
Shear stress induces changes in the morphology and cytoskeleton organisation of arterial endothelial...
7
Eur J Vasc Endovasc Surg 9, 86-92 (1995) Shear Stress Induces Changes in the Morphology and Cytoskeleton Organisation of Arterial Endothelial Cells Alessandra Cucina, Antonio V. Sterpetti, Guntars Pupelis, Alessandra Fragale, Sandro Lepidi, Antonino Cavallaro, Quinto Giustiniani, and Luciana Santoro D'Angelo First Department of Surgery and Institute of Histology and Embryology, School of Medicine, La Sapienza University, Rome, Italy Objectives: The aim of this study was to determine the changes in the morphology and cytoskeleton organisation of endothelial cells (EC) determined by exposure to a laminarflow. Cultured EC were exposed to a wall shear stress of 6 dyne/ cm z for 24 hours. Chief outcome measures: The morphology of EC was analysed by light and scanning electron microscopy. The organisation of the cytoskeleton was determined by doublefluorescence labelingwith antibody anti-vimentin, anti-vinculin, anti-tubulin, and with rhodamine-labeled phalloidin. Results: EC exposed to laminar flow become round-shaped with decreased area of adhesion to the substrate. There was a clear reorganisation of the cytoskeleton after exposure to shear stress; the distribution of actin changedfrom a stress fibre pattern to a more diffuse membrane-associated distribution. These changes in shape and cytoskeleton organisation were reversible after a 48-hour resting period. Conclusions: EC respond to laminarflow in a predictable manner and thesefindings may be correlated to the functional changes of EC observed after exposure to shear stress. Key Words: Endothelial cells; Cytoskeleton; Shear stress. Introduction Endothelial cells (EC) form a continuous layer that lines the entire vascular system, providing a non- thrombogenic surface and performing a variety of specialised metabolic and transport functions. EC function can be influenced by chemical and physical stimuli. EC are continuously subjected to a variety of haemodynamic forces. There is evidence that fluid dynamics play a major role in the development of atherosclerosis. Regions of arterial branching and sharp curvature have the greatest predilection to develop atherosclerosis. 1~ Haemodynamic injury to the endothelium may Pslay a contributory role in the atherosclerotic process. Much of our knowledge of EC biology has come from studies of EC cultured in static conditions. However, EC in vivo are continuously subjected to the blood flowing under pressure. Mechanical stress applied to cells in culture alters the functional charac- teristics of the cells. 64 Cytoskeleton elements have Please address all correspondence to: Alessandra Cucina, Via A. Scarpa, 14 00161Rome, Italy. been related to several cell functions. Quantitative changes of cytoskeletal components have been shown to correlate with adaption during pathological phe- nomena. Theoretically, shear stress might influence EC function through changes in cytoskeleton organisation. The purpose of this study was to determine the changes in the morphology and cytoskeleton organi- sation of EC determined by laminar flow. Material and Methods Cell isolation and culture Thoracic aortas of calves were removed aseptically at the abattoir and stored on ice in Phosphate Buffered Saline (PBS) and penicillin (100UI/ml) /streptomycin (100~g/ml) /fungizone (2.5~g/ml). The aortas were washed with Hanks' balanced salt solution and opened longitudinally. They were placed with the luminal surface downward in a 0.03% solution of collagenase type II (Sigma Chemical Co, St Louis, Mo, 1078-5884/95/010086 + 07 $08"00/0© 1995W. B. Saunders Company Ltd.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Shear stress induces changes in the morphology and cytoskeleton organisation of arterial endothelial...

Eur J Vasc Endovasc Surg 9, 86-92 (1995)
Shear Stress Induces Changes in the Morphology and Cytoskeleton Organisation of Arterial Endothelial Cells
Alessandra Cucina, Antonio V. Sterpetti, Guntars Pupelis, Alessandra Fragale, Sandro Lepidi, Antonino Cavallaro, Quinto Giustiniani, and Luciana Santoro D'Angelo
First Department of Surgery and Institute of Histology and Embryology, School of Medicine, La Sapienza University, Rome, Italy
Objectives: The aim of this study was to determine the changes in the morphology and cytoskeleton organisation of endothelial cells (EC) determined by exposure to a laminar flow. Cultured EC were exposed to a wall shear stress of 6 dyne/ cm z for 24 hours. Chief outcome measures: The morphology of EC was analysed by light and scanning electron microscopy. The organisation of the cytoskeleton was determined by double fluorescence labeling with antibody anti-vimentin, anti-vinculin, anti-tubulin, and with rhodamine-labeled phalloidin. Results: EC exposed to laminar flow become round-shaped with decreased area of adhesion to the substrate. There was a clear reorganisation of the cytoskeleton after exposure to shear stress; the distribution of actin changed from a stress fibre pattern to a more diffuse membrane-associated distribution. These changes in shape and cytoskeleton organisation were reversible after a 48-hour resting period. Conclusions: EC respond to laminar flow in a predictable manner and these findings may be correlated to the functional changes of EC observed after exposure to shear stress.
