. 3). These
landtypes include water, residential land, terrace, southern slope
(SS), northern slope (NS), burned terrace (BT), burned southern
slope (BSS), and burned northern slope (BNS). Because large
burned areas came into existence after the 1987 fire, fire/fuel
characteristics – such as fuel accumulation and the time elapsed
since the previous fire – were different for unburned areas.
Therefore, we differentiated BT, BSS, and BNS from unburned
areas. All landtypes were interpreted from the previous forest
inventory and the TM images taken in 1987. Non-active
landtypes (water and residential land) account for 0.90% of the
total area, while active landtypes (including terrace, SS, NS, BT,
BSS, and BNS) account for 0.94%, 23.57%, 17.39%, 3.39%,
31.43%, and 22.38%, respectively.
The species establishment coefficient is a critical feature of
each landtype. It is an estimate of the probability that a species
will successfully establish on a landtype, given the environ-
mental conditions encapsulated by that landtype. The species
establishment coefficients (Table 2) were derived from
available literature as well as the existing LANDIS para-
meterizations on northeastern China (Li et al., 1987; Zhao et al.,
1997; Liu et al., 1999; He et al., 2002a; Hu et al., 2004).

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–93 87
3.1.3. Fire regimes
The current fire regime for our simulations was parameter-
ized based on a database of 10-year fire dating from 1990 to
2000 (Hu et al., 2004). Current fire return intervals were
estimated by calculating the reciprocal of the annual proportion
of forest land burned within each landtype in the Bureau. Only
those fires greater than the resolution of the simulation recorded
in the database (0.81 ha) were used to parameterize the fire
regime (Table 2). Within the study area, the common planting
strategy consisted of planting coniferous trees over about 50%
of the total severely burned area (about 41300 ha). Two
endemic coniferous species of larch and Mongolian Scotch pine
(the ratio is about 7:3) were the main tree species planted.
3.2. Simulation scenarios
We began with the realistically parameterized forest
composition and landtype maps including species/age classes
that represent the initial status of 1987. Four levels of planting
intensity (10%, 30%, 50%, and 70% of the severely burned
area) and two spatial patterns of plantation (dispersed planting
and current/aggregated planting) were simulated in a 4 � 2
factorial design over a 300-year period (Table 3). Two
coniferous tree species (larch and Mongolian Scotch pine)
were planted in severely burned areas, and the proportion was
7:3. Five replicates of each scenario were simulated. The
current planting intensity of planting coniferous species over
about 50% of the severely burned area was simulated;
additionally, three other planting intensities (including 10%,
30%, and 70% of the severely burned area) were selected to
compare with the current planting intensity. The planting
strategy, dispersed planting, consists of planting coniferous
seedlings randomly throughout a severely burned area, whereas
aggregated planting consists of planting coniferous seedlings in
selected locations of severely burned areas where coniferous
forests were dominant prior to a fire; both strategies were
applied in our simulations. A ‘‘no reforestation’’ scenario was
also simulated. All scenarios were simulated up to 300 years to
examine the long-term effects of planting intensity and spatial
pattern of plantation on forest succession.
The study area was divided into four management areas
(MA) indicating the planting options practiced there: (1)
unburned area (MA 1), (2) severely burned area where conifers
were dominant before the 1987 fire (MA 2), (3) severely burned
Table 3
The scenarios simulated by LANDIS
Simulation scenarios Descriptions
I1M1 Planting coniferous trees, covering 10% of the severely bu
I2M1 Planting coniferous trees, covering 30% of the severely bu
I3M1 Planting coniferous trees, covering 50% of the severely bu
I4M1 Planting coniferous trees, covering 70% of the severely bu
I1M2 Planting coniferous trees, covering 10% of the severely bu
I2M2 Planting coniferous trees, covering 30% of the severely bu
I3M2 Planting coniferous trees, covering 50% of the severely bu
I4M2 Planting coniferous trees, covering 70% of the severely bu
area where conifers were not dominant before the fire (MA 3),
and (4) other burned area (MA 4) (Fig. 1). The goal of the
current planting strategy was to increase the proportion of
coniferous trees in the Bureau. Dispersed planting was applied
in MA 2 and MA 3, while current planting was restricted in MA
2. Within MA 2, planting activity was applied to 50% of the
severely burned area in 10 years to mimic the current planting
strategy designed to promote coniferous species. In MA 1 and
MA 4, natural regeneration was not simulated because there
were sufficient residual coniferous trees to provide seeds.
Table 3 gives an overview of different combinations of planting
intensities and spatial patterns of plantation.
