
Selection and their antitumor activity of antisense oligonucleotides targeting messenger RNA of...
10
Chinese Journal of Cancer Research 17(3):161-170, 2005 161 SELECTION AND THEIR ANTITUMOR ACTIVITY OF ANTISENSE OLIGONUCLEOTIDES TARGETING MESSENGER RNA OF VASCULAR ENDOTHELIAL GROWTH FACTOR RECEPTOR 2 ZHENG Su-jun (~.~)1, LIN Ru-xian (~k~lh) 1, XIA Yun (~)1, Bo Xiao-chen (~t ~J~) 1, REN Hong (~)2, ZHONG Sen (~)2, WANG Sheng -qi (q:~-~)l* 1Beijing Institute of Radiation Medicine, Beijing 100850; 21nstitute of Viral Hepatitis, Chongqing University of Medical Sciences, Chongqing 400016 CLC number: R730.5 Document code: A Article ID: 1000-9604(2005)03-0161-10 ABSTRACT Objective: To select the antisense oligonucleotides (asONs) which hybridize with the mRNA of vascular endothelial growth factor receptor2 (VEGFR2, also named as kinase insert domain-containing receptor:KDR) in an effective and specific way, and to investigate their antitumor activity in MCF-7 cells. Methods: The effective antisense oligonucleotides were chosen by computer prediction combined with oligonucleotide microarrays. The inhibition effect on MCF-7 cells proliferation was measured by MTT; and VEGFR2 expression was surveyed by Western-blotting and RT- PCR. Results: Using predicting secondary structure of VEGFR2 mRNA with RNA folding program, computer prediction designed 30 antisense oligonucleotide probes that were directed to local loose regions of RNA structure. In 30 probes, 4(4•30, 13.33%) antisense oligonucleotides showed strong hybridization intensities in oligonucleotide microarrays test and were selected. All these antisense oligonucleotides targeting 4 different sites of VEGFR2 mRNA lowered the level of VEGFR2 mRNA and protein present in MCF-7 cells. Proliferation of MCF-7 eel ls was reduced by 4 antisense oligonucleotides, respectively, in which asON1 was the most effective, with the inhibitory rates being 53.06% at 0.8 ~tmol/L. Conclusion: Combination of computer prediction with oligonucleotide microarrays is an effective way in selecting optimal antisense oligonucleotides. The antisense oligonucleotides showed good correlation between their antitumor activity and the hybridization intensities. The antisense oligonucleotides targeting VEGFR2 mRNA demonstrated prominent antitumor role in vitro. Key words: Target selection; Computer prediction; Oligonucleotide microarrays; Antisense oligonucleotide; VEGFR2/KDR Angiogenesis, the process by which capillaries sprout from preexisting blood vessels, is tightly regulated by a large number of proangiogenic and antiangiogenic factors. Tumor cells have an absolute requirement for a persistent supply of new blood vessels to nourish their Received date: Mar. 28, 2005; Accepted date: May 16, 2005 Foundation item: This work was supported by the National Natural Sciences Foundation of China (No. 3017111) and National Project "863" (No. 2001AA234041). *Author to whom correspondence should be addressed. Phone: (0086-10)-66932211; E-mail: [email protected] Biography: ZHENG Su-jun (1973-), male, attending physician, now working at Beijing Youan Hospital, Capital University of Medical Sciences, majors in liver disease. E-mall: zheng003 @ sina.com growth and to facilitate metastasis. Thus, tumor vascularization is a vital process for the progression of a neoplasm from a small localized tumor to an enlarging tumor with the ability to metastasize. Vascular endothelial growth factor (VEGF), the most potent direct-acting angiogenic protein known tll, is a diffusible endothelial ceil (EC) specific mitogen and angiogenic factor that also increases vascular permeability. Endothelial cell survival in newly formed vessel is VEGF-dependentt21, VEGF overproduction has been identified as a major factor underlying pathological angiogenesis for tumor in vivo. VEGF plays its most biological function through binding to its receptor: Vascular endothelial growth factor receptor 2 t3' 41 (VEGFR2, also named as kinase insert domain- containing receptor:KDR). The VEGFR2/KDR gene has been mapped to human chromosome 4ql 1-q12. The full
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Selection and their antitumor activity of antisense oligonucleotides targeting messenger RNA of...

Chinese Journal of Cancer Research 17(3):161-170, 2005 161
SELECTION AND THEIR ANTITUMOR ACTIVITY OF ANTISENSE OLIGONUCLEOTIDES TARGETING MESSENGER RNA OF VASCULAR ENDOTHELIAL GROWTH FACTOR RECEPTOR 2
ZHENG Su-jun ( ~ . ~ ) 1 , LIN Ru-xian ( ~ k ~ l h ) 1, XIA Yun ( ~ ) 1 , Bo Xiao-chen (~t ~ J ~ ) 1, REN Hong ( ~ ) 2 , ZHONG Sen ( ~ ) 2 , WANG Sheng -qi ( q : ~ - ~ ) l *
1Beijing Institute of Radiation Medicine, Beijing 100850; 21nstitute of Viral Hepatitis, Chongqing University of Medical Sciences, Chongqing 400016
CLC number: R730.5 Document code: A Article ID: 1000-9604(2005)03-0161-10
ABSTRACT
Objective: To select the antisense oligonucleotides (asONs) which hybridize with the mRNA of vascular endothelial growth factor receptor2 (VEGFR2, also named as kinase insert domain-containing receptor:KDR) in an effective and specific way, and to investigate their antitumor activity in MCF-7 cells. Methods: The effective antisense oligonucleotides were chosen by computer prediction combined with oligonucleotide microarrays. The inhibition effect on MCF-7 cells proliferation was measured by MTT; and VEGFR2 expression was surveyed by Western-blotting and RT- PCR. Results: Using predicting secondary structure of VEGFR2 mRNA with RNA folding program, computer prediction designed 30 antisense oligonucleotide probes that were directed to local loose regions of RNA structure. In 30 probes, 4(4•30, 13.33%) antisense oligonucleotides showed strong hybridization intensities in oligonucleotide microarrays test and were selected. All these antisense oligonucleotides targeting 4 different sites of VEGFR2 mRNA lowered the level of VEGFR2 mRNA and protein present in MCF-7 cells. Proliferation of MCF-7 eel ls was reduced by 4 antisense oligonucleotides, respectively, in which asON1 was the most effective, with the inhibitory rates being 53.06% at 0.8 ~tmol/L. Conclusion: Combination of computer prediction with oligonucleotide microarrays is an effective way in selecting optimal antisense oligonucleotides. The antisense oligonucleotides showed good correlation between their antitumor activity and the hybridization intensities. The antisense oligonucleotides targeting VEGFR2 mRNA demonstrated prominent antitumor role in vitro.