Key Words: Endothelial cells; Cytoskeleton; Shear stress.
Introduction
Endothelial cells (EC) form a continuous layer that lines the entire vascular system, providing a non- thrombogenic surface and performing a variety of specialised metabolic and transport functions. EC function can be influenced by chemical and physical stimuli. EC are continuously subjected to a variety of haemodynamic forces. There is evidence that fluid dynamics play a major role in the development of atherosclerosis. Regions of arterial branching and sharp curvature have the greatest predilection to develop atherosclerosis. 1~ Haemodynamic injury to the endothelium may Pslay a contributory role in the atherosclerotic process.
Much of our knowledge of EC biology has come from studies of EC cultured in static conditions. However, EC in vivo are continuously subjected to the blood flowing under pressure. Mechanical stress applied to cells in culture alters the functional charac- teristics of the cells. 64 Cytoskeleton elements have
Please address all correspondence to: Alessandra Cucina, Via A. Scarpa, 14 00161 Rome, Italy.
been related to several cell functions. Quantitative changes of cytoskeletal components have been shown to correlate with adaption during pathological phe- nomena. Theoretically, shear stress might influence EC function through changes in cytoskeleton organisation.
The purpose of this study was to determine the changes in the morphology and cytoskeleton organi- sation of EC determined by laminar flow.
Material and Methods
Cell isolation and culture
Thoracic aortas of calves were removed aseptically at the abattoir and stored on ice in Phosphate Buffered Saline (PBS) and penicillin (100UI/ml) /streptomycin (100~g/ml) /fungizone (2.5~g/ml). The aortas were washed with Hanks' balanced salt solution and opened longitudinally. They were placed with the luminal surface downward in a 0.03% solution of collagenase type II (Sigma Chemical Co, St Louis, Mo,
1078-5884/95/010086 + 07 $08"00/0 © 1995 W. B. Saunders Company Ltd.

Cytoskeleton Organisation and Shear Stress 87
U.S.A.). After incubation at 37°C for 30 min, the EC were removed by gentle scraping with a rubber policeman. The cells were cultured on polystyrene culture dishes in M199 supplemented with 10% fetal calf serum (FCS) and antibiotics. They were grown in an environment of 5% CO2 at 37°C. Culture medium was changed every other day. The cells were sub- cultured after removal with 0.01% trypsin-0.01% EDTA.
For shear stress tests, EC were grown in cov- erslips in complete culture medium, in a modified Leighton tube (Costar, Cambridge, Ma). Coverslips were used in shear stress tests after two days of culture; at this time, cultures were at a subconfluent density. 50.000 EC were cultured on the plastic coverslips of a Leighton tube. The coverslips were interposed in a continuous flow system to expose cultured EC to a predetermined level of shear stress under conditions of steady laminar flow. 9 The circuit was composed of a roller-type perfusion pump and a closed circulatory loop. The flow medium consisted of DMEM with 10% FCS and penicillin (100UI/ml) /streptomycin (100 btg/ml) /fungizone (2.5~g/ml). The entire apparatus was kept at 37°C in a 5% CO2 incubator.
A wall shear stress of 6 dyne /cm 2 was used for these experiments. The control EC were subjected to similar incubation conditions, without being subjected to a passive flow. All experiments were performed at least in triplicate. EC were exposed to laminar flow for 24 hours, In order to determine the changes after shear stress, the medium was changed and the EC were left undisturbed for 48 hours.
Immunofluorescence
The organisation of the cytoskeleton was analysed after 24 h of laminar flow and after a 48 h resting period. Coverslips were rinsed in PBS and fixed in cold methanol for 10 minutes and stored dry at -20°C. Before labeling, specimens were washed three times in PBS and then overlaid with mouse monoclonal anti- bodies anti-vimentin, anti-vinculin and anti-beta tubu- lin (SIGMA). Each antibody was used in a single experiment. These antibodies crossreact with bovine cells (unpublished data).
