
Elk decision-making rules are simplified in the presence of wolves

Secret lives of maned wolves (Chrysocyon brachyurus Illiger1815): as revealed by GPS tracking collars
L. F. Bandeira de Melo1, M. A. Lima Sabato1, E. M. Vaz Magni1, R. J. Young2 & C. M. Coelho1
1 Fundacao Zoo-Botanica, Belo Horizonte, Minas Gerais, Brazil
2 Conservation, Ecology and Animal Behaviour Group, Mestrado em Zoologia, Pontifıca Universidade Catolica de Minas Gerais, Belo Horizonte,
Minas Gerais, Brazil
Keywords
Chrysocyon brachyurus; maned wolves; GPS
collar; socio-spatial dynamics; home range.
Correspondence
Robert John Young, Conservation, Ecology
and Animal Behaviour Group, Predio 41,
Mestrado em Zoologia, Pontifıca
Universidade Catolica de Minas Gerais, Av.
Dom Jose Gaspar, 500 Coracao Eucarıstico,
30535-610 Belo Horizonte, Minas Gerais,
Brazil. Tel: +55 31 3319 4936;
Fax: +55 31 3319 4938
Email: [email protected]
Received 20 December 2005; accepted
11 April 2006
doi:10.1111/j.1469-7998.2006.00176.x
Abstract
The maned wolf Chrysocyon brachyurus is a nocturnal and shy species, which has
proven difficult to study in the field; consequently, data about its behavioural
biology are almost absent from the scientific literature. However, recent advances
in global positioning system (GPS) technology mean that it is now possible to
study the socio-spatial dynamics of this species in the field. In the present study
three related maned wolves (an adult pair and their juvenile female offspring) were
monitored by GPS tracking collars for a 6-month period, which included the birth
of a litter of pups to the adult pair. The three GPS collars were programmed to
record the position of the wearer every 2 h (simultaneously for all three indivi-
duals). Analyses of the data from the three individuals showed that the female had
the largest home range, then the male and finally the juvenile. Furthermore, there
was considerable overlap in home-range use by all three individuals. The home
ranges of the adult pair also varied in relation to the birth of pups (decreasing
dramatically and then showing a slow increase). During the night, which is the
normal period of activity for this species, the three individuals avoided contact
with each other. However, during the day, the adult pair usually slept together and
their juvenile slept within a few hundred metres of their location. The distance
travelled at night was greatest for the female, then the male and finally the juvenile.
These data suggest that (1) the social bond between the male and female is strong,
(2) maned wolves are highly tolerant of their juvenile offspring, even in the case of
new pups being born, and (3) that maned wolves do not hunt together and, in fact,
avoid each other when hunting.
Introduction
The maned wolf Chrysocyon brachyurus is the largest South
American canid with a wide distribution in the savannas
of Latin America. Despite being a characteristic species of
the Brazilian cerrado (savanna environment), relatively
little is known about the behavioural biology of this
species. The classic field study on the biology of the maned
wolf by Dietz (1984) is now more than 20 years old, and
arguably our knowledge about the wild behaviour
of this species has not increased greatly since this study.
The study by Dietz (1984) was based on a mixture of
methodologies, including radio-tracking, collection of
faeces and interviews with local people. Unfortunately,
much of the data we have on the species’ social and
reproductive behaviours comes from interviews of local
people and not from quantitative studies. Our lack of
knowledge concerning these aspects of the natural history
of maned wolves results largely from their secretive and
nocturnal habits.
The few field studies that have been published on maned
wolves have largely concentrated on analysing the species’
diet (i.e. trophic ecology; Dietz, 1984; Motta-Junior et al.,
1996; Motta-Junior, 2000; Bueno, Belentani & Motta-
Junior, 2002; Motta-Junior & Martins, 2002) owing to the
ease with which faeces can be collected (maned wolf faeces
have a strong and pungent odour), or have captured maned
wolves to investigate their health status (e.g. Carvalho &
Vasconcellos, 1995). Recently, Silva & Talamoni (2004)
tried to investigate the centre of activity of maned wolves in
the field from their defecation sites and tracks, and a few
authors have attempted to estimate the species’ home range
using radio-tracking studies (e.g. Dietz, 1984). Behavioural
studies of maned wolves have been restricted to captive
studies, for example studies on parental behaviour (Veado,
1997) and reproductive behaviour (Dietz, 1984). The field-
work conducted by Dietz (1984) has been hugely influential
in promoting the hypothesis that maned wolves are nor-
mally solitary animals, which come together only for repro-
duction. However, this conclusion is based on almost no
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London 27
Journal of Zoology. Print ISSN 0952-8369

observational data, and captive studies, including that
conducted by Dietz (1984), have suggested a reasonably
high level of sociability, for example male provisioning
of food to pups (Dietz, 1984; Rasmussen & Tilson, 1984;
Veado, 1997).
The maned wolf is presently listed as ‘near threatened’ in
the IUCN’s Red List of 2004 (Sillero-Zubiri & Hoffmann,
2004), and this classification is based largely on population
sizes and not on an in-depth understanding of the species’
biology. Therefore, presently in Brazil the maned wolf is
increasingly being studied both in situ and ex situ owing to
our lack of knowledge about this species and its potential as
a flagship or umbrella species.