Key words: Target selection; Computer prediction; Oligonucleotide microarrays; Antisense oligonucleotide; VEGFR2/KDR
Angiogenesis, the process by which capillaries sprout from preexisting blood vessels, is tightly regulated by a large number of proangiogenic and antiangiogenic factors. Tumor cells have an absolute requirement for a persistent supply of new blood vessels to nourish their
Received date: Mar. 28, 2005; Accepted date: May 16, 2005 Foundation item: This work was supported by the National Natural Sciences Foundation of China (No. 3017111) and National Project "863" (No. 2001AA234041). *Author to whom correspondence should be addressed. Phone: (0086-10)-66932211; E-mail: [email protected] Biography: ZHENG Su-jun (1973-), male, attending physician, now working at Beijing Youan Hospital, Capital University of Medical Sciences, majors in liver disease. E-mall: zheng003 @ sina.com
growth and to facilitate metastasis. Thus, tumor vascularization is a vital process for the progression of a neoplasm from a small localized tumor to an enlarging tumor with the ability to metastasize.
Vascular endothelial growth factor (VEGF), the most potent direct-acting angiogenic protein known tll, is a diffusible endothelial ceil (EC) specific mitogen and angiogenic factor that also increases vascular permeability. Endothelial cell survival in newly formed vessel is VEGF-dependent t21, VEGF overproduction has been identified as a major factor underlying pathological angiogenesis for tumor in vivo. VEGF plays its most biological function through binding to its receptor: Vascular endothelial growth factor receptor 2 t3' 41 (VEGFR2, also named as kinase insert domain- containing receptor:KDR). The VEGFR2/KDR gene has been mapped to human chromosome 4ql 1-q12. The full

162 Chinese Journal of Cancer Research 17(3): 161-170, 2005
length of VEGFR2 mRNA (GenBank Version: NM_002253.1, GI: 11321596) is 5830 nt, among which the fragment of 1-303 belongs to the 5'-nontranslated codon region; the fragment of 304-4374 is the coding sequence; 4375-5830 is 3'-nontranslated coding region sequence. VEGFR2/KDR is a type III receptor tyrosine kinase containing a kinase insert domain of 68 amino acids.
Interestingly, excepting that VEGFR2 is mostly expressed in endothelial cells, it is expressed in some tumor cells, and appears to play a pivotal role in vasculogenesis and tumor cell proliferation; some studies showed that VEGF is an autocrine growth factor for VEGF-receptor positive human tumors ts-71. So, inhibition of the VEGF tyrosine kinase signaling pathway could block new vessel formation in growing tumors, or inhibit the proliferation of tumor cells directly, leading to stasis or regression of tumor growth.
Antisense oligonucleotides (asONs) targeting VEGFR2 mRNA hold potential for tumor gene therapy. However, not all antisense oligonucleotides are equally effective in their ability to inhibit protein synthesis. There is now convincing evidence that binding of complementary oligonucleotide is mainly determined by the secondary and tertiary structure of the RNAs. Intramolecular base pairing into stable secondary and tertiary structures renders large parts of the molecule inaccessible to interacting with complementary nucleic acids t8]. The selection of effective antisense oligonucleotides pose a formidable technical challenge. In this study, we screened the accessible sites of VEGFR2 mRNA by combining computer prediction of mRNA secondary structure with oligonucleotide scanning arrays. Finally, in MCF-7 cells we investigate the antitumor activity of antisense oligonucleotides selected.
METHODS
Cloning VEGFR2/KDR Gene
The VEGFR2/KDR gene was divided into KDR (20-2823) and KDR2 (2711-5330) gene fragments to screen the mRNA antisense oligonucleotides accessible sites. Total RNA was isolated with TRIzol reagent (Life Technologies, USA) from A375 metastatic melanoma cells. VEGFR2/KDR cDNA was synthesized using a reverse transcription system (Invitrogen) with 2 ~tg total RNA. KDR1 and KDR2 gene fragments were amplified using the following primers:KDRl:(sense) 5'-GGA GAG CGG TCA GTG TGT GGT CGC TG-3'; (antisense): 5'-AGA GGC TTA CCT AGC T r c AGC CGG TCT CTG-3'; KDR2: (sense) 5'-CCA TCG TCA TGG ATC CAG ATG AAC TCC CAT-3'; (antisense): 5'-TGA
GTC GAC GAC ACC ACA CAC AGC TTC ACA TTG-3'; PCR was performed at 94°C 5 min, then 30 cycles at 94°C 45 S, at 58°C for 45 S, at 72°C for 2 min, and a final extension at 72°C for 5 rain. The PCR products were ligated with pGEMR-T vector and were named as pGEMR-T-KDR 1, pGEMR-T-KDR2, respectively. After sequencing analysis, the plasmids with the inserted fragments (direction: 5 ' -3 ' ) being the downstream of T7 promoter were selected for the following tests.
Fabrication of the Oligonucleotide Microarrays
The minimal free energy structures of KDR1 mRNA and KDR2 mRNA were analyzed with Mfold web server E91 under default folding conditions (37°C, 1 M NaC1, and no limit on distance between paired bases), respectively. 30 antisense oligonucleotides targeting to local loose structure of RNA, such as hairpin or stem-loop etc, were designed. Oligonucleotide probes were synthesized and modified with the 3' end amino to have a primary amino group for covalently binding to aldehyde-coated glass slides, and the amino group was linked through four polyethyleneglycol spacers to a specific sequence. All these probes were diluted with buffer (10 mmol/L Tris-HC1 pH 8.3, 50 mmol/L KC1 and 1.5 mmol/L MgC12) to a final concentration of 15 ~tmol/L, and were delivered to the microarray substrates (CSS-100 silylated glass slides) in single droplet with an average volume of 50 nl using a non-contract piezoelectric microdispenser (Cartisan). The spots were 500 Ixm in diameter.
Hybridization on Microarrays
The recombinant plasmids pGEMR-T-KDR1, pGEMR-T-KDR2 were lined with Nde I and KDR1 mRNA, KDR2 mRNA were transcribed by T7 RNA polymerase (Promega) in vitro, respectively. Internally radiolabelled transcripts for probing the microarrays were obtained in the presence of 100 Ix Ci [tx-32p] UTP (~3000 Ci/mmol, Amersham), 2 mmol/L each of ATP, CTP, GTP and 1 mmol/L unlabelled UTP (Promega). Microarrays were hybridized in a closed chamber as described tl°l. Briefly, approximately 5-8 pmol radiolabelled transcript was diluted in 10 ~tl hybridization solution (1 mol/L NaC1, 10 mmol/L, Tris-HC1 pH 8.0, 1 mmol/L EDTA, 0.01% SDS and 0.2 U/Ixl yeast tRNA) and hybridized at 37°C for 3 h. After washed in hybridization solution and dried, the microarray was exposed to a storage phosphor screen. Scanning the storage phosphor screen with Typhoon 9410 scanner system (Amersham Biosciences), the hybridization intensity was analyzed using program ImageQuant 5.2. Four pieces of antisense oligonucleotides with strong

Chinese Journal of Cancer Research 17(3):161-170, 2005 163
hybridization intensity were selected and synthesized with phosphorothioated modification. Their anti-tumor activity was investigated in the following tests.
