
Short-term environmental variability in cold-water coral habitat at Viosca Knoll, Gulf of Mexico
14
Short-term environmental variability in cold-water coral habitat at Viosca Knoll, Gulf of Mexico Andrew J. Davies a, , Gerard C.A. Duineveld b , Tjeerd C.E. van Weering b,c , Furu Mienis b , Andrea M. Quattrini d,1 , Harvey E. Seim e , John M. Bane e , Steve W. Ross d a School of Ocean Sciences, Bangor University, Menai Bridge, Anglesey LL59 5AB, UK b Royal Netherlands Institute for Sea Research, PO Box 59, 1790 AB Den Burg, Texel, The Netherlands c Free University of Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlands d University of North Carolina at Wilmington, Center for Marine Science, 5600 Marvin K. Moss Lane, Wilmington, NC 28409, USA e Department of Marine Sciences, University of North Carolina at Chapel Hill, 340 Chapman Hall, Chapel Hill, NC 27599-3300, USA article info Article history: Received 26 June 2009 Received in revised form 15 October 2009 Accepted 2 November 2009 Available online 10 November 2009 Keywords: Lophelia pertusa Deep-sea corals Ecosystem engineer Chemosynthetic communities Environmental parameters abstract The Lophelia pertusa community at Viosca Knoll (VK826) is the most extensive found to date in the Gulf of Mexico. As part of a multi-disciplinary study, the physical setting of this area was described using benthic landers, CTD transects and remotely operated vehicle observations. The site was broadly characterised into three main habitats: (1) dense coral cover that resembles biogenic reef complexes, (2) areas of sediment, and (3) authigenic carbonate blocks with sparse coral and chemosynthetic communities. The coral communities were dominated by L. pertusa but also contained numerous solitary coral species. Over areas that contained L. pertusa, the environmental conditions recorded were similar to those associated with communities in the north-eastern Atlantic, with temperature (8.5–10.6 1C) and salinity ( 35) falling within the known species niche for L. pertusa. However, dissolved oxygen concentrations (2.7–2.8 ml l 1 ) and density (s Y , 27.1–27.2 kg m 3 ) were lower and mass fluxes from sediment trap data appeared much higher (4002–4192 mg m 2 d 1 ). Yet, this species still appears to thrive in this region, suggesting that L. pertusa may not be as limited by lower dissolved oxygen concentrations as previously thought. The VK826 site experienced sustained eastward water flow of 10–30 cm s 1 over the 5-day measurement period but was also subjected to significant short- term variability in current velocity and direction. In addition, two processes were observed that caused variability in salinity and temperature; the first was consistent with internal waves that caused temperature variations of 0.8 1C over 5–11 h periods. The second was high-frequency variability (20–30 min periods) in temperature recorded only at the ALBEX site. A further pattern observed over the coral habitat was the presence of a 24 h diel vertical migration of zooplankton that may form part of a food chain that eventually reaches the corals. The majority of detailed studies concerning local environmental conditions in L. pertusa habitats have been conducted within the north-eastern Atlantic, limiting most knowledge of the niche of this species to a single part of an ocean basin. Data presented here show that the corals at VK826 are subjected to similar conditions in temperature, salinity, and flow velocity as their counterparts in the north-east Atlantic, although values for dissolved oxygen and density (sigma-theta: s Y ) are different. Our data also highlight novel observations of short-term environmental variability in cold-water coral habitat. & 2009 Elsevier Ltd. All rights reserved. 1. Introduction Communities dominated by the cold-water coral Lophelia pertusa are a common feature on hard substrata in the north- east Atlantic Ocean. Over 20 years of intensive study has revealed large reefs and carbonate mounds at a range of depths from 50 to over 2000 m (Freiwald and Roberts, 2005). In the north-west Atlantic Ocean, L. pertusa communities occur frequently on the upper continental slope (300–800 m) and are particularly abun- dant off the south-eastern United States (Ross and Nizinski, 2007). In contrast, L. pertusa in the Gulf of Mexico (GoM) appears to be more scattered and forms less dense communities than those off the south-eastern US or on the north-eastern Atlantic margins (Brooke and Schroeder, 2007; Ross and Nizinski, 2007). The ARTICLE IN PRESS Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/dsri Deep-Sea Research I 0967-0637/$ - see front matter & 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.dsr.2009.10.012 Corresponding author. Tel.: + 44 1248 382892 E-mail address: [email protected] (A.J. Davies). 1 Present address: Biology Department, Temple University, 1900 N 12th Street, Philadelphia, PA 19122, USA. Deep-Sea Research I 57 (2010) 199–212
Transcript of Short-term environmental variability in cold-water coral habitat at Viosca Knoll, Gulf of Mexico

ARTICLE IN PRESS
Deep-Sea Research I 57 (2010) 199–212
Contents lists available at ScienceDirect
Deep-Sea Research I
0967-06
doi:10.1
� Corr
E-m1 Pr
Street, P
journal homepage: www.elsevier.com/locate/dsri
Short-term environmental variability in cold-water coral habitat at VioscaKnoll, Gulf of Mexico
Andrew J. Davies a,�, Gerard C.A. Duineveld b, Tjeerd C.E. van Weering b,c, Furu Mienis b,Andrea M. Quattrini d,1, Harvey E. Seim e, John M. Bane e, Steve W. Ross d
a School of Ocean Sciences, Bangor University, Menai Bridge, Anglesey LL59 5AB, UKb Royal Netherlands Institute for Sea Research, PO Box 59, 1790 AB Den Burg, Texel, The Netherlandsc Free University of Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlandsd University of North Carolina at Wilmington, Center for Marine Science, 5600 Marvin K. Moss Lane, Wilmington, NC 28409, USAe Department of Marine Sciences, University of North Carolina at Chapel Hill, 340 Chapman Hall, Chapel Hill, NC 27599-3300, USA
a r t i c l e i n f o
Article history:
Received 26 June 2009
Received in revised form
15 October 2009
Accepted 2 November 2009Available online 10 November 2009
Keywords:
Lophelia pertusa
Deep-sea corals
Ecosystem engineer
Chemosynthetic communities
Environmental parameters
37/$ - see front matter & 2009 Elsevier Ltd. A
016/j.dsr.2009.10.012
esponding author. Tel.: +44 1248 382892
ail address: [email protected] (A.
esent address: Biology Department, Templ
hiladelphia, PA 19122, USA.
a b s t r a c t
The Lophelia pertusa community at Viosca Knoll (VK826) is the most extensive found to date in the Gulf
of Mexico. As part of a multi-disciplinary study, the physical setting of this area was described using
benthic landers, CTD transects and remotely operated vehicle observations. The site was broadly
characterised into three main habitats: (1) dense coral cover that resembles biogenic reef complexes,
(2) areas of sediment, and (3) authigenic carbonate blocks with sparse coral and chemosynthetic
communities. The coral communities were dominated by L. pertusa but also contained numerous
solitary coral species. Over areas that contained L. pertusa, the environmental conditions recorded were
similar to those associated with communities in the north-eastern Atlantic, with temperature
(8.5–10.6 1C) and salinity (�35) falling within the known species niche for L. pertusa. However,
dissolved oxygen concentrations (2.7–2.8 ml l�1) and density (sY, 27.1–27.2 kg m�3) were lower and
mass fluxes from sediment trap data appeared much higher (4002–4192 mg m�2 d�1). Yet, this species
still appears to thrive in this region, suggesting that L. pertusa may not be as limited by lower dissolved
oxygen concentrations as previously thought. The VK826 site experienced sustained eastward water
flow of 10–30 cm s�1 over the 5-day measurement period but was also subjected to significant short-
term variability in current velocity and direction. In addition, two processes were observed that caused
variability in salinity and temperature; the first was consistent with internal waves that caused
temperature variations of 0.8 1C over 5–11 h periods. The second was high-frequency variability
(20–30 min periods) in temperature recorded only at the ALBEX site. A further pattern observed over
the coral habitat was the presence of a 24 h diel vertical migration of zooplankton that may form part of
a food chain that eventually reaches the corals. The majority of detailed studies concerning local
environmental conditions in L. pertusa habitats have been conducted within the north-eastern Atlantic,
limiting most knowledge of the niche of this species to a single part of an ocean basin. Data presented
here show that the corals at VK826 are subjected to similar conditions in temperature, salinity, and flow
velocity as their counterparts in the north-east Atlantic, although values for dissolved oxygen and
density (sigma-theta: sY) are different. Our data also highlight novel observations of short-term
environmental variability in cold-water coral habitat.
& 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Communities dominated by the cold-water coral Lophelia
pertusa are a common feature on hard substrata in the north-
ll rights reserved.