For each of the three species (larch, Mongolian Scotch pine,
and white birch), the proportion of area (PA) was calculated for
the mapped output of species for each 10-year step in the 300
simulation years to qualitatively evaluate temporal trends in the
data. We divided the 300 simulation years into three stages: 10–
30 year as short-term stage, 40–100 year as mid-term stage, and
>100 years as long-term stage. The average PA in each of the
three stages was calculated for all three species. However, the
average PA for the three species is dependent on each other at
different simulation stages. Therefore, we analyzed the three
response variables (average area of the three species) together
in a multivariate analysis of variance (MANOVA) with planting
intensity (n = 4) and spatial pattern of plantation (n = 2) in
SPSS (SPSS 10.0) to test the global hypothesis that planting
intensity and spatial pattern of plantation affect the area of the
three species at different simulation stages. We used the Pillai’s
Trace statistic to test our hypotheses because it is the least
sensitive of the four multivariate tests provided by SPSS to the
heterogeneity of variance assumption of MANOVA (Zar,
1999).
In addition, to evaluate the effect of reforestation scenarios
on the spatial pattern of species distributions, we calculated an
aggregation index (AI) for the three species using the mapped
output of each species throughout the 300-year simulation. The
AI has a range between 0 and 1, with higher values indicating a
higher level of aggregation in the measured spatial pattern (He
et al., 2000). It assumes that a class with the highest level of
aggregation (AI = 1) is comprised of pixels sharing the most
possible edges. A class whose pixels share no edges
(completely disaggregated) has the lowest level of aggregation
(AI = 0). If ei,j represents total edges of class i adjacent to class
j, for class i of area Ai, aggregation index measures ei,i, the total
rned area (70% larch and 30% Mongolian Scotch pine); dispersed planting
rned area (70% larch and 30% Mongolian Scotch pine); dispersed planting
rned area (70% larch and 30% Mongolian Scotch pine); dispersed planting
rned area (70% larch and 30% Mongolian Scotch pine); dispersed planting
rned area (70% larch and 30% Mongolian Scotch pine); aggregated planting
rned area (70% larch and 30% Mongolian Scotch pine); aggregated planting
rned area (70% larch and 30% Mongolian Scotch pine); aggregated planting
rned area (70% larch and 30% Mongolian Scotch pine); aggregated planting

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–9388
edges shared by class i itself. The shared edges are counted only
once in AI, and currently only four neighbors are considered.
The level of aggregation of class i is calculated as:
AIi ¼ei;i
max ei;i:
Given a class is of area Ai, the maximum aggregation level is
reached when A clumps into one patch that has the largest ei,i
(e.g., it does not have to be a square). If n is the side of largest
integer square smaller than Ai, and m = Ai � n2, then the largest
number of shared edges for class i, max_ei,i, will take one of the
three forms:
max ei;i ¼ 2nðn� 1Þ; when m ¼ 0; or
max ei;i ¼ 2nðn� 1Þ þ 2m� 1; when m< n; or
max ei;i ¼ 2nðn� 1Þ þ 2m� 2; when m � n:
The two indices (PA and AI) were calculated in APACK
(Mladenoff and DeZonia, 1997). This analysis was conducted
for the entire study area.
4. Results
MANOVA results indicate that both planting intensity and
spatial pattern of plantation also significantly affect the PA of
the three species (larch, Mongolian Scotch pine and white
birch) across the study landscape at short-, mid-, and long-term
stages (Tables 4–6), and their interaction of both treatments has
also a significant influence. Planting intensity always has a
significant influence on the successional dynamic of the three
species for all simulation years. However, the effects include
both positive and negative effects: greater planting intensity
Table 4
MANOVA and individual ANOVA results for the average area of the three species (la
planting intensity and spatial pattern of plantation
Effect df (h, e)
MANOVA global test of hypotheses
Intensity (I) 9,96
Spatial pattern of plantation (P) 3,30
I � P 9,96
Individual ANOVA tests of hypotheses
Average area of larch at short-term stage–Model R2 = 0.96
Intensity (I) 3
Spatial pattern of plantation (P) 1
I � P 3
Error (e) 32
Average area of Mongolian Scotch pine at short-term stage–Model R2 = 0.998
Intensity (I) 3
Spatial pattern of plantation (P) 1
I � P 3
Error (e) 32
Average area of white birch at short-term stage–Model R2 = 0.988
Intensity (I) 3
Spatial pattern of plantation (P) 1
I � P 3
Error (e) 32
positively influences larch and Mongolian Scotch pine
abundance, but negatively influences white birch abundance.
Larch abundance is greater under high planting intensity
scenarios than under low planting intensity scenarios. For
example, under the 10% planting intensity scenario, it takes
roughly 200 years for larch to reach 70% of the landscape, but it
takes only 60 years to reach 70% of the landscape under the 70%
planting intensity scenario. The PA of Mongolian Scotch pine
responds to planting intensity in a similar way, with a greater
abundance under high planting intensity scenarios than that
under low planting intensity scenarios. In addition, the PA for
larch increases with simulation years in all simulation scenarios,
whereas the PA for Mongolian Scotch pine decreases with
simulation years after roughly 10–20 years (Fig. 4). Mongolian
Scotch pine has a relatively low abundance – less than 10% of the
study area during most simulation years – because it is a fire-
intolerant and shade-intolerant species and is less competitive
than either larch or white birch. Contrary to larch, white birch has
a greater abundance under low planting intensity scenarios than
that under high planting intensity scenarios. For example, under
the 10% planting intensity scenario, the PA for white birch
reaches 30% after only 10 years, and exceeds 40% after 40 years.