Cell Culture
A375 and MCF-7 cell lines were cultured in DMEM (Gibcol BRL) containing 10% fetal calf serum, 100 U/ml penicillin and 100 mg/L streptomycin at 37°C in a humidified 5% CO2 incubator.
MTT Assay
MCF-7 cells were seeded into 96-well plates with 4 x 103 cells/well. 18 h later, the cells were treated with 0.2, 0.4, 0.8 ~tmol/L antisense oligonucleotides mixed with lipofectin (Invitogen, Carksbad, CA) or lipofetin alone according to the manufacturer' s instructions, respectively. After treatment for 56 h, 20 p.1 MTS (Promega) were added into each well and incubated for another 90 min at 37°C in a humidified atmosphere of 5% CO2. The absorbance at 490 nm was detected using VICTOR T M 1420 multilabel counter. Inhibition of cells proliferation was calculated by normalized OD of control sample according to the following formula:
Inhibition percentage (%) = OD~u coaaol - ODsampl¢
OD~u ~o~orODbl~k control x 1 0 0 %
Analysis of VEGFR2/KDR mRNA Expression by RT-PCR
A total of 1.5x105 MCF-7 cells/well were seeded in a 6-well plate. The cells were transfected with 4 phosphorothioated antisense oligonucleotides at the concentration of 0.8 Ixmol/L, respectively, using lipofectin according to the manufacture's instruction. 36 h later, total RNA was prepared by TRIzol reagent. RT-PCR reaction was performed using a reverse transcription system same as above, with 2 ~tg RNA, and ~-actin as internal control. The sense and antisense primers used for PCR amplification of VEGFR2 are 5'-GAAGATATCCCGTTAGAAGAACCAGAAGTA-3' (4021-4050); and 5'-ATCGGAGTGATATCCGGA- CTGGTAGCCGCT-3' (4210-4239), respectively. The primers for ~-actin are (sense): 5'-TTCAGGTTT- ACTCACGTCATCC-3', (antisense): 5'-CCAAATG- CGGCATCTrCAAACCC-3'; PCR was performed at 94°C for 5 min, then 30 cycles at 94°C for 30 s, at 58°C for 30 s, at 72°C for 30 s and a final extension at 72°C for 5 min. The anticipated 13-actin PCR products are 317 bp in length, and that of VEGFR2 are 219 bp in length.
The levels of VEGFR2 protein expression in cells treated with antisense agents were measured by Western-blotting. The treatment was performed using the same method as for the VEGFR2 mRNA level analysis described above. 56 h later, the cells were lysed in a lysis buffer containing 9.1 mmol/L Na2HPO4, 1.7 mmol/L
NaH2PO4 (pH: 7.4), 150 mmol/L NaC1, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, protease inhibitor cocktail tablets (Roche Diagnostics GmbH Mannheim, Germany) for 1 h on ice. Equal amount (22 ~tg) of protein was run on a 8% SDS-PAGE gel, and transferred onto PVDF membrane. The membrane was blocked with 5% nonfat-milk and incubated with primary antibody (1:1000) against VEGFR2 (Sigma) or primary antibody (1:2000) against [3-actin (Sigma), and followed with peroxidase-conjugated secondary antibodies (1:3000). Protein bands were visualized using ECL detection system (Promega). The intensity of the bands was scanned by Gel Doc 1000 (Bio-RAD), and the inhibition percentage (%) was calculated according to the following formula:
Inhibition percentage = (1-Asample/Acontrol) × 100%
Statistics Analysis
The data were subjected to ANOVA and expressed as means+ standard deviation (SD). A P value <0.05 was considered to be significant.
RESULTS
Plasmids Construction
The PCR products of KDR1, KDR2 were cloned into pGEMR-T vector, and named as pGEMR-T-KDR1, pGEMR-T-KDR2, respectively. Sequencing analysis showed that the VEGFR2/KDR gene fragments were correct.
Mfold-predicted Structures of the Transcripts
The minimal free energy structures of VEGFR2 mRNAs were predicted using the program mfold version 3.1. Base-pairing loose regions in RNA secondary structure are likely to be accessible for RNA-targeting nucleic acids. According to this principle, 30 antisense probes were designed (Table 1, Table 2). Figure 1 shows the target location of some antisense oligonucleotides.