J. Davies).
e University, 1900 N 12th
east Atlantic Ocean. Over 20 years of intensive study has revealedlarge reefs and carbonate mounds at a range of depths from 50 toover 2000 m (Freiwald and Roberts, 2005). In the north-westAtlantic Ocean, L. pertusa communities occur frequently on theupper continental slope (300–800 m) and are particularly abun-dant off the south-eastern United States (Ross and Nizinski, 2007).In contrast, L. pertusa in the Gulf of Mexico (GoM) appears to bemore scattered and forms less dense communities than those offthe south-eastern US or on the north-eastern Atlantic margins(Brooke and Schroeder, 2007; Ross and Nizinski, 2007). The

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212200
importance of L. pertusa in the deep sea is becoming increasinglyclear (Davies et al., 2007). For example, the ecosystem servicesprovided by L. pertusa reefs are now believed to mirror those ofshallow-water corals in terms of habitat provision and associateddiversity (e.g. Jensen and Frederiksen, 1992; Rogers, 1999; Reed,2002; Henry and Roberts, 2007).
L. pertusa is found at temperatures between 4 and 12 1C andsalinities of around 35 on the practical salinity scale (Freiwald,2002; Davies et al., 2008). Hard substrata are needed for initialgrowth, and water motion is required to provide food particles,keep the coral clear of sediment and resupply depleted dissolvedoxygen (Wilson, 1979; Duineveld et al., 2004; White et al., 2005;Davies et al., 2009). The chemistry of the surrounding water isalso important, with L. pertusa mostly found in areas saturatedwith aragonite (Oarag41) and within a density (sigma-theta: sY)envelope of 27.35–27.65 kg m�3 (Guinotte et al., 2006; Dulloet al., 2008). Given that L. pertusa growth extends over severalthousand years in many areas (Hall-Spencer et al., 2002; Freiwaldet al., 2004), the environment that L. pertusa occupies could beexpected to be relatively stable. However, recent studies haveshown that short-term variability, generated mostly by internalwaves, occurs in L. pertusa habitats (Duineveld et al., 2007; Mieniset al., 2007; Davies et al., 2009). These patterns are often missedbecause of the coarse sampling frequency and long deploymentsthat characterise many oceanographic studies in the deep sea.Deployments of short-duration instrumentation and high-frequency sampling have led to the discovery that tidal variabilitycan increase mixing across buoyancy layers or can act as a pumpthat draws surface or intermediate water to depth in coral habitat(Davies et al., 2009). The breaking of internal waves can alsosuspend local sediments, promote nutrient mixing and can driveupslope resuspension events (Frederiksen et al., 1992; Bonninet al., 2002; Hosegood et al., 2004). How these events drive thefunctioning and distribution of cold-water coral habitats arelargely unknown; however, an increasing body of work highlightsthe importance of short-term variability within shallow-watercoral ecosystems (Leichter et al., 2005, 2006; Wang et al., 2007;Piniak and Storlazzi, 2008).
In the present study, we describe the physical habitat of aportion of the Minerals Management Service lease block VioscaKnoll 826 (VK826), a site that contains the most extensive cold-water coral communities known in the GoM (Schroeder, 2002;Brooke and Schroeder, 2007). In order to address how the physicaloceanographic environment interacts with coral communities atVK826, we investigated the variability from high-frequencyphysical observations using benthic landers and also provideinformation on oceanographic conditions from CTD data. Pre-sently, our knowledge of the environmental conditions for cold-water corals in the GoM is incomplete. Hence, this area iscurrently the focus of a four year, multi-agency effort that buildsupon previous multi-disciplinary studies of cold-water coralecosystems on the slope of the GoM (Continental Shelf Associates,2007; Sulak et al., 2008).
2. Study area
2.1. Regional setting
The GoM is a semi-enclosed basin that has an area ofapproximately 1.5�106 km2 (Brooke and Schroeder, 2007) andis encircled by a broad, relatively flat shelf (o200 m). Moving tothe slope, the topography becomes increasingly complex, withnumerous knolls, basins, and canyons (Bryant et al., 1990). Thesubstrata of the area mostly consist of fine sediments; however,there is significant hard bottom in the form of precipitated
authigenic carbonates that are formed through biogeochemicalactivity in areas of active hydrocarbon seepage. These can persistafter seepage has discontinued (Formolo et al., 2004). Thesecarbonates are often colonised by cold-water corals and chemo-synthetic communities (Kennicutt et al., 1988; Schroeder et al.,2005; Cordes et al., 2008).
On the upper De Soto slope, located along the northern marginof the GoM basin (Fig. 1), a cold-water coral community known asVK826 has been intensively surveyed with remotely and human-operated vehicles (e.g. Schroeder, 2002; Continental ShelfAssociates, 2007; Cordes et al., 2008, 2009; Becker et al., 2009).There are two main topographic features at VK826 (Fig. 1); thefirst is an isolated 90-m-high knoll that contains outcrops ofauthigenic carbonate amongst sediment that ranges from gravellysandy mud to gravelly mud. The second knoll is a small moundthat lies approximately 1000 m north-east of the isolated knolland contains hard carbonate substrata amongst shell pavementsand fine sediments (Continental Shelf Associates, 2007). The closeproximity of two habitat-defining communities in this area,chemosynthetic species and cold-water corals, arises from twofactors, the presence of authigenic carbonate blocks amongstcomplex topography and the migration of sulphide-rich fluidsthat are associated with the uplift of these blocks (Schroeder,2002; Cordes et al., 2006). This is not the first report of cold-watercoral communities growing amongst seeps; parallels can bedrawn with observations from the Norwegian shelf (Hovlandet al., 1998). However, it is unlikely that these communities areconnected, as most evidence suggests that there are no trophiclinkages between cold-water coral and chemosyntheticcommunities (Duineveld et al., 2007; Becker et al., 2009).
2.2. Oceanographic setting
The complex topography of the GoM interacts with currentand water mass density fields that are equally complex. Circula-tion is mostly driven by the Loop Current, which enters throughthe Straits of Yucatan, flows in a looping, clockwise paththrough the eastern GoM and exits through the Straits of Floridain the south-eastern corner of the Gulf (Fig. 1; Sturges andLugo-Fernandez, 2005). Every several months, an anticyclonicallyswirling eddy (known as a Loop Current Ring (LCR)) detaches fromthe Loop Current and propagates generally westward through thebasin, impinging on the western continental slope and shelfbefore decaying in the western GoM (Smith, 1986; Hamilton et al.,1999; Sturges and Lugo-Fernandez, 2005). Cyclonically swirlingeddies are typically found along the outer boundaries of the LoopCurrent and LCRs. These ‘‘boundary eddies’’ are usually severaltens to around 100 km in diameter, with swirl speeds of 10 cm s�1
or more. In addition to the complexities of the Loop Current andits eddy fields, currents along the shelf edge in the northern andnorth-eastern GoM are impacted by winds and buoyancy-drivencoastal circulation (Carnes et al., 2008). Near the seabed on thecontinental slope, bottom currents are often intensified by aninterplay between the complex topography and density stratifica-tion, and speeds can reach maxima of 50 cm s�1 or more. Jochensand DiMarco (2008) suggested that such currents may beresponsible for particle transport in benthic areas.
Data from earlier observations at VK826 showed the coralcommunity was established within an oxygen minimum layer(between 2.6 and 3.2 ml l�1) in the 400–500 m depth range(Schroeder, 2002), a defining characteristic of Tropical AtlanticCentral Water (Jochens and DiMarco, 2008). During an 11-dayrecord (22 July–02 August 2004), water temperature was7.5–9.0 1C with a mean of 8.3 1C. A northward current wasrecorded during this period with a mean speed of 5.8 cm s�1

ARTICLE IN PRESS
88°3.0’W 88°2.0’W 88°1.0’W 88°0.0’W 87°59.0’W
29°9.0’N
29°10.0’N
29°11.0’N
ALBEX lander
BOBO lander
High resolution CTD transect
CTD yo-yo
ROV-2008-001
ROV-2008-002
ROV-2008-006
ROV-2008-004
ROV-2008-005
a b
Fig. 1. Location, bathymetry (5 m resolution multibeam), and sampling stations in the Gulf of Mexico. Inset (a) shows the location of the VK826 area in the Gulf of Mexico
and the generalised flow of the Loop Current. The main map shows the locations of the ROV transects (the red box is shown as inset (b) for clarity), the two benthic landers,
the CTD surveys (the cross-shelf CTD transect is not shown), and seismic lines (Lines A, B, C) including their labelled structures in Fig. 2. (For interpretation of the references
to colour in this figure legend, the reader is referred to the web version of this article.)