However, under the 70% planting intensity scenario, it requires
between 30 and 40 years attaining 30% abundance, and never
reaches 40% in 300 simulation years.
Individual ANOVA results also show that spatial pattern of
plantation has a significant influence on the short-, mid-, and
long-term PA for larch, and has a significant influence on the
long-term PA for both Mongolian Scotch pine and white birch
(Tables 4–6). In other words, spatial pattern of plantation has a
significant influence on the abundance of larch during all
simulation years, but has no short- or mid-term influence on
rch, Mongolian Scotch pine and white birch) at short-term stage as a function of
Pillai’s trace/type III SS F P
1.33 8.53 <0.001
0.92 112.69 <0.001
0.72 3.39 0.001
441.49 253.86 <0.001
3.32 5.73 0.023
0.39 0.23 0.88
18.55
73.49 6643.83 <0.001
0.0006 0.16 0.69
0.003 0.29 0.834
0.118
309.30 880.75 <0.001
0.034 0.289 0.594
0.014 0.041 0.989
3.75

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–93 89
Table 5
MANOVA and individual ANOVA results for the average area of the three species (larch, Mongolian Scotch pine and white birch) at mid-term stage as a function of
planting intensity and spatial pattern of plantation
Effect df (h, e) Pillai’s trace/type III SS F P
MANOVA global test of hypotheses
Intensity (I) 9,96 1.65 13.00 <0.001
Spatial pattern of plantation (P) 3,30 0.96 237.22 <0.001
I � P 9,96 0.87 4.37 <0.001
Individual ANOVA tests of hypotheses
Average area of larch at mid-term stage–Model R2 = 0.994
Intensity (I) 3 521.9 1519.73 <0.001
Spatial pattern of plantation (P) 1 52.16 455.65 <0.001
I � P 3 6.12 17.83 <0.001
Error (e) 32 3.66
Average area of Mongolian Scotch pine at mid-term stage–Model R2 = 0.996
Intensity (I) 3 81.37 2610.86 <0.001
Spatial pattern of plantation (P) 1 0.027 2.59 0.118
I � P 3 0.06 2.03 0.13
Error (e) 32 0.33
Average area of white birch at mid-term stage–Model R2 = 0.975
Intensity (I) 3 631.96 423.24 <0.001
Spatial pattern of plantation (P) 1 0.16 0.32 0.574
I � P 3 0.75 0.50 0.684
Error (e) 32 15.93
Table 6
MANOVA and individual ANOVA results for the average area of the three species (larch, Mongolian Scotch pine and white birch) at long-term stage as a function of
planting intensity and spatial pattern of plantation
Effect df (h, e) Pillai’s trace/type III SS F P
MANOVA global test of hypotheses
Intensity (I) 9,96 1.64 12.8 <0.001
Spatial pattern of plantation (P) 3,30 0.99 796.98 <0.001
I � P 9,96 0.965 5.06 <0.001
Individual ANOVA tests of hypotheses
Average area of larch at long-term stage–Model R2 = 0.993
Intensity (I) 3 179.2 886.72 <0.001
Spatial pattern of plantation (P) 1 132.73 1970.36 <0.001
I � P 3 11.11 54.97 <0.001
Error (e) 32 2.16
Average area of Mongolian Scotch pine at long-term stage–Model R2 = 0.989
Intensity (I) 3 38.63 2610.86 <0.001
Spatial pattern of plantation (P) 1 0.20 15.30 <0.001
I � P 3 0.05 1.15 0.346
Error (e) 32 0.42
Average area of white birch at long-term stage–Model R2 = 0.969
Intensity (I) 3 287.37 310.41 <0.001
Spatial pattern of plantation (P) 1 16.88 54.69 <0.001
I � P 3 3.38 3.65 0.023
Error (e) 32 9.88
either Mongolian Scotch pine or white birch. The interaction of
treatment intensity and treatment method has no effect on the
dynamics of Mongolian Scotch pine throughout the simulation
years, and only affects the long-term stage of white birch
abundance. The PA for larch under the dispersed planting
scenario is higher than that under the aggregated planting
scenario, very likely because dispersed planting increases the
cover range of seed dispersal and benefits the regeneration of
larch. However, Mongolian Scotch pine does not respond to
spatial pattern of plantation in a similar way. Spatial pattern of
plantation has no influence on the abundance of Mongolian
Scotch pine at the early simulation stage because the pine has a
low proportion in the landscape and only relatively short
dispersal distances. At the long-term simulation stage, spatial
pattern of plantation shows an influence on Mongolian Scotch
pine because such a long time frame captures its slow and

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–9390
Fig. 4. Proportion of sites in the study landscape occupied by larch, Mongolian Scotch pine, and birch during each decade for all simulation scenarios. I1, I2, I3, and I4
represent simulation scenarios where the planting intensity covers 10%, 30%, 50%, and 70% of the severely burned area, respectively.