Fabrication of the Oligonudeotide Microarrays
Western-blottingAnalysis Two sorts of microarrays were fabricated. The
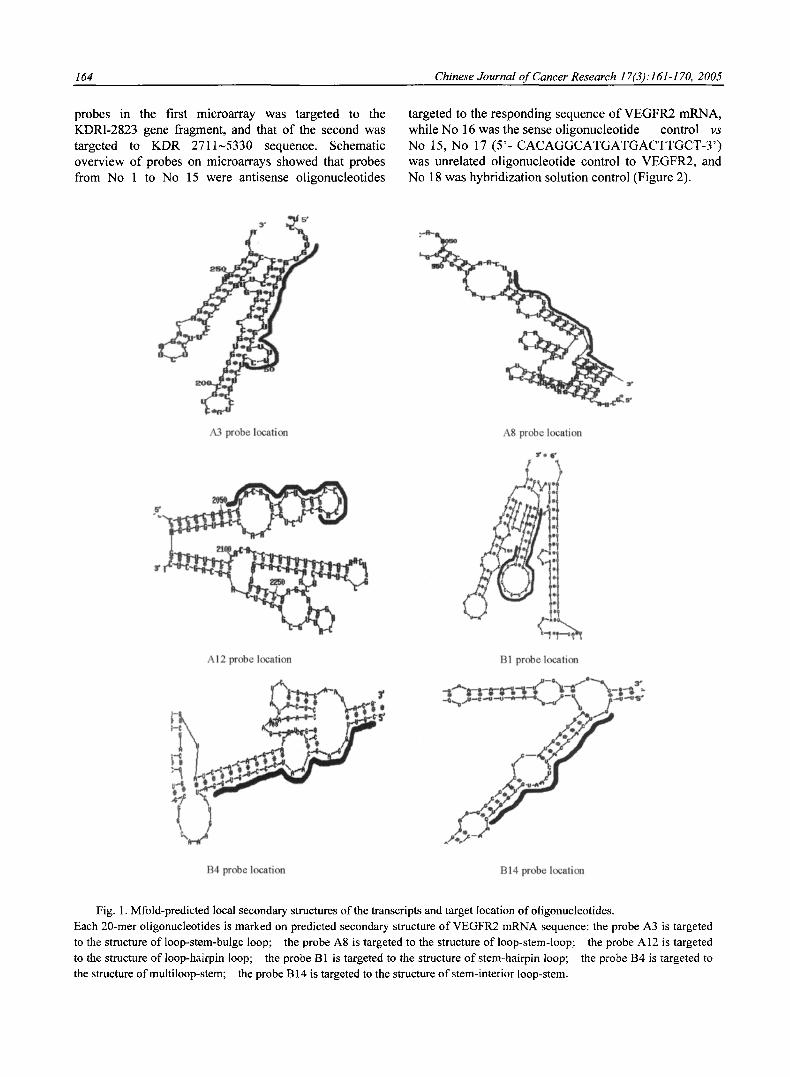
164 Chinese Journal of Cancer Research 17(3): 161-170, 2005
probes in the first microarray was targeted to the KDR1-2823 gene fragment, and that o f the second was targeted to KDR 2711~5330 sequence. Schematic overview o f probes on microarrays showed that probes from No 1 to No 15 were antisense oligonucleotides
targeted to the responding sequence o f VEGFR2 mRNA, while No 16 was the sense oligonucleotide control vs
No 15, No 17 ( 5 ' - C A C A G G C A T G A T G A C T T G C T - 3 ' ) was unrelated oligonucleotide control to VEGFR2, and No 18 was hybridization solution control (Figure 2).
5"
A3 probe location
A12 probe location
B4 probe location
A8 probe location
B1 probe location
G )J-
B 14 probe location
Fig. 1. Mfold-predicted local secondary structures of the transcripts and target location ofoligonucleotides. Each 20-mer oligonucleotides is marked on predicted secondary structure of VEGFR2 mRNA sequence: the probe A3 is targeted to the structure of loop-stem-bulge loop; the probe A8 is targeted to the structure of loop-stem-loop; the probe A12 is targeted to the structure of loop-hairpin loop; the probe B 1 is targeted to the structure of stem-hairpin loop; the probe B4 is targeted to the structure ofmultiloop-stem; the probe B 14 is targeted to the structure of stem-interior loop-stem.
Chinese Journal of Cancer Research 17(3):161-170, 2005 165
Analysis of Mieroarrays Hybridization
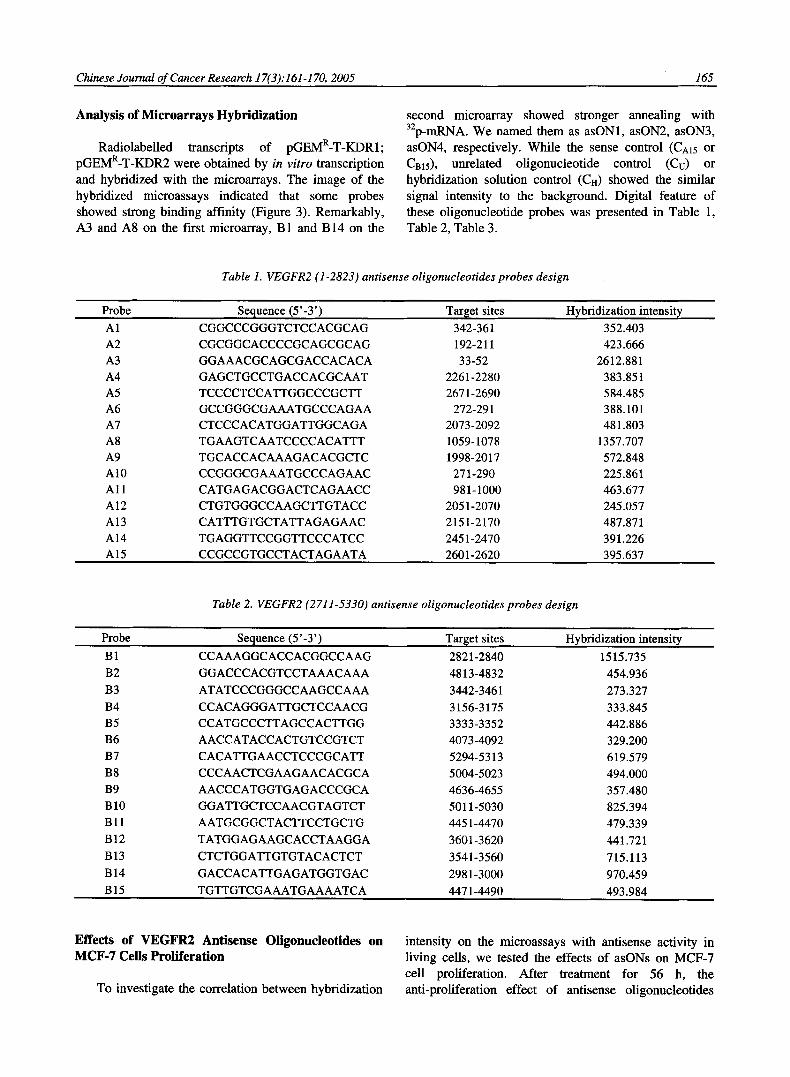
Radiolabelled transcripts of pGEMR-T-KDR1; pGEMR-T-KDR2 were obtained by in vitro transcription and hybridized with the microarrays. The image of the hybridized microassays indicated that some probes showed strong binding affinity (Figure 3). Remarkably, A3 and A8 on the first microarray, B 1 and B 14 on the
second microarray showed stronger annealing with 32p-mRNA. We named them as asON1, asON2, asON3, asON4, respectively. While the sense control (CA15 or Cms), unrelated oligonucleotide control (Ct:) or hybridization solution control (CH) showed the similar signal intensity to the background. Digital feature of these oligonucleotide probes was presented in Table 1, Table 2, Table 3.