Classically, large mammal species with secretive and/or
nocturnal habits have been studied in the wild with the
aid of radio-telemetry devices. However, such devices
have a number of limitations, for example the need to
triangulate the position of the animal (Saltz, 1994)
and the risk of disturbing the animal. In a species
such as the maned wolf, which when active is constantly
in rapid locomotion, the process of triangulation to
locate the animal is not only difficult but also relat-
ively inaccurate (Samuel & Fuller, 1994; Theuerkauf &
Jedrzejewski, 2002). Other methods of assessing areas
of spatial activity have been attempted using location
of faeces and tracks, and the marking of collection
points with a global positioning system (GPS) device
(Silva & Talamoni, 2004). Recently, GPS technology
has advanced to the point where GPS devices can be
included in the collars placed on wild animals (Rutter,
Beresford & Roberts, 1997; Blake, Douglas-
Hamilton & Karesh, 2001; McCarthy et al., 2005) and
thereby allow tracking. If GPS collars are placed
on several individuals of the same social group, it
becomes possible to measure group socio-spatial
dynamics. The spatial distribution of individuals of a
species can reveal interesting information about the
species’ social organization. For example, a number of
authors have asserted that the maned wolf is monogamous
and territorial, but this is based on fortuitous (ad hoc)
field observations and radio-telemetry studies (e.g. Dietz,
1984, 1985).
Although ad hoc field observations of maned wolves can
be helpful in understanding this species, these are frequently
limited to daylight hours (e.g. Dietz, 1984). A number of
species show temporal variation in their expression of social
behaviour. The classic example is the domestic cat Felis
catus, which during its period of inactivity, daylight hours,
often does not seek the company of conspecifics. However,
at night, during its period of activity, it seeks encounters
with conspecifics and can, under certain environmental
conditions, be highly social (Crowell-Davis, Curtis &
Knowles, 2004).
In this paper we report the spatial relationships and home
range of three related maned wolves (an adult pair and
a juvenile female), all monitored simultaneously by GPS
collars before and after the adult pair produced a litter
of pups.
Methods
Animals
The subjects of this study were three wild maned wolves
C. brachyurus: one adult female, one adult male (both of
unknown ages) and one juvenile female (estimated to be
1 year old, not sexually mature and assumed to be
the offspring of the adult male pair). Before the start of
the study, beginning the wolves were captured using a
wooden trap as described by Dietz (1984); the male
was caught on 7 May 2003, the female on 8 May 2003 and
the juvenile on 26 February 2003. The use of this type of trap
is considered a humane way of capturing wild maned
wolves.
Immediately after being captured, each maned wolf was
anaesthetized by a qualified veterinary surgeon, who then
clinically examined the health status of the individual
(e.g. presence of skin parasites), and biometric measure-
ments were taken. Following this a GPS radio-transmitting
collar (see below) was placed on the individual, the animal
was then allowed to recover from the anaesthesia and, when
the veterinary surgeon considered the animal to have fully
recovered, it was released. At the end of the study the
subjects were recaptured to have their collar removed or
the battery changed.
Study site
Our study was conducted at the Galheiros Reserve [RPPN
Estacao Ambiental de Galheiro, which is a private nature
reserve belonging to and managed by CEMIG (Companhia
Energetica de Minas Gerais)] in Perdizes, in the west of
Minas Gerais State, Brazil. The reserve has an area of
2847 ha and is largely a cerrado (Brazilian savanna) habitat
with some Atlantic rainforest influence. The reserve undu-
lates substantially with minimum altitudes of 815m and
maximum altitudes of 929m. The vegetation can be mainly
classified as cerrado fields or dense cerrado shrubland; there
is also some gallery forest within the reserve. The low-
altitude section of the reserve is comprised of forests and an
artificial reservoir that provides water for Nova Ponte
hydro-electricity station. The reserve is well protected from
illegal environmental exploitation, such as hunting. It can be
considered an ‘island’ of natural habitat, because it is
completely surrounded by farmland where soya beans are
grown and cattle are raised. Inside the reserve are a number
of administrative buildings and a visitor centre, which
provides environmental education opportunities for the
schools of the region.
The study was conducted in the dry season from
the month of May to October, with total rainfall for
this period usually having a mean value of less than 30mm.
During this season in the cerrado, food in the form of
small rodents is considered to be more abundant than
in the wet season (Dietz, 1984; Alho, Pereira & Paula,
1986).
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London28
The secret lives of maned wolves L. F. Bandeira de Melo et al.

GPS collar
The GPS collar was model ‘GPS-Simplex’ manufactured by
TELEVILT (Sweden) and had a total weight of 700 g, which
corresponds to c. 2% of the body weight of an adult maned
wolf. This type of telemetry device permits two types of data
collection: (1) the manual use of a receiver (RX-900; Tele-
vilt, Lindesberg, Sweden) to locate the wearer in the classic
manner of radio-telemetry studies (i.e. through triangula-
tion) and (2) the automatic storage of satellite position data
(i.e. GPS coordinates of the wearer), which can be subse-
quently downloaded by radio transmission to a receiver in
the field (when within a few kilometres of the animal’s
location) for subsequent analysis (using the software ‘Sim-
plex Project Manager V 1.2.5’). The data reported in this
study were collected using only the second method. The
collar of each animal was programmed to store the wearer’s
geographical coordinates every 2 h (1, 3, 5 until 23 h) during
the life of the battery (until 11 October 2003 in the case of
the male and female). Once a month during the field study,
the field biologists downloaded the data to the receiver. The
GPS collars provided us with the identity of the wearer, its
geographical location (hours, minutes and seconds, later
converted to UTM format by the management software),
and the date and time when location was recorded. Unfor-
tunately, if the wearer of the collar was in dense under-
growth the GPS device was not able to locate itself and,
therefore, the data do not correspond perfectly to the
number of days of the study multiplied by 12 two-hourly
data points (see Results).