After rinsing in PBS, coverslips were overlaid with fluorescein-conjugated antibody to immunoglo- bulin G for 30 minutes at room temperature. The coverslips were then washed three times in PBS, mounted in PBS/glycerol (50 v/v) . For rhodamine- labeled phalloidin stain coverslips were rinsed in PBS and fixed in 3% formaldehyde in PBS (pH 7.6) for 5 rain at room temperature. After rinsing in PBS, coverslips were soaked in HEPES-Triton X-100 buffer for 5 min at 0°C. 1° Fixed and permeabilised cells were stained with 2 btg/ml rhodamine-labeled phalloidin for 30 min at 37°C and mounted in PBS/glycerol (50 v/v).
Negative control staining was performed by replacing the primary antiserum with non-reacting mouse IgG. Coverslips were examined with a Zeiss photomicroscope. For fluorescein fluorescence a selec- tive interference excitation filter (410 to 485 nm) was used. For rhodamine fluorescence a selective inter- ference excitation filter (535 t ° 550 nm) was used.
Results
Microscopy
The EC monolayer grown on coverslips were exam- ined at 24 hours after laminar flow by a Zeiss photo- microscope for phase-contrast microscopy.
For electron microscopy EC were fixed in glutar- aldehyde (2.5% in 0.1 M cacodylate buffer) for 3 h at 4°C, washed 3 times in 0.1 M cacodylate buffer, postfixed in OsO4 (2% in 0.1 M cacodylate buffer) for 1 h and then washed three times in 0.1 M cacodylate buffer. EC were then dehydrated in a graded ethanol series and dried by the critical,point method (Emscope CPD 750). The cells were mounted on stubs, coated with gold plate by a sputterer (Emscope SC 500; 15 mA, 44 s) and examined with a scanning electron microscope (Hitachi S-570) operating at 15 Kv.
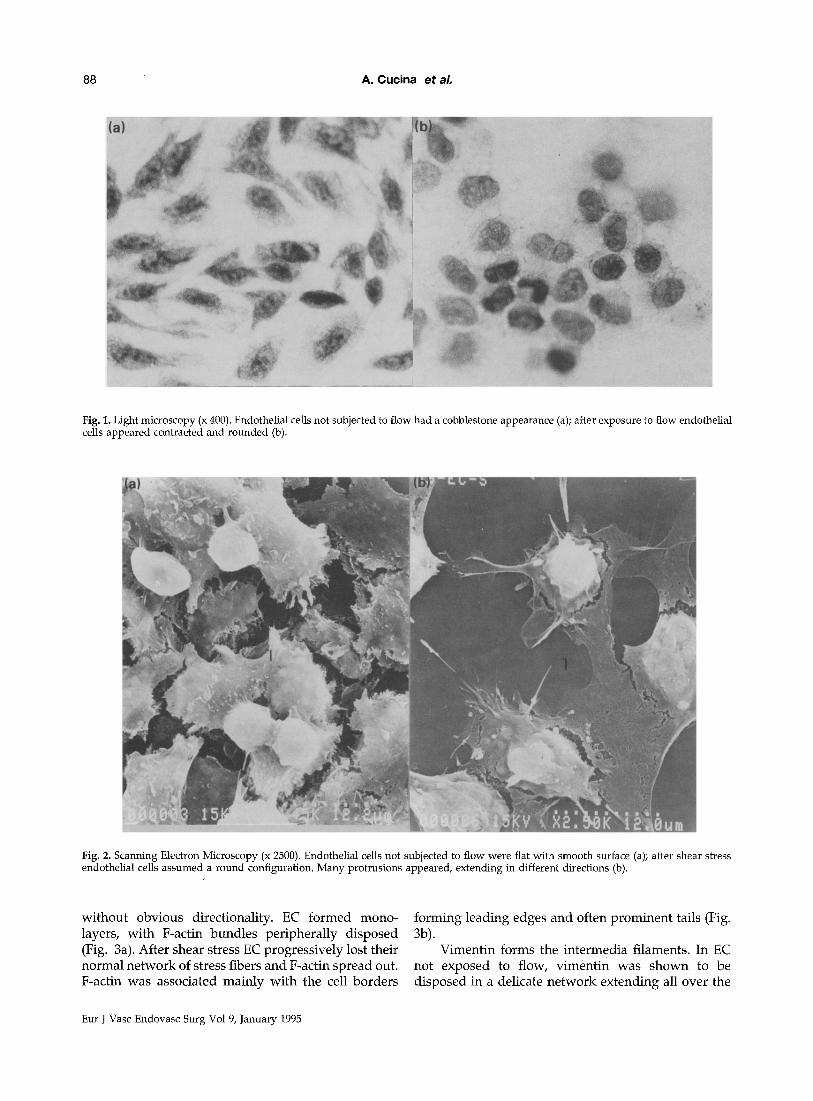
EC not subjected to flow had a cobblestone appear- ance on light microscopy, whereas EC subjected to shear stress appeared contracted and lost contact with neighbour cells (Fig. la, lb). The changes observed by light microscopy were confirmed by scanning electron microscopy. EC not subjected to flow were flat with a rather smooth surface and in contact by means of rounded shaped protrusions. After 24 h of laminar flow, EC assumed a round configuration with nucle- ous protruding from the even surface. Many protru- sions appeared, extending in different directions (Fig. 2a, 2b).
