
SAMPLE THESIS TITLE WITH A CONCISE AND ACCURATE ...
176
CONTRIBUTION OF PIEZO1 MECHANOSENSITIVE CHANNELS TO ASTROCYTE CALCIUM SIGNALING by Rebecca Wai Yin Ko B.Sc., The University of British Columbia, 2009 A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in THE FACULTY OF GRADUATE AND POSTDOCTORAL STUDIES (Neuroscience) THE UNIVERSITY OF BRITISH COLUMBIA (Vancouver) June 2018 © Rebecca Wai Yin Ko, 2018
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of SAMPLE THESIS TITLE WITH A CONCISE AND ACCURATE ...

CONTRIBUTION OF PIEZO1 MECHANOSENSITIVE CHANNELS TO ASTROCYTE
CALCIUM SIGNALING
by
Rebecca Wai Yin Ko
B.Sc., The University of British Columbia, 2009
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF
THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
in
THE FACULTY OF GRADUATE AND POSTDOCTORAL STUDIES
(Neuroscience)
THE UNIVERSITY OF BRITISH COLUMBIA
(Vancouver)
June 2018
© Rebecca Wai Yin Ko, 2018

ii
The following individuals certify that they have read, and recommend to the Faculty of Graduate
and Postdoctoral Studies for acceptance, the dissertation entitled:
Contribution of Piezo1 mechanosensitive channels to astrocyte calcium signaling i
submitted by Rebecca Wai Yin Ko in partial fulfillment of the requirements for
the degree of Doctor of Philosophy I
in Neuroscience i
Examining Committee:
Brian MacVicar, Neuroscience I
Supervisor
Terrance Snutch, Neuroscience I
Supervisory Committee Member
Eric Accili, Neuroscience I
University Examiner
Vanessa Auld, Neuroscience I
University Examiner
Additional Supervisory Committee Members:
Ann Marie Craig, Neuroscience I
Supervisory Committee Member
Yu Tian Wang, Neuroscience I
Supervisory Committee Member

iii
Abstract
Astrocyte calcium (Ca2+) signaling is involved in the regulation of physiological processes such
as synaptic activity and vascular tone in the brain. Recent developments in tools to monitor Ca2+
signals have revealed a novel type of spontaneous Ca2+ transient that is localized to
microdomains in the fine astrocytic processes. However, the molecular mechanisms underlying
these signals have not been fully characterized. Based on data from rodent brain transcriptomic
and proteomic studies, we identified Piezo1, a mechanosensitive cation channel, as a potential
candidate for mediating these Ca2+ transients.
In Chapter 2, we performed two-photon imaging of the membrane-tethered, genetically encoded
Ca2+ indicator Lck-GCaMP5 in cultured astrocytes and developed an algorithm for extracting
and analyzing the microdomain Ca2+ signals. Using a combination of pharmacological and
siRNA approaches, we showed that Piezo1 channels contribute to these spontaneous Ca2+
transients. We also conducted preliminary imaging experiments in brain slice astrocytes and
found that spontaneous Ca2+ signals in the endfoot compartments were sensitive to
pharmacological modulators of Piezo1. In Chapter 3, we performed immunostaining using
strategies that were optimized to target subcellular locations where Piezo1 expression had
previously been reported in other cell types. Our results indicated that Piezo1 is localized to
subcellular compartments relevant to mechanosensation; Piezo1 immunoreactivity was localized
to discrete clusters on the plasma membrane and associated with focal adhesion and actin stress
fibers in cultured astrocytes, and Piezo1 expression was observed within the endfoot processes of

iv
brain slice astrocytes. Lastly, in Chapter 4, we showed that an osmotic stress model of astrocyte
swelling could activate Piezo1-mediated Ca2+ microdomain signals in cultured astrocytes.
Taken together, the data provide evidence that Piezo1 contributes to spontaneous Ca2+
microdomain signals in astrocytes in both cell culture and acute brain slices, and suggest that
astrocyte Ca2+ signaling may play a role in integrating mechanical stimuli to regulate brain
function in physiological and pathological processes involving changes in mechanical force.

v
Lay Summary
Astrocytes are brain cells that display intracellular calcium signals, and these signals have been
shown to regulate physiological processes such as neuronal activity and cerebral blood flow.
Recent advances in imaging tools revealed a novel type of spontaneous calcium signals within
astrocytes, but the underlying mechanisms have not been fully characterized. Here we present
evidence that Piezo1, a channel sensitive to mechanical forces, is involved in mediating these
signals. We performed imaging experiments and showed that the signals can be modulated by
strategies that activate or block Piezo1. We conducted staining experiments and demonstrated
that Piezo1 is located in astrocyte regions associated with force sensing. Furthermore, we found
that cell swelling could activate Piezo1 in astrocytes. Taken together, the data suggest that these
calcium signals may represent astrocyte integration of mechanical stimuli, raising interesting
possibilities for the role of astrocytes in physiological and pathological processes involving
changes in mechanical force.

vi
Preface
The experiments in this dissertation were jointly designed with Dr. Brian MacVicar. I conducted
all the experiments and data analyses, with the following exceptions:
In Chapter 2, Alexa Nelson assisted with preparation of the Lck-GCaMP5 plasmid. Dr. Leigh
Wicki-Stordeur conducted the immunoblotting experiments. Dr. John Tyson from Dr. Terry
Snutch’s lab designed the luciferase and Piezo1 siRNA sequences, and performed the qPCR
analyses. Joslyn Quick from Dr. Pieter Cullis’ lab prepared the lipid nanoparticle plasmid DNA
and siRNA encapsulations. Jeffrey Ledue contributed greatly to the Matlab algorithms used for
data analysis. Dr. Xiling Zhou from Dr. Ann Marie Craig’s lab provided the astrocyte cultures.
In Chapter 3, Dr. Esperanza Garcia cultured the Neuro2A cells and assisted with designing the
immunostaining experiments. Karen Jones performed immunoblotting and Neuro2A cell
staining experiments. Dr. John Tyson conducted the qPCR analyses. All collaborators were
from Dr. Terry Snutch’s lab.
All animal procedures performed for this dissertation were approved by the University of British
Columbia Animal Care Committee under certificates A15-0209 and A15-0086.

vii
Table of Contents
Abstract ......................................................................................................................................... iii
Lay Summary .................................................................................................................................v
Preface ........................................................................................................................................... vi
Table of Contents ........................................................................................................................ vii
List of Tables ............................................................................................................................... xii
List of Figures ............................................................................................................................. xiii
List of Abbreviations ...................................................................................................................xv
Acknowledgements .................................................................................................................. xviii
Chapter 1: Introduction ................................................................................................................1
1.1 Astrocyte Ca2+ signaling ................................................................................................. 1
1.1.1 Evolution of approaches to study astrocyte Ca2+ signaling ........................................ 2
1.1.2 Spontaneous Ca2+ microdomain signals in astrocytes ................................................ 6
1.2 Astrocytes as mechanosensitive cells ........................................................................... 10
1.2.1 Astrocytes detect mechanical stimuli to regulate cell physiology ............................ 10
1.2.2 Membrane proteins linked to mechanosensitivity in astrocytes ............................... 12
1.3 Piezo1: a mechanosensitive cation channel .................................................................. 16
1.3.1 Piezo1 channel topology and the structural basis of function................................... 18
1.3.2 Properties of the Piezo1 channel ............................................................................... 20
1.3.3 Modulation of Piezo1 channel activity by second messengers ................................. 23
1.3.4 Piezo1 pharmacology ................................................................................................ 24
1.3.5 Piezo1 expression and physiological functions ........................................................ 26

viii
1.3.6 Pathologies associated with Piezo1 dysfunction....................................................... 27
1.4 Rationale and hypotheses .............................................................................................. 29
1.4.1 Objective 1: To determine the contribution of Piezo1 to spontaneous Ca2+ transients
in astrocytes .......................................................................................................................... 30
1.4.2 Objective 2: To characterize the subcellular localization of Piezo1 in astrocytes .... 30
1.4.3 Objective 3: To examine the activation of Piezo1 in hypotonicity-induced astrocyte
swelling ................................................................................................................................. 31
Chapter 2: Piezo1 mediates astrocyte Ca2+ microdomain signals ...........................................32
2.1 Overview ....................................................................................................................... 32
2.2 Methods......................................................................................................................... 34
2.2.1 Chemicals and reagents............................................................................................. 34
2.2.2 Astrocyte cultures ..................................................................................................... 35
2.2.3 siRNA sequences for Piezo1 knockdown ................................................................. 36
2.2.4 Lck-GCaMP5 plasmid preparation ........................................................................... 36
2.2.5 Preparation of siRNA- and plasmid DNA-containing lipid nanoparticles ............... 37
2.2.6 Astrocyte transfection with LNPs ............................................................................. 38
2.2.7 Evaluation of transfection viability and efficiency ................................................... 38
2.2.8 Evaluation of Piezo1 siRNA effect on mRNA levels ............................................... 39
2.2.9 Evaluation of Piezo1 siRNA effect on protein levels ............................................... 39
2.2.10 Culture imaging .................................................................................................... 40
2.2.11 Transgenic GLAST-GCaMP5 mice and tamoxifen treatment.............................. 41
2.2.12 Hippocampal slice preparation ............................................................................. 41
2.2.13 Slice imaging ........................................................................................................ 42

ix
2.2.14 Image processing and data analysis ...................................................................... 42
2.2.15 Statistics ................................................................................................................ 43
2.3 Results ........................................................................................................................... 44
2.3.1 LNP delivery of plasmid DNA and siRNA in cultured astrocytes ........................... 44
2.3.2 Development of a novel method to analyze astrocyte Ca2+ signals .......................... 47
2.3.3 Piezo1 activation selectively increases the frequency of microdomain Ca2+ signals 49
2.3.4 Yoda1-induced Ca2+ microdomain signals are sensitive to pharmacological and
genetic Piezo1 inhibition....................................................................................................... 50
2.3.5 Yoda1-induced Ca2+ microdomain signals depend on extracellular Ca2+ entry ....... 54
2.3.6 Spontaneous Ca2+ microdomain signals are sensitive to Piezo1 antagonists ........... 55
2.3.7 Preliminary imaging of brain slice astrocyte Ca2+ signals show sensitivity to
pharmacological Piezo1 modulations ................................................................................... 57
2.4 Discussion ..................................................................................................................... 59
Chapter 3: Subcellular localization of Piezo1 in astrocytes .....................................................65
3.1 Overview ....................................................................................................................... 65
3.2 Methods......................................................................................................................... 66
3.2.1 Cell culture ................................................................................................................ 66
3.2.2 Astrocyte transfection and treatment ........................................................................ 67
3.2.3 qPCR for Neuro2A cells ........................................................................................... 68
3.2.4 Immunoblotting......................................................................................................... 68
3.2.5 Immunocytochemistry .............................................................................................. 69
3.2.6 Immunohistochemistry ............................................................................................. 71
3.2.7 Confocal image acquisition and analysis .................................................................. 72

x
3.3 Results ........................................................................................................................... 73
3.3.1 Antibody testing in Neuro2A cells that endogenously express Piezo1 .................... 73
3.3.2 Piezo1 immunoreactivity clusters in discrete domains in the astrocyte plasma
membrane .............................................................................................................................. 74
3.3.3 Piezo1 immunoreactivity is present in actin filaments and focal adhesions ............. 79
3.3.4 Forskolin-induced disorganization of the cytoskeleton alters Piezo1 distribution ... 82
3.3.5 Piezo1 immunoreactivity colocalizes with astrocyte markers in the mouse brain ... 84
3.4 Discussion ..................................................................................................................... 86
Chapter 4: Role of Piezo1 in astrocyte swelling ........................................................................93
4.1 Overview ....................................................................................................................... 93
4.2 Methods......................................................................................................................... 96
4.2.1 Cell culture and transfection ..................................................................................... 96
4.2.2 Culture imaging ........................................................................................................ 96
4.2.3 Data analysis and statistics........................................................................................ 97
4.3 Results ........................................................................................................................... 97
4.3.1 Piezo1 mediates Ca2+ microdomain signals in astrocytes under hypotonic stress .... 97
4.3.2 Forskolin-induced reorganization of the cytoskeleton does not alter the
pharmacological profile of Piezo1 or affect Piezo1-mediated Ca2+ transients to hypotonic
challenge ............................................................................................................................... 99
4.3.3 Piezo1 mediates hypotonicity-induced swelling in astrocytes ................................ 103
4.4 Discussion ................................................................................................................... 104
Chapter 5: Conclusion ...............................................................................................................109
5.1 Summary of research findings .................................................................................... 109

xi
5.2 Research significance.................................................................................................. 109
5.3 Future directions ......................................................................................................... 113
5.3.1 Transgenic mice for investigating Piezo1-mediated Ca2+ signaling in vivo ........... 114
5.3.2 Role of Piezo1 in regulation of cerebral vasculature tone ...................................... 115
5.3.3 Role of Piezo1 in astrocyte differentiation ............................................................. 116
5.3.4 Role of Piezo1 in transmitter release ...................................................................... 117
5.3.5 Piezo1 and pathology .............................................................................................. 118
References ...................................................................................................................................120

xii
List of Tables
Table 2-1. LNP properties. ........................................................................................................... 38

xiii
List of Figures
Figure 1-1. Genetically-encoded Ca2+ indicators (GECIs) reveal distal astrocyte processes that
are not detected by bulk-loaded Ca2+ indicator dyes. ..................................................................... 5
Figure 1-2. Structure of mouse Piezo1. ....................................................................................... 18
Figure 2-1. Ca2+ signal analysis algorithm flowchart. ................................................................. 43
Figure 2-2. Lipid nanoparticle (LNP) delivery of Lck-GCaMP5 plasmid DNA and Piezo1
siRNA is non-toxic and effective in cultured astrocytes. ............................................................. 46
Figure 2-3. The Ca2+ signal analysis algorithm isolates Ca2+ microdomain events by masking
Ca2+ waves. ................................................................................................................................... 48
Figure 2-4. The Piezo1 agonist, Yoda1, selectively increases the number of Ca2+ microdomain
signals. .......................................................................................................................................... 50
Figure 2-5. Yoda1-induced increases in Ca2+ microdomain signals are partially inhibited by
Piezo1 antagonists GsMTx4 and Gd3+. ......................................................................................... 52
Figure 2-6. Yoda1-induced increases in Ca2+ microdomain signals are inhibited by Piezo1
siRNA. .......................................................................................................................................... 53
Figure 2-7. Yoda1-induced increases in Ca2+ microdomain signals depend on extracellular Ca2+
entry. ............................................................................................................................................. 55
Figure 2-8. Spontaneous Ca2+ microdomain signals are inhibited by Piezo1 antagonists GsMTx4
and Gd3+. ....................................................................................................................................... 57
Figure 2-9. The frequency of astrocyte endfeet Ca2+ transients are sensitive to pharmacological
modulators of Piezo1. ................................................................................................................... 58

xiv
Figure 3-1. Flowchart of immunocytochemistry protocols for optimization of Piezo1 staining in
different subcellular regions. ........................................................................................................ 70
Figure 3-2. Schematic illustrating the epitopes targeted by the Alomone and Proteintech Piezo1
antibodies. ..................................................................................................................................... 71
Figure 3-3. Neuro2A cells show distinct clusters of Piezo1 immunoreactivity. ......................... 74
Figure 3-4. Astrocytes exhibit different patterns of Piezo1 immunoreactivity at the plasma
membrane. ..................................................................................................................................... 75
Figure 3-5. Piezo1 immunoreactivity is distributed in clusters located on astrocyte plasma
membranes. ................................................................................................................................... 77
Figure 3-6. Piezo1 siRNA reduces the number of Piezo1 clusters in astrocytes. ........................ 79
Figure 3-7. Piezo1 immunoreactivity is found in actin filaments and focal adhesions of the
cytoskeleton. ................................................................................................................................. 81
Figure 3-8. Forskolin induces astrocyte stellation and Piezo1 redistribution. ............................. 84
Figure 3-9. Piezo1 immunoreactivity co-localizes with astrocyte markers in brain slices. ......... 86
Figure 4-1. Hypotonic solution induces an increase in astrocyte microdomain Ca2+ signals that is
inhibited by Piezo1 siRNA treatment. .......................................................................................... 99
Figure 4-2. Forskolin treatment does not affect the pharmacological profile of Piezo1. .......... 101
Figure 4-3. Hypotonic solution induces an increase in forskolin-treated astrocyte microdomain
Ca2+ signals that is inhibited by Piezo1 siRNA treatment. ......................................................... 102
Figure 4-4. Piezo1 siRNA treatment inhibits hypotonicity-induced swelling in forskolin-treated
astrocytes..................................................................................................................................... 104

xv
List of Abbreviations
AQP4 Aquaporin-4
ASIC Acid-sensing ion channel
ATP Adenosine triphosphate
BAPTA 1,2-bis(o-aminophenoxy)ethane-N,N,N,N-tetraacetic acid
Ca2+ Calcium
cAMP Cyclic adenosine monophosphate
CED C-terminal extracellular loop domain
CHO-K1 Chinese hamster ovary K1 cell line
CNS Central nervous system
CTD C-terminal domain
DEG/ENaC Degenerin/epithelial Na+ channel
DiIC18 1,1’-dioctadecyl-3,3,3’,3’-tetramethylindocarbocyanine
perchlorate
DLin-KC2-DMA 2,2-dilinoleyl-4-(2-dimethylaminoethyl)-[1,3]-dioxolane
DLin-MC3-DMA heptatriaconta-6,9,28,31-tetraen-19-yl 4-(dimethylamino)butanoate
DSPC 1,2-distearoyl-sn-glycero-3-phosphocholine
ECM Extracellular matrix
ECS Extracellular space
EGTA Ethylene glycol tetraacetic acid
ER Endoplasmic reticulum
GABA Gamma-aminobutyric acid

xvi
Gd3+ Gadolinium
GECI Genetically-encoded Ca2+ indicator
GFAP Glial fibrillary acidic protein
GFP Green fluorescent protein
GPCR G-protein-coupled receptor
GsMTx4 Grammostola spatulata mechanotoxin 4
HEK293 Human embryonic kidney 293 cell line
HPLC High-performance liquid chromatography
IP3 Inositol-1,4,5-trisphosphate
IP3R2 IP3 type 2 receptor
K2P Two-pore domain K+ channel
La3+ Lanthanum
LNP Lipid nanoparticle
mPTP Mitochondrial permeability transition pore
mRNA Messenger RNA
NMDAR N-methyl-D-aspartate receptor
PBS Phosphate buffered saline
PEG-DMG Polyethylene glycol (2K)-dimyristolglycerol
PI(4,5)P2 Phosphatidylinositol-4,5-bisphosphate
qPCR Quantitative polymerase chain reaction
ROI Region of interest
ROS Reactive oxygen species
RVD Regulatory volume decrease

xvii
SERCA Sarco/endoplasmic reticulum Ca2+ ATPase
siRNA Small interfering ribonucleic acid
SOPC 1-stearoyl-2-oleoyl-sn-glycero-3-phosphocholine
STOML3 Stomatin-like protein 3
TRP Transient receptor potential
VRAC Volume-regulated anion channel

xviii
Acknowledgements
I would like to thank my supervisor, Dr. Brian MacVicar, for his mentorship and guidance. His
scientific insight and passion for research have been truly inspiring. Thank you for providing me
with opportunities to attend courses, participate in collaborations, and present my work in both
national and international settings, which have greatly benefitted my growth as a scientist.
I would also like to thank my committee members Dr. Ann Marie Craig, Dr. Terry Snutch, and
Dr. Yu Tian Wang for sharing their expertise and providing invaluable feedback for my projects
throughout the years.
My PhD training was financially supported by the University of British Columbia Four-Year
Fellowship and the Canadian Institutes of Health Research Master’s and Doctoral Research
Awards.
To all past and present members of the MacVicar lab, I am fortunate to have crossed paths with
you. Thank you for the scientific collaborations, troubleshooting advice, and moral support,
especially during the inevitable failures that are a part of scientific research.
Lastly, thank you to my family and friends for your unconditional support and encouragement.
This thesis would not be possible without you.