cumulative dispersal behavior. However, the abundance of
Mongolian Scotch pine is only slightly higher under the
dispersed planting scenario than that under the aggregated
planting scenario (Fig. 4). Due to its strong dispersal and
colonization ability, white birch can invade open spaces created
by fire. This results in no significant influence of the two spatial
patterns of plantation on white birch abundance at the early
simulation stage. However, in the later simulation stages, the
abundance of white birch under the dispersed planting scenario
is lower than that under the aggregated planting scenario due to
strong competition with larch. The environment becomes more
unfavorable for white birch in a dispersed planting as the
canopy closure of larch forests increases.
Compared with the no planting scenario, the abundance of
larch in these planting scenarios was greater, which suggests
that planting activity is effective for increasing the abundance
of larch throughout these simulation years. Given its vital
attributes and low abundance, the decreasing abundance of
Mongolian Scotch pine with greater simulation years in all
simulation scenarios is not surprising. The difference between
these planting simulation scenarios and the no planting scenario
– especially in the later simulation stage – is discernable, with
the abundance of white birch under the no planting scenario
higher than that in the planting simulation scenarios due to its
diminished competition with larch.
The simulated results indicate that planting intensity and
spatial pattern of plantation not only significantly influence
the abundance of the three species, but also influence the
spatial pattern of these species (Fig. 5). However, the degree
of aggregation of species distribution differs by different
planting scenarios. The aggregation levels for larch under all
planting scenarios are similar, probably because larch has
relatively high proportions under all simulation scenarios;
high proportions tend to have aggregated distributions in the
study landscape. The aggregation level of larch is consider-
able high, with an AI larger than 0.7 at the start of the
simulation, and stabilizing at 0.8 after only a few simulation
decades. The distribution of Mongolian Scotch pine is more
aggregated under the high planting intensity scenarios than
that under the low planting intensity scenarios, whereas the
aggregation level of white birch is negatively correlated with
greater planting intensity. Also, the distribution of pine under
the dispersed planting scenarios has a lower aggregation level
than that under the aggregated planting scenarios. However,
the difference in the level of aggregation between the two
spatial patterns of plantation increases with the planting
intensity. There is a sudden decrease in the AI for pine after
about 200 simulation years; this is because the large number
of pines planted at the beginning of the simulation began to
die when they approached their 250-year longevity. The
aggregation level of white birch responds to spatial pattern of
plantation in a way similar to that for larch, with its AI lower
under the dispersed planting scenarios than under the
aggregated planting scenarios.
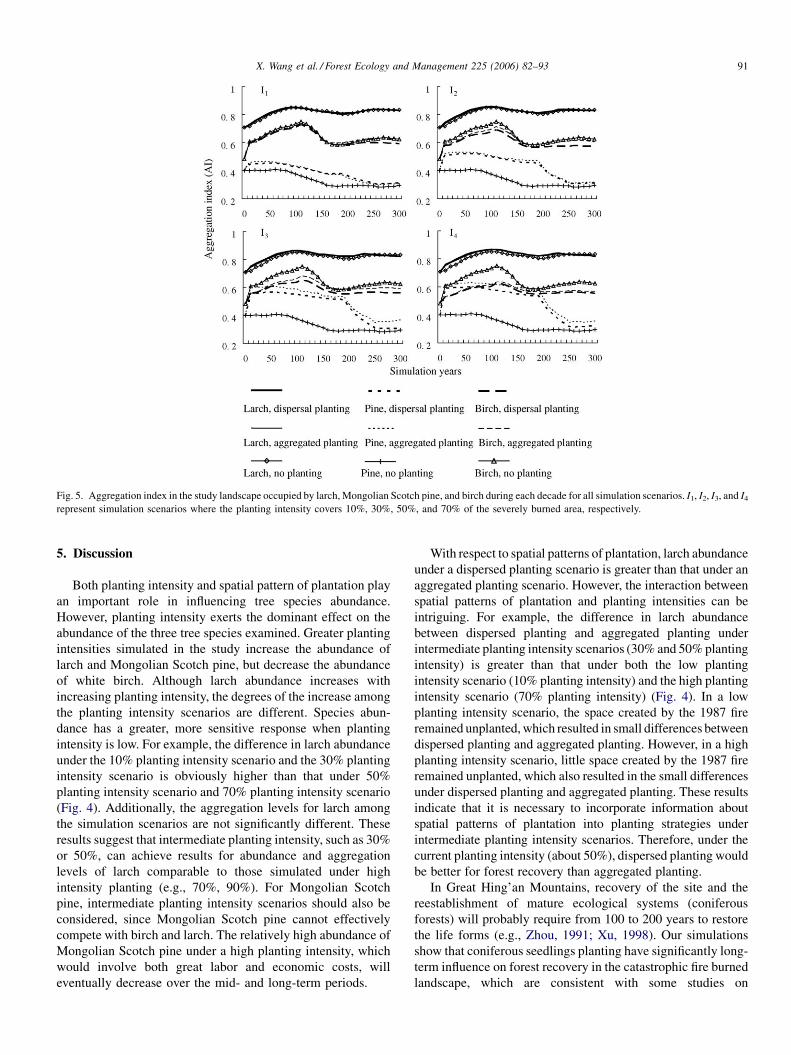
X. Wang et al. / Forest Ecology and Management 225 (2006) 82–93 91
Fig. 5. Aggregation index in the study landscape occupied by larch, Mongolian Scotch pine, and birch during each decade for all simulation scenarios. I1, I2, I3, and I4
represent simulation scenarios where the planting intensity covers 10%, 30%, 50%, and 70% of the severely burned area, respectively.