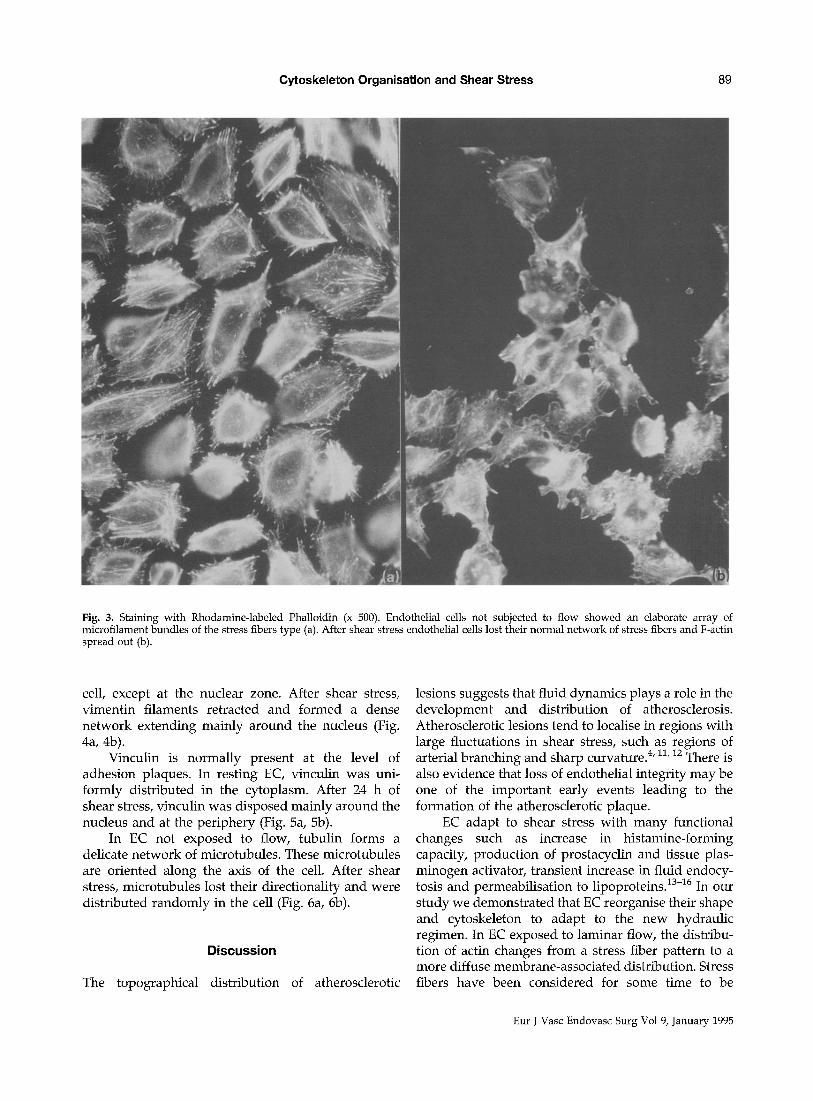
After staining with R-PHD (Rhodanine-labeled Phalloidin) that specifically binds to F-actin, resting EC showed an elaborate array of microfilament bundles of the stress fibers type. Within the central cellular regions, filaments were randomly oriented
Eur J Vasc Endovasc Surg Vol 9, January 1995
88 A. Cucina et aL
Fig. 1. Light microscopy (x 400). Endothelial cells not subjected to flow had a cobblestone appearance (a); after exposure to flow endothelial cells appeared contracted and rounded (b).