1
Chapter 1: Introduction
1.1 Astrocyte Ca2+ signaling
Astrocytes are a type of glial cell found throughout the vertebrate central nervous system,
accounting for approximately 20 – 40% of the cells in the brain (Herculano-Houzel, 2014). They
were first described by Rudolf Virchow in 1846 as a connective substance, or ‘glue’, that acts as
a scaffold to hold neurons together (García-Marín et al., 2007). Later developments in staining
methods allowed the star-like morphology of these cells to be visualized, leading to the
introduction of the term ‘astrocyte’ (García-Marín et al., 2007). However, researchers lacked
techniques to study dynamic processes in living astrocytes; in contrast to neuronal activity,
which could be probed with electrophysiology, astrocytes were electrically unexcitable.
Therefore, it was assumed that astrocytes were uniformly passive cells that merely provided
structural and trophic support for neurons (Navarrete and Araque, 2014).
Despite the lack of sophisticated technology, early researchers studied static images of astrocyte
morphology and made important observations revealing the diversity of these cells, as well as
predictions about their relevance in brain function. For example, Andriezen (1893) found that
astrocytes could be classified based on different process morphologies and distribution in the
brain. Furthermore, by refining staining methods, Ramón y Cajal was able to observe the highly
complex organization of fine astrocyte processes; based on their close association with synapses
and blood vessels, he proposed that astrocytes may release substances that affect neuronal
activity and mediate vasoconstriction or dilation (García-Marín et al., 2007; Navarrete and
Araque, 2014). Advances in imaging and genetic tools have since demonstrated that astrocytes

2
are molecularly and functionally heterogeneous (Khakh and Sofroniew, 2015; Haim and
Rowitch, 2017), and provided evidence supporting the role of astrocytes in regulation of synaptic
transmission and blood flow (Araque and Navarrete, 2010; Attwell et al., 2010).
Calcium (Ca2+) is a ubiquitous second messenger that regulates multiple cellular functions. A
key development in the field of astrocyte biology was the introduction of fluorescence
microscopy and Ca2+-sensitive fluorescent dyes, which allowed researchers to monitor changes
in cytosolic Ca2+ in response to synaptic activity and sensory stimuli (Verkhratsky et al., 2012;
Volterra et al., 2014). These observations suggested that astrocytes exhibit a form of excitability
based upon variations in intracellular Ca2+ concentrations rather than through action potential
propagation, and indicated that astrocytes may also be involved in information processing in the
brain (Volterra et al., 2014). The role of astrocytes is now established in a number of processes
including cellular homeostasis, synapse formation and pruning, synaptic modulation, and
neurovascular coupling (Shigetomi et al., 2016). New insights in the field of astrocyte research
are intimately linked to the improving tools for monitoring Ca2+ signals, and recent
developments in imaging techniques, such as genetically encoded Ca2+ indicators (GECIs), have
revealed further complexities in astrocyte Ca2+ signaling and the functions they govern.
1.1.1 Evolution of approaches to study astrocyte Ca2+ signaling
The discovery of Ca2+ signaling as the mode of astrocyte excitability changed the perception of
astrocytes from passive support cell to potential partner in brain information processing. This
important advance was possible due to tools such as bulk-loaded Ca2+ indicator dyes that
preferentially loaded astrocytes in brain slices, allowing astrocyte Ca2+ signals to be monitored

3
(Porter and McCarthy, 1995), and caged Ca2+ compounds that when activated selectively elevate
Ca2+ levels in astrocyte somata. By using these strategies, pioneering studies in the early 1990s
demonstrated that astrocytes in culture and brain slices respond to exogenous or synaptically
released glutamate with oscillatory increases in somatic Ca2+ concentrations and propagating
cytoplasmic Ca2+ waves (Cornell-Bell et al., 1990; Dani et al., 1992; Porter and McCarthy,
1996). In addition to glutamate, other neurotransmitters that act on Gq-linked G protein-coupled
receptors, including norepinephrine, acetylcholine, and ATP, can also evoke astrocyte Ca2+
signals through inositol-1,4,5-trisphosphate (IP3) – dependent release from endoplasmic
reticulum stores, suggesting that astrocytes can respond to a broad range of neuronal activity
(Perea et al., 2009). The use of techniques to selectively monitor and manipulate astrocyte
cytosolic Ca2+ levels led to findings implicating astrocyte Ca2+ signaling in physiological
processes such as modulation of synaptic strength and cerebral blood flow (Bazargani and
Attwell, 2016).
The physiological relevance of astrocyte Ca2+ signaling has been challenged in subsequent
studies examining the mechanisms of Ca2+ elevation in astrocytes, and the functional
consequences of inhibiting such elevations. For instance, Sun et al. (2013) showed that the Gq-
linked metabotropic glutamate receptor, mGluR5, is both developmentally downregulated and
that mGluR5 agonists failed to increase intracellular Ca2+ levels in adult mouse astrocytes. The
authors therefore speculated that astrocytes may no longer respond to synaptic glutamate release
beyond the juvenile developmental stage in rodents. Furthermore, genetic deletion of the IP3
type 2 receptor (IP3R2), which is known to be highly expressed in astrocytes, abolished astrocyte
Ca2+ elevations to Gq-linked GPCR agonists, but the transgenic mice still exhibited excitatory

4
synaptic activity and neurovascular coupling, leading to the proposal that astrocyte intracellular
Ca2+ changes may be functionally irrelevant (Petravicz et al., 2008; Nizar et al., 2013; Bonder
and McCarthy, 2014).
A critical insight in addressing these discrepancies was the realization that bulk-loaded Ca2+
indicators were restricted to the astrocyte soma and proximal processes, leading to severe
undersampling of ~90% of the astrocyte volume that is comprised of fine processes (Reeves et
al., 2011). Recent developments utilizing GECI proteins or dialysis of patch-loaded Ca2+ dyes
allowing for Ca2+ fluctuations to be monitored in the fine processes (Fig. 1-1) have revealed a
previously unappreciated type of astrocyte Ca2+ signaling differing from soma signals in their
spatial and temporal characteristics (Shigetomi et al., 2013a; Rungta et al., 2016). For example,
Srinivasan et al. (2015) showed that while soma Ca2+ signals were greatly reduced in IP3R2
knockout mice, a substantial proportion of microdomain transients persisted in the fine
processes. Removal of extracellular Ca2+ did not affect soma Ca2+ transients, but reduced the
frequency of fine process signals. Furthermore, Gq-linked GCPR stimulation evoked Ca2+
elevations in the processes but not in the soma. Taken together, these data indicate that
astrocytes exhibit a diverse range of Ca2+ signals that vary in their subcellular
compartmentalization, induction mechanism, and Ca2+ source (Khakh and Sofroniew, 2015).
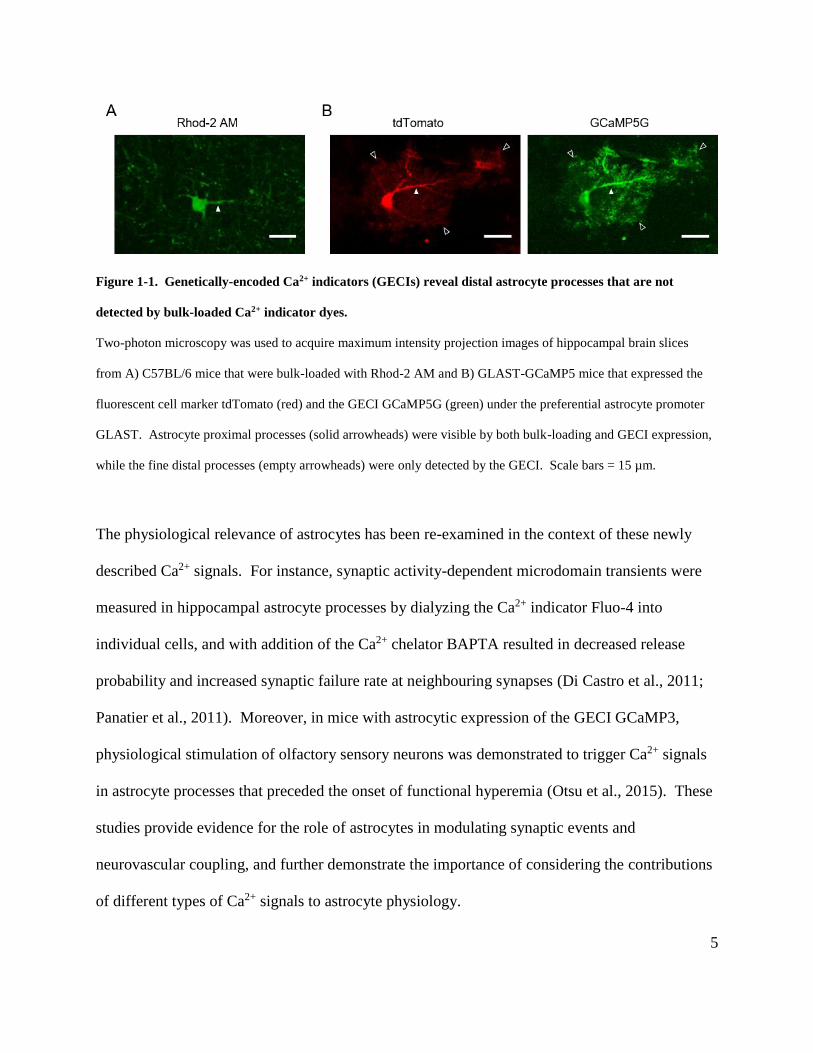
5
Figure 1-1. Genetically-encoded Ca2+ indicators (GECIs) reveal distal astrocyte processes that are not
detected by bulk-loaded Ca2+ indicator dyes.
Two-photon microscopy was used to acquire maximum intensity projection images of hippocampal brain slices
from A) C57BL/6 mice that were bulk-loaded with Rhod-2 AM and B) GLAST-GCaMP5 mice that expressed the
fluorescent cell marker tdTomato (red) and the GECI GCaMP5G (green) under the preferential astrocyte promoter
GLAST. Astrocyte proximal processes (solid arrowheads) were visible by both bulk-loading and GECI expression,
while the fine distal processes (empty arrowheads) were only detected by the GECI. Scale bars = 15 µm.
The physiological relevance of astrocytes has been re-examined in the context of these newly
described Ca2+ signals. For instance, synaptic activity-dependent microdomain transients were
measured in hippocampal astrocyte processes by dialyzing the Ca2+ indicator Fluo-4 into
individual cells, and with addition of the Ca2+ chelator BAPTA resulted in decreased release
probability and increased synaptic failure rate at neighbouring synapses (Di Castro et al., 2011;
Panatier et al., 2011). Moreover, in mice with astrocytic expression of the GECI GCaMP3,
physiological stimulation of olfactory sensory neurons was demonstrated to trigger Ca2+ signals
in astrocyte processes that preceded the onset of functional hyperemia (Otsu et al., 2015). These
studies provide evidence for the role of astrocytes in modulating synaptic events and
neurovascular coupling, and further demonstrate the importance of considering the contributions
of different types of Ca2+ signals to astrocyte physiology.

6
1.1.2 Spontaneous Ca2+ microdomain signals in astrocytes
The use of GECIs and patch-loaded Ca2+ indicator dyes revealed a diverse range of astrocyte
Ca2+ signals in different subcellular compartments; up to seven distinct kinds of signals have
been described (Khakh and Sofroniew, 2015). Previous studies have shown that neuronal
activity-associated astrocyte Ca2+ signals can be observed with GECIs or bulk- and patch-loaded
Ca2+ dyes (Perea et al., 2009; Khakh and Sofroniew, 2015). Intriguingly, the ability to monitor
Ca2+ in the distal astrocyte processes with the new approaches has revealed a spontaneous Ca2+
transient that is localized to the fine processes (Shigetomi et al., 2010). However, the
mechanisms underlying these spontaneous Ca2+ signals are incompletely characterized.
Spontaneous Ca2+ transients were first described by Nett et al. (2002) in juvenile mouse brain
slices wherein individual astrocytes were dialyzed with the Ca2+ dye Oregon Green BAPTA-1.
The authors imaged both proximal and distal processes, and found that astrocyte Ca2+
fluctuations persisted in the presence of the Na+ channel blocker tetrodotoxin and the vesicular
H+-ATPase inhibitor bafilomycin A1, suggesting that the signals are independent of neuronal
action potentials and spontaneous vesicular glutamate release. The Ca2+ transients in astrocyte
processes occurred at a higher frequency compared to soma signals. Furthermore, the process
signals were localized as microdomains, and the signals occurred asynchronously as if each
microdomain was functionally independent. Similar observations have since been described in
astrocytes from culture, brain slices, and in vivo, using both dye dialysis and GECI-based
approaches (Wang et al., 2006; Di Castro et al., 2011; Panatier et al., 2011; Shigetomi et al.,
2012, 2013a; Rungta et al., 2016).

7
The mechanisms mediating the spontaneous Ca2+ transients are under investigation with three
components thus far proposed as the Ca2+ source: release from the endoplasmic reticulum,
release from mitochondria, and influx across the plasma membrane. Pharmacological and
genetic tools have indicated that multiple pathways are likely to be involved. For example, the
microdomain signals are inhibited by cyclopiazonic acid and thapsigargin, agents that deplete
intracellular endoplasmic reticulum Ca2+ stores (Nett et al., 2002; Agarwal et al., 2017).
However, the observation that these Ca2+ signals are reduced, but not abolished, in IP3R2 KO
mice, suggests contributions from other sources. Agarwal et al. (2017) suggested that the
spontaneous signals may be mediated by mitochondria, as signal frequency and amplitude were
decreased by blockade of the mitochondrial permeability transition pore with cyclosporine A and
rotenone. Another possibility is Ca2+ entry through plasma membrane channels or transporters,
since the spontaneous transients are blocked by removal of Ca2+ from the extracellular solution
(Shigetomi et al., 2012; Rungta et al., 2016). Shigetomi et al. (2012) has proposed the transient
receptor potential channel TRPA1 as a candidate for mediating this transmembrane Ca2+ influx,
based on the observations that treatment with TRPA1 antagonists or siRNA reduced the
microdomain signals in cultured astrocytes, while TRPA1 agonists or overexpression in a
heterologous system increased the transients. However, subsequent experiments in hippocampal
brain slices showed that spontaneous Ca2+ transients persisted in the presence of TRPA1
blockers, as well as in TRPA1 knockout mice (Shigetomi et al., 2013b; Rungta et al., 2016;
Agarwal et al., 2017). Inhibition of other plasma membrane proteins implicated in Ca2+ flux in
astrocytes, such as voltage-gated Ca2+ channels, Na+-Ca2+ exchangers, ryanodine receptors, and
Ca2+ release-activated channels have also failed to block the Ca2+ transients (Rungta et al., 2016;

8
Agarwal et al., 2017). Therefore, the exact Ca2+ entry pathways contributing to the spontaneous
Ca2+ microdomain signals remain to be determined.
Several studies have examined the physiological significance of these ubiquitous spontaneous
Ca2+ signals. Experiments conducted by Agarwal et al. (2017) demonstrated that microdomain
signaling is enhanced by elevated reactive oxygen species (ROS) levels through Ca2+ efflux from
mitochondria stores. Moreover, since ROS are produced during the process of oxidative
phosphorylation (Mailloux and Harper, 2012), the authors speculated that the spontaneous
signals may reflect the metabolic state of the cell. Interestingly, increased spontaneous Ca2+
transients have also been observed in astrocytes from mouse models of Alzheimer’s disease
(Kuchibhotla et al., 2009; Delekate et al., 2014). Together with the observation that Ca2+ can
increase ATP production by acting on enzymes involved in glycolysis and oxidative
phosphorylation (Ververken et al., 1982; McCormack et al., 1990), it is possible that Ca2+ signals
facilitate coupling of ATP synthesis to disease states wherein energy metabolism is likely
compromised.
Spontaneous Ca2+ signals have also been implicated in the control of basal Ca2+ levels in
astrocytes, which may be important in regulation of synaptic activity and resting vessel tone. For
instance, as measured by ratiometric Fura-2 imaging, Shigetomi et al. (2012) demonstrated that
pharmacological inhibition of TRPA1-mediated Ca2+ transients was associated with a reduction
in basal Ca2+ concentrations from 120 nM to 50 nM. This result is supported by the observations
from Rungta et al. (2016) that removal of extracellular Ca2+ abolished Ca2+ transients in Fluo-4-
dialyzed astrocytes, and resulted in a ~40% decrease in baseline fluorescence. Reduction of

9
astrocyte intracellular free Ca2+ concentration by TRPA1 block or by dialysis of the Ca2+
chelator BAPTA was found to reduce the amplitude of interneuron mini inhibitory postsynaptic
currents. The authors demonstrated that decreased basal Ca2+ was associated with decreased
expression of the GABA transporter GAT-3 in astrocytes, leading to increased extracellular
levels of the neurotransmitter GABA and subsequent GABAA receptor desensitization
(Shigetomi et al., 2012). Pharmacological inhibition or genetic deletion of TRPA1 also
suppressed LTP in hippocampal CA1 pyramidal neurons, which was rescued by exogenous
application of the NMDA receptor (NMDAR) coagonist D-serine (Shigetomi et al., 2013b).
Similar results were obtained when astrocyte intracellular free Ca2+ was reduced to 50 – 80 nM
with the chelator EGTA (Henneberger et al., 2010), suggesting that a certain level of basal Ca2+
must be maintained for constitutive D-serine release and NMDAR-dependent plasticity. Finally,
Rosenegger et al. (2015) showed that introducing BAPTA into astrocytes with a patch pipette
resulted in vasoconstriction of adjacent arterioles. This effect was occluded by cyclooxygenase
inhibition, indicating that astrocytes may constitutively release prostaglandins in a Ca2+-
dependent manner to maintain constant vasodilation. Taken together, these examples
demonstrate that spontaneous Ca2+ transients in astrocytes may play important roles in the
modulation of metabolism, synaptic activity, and vascular tone in the brain.
In conclusion, continual improvements in the techniques employed to monitor Ca2+ levels have
demonstrated that astrocytes exhibit a complex range of Ca2+ signals within distinct subcellular
compartments, with different contributions from intracellular and transmembrane Ca2+ sources.
This nuanced view of astrocyte Ca2+ signaling is critical for understanding the role of astrocytes
in brain function.