5. Discussion
Both planting intensity and spatial pattern of plantation play
an important role in influencing tree species abundance.
However, planting intensity exerts the dominant effect on the
abundance of the three tree species examined. Greater planting
intensities simulated in the study increase the abundance of
larch and Mongolian Scotch pine, but decrease the abundance
of white birch. Although larch abundance increases with
increasing planting intensity, the degrees of the increase among
the planting intensity scenarios are different. Species abun-
dance has a greater, more sensitive response when planting
intensity is low. For example, the difference in larch abundance
under the 10% planting intensity scenario and the 30% planting
intensity scenario is obviously higher than that under 50%
planting intensity scenario and 70% planting intensity scenario
(Fig. 4). Additionally, the aggregation levels for larch among
the simulation scenarios are not significantly different. These
results suggest that intermediate planting intensity, such as 30%
or 50%, can achieve results for abundance and aggregation
levels of larch comparable to those simulated under high
intensity planting (e.g., 70%, 90%). For Mongolian Scotch
pine, intermediate planting intensity scenarios should also be
considered, since Mongolian Scotch pine cannot effectively
compete with birch and larch. The relatively high abundance of
Mongolian Scotch pine under a high planting intensity, which
would involve both great labor and economic costs, will
eventually decrease over the mid- and long-term periods.
With respect to spatial patterns of plantation, larch abundance
under a dispersed planting scenario is greater than that under an
aggregated planting scenario. However, the interaction between
spatial patterns of plantation and planting intensities can be
intriguing. For example, the difference in larch abundance
between dispersed planting and aggregated planting under
intermediate planting intensity scenarios (30% and 50% planting
intensity) is greater than that under both the low planting
intensity scenario (10% planting intensity) and the high planting
intensity scenario (70% planting intensity) (Fig. 4). In a low
planting intensity scenario, the space created by the 1987 fire
remained unplanted, which resulted in small differences between
dispersed planting and aggregated planting. However, in a high
planting intensity scenario, little space created by the 1987 fire
remained unplanted, which also resulted in the small differences
under dispersed planting and aggregated planting. These results
indicate that it is necessary to incorporate information about
spatial patterns of plantation into planting strategies under
intermediate planting intensity scenarios. Therefore, under the
current planting intensity (about 50%), dispersed planting would
be better for forest recovery than aggregated planting.
In Great Hing’an Mountains, recovery of the site and the
reestablishment of mature ecological systems (coniferous
forests) will probably require from 100 to 200 years to restore
the life forms (e.g., Zhou, 1991; Xu, 1998). Our simulations
show that coniferous seedlings planting have significantly long-
term influence on forest recovery in the catastrophic fire burned
landscape, which are consistent with some studies on

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–9392
reforestation (e.g., Miyawaki and Golley, 1993; Chen et al.,
2003), for coniferous seedlings planting will eventually
replicate the mature ecosystems of the site without going
through the lengthy process of succession. Residual trees play
an important role in forest restoration (e.g., Keeton and
Franklin, 2005). However, only few residual trees remained in
the severely burned area after the 1987 fire, which resulted in a
lower abundance of coniferous trees under the no planting
scenario than that under the planting scenarios. Therefore, large
area clear-cutting should not be applied in forest management
for this region. Coniferous trees planted in the severely burned
area serve as residual trees, which further disperse seeds for
forest recovery—that is, more residual trees are left with
increasing planting intensity. In addition, the simulation results
demonstrate that dispersed planting yields greater abundance of
larch forests than aggregated planting, suggesting that some
dispersed coniferous trees or seedlings should be left as seed
sources for forest recovery when timber harvesting is applied in
a forest landscape.