Fig. 2. Scanning Electron Microscopy (x 2500). Endothelial cells not subjected to flow were flat with smooth surface (a); after shear stress endothelial cells assumed a round configuration. Many protrusions appeared, extending in different directions (b).
wi thout obvious directionality. EC formed mono- layers, wi th F-actin bundles per iphera l ly d isposed (Fig. 3a). After shear stress EC progress ively lost their normal ne twork of stress fibers and F-actin spread out. F-actin was associated main ly wi th the cell borders
fo rming leading edges and often p rominen t tails (Fig. 3b).
Vimentin forms the in termedia filaments. In EC not exposed to flow, v iment in was shown to be d isposed in a delicate ne twork extending all over the
Eur J Vasc Endovasc Surg Vol 9, January 1995
Cytoskeleton Organisation and Shear Stress 89
Fig. 3. Staining with Rhodamine-labeled Phalloidin (x 500). Endothelial cells not subjected to flow showed an elaborate array of microfilament bundles of the stress fibers type (a). After shear stress endothelial cells lost their normal network of stress fibers and F-actin spread out (b).
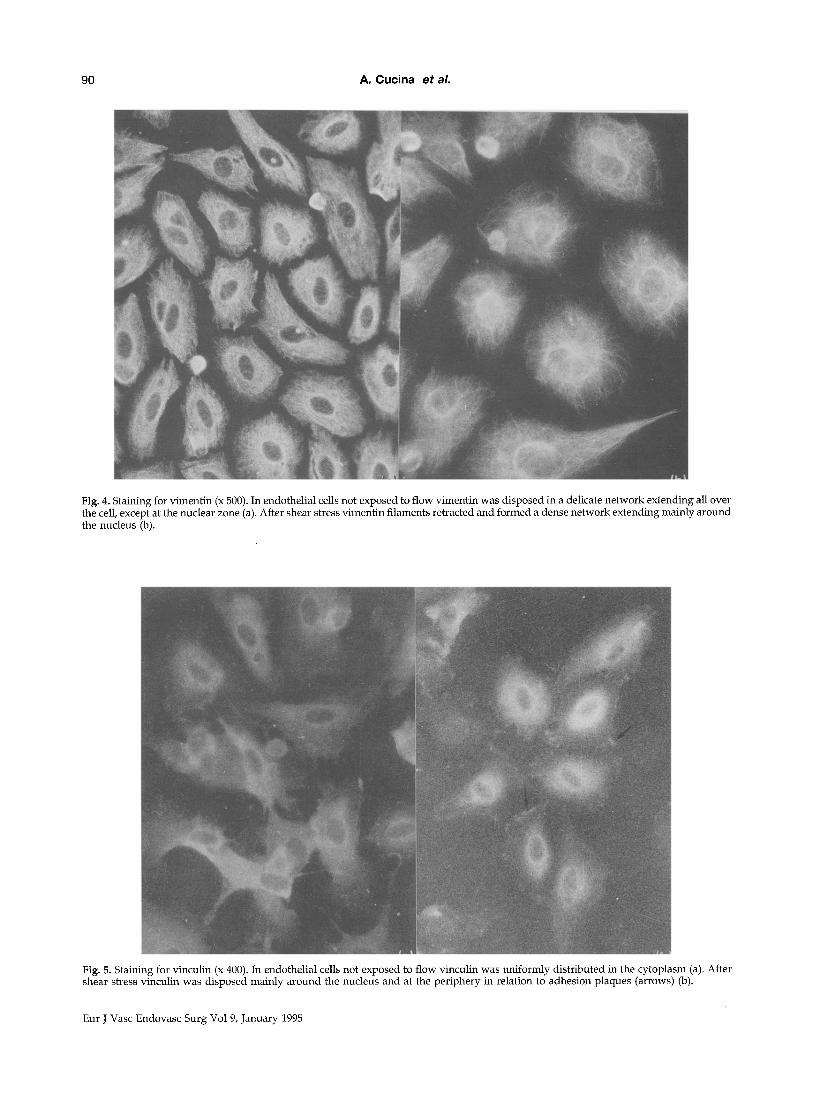
cell, except at the nuclear zone. After shear stress, vimentin filaments retracted and formed a dense network extending mainly around the nucleus (Fig. 4a, 4b).
Vinculin is normally present at the level of adhesion plaques. In resting EC, vinculin was uni- formly distributed in the cytoplasm. After 24 h of shear stress, vinculin was disposed mainly around the nucleus and at the periphery (Fig. 5a, 5b).