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212 201
and a maximum speed of 18.3 cm s�1 (Continental Shelf Associ-ates, 2007). In a longer term study (12 months duration), Teagueet al. (2006) and Hallock et al. (2009) observed currents to flowgenerally along isobaths and fluctuate with several importantperiods, ranging from near-inertial (about 1 day) to over 60 days.Significant energy fluctuation was found at periods near 16 and21 days, and these motions were interpreted to be bottom-intensified topographically trapped waves that propagate towardsthe west (Hallock et al., 2009). The fact that near-bottom currentsduring this one-year study were observed to flow generally alongthe isobaths suggests that the observations of consistentlynorthward (across-isobath) flow in the shorter duration, 11-daystudy (Continental Shelf Associates, 2007) were not representa-tive of usual conditions at VK826, a site that is situated in a regioncharacterised by considerable hydrodynamic variability.
3. Materials and methods
3.1. Acoustic mapping
Multibeam mapping was conducted using a Kongsberg-SimradEM1002 capable of mapping to 1000 m water depth. The systemwas a 95 kHz echosounder that used 111 beams per ping over amaximum coverage sector of 1501 (beam spacing was equidi-stant). Raw data were tidally corrected and postprocessed usingCARIS HIPS and SIPS (version 6.1) to produce bathymetric andbackscatter maps. To supplement the multibeam mapping, threesingle-beam sonar transects were conducted over the majorVK826 coral mounds to determine topography and surficialsediments. Using a 3 kW Knudsen 3200 single-beam sounder
operating at 12 kHz, three parallel tracks were surveyed with thesounder to cover: (1) the northern mounds east of a pockmark,(2) the soft sediment plain between mounds (passing over theplotted position of the BOBO lander, see Landers below), and(3) the authigenic carbonate blocks to the south (passing over theplotted position of the ALBEX lander; Fig. 1).
3.2. ROV data
Video ground truthing data were acquired using a SeaeyeFalcon DR remotely operated vehicle (ROV) deployed for a totalof five dives. Initially, each dive was targeted towards areaswith known or suspected coral occurrence but once at bottom,the dive attempted to transect across as many habitats aspossible. The main visual information was provided by a bowmounted video camera (SeaEye low-light, 0.2 lux) lit by a largeLED array. Digital still images of habitat and fauna were takenperiodically using a seven megapixel camera with internalstrobe (Imenco). The ROV was tracked using a LinkQuestTrackLink USBL system, with the resulting feed of time, depth,latitude, and longitude overlaid onto the recorded video.Transect distances varied depending on habitat, bottom currents,and surface seas. On each ROV deployment, a conductivity,temperature, depth (CTD) system (SBE 19+ profiler, Sea-Bird)including auxiliary sensors for turbidity (Seapoint), pH (SBE18),and fluorescence (Seapoint) was mounted on top of the ROVgarage. The garage was kept close to the bottom during alldeployments. Readings were edited after collection to show onlythe data from when the garage was on or near bottom (0–30 mabove bottom).

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212202
3.3. CTD
On 05 and 06 October 2008, a CTD transect was made acrossthe shelf of the GoM using an SBE 911+ profiler includingauxiliary sensors for turbidity (Seapoint), fluorescence (Seapoint),and oxygen (SBE43). The transect originated at 29124.610N,87159.260W (2037 CDT or 0137 UTC) at 74 m depth, with fiveCTD stations at approximately 10 km intervals, ultimately endingat 29104.3830N, 88101.9730W (0401 CDT) at 1100 m depth. On 12October 2008 at 0754 CDT, an eight-station CTD transect wasconducted at a higher spatial resolution (500 m) over the VK826
I IISediment drape Carbonate blocks and slabs and c
coral patches
Knolls
Erosionalchannel
I IISediment drape Thin sediment drape
covering carbonate blocks
Sediment drape Carbthin
I
Line A
Line B
Line C
400 m
420 m
450 m
460 m
480 m
500 m
520 m
540 m
560 m
580 m
600 m
400 m420 m450 m460 m480 m500 m520 m540 m560 m580 m600 m
400 m
420 m
450 m
460 m
480 m
500 m
520 m
540 m
560 m
580 m
600 m
Fig. 2. Interpreted single-beam echosounder lines crossing the VK826 knolls (see Fig. 1
panels are orientated from west to east.
coral mounds (Fig. 1) and took approximately 9.5 h to complete.At each station along the transect, the CTD was lowered andraised from the seafloor before moving to the next station. Todetermine any temporal trends and possibly identify any regularactivity such as internal waves, a repeated stationary yo–yo CTD(12 casts in total) was conducted over a 5 h period at 29109.390N,88101.210W (starting at 0004 CDT) directly over the coral moundsat approximately 530 m depth. Vertical profiles of temperature,fluorescence, oxygen, density (sY), and turbidity were plottedagainst distance for transects and time for the yo–yo usingSurfer 8 (Golden Software).
III IVold water
Thin cover of sedimenton carbonate blocks
Sediment drape
III IVSediment drape
Knoll structurepossibly with corals
Thin sediment drapecovering carbonate blocks
onate blocks with sediment cover
Knoll structure
Cold water coralreef-like growth
Sediment drape
II III IV
for location of each line). Structures described within the text are labelled I–IV; all

ARTICLE IN PRESS
Fig. 3. Images of habitats taken by the ROV at VK826. The following are from areas with a dense cover of L. pertusa and antipatharians that resemble biogenic reefs:
(a) L. pertusa and unidentified crinoids. (b) L. pertusa and the antipatharian Leiopathes c.f. glaberrima. (c) Anemones, Family Actinostolidae, the gorgonian Keratoisis sp. and
the fish Gephyroberyx darwinii. (d) The brisingid Novodinia antillensis and L. pertusa. The following are from areas with sediments: (e) the fish Chaunax suttkusi and
unidentified anemones. (f) The fish Nezumia aequalis and unidentified anemones. The following are from authigenic carbonate blocks, colonised by solitary coral colonies,
small patches of L. pertusa and some tubeworms and bivalve shells that may indicate some chemosynthetic activity. (g) L. pertusa, the chirostylid Eumunida picta, the urchin
Echinus tylodes, and the fish Conger oceanicus. (h) L. pertusa on eroded authigenic carbonate and bivalve shells. (i) Unidentified vestimentiferan tubeworms on authigenic
carbonate. (j) Authigenic carbonate.
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212 203
ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212204
3.4. Landers
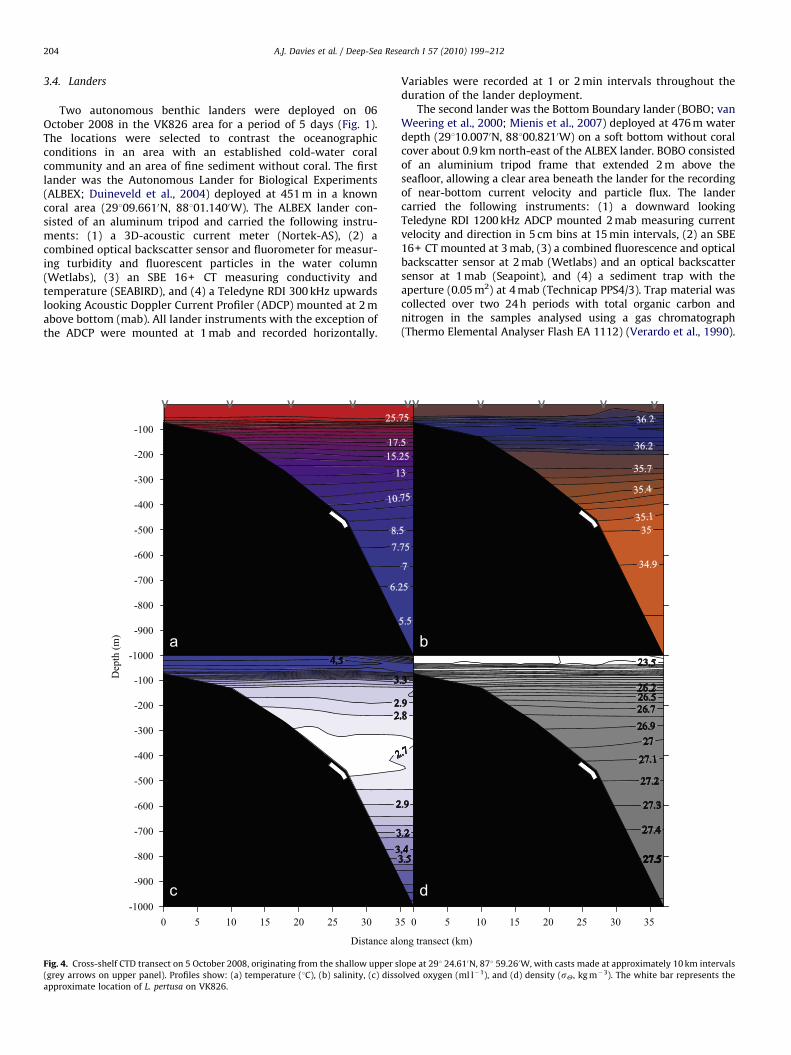
Two autonomous benthic landers were deployed on 06October 2008 in the VK826 area for a period of 5 days (Fig. 1).The locations were selected to contrast the oceanographicconditions in an area with an established cold-water coralcommunity and an area of fine sediment without coral. The firstlander was the Autonomous Lander for Biological Experiments(ALBEX; Duineveld et al., 2004) deployed at 451 m in a knowncoral area (29109.6610N, 88101.1400W). The ALBEX lander con-sisted of an aluminum tripod and carried the following instru-ments: (1) a 3D-acoustic current meter (Nortek-AS), (2) acombined optical backscatter sensor and fluorometer for measur-ing turbidity and fluorescent particles in the water column(Wetlabs), (3) an SBE 16+ CT measuring conductivity andtemperature (SEABIRD), and (4) a Teledyne RDI 300 kHz upwardslooking Acoustic Doppler Current Profiler (ADCP) mounted at 2 mabove bottom (mab). All lander instruments with the exception ofthe ADCP were mounted at 1 mab and recorded horizontally.