Two key assumptions likely affect our conclusions. The first is
that the species establishment coefficients do not vary at the scale
of landtypes. Fine scale heterogeneity, such as soil moisture and
temperature availability, are important variables to affect forest
dynamics because they would affect plant establishment and
growth, species diversity, nutrient cycling, rates of soil formation
and other biotic activity (e.g., Xu, 1998). In reality, drought sites
are often colonized by Mongolian Scotch pine rather than larch,
for the pine has a lower shade tolerance than larch. Hence, the
abundance of Mongolian Scotch pine may be underestimated in
these scenarios. However, given its vital attributes discussed
above, we believe that Mongolian Scotch pine could not have a
high proportion of the study landscape. In addition, species
establishment coefficients may vary with fire severity. In low-
severity burned areas, burning prepares seedbeds by removing
duff and competing vegetation, and by altering the thermal
regime of the soil, which favors germination. But in severely
burned areas, organic layers of soil were almost completely
consumed—an unfavorable condition for the regeneration of
larch and Mongolian Scotch pine (Xiao et al., 1988; Xu, 1998).
Using finer-scale land units to parameterize our simulations
could alleviate these problems. However, no available data
currently exists to estimate species establishments at a finer
spatial scale.
A second assumption is that once established, trees die only
because of disturbance-caused mortality, approaching their
longevity, and a small chance of random mortalities simulated in
the model. In other words, other types of mortality such as stress
related in the early stage of their life span are not considered in the
LANDIS model. This assumption is appropriate, given that
human interference is not considered. In this study, intensive
planting is employed in the burned area, but the success rates of
species establishment are also affected by the quality of the
seedlings and aftermath care after planting (e.g., Xiao et al.,
1988; Zhao, 1988; Tan, 1994). These factors are not addressed in
LANDIS modeling. In addition, LANDIS did not simulate the
effects of shrubs and herbs on forest recovery, which could have
significant influence on coniferous seedlings. For example, some
studies (Turner et al., 1997, 1999) have shown that shrub and herb
recruitment following forest fires in burned areas where conifer
seedlings were scarce may lengthen the time required for forest
development, or even preclude them entirely, for shrub and herb
may compete with trees for resources (e.g., light, nutrients).
Previous studies in the Great Hing’an Mountains showed that
shrubs and herbs would soon invade these open spaces created by
the catastrophic fire in 1987. In some cases, if vegetation
becomes too dense, light levels beneath shrubs and grasses may
be insufficient for tree seedlings to maintain a net positive carbon
balance, and thus inadequate radiation may cause low coniferous
seedling survivorship. Therefore, coniferous seedlings could not
regenerate in some places because of competition from the well-
developed shrub and herb community (e.g., Zhou et al., 1989;
Yang et al., 1998; Wang et al., 2003). If shrubs and herbs were not
cleared before coniferous seedling plantation, some seedlings
planted could not survival, which would cause lower abundance
of coniferous tree species (larch and Mongolian Scotch pine).
However, this may not be a significant factor: in our study areas,
shrubs and herbs must be cleared prior to the coniferous seedling
plantation (Shao, 1988; Wang et al., 2003). When coniferous
seedlings grow older, they will out-compete shrubs and herbs.
Thus, the simulated results could effectively reflect the long-term
forest dynamics in the landscape based on this assumption.
6. Conclusion
Landscape models are an efficient tool for evaluating the
effects of different planting strategies on landscape-scale forest
recovery. They provide insights into forest dynamics under
natural disturbances and reforestation. In summary, we found
that planting intensity can modify the abundance of the three
species in the study (larch, Mongolian Scotch pine, and white
birch) by encouraging larch and pine establishment that will
eventually decrease white birch abundance. Although larch
abundance increases with increasing planting intensity, the
degree of increased abundance among these planting intensity
scenarios is different. Given considerable labor inputs and
economic costs, intermediate planting intensity (30% or 50%)
would better facilitate forest recovery. In addition, spatial
pattern of plantation also influences the three species, with
higher larch and pine abundance and lower white birch
abundance under dispersed planting than that under aggregated
planting. This indicates that it is necessary to incorporate
information about spatial patterns of plantation into planting
strategies. These results have important implications for forest
managers to design sound forest restoration projects for
landscapes affected by large infrequent disturbances.
Acknowledgements
This paper is sponsored by the Chinese Academy of
Sciences, National Natural Science Foundation of China
(40331008, 30270225), and the Project of Chinese Academy
of Sciences (No. KSCX2-SW-133) and Sino-Russian Joint
Research Center on Natural Resources and Eco-Environmental
Sciences.

X. Wang et al. / Forest Ecology and Management 225 (2006) 82–93 93
References
Ai, C.L., Wang, C.D., Ai, S.H., 1985. Study on the seed dispersal of Larix
gmelinii. Forest. Sci. Technol. 4, 2–5 (in Chinese).
Beyers, J.L., 2004. Postfire seeding for erosion control: effectiveness and
impacts on native plant communities. Conserv. Biol. 18 (4), 947–956.
Borchert, M., Johnson, M., Schreiner, D.S., Wall, S.B.V., 2003. Early postfire
seed dispersal, seedling establishment and seedling mortality of Pinus
coulterri (D. Don) in central coastal California, USA. Plant Ecol. 168,
207–220.