Table 1. VEGFR2 (1-2823) antisense oligonucleotides probes design
Probe
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 All A12 A13 A14 A15
Sequence (5'-3') CGGCCCGGGTCTCCACGCAG CGCGGCACCCCGCAGCGCAG GGAAACGCAGCGACCACACA GAGCTGCCTGACCACGCAAT TCCCCTCCATTGGCCCGCTT GCCGGGCGAAATGCCCAGAA CTCCCACATGGATTGGCAGA TGAAGTCAATCCCCACATTT TGCACCACAAAGACACGCTC CCGGGCGAAATGCCCAGAAC CATGAGACGGACTCAGAACC CTGTGGGCCAAGCTTGTACC CATTTGTGCTATTAGAGAAC TGAGGTTCCGGTTCCCATCC CCGCCGTGCCTACTAGAATA
Tar~etsites
342-361 192-211 33-52
2261-2280 2671-2690
272-291 2073-2092 1059-1078 1998-2017
271-290 981-1000
2051-2070 2151-2170 2451-2470 2601-2620
Hybfidizationintensit~
352.403 423.666
2612.881 383.851 584.485 388.101 481.803
1357.707 572.848 225.861 463.677 245.057 487.871 391.226 395.637
Table 2. VEGFR2 (2711-5330) antisense oligonucleotides probes design
Probe
B1 B2 B3 B4 B5 B6 B7 B8 B9 B10 Bl l B12 B13 B14 B15
Sequence (5'-3') CCAAAGGCACCACGGCCAAG GGACCCACGTCCTAAACAAA ATATCCCGGGCCAAGCCAAA CCACAGGGATTGCTCCAACG CCATGCCCTTAGCCACTTGG AACCATACCACTGTCCGTCT CACATTGAACCTCCCGCATT CCCAACTCGAAGAACACGCA AACCCATGGTGAGACCCGCA GGATTGCTCCAACGTAGTCT AATGCGGCTACTTCCTGCTG TATGGAGAAGCACCTAAGGA CTCTGGATTGTGTACACTCT GACCACATTGAGATGGTGAC TGTTGTCGAAATGAAAATCA
Targetsites Hybddizafionin~nsity
2821-2840 1515.735 4813-4832 454.936 3442-3461 273.327 3156-3175 333.845 3333-3352 442.886 4073-4092 329.200 5294-5313 619.579 5004-5023 494.000 4636-4655 357.480 5011-5030 825.394 4451-4470 479.339 3601-3620 441.721 3541-3560 715.113 2981-3000 970.459 4471-4490 493.984
Effects of VEGFR2 Antisense Oligonucleotides on MCF-7 Cells Proliferation
To investigate the correlation between hybridization
intensity on the microassays with antisense activity in living cells, we tested the effects of asONs on MCF-7 cell proliferation. After treatment for 56 h, the anti-proliferation effect of antisense oligonucleotides

166 Chinese Journal of Cancer Research 17(3):161-170, 2005
(asONl~asON4) targeting 4 different sites on VEGFR2 mRNA is showed in Figure 4. The antisense oligonucleotides decreased the proliferation o f MCF-7 cells in a concentration dependent manner, in which asON1 was the most effective. At 0.8 gmol/L, the inhibitory rate o f asON1, asON1, asON2, asON3 asON4 were 53.06%, 35.18%, 42.55%, 24.63%, respectively. It was also showed that the inhibitory rates o f antisense oligonucleotides were posi t ively related to their hybridization intensity (Table 3). The antisense ol igonucleotide with stronger hybridizat ion intensity had more powerful anti-proliferation potency. Ol igonucleo t ide microarrays is an effective tool o f antisense oligonucleotides selection.
©@©@@@ © Q Q Q @ © @(gO©@®
, Fig. 2. Schematic overview of probes on microarrays. Probes from No 1 to No 15 were antisense oligonucleotides targeted to the responding sequence of VEGFR2 mRNA, while No 16 was the sense oligonucleotide control vs No 15, No 17 (Cu) was unrelated oligonucleotide control to VEGFR2, and No 18(CH) was hybridization solution control.
Table 3. The selection of VEGFR2 mRNA antisense oligonucleotides by microarrays
asON Sequence (5 ' - 3 ' ) Target sites Hybridization intensity Inhibitory rate (%)*
1 GGAAACGCAGCGACCACACA 33-52 2612.881 53.06%
2 TGAAGTCAATCCCCACATTT 1059-1078 1357.707 35.18%
3 CCAAAGGCACCACGGCCAAG 2821-2840 1515.735 42.55%
4 GACCACATTGAGATGGTGAC 2981-3000 970.459 24.63%
The inhibitory rates of MCF-7 cells proliferation was calculated when the cells were treated with antisense
oligonueleotides at 0.8~mol/L in MTT assay.
A1 A4 A7 A10 A13 CA15 B1 B4 B7 B10 B13 Cm5
A2 A5 A8 Al l A14 Cu B2 B5 B8 Bll B14 Cu
A3 A6 A9 A12 AI5 CH B3 B6 B9 B12 B15 CH
A B
Fig. 3. The image obtained after hybridization of the radiolabelled transcripts to the microarrays. A: The antisense probes were designed targeting to the different sites of KDR1-2823 mRNA. pGEMR-T-KDRI lined
with Nde I was transcribed in vitro and hybridization on microarrays. B: The antisense probes were designed targeting to the responding sites of KDR2711-5330 mRNA. pGEMR-T-KDR2
lined with Nde I was transcribed in vitro and hybridization on microarrays. Analysis of hybridization intensities by computer program showed that the No 3 (A3), No 8 (A8) in Figure A and No 1 (B1), No 14 (B14) in Figure B showed stronger binding affinity, named as asON1, asON2, asON3, asON4, respectively.