-1000
-900
-800
-700
-600
-500
-400
-300
-200
-100
V V V V
0 10 15 20 25 30 3-1000
-900
-800
-700
-600
-500
-400
-300
-200
-100
Dep
th (m
)
Distance al
Dep
th (m
)
5
Fig. 4. Cross-shelf CTD transect on 5 October 2008, originating from the shallow upper s
(grey arrows on upper panel). Profiles show: (a) temperature (1C), (b) salinity, (c) diss
approximate location of L. pertusa on VK826.
Variables were recorded at 1 or 2 min intervals throughout theduration of the lander deployment.
The second lander was the Bottom Boundary lander (BOBO; vanWeering et al., 2000; Mienis et al., 2007) deployed at 476 m waterdepth (29110.0070N, 88100.8210W) on a soft bottom without coralcover about 0.9 km north-east of the ALBEX lander. BOBO consistedof an aluminium tripod frame that extended 2 m above theseafloor, allowing a clear area beneath the lander for the recordingof near-bottom current velocity and particle flux. The landercarried the following instruments: (1) a downward lookingTeledyne RDI 1200 kHz ADCP mounted 2 mab measuring currentvelocity and direction in 5 cm bins at 15 min intervals, (2) an SBE16+ CT mounted at 3 mab, (3) a combined fluorescence and opticalbackscatter sensor at 2 mab (Wetlabs) and an optical backscattersensor at 1 mab (Seapoint), and (4) a sediment trap with theaperture (0.05 m2) at 4 mab (Technicap PPS4/3). Trap material wascollected over two 24 h periods with total organic carbon andnitrogen in the samples analysed using a gas chromatograph(Thermo Elemental Analyser Flash EA 1112) (Verardo et al., 1990).
V
5 0 10 15 20 25 30 35
ong transect (km)
V V V V
5
V
lope at 291 24.610N, 871 59.260W, with casts made at approximately 10 km intervals
olved oxygen (ml l�1), and (d) density (sY, kg m�3). The white bar represents the

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212 205
4. Results
4.1. Physical habitat
The VK826 knolls are elevated features that rise fromapproximately 450–550 m water depth from the slope in theGoM. The remotely sensed data including ROV footage, single-beam, and multibeam echosounder, defined three distinct macro-habitats in the area, characterised by differences in topographicmorphology and cold-water coral density: (1) dense coral coverresembling biogenic reef, (2) well-layered sediment habitat, and(3) authigenic carbonate blocks with sparse coral and chemosyn-thetic communities (Figs. 1–3).
The first seismic line was recorded along the southern edge ofthe pockmark (Figs. 1 and 2a). This line was characterised by awell-developed sediment drape (Fig. 2a, structure I), followed bycarbonate blocks that had a thin sediment cover (structure II). Tothe east of the pockmark was a small knoll with a summit at470 m water depth (structure III). This knoll was covered with adense cold-water coral community and associated fauna thatresembles a typical biogenic reef structure (Figs. 3a–d). L. pertusa
was the dominant framework building coral, but other species,including the antipatharian Leiopathes c.f. glaberrima andgorgonian Keratoisis sp., were often observed in ROV footage.The lower flank of the structure was characterised by coral rubble,partially covered by sediment, solitary coral colonies, and fields ofanemones (Family: Actinostolidae; Figs. 3a–d).
The second seismic line crossed a low relief area between thepockmark and the two knolls situated to the south, and transectedalmost over the BOBO lander site (Fig. 2b). The line was flanked tothe west and east by well-layered sediments (Fig. 2b, structures I
-600
-500
-400
-300
-200
-100
0 1000 2000 3000
-600
-500
-400
-300
-200
-100
0 1000
Distance al
Dep
th (
m)
V V V V V VV V V V
Fig. 5. High resolution CTD transect taken on 12 October 2008. The white bar represent
show individual CTD locations at 500 m spacing. The values along the x-axis represent t
temperature (1C), (b) salinity, (c) turbidity (FTU, formazin turbidity units), (d) dissolve
and IV) with a central feature consisting of thin sediment drapeover carbonate blocks (structures II and III). This was capped by aslightly elevated knoll-like structure that may contain corals(structure III). ROV images taken in the area of the BOBO landerconfirm the presence of surficial sediments colonised bycnidarians and an evidence of biological activity (lebensspuren;Figs. 3e and f). The third seismic line was recorded over the top oftwo knolls and the location of the ALBEX lander (Fig. 2c). Thesummits of the knolls were at 440 and 460 m water depth andwere covered by broken and eroded carbonate blocks and slabs(Fig. 2c, structures II and III). These features were often severalmeters high, providing steep local relief (Figs. 3i and j). In severalROV images, the presence of vestimentiferan tubeworms andaggregations of bivalve shells are likely related to a chemosyn-thetic activity (Figs. 3h–j). Although the authigenic carbonatesand elevated topography in this area form suitable substrata forcoral colonisation, the overall density of colonies of solitary coralsand L. pertusa was much less than that observed in the area of thefirst seismic line (Figs. 3g and h).
4.2. CTD
The cross-shelf CTD transect showed a relatively stablehydrography downslope with a prominent thermocline at 55 m(Fig. 4). Temperatures ranged from 27 1C at the surface to 4.7 1C at1083 m. Salinity ranged between 34.9 and 36.5, and sY wasbetween 23.4 and 29.6 kg m�3. Dissolved oxygen was 4.3 ml l�1 atthe surface, decreasing below the thermocline to a minimum of2.6 ml l�1 at approximately 500 m before rising to 4.4 ml l�1 at thedeepest measurement of 1083 m. Intersecting the CTD data at thedepth contour where corals were found at VK826 (between 400
2000 3000 0 1000 2000 3000
ong transect (m)
V V V V VV V V V V V V
s the approximate location of coral framework and grey arrows in the upper panel
he distance from the start at the north end of the transect. Recorded variables: (a)
d oxygen (ml l�1), (e) density (sY, kg m�3), and (f) fluorescence (Chl a, mg l�1).

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212206
and 500 m), temperature was 8.5–10.6 1C, salinity 35.0–35.3, sY27.1–27.2 kg m�3, and dissolved O2 was 2.6–2.8 ml l�1 (Fig. 4).
Smaller scale patterns were resolved by a higher resolutionCTD transect over the knolls (Fig. 5). Most evident was anintermediate nepheloid layer that occurred between 300 and400 m, directly over the VK826 mounds. There also appeared to bean increase in turbidity downslope of the knolls (Fig. 5c), but nofluorescent signal was observed below 150 m (Fig. 5f). The 5 h yo–yo CTD showed no temporal variation in fluorescence and someminor fluctuations in temperature, salinity, sY, and dissolved
-500
-450
-400
-350
-300
-250
-200
-150
-100 V V V V VV V V VV
0 100 200 300
-500
-450
-400
-350
-300
-250
-200
-150
0 100
Dep
th (
m)
Time since
V V
Fig. 6. Five hour CTD yo–yo series made on 7 October 2008 on top of VK826 (see Fig. 1).
(b) salinity, (c) turbidity (FTU, formazin turbidity standard), (d) dissolved oxygen (ml l
oxygen (Fig. 6). Turbidity showed three consistent highs at 150,240, and 450 m that all appeared to strengthen towards the end ofthe yo–yo (Fig. 6c). The ROV garage mounted CTD recorded datanear the seafloor on all five dives (Table 1). The first dive(ROV-2008-001) transected over the large knoll at the south nearthe location of the ALBEX lander and measured variables over a37 m vertical range, recording a mean temperature of 9.3 1C andpH of 7.96. The second dive (ROV-2008-002) crossedapproximately south across the plain between the pockmarkand the location of the BOBO lander. This dive covered the
V V V V V V VV
200 300 0 100 200 300
2245 UTC (min)
V V V V
Grey arrows represent the individual CTD casts. Profiles show (a) temperature (1C),�1), (e) density (sY, kg m�3), and (f) fluorescence (Chl a, mg l�1).