10
1.2 Astrocytes as mechanosensitive cells
Astrocytes exhibit fluctuations in intracellular Ca2+ concentration thought to represent a form of
cellular excitability (Parpura and Verkhratsky, 2012). While neuronal activity-associated
astrocyte Ca2+ signaling is well-established, other studies have indicated that astrocytes are also
sensitive to mechanical stimulation. Early studies demonstrated that membrane deformation or
substrate stretch increased intracellular Ca2+ levels in cultured astrocytes, raising the possibility
that they can transduce mechanical stress into chemical signals (Charles et al., 1991; Ostrow et
al., 2000). Mounting evidence suggests that astrocytes express mechanosensitive channels and
can respond to a variety of mechanical signals including blood flow, matrix stiffness, and
osmotic stress (Kirischuk, 2009).
1.2.1 Astrocytes detect mechanical stimuli to regulate cell physiology
Cells are continuously exposed to mechanical forces that are integrated by the plasma membrane,
cytoskeleton, extracellular matrix (ECM), cell adhesion proteins, and ion channels to influence
cell functioning (Tyler, 2012). In particular, astrocytes are structurally well-suited to detecting a
diverse range of mechanical stimuli. For example, astrocyte membranes form specialized
processes known as endfeet that ensheath the blood vessels in the brain (Gordon et al., 2011).
Benfenati et al. (2007) described enriched expression of the mechanosensitive cation channel
TRPV4 in these endfeet compartments, raising the possibility that astrocyte endfeet may be
involved in detecting hemodynamic forces such as blood flow. The hypothesis was supported by
experiments in cannulated brain slice parenchymal arterioles demonstrating that increases in
blood flow stimulated astrocyte Ca2+ signals and subsequent vasoconstriction; Ca2+ elevation and
vessel response were both attenuated by patch-dialysis of the Ca2+ chelator BAPTA into

11
astrocytes, or by pharmacological inhibition or genetic deletion of TRPV4 (Kim et al., 2015).
Hence, astrocytic TRPV4 was suggested to play a significant role in sensing blood flow and
regulating parenchymal arteriole tone.
Astrocyte functions have also been intimately linked to the extracellular environment.
Astrocytes secrete a number of ECM proteins, including hevin, tenascin-C, and fibronectin
which provide a microenvironment promoting proper synaptogenesis, synaptic plasticity, and
blood-brain barrier integrity (Faissner et al., 2010; Benarroch, 2015). Interestingly, astrocyte
morphology and function are also regulated by interactions with the ECM. Astrocytes express
integrins, a family of transmembrane cell-adhesion molecules that act as receptors for various
ECM proteins (Milner et al., 1999; Tanigami et al., 2012). The cytoplasmic domains of integrin
proteins are associated with signaling complexes and cytoskeletal components such as actin
stress fibers (Geiger et al., 2009; Campbell and Humphries, 2011). Thus, astrocytes can sense
physical properties of their environment through the forces generated from integrin-mediated
adhesions to the ECM, which are then transduced intracellularly to modulate cellular response
(Schwartz, 2010). For instance, the molecular composition and mechanical stiffness of the cell
substrate affect proliferation, reactive oxygen species production, glutamate uptake, and
migration in cultured astrocytes (Johnson et al., 2015; Wilson et al., 2016). Since the protein
composition of the ECM and tissue stiffness vary with age and disease state (Lau et al., 2013;
Arani et al., 2015; Murphy et al., 2016), the ability of astrocytes to discriminate between
different extracellular environments may play an important role in regulating both physiological
and pathological astrocyte functions.

12
Astrocytes have also been implicated in the maintenance of volume and ion homeostasis in the
brain (Simard and Nedergaard, 2004). Electron microscopy studies revealed that aquaporin-4
(AQP4), the predominant water channel in the brain (Nagelhus and Ottersen, 2013), is
selectively localized to astrocytes, with the strongest staining observed in brain regions
associated with detection and regulation of osmolarity (Oliet and Bourque, 1993; Nielsen et al.,
1997). AQP4 expression is enriched in astrocyte endfeet lining blood vessels and the pial
surface, and these endfeet compartments have been proposed to form a paravascular
compartment as part of a ‘glymphatic’ system that promotes interstitial fluid and osmolyte
clearance from the brain (Thrane et al., 2014). The selective expression of AQP4 in astrocytes
may underlie the cell’s sensitivity to osmotic changes. Wasterlain and Torack (1968) examined
rat brain cell morphology two hours following intraperitoneal injections of water; this model of
water intoxication induced astrocyte swelling while neurons and oligodendrocytes were
unaffected. In addition to AQP4 channels, astrocytes also express a range of channels and
transporters involved in uptake of ions and neurotransmitters from the extracellular space (ECS),
which is followed by obligatory water entry to maintain the osmotic gradient across the
membrane (Simard and Nedergaard, 2004; Vargova and Sykova, 2014). Therefore, astrocyte
swelling is observed in response to both physiological and pathological perturbations to ECS
homeostasis (Kimelberg et al., 1993; Florence et al., 2012). Astrocyte swelling can activate
mechanosensitive membrane channels, which will be discussed in Section 1.2.2.
1.2.2 Membrane proteins linked to mechanosensitivity in astrocytes
Mechanosensitive channels in the CNS can be activated by plasma membrane perturbations such
as osmotic stress (Tyler, 2012). Initial studies by Kimelberg and O’Connor (1988)

13
demonstrated that perfusion with hypoosmotic solution induced cell swelling and reversible
membrane depolarization in cultured astrocytes, and subsequent ion-replacement studies
indicated that both cation- and anion-permeable channels may contribute to the hypotonic-
induced membrane potential changes (Kimelberg et al., 1990; Pasantes-Morales et al., 1994).
Furthermore, the mechanosensitive channel blocker GsMTX4 inhibited swelling-activated
currents in astrocytes (Suchyna et al., 2000). Together, the data support that astrocytes express
channels that can be activated by hypoosmotic stimuli-induced swelling. While their molecular
identities have not been fully characterized, putative astrocyte mechanosensitive channels are
described below.
The transient receptor potential (TRP) family is a group of non-selective cation channels that
responds to a diverse range of stimuli, including temperature, pressure, and inflammatory agents
(Moran et al., 2004). Six types of TRP channels have been identified in mammals: the classical
TRPs (TRPCs), the vanilloid receptor TRPs (TRPVs), the melastatin TRPs (TRPMs), the
mucolipins (TRPMLs), the polycystins (TRPPs), and the ankyrin transmembrane protein 1
(TRPA1) (Moran et al., 2004). Amongst the TRPs, the best candidate for sensing osmotic
stimuli in astrocytes is the TRPV4 channel (Liedtke and Kim, 2005; Plant, 2014). The earliest
evidence of TRPV4 as an osmosensor came from Ca2+ imaging and electrophysiological studies.
In HEK293 or CHO-K1 cells transfected with TRPV4, perfusion of hypoosmotic solution
induced an increase in intracellular Ca2+ and activated cationic currents, which were both
inhibited by non-selective mechanosensitive channel blockers such as ruthenium red and La3+
(Liedtke et al., 2000; Strotmann et al., 2000). TRPV4 expression has been identified using
immunofluorescence approaches in both cultured and brain slice rat cortical astrocytes

14
(Benfenati et al., 2007). Electron microscopy studies suggest that TRPV4 is enriched at
astrocyte endfeet bordering blood vessels and the pial surface (Benfenati et al., 2007). Astrocyte
TRPV4 channels are functional, as application of a TRPV4 agonist or hypotonicity-induced
swelling caused intracellular Ca2+ elevations in cultured astrocytes, and these signals were
inhibited by ruthenium red (Benfenati et al., 2007). Interestingly, TRPV4 has been shown to be
involved in a homeostatic mechanism termed regulatory volume decrease (RVD), where cell
volume recovers following hypotonicity-induced swelling. Overexpression of recombinant
TRPV4 conferred RVD to a cell line that could not reduce its volume following hypoosmotic
stress (Becker et al., 2005), while RVD was impaired in astrocytes with siRNA-mediated TRPV4
knockdown (Benfenati et al., 2011). Therefore, TRPV4 may contribute to both detection of
volume change and subsequent volume recovery in response to osmotic stimuli in astrocytes.
Mechanosensitive K+-selective channels have also been found expressed in astrocytes, although
they remain to be fully characterized at the molecular level (Bowman et al., 1992; Kirischuk,
2009). These channels belong to the two-pore domain K+ (K2P) channel family, a group of
structurally similar K+ channels that are formed by dimerization (Feliciangeli et al., 2015). Of
the fifteen K2P channels identified in mammals, only the TREK-1, TREK-2, and TRAAK
channels are mechanosensitive (Ryoo and Park, 2016). Gnatenco et al. (2002) found that
cultured astrocytes contained TREK-2 mRNA, and functional expression was confirmed with
single-channel recordings. Of note, TREK-2 activity was increased in astrocytes exposed to
hypotonic solution (Gnatenco et al., 2002), consistent with previous studies suggesting that
TREK channels are activated by membrane stretch (Patel et al., 1998). Although TREK-1

15
expression has also been reported in astrocytes (Zhou et al., 2009), its functions related to
mechanotransduction have not been described.
Previous studies have also recognized that osmotic swelling activates Cl- currents in astrocytes
(Lascola and Kraig, 1996; Crépel et al., 1998). The currents were proposed to be mediated by
volume-regulated anion channels (VRACs) that, in addition to being permeable to Cl-, can also
flux organic osmolytes such as glutamate, aspartate, and taurine (Kimelberg et al., 2006). Recent
work has identified the LRRC8 family of transmembrane proteins as the molecular constituents
of VRACs (Qiu et al., 2014; Voss et al., 2014), an important advance that has allowed for
development of genetic tools to specifically target these channels. LRRC8 mRNA has since
been identified in cultured astrocytes, and siRNA-mediated LRRC8 knockdown was found to
inhibit hypoosmotic solution-induced amino acid efflux, suggesting that astrocytes express
functional LRRC8 proteins (Hyzinski-García et al., 2014; Schober et al., 2017). However,
VRACs have been demonstrated to be activated by reduction of intracellular ionic strength in
cells maintained at constant volume (Pedersen et al., 2015). Hence, while VRACs may respond
to hypotonic stimuli, the channels may not be intrinsically mechanosensitive.
In summary, astrocytes express a variety of channels activated by osmotic stress, and the
examples described above may represent just a subset. Future experiments examining the
concerted activation of these channels in response to hypotonic stimuli are required to determine
whether the channels are functionally redundant, or if they act in a complementary fashion to
encode different aspects of the mechanical force.

16
1.3 Piezo1: a mechanosensitive cation channel
Mechanosensitive cation channels mediate the transduction of mechanical forces into chemical
or electrical signals that allow cells to sense and adapt to changes in their environment (Gillespie
and Walker, 2001). Studies in Caenorhabditis elegans and Drosophila melanogaster mutants
initially implicated mechanosensitive channels in sensory modalities such as touch and hearing,
and identified three classes of channel proteins as putative mechanosensory channels: the
degenerin/epithelial Na+ channel (DEG/ENaC) family, the transient receptor potential (TRP)
channels, and the two-pore-domain K+ (K2P) channels (reviewed in Chalfie, 2009). In mammals,
mechanical stimuli such as touch, pressure, and tendon stretch are converted to electrical signals
by mechanosensory transducers on specialized somatosensory neurons that can lead to
depolarization and generation of action potentials that propagate to the CNS (Ernstrom and
Chalfie, 2002; Delmas et al., 2011). However, given the inaccessibility of the small sensory
nerve endings and the sparseness of the mechanosensing molecules, the molecular entities
responsible for mammalian mechanotransduction have been difficult to study (Chalfie, 2009;
Delmas et al., 2011).
Early experiments measuring the speed of sensory transduction in the hair cells of the bullfrog
sacculus found that an electrical response was produced within 40 µs, suggesting that the
mechanism was too rapid to involve chemical signaling; rather, the electrical response was
proposed to arise from direct activation of an ion channel (Corey and Hudspeth, 1979).
Researchers therefore focused on channel proteins that may be directly gated by mechanical
stimuli in their search for the molecules underlying mechanotransduction. However,
investigations of homologous proteins identified by the invertebrate mutant screens yielded

17
mixed results, as deletions of the genes in mouse models were rarely associated with a clear loss
of phenotype (Alloui et al., 2006; Bautista et al., 2006). For example, while DEG/ENaC
channels mediate touch sensitivity in C. elegans (Chalfie and Sulston, 1981), loss of the
homologous ASIC2 in knockout mice did not produce touch defects, suggesting that the
invertebrate channels may not be functionally conserved in mammals (Drew et al., 2004).
Therefore, the molecular identities of mechanosensitive channels in mammalian cells remain
incompletely characterized.
In 2010, Coste et al. identified a novel class of mammalian mechanosensitive channels by
screening siRNA sequences against candidate channels for efficacy in inhibiting endogenous
mechanically activated ionic currents in the Neuro2A mouse neuroblastoma cell line. The
authors found that knockdown of the Fam38A gene reduced pressure- and stretch-induced
currents in the Neuro2A cells, and renamed the gene Piezo1, from the Greek word píesi meaning
‘pressure’. A second related gene, Piezo2, was later cloned from dorsal root ganglia neurons.
While the Piezo proteins are conserved amongst animal, plant, and other eukaryotic species, they
lack homology to other channels. Piezo1 or Piezo2 overexpression in multiple mammalian cell
lines demonstrated large mechanically activated cation currents that were sensitive to ruthenium
red and gadolinium, two non-specific blockers of stretch-activated channels. Taken together, the
data suggest that the proteins are involved in generating currents in response to mechanical force.
The role of Piezo proteins in mechanotransduction has since been demonstrated in a number of
organ systems including the bladder, the lung, and the vasculature (Miyamoto et al., 2014;
Ranade et al., 2014; Nonomura et al., 2016).

18
1.3.1 Piezo1 channel topology and the structural basis of function
Mammalian Piezo1 is a large transmembrane protein of over 2500 amino acids (Coste et al.,
2010). Piezo1 was first proposed to assemble as an oligomeric protein based on evidence from
photobleaching experiments of mouse Piezo1 – GFP fusion proteins expressed in Xenopus
oocytes, and size analysis of glutathione S-transferase-tagged Piezo1 proteins resolved by gel
electrophoresis (Coste et al., 2012; Ge et al., 2015). The model was later refined by cryo-
electron microscopy that allowed the three-dimensional structure of Piezo1 to be determined at a
resolution of 4.8 Å. Mouse Piezo1 was found to be a homotrimer, with three propeller-like
wings consisting of transmembrane helices and extracellular blade domains, supported by
intracellular beam and anchor domains that surround an ion-conducting pore (Fig. 1-2) (Ge et al.,
2015).
Figure 1-2. Structure of mouse Piezo1.
A) Extracellular, B) side, and C) intracellular views of the cyro-electron microscopy structures of Piezo1. D)
Schematic diagram of Piezo1 showing the major structural features of the channel, with the putative
mechanotransducing and pore modules illustrated in blue and red, respectively. CED, C-terminal extracellular loop
domain; CTD, C-terminal domain. Adapted from Geng et al., 2017.
Piezo1 topology was further characterized with biochemical mapping of Flag- or Myc-tagged
amino acids to intracellular and extracellular domains, leading to a prediction of 14 – 18

19
transmembrane helices for each wing domain (Coste et al., 2015; Ge et al., 2015). Subsequent
studies suggested that the C-terminal extracellular loop domain (CED) and the intracellular C-
terminal domain (CTD) form the central pore region of Piezo1, as replacing this region of the
mouse protein with the analogous sequence from Drosophila resulted in conductance properties
that resembled the characteristics of the fly Piezo (Zhao et al., 2016). Site-directed mutagenesis
studies also indicated that this pore region determines the conduction properties of Piezo1,
including unitary conductance, ion selectivity, and sensitivity to pore blockers (Coste et al.,
2015; Zhao et al., 2016).
In order to elucidate the structural components of Piezo1 that mediate mechanotransduction,
Zhao et al. (2016) replaced the Piezo1 pore region with the mechano-insensitive ASIC1 channel,
resulting in a chimeric protein that can respond to both mechanical force and pH. This indicated
that the non-pore-containing region of Piezo1 can confer mechanosensitivity to the ion-fluxing
pore (Zhao et al., 2016). The authors proposed that the extracellular blades and transmembrane
wing domains may sense mechanical stimuli, which are then transmitted through the beam and
interfacing anchor domains to cause conformational changes of the pore module (Zhao et al.,
2016). Hence, the structure of Piezo1 suggests that the channel contains mechanotransduction
components and raises the possibility that Piezo1 may be inherently mechanosensitive (Wang
and Xiao, 2017). Together, these topological and structural studies of Piezo1 provide insight
into the mechanotransduction mechanism of the channel, and form a foundation for designing
pharmacological agents to modify Piezo1 activity.

20
1.3.2 Properties of the Piezo1 channel
Piezo1 channels, like all mechanosensitive channels, mediate the transduction of physical forces
into conformational changes associated with channel opening. Two models have been proposed
to explain the mechanism of molecular force transduction: the “force through lipid” model
wherein bilayer tension directly acts on the channel, and the “force through filaments” model
where mechanical stimuli are transduced through physical tethers to the cytoskeleton or the
extracellular matrix (ECM) (Ranade et al., 2015). While some mechanosensitive channels have
been shown to be exclusively gated by one mechanism, most membrane channels are also
associated with scaffolding proteins that are linked to cytoskeletal and ECM proteins (Zhang et
al., 2000; Cox et al., 2017), suggesting that both models may contribute to channel activation.
To address whether Piezo1 can be activated by membrane tension alone, Piezo1-transfected
HEK293 cells were treated with hypoosmotic solution to create membrane blebs, protrusions of
the bilayer that are uncoupled to the underlying cytoskeleton. Mechanically stimulated Piezo1
currents could be recorded from patched blebs, and Piezo1 activity persisted in cells that had
been pretreated with drugs to disrupt microtubule assembly and actin depolymerization (Cox et
al., 2016). In addition, spontaneous Piezo1 activity was observed in channels reconstituted in
asymmetric bilayers (Syeda et al., 2015). The data suggest that Piezo1 may be inherently
mechanosensitive in the absence of other cellular components, as predicted by its cryo-electron
microscopy structure (Zhao et al., 2016), and support that mechanical force can be transmitted
directly from the membrane to the channel.