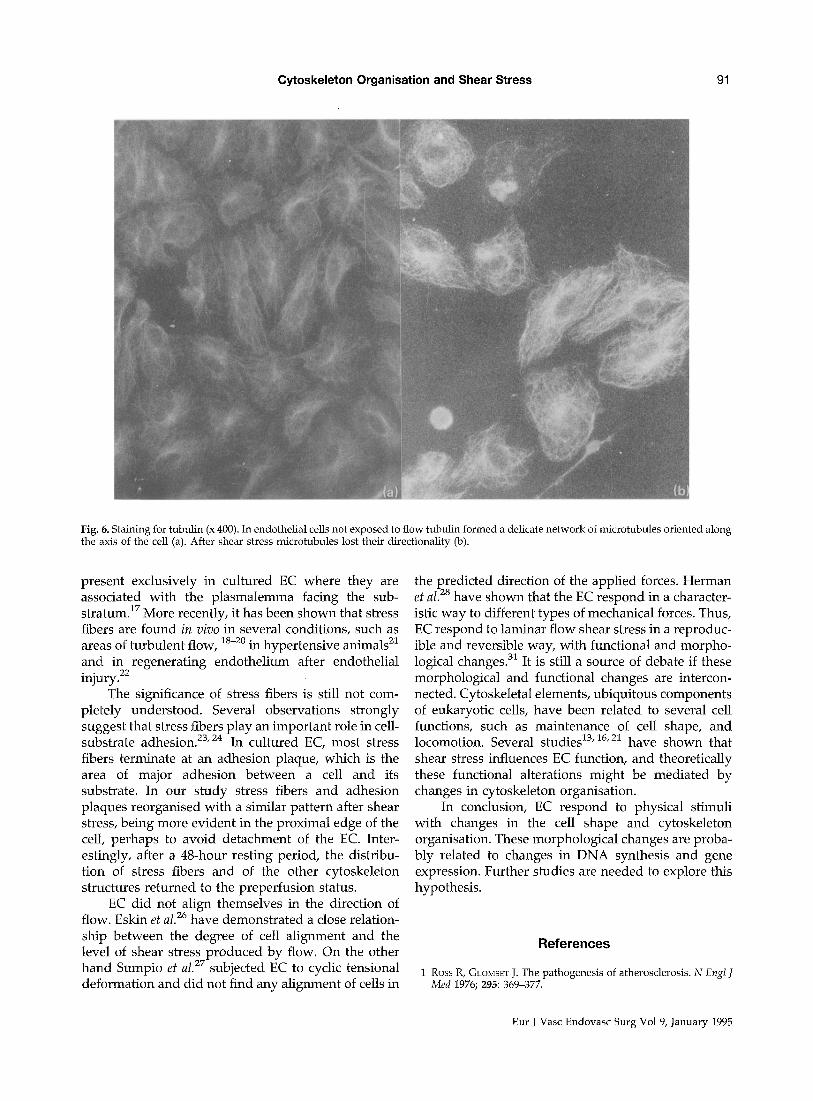
In EC not exposed to flow, tubulin forms a delicate network of microtubules. These microtubules are oriented along the axis of the cell. After shear stress, microtubules lost their directionality and were distributed randomly in the cell (Fig. 6a, 6b).
Discussion
The topographical distribution of atherosderotic
lesions suggests that fluid dynamics plays a role in the development and distribution of atherosclerosis. Atherosclerotic lesions tend to localise in regions with large fluctuations in shear stress, such as regions of arterial branching and sharp curvature. 4' 11, 12 There is also evidence that loss of endothelial integrity may be one of the important early events leading to the formation of the atherosclerotic plaque.
EC adapt to shear stress with many functional changes such as increase in histamine-forming capacity, production of prostacyclin and tissue plas- minogen activator, transient increase in fluid endocy- tosis and permeabilisation to lipoproteins. ~3-~6 In our study we demonstrated that EC reorganise their shape and cytoskeleton to adapt to the new hydraulic regimen. In EC exposed to laminar flow, the distribu- tion of actin changes from a stress fiber pattern to a more diffuse membrane-associated distribution. Stress fibers have been considered for some time to be
Eur J Vasc Endovasc Surg Vol 9, January 1995
90 A. Cucina et aL
Fig. 4. Staining for vimentin (x 500). In endothelial cells not exposed to flow vimentin was disposed in a delicate network extending all over the cell, except at the nuclear zone (a). After shear stress vimentin filaments retracted and formed a dense network extending mainly around the nucleus (b).