Field study
During the study period fromApril 2003 to the present date,
two biologists made monthly visits to the study area to
download the GPS data and to collect other information
concerning the biology of maned wolves. For example,
camera trapping was used to detect the presence of large
predators, along with other species in the reserve, and to
observe the movement of the maned wolves, especially with
their pups. Maned wolves were not radio-tracked during
these field campaigns to avoid disturbance of their natural
behaviour.
Data analysis
The data used in this article only pertain to the period when
all three GPS collars were functioning, which was from
8 May 2003 to 11 October 2003, as we wish to analyse the
social spatial dynamics of the three related individuals. To
calculate sizes of home ranges and distances moved, we used
all the data available for the adult female (1668 fixes), adult
male (1676) and juvenile female (1553). To calculate spatial
relationships between individuals, we used only data when
the three GPS collars recorded the three individuals on the
same day at the same time (total number of fixes=1502).
Although we did not test the accuracy of our GPS collars,
a recent study has reviewed the accuracy of consumer-grade
GPS devices and found them to have a mean accuracy of 5m
in open habitats and a maximum error of 15m (Wing,
Eklund & Kellogg, 2005).
The home-range areas and their areas of overlap were
calculated using the software ‘Biota 1.03 alpha’. The dis-
tances between individuals and their movement between
fixes were calculated in Microsoft Excel (XP Office Suite)
using the following equation after the data had been
converted to UTM format:
d2ab ¼ ðXb� XaÞ2 þ ðYb� YaÞ2
where d is the distance between the points, Xa is the X
coordinate of the first point, Ya is the Y coordinate of the
first point,Xb is theX coordinate of the second point andYb
is the Y coordinate of the second point.
These data were then subsequently statistically analysed
in MINITAB 13 for Windows. The data were tested for
normality using the Anderson–Darling test and found not to
be normally distributed; therefore we opted to analyse the
data using non-parametric statistical tests.
Results
Home-range size
The estimated home-range sizes for the three individuals
varied considerably between individuals and by method of
estimation [minimum convex polygon (MCP) or fixed ker-
nel; Table 1]. Furthermore, the number of fixes (locations)
required before the home ranges of individuals reached an
asymptote also varied greatly (from c. 100 for the adult
female, 375 for the adult male and 1075 for the juvenile
female; see Fig. 1).
Effect of a birth on home-range size and dailyranging
The data showed that the adult female gave birth to an
unknown number of pups on 17 June 2003. To investigate
the effects of a birth on home range, the home ranges of the
three individuals were calculated every 5 days from 40 days
before the birth and for 110 days after the birth (Fig. 2). A
5-day period was chosen as this is a significant period of time
in relation to the gestation period of maned wolves (66 days;
Dietz, 1984), and the large quantity of data we collected
allowed such analyses to be undertaken. We can see that the
home-range sizes of the adult male and juvenile female
remained fairly constant during the observation period,
whereas the female’s home range reduced dramatically after
Table 1 Number of fixes (location) and home ranges (km2) of three
maned wolves Chrysocyon brachyurus calculated using minimum
convex polygon (MCP) and fixed kernel estimators
Individual Fixes MCP 100 MCP 99 MCP 95 Kernel 95
Female 1668 69.10 55.62 48.70 40.17
Male 1676 50.10 40.95 28.97 36.10
Juvenile 1553 39.40 33.78 20.93 7.19
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London 29
The secret lives of maned wolvesL. F. Bandeira de Melo et al.

the birth of the pups and thereafter started to increase with
time. Daily ranging did not vary greatly between individuals
or months, even during the period of the birth (Fig. 3),
except during the month of June when the adult female
dramatically reduced her ranging when she gave birth. Daily
ranging returned to pre-birth levels only 5 days after the
birth, whereas home-range size returned to normal only
15–20 days after the birth (see Fig. 2).
Home-range overlap
The data showed that there was considerable overlap
(median=97%; minimum=80% and maximum=100%)
in the use of home ranges by the three individuals. A
Kruskal–Wallis non-parametric analysis of variance showed
that there was no significant difference in the amount of
monthly home-range overlap between the three possible
combinations of individuals (H=1.13; d.f.=2; P=0.568).
There were no significant correlations between home-range
overlap and time (5-day measurements or months), and no
discernible pattern of home-range overlap in relation to time
was observed.
Spatial relationships between individuals
A Friedman test showed that there were no significant
differences between monthly inter-individual distances
(Fr=1.33; d.f.=2; P=0.5134). However, there were sig-
nificant differences between months (Fr=12.33; d.f.=5;
P=0.0305; see Fig. 4), and the post hoc Tukey-type tests
0
10
20
30
40
50
60
70
25 100
175
240
310
385
460
535
615
690
770
845
92010
0010
7511
4512
2012
9013
6514
3515
0515
7516
25
Cumulative number of fixes
Female Male Juvenile
Hom
e ra
nge
(km
)
Figure 1 Cumulative curves of home-range
areas (MCP 100) plotted against cumulative
number of fixes (locations) for the three
maned wolves Chrysocyon brachyurus. MCP,
minimum convex polygon.
05
101520253035404550
−40 −35
−30
−25
−20
−15
−10 −5 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 10
010
511
0
Days in relation to birth
Hom
e ra
nge
(km
)MaleFemale Juvenile
Figure 2 Relationship between home-range
sizes (MCP 100) and birth for three maned
wolves Chrysocyon brachyurus. MCP, mini-
mum convex polygon.