Brown, J.K., See, T.E., 1981. Downed dead woody fuel and biomass in the
northern Rocky Mountains. USDA Forest Service General Technical Report
INT-117, pp. 1–47.
Chen, X., Li, B., Lin, Z., 2003. The acceleration of succession for the restoration
of the mixed-broadleaved Korean pine forests in Northeast China. Forest
Ecol. Manage. 177, 503–514.
Duan, X., 1991. Study on fire tolerance of Larix gmelinii. Forest Fire Prevent. 31
(4), 7–11 (in Chinese).
Franklin, J., Syphard, A.D., Mladenoff, D.J., He, H.S., Simons, D.K., Martin,
R.P., Deutschman, D., O_Leary, J.F., 2001. Simulating the effects of
different fire regimes on plant functional groups in Southern California.
Ecol. Model. 142, 261–283.
Grimm, V., 1999. Ten years of individual-based modelling in ecology: what
have we learned and what could we learn in the future? Ecol. Model. 115,
129–148.
Gustafson, E.J., Shifley, S.R., Mladenoff, D.J., Nimerfro, K.K., He, H.S., 2000.
Spatial simulation of forest succession and timber harvesting using
LANDIS. Can. J. Forest Res. 30, 32–43.
He, H.S., Mladenoff, D.J., Boeder, J., 1999. An object-oriented forest landscape
model and its representation of tree species. Ecol. Model. 119, 1–19.
He, H.S., DeZonia, B., Mladenoff, D.J., 2000. An aggregation index (AI) to
quantify spatial patterns of landscapes. Landscape Ecol. 15 (7),
591–601.
He, H.S., Hao, Z., Larsen, D.R., Dai, L., Hu, Y.M., Chang, Y., 2002a. A
simulation study of landscape scale forest succession in China. Ecol. Model.
156, 153–166.
He, H.S., Mladenoff, D.J., Gustafson, E.J., 2002b. Study of landscape change
under forest harvesting and climate warming-induced fire disturbance.
Forest Ecol. Manage. 155, 257–270.
He, H.S., Li, W., Sturtevant, B.R., Yang, J., Shang, B.Z., Gustafson, E.J.,
Mladenoff, D.J., 2004. LANDIS 4.0 User’s Guide. School of Natural
Resources, University of Missouri-Columbia, Columbia, pp. 1–91.
He, H.S., Mladenoff, D.J., 1999. Spatially explicit and stochastic simulation of
forest landscape fire disturbance and succession. Ecology 80, 81–99.
Hu, H.Q., Yao, S.R., Shang, D.Y., 1991. The characteristics and effect of the
forest fire in Northeast China. Forest Fire Prevent. 31 (4), 13–16 (in Chinese).
Hu, Y., Xu, C., Chang, Y., 2004. Application of spatially explicit landscape
model (LANDIS): a case study in Huzhong area, Mt. Great Hing’an. Acta
Ecol. Sinica 24 (9), 1846–1856 (in Chinese).
Kauffman, J.B., Uhl, C., Cummings, D.L., 1988. Fire in the Venezuelan
Amazon. 1: Fuel biomass and tree chemistry in the evergreen rainforest
of Venezuela. Oikos 53, 167–175.
Keeton, W.S., Franklin, J.E., 2005. Do remnant old-growth trees accelerate rates
of succession in mature Douglas-fir forests? Ecol. Monogr. 75 (1), 103–118.
Lamb, D., 1998. Large-scale ecological restoration of degraded tropical forest
lands: the potential role of timber plantations. Restor. Ecol. 6 (3), 271–279.
Li, R.X., Zhao, H.X., Wang, F.Y., Zhang, D.L., 1987. The relationship between
growth of natural Larix gmelinii and land type. J. Northeast Forest. Univ. 15,
55–59 (in Chinese).
Liu, G.F., Yang, C.P., Liu, G.J., Yang, R.H., 1999. The configuration characters
and germination percentage of various provenance seeds of Betula platy-
phylla. J. Northeast Forest. Univ. 27 (4), 1–4 (in Chinese).
Mehta, S., Frelich, L.E., Jones, M.T., Manolis, J., 2004. Examining the effects of
alternative management strategies on landscape-scale forest patterns in
northeastern Minnesota using LANDIS. Ecol. Model. 180, 73–87.
Mladenoff, D.J., 2004. LANDIS and forest landscape models. Ecol. Model.
180, 7–19.
Mladenoff, D.J., DeZonia, B., 1997. APACK 2.0 User’s Guide. Department of
Forest Ecology and Management. University of Wisconsin-Madison, Madi-
son, WI, USA.
Miyawaki, A., Golley, F.B., 1993. Forest reconstruction as ecological engineer-
ing. Ecol. Eng. 2, 333–345.