Fig. 5. Staining for vinculin (x 400). In endothelial cells not exposed to flow vinculin was Uniformly distributed in the cytoplasm (a). After shear stress vinculin was disposed mainly around the nucleus and at the periphery in relation to adhesion plaques (arrows) (b).
Eur J Vasc Endovasc Surg Vol 9, January 1995
Cytoskeleton Organisation and Shear Stress 91
Fig. 6. Staining for tubulin (x 400). In endothelial cells not exposed to flow tubulin formed a delicate network of microtubules oriented along the axis of the cell (a). After shear stress microtubules lost their directionality (b).
present exclusively in cultured EC where they are associated with the plasmalemma facing the sub- stratumJ 7 More recently, it has been shown that stress fibers are found in vivo in several conditions, such as areas of turbulent flow, 18-20 in hypertensive animals 21 and in regenerating endothelium after endothelial injury. 22
The significance of stress fibers is still not com- pletely understood. Several observations strongly suggest that stress fibers play an important role in cell- substrate adhesion. 23' 24 In cultured EC, most stress fibers terminate at an adhesion plaque, which is the area of major adhesion between a cell and its substrate. In our study stress fibers and adhesion plaques reorganised with a similar pattern after shear stress, being more evident in the proximal edge of the cell perhaps to avoid detachment of the EC. Inter- estingly, after a 48-hour resting period, the distribu- tion of stress fibers and of the other cytoskeleton structures returned to the preperfusion status.
EC did not align themselves in the direction of flow. Eskin et al. 26 have demonstrated a close relation- ship between the degree of cell alignment and the level of shear stress produced by flow. On the other hand Sumpio et al. 27 subjected EC to cyclic tensional deformation and did not find any alignment of cells in
the predicted direction of the applied forces. Herman et a l J s have shown that the EC respond in a character- istic way to different types of mechanical forces. Thus, EC respond to laminar flow shear stress in a reproduc- ible and reversible way, with functional and morpho- logical changes. 31 It is still a source of debate if these morphological and functional changes are intercon- nected. Cytoskeletal elements, ubiquitous components of eukaryotic cells, have been related to several cell functions, such as maintenance of cell shape, and locomotion. Several studies 13' 16,22 have shown that shear stress influences EC function, and theoretically these functional alterations might be mediated by changes in cytoskeleton organisation.
In conclusion, EC respond to physical stimuli with changes in the cell shape and cytoskeleton organisation. These morphological changes are proba- bly related to changes in DNA synthesis and gene expression. Further studies are needed to explore this hypothesis.
References
1 Ross R, GLOMSET J. The pathogenesis of atherosclerosis. N Engl J Med 1976; 295: 369-377.
Eur J Vasc Endovasc Surg Vol 9, January 1995

92 A. Cucina et aL
2 SCHWARTZ SM. Role of endothelial integrity in atherosclerosis. Artery 1980; 8: 305-314.
3 REIDY MA, SCHWARTZ SM. Recent advances in molecular pathol- ogy. Arterial endothelium-assessment of in vivo injury. Exp Mol Pathol 1984; 41: 419-434.
4 SABBAH HN, KHRAJA F, BRYIVIER JF, HAWKINS ET, STEIN PD. Blood flow velocity in the right coronary artery: Relation to the distribution of atherosclerotic lesions. Am J CardioI 1984; 53: 1008-1012.
5 REIDY MA, SCHWARTZ SM. Endothelial regeneration III Time course of intimal changes after small defined injury to rat aortic endothelium. Lab Invest 1981; 44: 301-306.
6 LEUNG DY, GLAGOV S, MATHEWS MB. Cyclic stretching stimulates synthesis of matrix components by arterial smooth muscle cells in vitro. Science 1976; 191: 475-477.
7 SuMPIO BE, BANES AJ, LEVIN LG, JOHNSON G. Mechanical stress stimulates aortic endothelial cells to proliferate. ]Vasc Surg 1987; 6: 252-256.
8 SUMPIO BE, BANES AJ. Response of porcine aortic smooth muscle cells to cydic tensional deformation in culture. J Surg Res 1988; 44: 696-701.
9 STERPETTI AV, CI:CINA A, SANTORO L, CARDILLO B, CAVALLARO A. Modulation of arterial smooth muscle cell growth by haemody- namic forces. Eur J Vasc Surg 1992; 6: 16-20.
10 BUSSOLINO F, CAMUSSI G, AGLIETTA Mr BRAQUET P, BosiA A, PESCARMONA G, SANAVIO F, D'URso N, MARCHISIO PC. Human endothelial cells are target for platelet-activating factor. I. Platelet-activating factor induces changes in cytoskeleton struc- tures. J ImmunoI 1987; 139: 2439-2446.