ARTICLE IN PRESS
Table 1ROV stations sampled at VK826, including a summary of data recorded from the garage mounted CTD (depth, m; temperature, 1C; salinity; pH).
ROV dive Date Location Depth Temperature Salinity pH
ROV-2008-001 07 October 2008 Start Minimum 414.52 8.83 35.01 7.92
29109.664/88101.095 Maximum 451.96 10.14 35.25 8.01
End Mean 440.42 9.30 35.11 7.96
29109.939/88101.009 SD 8.27 0.36 0.05 0.03
ROV-2008-002 08 October 2008 Start Minimum 434.44 8.25 34.95 8.04
29109.951/88100.793 Maximum 485.09 11.10 35.39 8.10
End Mean 458.67 9.52 35.14 8.07
29109.386/88100.911 SD 14.00 0.73 0.09 0.02
ROV-2008-004 10 October 2008 Start Minimum 455.72 8.30 34.97 7.94
29110.185/88100.683 Maximum 480.62 8.90 35.11 7.97
End Mean 470.63 8.43 35.01 7.96
29110.289/88100.604 SD 3.49 0.07 0.01 0.01
ROV-2008-005 11 October 2008 Start Minimum 462.83 8.68 34.99 7.88
29110.196/88100.101 Maximum 480.07 9.15 35.17 7.99
End Mean 472.23 8.84 35.05 7.95
29110.216/88100.730 SD 3.47 0.06 0.01 0.04
ROV-2008-006 11 October 2008 Start Minimum 467.32 8.24 34.99 7.92
29110.221/88100.647 Maximum 478.53 8.71 35.08 7.98
End Mean 477.36 8.41 35.01 7.97
29110.215/88100.623 SD 0.54 0.07 0.01 0.01
10 15 20 250
20
40
60
80
100
120
140
Hei
ght a
bove
bot
tom
(m
)
-5 0 5
Current velocity (cms-1)
Fig. 7. The average vertical profiles of currents in the east-west (a) and north-south (b) direction measured by the upward-looking ALBEX ADCP. The slanting dashed line
shows an approximate linear fit to the upper section of the profile and is used to estimate the zero-crossing elevation (roughly 200 mab) and the height to which friction
influences the velocity structure at about 70 mab. It also highlights the bottom-intensified character of the current. The horizontal dashed line marks the top of the
assumed friction layer, below which the eastward velocity departs from its linearly increasing speed and the current vector veers counter-clockwise with increasing depth.
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212 207
greatest vertical range of over 50 m and recorded a slightly highermean temperature of 9.5 1C and pH of 8.07 compared to the othersites. The remaining three dives (ROV-2008-004-006) wereconcentrated in the densest coral area at the small knoll east ofthe pockmark and recorded similar values for all variables. Thesedives were concentrated over the smallest vertical range, between11 and 25 m, and recorded mean temperatures of between 8.4 and8.8 1C and salinities of around 35. pH was consistent throughoutall three dives with values between 7.95 and 7.97.
4.3. Landers
It is possible to define three aspects of the water flow at VK826based on the 5 days of high-frequency data collected by thelanders: (1) the sub-inertial flow (low-frequency), (2) thefrictional bottom boundary layer, and (3) super-inertial flow(high-frequency). The upward-looking ADCP mounted on theALBEX lander indicated the presence of a bottom-intensifiedeastward current that had a vertical scale of approximately 200 m

ARTICLE IN PRESS
-10
0
10
20
30
40
-20
-10
0
10
20
8
8.5
9
9.5
10
06 Oct00:00
07 Oct00:00
08 Oct00:00
09 Oct00:00
11 Oct00:00
12 Oct00:00
13 Oct00:00
34.8
35
35.2
35.4
Eas
t-w
est c
urre
ntve
loci
ty (
cms-
1 )
Nor
th-s
outh
cur
rent
velo
city
(cm
s-1
)Te
mpe
ratu
re (
°C)
Salin
ity
Date/Time (UTC)
Fig. 8. Time-series of east-west (a) and north-south (b) current components at 50 mab from the upward looking ALBEX ADCP (red lines), and from the BOBO ADCP at 1 mab
(blue lines), and the 40 hour low-passed of the ALBEX ADCP record (dashed). Time-series of temperature (c) and salinity (d) from the landers. (For interpretation of the
references to colour in this figure legend, the reader is referred to the web version of this article.)
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212208
(the estimated height above bottom of the zero-crossing of theeastward current, Fig. 7a). This current strengthened from 15 to30 cm s�1 on the first day of the deployment and maintained30 cm s�1 over the subsequent days (Fig. 8a). This increase inflow speed, probably linked to a different water mass, wasaccompanied by corresponding decreases in temperature andsalinity (Fig. 8c).
The ALBEX lander was positioned amongst the coral frame-work, and the horizontally looking current meter (1 mab)recorded a median near-bottom current speed of 6.7 cm s�1
(SD 3.1) with the majority of current speeds below 15 cm s�1
and a maximum of 34 cm s�1. Currents recorded at the BOBOlander site were stronger, with a median current speed of20.6 cm s�1 (SD 5.2) at 1 mab. Contrasting the results from theADCP instruments mounted on both landers showed that the
currents at 1 mab on the BOBO were similar, but were of a lessermagnitude and with a more counter-clockwise heading relative tothe currents recorded 50 m above the ALBEX lander (Figs. 8aand b). The shape of the vertical profiles of velocity componentsfrom the ADCP on ALBEX suggested that veering began atapproximately the same elevation as the velocity maximum,and this feature could be interpreted as the height of a frictionallyinfluenced bottom boundary layer, approximately 70 m above thepeak of the larger knoll (Fig. 7). Both characteristics wereconsistent with a bottom Ekman layer with a thickness greaterthan the height of the topographic features of the knoll.
Super-inertial current variability was evident in the observa-tions from the landers, but it was relatively weak over the shorttime-series. Root-mean-square currents of 40 h high-pass filteredcurrent components were approximately 3 cm s�1. A temperature

ARTICLE IN PRESS
12:00 15:00 18:00 21:00 00:008.4
8.6
8.8
9
Tem
pera
ture
(°C
)
Time (hours)
Fig. 9. Time-series of high-frequency temperature records (sampling interval 2 min, with the exception of the BOBO ADCP at 15 min) from the landers on 10 October 2008.
The three temperature sensors on the ALBEX lander register high-frequency fluctuations (blue line, CTD; green, current meter; aquamarine, ADCP) but these were not
observed at the BOBO lander site (red line, ADCP; purple, CTD). (For interpretation of the references to colour in this figure legend, the reader is referred to the web version
of this article.)
(dB
)
Period (hours)
Spec
tral
uni
ts
45
50
55
60
65
7006 Oct 00:00
07 Oct 00:00
08 Oct 00:00
09 Oct 00:00
10 Oct 00:00
11 Oct 00:00
180
160
140
120
100
20
40
60
80
Hei
ght a
bove
bot
tom
(m
)
12 Oct 00:00
302418126
0
1
2
3
4
5
Date / Time (UTC)
Fig. 10. (a) Volume scattering strength (in dB) as a function of time from the upwards looking ADCP mounted on the ALBEX lander within the coral mounds (dashed line
shows the bin used for the spectrogram). (b) Spectrogram showing the synchronicity of diel vertical migration shown using CLEAN spectral analysis (Rosato and Kyriacou,
2006) of backscatter from the 120 m bin (shown as dashed line in Fig. 8a). There are two periodic components above the 99% confidence limit (dashed line): 8.2 and 24 h
(the daily migration pattern).
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212 209
minimum of 8 1C was recorded by the BOBO lander starting at0000 UTC on 09 October 2008 and was also reflected to a lesserextent at the ALBEX site (Figs. 8c). The southward flow thataccompanied the temperature decrease suggests an origin to thenorth of the BOBO lander (Fig. 8b and c). The pockmark featurewas the most likely source of the colder water, as it has atopographic connection to deeper waters through a trough to thewest (Fig. 1). After the temperature minimum event, temperatureand salinity at both sites exhibited coherent fluctuations atperiods between 5 and 11 h (Fig. 8c). These fluctuations are likelycaused by internal waves, and reflect the stratification of the areabut are probably episodic. Throughout the entire 5-day time-series, temperature ranged between 8.5 and 9.5 1C (maximumchange was 0.8 1C over 8 h, Fig. 8c), and salinity varied between35.03 and 35.14 (the offset between the salinity readings of thelanders may be the result of calibration error; Fig. 8d). Very high-frequency temperature oscillations (20–30 min periods) wererecorded by all three temperature sensors on the ALBEX lander,but were not observed in sensors on the BOBO lander (Fig. 9).These fluctuations were asymmetric, typically beginning with arapid temperature rise of 0.1–0.2 1C, followed by a slowertemperature decline.