Effects of Ant isense Ol igonucleot ides on V E G F R 2 m R N A Express ion
It has been sugges ted that ant isense
o l igonuc leo t ides may inhibi t gene express ion th rough d iverse effects on t ranscr ip t ion and t rans la t ion , inhibi t ion o f m R N A t ranscr ip t ion can occur through format ion o f t r iple hel ixes wi th c o m p l e m e n t a r y
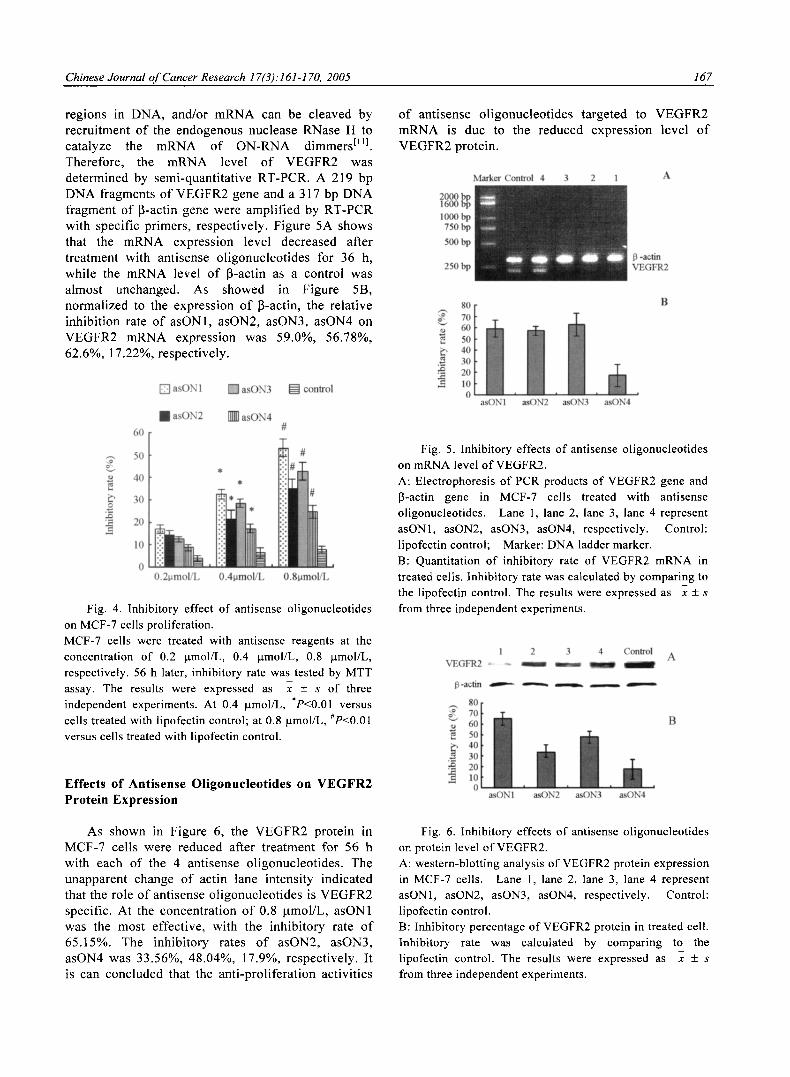
Chinese Journal o f Cancer Research 17(3).'161-170, 2005 167
regions in DNA, and/or m R N A can be c leaved by recrui tment o f the endogenous nuclease RNase H to ca ta lyze the m R N A o f O N - R N A dimmersElll . Therefore , the m R N A level o f V E G F R 2 was de te rmined by semi-quant i ta t ive RT-PCR. A 219 bp D N A fragments o f V E G F R 2 gene and a 317 bp D N A fragment o f 13-actin gene were ampl i f i ed by R T - P C R with specif ic pr imers , respect ive ly . F igure 5A shows that the m R N A express ion level decreased after t reatment with ant isense o l igonuc leo t ides for 36 h, whi le the m R N A level o f 13-actin as a control was a lmost unchanged. As showed in Figure 5B, normal i zed to the express ion o f 13-actin, the re la t ive inhibi t ion rate o f asON1, asON2, asON3, asON4 on V E G F R 2 m R N A express ion was 59.0%, 56.78%, 62.6%, 17.22%, respect ive ly .
[ ] asON 1 [ ] asON3 INt control
Z v
o
60
50
40
30
20
10
0
• asON2 [ ] asON4 #
#
.
0.2i~mol/L 0.4~mol/L 0.8~mol/L
Fig. 4. Inhibitory effect of antisense oligonucleotides on MCF-7 cells proliferation. MCF-7 cells were treated with antisense reagents at the concentration of 0.2 gmol/L, 0.4 gmol/L, 0.8 p.mol/L, respectively. 56 h later, inhibitory rate was tested by MTT assay. The results were expressed as x + s of three independent experiments. At 0.4 gmol/L, *P<0.01 versus cells treated with lipofectin control; at 0.8 p.mol/L, #P<0.01 versus cells treated with lipofectin control.
Effects o f A n t i s e n s e O l i g o n u c l e o t i d e s on V E G F R 2 Prote in Express ion
As shown in Figure 6, the V E G F R 2 prote in in MCF-7 cel ls were reduced after t rea tment for 56 h wi th each o f the 4 ant isense o l igonuc leo t ides . The unapparent change o f act in lane intensi ty indica ted that the role o f ant isense o l igonuc leo t ides is V E G F R 2 specif ic . At the concentra t ion o f 0.8 lamol/L, asON1 was the most effect ive, with the inhib i tory rate o f 65.15%. The inhibi tory rates o f asON2, asON3, asON4 was 33.56%, 48.04%, 17.9%, respect ive ly . It is can conc luded that the an t i -p ro l i fe ra t ion act iv i t ies
o f ant isense o l igonuc leo t ides ta rge ted to V E G F R 2 m R N A is due to the reduced express ion level o f V E G F R 2 protein.
3 2 1 A
t3 -actin VEGFR2
Z 0~
e~
80 70 60 50 40 30 20 10 0
asONl asON2 asON3 asON4
B
Fig. 5. Inhibitory effects of antisense oligonucleotides on mRNA level of VEGFR2. A: Electrophoresis of PCR products of VEGFR2 gene and fi-actin gene in MCF-7 cells treated with antisense ol!gonucleotides. Lane 1, lane 2, lane 3, lane 4 represent asONl, asON2, asON3, asON4, respectively. Control: lipofectin control; Marker: DNA ladder marker. B: Quantitation of inhibitory rate of VEGFR2 mRNA in treated cells. Inhibitory rate was calculated by comparing to the lipofectin control. The results were expressed as x + s from three independent experiments.
1 2 3 4 Control A VEGFR2 ~ ~ ~ ~ ~
13-acfin ~ " - ' - - , - ~ o r 1
B
e~
asON1 asON2 a~ON3 asON4
Fig. 6. Inhibitory effects of antisense oligonucleotides on protein level of VEGFR2. A: western-blotting analysis of VEGFR2 protein expression in MCF-7 cells. Lane 1, lane 2, lane 3, lane 4 represent asON1, asON2, asON3, asON4, respectively. Control: lipofectin control. B: Inhibitory percentage of VEGFR2 protein in treated cell. Inhibitory rate was calculated by comparing to the lipofectin control. The results were expressed as x +_ s from three independent experiments.

168 Chinese Journal of Cancer Research 17(3):161-170, 2005
DISCUSSION
VEGF binds to VEGFR/KDR, and regulates the neovascularization, progress, metastasis and proliferation of tumor. Most functional endothelial VEGF cell signaling described to date is mediated via VEGFR2 or strongly suspected to involve VEGFR2 on the basis of ligand specificity E12-141. VEGF plays its role through paracrine or autocrine mechanism. On the one hand, tumor cells secret VEGF, which binds to VEGFR2 on vascular endothelial cells and activates the signaling pathway, resulting to the neovascularization. On the other hand, some researches show that VEGF is an autocrine growth factor for VEGF receptor positive human tumors, such as Kaposi sarcoma, melanoma, ovarian carcinoma, pancreatic carcinoma, prostate carcinoma, etc. Inhibition of VEGF (VEGF antisense oligonucleotides) or VEGFR2 (neutralizing antibodies) decreased the proliferation of these cells in vitro t61.