21
Mechanical stresses might affect cell shape and produce distortion of the cell surface and several
studies have examined the involvement of the cytoskeleton and associated scaffolding proteins in
the transduction mechanism responsible for the activation of Piezo1 in response to mechanical
forces propagated from the ECM. For example, mechanically-stimulated Piezo1 whole-cell
currents were inhibited by actin disruption in a heterologous expression system (Gottlieb et al.,
2012). The scaffold protein stomatin-like protein 3 (STOML3), a molecule necessary for
mechanosensitive ion channel function in mouse sensory neurons, was found to sensitize Piezo1
channels in Neuro2A cells, possibly via cholesterol binding and modulation of plasma membrane
stiffness (Wetzel et al., 2007; Poole et al., 2014; Qi et al., 2015). Gaub and Müller (2017) found
that plating Piezo1-transfected HEK293 cells onto ECM proteins sensitized Ca2+ signals induced
by membrane deformation compared to using a glass substrate. Furthermore, Pathak et al.
(2014) described spontaneous Piezo1-mediated Ca2+ signals in neural stem cells that were
modified by substrate stiffness, and disruption of the traction force used by the cells to sense
ECM rigidity was associated with loss of Piezo1 activity. Taken together, the data suggest that
force from both membrane tension and physical tethers can affect Piezo1 activity, and that the
contribution from each may depend on the cell type and/or mechanical stimulus experienced by
the cell (Nourse and Pathak, 2017).
Piezo1-mediated cationic currents decay rapidly after activation by a mechanical stimulus (Coste
et al., 2010). Previous studies have shown that Piezo1 inactivation regulates the ability of the
channel to act as a frequency filter of repetitive mechanical stimuli (Lewis et al., 2017). In
contrast to activation, Piezo1 inactivation is independent of membrane tension and actin
cytoskeleton modulation (Gottlieb et al., 2012; Wu et al., 2017). Rather, channel inactivation

22
was observed to be voltage-dependent, with faster inactivation occurring at more hyperpolarized
membrane potentials (Bae et al., 2013b). Therefore, inactivation-dependent modulation of
Piezo1 may be particularly important in excitable cells such as neurons and cardiomyocytes.
Mutations in the C-terminal extracellular loop domain (CED) and intracellular C-terminal
domain (CTD) were associated with slowing of inactivation (Bae et al., 2013b). The
involvement of the CED in regulating Piezo1 inactivation kinetics was further observed in
chimeric proteins where the CEDs of Piezo1 and Piezo2 were exchanged (Wu et al., 2017). In
addition, mutation of a single lysine residue within the inner pore helix abolished the voltage-
dependency of Piezo1 inactivation (Wu et al., 2017). The data suggest that the CED and inner
pore module may act in concert to mediate Piezo1 inactivation.
Electrophysiological recordings from Piezo1-overexpressing HEK293 cells revealed
mechanically activated inward currents that were blocked when cations in the extracellular
solution were substituted by non-permeant N-methyl-D-glucamine, suggesting that Piezo1 is a
non-selective cation channel (Coste et al., 2010). Piezo1 was found to be permeable to both
monovalent and divalent cations, with the highest permeability for K+ (Gnanasambandam et al.,
2015). Site-directed mutagenesis studies have indicated that the ion selectivity and single-
channel conductance of Piezo1 is determined by specific glutamate residues in the C-terminal
region forming the pore module (Coste et al., 2015; Zhao et al., 2016), where the negatively
charged amino acids may act as a selectivity filter to allow for preferential flux of cations over
anions through the Piezo1 channel.

23
1.3.3 Modulation of Piezo1 channel activity by second messengers
While mechanosensitive channels are primarily gated by mechanical stimuli, their functions can
be modulated by intracellular second messengers such as phospholipid-derived signaling
molecules and protein kinases. For instance, the mechanosensitive K+ channel TREK-1 has been
shown to be inhibited by protein kinase A- and protein kinase C-mediated phosphorylation
downstream of G-protein-coupled receptor (GPCR) activity (Noël et al., 2011).
Phosphatidylinositol-4,5-bisphosphate (PI(4,5)P2), the most abundant inositol phospholipid in
the plasma membrane (Fruman et al., 1998), has also been demonstrated to regulate TREK-1
activity (Chemin et al., 2005). These data raise the possibility that Piezo1 activity may also be
similarly sensitive to modulation by second messenger pathways.
Borbiro et al. (2015) published one of the first reports documenting the effects of PI(4,5)P2 on
Piezo1 activity. Using a heterologous expression system coexpressing the heat-sensitive TRPV1
channel with Piezo1, the researchers found that Piezo1-mediated mechanosensitive currents were
attenuated by TRPV1 activation with capsaicin. This inhibition was dependent on TRPV1-
mediated Ca2+ influx, activation of the phospholipase C δ (PLCδ) isoform, and subsequent
decrease in PI(4,5)P2 levels. Depletion of PI(4,5)P2 with a chemically inducible phosphatase
system decreased Piezo1 currents, while intracellular application of excess PI(4,5)P2 rescued the
capsaicin-induced inhibition of Piezo1 currents. The data suggest a functional link between
TRPV1 and Piezo1, and that PI(4,5)P2 is required for Piezo1 activity.
A recent study suggested that Piezo1 activity can also be modified by GPCR signaling pathways
(Lawrence et al., 2017). Piezo1 was found to promote Ca2+ accumulation and cell death of

24
human chondrocytes, and that was blocked by either stimulation of endogenous Gαs-linked
receptors or pharmacological elevation of cAMP production. In addition, phospholipase A2
inhibition was observed to reduce Piezo1-mediated cell death, leading to the speculation that a
yet-unidentified lipid pathway may contribute to Piezo1 activation. However, it is currently
unclear whether these putative lipid modifiers of Piezo1 activity interact directly or indirectly
with the channels (Borbiro and Rohacs, 2017). The factors regulating the relevance of these
signaling pathways in different Piezo1-expressing cells also remain to be determined.
1.3.4 Piezo1 pharmacology
To establish Piezo1 as a mechanosensitive channel, initial studies tested the sensitivity of Piezo1-
mediated currents to the non-specific mechanosensitive ion channel blockers, gadolinium (Gd3+)
and ruthenium red (Coste et al., 2010). Gd3+ has been proposed to bind to negatively charged
headgroups of membrane phospholipids, affecting membrane curvature and force distribution
within the bilayer, with direct consequences for channel gating (Ermakov et al., 2010). This
membrane-based mechanism of channel inhibition supports previous observations that the lipid
environment can modulate Piezo1 activity (see section 1.3.3). In contrast, ruthenium red likely
acts as a pore blocker, since mutations of specific glutamate residues within the Piezo1 pore
module resulted in loss of ruthenium red blockade (Coste et al., 2012; Zhao et al., 2016). This
pharmacological approach has been used to demonstrate the role of Piezo1 in mediating
functions such as ATP release from red blood cells and stretch-induced proliferation of epithelial
cells (Cinar et al., 2015; Gudipaty et al., 2017). However, both inhibitors block a variety of other
targets, such as voltage-gated Ca2+ and Na+ channels (Biagi and Enyeart, 1990; Elinder and

25
Arhem, 1994; Malécot et al., 1998), and therefore cannot be used as the sole strategy to rule out
contributions from other channels.
The first specific blocker for mechanosensitive channels was isolated from the venom of the
tarantula Grammostola spatulata, which had previously been reported to block mechanically
activated currents in Xenopus oocyte and chick cardiac cells (Niggel et al., 1996). By separating
venom components using reverse phase HPLC and testing each fraction against stretch-activated
channels in astrocytes, Suchyna et al. (2000) identified a 34-amino acid peptide Grammostola
spatulata mechanotoxin 4 (GsMTx4) that inhibited mechanically activated currents without
affecting voltage-sensitive currents. GsMTx4 was found to inhibit mechanically induced
currents in Piezo1-transfected HEK293 cells, with a rightward shift in the activation curve,
suggesting the toxin acts as a gating modifier (Bae et al., 2011). Interestingly, both D and L
enantiomers of GsMTx4 were effective, indicating that the toxin does not bind to a specific
complementary binding site on Piezo1 (Bae et al., 2011). Instead, GsMTx4 may mediate Piezo1
inhibition by modulating the lipids surrounding the channel to decrease the efficiency of force
transduction between the membrane bilayer and the channel (Gnanasambandam et al., 2017).
While GsMTx4 is an important tool for studying the physiological and pathological roles of
mechanosensitive channels, the toxin does not exclusively block Piezo1 (Bowman et al., 2007)
and to date, no specific Piezo1 pharmacological antagonists have been identified.
Recently, a Piezo1 activator, Yoda1, was identified by screening synthetic compounds for their
ability to induce Ca2+ entry in Piezo1-transfected HEK293 cells (Syeda et al., 2015).
Electrophysiological recordings demonstrated that Yoda1 could modify Piezo1-mediated

26
mechanically activated currents by slowing their inactivation and shifting the activation curve to
the left, indicating sensitization of Piezo1 to mechanical stimulation (Syeda et al., 2015). To
study the mechanism of Yoda1 action, its effects on Piezo1 activity were examined on channels
reconstituted in an artificial cell membrane. Yoda1 was found to induce single-channel currents
with increased open time (Syeda et al., 2015). Of note, Yoda1 did not affect Piezo2 activity.
Since Yoda1 can specifically activate Piezo1 in the absence of other cellular components, Syeda
et al. (2015) proposed that Yoda1 may interact directly with Piezo1. The development of Yoda1
suggests that Piezo1 is not exclusively gated by mechanical forces (Syeda et al., 2015).
1.3.5 Piezo1 expression and physiological functions
Piezo1 is widely expressed in various mechanically sensitive cells. In the mouse, Piezo1 mRNA
was identified in tissues such as the lung, bladder, kidney, skin, and colon (Coste et al., 2010),
with subsequent studies characterizing the channel’s functional role in different organs. For
instance, Piezo1 has been shown to mediate Ca2+ influx in bladder urothelial cells upon
mechanical stretch, leading to downstream ATP release that may act as signal for bladder
contractions (Miyamoto et al., 2014). Piezo1 is also expressed by chondrocytes in cartilage,
where it mediates compression-evoked Ca2+ transients with possible consequences for
cytoskeleton remodeling and energy homeostasis (Lee et al., 2014). In addition, the function of
the channel in vivo has been demonstrated in the vascular system, where Piezo1 in endothelial
cells can detect shear stress to mediate proper cell alignment; Piezo1 knockdown with siRNA
was associated with abnormal endothelial cell alignment, while genetic deletion led to vascular
abnormalities and embryonic lethality (Ranade et al., 2014).

27
Previous studies have also raised the possibility for Piezo1-mediated mechanosensing in the
CNS. For example, cultured rat hippocampal neurons associate with astrocytes in a manner
dependent on the roughness of the cell substrate, an association lost with GsMTx4 treatment,
suggesting that Piezo1 may mediate neuronal sensing of substrate topology (Blumenthal et al.,
2014). In neural stem cells, Piezo1-mediated Ca2+ influx was demonstrated to be affected by
substrate stiffness, with downstream consequences for differentiation along either a neuronal or
astrocytic lineage (Pathak et al., 2014). Furthermore, Xenopus retinal ganglion cells were
proposed to use Piezo1 as a detector for sensing brain tissue stiffness and promoting axon growth
toward softer tissues, as Piezo1 knockdown or GsMTx4 treatment led to disrupted axonal
pathfinding (Koser et al., 2016).
Piezo1 expression in astrocytes is supported by transcriptomic and proteomic studies. Piezo1
mRNA was found in cortical astrocytes from both P7 and P80 mice (Zhang et al., 2014;
Srinivasan et al., 2016), while the protein was detected in primary astrocyte cultures as well as in
adult mouse brains (Sharma et al., 2015). An early report by Satoh et al. (2006) suggested that
Piezo1 may be upregulated in reactive astrocytes associated with amyloid-beta plaques in
Alzheimer’s disease. However, the physiological functions of Piezo1 in astrocytes has not been
characterized.
1.3.6 Pathologies associated with Piezo1 dysfunction
Consistent with the role of Piezo1 in mechanosensing, mutations in human Piezo1 have been
linked to multiple hereditary disorders (Alper, 2017). While most mutations remain to be fully
characterized, Piezo1 loss-of-function mutations are associated with congenital lymphatic

28
dysplasia, wherein abnormal function of the lymphatic system results in symptoms such as
generalized edema, seizures, and microencephaly (Fotiou et al., 2015; Lukacs et al., 2015).
HEK293 cells overexpressing the G2029R Piezo1 loss-of-function mutant displayed impaired
Yoda1-induced Ca2+ responses, minimal mechanically activated currents, and reduced Piezo1
surface expression (Lukacs et al., 2015). The mutation may also have hematological effects, as
some patients exhibited red blood cell swelling and mild hemolysis (Fotiou et al., 2015).
In contrast, Piezo1 gain-of-function mutations have been linked to a type of hemolytic anemia
known as autosomal dominant hereditary xerocytosis, or dehydrated stomatocytosis
(Zarychanski et al., 2012; Andolfo et al., 2013; Alper, 2017). In HEK293 cells overexpressing
Piezo1 with two gain-of-function mutations, pressure- or osmotic stress-induced cationic
currents exhibited delayed inactivation (Albuisson et al., 2013). Mechanistically, this results in
increased Ca2+ influx into red blood cells, leading to K+ channel activation and K+ efflux,
followed by osmotically driven dehydration (Cahalan et al., 2015). The clinical phenotypes of
both gain-of-function and loss-of-function Piezo1 mutants suggest that a balanced level of
mechanical sensitivity is likely required to maintain normal cell health.
Since Piezo1 was first identified as a mechanosensitive cation channel, the understanding of
force transduction in mammalian cells has advanced rapidly. These developments were aided by
structure-function studies and pharmacological tools, and it is now well-established that Piezo1
is involved in many physiological responses in a wide variety of cell types. The importance of
Piezo1 function is further supported by examples of Piezo1 mutations that lead to pathologies
associated with impaired mechanical sensing.

29
1.4 Rationale and hypotheses
Over the last few decades of research, technological advances in the field of astrocyte Ca2+
signaling have allowed Ca2+ levels to be monitored in fine astrocyte arborizations, revealing a
new and diverse repertoire of Ca2+ signals and refining our understanding of the physiological
roles they mediate. These new approaches have revealed an intriguing spontaneous Ca2+ signal
localized to microdomains in the fine astrocytic processes. Recent studies have provided
evidence for the role of these signals in regulating astrocyte basal Ca2+ levels, with implications
for synaptic activity and vessel tone. However, the molecular mechanisms underlying the
spontaneous Ca2+ transients have not been fully characterized. While the signals are dependent
on transmembrane Ca2+ influx, they are resistant to pharmacological block or genetic deletion of
a range of Ca2+-permeable candidate channels. Therefore, determining the molecular identity of
the channel or transporter mediating these signals would be an important contribution to the
overall understanding of the role of astrocytes in the physiology and pathology of the CNS.
We initially searched for novel Ca2+ fluxing proteins in recently published rodent brain
transcriptomic and proteomic databases, and found that both Piezo1 mRNA and protein were
expressed in astrocytes. Piezo1 is a non-selective cationic channel that is a potential candidate
for mediating Ca2+ entry across the plasma membrane. Importantly, astrocytes are known to be
mechanosensitive cells that respond to physical forces such as extracellular matrix stiffness and
hypotonicity-induced swelling, mechanical stimuli previously shown to activate Piezo1 in other
cell types. We therefore decided to investigate the role of Piezo1 in astrocytes, from the
perspective of a Ca2+ signal mediator as well as a force transducer. We hypothesize: 1) that
spontaneous Ca2+ transients in astrocytes are mediated by Piezo1 channel activity, 2) that

30
Piezo1 is expressed in astrocyte subcellular compartments relevant to
mechanotransduction, and 3) that Piezo1 can be activated by hypoosmotic stress-induced
astrocyte swelling.
1.4.1 Objective 1: To determine the contribution of Piezo1 to spontaneous Ca2+
transients in astrocytes
Astrocytes exhibit spontaneous Ca2+ microdomain signals that are abolished in the absence of
extracellular Ca2+, suggesting the involvement of a yet-undefined Ca2+ entry pathway. I will
monitor Ca2+ microdomain signals in cultured astrocytes using the membrane-tethered GECI,
Lck-GCaMP5, and assess the effects of pharmacological Piezo1 agonists and antagonists and
siRNA-mediated Piezo1 knockdown to examine the role of Piezo1 in mediating the spontaneous
Ca2+ signals.
1.4.2 Objective 2: To characterize the subcellular localization of Piezo1 in astrocytes
Piezo1 expression has been identified in various subcellular compartments in different cell types;
some compartments, such as focal adhesions, are associated with mechanotransduction, in
contrast with other structures such as the endoplasmic reticulum. I will identify the subcellular
distribution of Piezo1 in cultured astrocytes using antibody labeling and fluorescent-tagged
protein approaches, and compare the staining patterns to those of the organelle-specific markers.

31
1.4.3 Objective 3: To examine the activation of Piezo1 in hypotonicity-induced astrocyte
swelling
Previous studies found that Piezo1 responds to a range of mechanical forces, including
extracellular matrix rigidity, shear stress, membrane stretch, and osmotic stress. Astrocytes are
involved in maintaining CNS volume homeostasis and exhibit swelling in response to both
physiological and pathological stimuli. I will utilize hypoosmotic solution as a tool to induce
cell swelling and perform Ca2+ imaging simultaneously to test the possibility that a mechanical
force relevant to astrocytes is capable of activating Piezo1 channels.