Mladenoff, D.J., He, H.S., 1999. Design and behavior of LANDIS, an object-
oriented model of forest landscape disturbance and succession. In: Mladen-
off, D.J., Baker, W.L. (Eds.), Advances in spatial Modeling of Forest
Landscape Change: Approaches and Applications. Cambridge University
Press, Cambridge, UK, pp. 1–13.
Nagashima, K., Sands, R., Whyte, A.G.D., Bilek, E.M., Nakagoshi, N., 2002.
Regional landscape change as a consequence of plantation forestry expan-
sion: an example in the Nelson region, New Zealand. Forest Ecol. Manage.
163, 245–261.
Romme, W.H., 1982. Fire and landscape diversity in subalpine forests of
Yellowstone National Park. Ecol. Monogr. 52 (2), 199–221.
Shao, C.H., 1988. Suggestions for the regeneration of burned areas in Tuqiang
Forest Bureau. Chin. J. Ecol. 7 (sup.), 7–74.
Shifley, S.R., Thompson III, F.R., Larsen, D.R., Dijak, W.D., 2000. Modeling
forest landscape change in the Missouri Ozarks under alternative manage-
ment practices. Comput. Electr. Agric. 27, 7–27.
Shu, L.F., Zuo, J.C., Tian, W.T., 1996. Discussion on natural regeneration in
burned areas of Great Hing’an Mountains. Manage. Forest. Resources 6,
43–45 (in Chinese).
Smale, M.C., Whaley, P.T., Smale, P.N., 2001. Ecological restoration of native
forest at Aratiatia, North Island, New Zealand. Restoration Ecol. 9, 28–37.
Sturatevant, B.R., Zollner, P.A., Gustafson, E.J., Cleland, D.T., 2004. Human
influence on the abundance and connectivity of high-risk fuels in mixed
forests of northern Wisconsin, USA. Landscape Ecol. 19, 235–253.
Tan, J., 1994. Reason and strategy on forest land reduction in Daxing’anling
Mountains. Territory Natural Resour. Study 2, 15–19 (in Chinese).
Turner, M.G., Romme, W.H., Gardner, R.H., Hargrove, W.W., 1997. Effects of
fire size and pattern on early succession in Yellowstone National Park. Ecol.
Monogr. 4 (67), 411–433.
Turner, M.G., Romme, W.H., Gardner, R.H., 1999. Prefire heterogeneity, fire
severity, and early postfire plant reestablishment in subalpine forests of
Yellowstone National Park, Wyoming. Int. J. Wildland Fire 9 (1), 21–36.
Wang, X., Li, X., Kong, F., 2003. Model of vegetation restoration under natural
regeneration and human interference in the burned area of northern Great
Hing’an Mountains. Chin. J. Ecol. 22 (5), 30–34 (in Chinese).
Wimberly, M.C., 2004. Fire and forest landscapes in the Georgia Piedmont: an
assessment of spatial modeling assumptions. Ecol. Model. 180, 41–56.
Xiao, D., Tao, D., Xu, Z., 1988. Impacts of an extra-ordinarily disastrous fire on
forest resources and environment. Chin. J. Ecol. 7 (sup.), 5–9 (in Chinese).
Xu, H., 1998. Forest in Great Hing’an Mountains of China. Science Press,
Beijing, pp. 1–231, (in Chinese).
Xu, C., He, H.S., Hu, Y., Chang, Y., Larsen, D.R., Li, X., Bu, R., 2004.
Assessing the effect of cell-level uncertainty on a forest landscape model
simulation in Northeastern China. Ecol. Model. 180, 57–72.
Yang, S., Liu, X., Cao, H., 1998. Study on vegetational change in burned areas
of Great Hing’an Mountains. J. Northeast Forest. Univ. 26 (1), 19–23 (in
Chinese).
Yang, J., He, H.S., Gustafson, E.J., 2004. A hierarchical statistical approach to
simulate the temporal patterns of forest fire disturbance in LANDIS model.
Ecol. Model. 180, 119–133.
Zar, J.H., 1999. Biostatistical Analysis. Prentice-Hall Inc., Upper Saddle River,
USA.
Zhao, D., 1988. Vegetations and their restoration after the disastrous fire in the
Daxing’anling Mountains. Chin. J. Ecol. 7 (sup.), 35–40 (in Chinese).
Zhao, K.T., Li, Y.C., Zhang, G.L., 1997. The development and protection of
Pinus pumila forest in Da Hingan Mountain. Territory Natural Resour.
Study 2, 55–59 (in Chinese).
Zhou, Y., 1991. Vegetation in Great Hing’an Mountains of China. Science Press,
Beijing, pp. 1–216, (in Chinese).
Zhou, Y., Wu, H., Chen, T., Zheng, Y., 1989. Accelerating the recovery of forest
in the burned area in Great Hing’an Mountains according to the syneco-
logical characteristics. J. Northeast Forest. Univ. 17 (3), 1–10 (in Chinese).




