0
2000
4000
6000
8000
10 000
12 000
14 000
−21
−19
−17
−15
−13
−11 −9 −7 −5 −3 −1 1 3 5 7 9 11 13 15 17 19 21
Days in relation to birth event
Tot
al d
ista
nce
trav
elle
din
24
h (m
)
Female Macho Juvenile
Figure 3 Distances travelled in 24-h periods
by the three maned wolves Chrysocyon
brachyurus in relation to the birth of pups.
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London30
The secret lives of maned wolves L. F. Bandeira de Melo et al.
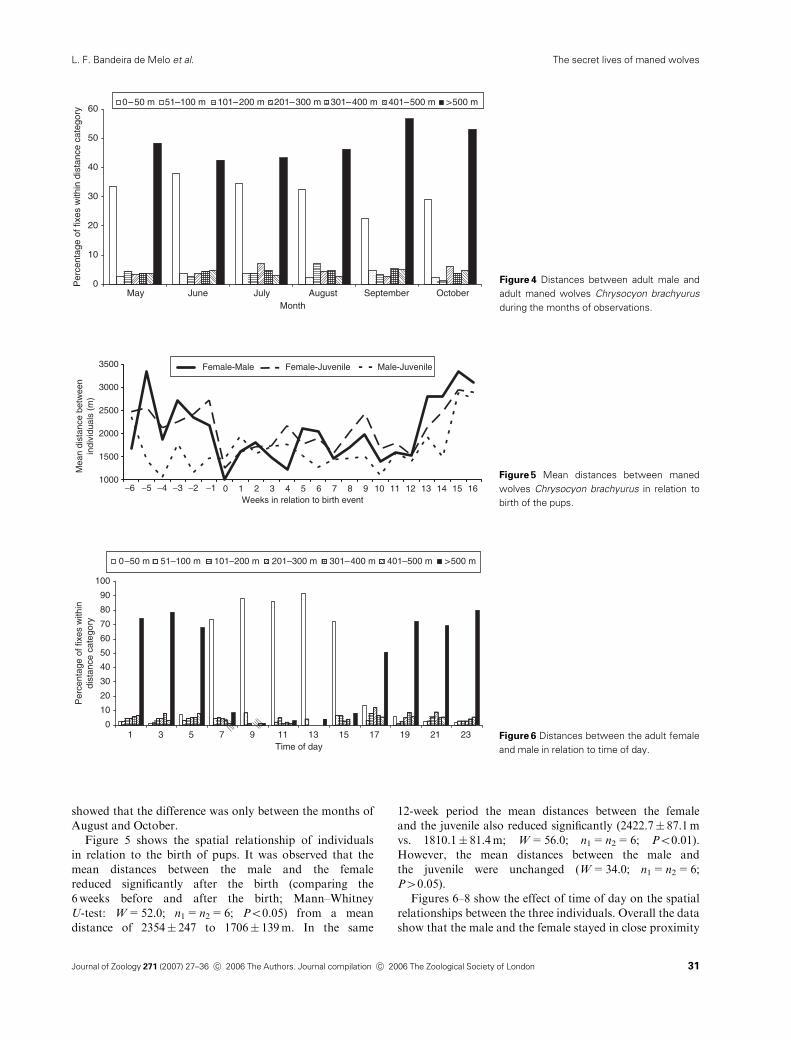
showed that the difference was only between the months of
August and October.
Figure 5 shows the spatial relationship of individuals
in relation to the birth of pups. It was observed that the
mean distances between the male and the female
reduced significantly after the birth (comparing the
6weeks before and after the birth; Mann–Whitney
U-test: W=52.0; n1=n2=6; Po0.05) from a mean
distance of 2354� 247 to 1706� 139m. In the same
12-week period the mean distances between the female
and the juvenile also reduced significantly (2422.7� 87.1m
vs. 1810.1� 81.4m; W=56.0; n1=n2=6; Po0.01).
However, the mean distances between the male and
the juvenile were unchanged (W=34.0; n1=n2=6;
P40.05).
Figures 6–8 show the effect of time of day on the spatial
relationships between the three individuals. Overall the data
show that the male and the female stayed in close proximity
0
10
20
30
40
50
60
May June July August September OctoberMonth
Per
cent
age
of fi
xes
with
in d
ista
nce
cate
gory
0–50 m 51–100 m 101–200 m 201–300 m 301–400 m 401–500 m >500 m
Figure 4 Distances between adult male and
adult maned wolves Chrysocyon brachyurus
during the months of observations.
1000
1500
2000
2500
3000
3500
−6 −5 −4 −3 −2 −1 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16Weeks in relation to birth event
Mea
n di
stan
ce b
etw
een
indi
vidu
als
(m)
Female-Male Female-Juvenile Male-Juvenile
Figure 5 Mean distances between maned
wolves Chrysocyon brachyurus in relation to
birth of the pups.
0
10
20
30
40
50
60
70
80
90
100
1 3 5 7 9 11 13 15 17 19 21 23Time of day
Per
cent
age
of fi
xes
with
indi
stan
ce c
ateg
ory
0–50 m 51–100 m 101–200 m 201–300 m 301–400 m 401–500 m >500 m
Figure 6 Distances between the adult female
and male in relation to time of day.
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London 31
The secret lives of maned wolvesL. F. Bandeira de Melo et al.

to each other during daylight hours, whereas all individuals
avoided each other at night.