32
Chapter 2: Piezo1 mediates astrocyte Ca2+ microdomain signals
2.1 Overview
Pioneering studies in astrocyte Ca2+ signaling have established the functional importance of these
signals in the brain (Shigetomi et al., 2016). By combining techniques such as bulk-loaded Ca2+
indicator dyes and caged Ca2+ compounds, researchers could selectively monitor and manipulate
Ca2+ levels in the astrocyte soma (Mulligan and MacVicar, 2004). This has led to findings such
as the ability of various neurotransmitters to evoke Ca2+ release from astrocyte intracellular
stores (Dani et al., 1992; Duffy and MacVicar, 1995; Porter and McCarthy, 1996; Pasti et al.,
1997), resulting in gliotransmitter and arachidonic acid metabolite release from astrocytes
(Parpura et al., 1994; Bezzi and Volterra, 2001) that in turn acts on neurons and vascular smooth
muscle to regulate synaptic plasticity and blood flow (reviewed in Volterra et al., 2014;
Bazargani and Attwell, 2016). However, subsequent studies demonstrated that IP3 type 2
receptor (IP3R2) knockout mice had normal synaptic activity and neurovascular coupling, despite
impaired astrocyte soma Ca2+ responses (Petravicz et al., 2008; Nizar et al., 2013; Bonder and
McCarthy, 2014). Therefore, the physiological relevance of astrocyte Ca2+ signals remained
controversial.
Recent developments in Ca2+ imaging techniques have revealed both spontaneous and
neurotransmitter-evoked microdomain Ca2+ signals within astrocyte fine processes (Shigetomi et
al., 2010; Rungta et al., 2016; Agarwal et al., 2017). These Ca2+ transients, detected by patch-
loaded dialysis of Ca2+ dyes or genetically encoded Ca2+ indicators (GECIs), show different
spatial and temporal characteristics compared to the somatic signals. Importantly, experiments

33
with IP3R2 knockout mice indicated that a substantial proportion of microdomain transients
persisted in the fine processes while Ca2+ signals in the astrocyte soma were largely abolished
(Srinivasan et al., 2015), suggesting that Ca2+ signals in different subcellular compartments may
have different underlying mechanisms and functions. These Ca2+ microdomain signals have
since been linked to regulation of neuronal and vascular function via modulation of basal
astrocyte Ca2+ levels and gliotransmitter release or uptake (Di Castro et al., 2011; Panatier et al.,
2011; Shigetomi et al., 2012; Otsu et al., 2015; Rosenegger et al., 2015). However, the
mechanisms mediating Ca2+ transients remain incompletely described.
Previous studies have shown that the spontaneous microdomain Ca2+ signals depend on
extracellular Ca2+ entry (Shigetomi et al., 2012; Rungta et al., 2016). One possible mediator of
these Ca2+ transients is the TRPA1 channel, as indicated by the potency of a TRPA1 antagonist
in blocking the signals in cultured astrocytes (Shigetomi et al., 2012). However, subsequent
experiments found that a proportion of the signals persisted in brain slices from TRPA1
knockout mice (Shigetomi et al., 2013b; Rungta et al., 2016), indicating that other mediators for
Ca2+ sparks exist. Given that no treatment thus far has completely abolished the signals, the data
suggest that an unidentified transmembrane channel or transporter may also be involved. Thus,
elucidating the molecular pathway mediating these spontaneous Ca2+ microdomain signals will
contribute to the overall understanding of astrocyte physiology.
Piezo1 is a mechanosensitive cation channel that was first cloned from the Neuro2A
neuroblastoma cell line using an siRNA approach (Coste et al., 2010). Piezo1 channels have
since been identified in the brain as mechanosensors for detecting extracellular matrix

34
topography and rigidity, with consequences for neural stem cell differentiation and retinal
ganglion cell axon growth (Pathak et al., 2014; Koser et al., 2016). Intriguingly, transcriptome
and proteome analyses have also suggested that Piezo1 is expressed in astrocytes (Zhang et al.,
2014; Sharma et al., 2015; Srinivasan et al., 2016), although little is known about the
contribution of the channels to astrocyte physiology.
In this chapter, we present evidence for the role of Piezo1 in mediating astrocyte Ca2+
microdomain transients. Two-photon imaging was performed in cultured astrocytes transfected
with the Ca2+ indicator Lck-GCaMP5. We developed an analysis algorithm that allowed
microdomain signals to be quantified independently of other types of Ca2+ signals, and assessed
the effect of both pharmacological and genetic manipulations of Piezo1. Finally, we repeated
pharmacological experiments in brain slices to extend the findings to astrocytes in situ.
2.2 Methods
2.2.1 Chemicals and reagents
Stock solutions (1000x) of BAPTA-AM (Invitrogen), HC067047 (Tocris), thapsigargin (Tocris),
and Yoda1 (Tocris) were dissolved in dimethyl sulfoxide, while EGTA (Sigma) and gadolinium
(III) chloride (Tocris) were prepared in water. GsMTx4 (Smartox) was dissolved in water as a
50x stock solution. All chemicals were diluted in aCSF immediately prior to use.

35
2.2.2 Astrocyte cultures
Rat astrocytes were prepared using a protocol adapted from Kaech and Banker (2006). Brains
were harvested from E19 Sprague Dawley rat pups, minced with micro-scissors, and incubated at
37°C for 15 minutes in balanced salt solution (Gibco) with 0.25% trypsin (Gibco) and 0.1%
DNAse (Roche Applied Science). The dissociated tissue was filtered through 75 µm nylon
mesh, centrifuged, and resuspended in glia media (MEM (ThermoFisher Scientific)
supplemented with 14% fetal bovine serum (FBS; ThermoFisher Scientific), 0.6% glucose
(Sigma), penicillin (100 units/mL, ThermoFisher Scientific) and streptomycin (0.1 mg/mL,
ThermoFisher Scientific)). Cells were plated at 2-3 x 106 cells per 75 cm2 flask and grown for 7
– 9 days until confluent. Cells were then detached with 0.25% trypsin, centrifuged, resuspended
with glia media, and aliquoted into cryogenic vials for storage in liquid nitrogen. For
experiments, astrocytes were thawed, resuspended in glial medium, and plated onto 35 mm glass
bottom petri dishes (poly-D-lysine-coated, 10 mm glass diameter; MatTek) at a density of 4 x
103 cells / dish. Cells were grown for 3 to 5 days in a 37°C incubator with 5% CO2 in glia media.
Mouse astrocytes were prepared from heterozygous Piezo1-tdTomato mice expressing Piezo1
fused to the fluorescent marker tdTomato (Ranade et al., 2014; Jax stock no. 029214). Cortices
were harvested between P0 – P10 and triturated. Cells were plated in DMEM/F12 (Gibco)
supplemented with 10% FBS (Gibco) and 1% penicillin/streptomycin (Gibco) and grown for 2 –
3 weeks. The mixed glial culture was then shaken for 4 -5 hours at 120 rpm to dislodge
microglial cells, resulting in an enriched astrocyte culture. Astrocytes were passaged 1 – 3 times
and maintained in a 37°C incubator with 5% CO2 in glia media.

36
2.2.3 siRNA sequences for Piezo1 knockdown
An siRNA approach was used for Piezo1 knockdown. All siRNA oligonucleotides were ordered
from Integrated DNA Technologies. Uppercase letters represent unmodified bases, lowercase
letters represent 2’-O-methyl-modified bases, and dTsdT represents two phosphorothioate-linked
deoxythymidine bases. These modifications increase siRNA resistance to nuclease degradation
(Deleavey and Damha, 2012).
Luciferase siRNA was described by Addepalli et al. (2010):
Sense strand, 5’-cuuAcGcuGAGuAcuucGAdTsdT-3’
Antisense strand, 5’-UCGAAGuACUcAGCGuAAGdTsdT-3’
Piezo1 siRNA:
Sense strand, 5’-GAgGUUUAgAuGaUUGUACUUCuUggu-3’
Antisense strand, 5’-caAgAaGUACAaUcAuCuAAACCuC-3’
2.2.4 Lck-GCaMP5 plasmid preparation
Ca2+ signals were monitored with the membrane-targeted GECI Lck-GCaMP5. The pN1 Lck-
GCaMP5 plasmid was a gift from Baljit Khakh (Addgene plasmid #34924; Akerboom et al.,
2012). The plasmid was expanded in E. coli (DH5alpha strain) grown on agar plates (1.5% agar
(InvivoGen) with kanamycin (50 µg/mL; Alfa Aesar)) overnight at 37 °C, and colonies were
selected and grown at 37°C in LB broth (Sigma) for 8 hours to create a starter culture. The
starter culture was used to inoculate a flask of LB broth and grown overnight at 37 °C. The
broth was then centrifuged, and plasmid DNA was isolated from the pelleted bacteria by midi
prep (Qiagen) according to the manufacturer’s instructions.

37
2.2.5 Preparation of siRNA- and plasmid DNA-containing lipid nanoparticles
Lipid nanoparticles (LNPs), the ionizable cationic lipid system used to package and deliver
siRNA and plasmid DNA to astrocytes, were synthesized as previously described (Rungta et al.,
2013; Kulkarni et al., 2017). For siRNA encapsulation, LNPs were composed of heptatriaconta-
6,9,28,31-tetraen-19-yl 4-(dimethylamino)butanoate (DLin-MC3-DMA; Biofine International),
1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC; Avanti Polar Lipids), cholesterol (Sigma),
(R)-2,3-bis(octadecyloxy)propyl-1-(methoxy polyethylene glycol 2000) carbamate (PEG-DMG;
synthesized as previously described in Akinc et al., 2008), and the lipophilic carbocyanine dye
1,1’-dioctadecyl-3,3,3’,3’-tetramethylindocarbocyanine perchlorate (DiIC18; Invitrogen) at a
molar ratio of 50/10/37.5/1.5/1. Plasmid DNA was encapsulated in LNPs consisting of 2,2-
dilinoleyl-4-(2-dimethylaminoethyl)-[1,3]-dioxolane (DLin-KC2-DMA; Biofine International),
1-stearoyl-2-oleoyl-sn-glycero-3-phosphocholine (SOPC; Avanti Polar Lipids), cholesterol, and
PEG-DMG at a molar ratio of 50/10/39/1.
Lipids were solubilized in ethanol and mixed with siRNA or plasmid DNA dissolved in sodium
acetate buffer (25 mM, pH 4.0) at a volumetric ratio of 1:3 using a microfluidic micromixer
(Precision NanoSystems) or T-junction mixer. Ethanol was removed from the resulting
nanoparticles and replaced with phosphate-buffered saline (PBS, pH 7.4; Sigma) by dialysis.
LNPs were concentrated using Amicon Ultra centrifugal filter units (Millipore). Particle sizes
and polydispersity index (PDI) were quantified by dynamic light scattering (Malvern Zetasizer
NanoZS). Total lipid concentration of LNPs was determined by measuring cholesterol content
using the Cholesterol E assay (Wako Chemicals). Nucleic acid concentration was measured by
absorbance at 260 nm, and encapsulation efficiency was measured using the Quanti-IT

38
Ribogreen RNA and Picogreen dsDNA assays (Life Technologies) for siRNA and plasmid
DNA, respectively. LNP properties are summarized in Table 2-1.
Table 2-1. LNP properties.
Nucleic acid Size (nm) PDI % encapsulated
Lck-GCaMP5 plasmid 72.67 ± 3.34 0.073 ± 0.020 94.92 ± 2.84
Piezo1 siRNA 43.53 ± 3.85 0.072 ± 0.053 95.16 ± 1.84
Luciferase siRNA 40.78 ± 3.28 0.088 ± 0.031 95.25 ± 3.45
2.2.6 Astrocyte transfection with LNPs
Astrocyte cultures were treated with LNP-siRNA complexes (0.125 µg/mL) for five days prior to
experiments. Lck-GCaMP5 plasmids were either packaged in LNPs or transfected using
Effectene (Qiagen) in accordance with the manufacturer’s instructions. Astrocytes were exposed
to the plasmid (500 ng/mL) for two hours, and experiments were conducted after 48 hours. The
treatments were combined by first treating astrocytes with LNP-siRNA for three days, followed
by a two-hour incubation with LNP-Lck-GCaMP5, and experiments were performed after 48
hours.
2.2.7 Evaluation of transfection viability and efficiency
Fluorescence microscopy images were acquired using the EVOS FL Cell Imaging System
(Invitrogen) to evaluate the optimal conditions for Lck-GCaMP5 plasmid transfection efficiency
and cell viability. Lck-GCaMP5 plasmid transfection efficiency was determined by the number
of cells expressing green fluorescence. Cells were then fixed with 4% paraformaldehyde (Ted
Pella) at room temperature for ten minutes and stained with DAPI (Sigma) for visualization of

39
cell nuclei. Green fluorescence and DAPI images were taken with a 20x/0.40 NA and 4x/0.13
NA objective lens, respectively. Five fields from each coverslip were sampled.
2.2.8 Evaluation of Piezo1 siRNA effect on mRNA levels
To measure the effect of Piezo1 siRNA on mRNA levels, quantitative polymerase chain reaction
(qPCR) was performed on primary astrocyte cultures treated with 0.125 µg/mL LNP-siRNA
complexes (luciferase or Piezo1) for five days. Cells were treated with TRIzol (Invitrogen), and
total RNA was purified with the MagMAX-96 Microarray Total RNA Isolation Kit (Life
Technologies) followed by cDNA synthesis using the High Capacity cDNA Reverse
Transcription Kit (Applied Biosystems). Gene-specific qPCR was performed with the KAPA
Probe Fast Universal qPCR Kit (KAPA Biosystems) using a CFX384 Real-Time PCR System
(Bio-Rad). TaqMan probes were used to amplify PIEZO1 (rPiezo1-1 Rn.PT.58.12896870;
rPiezo1-2 Rn.PT.58.46120084), with GAPDH (rGAPDH Rn.PT.58.35727291) as the internal
control.
2.2.9 Evaluation of Piezo1 siRNA effect on protein levels
Immunoblot analysis was conducted to assess the effect of Piezo1 siRNA knockdown on protein
levels. Primary astrocyte cultures were treated with 0.125 µg/mL LNP-siRNA complexes
(luciferase or Piezo1) for five days. Cells were collected and homogenized on ice in RIPA buffer
(Tris Buffered Saline with 1% IGEPAL, 0.5% sodium deoxycholate, 0.1% SDS) supplemented
with cOmplete ULTRA protease inhibitor cocktail (Sigma Aldrich), phosSTOP phosphatase
inhibitors (Roche), and 1 mM EDTA. Cell homogenates were passed three times through a 27-
gauge syringe, incubated on ice for 30 minutes, then centrifuged at 4°C for 20 min at 12,000 rpm

40
to remove debris. Protein concentration was determined using a DC Protein Assay (Bio-Rad),
and equal protein amounts were incubated at room temperature for two hours in Laemmli
Sample buffer under reducing conditions (5% β-mercaptoethanol) before loading onto gels. Gels
were transferred to 0.2 µm pore-size polyvinyldene fluoride (PVDF) membrane using Trans-Blot
Turbo semi-dry transfer system (Bio-Rad). Blocking and antibody incubations took place in
blocking buffer (5% skim milk in TBS with 0.1% Tween 20). Primary antibodies used were:
1/3000 Piezo1 (Alomone); 1/500 tdTomato (Biorbyt); 1/4000 Actin (Santa Cruz Biotechnology);
1/1000 GAPDH (Santa Cruz Biotechnology). Secondary antibodies used were: 1/2000
horseradish peroxidase (HRP)-conjugated donkey anti-rabbit; 1/2000 HRP-conjugated donkey
anti-goat (both from Jackson Immunoresearch). Blots were imaged using a C-DiGit western blot
scanner (LI-COR).
2.2.10 Culture imaging
Experiments were performed in static baths (35 mm petri dishes) at room temperature with
HEPES-aCSF containing (in mM): NaCl, 140.4; HEPES, 10.8; KCl, 2.5; glucose, 10; MgCl2,
2.0; CaCl2, 2.0. HEPES buffer was used in place of NaHCO3 and NaH2PO4 was omitted to
prevent precipitation with gadolinium. Cells were imaged using a two-photon laser scanning
microscope (LSM 7 MP; Zeiss) with a 20x-W/1.0 numerical aperture objective lens coupled to a
Chameleon Ultra2 laser (Coherent). Images were acquired as time series of 256 x 256 pixels
using 8-line averaging with a frequency of 0.41 Hz. Lck-GCaMP5 was excited at 920 nm, and
the emission was detected by a photomultiplier tube after passing through a 500-550 nm filter.

41
2.2.11 Transgenic GLAST-GCaMP5 mice and tamoxifen treatment
Slc1a3(GLAST)-cre/ERT (Jax stock no. 012586) and PC-G5-tdT (Jax stock no. 024477) mice
were bred according to protocols approved by the University of British Columbia Animal Care
Committee. The resulting GLAST-GCaMP5 mice expressed the Ca2+ indicator GCaMP5 and the
red fluorescent protein tdTomato in astrocytes upon tamoxifen administration. Mice were
housed under standard conditions (12-hour light/dark cycle), and food and water were provided
ad libitum. Tamoxifen (Sigma) was dissolved in peanut oil (Sigma; 20 mg/mL), and mice
(postnatal day 30) were given three doses (200 mg/kg intraperitoneally) at 48-hour intervals.
Experiments were performed at least three weeks after the last tamoxifen injection.
2.2.12 Hippocampal slice preparation
Mice (postnatal day 50 to 60) were decapitated without anesthesia according to protocols
approved by the University of British Columbia Animal Care Committee. Brains were rapidly
extracted and submerged in ice-cold slicing solution saturated with carbogen (95% O2 / 5%
CO2). The slicing solution contained (in mM): N-methyl-D- glucamine, 120; KCl, 2.5;
NaH2PO4, 1.2; NaHCO3, 25; CaCl2, 1.0; MgCl2, 7.0; sodium pyruvate, 2.4; sodium ascorbate,
1.3; glucose, 20. Transverse hippocampal slices, 300 µm thick, were cut using a Leica VT
1200S vibratome and recovered in artificial cerebrospinal fluid (aCSF) saturated with carbogen
at room temperature for 1 hour. aCSF contained (in mM): NaCl, 126; NaHCO3, 26; KCl, 2.5;
glucose, 10; NaH2PO4, 1.25; MgCl2, 2.0; CaCl2, 2.0.