From the lack of a discernable pattern in fluorescence fromlanders or CTD data (Figs. 4–6), it appears that there was no freshmatter input from surface sources during our observations.However, one alternative food source may be migrating zoo-plankton above the coral mounds. The upwards looking ADCPmounted on the ALBEX lander located within the coral mounds
showed a clear diel vertical migration in the volume scatteringstrength between 100 and 180 m above bottom (Figs. 10a). Thescatterers were likely migrating nekton (probably zooplankton)that moved downwards at sunrise and rose through the watercolumn at sunset following a 24 h cycle (Fig. 10a and b). Materialcollected in the sediment trap of the BOBO lander showed highmass fluxes up to 4192 g m�2 d�1. The sizing of these particlesshowed they were mostly fine-grained aggregates (95% waso63mm) and included particles that consisted of 2.4% totalorganic carbon and 0.4% total nitrogen.
5. Discussion
The cold-water coral communities at VK826 are dominated byL. pertusa interspersed with colonies of octocorals, antipatharians,and other filter-feeders. Our investigation of the knolls showedthat they are generally not similar in composition to thecarbonate mounds found throughout the north-eastern Atlantic(e.g. De Mol et al., 2002). Rather, the cold-water coral commu-nities colonised available hard substrata in the form of authigeniccarbonate blocks (Schroeder, 2002; Cordes et al., 2008). L. pertusa
formed dense biogenic structures east of the pockmark (approxi-mately 29110.230N, 8810.690W). This area contained the mostextensive coral community at VK826, and whilst carbonate blocksoccurred there, the habitat (consisting of living coral framework,trapped sediments, and surrounding coral rubble) was similar tothat of bioherms off North Carolina and the north-east Atlantic

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212210
(e.g. Wilson, 1979; Ross and Nizinski, 2007). Sulak et al. (2007)claimed that coral rubble was essentially absent at Vioska Knoll.In our observations, coral rubble (medium to small fragments ofdead, detached L. pertusa) was fairly common within the coralhabitat to the east of the pockmark (referred to as the ‘‘Big BlueReef’’ site in Sulak et al., 2007), but was less common where coralscolonised existing hard substrata. The larger knolls to the southcontained less extensive coral communities but contained someevidence of chemosynthetic communities in the form of vesti-mentiferan tubeworms (likely to be Lamellibrachia luymesi andSeepiophila jonesi, after Cordes et al., 2006) and accumulations offossil bivalve shells. The lack of new chemosynthetic growthobserved in the VK826 area likely suggests that active seepage isreduced and these communities are entering a senescent phase(Continental Shelf Associates, 2007).
The environmental conditions at VK826 were relatively similar tomany previously studied L. pertusa sites, and broadly reflectedsimilar values reported for communities within the North Atlantic.Salinity and temperature were both consistent with observationswithin the north-east Atlantic and with meta-analyses of the speciesniche (Freiwald et al., 1997; Duineveld et al., 2004; Davies et al.,2008; Dullo et al., 2008). Notable exceptions were the lower sY of27.1–27.2 kg m�3 that fell outside of the envelope found in theNorth Atlantic (27.35–27.65 kg m�3; Dullo et al., 2008), and thelower levels of dissolved oxygen at the VK826 mounds relative tocommunities found elsewhere. Dissolved oxygen concentrationsrecorded at VK826 were 2.7 ml l�1 from ship-taken CTD casts. Thiswas similar to previous records for dissolved oxygen in this region(Schroeder, 2002), and was within the range of concentrationsobserved at this depth in the GoM (see Jochens and DiMarco, 2008;Vidal et al., 1994). Values from other sites throughout the NorthAtlantic are often higher. For example, the Sveinsgrunnen andFugløy Reefs (Norway) have recorded values between 6.52 and6.64 ml l�1. In the Porcupine Seabight, dissolved oxygen concentra-tions were closer to the GoM, with values between 3.76 and4.60 ml l�1 (Dullo et al., 2008). Experimental analysis withL. pertusa samples collected in the north-east Atlantic suggests thatthe critical PO2 at 9 1C is 3.26 ml l�1. Below this, the normal aerobicfunction will be impaired (Dodds et al., 2007). It is possible thatL. pertusa communities throughout the GoM are limited by thedissolved oxygen concentration. Recently, a mean growth rate of3.77 mm yr�1 was reported for L. pertusa at VK826 (Brooke andYoung, in press), which is among the lowest published for thisspecies (Brooke and Young, in press), possibly indicating thatL. pertusa at VK826 may be occurring towards the extreme of itstolerance with respect to dissolved oxygen. However, comparison ofL. pertusa growth rates and eco-physiology is problematic at presentdue to a lack of standardisation, lack of direct in situ measurements,and only few published studies.
The possibility that L. pertusa could have a lower metabolicrate in the GoM due to lower oxygen availability may also reflectlinkages with other physical characteristics found in the area. Thelack of a fluorescent signal at depth at VK826 is surprising,considering the fact that the surface productivity above the localarea is relatively high compared to the general conditionsthroughout the GoM (Continental Shelf Associates, 2007). Thissuggests that the physical mechanisms of surface to depthtransport observed throughout the north-east Atlantic coralcommunities may not be present at VK826 (e.g. Duineveld et al.,2004, 2007; Mienis et al., 2007; Davies et al., 2009), althoughlonger term benthic observations are needed to test thishypothesis. It is possible that the majority of food particles aretransported by lateral advection or sinking. One process revealedby the upwards looking ADCP at VK826 was a diel verticalmigration of what appears to be zooplankton that deepens duringdaylight (to near bottom) and rises during darkness (over a 24 h
cycle). These migrants can be an important vector for the flux ofcarbon and nitrogen to depth (Angel and Pugh, 2000). Data fromthe sediment trap on the BOBO lander showed that fluxes atVK826 were high compared to other coral locations on the RockallTrough margin (on-mound mean of 381 mg m�2 d�1, off-moundmean of 1411 mg m�2 d�1; Mienis et al., 2009). Another study inthe GoM also found high fluxes at VK826 when compared to othersites in the area, but the report did not provide sufficient data fordirect comparison with the BOBO sediment trap data (ContinentalShelf Associates, 2007). The BOBO sediment trap materialconsisted of fine-grained aggregates of riverine origin amongstparticles with high organic carbon content. It is possible thatfaecal pellets and other organic material from the migrants (orother nekton) may be responsible for increased carbon content,potentially providing a food source for cold-water corals. This issupported by recent isotopic analyses of L. pertusa communities inthe GoM that show no reliance upon the seeping hydrocarbonsthat are widespread throughout the area; instead they appear tobe part of a food web based on phytoplankton-derived particulateorganic matter (Becker et al., 2009).
During the 5-day deployment at VK826 the steady flowtowards the east was likely a portion of one cycle of a topographicwave with a two- to three-week-long period, consistent withprevious observations at VK826 of bottom-trapped topographicwaves that can dominate sub-inertial variability (Hallock et al.,2009). From the short-term data, the flow velocities in coral areasat VK826 were similar to observations from other cold-watercoral habitats in the North Atlantic (median 6.7 cm s�1, maximum34 cm s�1). For example, on the Rockall Bank, median flowswithin the coral habitat were 9 cm s�1 with a maximum flow of30 cm s�1 (Duineveld et al., 2007). One consistent theory regard-ing food supply mechanisms at cold-water coral habitats is thedependence upon advection from intensified currents that areaccelerated by topography or the presence of features such asinternal waves that promote mixing (Genin et al., 1986;Frederiksen et al., 1992; White et al., 2005, 2007; Davies et al.,2009). Often overlooked is the strength of the accelerated currentand how coral responds to different velocities. Several studieshave shown that flow velocities within coral habitat are oftenlower than areas without it (Duineveld et al., 2007), but furtherwork is required to show the finer scale interaction between coraland flow velocity.
Whether cold-water corals are subjected to short-termvariability and how they interact with such variability are ofincreasing interest (Duineveld et al., 2004, 2007; Mienis et al.,2007; Davies et al., 2009). Our data from VK826 uncoveredseveral examples of variability over short timescales, fromminutes to days. Firstly, consistent high-frequency variabilityin temperature over 30 min periods was observed at thelocation of the ALBEX lander (within coral habitat). The originof this variability is currently unclear. It could be a high-frequency internal wave; however, the lack of coherencebetween the two lander sites suggests that this is a localphenomenon. The second example was the presence of varia-tion driven by internal waves that interact with abrupttopography in the area. These waves resulted in temperaturevariations of 0.5 1C over 30 min and ultimately 0.8 1C over 8 h,with fluctuations exhibiting 5–11 h cycles. Within theL. pertusa community, these high-frequency variations couldpotentially influence ecological and behavioural processes suchas feeding and thereby ultimately effect the physiology ofL. pertusa, which is known to be strongly linked to temperature(Dodds et al., 2007). However, the general lack of studies onhow L. pertusa interacts with its local environment needs to beaddressed in order to identify the processes that control orinfluence this species.