Daily range
The female maned wolf moved the greatest distances
between sampling points per month, except for the month
of June when she gave birth (see Fig. 3). However, a
Kruskal–Wallis non-parametric analysis of variance showed
that these differences between individuals were not statisti-
cally significant (H=2.95; d.f.=2; P=0.229). The data for
the whole study period showed that during an average day
(12 sample points), the female moved a mean total of
9039� 238m (mean� SEM; minimum=112 and max-
imum=15 807), the male 7012� 192m (mean� SEM; mini-
mum=857 and maximum=15 388) and the juvenile
6165� 161m (mean� SEM; minimum=2588 and max-
imum=12 062). A Friedman repeated-measures analysis of
0102030405060708090
100
Time of day
Per
cent
age
of fi
xes
indi
stan
ce c
ateg
ory
0–50 m 51–100 m 101–200 m 201–300 m 301–400 m 401–500 m >500 m
1 3 5 7 9 11 13 15 17 19 21 23Figure 7 Distances between the adult male
and the juvenile female in relation to time
of day.
0102030405060708090
100
Time of day
Per
cent
age
of fi
xes
indi
stan
ce c
ateg
ory
0–50 m 51–100 m 101–200 m 201–300 m 301–400 m 401–500 m >500 m
1 3 5 7 9 11 13 15 17 19 21 23Figure 8 Distances between the adult female
and the juvenile female in relation to time
of day.
0
200
400
600
800
1000
1200
1400
1600
1 3 5 7 9 11 13 15 17 19 21 23Time of day (h)
Med
ian
dist
ance
mov
ed (
m)
Female Male Juvenile
Figure 9 Median distance moved (m) between
sample points in relation to time of day.
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London32
The secret lives of maned wolves L. F. Bandeira de Melo et al.

variance showed that these daily median totals were signifi-
cantly different (Fr=75.38; d.f.=2; Po0.001), and a post
hoc Tukey-type test showed that the female’s median daily
total was significantly greater than that of both the male and
the juvenile (there was no statistically significant difference
between the juvenile and the male). A Spearman rank
correlation of the daily median total distance travelled
against month showed significant positive correlations for
the female (rs=0.943; n=6; Po0.01) and the juvenile
(rs=1.000; n=6; Po0.001) but not for the male
(rs=0.558; n=6; P=0.228).
Activity during 24 h
Figure 9 shows that the three individuals displayed predo-
minantly nocturnal activity throughout the study period
from May to October. During this period sunrise would
have occurred between 06:00 and 07:00 h, and sunset
between 17:30 and 18:30 h. In the case of the female she
showed two peaks of activity: one at sunrise and the other
1–2 h after sunset. The juvenile female also showed a peak of
activity at sunrise but not after sunset, whereas the male
showed a small peak of activity 1–2 h before sunrise but
again no peak after sunset.
Discussion
The results of this paper strongly challenge the currently
held theory that maned wolves live largely solitary lives
(Dietz, 1984). Here we show for the first time that an adult
pair associates spatially with each other before, during and
after a birth of pups (Figs 6–8). This spatial association has a
diurnal nature, with the pair spending an average of 82% of
their time during daylight hours within 50m of each other.
However, during the night, when they are hunting, the adult
pair seemed to be actively avoiding each other (see Figs 6–8).
Therefore, our data strongly suggest that maned wolves may
indeed be social animals, but we do not have direct beha-
vioural observations to confirm what type of social beha-
viour they are expressing. Certainly, the spatial relationships
between individuals and the female were stronger (as mea-
sured by the distance between individuals; Fig. 5) when the
female had pups, which strongly suggests parental care
behaviour being expressed by the male and potentially the
juvenile female as a helper. The male also showed a
protracted and strong spatial association with the female
before her pups were born. This may reflect mate guarding
by the male, as occurs in the red fox Vulpes vulpes, which
was once thought to be monogamous (Harris & White,
1994; Baker et al., 2004).
Home-range size
The home-range sizes reported in this study are within the
limits of home ranges reported by other authors for maned
wolves, which vary between 21.7 and 115 km2 (Dietz, 1984,
1985; Carvalho and Vasconcellos, 1995). For many species,
home-range size is obviously a function of environmental
quality; that is, in general we find a negative correlation
between home-range size and the level of food resources
within any given environment (Herfindal et al., 2005). One
interesting methodological issue we had in relation to
calculating home-range size was whether MCP 100 or MCP
95 was most appropriate because of our large sample sizes
per animal (more than 1500 fixes). Using MCP 95, we were
discarding between 75 and 80 fixes per animal, which is the
number of fixes frequently used to calculate home ranges in
conventional radio-telemetry studies (e.g. Dietz, 1984).
Therefore, we also calculated MCP 99, whereby only 15–16
of the most outlying points were not included in our
analyses.
Given the large quantity of data we had available, we also
calculated home ranges in 5-day blocks, which gave means
of between 7.5 and 28.5 km2 (Fig. 2). Obviously, here we are
using a limited number of data points (60), but this magni-
tude of sample size has been used by other researchers to
calculate home ranges (Dietz, 1984). These data, when
compared with the final estimates of home range (Table 1),
show the importance of collecting many fixes and over a
prolonged period of time, thereby allowing home-range data
to reach an asymptote. It is important to collect home-range
data over long periods of time, because this will avoid
problems of autocorrelations between fixes (Harris et al.,
1990).