42
2.2.13 Slice imaging
Experiments were performed at room temperature with continuous perfusion (2.5 mL/min) of
HEPES-aCSF saturated with oxygen. Hippocampal slices were imaged using a two-photon laser
scanning microscope as described in section 2.2.10. Cells were imaged in the CA1 stratum
radiatum at depths between 70 – 90 µm. Frame images for time-lapse analysis were collected at
256 x 160 pixels using 8-line averaging with a frequency of 0.26 Hz. GCaMP5 and tdTomato
were excited at 920 nm, and signals were detected with photomultiplier tubes after passing
through 500-550 nm and 600-660 nm emission filters, respectively.
2.2.14 Image processing and data analysis
Time-lapse images of astrocyte culture Ca2+ signals were processed using ImageJ (NIH). Stacks
were aligned to remove drift in the x and y axes, and cell borders were delimited by hand.
Images were median filtered to remove noise, and thresholding was applied to identify the
signals from the background. Signals were then analyzed with the 3D components function in
MATLAB (MathWorks), which recognizes signals continuous in the x, y, and t axes. Large
spreading signals, such as Ca2+ waves, that exceeded the empirically-determined size threshold
of 1000 pixels were assigned a value of zero in a binary mask, and the mask was applied to the
aligned images to generate a stack containing only microdomain Ca2+ transients. Microdomain
regions of interest (ROIs; 6 x 6 pixels) were then detected based on maximum fluorescence
intensity, and the intensity profile of each ROI was baseline-corrected using the rolling ball filter.
Microdomain events were defined as signals larger than 10x the standard deviation of the ROI
baseline intensity, and parameters of frequency, amplitude, and duration were calculated and
binned over five-minute intervals. The analysis algorithm is summarized in Fig. 2-1.
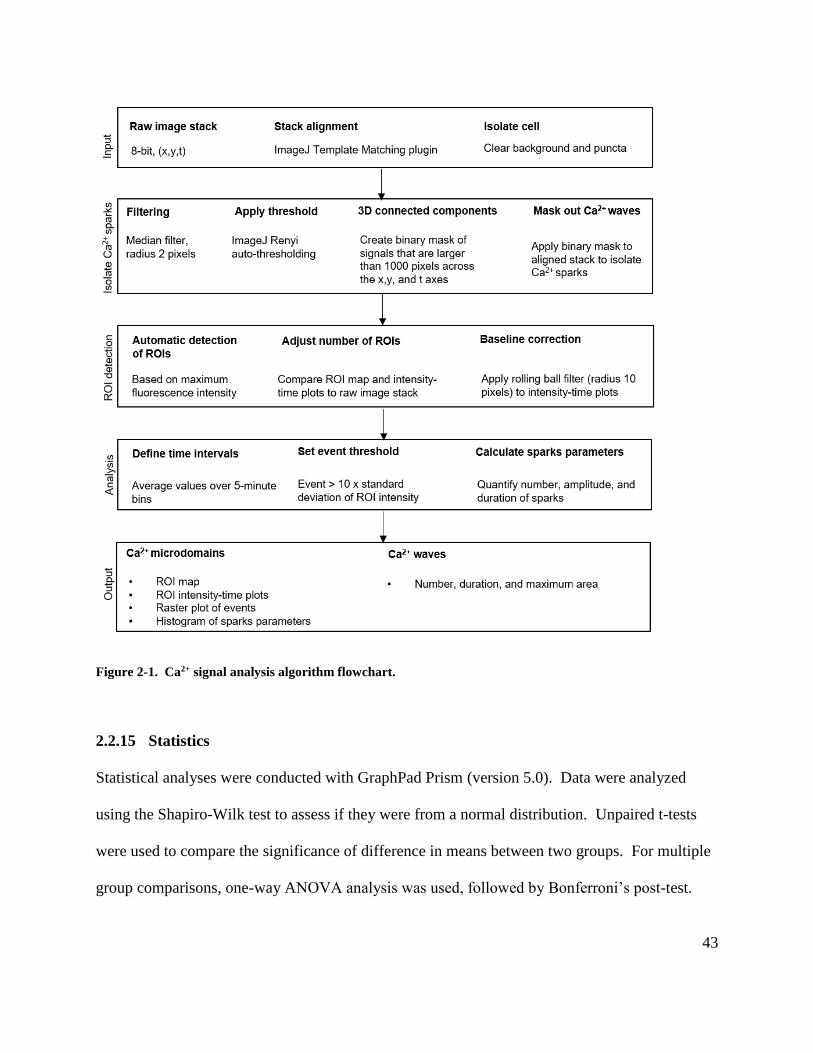
43
Figure 2-1. Ca2+ signal analysis algorithm flowchart.
2.2.15 Statistics
Statistical analyses were conducted with GraphPad Prism (version 5.0). Data were analyzed
using the Shapiro-Wilk test to assess if they were from a normal distribution. Unpaired t-tests
were used to compare the significance of difference in means between two groups. For multiple
group comparisons, one-way ANOVA analysis was used, followed by Bonferroni’s post-test.

44
When comparing different treatments conducted on the same cell, a paired t-test or repeated-
measures ANOVA analysis followed by Bonferroni’s post hoc test was performed. The n values
represent the number of cells imaged. Data are reported as mean ± 95% confidence interval, and
p < 0.05 was considered statistically significant.
2.3 Results
2.3.1 LNP delivery of plasmid DNA and siRNA in cultured astrocytes
To study the role of Piezo1 in mediating astrocyte Ca2+ signals, a safe and efficient strategy was
necessary for transfecting the Lck-GCaMP5 Ca2+ indicator into astrocytes. The LNP technology,
a system that packages molecules within inverted micelles of ionizable cationic lipids for
delivery into cells (Leung et al., 2012), was chosen as our lab had previously demonstrated its
effectiveness in delivering nucleic acids for both in vitro and in vivo applications in the brain
(Rungta et al., 2013, 2015). To determine whether LNPs are a feasible approach for transfecting
astrocytes with Lck-GCaMP5 plasmid DNA, we compared its cytotoxicity and transfection
efficiency with the commercially available transfection reagent Effectene, which had previously
been used to transfect cultured astrocytes with the Lck-GCaMP3 plasmid (Shigetomi et al.,
2012). Cytotoxicity, assessed by the number of adherent cells following transfection, was
quantified by counting the number of DAPI-positive cell nuclei per visual field, while
transfection efficiency was determined by quantifying the number of astrocytes with green
fluorescence from Lck-GCaMP5 expression (Fig. 2-2A). We found that LNP-packaged Lck-
GCaMP5 plasmid DNA transfection had no effect on cell viability (approximately 1000 cells
counted per coverslip; untreated: 100 ± 76%; LNP: 106 ± 31%; Effectene: 130 ± 70%; p > 0.05)

45
(Fig. 2-2B), with similar transfection efficiency achieved by both LNP and Effectene
(approximately 125 cells counted per coverslip; untreated: 0%; LNP: 10.6 ± 9.1%; Effectene: 9.2
± 10.3 %; p < 0.05 compared to untreated) (Fig. 2-2C).
Previous studies in our lab have shown that LNP-siRNA complexes can safely and effectively
knock down target genes in neurons (Rungta et al., 2013). Here we validate the use of LNP-
packaged Piezo1 siRNA in astrocytes by performing qPCR and immunoblotting to measure
mRNA and protein levels respectively. Using two probes targeted to different locations of the
Piezo1 sequence, we found that five-day treatment of astrocytes in cell culture with LNP-
packaged Piezo1 siRNA significantly reduced Piezo1 mRNA levels relative to the luciferase
siRNA-treated control (Probe 1, luciferase: 100.0 ± 4.2%; Probe 1, Piezo1: 18.2 ± 1.2%; Probe 2,
luciferase: 100 ± 5.6%; Probe 2, Piezo1: 15.6 ± 0.5%; p < 0.001 compared to luciferase
treatment) (Fig. 2-2D). Similarly, LNP-Piezo1 siRNA treatment knocked down Piezo1 protein
expression in cultured wild-type rat astrocytes relative to the untreated and luciferase siRNA
controls, and this knockdown was observed in the Piezo1-tdTomato mouse astrocytes as detected
by both the Piezo1 and tdTomato antibodies (Fig. 2-2E). The single and double band patterns
have been demonstrated in immunoblots of other rodent tissue samples, although it remains to be
determined whether the molecular weight differences are due to post-translational modifications
or splice variants (Andolfo et al., 2013; Ranade et al., 2014). Taken together, the data suggest
that LNPs are a safe and effective method for transfecting plasmid DNA and siRNA into
astrocyte cultures.
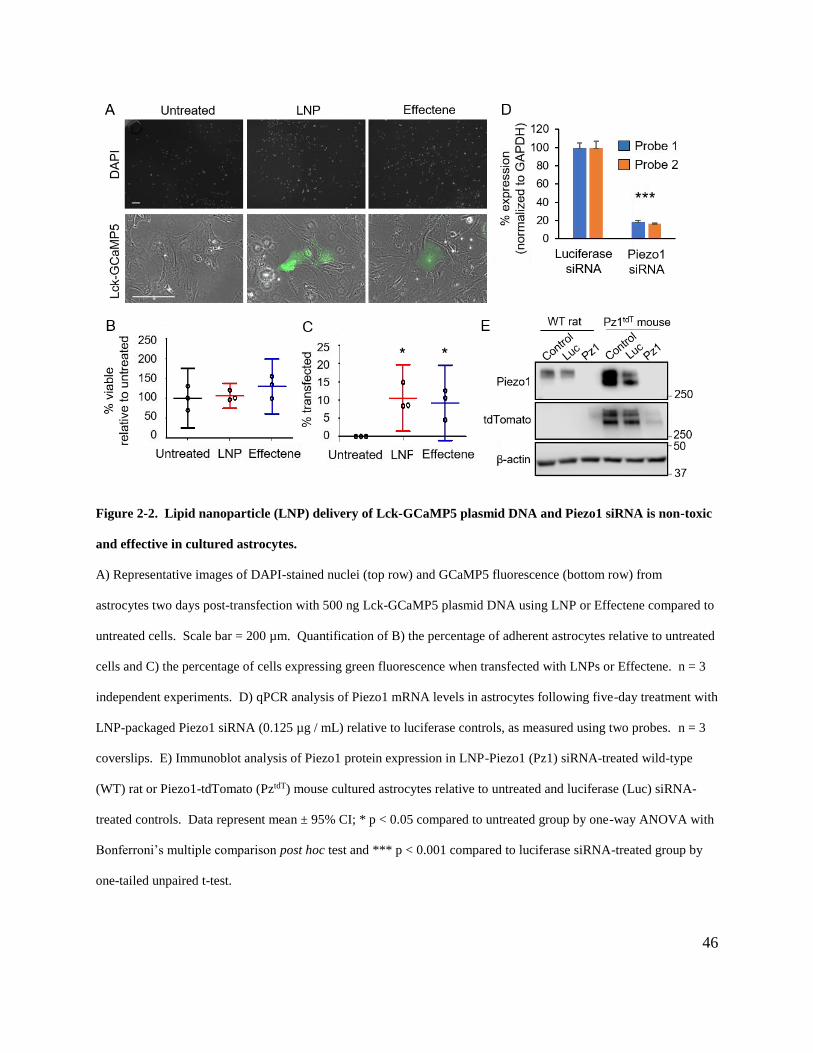
46
Figure 2-2. Lipid nanoparticle (LNP) delivery of Lck-GCaMP5 plasmid DNA and Piezo1 siRNA is non-toxic
and effective in cultured astrocytes.
A) Representative images of DAPI-stained nuclei (top row) and GCaMP5 fluorescence (bottom row) from
astrocytes two days post-transfection with 500 ng Lck-GCaMP5 plasmid DNA using LNP or Effectene compared to
untreated cells. Scale bar = 200 µm. Quantification of B) the percentage of adherent astrocytes relative to untreated
cells and C) the percentage of cells expressing green fluorescence when transfected with LNPs or Effectene. n = 3
independent experiments. D) qPCR analysis of Piezo1 mRNA levels in astrocytes following five-day treatment with
LNP-packaged Piezo1 siRNA (0.125 µg / mL) relative to luciferase controls, as measured using two probes. n = 3
coverslips. E) Immunoblot analysis of Piezo1 protein expression in LNP-Piezo1 (Pz1) siRNA-treated wild-type
(WT) rat or Piezo1-tdTomato (PztdT) mouse cultured astrocytes relative to untreated and luciferase (Luc) siRNA-
treated controls. Data represent mean ± 95% CI; * p < 0.05 compared to untreated group by one-way ANOVA with
Bonferroni’s multiple comparison post hoc test and *** p < 0.001 compared to luciferase siRNA-treated group by
one-tailed unpaired t-test.

47
2.3.2 Development of a novel method to analyze astrocyte Ca2+ signals
Astrocytes exhibit various types of Ca2+ signals with distinct spatial and temporal characteristics
that reflect different mechanisms of Ca2+ release or entry (Srinivasan et al., 2015). In cultured
astrocytes transfected with the Lck-GCaMP5 construct, we observed at least two distinct patterns
of spontaneous Ca2+ signals- Ca2+ waves with long duration that appeared to expand and contract
from their point of origin, and Ca2+ transients that were spatially restricted in microdomains (Fig.
2-3A). Since Ca2+ waves and transients overlap in the xy-plane, we developed an algorithm to
distinguish and independently analyze these signals.
A key feature of the algorithm involves using the MATLAB 3D connected components function
to identify Ca2+ signals that are continuous in the x, y, and t axes, allowing the signals to be
classified based on total area (Fig. 2-3B). The masking threshold was set at 1000 pixels (1186
µm2) empirically to separate Ca2+ microdomain events from the Ca2+ waves (Fig. 2-3C). The
algorithm then identifies microdomain regions of interest (ROIs) based on fluorescence intensity
(Fig. 2-3D), and generates intensity-time course traces (Fig. 2-3E) and raster plots (Fig. 2-3F) of
the Ca2+ signals detected for each ROI. Parameters such as event frequency, intensity, and
duration can then be quantified to assess the effect of different interventions on the microdomain
transients selectively.
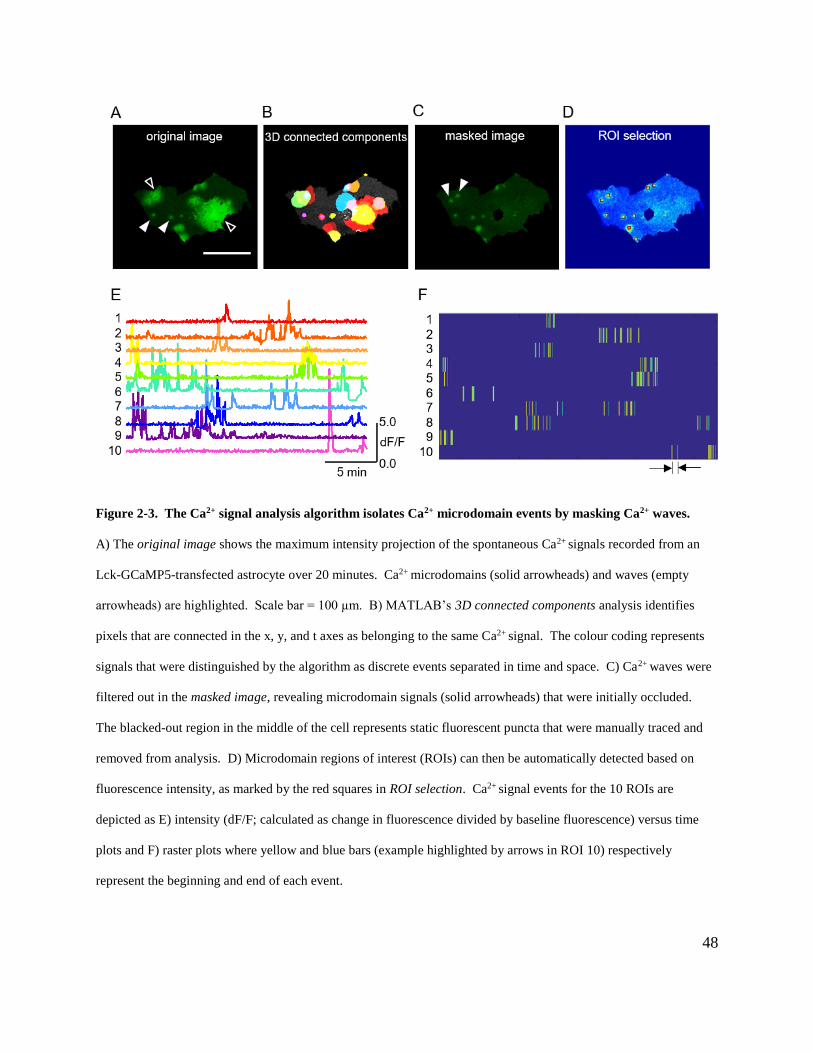
48
Figure 2-3. The Ca2+ signal analysis algorithm isolates Ca2+ microdomain events by masking Ca2+ waves.
A) The original image shows the maximum intensity projection of the spontaneous Ca2+ signals recorded from an
Lck-GCaMP5-transfected astrocyte over 20 minutes. Ca2+ microdomains (solid arrowheads) and waves (empty
arrowheads) are highlighted. Scale bar = 100 µm. B) MATLAB’s 3D connected components analysis identifies
pixels that are connected in the x, y, and t axes as belonging to the same Ca2+ signal. The colour coding represents
signals that were distinguished by the algorithm as discrete events separated in time and space. C) Ca2+ waves were
filtered out in the masked image, revealing microdomain signals (solid arrowheads) that were initially occluded.
The blacked-out region in the middle of the cell represents static fluorescent puncta that were manually traced and
removed from analysis. D) Microdomain regions of interest (ROIs) can then be automatically detected based on
fluorescence intensity, as marked by the red squares in ROI selection. Ca2+ signal events for the 10 ROIs are
depicted as E) intensity (dF/F; calculated as change in fluorescence divided by baseline fluorescence) versus time
plots and F) raster plots where yellow and blue bars (example highlighted by arrows in ROI 10) respectively
represent the beginning and end of each event.