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212 211
6. Conclusion
The majority of detailed studies into the environmentalconditions within L. pertusa habitat have been conducted in thenorth-east Atlantic, limiting most of our knowledge to a singlepart of an ocean basin. Our data show that the corals at VK826 aresubjected to similar conditions to those recorded in the north-eastAtlantic and fall within the proposed species niche (Davies et al.,2008). One major difference is that the dissolved oxygenconcentrations at VK826 are lower than those found in thenorth-east Atlantic, but cold-water corals still appear to thrivehere with dense frameworks and high associated diversity(Cordes et al., 2008). It does appear that the lower oxygenconcentrations may be reflected in a reduced growth ratecompared to other sites (Brooke and Young, in press). However,further observations that include measures of food supply areneeded to determine whether it is low dissolved oxygen thatconstrains growth in this area. Our data are part of longer termobservations into the cold-water coral communities at VK826specifically aimed to address several hypotheses about theirinteraction with environmental conditions in the GoM. Thesefurther in situ observations will reveal the seasonal processes thatoccur in the GoM, determine particle flux and origin, andultimately lead to a better understanding of how this cold-watercoral habitat functions and how it relates to other habitatsthroughout the world.
Acknowledgements
General funding was provided by the US Geological SurveyOuter Continental Shelf Ecosystem Studies Program, and shiptime was provided by the NOAA Undersea Research Center(UNCW) through a grant to SWR. Funding to SWR was providedby USGS (Cooperative Agreement 05HQAG0099, Subagreement5099HS0013). AJD was sponsored through a grant from theWinston Churchill Memorial Trust. We appreciate the efforts ofthe ship’s officers and crew on the R.V. Nancy Foster. The scientificteam is thanked for their assistance (Stephen Artabane, MikeCarlson, Amanda Demoploulos, Mike Grey, Martha Nizinski, andAdela Roa-Var �on). The ROV was provided by SeaVision throughfunding by USGS. Multibeam image was provided by Erik Cordes,courtesy of the US Minerals Management Service and NOAA OERLophelia II project. We also thank three anonymous reviewers andassociate editor Prof. A. Gooday for valuable comments on themanuscript.
References
Angel, M.V., Pugh, P.R., 2000. Quantification of diel vertical migration bymicronektonic taxa in the northeast Atlantic. Hydrobiologia 440, 161–179.
Becker, E.L., Cordes, E.E., Macko, S.A., Fisher, C.R., 2009. Importance of seep primaryproduction to Lophelia pertusa and associated fauna in the Gulf of Mexico.Deep-Sea Research Part I: Oceanographic Research Papers 56, 786–800.
Bonnin, J., van Raaphorst, W., Brummer, G.-J., van Haren, H., Malschaert, H., 2002.Intense mid-slope resuspension of particulate matter in the Faeroe-ShetlandChannel: short-term deployment of near-bottom sediment traps. Deep-SeaResearch Part I: Oceanographic Research Papers 49, 1485–1505.
Brooke, S., Schroeder, W.W., 2007. State of deep coral ecosystems in the Gulf ofMexico Region: Texas to the Florida Straits. In: Lumsden, S.E., Hourigan, T.F.,Bruckner, A.W., Dorr, G. (Eds.), The State of Deep Coral Ecosystems of theUnited States NOAA Technical Memorandum CRCP-3 Silver Spring, MD,pp. 271–306.
Brooke, S., Young, C.M.. Direct measurements of in situ survival and growth ofLophelia pertusa in the Northern Gulf of Mexico. Marine Ecology ProgressSeries, doi:10.3354/meps08344.
Bryant, W.R., Bryant, J.R., Feeley, M.H., Simmons, G.R., 1990. Physiographic andbathymetric characteristics of the continental slope, northwest Gulf of Mexico.Geo-Marine Letters 10, 182–199.
Carnes, M.R., Teague, W.J., Jarosz, E., 2008. Low-frequency current variabilityobserved at the shelfbreak in the northeastern Gulf of Mexico: November2004–May 2005. Continental Shelf Research 28, 399–423.
Continental Shelf Associates, 2007. Characterization of Northern Gulf of Mexicodeepwater hard bottom communities with emphasis on Lophelia coral. USDept Interior, Minerals Management Service, Gulf of Mexico OCS Region, NewOrleans, LA OCS Study MMS 2007–044, pp. 169 and appendices.
Cordes, E.E., Bergquist, D.C., Predmore, B.L., Jones, C., Deines, P., Telesnicki, G.,Fisher, C.R., 2006. Alternate unstable states: convergent paths of succession inhydrocarbon-seep tubeworm-associated communities. Journal of Experimen-tal Marine Biology and Ecology 339, 159–176.
Cordes, E.E., McGinley, M.P., Podowski, E.L., Becker, E.L., Lessard-Pilon, S., Viada,S.T., Fisher, C.R., 2008. Coral communities of the deep Gulf of Mexico. Deep-SeaResearch Part I: Oceanographic Research Papers 55, 777–787.
Cordes, E.E., Bergquist, D.C., Fisher, C.R., 2009. Macro-ecology of Gulf of Mexicocold seeps. Annual Review of Marine Science 1, 143–168.
Davies, A.J., Roberts, J.M., Hall-Spencer, J., 2007. Preserving deep-sea naturalheritage: emerging issues in offshore conservation and management. Biolo-gical Conservation 138, 299–312.
Davies, A.J., Wisshak, M., Orr, J.C., Roberts, J.M., 2008. Predicting suitable habitat forthe cold-water reef framework-forming coral Lophelia pertusa (Scleractinia).Deep-Sea Research Part I: Oceanographic Research Papers 55, 1048–1062.
Davies, A.J., Duineveld, G.C.A., Lavaleye, M.S.S., Bergman, M.J.N., Van Haren, H.,Roberts, J.M., 2009. Downwelling and deep-water bottom currents as foodsupply mechanisms to the cold-water coral Lophelia pertusa (Scleractinia) atthe Mingulay Reef Complex. Limnology and Oceanography 54, 620–629.
De Mol, B., Van Rensbergen, P., Pillen, S., Van Herreweghe, K., Van Rooij, D.,McDonnell, A., Huvenne, V., Ivanov, M., Swennen, R., Henriet, J.P., 2002. Largedeep-water coral banks in the Porcupine Basin, southwest of Ireland. MarineGeology 188, 193–231.
Dodds, L.A., Roberts, J.M., Taylor, A.C., Marubini, F., 2007. The cold-water coralLophelia pertusa (Scleractinia) reveals metabolic tolerance to temperature anddissolved oxygen change. Journal of Experimental Marine Biology and Ecology349, 205–214.
Duineveld, G.C.A., Lavaleye, M.S.S., Berghuis, E.M., 2004. Particle flux and foodsupply to a seamount cold-water coral community (Galicia Bank, NW Spain).Marine Ecology-Progress Series 277, 13–23.
Duineveld, G.C.A., Lavaleye, M.S.S., Bergman, M.I.N., de Stigter, H., Mienis, F., 2007.Trophic structure of a cold-water coral mound community (Rockall Bank, NEAtlantic) in relation to the near-bottom particle supply and current regime.Bulletin of Marine Science 81, 449–467.
Dullo, W.C., Flogel, S., Ruggeberg, A., 2008. Cold-water coral growth in relation tothe hydrography of the Celtic and Nordic European continental margin. MarineEcology Progress Series 371, 165–176.
Formolo, M.J., Lyons, T.W., Zhang, C., Kelley, C., Sassen, R., Horita, J., Cole, D.R.,2004. Quantifying carbon sources in the formation of authigenic carbonates atgas hydrate sites in the Gulf of Mexico. Chemical Geology 205, 253–264.
Frederiksen, R., Jensen, A., Westerberg, H., 1992. The distribution of thescleractinian coral Lophelia pertusa around the Faeroe Islands and the relationto internal tidal mixing. Sarsia 77, 157–171.
Freiwald, A., Henrich, R., Patzold, J., 1997. Anatomy of a deep-water coral reefmound from Stjernsund, west Finnmark, northern Norway. In: James, N.P.,Clarke, J.A.D. (Eds.), Cool-water Carbonates. Society of Sedimentary Geology(SEPM) Special Publication, pp. 140–161.
Freiwald, A., 2002. Reef-forming cold-water corals. In: Wefer, G., Billett, D.,Hebbeln, D., Jorgensen, B.B., Schluter, M., Van Weering, T. (Eds.), Ocean MarginSystems. Springer-Verlag Berlin Heidelberg, Berlin, pp. 365–385.
Freiwald, A., Fossa, J.H., Grehan, A., Koslow, T., Roberts, J.M., 2004. Cold-water CoralReefs. UNEP-WCMC, Cambridge, UK (pp. 84).
Freiwald, A., Roberts, J.M. (Eds.), 2005. Cold-water Corals and Ecosystems.Springer-Verlag, Berlin Heidelberg, pp. 1243.