A further interesting point arose in relation to the number
of fixes before reaching asymptotic home-range size. We
saw in Fig. 1 that the female’s home-range size became
asymptotic quickly (around 100 fixes). However, this
asymptote may be an artefact because the female appeared
to be searching for a nest site before giving birth and was
therefore increasing her home range. It is interesting to note
that both the male and the juvenile needed hundreds of fixes
before their home ranges stabilized. In the case of the
juvenile, this may reflect the increasing physiological
needs of a growing animal or her need to explore and
gather information about her environment. In the case of
the male, the lack of reaching an asymptotic home
range may be related to the breeding season or may simply
reflect the dynamic nature of environmental use by
adult maned wolves. These data certainly suggest that
other researchers should be extremely cautious when
calculating and interpreting home-range sizes when less
than several hundred location fixes are available, especially
if fixes were collected in a short period of time (Harris et al.,
1990).
Home range and birth
For the 40 days before the birth of her pups, the female in
our study was seen to be increasing and decreasing her home
range (Fig. 2). There are three possible explanations for this
pattern of home range: (1) the female was procuring more
food due to increasing energetic demands because of her
pregnancy, (2) the female was procuring a birth site (nest/
den) and her increase in home range reflects search effort,
and (3) the female was responding to decreased prey avail-
ability by increasing her home range. The data best fit
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London 33
The secret lives of maned wolvesL. F. Bandeira de Melo et al.

hypothesis (2), the female searching for an appropriate
birthing site, because if she was simply responding to
increasing energetic demands she could simply increase her
activity levels within her existing home range, assuming prey
density was not limiting. However, in Fig. 3 we can see that
the distanced travelled within a 24-h period did not increase
before the female gave birth.
Immediately following the birth of her pups, the female
dramatically reduced her home-range size and distance
travelled within a 24-h period. This was obviously in
response to her need to dedicate time for parental care of
her pups. Simultaneously, the male also reduced his home-
range size and activity levels, suggesting that the male was
also providing parental care. This reduction in home-range
size was not observed in the juvenile (see Fig. 2). These data
support observations of captive studies, which suggest that
the male plays an important role in the care of offspring
(Dietz, 1984; Rasmussen & Tilson, 1984; Veado, 1997).
Furthermore, before the birth of the pups on 17 June
2003, the juvenile female spent only 2 h at exactly the
same location as that of the female in 40 days, whereas
after the birth of the pups she spent 48 h at the same
location in the following 70 days. Interestingly, on the day
after the birth she spent 6 continuous hours with the female
and presumably, therefore, the pups. The exact function of
the juvenile female’s visit remains unknown, but we can
propose various hypotheses to be tested: from helping
behaviour to simple curiosity. Undoubtedly, the birth of
pups created the emission of new stimuli, such as scents in
the environment, which may have attracted the juvenile
female. The male at this point remained close to the birth
site, but our GPS collars did not detect him at the same
point as the birth site (but we need to point out that our
sampling was not continuous). During the weeks and
months after the birth of the three pups (as confirmed by
direct observations and camera traps), the juvenile female
visited her mother and presumably the pups on several
occasions (five times in the first month, four times in the
second month, etc.). The purpose of such visits remains
unclear, and in future studies we hope to place video
cameras at the nest site to confirm whether ‘helping’ occurs
in maned wolves.
The area chosen for the birth site was an open area of
cerrado covered in tall grass species (at least 1m high) with a
small tree beside the birth site. Visual inspection of the birth
site, after the female had left it with her pups, showed that it
comprised a maze of grass tunnels. In this study we encoun-
tered many sleeping sites for maned wolves, and their
structure was different from the birth site in that they did
not contain any tunnels within the local vegetation. We
suspect that the female used these tunnels to move the pups
around, probably as an anti-predator strategy. Our camera
traps photographed several potential predators of maned
wolf pups, such as pumas Puma concolor and feral dogs
Canis familiaris. A similar pattern of cub movement has
been observed in female cheetahs, which may be in response
to predators and the build-up of parasites in the den
(Laurenson, 1993).
Home-range overlap
The female had the largest home range of the individuals in
our study (see Table 1), and the data show that the male and
the juvenile lived within the female’s home range. However,
both the male and the juvenile did have very small areas
where the female did not visit. Therefore, we conclude that
the degree of tolerance between related individuals in the
wild may be high when environmental conditions permit
(i.e. good food abundance). Why the female has a home
range much larger than the male is not clear. This could
reflect a greater energetic demand or the simple fact that she
needs to procure certain sex-specific resources, such as a
nesting site. Another interesting observation is that our
camera traps photographed other males in the area sur-
rounding our study subjects’ home ranges. It could be that
the presence of other males nearby limited the home-range
size of the male in our study, as has been observed in white
rhinoceros Ceratotherium simum (Rachlow, Kie & Berger,
1999).
The fact that one (we only observed one juvenile female
within the female’s home range) and possibly more juveniles
stay within the home range of their mother is interesting,
and this has also been observed in other reserves (e.g. RPPN
Serra do Caraca; R. J. Young, pers. obs.). This suggests an
extended learning period in maned wolves, which is most
probably related to learning the spatial distribution of
resources, and not hunting skills, as in other carnivore
species, such as cheetahs Acinonyx jubatus (Caro, 1994).
We feel confident in saying this because maned wolves are
spatially separated at night when hunting (see Figs 6–8) and,
therefore, a hunting juvenile cannot learn from an adult.
Furthermore, in comparison with other carnivores the
maned wolf has a much larger area of its brain dedicated
to spatial memory (Dunbar & Bever, 1998), suggesting a
necessity to learn about its environment.
Spatial relationships between individuals
Figure 6 clearly shows that the adult pair normally sleep
together or in close proximity during daylight hours, and
then when active at night they strongly avoid one another.