Similarly, many studies indicated that VEGF and VEGFR2 are co-expressed in breast cancer. The presence of VEGF autocrine growth factor activity was demonstrated in some breast cancer cell lines (T-47D breast cancer cell) and in invasive breast cancer tissue t7' 15, 16]. VEGF increases the cellular invasion of T-47D cells on Matrigel/fibronectin- coated transwell membranes. VEGF induces the activation of the extracellular regulated kinases 1, 2 as well as activation of phosphatidylinositol 3'-kinase, Akt, and Forkhead receptor L1. VEGFR2 activation is enhanced in breast cancer in vivo and demonstrate the functional role of VEGF and VEGFR2 in the development of malignant breast disease. VEGF can induce breast cancer cel 1 proliferation directly t17" 181.
Compared with normal tissue, the expression level of VEGF and VEGFR2 in tumor are increased. Thus, VEGFR2 is a good target for gene therapy. Antisense oligonucleotide is an effective tool to block gene expression. The presence of autocrine growth pathway in breast cancer implies that VEGFR2 antisense therapy is acting on 2 levels: antiangiogenic effects on the tumor vasculature and antineoplastic effects on the tumor cell population. VEGFR2 expression in the tumor cells may thus predict for better response to VEGFR2 antisense therapy.
However, selection of asONs accessible gene target site is crucial to the success of antisense experiment. The selection of effective antisense sequence is difficult. Conventional approaches including empirical selection such as "sequence walking", oligonucleotide scanning arrays, RNase H mapping, have some defects. "Sequence walking" synthesize several oligonucleotides (usually 50-100), targeted to various region of an mRNA, and measure
their antisense activity. Only 2%-5% of the oligonucleotides are generally found to be effective. This method is always expensive, time-consuming, laborious and almost impossible to perform for some laboratory room. Using oligonucleotide scanning arrays, radiolabelled transcription of target gene mRNA are hybridized to the scanning arrays. Those oligonucleotides showing strong binding can be analyzed for antisense activity tl°l. However, it is difficult to fabricate long gene fragment oligonucleotides arrays, to date the longest sequence stretch analyzed by scanning arrays is 800 nt t191. The use of RNase H mapping is limited by its radioactive pollution and location of target sequence not precise enough.
There is compelling evidence that duplex formation between oligonucleotides and long nucleic acids is constrained by the secondary folding of the RNA and is not determined primarily by base composition or sequence ES' 2ol. It may be another choice to screen the target sites of antisense oligonucleotides according to the RNA structure. Using predicting secondary structure of target RNA with RNA folding programs, Computer prediction designs antisense oligonucleotides that binds to regions that are predicted free from intramolecular base pairing. Some researches showed that the secondary structure predictions of RNAs were mostly effective using these programs, and the antisense oligonucleotides designed with RNA accessibility prediction is consistent with experimental studies t21,22J. However, some studies reported that accessible sites can not be mapped on predicted structures and that there is no obvious structural difference between accessible and inaccessible regions on the computer folded structures, and just use of computer prediction methods are not reliable for designing antisense agents[23. 241
In order to avoid the limitations and utilize the rational parts of above screening approaches, in this study, we use the combination of computer prediction with microarrays to select the effective antisense oligonucleotides that blocks VEGFR2 expression. First, we predict the secondary structure of VEGFR2 mRNA, then selected the RNA loose structure regions, such as hairpin, stem-loop sequence to design antisense agents. Computer prediction decrease the selection regions, and reduced the expense of screening antisense reagents. Then we use oligonucleotides microarrays to choose the antisense probes designed by computer prediction again. On conventional oligonucleotides arrays, mask is used to retain the synthesis reagents and is moved along the surface by predetermined step size after each base coupling, with overlapping footprints. This kind of

Chinese Journal of Cancer Research 17(3): 161-170, 2005 169
oligonucleotide arrays is difficult to fabricate and expensive. Here, we make a microarray that contains just the antisense probes designed by computer prediction, and the probes are sprayed to glass slide in single droplet using a non-contract piezoelectric microdispenser. All this decreases the population of antisense probes and costs less. Combination of computer prediction with microarrays is performed easily and feasibly.
The results indicated that 4 probes (4/30, 13.33%), but not all antisense oligonucleotides targeted to the loose regions of predicted VEGFR2 mRNA showed strong hybridization intensity. In Figure 1, compared with the "loop-stem-bulge loop" structure of A3 probes, the "loop-hairpin loop" structure of A12 should be less stable. However, A12 got weaker hybridization intensity (Table 1). Similarly, the hybridization intensity of probe B4 was weaker than B14 (Table 2). All these revealed that it is not correct enough to predict the hybridization ability of antisense oligonucleotides just according to the secondary structure of RNA analyzed with computer. The possible mechanism is for long RNA molecules, energy calculations often return a number of different structures with similar free energies: there is difficulty in choosing the correct one [19]. To select effective antisense oligonucleotide, the increase in population of antisense oligonucleotides probes may be helpful. This also means that it is indispensable to further measure the antisense oligonucleotides hybridization ability with target mRNA using another empirical approach, such as oligonucleotide scanning arrays.
Using the way of combination, 4 pieces of antisense oligonucleotides with strong hybridization intensity were selected, blocked the VEGFR2 gene expression on the level of mRNA and protein, and reduced the proliferation of MCF-7 cells. The inhibition effect on VEGFR2 expression and MCF-7 cells proliferation correlated well with hybridization intensities, and this revealed that the oligonucleotide scanning arrays method is reliable. MCF-7 cells secret VEGF and also express VEGFR2, which means the autocrine mechanism regulates the happening and progressing of breast cancer. VEGF could stimulated PLC- T and ERK activity in VEGFR2-expressing NIH3T3 cells and VEGFR2 is strong implicated in activation of the P13K/AKT pathway tT' 2s, 261. The possible mechanism of inhibition MCF-7 proliferation by asONs may be the blockage of VEGFR2 gene expression results in that the PLC-y or AKT pathway could not be effectively activated, then inducing tumor cell happening apoptosis or necrosis.