49
2.3.3 Piezo1 activation selectively increases the frequency of microdomain Ca2+ signals
To determine if Piezo1 is involved in mediating Ca2+ signals in astrocytes, we analyzed the effect
of the Piezo1 agonist, Yoda1, on Ca2+ waves and microdomain transients in LNP-Lck-GCaMP5-
transfected cells. Yoda1 was applied at 1 µM, corresponding to the concentration found to
induce single-channel currents in Piezo1 channels reconstituted in artificial bilayers (Syeda et al.,
2015). The maximum intensity projection images from the 3D connected components analysis
show that discrete Ca2+ wave and microdomain signals occurred both spontaneously at baseline
and following Yoda1 treatment (Fig. 2-4A). Yoda1-mediated Piezo1 activation did not affect
astrocyte Ca2+ waves as monitored by frequency (baseline: 1.4 ± 1.2 events/10 min; Yoda1: 1.7 ±
1.1 events/10 min; p > 0.05) (Fig. 2-4B), maximum area of spread in the xy-plane (baseline:
2100 ± 2200 µm2; Yoda1: 1000 ± 520 µm2; p > 0.05)(Fig. 2-4C), and duration (baseline: 98 ± 65
s; Yoda1: 110 ± 76 s; p > 0.05)(Fig. 2-4D). In contrast, Yoda1 increased the frequency of
microdomain signals (baseline: 0.98 ± 0.77 events/ROI/5 min; Yoda1: 3.3 ± 1.2 events/ROI/5
min; p < 0.01)(Fig. 2-4E) without altering their amplitude (baseline: 1.5 ± 0.6 a.u.; Yoda1: 1.5 ±
0.3 a.u.; p > 0.05)(Fig. 2-4F) or duration (baseline: 6.7 ± 4.0 s; Yoda1: 7.1 ± 2.1 s; p > 0.05)(Fig.
2-4G).
Our results suggest that Piezo1 activation in astrocytes causes localized increases in Ca2+
concentration proximal to the plasma membrane, as detected by the membrane-targeted Lck-
GCaMP5 indicator. Furthermore, the selective effect of Yoda1 supports the interpretation that
Ca2+ waves and microdomain transients are physiologically distinct, and validates the importance
of analyzing these signals separately. We focused on the Ca2+ signal frequency in subsequent
experiments, as this was the only parameter that was affected by Yoda1 application.
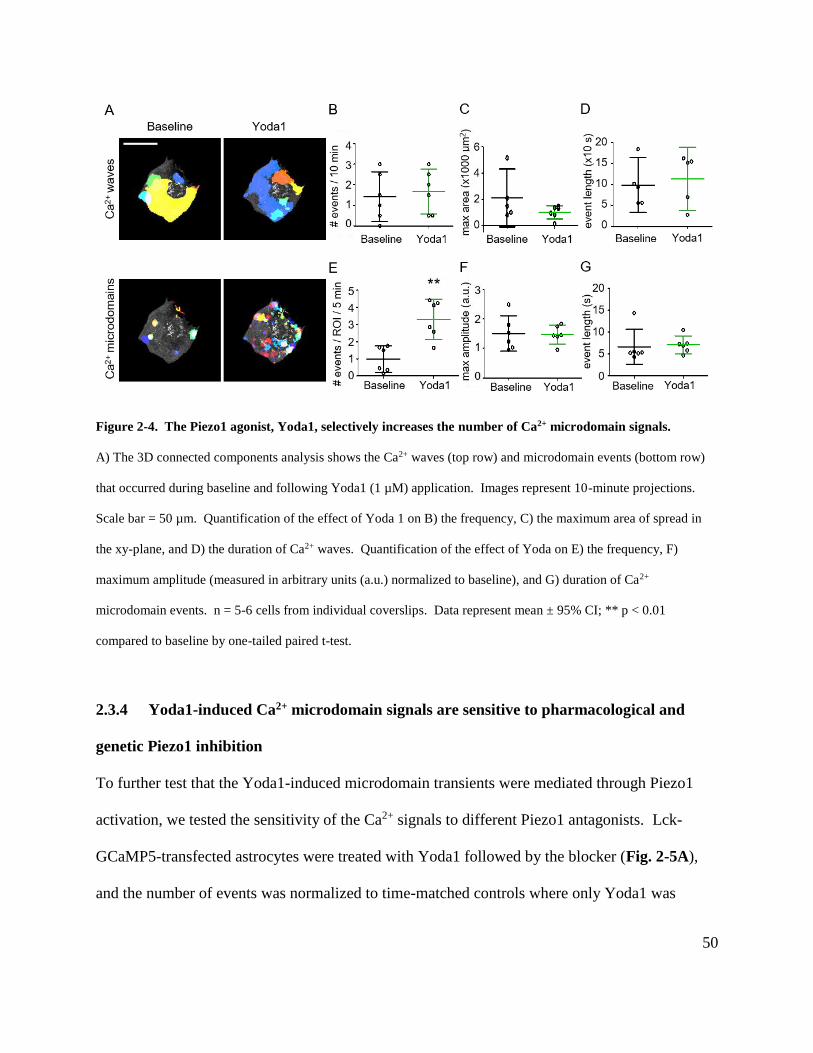
50
Figure 2-4. The Piezo1 agonist, Yoda1, selectively increases the number of Ca2+ microdomain signals.
A) The 3D connected components analysis shows the Ca2+ waves (top row) and microdomain events (bottom row)
that occurred during baseline and following Yoda1 (1 µM) application. Images represent 10-minute projections.
Scale bar = 50 µm. Quantification of the effect of Yoda 1 on B) the frequency, C) the maximum area of spread in
the xy-plane, and D) the duration of Ca2+ waves. Quantification of the effect of Yoda on E) the frequency, F)
maximum amplitude (measured in arbitrary units (a.u.) normalized to baseline), and G) duration of Ca2+
microdomain events. n = 5-6 cells from individual coverslips. Data represent mean ± 95% CI; ** p < 0.01
compared to baseline by one-tailed paired t-test.
2.3.4 Yoda1-induced Ca2+ microdomain signals are sensitive to pharmacological and
genetic Piezo1 inhibition
To further test that the Yoda1-induced microdomain transients were mediated through Piezo1
activation, we tested the sensitivity of the Ca2+ signals to different Piezo1 antagonists. Lck-
GCaMP5-transfected astrocytes were treated with Yoda1 followed by the blocker (Fig. 2-5A),
and the number of events was normalized to time-matched controls where only Yoda1 was

51
applied. We first tested GsMTx4, a toxin purified from the spider Grammostola spatulata that
blocked mechanosensitive currents in Piezo1-transfected HEK cells (Bae et al., 2011). As shown
in Fig. 2-5B, Yoda1-evoked microdomain Ca2+ transients were partially inhibited by GsMTx4
(20 µM) (Yoda1: 100 ± 29%; Yoda1 + GsMTx4: 54 ± 19%; p < 0.05). Application of Gd3+ (100
µM), a non-specific blocker of stretch-activated channels (Hamill and McBride, 1996), also
partially inhibited the signals (Yoda1 + Gd3+: 37 ± 45%; p < 0.05). As a negative control, we
tested HC067047 (10 µM), a selective inhibitor of the Ca2+-permeable mechanosensitive TRPV4
channel expressed in astrocytes (Benfenati et al., 2007; Dunn et al., 2013) and found it had no
effect on the frequency of Ca2+ signals (Yoda1 + HC067047: 77 ± 20%; p > 0.05).

52
Figure 2-5. Yoda1-induced increases in Ca2+ microdomain signals are partially inhibited by Piezo1
antagonists GsMTx4 and Gd3+.
A) Images represent masked 10-minute maximum intensity projections of astrocytes transfected with Lck-GCaMP5
during baseline, following Yoda1 (1 µM) application, and after subsequent addition of either GsMTx4 (20 µM),
Gd3+ (100 µM) or HC067047 (10 µM). Scale bars = 10 µm. B) Quantification of the effects of the antagonists on
Yoda1-induced Ca2+ microdomain transients. n = 6 – 22 cells from individual coverslips. Data represent mean ±
95% CI; * p < 0.05 compared to Yoda1 by one-way ANOVA with Bonferroni’s multiple comparison post hoc test.
Since pharmacological blockers can have non-specific effects, we further studied the
involvement of Piezo1 in Yoda1-induced microdomain signals using an siRNA approach to
knock down Piezo1 expression. Astrocyte cultures were treated with LNP-packaged siRNA
(0.125 µg/mL) for five days based on the mRNA and protein knockdown data (Fig. 2-2D and
E). Yoda1-induced Ca2+ microdomain transients were significantly reduced in Piezo1 siRNA-
treated astrocytes relative to the control luciferase siRNA condition (luciferase siRNA baseline:
0.3 ± 0.3 events/ROI/5 min; luciferase siRNA Yoda1: 5.0 ± 1.5 events/ROI/5 min; p < 0.001;
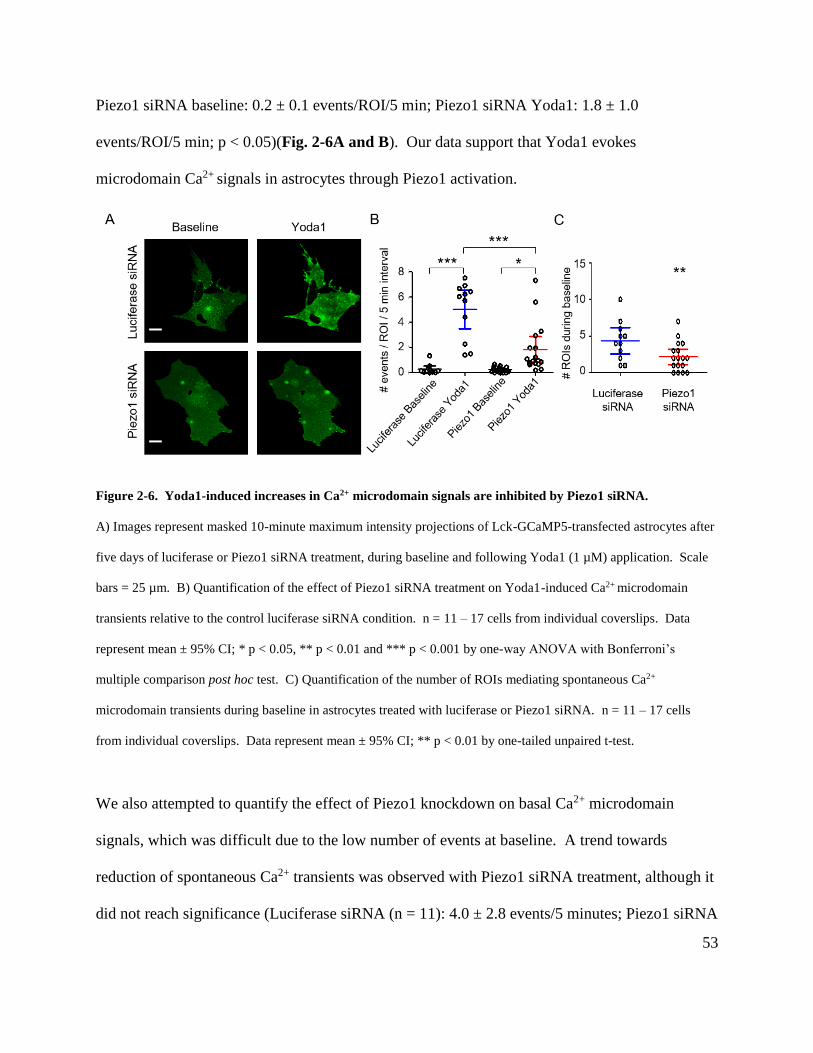
53
Piezo1 siRNA baseline: 0.2 ± 0.1 events/ROI/5 min; Piezo1 siRNA Yoda1: 1.8 ± 1.0
events/ROI/5 min; p < 0.05)(Fig. 2-6A and B). Our data support that Yoda1 evokes
microdomain Ca2+ signals in astrocytes through Piezo1 activation.
Figure 2-6. Yoda1-induced increases in Ca2+ microdomain signals are inhibited by Piezo1 siRNA.
A) Images represent masked 10-minute maximum intensity projections of Lck-GCaMP5-transfected astrocytes after
five days of luciferase or Piezo1 siRNA treatment, during baseline and following Yoda1 (1 µM) application. Scale
bars = 25 µm. B) Quantification of the effect of Piezo1 siRNA treatment on Yoda1-induced Ca2+ microdomain
transients relative to the control luciferase siRNA condition. n = 11 – 17 cells from individual coverslips. Data
represent mean ± 95% CI; * p < 0.05, ** p < 0.01 and *** p < 0.001 by one-way ANOVA with Bonferroni’s
multiple comparison post hoc test. C) Quantification of the number of ROIs mediating spontaneous Ca2+
microdomain transients during baseline in astrocytes treated with luciferase or Piezo1 siRNA. n = 11 – 17 cells
from individual coverslips. Data represent mean ± 95% CI; ** p < 0.01 by one-tailed unpaired t-test.
We also attempted to quantify the effect of Piezo1 knockdown on basal Ca2+ microdomain
signals, which was difficult due to the low number of events at baseline. A trend towards
reduction of spontaneous Ca2+ transients was observed with Piezo1 siRNA treatment, although it
did not reach significance (Luciferase siRNA (n = 11): 4.0 ± 2.8 events/5 minutes; Piezo1 siRNA

54
(n = 17): 2.9 ± 1.5 events/5 minutes; p > 0.05). However, the number of ROIs that mediated the
spontaneous microdomain Ca2+ signals was significantly decreased by Piezo1 siRNA treatment
(Luciferase siRNA: 4.4 ± 1.8 ROIs; Piezo1 siRNA: 2.2 ± 1.0 ROIs; p < 0.01)(Fig. 2-6C). The
data suggest that Piezo1 siRNA knockdown reduces the number of basally active Ca2+
microdomains in astrocytes.
2.3.5 Yoda1-induced Ca2+ microdomain signals depend on extracellular Ca2+ entry
To elucidate the Ca2+ source underlying Yoda1-induced microdomain transients, we first tested
the effect of chelating intracellular Ca2+ with BAPTA-AM (1 µM). As shown in Fig. 2-7A, Ca2+
signals were completely inhibited by BAPTA-AM, consistent with the intracellular localization
of the Lck-GCaMP5 indicator (Yoda1: 3.5 ± 1.5 events/ROI/5 min; BAPTA-AM: 0
events/ROI/5 min; p < 0.05). Yoda1-evoked transients were also blocked in the absence of
extracellular Ca2+ (aCSF containing 0 mM Ca2+ and 2 mM EGTA)(0 Ca2+/EGTA: 0
events/ROI/5 min; p < 0.01 compared to Yoda1 alone). In contrast, depleting intracellular Ca2+
stores by blocking SERCA with thapsigargin (1 µM) had no effect (thapsigargin: 5.2 ± 1.8
events/ROI/5min; p > 0.05 compared to Yoda1 alone). The results from sections 2.3.4 and 2.3.5
are summarized in the schematics in Fig. 2-7B, which illustrate Yoda1-induced Ca2+
microdomain signals in astrocytes occur via Ca2+ influx through the Piezo1 channel.
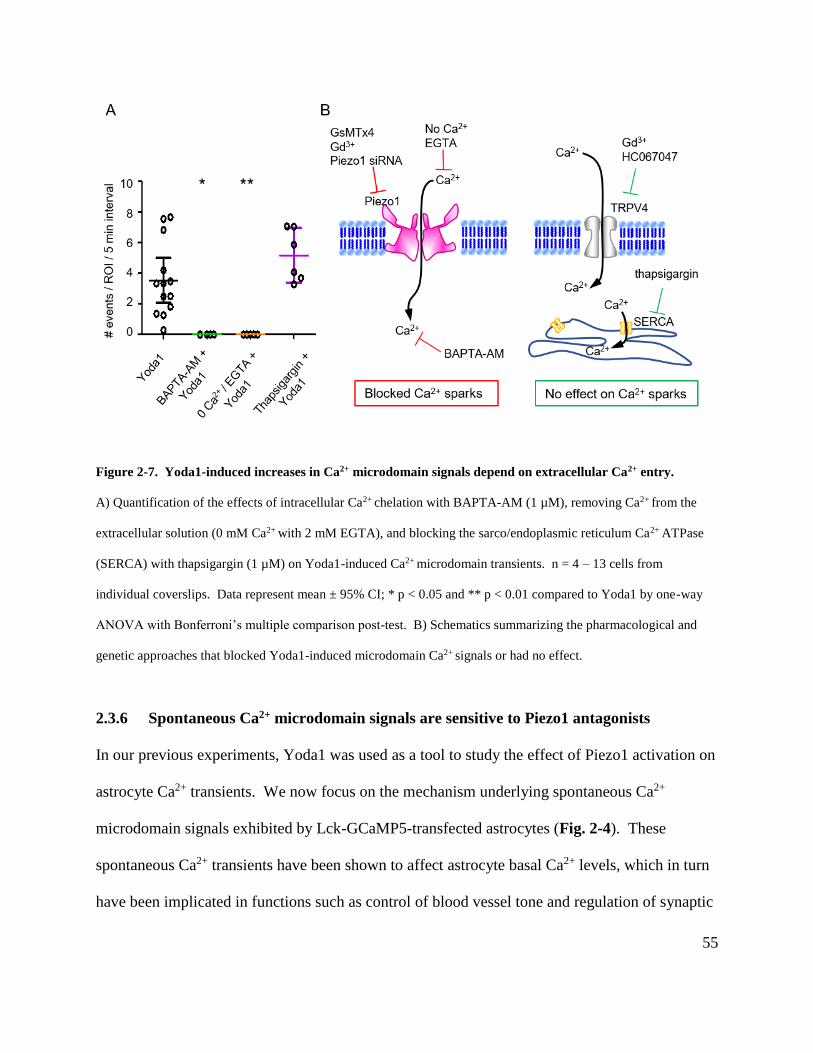
55
Figure 2-7. Yoda1-induced increases in Ca2+ microdomain signals depend on extracellular Ca2+ entry.
A) Quantification of the effects of intracellular Ca2+ chelation with BAPTA-AM (1 µM), removing Ca2+ from the
extracellular solution (0 mM Ca2+ with 2 mM EGTA), and blocking the sarco/endoplasmic reticulum Ca2+ ATPase
(SERCA) with thapsigargin (1 µM) on Yoda1-induced Ca2+ microdomain transients. n = 4 – 13 cells from
individual coverslips. Data represent mean ± 95% CI; * p < 0.05 and ** p < 0.01 compared to Yoda1 by one-way
ANOVA with Bonferroni’s multiple comparison post-test. B) Schematics summarizing the pharmacological and
genetic approaches that blocked Yoda1-induced microdomain Ca2+ signals or had no effect.
2.3.6 Spontaneous Ca2+ microdomain signals are sensitive to Piezo1 antagonists
In our previous experiments, Yoda1 was used as a tool to study the effect of Piezo1 activation on
astrocyte Ca2+ transients. We now focus on the mechanism underlying spontaneous Ca2+
microdomain signals exhibited by Lck-GCaMP5-transfected astrocytes (Fig. 2-4). These
spontaneous Ca2+ transients have been shown to affect astrocyte basal Ca2+ levels, which in turn
have been implicated in functions such as control of blood vessel tone and regulation of synaptic