Figures 7 and 8 show a similar pattern, with the juvenile
attempting to sleep close to her parents during the day and
apparently actively avoiding them at night. These data
suggest that social bonds between adult pairs may be
stronger than previously thought (see Dietz, 1984), espe-
cially because this pattern of association was seen before the
birth of the pups (Fig. 5). It could be argued that the maned
wolves slept together because of the scarce availability of
suitable sleeping sites. However, then we would expect all
pairwise comparisons between individuals to show the same
result, which they clearly did not. These data strongly
suggest that social behaviour is important to maned wolves,
and this is a topic that should be addressed in the field
through direct behavioural observations. The relationship
between the juvenile and her parents is intriguing and may
suggest that the juvenile had an important role in the
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London34
The secret lives of maned wolves L. F. Bandeira de Melo et al.

reproductive success of her parents, perhaps acting as a
helper.
The apparent active avoidance of other individuals at
night seems most probably a strategy that wild maned
wolves adopt to avoid one animal interfering with the
hunting behaviour of another individual. For example, after
a predator has recently moved through a specific area, many
prey species may have already sought cover, and therefore
would be more difficult to capture. The hunting method of
maned wolves may also encourage mutual avoidance; the
use of their large ears to pinpoint the sounds of rodent prey
in tall grass would soon be compromised by the presence of
others. In otters Lontra canadensis, individuals avoid hunt-
ing close together to increase individual capture rates and to
reduce intra-specific competition (Melquist & Hornocker,
1983).
For the 6weeks after the birth of the pups, both the male
and the juvenile reduced their mean distances to the female,
and increased their frequency of encounters with the female.
These data suggest that both individuals may have been
involved in the care of the pups. Studies in captivity have
demonstrated that the male will often bring food to a female
who has pups (Dietz, 1984; Rasmussen & Tilson, 1984;
Veado, 1997).
Daily range
The daily ranges of maned wolves varied considerably,
ranging from 0.07 to 16 km, and obviously the birth of pups
greatly affected the ranging of the female. On average, adult
and juvenile maned wolves ranged between 6 and
9 kmday�1. The ranging of both the female and the juvenile
generally increased during our study, especially towards the
end of the study. It should be noted that our study
terminated as the rainy season began in 2003. During the
rainy season, prey availability is reported to be significantly
lower than in the dry season (Alho et al., 1986), which
implies that animals need to expend greater hunting effort in
the wet season. This may explain why animals increased
their home ranges at the end of this study (see Fig. 2).
Activity during 24 h
From the data in this study, we show that, as found in other
studies (Dietz, 1984), maned wolves are strongly nocturnal
with peaks of activity at dawn and dusk (i.e. crepuscular).
The crepuscular activity peaks seemed to be more pro-
nounced in the adult female in comparison with the other
two individuals. However, in other areas of Brazil maned
wolves can be active during daylight hours (e.g. in Serra da
Canastra National Park, Minas Gerais).
Conclusions
The conclusions presented here are based on data pertaining
to three individuals. Therefore, some caution must be used
so as not to overgeneralize, because individual variation in
the variables described in this study could occur (see Lott,
1984). The results presented in this article have implications
for both in situ and ex situ conservation of this species. The
data presented here suggest strongly that maned wolves are
more social than previously thought and may, like other
canid species, have rich and complex social lives. However,
to reveal more details about the lives of wild maned wolves,
direct behavioural field observations are required.
Acknowledgements
We thank CEMIG (Companhia Energetica de Minas Ger-
ais) for their financial and logistical support, without which
this project would not have been possible. We are grateful to
the group of CEMIG from RPPN Galheiro Environmental
Station for their support. This article was greatly improved
by the comments of Dr Andrew Kitchener, Dr Ron Swais-
good and Prof. Sonia Talamoni. Televilt’s staff was most
helpful in answering our queries about the use of GPS
collars. This study was conducted in compliance with the
appropriate Brazilian laws in enforcement at the time of the
study, and we wish to thank IBAMA for their permission to
carry out this study.
References
Alho, C.J.R., Pereira, L.A. & Paula, A.C. (1986). Patterns of
habitat utilization by small mammal populations in cerra-
do biome of central Brazil. Mammalia 50, 447–460.
Baker, P.J., Funk, S., Bruford, M.W. & Harris, S. (2004).
Polygynandry in a red fox population: implications for the
evolution of group living in canids? Behav. Ecol. 15,
766–778.
Blake, S., Douglas-Hamilton, I. & Karesh, W.B. (2001). GPS
telemetry of forest elephants in Central Africa: results of a
preliminary study. Afr. J. Ecol. 39, 178–186.
Bueno, A.A., Belentani, S.C.S. & Motta-Junior, J.C. (2002).
Feeding ecology of the maned wolf, Chrysocyonbrachyurus
(Illiger, 1815) (Mammalia: Canidae), in the ecological
station of Itirapina, Sao Paulo state, Brazil. Biota Neotro-
pica 2, 1–9.
Caro, T.M. (1994). Cheetahs of the Serengeti plains. Chicago:
University of Chicago Press.
Carvalho, C.T. & Vasconcellos, L.E.M. (1995). Disease, food
and reproduction of the maned wolf Chrysocyon bra-
chyurus (Illiger) (Carnivora, Canidae) in southeast Brazil.
Rev. Brasil. Zool. 12, 627–640.
Crowell-Davis, S.L., Curtis, T.M. & Knowles, R.J. (2004).
Social organization in the cat: a modern understanding.