11 ZARINS CK, G1DDENS DP, BHARADVAY BK, SOTTIURAI MS, MABON RE, GLAGOV S. Carotid bifurcation atherosclerosis. Quantitative correlation of plaque localization with flow velocity profiles and wall shear stress. Circ Res 1983; 53: 502-514.
12 ZARINS CK, BOMBERGER RA, GLAGOV S. Local effects of stenoses. Increased flow velocity inhibits atherogenesis. Circulation 1981; 64(II): 221-227.
13 FRANGOS JA, ESKIN SG, MCINTIRE LV, IVES CL. Flow effects on prostacyclin production by cultured human endothelial cells. Science 1985; 227: 1477-1479.
14 DIAMOND SL, SHAREFKIN JB, DEFFENBACH C, FRASIER-SCOTT K r
MCINTIRE LV, ESKIN SG. Tissue plasminogen activator messenger RNA levels increase in cultured human endothelial cells exposed to laminar shear stress. J Cell PhysioI 1990; 143: 364-371.
15 DEWEY CF, BUSSOLAKI SRr GIMBRONE MA, DAVIES PF. The dynamic response of vascular endothelial cells to fluid shear stress. ] Biomech Eng 1981; 103: 177-185.
16 DAVIES PF, DEWEY CF, BUSSOLARI SR, GORDON EJ, GIMBRONE MA. Influence of hemodynamic forces on vascular endothelial func- tion. In vitro studies of shear stress and pinocytosis in bovine aortic cells. J Clin Invest 1984; 73: 1121-1129.
17 BYERS HR, FUJIWARA SR. Stress fibers in ceils in situ: immuno- fluorescent visualization with antiactin, antimyosin, and anti- alfa-actin. ] Cell BioI 1982; 93: 804-811.
18 WHITE GE, GIMBRONE MA, FUJIWARA K. Factors influencing the expression of stress fibers in vascular endothelial cells in situ. J Cell Biol 1983; 97: 416-424.
19 KIM DW, LANGILLE L, WONG MKK, GOTLIEB AI. Patterns of endothelial microfilament distribution in the rabbit aorta in situ. Circ Res 1989; 64: 21-31.
20 WECHEZAK AR, VIGGERS RE, SAUVAGE LR. Fibronectin and F-actin redistribution in cultured endothelial cells exposed to shear stress. Lab Invest 1985; 53: 639~47.
21 WHITE GE, FUJIWARA K. Expression and intracellular distribution of shear stress fibers in aortic endothelium. J Cell Biol 1986; 103: 63-70.
22 GABBIANI G, GABBIANI F, LOMBARDI D, SCHWARTZ SM. Organiza- tion of actin cytoskeleton in normal and regenerating arterial endothelial cells. Proc Natl Acad Sci USA 1983; 80: 2361-2364.
23 WONG AJ, POLLARD TD, HERMAN lIVE Actin filament stress fibers in vascular endothelial cells in vivo. Science 1983; 219: 867-869.
24 WECHEZAK AR, WIGHT TN, VIGGERS RE, SAUVAGE LR. Endothelial adherence under shear stress is dependent upon microfilament reorganization. J Ceil Physiol 1989; 139: 136-146.
25 ESKIN SG, IVES CL, MCINTIRE LV, NAVARRO LT. Response of cultured endothelial cells to steady flow. Microvasc Res 1984; 28: 87-94.
26 SUMPIO BE, BANES AJ, BUCKLEY M, JOHNSON G. Alterations in aortic endothelial cell morphology and cytoskeleton protein synthesis during cyclic tensional deformation, l Vasc Surg 1988; 7: 130-138.
27 HERMAN IM, BRANT AM, WARTY VS, BONACCORSO Jr KLEIN EC, KOP, MOS RL, BOROVETZ HS. Hemodynamics and vascular endo- thelial cytoskeleton. J Cell Biol 1987; 105: 291-302.
28 STERPETTI AV, CUCINA A, MORENA A, DI DONNA S, D'ANGELO SANTORO L, CAVALLARO A, STIPA S. Shear stress increases the release of Interleukin 1 and Interleukin 6 by aortic endothelial cells. Surgery 1993; 114: 911-914.
Accepted 26 May 1994
Eur J Vasc Endovasc Surg Vol 9, January 1995




