However, as shown in Figure 5 and Figure 6, there were some inconsistency between the asONs
inhibitory rate of VEGFR2 mRNA expression with that of VEGFR2 protein expression. Despite the inhibitory rate of asON1 is lower than that of asON3 (59.0% vs 62.6%) on VEGFR2 mRNA expression, the inhibitory rate of asON1 is higher than that of asON3 (65.15% vs 48.04%) on VEGFR2 protein expression. The possible mechanism is that asON1 is targeted to VEGFR2 mRNA 5'-nontranslated codon region (target position: 33-52), and inhibits VEGFR2 protein expression through two aspects: RNase H-dependent reduction of targeted RNA and translational machinery prevention by oligonucleotides steric- blockage; whereas asON3 targeted to RNA coding region (2821-2840 nt) reduced VEGFR2 protein expression mostly by RNase H-dependent degradation of mRNA t27]. But the mechanisms by which asONs modulating transferring of the genetic information to protein are subtle and complex, there maybe other unknown reasons.
Antisense oligonucleotides targeted to VEGFR2 are hopeful to become new reagents of antitumor. We will further to investigate their antitumor mechanism, and to explore their antitumor role in vivo.
REFERENCES
[1] Ferrara N, Davis-Smyth T. The biology of vascular endothelial growth factor [J]. Endocr Rev 1997; 18:4-25.
[2] Alon T, Hemo I, Ltin A, et al. Vascular endothelial growth factor acts as a survival factor for newly formed retinal vessels and has implications for retinopathy of prematurity [J]. Nat Med 1995; 1:1024-8.
[3] Terman BI, Dougher-Vermazen M, Carrion ME, et al. Identification of the KDR tyrosine kinase as a receptor for vascular endothelial cell growth factor [J]. Biochem Biophys Res Commun 1992; 187:1579-86.
[4] Klagsbrun M, D'Amore PA. Vascular endothelial growth factor and its receptors [J]. Cytokine Growth Factor Rev 1996; 7:259-70.
[5] Zhang H, Li Y, Li H, et al. Inhibition of both the autocrine and the paracrine growth of human leukemia with a fully human antibody directed against vascular endothelial growth factor receptor 2 [J]. LeukLymphoma 2004; 45:1887-97.
[6] Masood R, Cai J, Zheng T, et al. Vascular endothelial growth factor (VEGF) is an autocrine growth factor for VEGF receptor- positive human tumors [J]. Blood 2001; 98:1904-13.
[7] Price D J, Miralem T, Jiang S, et al. Role of vascular endothelial growth factor in the stimulation of cellular invasion and signaling of breast cancer cells

170 Chinese Journal of Cancer Research 17(3):161-170, 2005
[J]. CellGrowth Differ 2001; 12:129-35. [8] Frauendorf A, Engels JW. Interaction of linear and
folded modified antisense oligonucleotides with sequences containing secondary structure elements [J]. Bioorg Med Chem Lett 1994; 4:1019-24.
[9] Zuker M. Mfold web server for nucleic acid folding and hybridization prediction [J]. Nucleic Acids Res 2003; 31:3406-15.
[10] Milner N, Mir KU, Southern EM. Selecting effective antisense reagents on combinatorial oligonucleotide arrays [J]. Nat Biotechnol 1997; 15:537-41.
[11] Green DW, Roh H, Pippin J, et al. Antisense oligonucleotides: an evolving technology for the modulation of gene expression in human disease [J]. J Am Col l Surg 2000; 191:93-105.
[12] Zachary I, Gliki G. Signaling transduction mechanisms mediating biological actions of the vascular endothelial growth factor family [J]. Cardiovasc Res 2001; 49:568-81.
[13] Cross MJ, Dixelius J, Matsumoto T, et al. VEGF-receptor signal transduction [J]. Trends Biochem Sci 2003; 28: 488-94.
[14] Waltenberger J, Claesson-Welsh L, Siegbahn A, et al. Different signal transduction properties of KDR and Fit-l, two receptors for vascular endothelial growth factor [J]. J Biol Chem 1994; 269:26988-95.
[15] Ryden L, Linderholm B, Nielsen NH, et al. Tumor specific VEGF-A and VEGFR2/KDR protein are co-expressed in breast cancer [J]. Breast Cancer Res Treat 2003; 82:147-54.
[16] de Jong JS, van Diest PJ, van der Valk P, et al. Expression of growth factors, growth inhibiting factors, and their receptors in invasive breast cancer. I: An inventory in search of autocrine and paracrine loops [J]. J Pathol 1998; 184:44-52.
[17] Kranz A, Mattfeldt T, Waltenberger J. Molecular mediators of tumor angiogenesis: enhanced expression and activation of vascular endothelial growth factor receptor KDR in primary breast cancer [J]. Int J Cancer 1999; 84:293-8.
[18] Nakopoulou L, Stefanaki K, Panayotopoulou E, et al.
Expression of the vascular endothelial growth factor receptor-2/Flk-1 in breast carcinomas: correlation with proliferation [J]. Hum Pathol 2002; 33:863-70.
[19] Sohail M, Southern EM. Selecting optimal antisense reagents [J]. Adv Drug Deliv Rev 2000; 44:23-34.
[20] Southern EM, Case-Green SC, Elder JK, et al. Arrays of complementary oligonucleotides for analysing the hybridisation behaviour of nucleic acids [J]. Nucleic Acids Res 1994; 22:1368-73.
[21] Kretschmer-Kazemi Far R, SczaKiel G. The activity of siRNA in mammalian cells is related to structural target accessibility: a comparison with antisense oligonucleotides [J]. Nucleic Acids Res 2003; 31:4417-24.
[22] Scherr M, Rossi JJ, Sczakiel G, et al. RNA accessibility prediction: a theoretical approach is consistent with experimental studies in cell extracts [J]. Nucleic Acids Res 2000; 28:2455-61.
[23] Sohail M, Akhtar S, Southern EM. The folding of large RNAs studied by hybridization to arrays of complementary oligonucleotides [J]. RNA 1999; 5:646-55.
[24] Schuster P, Stadler PF, Renner A. RNA structure and folding: From conventional to new issues in structure predictions [J]. Curr Opin Struct Biol 1997; 7:229-35.
[25] Takahashi T, Shibuya M. The 230 kDa mature form of KDR/Flk-1 (VEGF receptor-2) activates the PLC-gamma pathway and partially induces mitotic signals in NIH3T3 fibroblasts [J]. Oncogene 1997; 14:2079-89.
[26] Gerber HP, McMurtrey A, Kowalski J, et al. Vascular endothelial growth factor regulates endothelial cell survival through the phosphatidylinositol 3'-kinase/AKT signal transduction pathway. Requirement for Flk-1/KDR activation [J]. J Bio Chem 1998; 273:30336-43.
[27] Dias N, Stein CA. Antisense oligonucleotides: basic concepts and mechanisms [J]. Mol Cancer Ther 2002; 1:347-55.




