J. Feline Med. Surg. 6, 19–28.
Dietz, J.M. (1984). Ecology and social organization of the
maned wolf (Chrysocyon brachyurus). Smithsonian Contrib.
Zool. 392, 1–51.
Dietz, J.M. (1985). Chrysocyon brachyurus. Mammal. Species
234, 1–4.
Dunbar, R.I.M. & Bever, J. (1998). Neocortex size determines
group size in insectivores and carnivores. Ethology 104,
695–708.
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London 35
The secret lives of maned wolvesL. F. Bandeira de Melo et al.

Harris, S., Creswell, W.J., Forde, P.G., Trewhella, W.J.,
Woollard, T. & Wray, S. (1990). Home-range analysis
using radio-tracking data – a review of problems and
techniques particularly as applied to the study of mammals.
Mammal. Rev. 20, 95–123.
Harris, S. & White, P.C.L. (1994). The red fox. 2nd enlarged
edn. London: The Mammal Society.
Herfindal, I., Linnell, J.D.C., Odden, J., Birkeland Nilsen, E.
& Andersen, R. (2005). Prey density, environmental
productivity and home-range size in the Eurasian lynx
(Lynx lynx). J. Zool. (Lond.) 265, 63–71.
Laurenson, M.K. (1993). Early maternal-behavior of wild
cheetahs – implications for captive husbandry. Zoo Biol.
12, 31–43.
Lott, D.F. (1984). Intraspecific variation in the social-systems
of wild vertebrates. Behaviour 88, 266–325.
McCarthy, T.M., Todd, K., Fuller, T.K. & Munkhtsog, B.
(2005). Movements and activities of snow leopards in
southwestern Mongolia. Biol. Conserv. 124, 527–537.
Melquist, W.E. & Hornocker, M.G. (1983). Ecology of river
otters in west central Idaho.Wildl. Monogr. 83, 1–60.
Motta-Junior, J.C. (2000). Variacao temporal e selecao de
presas na dieta do lobo-guara, Chrysocyon brachyurus
(Mammalia: Canidae), na Estacao Ecologica de Jataı, Luiz
Antonio, SP. In Estudos integrados em ecossistemas:
331–346. Santos, J.E. & Pires, J.S.R. (Eds). Estacao
Ecologica de Jataı. v.1. Sao Carlos: Rima Editora.
Motta-Junior, J.C. & Martins, K. (2002). The frugivorous
diet of the maned wolf, Chrysocyon brachyurus, in Brazil:
ecology and conservation. In Seed dispersal and frugivory:
ecology, evolution and conservation: 291–303. Levey, D.J.,
Silva, W.R. & Galetti, M. (Eds). Wallingford, Oxfordshire:
CABI Publishing.
Motta-Junior, J.C., Talamoni, S.A., Lombardi, J.A. & Simo-
lomaki, K. (1996). Diet of the maned wolf, Chrysocyon
brachyurus, in central Brazil. J. Zool. (Lond.) 240,
277–284.
Rachlow, J.L., Kie, J.G. & Berger, J. (1999). Territoriality
and spatial patterns of white rhinoceros in Matobo
National Park, Zimbabwe. J. Afr. Ecol. 37, 295–304.
Rasmussen, J.L. & Tilson, R.L. (1984). Food provisioning by
adult maned wolves (Chrysocyon brachyurus). Z. Tierpsy-
chol. 65, 346–352.
Rutter, S.M., Beresford, N.A. & Roberts, G. (1997). Use of
GPS to identify the grazing areas of hill sheep. Comp.
Electron. Agri. 17, 177–188.
Saltz, D. (1994). Reporting error measurements in radio
location by triangulation: a review. J. Wildl. Mgmt. 58,
181–184.
Samuel, M.D. & Fuller, M.R. (1994). Wildlife telemetry. In
Research and management techniques for wildlife and habi-
tats: 370–418. Bookhout, T.A. (Ed.). Bethesda, MD: The
Wildlife Society.
Sillero-Zubiri, C. & Hoffmann, M. (2004). Chrysocyon bra-
chyurus. In IUCN 2004. 2004 IUCN Red List of threatened
species. http://www.redlist.org. Downloaded on 7 October
2005.
Silva, J.A. & Talamoni, S.A. (2004). Core area and centre of
activity of maned wolves, Chrysocyon brachyurus (Illiger)
(Mammalia, Canidae), submitted to supplemental feeding.
Rev. Brasil. Zool. 21, 391–395.
Theuerkauf, J. & Jedrzejewski, W. (2002). Accuracy of radio-
telemetry to estimate wolf activity and locations. J. Wildl.
Mgmt. 66, 859–864.
Veado, B.V. (1997). Parental behaviour in maned wolf Chry-
socyon brachyurus at Belo Horizonte Zoo. Int. Zoo Year-
book 35, 279–286.
Wing, M.G., Eklund, A. & Kellogg, L.D. (2005). Consumer-
grade global positioning system (GPS) accuracy and
reliability. J. Fores. 103, 169–173.
Journal of Zoology 271 (2007) 27–36 c� 2006 The Authors. Journal compilation c� 2006 The Zoological Society of London36
The secret lives of maned wolves L. F. Bandeira de Melo et al.
Copyright © 2022 FDOKUMEN










![Ezra Pound e il "canto" del 1815 napoleonico [Ezra Pound and the Napoleonic 1815 Canto]](https://static.fdokumen.com/doc/165x107/631afa4a80cc3e9440059dad/ezra-pound-e-il-canto-del-1815-napoleonico-ezra-pound-and-the-napoleonic-1815.jpg)