Genin, A., Dayton, P.K., Lonsdale, P.F., Speiss, F.N., 1986. Corals on seamount peaksprovide evidence of current acceleration over deep-sea topography. Nature322, 59–61.
Guinotte, J.M., Orr, J.C., Cairns, S.D., Freiwald, A., Morgan, L., George, R., 2006. Willhuman-induced changes in seawater chemistry alter the distribution ofdeep-sea scleractinian corals? Frontiers in Ecology and Environment 4,141–146.
Hall-Spencer, J., Allain, V., Fossa, J.H., 2002. Trawling damage to Northeast Atlanticancient coral reefs. Proceedings of the Royal Society of London SeriesB—Biological Sciences 269, 507–511.
Hallock, Z.R., Teague, W.J., Jarosz, E., 2009. Subinertial slope-trapped waves inthe northeastern Gulf of Mexico. Journal of Physical Oceanography 39,1475–1485.
Hamilton, P., Fargion, G., Biggs, D., 1999. Loop Current eddy paths in the westernGulf of Mexico. Journal of Physical Oceanography 29, 1180–1207.
Henry, L.-A., Roberts, J.M., 2007. Biodiversity and ecological composition ofmacrobenthos on cold-water coral mounds and adjacent off-mound habitat inthe bathyal Porcupine Seabight, NE Atlantic. Deep-Sea Research Part I:Oceanographic Research Papers 54, 654–672.
Hosegood, P., Bonnin, J., van Haren, H., 2004. Solibore-induced sedimentresuspension in the Faeroe-Shetland Channel. Geophysical Research Letters31, L09301.
Hovland, M., Mortensen, P.B., Brattegard, T., Strass, P., Rokoengen, K., 1998.Ahermatypic coral banks off Mid-Norway: evidence for a link with seepage oflight hydrocarbons. Palaios 13, 189–200.

ARTICLE IN PRESS
A.J. Davies et al. / Deep-Sea Research I 57 (2010) 199–212212
Jensen, A., Frederiksen, R., 1992. The fauna associated with the bank-formingdeep-water coral Lophelia pertusa (Scleractinaria) on the Faroe Shelf. Sarsia 77,53–69.
Jochens, A.E., DiMarco, S.F., 2008. Physical oceanographic conditions in thedeepwater Gulf of Mexico in Summer 2000–2002. Deep-Sea Research Part II:Topical Studies in Oceanography 55, 2541–2554.
Kennicutt, M.C., Brooks, J.M., Bidigare, R.R., Denoux, G.J., 1988. Gulf of Mexicohydrocarbon seep communities 1: regional distribution of hydrocarbonseepage and associated fauna. Deep-Sea Research 35, 639–651.
Leichter, J.J., Deane, G.B., Stokes, M.D., 2005. Spatial and temporal variability ofinternal wave forcing on a coral reef. Journal of Physical Oceanography 35,1945–1962.
Leichter, J.J., Helmuth, B., Fischer, A.M., 2006. Variation beneath thesurface: quantifying complex thermal environments on coral reefs inthe Caribbean, Bahamas and Florida. Journal of Marine Research 64,563–588.
Mienis, F., de Stigter, H.C., White, M., Duineveld, G., de Haas, H., van Weering,T.C.E., 2007. Hydrodynamic controls on cold-water coral growth andcarbonate-mound development at the SW and SE Rockall Trough margin, NEAtlantic ocean. Deep-Sea Research Part I: Oceanographic Research Papers 54,1655–1674.
Mienis, F., de Stigter, H.C., de Haas, H., van Weering, T.C.E., 2009. Near-bed particledeposition and resuspension in a cold-water coral mound area at theSouthwest Rockall Trough margin, NE Atlantic. Deep-Sea Research Part I:Oceanographic Research Papers 56, 1026–1038.
Piniak, G.A., Storlazzi, C.D., 2008. Diurnal variability in turbidity and coralfluorescence on a fringing reef flat: Southern Molokai, Hawaii. Estuarine,Coastal and Shelf Science 77, 56–64.
Reed, J.K., 2002. Deep-water Oculina coral reefs of Florida: biology, impacts, andmanagement. Hydrobiologia 471, 43–55.
Rogers, A.D., 1999. The biology of Lophelia pertusa (Linnaeus, 1758) and otherdeep-water reef-forming corals and impacts from human activities. Interna-tional Review of Hydrobiology 84, 315–406.
Rosato, E., Kyriacou, C.P., 2006. Analysis of locomotor activity rhythms inDrosophila. Nature Protocols 1, 559–568.
Ross, S.W., Nizinski, M.S., (Eds.), 2007. State of Deep Coral Ecosystems in the USSoutheast Region: Cape Hatteras to southeastern Florida. In: Lumsden, S.E.,Hourigan, T.F., Bruckner, A.W., Dorr, G., (Eds.), The State of Deep CoralEcosystems of the United States NOAA Technical Memorandum CRCP-3 SilverSpring MD, pp. 233–270.
Schroeder, W.W., 2002. Observations of Lophelia pertusa and the surficial geologyat a deep-water site in the northeastern Gulf of Mexico. Hydrobiologia 471,29–33.
Schroeder, W., Brooke, S., Olson, J., Phaneuf, B., McDonough, J., Etnoyer, P., 2005.Occurrence of deep-water Lophelia pertusa and Madrepora oculata in the Gulf of
Mexico. In: Freiwald, A., Roberts, J.M. (Eds.), Cold-water Corals and Ecosys-tems. Springer, pp. 297–307.
Smith, D.C., 1986. A numerical study of Loop Current Eddy interaction withtopography in the western Gulf of Mexico. Journal of Physical Oceanography16, 1260–1272.
Sturges, W., Lugo-Fernandez, A., (Eds.), 2005. Circulation in the Gulf of Mexico:Observations and Models, vol 161. American Geophysical Union, pp. 360 andCD appendices.
Sulak, K.J., Brooks, R.A., Luke, K.E., Norem, A.D., Randall, M.T., Quaid, A.J., Yeargin,G.E., Miller, J.M., Harden, W.M., Caruso, J.H., Ross, S.W., 2007. Demersal fishesassociated with Lophelia pertusa coral and hard-substrate biotopes on thecontinental slope, northern Gulf of Mexico. In: George, R.Y., Cairns, S.D. (Eds.),Conservation and Adaptive Management of Seamount and Deep-sea CoralEcosystems. University of Miami, pp. 65–92.
Sulak, K.J., Randall, M.T., Luke, K.E., Norem, A.D., Miller, J.M., (Eds.), 2008Characterization of northern Gulf of Mexico deepwater hard bottom commu-nities with emphasis on Lophelia coral: Lophelia reef megafaunal communitystructure, biotopes, genetics, microbial ecology, and geology (2004–2006),USGS Open-File Report 2008-1148, OCS Study MMS 2008-015.
Teague, W.J., Jarosz, E., Carnes, M.R., Mitchell, D.A., Hogan, P.J., 2006.Low-frequency current variability observed at the shelfbreak in the north-eastern Gulf of Mexico: May–October, 2004. Continental Shelf Research 26,2559–2582.
van Weering, T.C.E., Koster, B., Heerwaarden, J., Thomsen, L., Viergutz, T., 2000.New technique for long term deep seabed studies. Sea Technology 41, 17–26.
Verardo, D., Froelich, P., McIntyre, A., 1990. Determination of organic carbon andnitrogen in marine sediments using the Carlo Erba NA-1500 Analyzer. Deep-Sea Research Part I: Oceanographic Research Papers 37, 157–165.
Vidal, V.M.V., Vidal, F.V., Hernandez, A.F., Meza, E., Zambrano, L., 1994. Winterwater mass distributions in the Western Gulf of Mexico affected by a collidinganticyclonic ring. Journal of Oceanography 50, 559–588.
Wang, Y.H., Dai, C.F., Chen, Y.Y., 2007. Physical and ecological processes of internalwaves on an isolated reef ecosystem in the South China Sea. GeophysicalResearch Letters 34, 7.
White, M., Mohn, C., de Stigter, H., Mottram, G., 2005. Deep-water coraldevelopment as a function of hydrodynamics and surface productivity aroundthe submarine banks of the Rockall Trough, NE Atlantic. In: Freiwald, A.,Roberts, J.M. (Eds.), Cold-water Corals and Ecosystems. Springer-Verlag, BerlinHeidelberg, pp. 503–514.
White,, M., 2007. Benthic dynamics at the carbonate mound regions of thePorcupine Seabight continental margin. International Journal of Earth Sciences96, 1–9.
Wilson, J.B., 1979. ‘Patch’ development of the deep-water coral Lophelia pertusa (L.)on Rockall Bank. Journal of the Marine Biological Association of the UnitedKingdom 59, 165–177.




