56
activity via neurotransmitter uptake (Shigetomi et al., 2012; Rosenegger et al., 2015). We tested
whether spontaneous Ca2+ signals have a Piezo1-mediated component using the same
pharmacological tools as in section 2.3.4. The Piezo1 antagonists GsMTx4 (20 µM) and Gd3+
(100 µM) decreased the basal Ca2+ signals (Baseline: 0.8 ± 0.2 events/ROI/5 min; GsMTx4: 0.5
± 0.3 events/ROI/5 min; p < 0.01; Baseline: 1.2 ± 0.9 events/ROI/5 min; Gd3+: 0.2 ± 0.2
events/ROI/5 min; p < 0.05)(Fig. 2-8A – C), while HC067047 (10 µM) had no effect (Baseline:
1.4 ± 1.2 events/ROI/5 min; HC067047: 0.9 ± 0.5 events/ROI/5 min; p > 0.05)(Fig. 2-8A and
D). Together with the observation that Piezo1 siRNA reduces the number of basally active Ca2+
microdomains (Fig. 2-6C), the data suggest that Piezo1 may contribute to astrocyte spontaneous
Ca2+ transients.
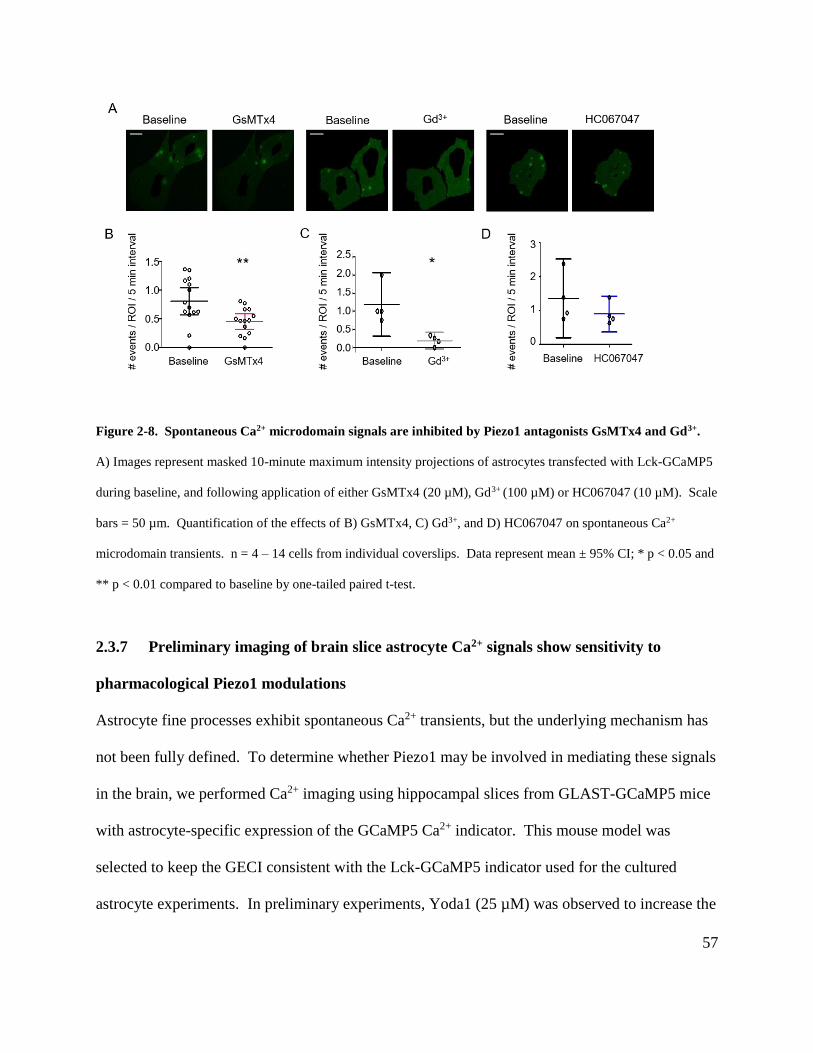
57
Figure 2-8. Spontaneous Ca2+ microdomain signals are inhibited by Piezo1 antagonists GsMTx4 and Gd3+.
A) Images represent masked 10-minute maximum intensity projections of astrocytes transfected with Lck-GCaMP5
during baseline, and following application of either GsMTx4 (20 µM), Gd3+ (100 µM) or HC067047 (10 µM). Scale
bars = 50 µm. Quantification of the effects of B) GsMTx4, C) Gd3+, and D) HC067047 on spontaneous Ca2+
microdomain transients. n = 4 – 14 cells from individual coverslips. Data represent mean ± 95% CI; * p < 0.05 and
** p < 0.01 compared to baseline by one-tailed paired t-test.
2.3.7 Preliminary imaging of brain slice astrocyte Ca2+ signals show sensitivity to
pharmacological Piezo1 modulations
Astrocyte fine processes exhibit spontaneous Ca2+ transients, but the underlying mechanism has
not been fully defined. To determine whether Piezo1 may be involved in mediating these signals
in the brain, we performed Ca2+ imaging using hippocampal slices from GLAST-GCaMP5 mice
with astrocyte-specific expression of the GCaMP5 Ca2+ indicator. This mouse model was
selected to keep the GECI consistent with the Lck-GCaMP5 indicator used for the cultured
astrocyte experiments. In preliminary experiments, Yoda1 (25 µM) was observed to increase the

58
frequency of Ca2+ signals in the astrocyte endfeet (Fig. 2-9A), while Gd3+ (100 µM) application
inhibited the transients (Fig. 2-9B). These results are consistent with the pharmacological profile
of the Ca2+ signals observed in the Lck-GCaMP5-transfected astrocyte cultures, suggesting that
Piezo1 contributes to spontaneous Ca2+ transients in the intact brain.
Figure 2-9. The frequency of astrocyte endfeet Ca2+ transients are sensitive to pharmacological modulators of
Piezo1.
A) Two-photon imaging of brain slices from mice with inducible astrocyte-specific expression of GCaMP5 showing
the effect of Yoda1 (25 µM) and B) Gd3+ (100 µM) on the frequency of Ca2+ transients in the endfeet, as highlighted
with the dashed circles. The representative baseline, Yoda1, and Gd3+ images correspond to the black, green, and
grey arrowheads, respectively, on the intensity-time traces to the right. Scale bar = 10 µm.

59
2.4 Discussion
In these studies, we demonstrated that Piezo1 channels contribute to microdomain Ca2+ signals in
cultured astrocytes, suggesting that Piezo1 may be physiologically important in regulating
astrocyte basal Ca2+ level and function. The Piezo1 agonist Yoda1 increased the number of
microdomain Ca2+ transients, which were completely abolished by removal of extracellular Ca2+
but unaffected by depletion of intracellular Ca2+ stores with thapsigargin. In addition, both
Yoda1-evoked and spontaneous signals were inhibited by the Piezo1 antagonists GsMTx4 and
Gd3+. The pharmacological data were further supported by the sensitivity of Yoda1-induced
Ca2+ signals to Piezo1 siRNA treatment. Furthermore, we performed preliminary imaging
experiments using brain slices from GLAST-GCaMP5 mice, and our observations suggest that
astrocyte endfeet Ca2+ transients may also involve a Piezo1-mediated component.
Numerous chemical, physical, and viral strategies have been developed and refined to maximize
transfection efficiency while preserving cell viability (Gresch and Altrogge, 2012).
Differentiated mammalian cells in primary culture are often difficult to transfect, with
transfection efficiency rates of ~3% previously reported for liposome-mediated transfection of
primary astrocytes (Wu et al., 2000). In our experiments, we utilized LNP technology for
delivery of both plasmid DNA and siRNA into cultured astrocytes due to its advantages in ease
of preparation, low immunogenicity, and adaptability for in vivo gene delivery and knockdown
(Nayerossadat et al., 2012; Rungta et al., 2013, 2015; Zatsepin et al., 2016). In preliminary
control experiments, we found LNPs were well-tolerated and equally effective as the non-
liposomal transfection reagent Effectene in delivering plasmid DNA to astrocytes (Shigetomi et
al., 2012), with transfection efficiency rate ~10%. We also verified that LNP-packaged Piezo1

60
siRNA was effective in silencing mRNA and reducing protein expression in astrocytes, as we
have previously shown for neurons (Rungta et al., 2013). While multiple approaches are
available for gene knockdown in the brain (Heidenreich and Zhang, 2016), an siRNA approach
was selected for its simplicity in generating sequences against specific targets, ease of delivery
into cells, and relatively low cost. Furthermore, effective Piezo1 knockdown by siRNA has been
demonstrated in various cell types (Coste et al., 2010; McHugh et al., 2010; Lee et al., 2014;
Pathak et al., 2014).
Studies on Piezo1 activity as a cationic channel have relied on patch clamp electrophysiological
techniques. However, these methods inherently apply a mechanical force to the cell, resulting in
alterations of membrane tension or cytoskeletal structure that may affect the properties of
mechanosensitive channels (Nourse and Pathak, 2017). In contrast, several groups have used
Ca2+ imaging as a non-invasive technique to measure Piezo1 function. For instance, Piezo1-
mediated intracellular Ca2+ elevations have been recorded in Piezo1-transfected HEK293 cells
and chondrocytes in response to mechanical deformation of the plasma membrane (Lee et al.,
2014; Gaub and Müller, 2017). Intriguingly, Pathak et al. (2014) observed Piezo1-dependent
spontaneous Ca2+ microdomain signals in neural stem cells that were loaded with Fluo-4 and
imaged with total internal reflection fluorescence microscopy (TIRFM), which allowed the
authors to selectively measure signals at the plasma membrane.
Alternatively, we used the membrane-tethered GECI, Lck-GCaMP5, to monitor near-membrane
Ca2+ microdomain signals in cultured astrocytes. While patch-dialysis of organic dyes is a viable
strategy for recording microdomain Ca2+ signals in fine astrocyte processes, cultured astrocytes

61
have a polygonal morphology that lacks the highly branched processes characteristic of in vivo
astrocytes. The Lck domain consists of tandem palmitoylation and myristoylation domains from
the N-terminal of a Src tyrosine kinase, which targets the GCaMP5 to the plasma membrane
(Shigetomi et al., 2010). Using such Lck-linked constructs, Shigetomi et al. (2010) previously
demonstrated that both global and localized Ca2+ signals could be detected in cultured astrocytes.
Furthermore, while GECIs are Ca2+ buffers that may alter endogenous Ca2+ homeostasis, no
studies to date have been published on GECI-induced alterations to astrocyte physiology
(Shigetomi et al., 2016). Importantly, the microdomain Ca2+ signals reported here share a
number of characteristics with the transients recorded from the fine processes of brain slice and
in vivo astrocytes, including dependence on extracellular Ca2+ and resistance to treatments that
inhibit Ca2+ release from intracellular stores (Shigetomi et al., 2010; Srinivasan et al., 2015;
Rungta et al., 2016).
Initial studies of astrocyte Ca2+ signaling relied on measuring large, robust responses in the soma
(Volterra et al., 2014). However, the discovery of microdomain Ca2+ transients, with different
spatial and temporal characteristics, presented a new challenge for analysis. Early work on
developing algorithms to detect and quantify Ca2+ transients was conducted in muscle cells,
where Ca2+ release from intracellular stores appeared as ‘sparks’ in confocal line scan images
(Cheng et al., 1999). Researchers recognized the need for an automatic analysis program to
quantify many events, and to prevent detection bias against events with low signal-to-noise ratio
(Cheng et al., 1999; Picht et al., 2007). In the astrocyte Ca2+ imaging field, algorithms such as
automatic segmentation, GECIquant, and CaSCaDe have been developed to analyze

62
microdomain signals (Di Castro et al., 2011; Srinivasan et al., 2015; Agarwal et al., 2017;
Bindocci et al., 2017).
Here we developed a semi-automatic method to quantify astrocyte Ca2+ microdomain signals.
The analysis algorithm shares several similarities, such as image alignment to eliminate x-y axis
drift, median filtering to remove noise, separation of microdomain and wave signals based on
area, and thresholding to identify microdomains events. One key feature is our use of the 3D
connected components function in MATLAB, which allows discrete Ca2+ signals to be identified
across time. Hence, Ca2+ waves that expanded and contracted from their point of origin could be
masked effectively, revealing microdomain signals that occupied the overlapping space in the
xy-plane. Intracellular Ca2+ wave events and localized microdomain transients were analyzed
separately due to differences in their spatial and temporal characteristics, as well as their Ca2+
source. Previous studies have found that intrinsic somatic Ca2+ waves have a greater dependence
on IP3R-mediated release from endoplasmic reticulum (ER) stores, and the extent of their spread
may be determined by the spatial distribution of IP3Rs in the ER (Khakh and McCarthy, 2015;
Srinivasan et al., 2015). In contrast, microdomain Ca2+ signals involve a transmembrane Ca2+
entry pathway rather than an IP3R-mediated mechanism (Shigetomi et al., 2012; Rungta et al.,
2016), consistent with the observation that fine astrocyte processes have little ER content
(Verkhratsky and Nedergaard, 2014). Using the Piezo1 agonist Yoda1, we were able to
selectively increase the frequency of microdomain transients, underscoring the need to consider
different types of Ca2+ signals separately.

63
Analysis of cultured astrocytes showed that spontaneous microdomain Ca2+ events occurred with
a frequency of 0.20 ± 0.06 events/ROI/min (mean ± SEM), comparable to the 0.5 ± 0.1 and ~0.4
events/ROI/min reported by other groups from imaging astrocytes in brain slices (Srinivasan et
al., 2015; Agarwal et al., 2017). Furthermore, we observed that microdomain transients
exhibited slow kinetics with an average duration of 6.7 ± 1.5 s (mean ± SEM), which is
consistent with the previously reported duration of 8.5 ± 1.7 s (Agarwal et al., 2017). The
convergence of the parameters across different models, Ca2+ indicators, and analysis algorithms
suggests our methods for imaging and quantifying Ca2+ microdomain signals are reliable.
The data suggest a role for Piezo1 in mediating spontaneous Ca2+ microdomain transients, in
agreement with previous studies of reconstituted Piezo1 channels that identified Piezo1-mediated
currents in the absence of applied pressure (Syeda et al., 2015). Yoda1 induced an increase in
the frequency of Ca2+ signals, consistent with reports that Yoda1 lowers the threshold for Piezo1
activation (Syeda et al., 2015). Yoda1 has also been shown to slow Piezo1 inactivation kinetics
(Syeda et al., 2015); however, we did not observe any changes in the duration of the Ca2+
transients, possibly because our scanning rate of 2.4 s/frame was too slow to capture any
differences. Our finding that Yoda1-evoked microdomain Ca2+ signals are dependent on Ca2+
entry recapitulates other studies demonstrating that such localized Ca2+ transients in astrocytes
are mediated by a transmembrane Ca2+ pathway rather than by release from intracellular stores
(Srinivasan et al., 2015; Rungta et al., 2016), and suggests that subcellular compartmentalization
of Ca2+ signaling is preserved in cultured astrocytes.

64
While our work suggests that Piezo1 is involved in mediating astrocyte Ca2+ microdomain
signals, it is interesting to note that only ~40% of the spontaneous signals were blocked by the
Piezo1 antagonist GsMTx4, while ~80% block was attained with the nonspecific
mechanosensitive channel blocker Gd3+. Furthermore, siRNA-mediated Piezo1 knockdown
reduced the number of Ca2+ microdomains but did not completely abolish the signals. This
suggests that in addition to Piezo1, other Ca2+-fluxing pathways, such as TRPA1 (Shigetomi et
al., 2012), are likely to contribute to the Ca2+ transients. Another candidate is Ca2+-induced Ca2+
release through the mitochondrial permeability transition pore (mPTP); Agarwal et al. (2017)
found that mPTP block was associated with a 70% reduction in the frequency of Ca2+ transients.
These transporters or channels could act in concert or contribute in different proportions under
different physiological conditions to mediate the spontaneous Ca2+ transients.

65
Chapter 3: Subcellular localization of Piezo1 in astrocytes
3.1 Overview
As reported in the previous Chapter, pharmacological and genetic manipulation of Piezo1 in
astrocytes affects the frequency of microdomain Ca2+ transients, suggesting that astrocytes
express functional Piezo1 channels. This finding is supported by recent transcriptomic and
proteomic studies of both cultured and acutely isolated rodent astrocytes demonstrating Piezo1
mRNA and protein expression (Zhang et al., 2014; Sharma et al., 2015; Srinivasan et al., 2016).
While our data point to the presence of Piezo1 at the plasma membrane to mediate
transmembrane Ca2+ influx, the channel may also be expressed in other cellular domains that are
relevant for mechanotransduction.
Li et al. (2014) observed green fluorescent protein-tagged Piezo1 (Piezo1-GFP) localized to
cytoskeleton-associated focal adhesion zones in cultured human umbilical vein endothelial cells,
where the channels may sense shear stress. The authors further suggested that this association
with the cytoskeleton may be related to the role of Piezo1 in mediating Ca2+ entry to activate
calpain-dependent reorganization of the cytoskeleton for proper endothelial cell alignment.
Furthermore, McHugh et al. (2010) reported colocalization of GFP-tagged Piezo1 or Piezo1
immunoreactivity with the endoplasmic reticulum (ER) marker SERCA2 in Chinese hamster
ovary and HeLa cells, and suggested Piezo1 involvement in recruiting R-Ras GTPase to the ER,
followed by Ca2+ release from ER stores and subsequent activation of integrin to mediate cell
adhesion. Taken together, the studies reveal that while cell type-specific Piezo1 expression
pattern in different organelles can mediate a diverse range of functions, there appears to be a

66
common association with structural locations related to mechanosensing and
mechanotransduction. Of note, astrocytes have been shown to express mechanosensitive
channels (Bowman et al., 1992), and the cells exhibit changes in morphology and function in
response to stimuli such as extracellular matrix stiffness (Summers et al., 2010; Wilson et al.,
2016). Therefore, it is interesting to study the subcellular localization of Piezo1 as it can provide
information about the protein’s interaction partners and physiological roles in astrocytes (Stadler
et al., 2013).
In this chapter, we provide evidence for Piezo1 association with the plasma membrane and
cytoskeletal components in astrocytes. Subcellular localization of Piezo1 was determined by
optimized immunocytochemistry protocols, expression of exogenous fluorescent protein –
tagged Piezo1, and verified by siRNA-mediated Piezo1 knockdown. To further investigate the
link between Piezo1 and the cytoskeleton, astrocytes were treated with forskolin to induce
cytoskeleton reorganization followed by immunostaining. Finally, we performed preliminary
immunohistochemistry experiments in fixed brain slices to assess Piezo1 localization in
astrocytes in situ.
3.2 Methods
3.2.1 Cell culture
Astrocyte cultures were prepared as described in section 2.2.2. For staining experiments, P1
astrocytes were thawed, resuspended in glial medium, and plated onto sterile 15 mm No. 1.5
glass coverslips (MatTek) at a density of ~9 x 103 cells per coverslip. For immunoblotting

67
experiments, cells were seeded at a density of 4.5 x 104 cells per T25 flask. Cells were grown for
3 to 7 days in a 37°C incubator with 5% CO2 in glia media.
Neuro2A cells (ATCC CCL-131, P185 at arrival) were cultured in Eagle’s Minimum Essential
Medium (SAFC Biosciences) supplemented with 10% FBS and 0.5 mM sodium pyruvate. Cells
were seeded at a density of 5 x 104 cells/mL in T25 flasks for immunoblotting and qPCR
experiments, and onto sterile 15 mm No. 1.5 glass coverslips for immunostaining experiments.
Cells were grown for 24 hours in a 37°C incubator with 5% CO2 prior to experiments.
3.2.2 Astrocyte transfection and treatment
For Piezo1 knockdown experiments, astrocyte cultures were treated with LNP-siRNA complexes
as described in section 2.2.6. The Piezo1-1591-GFP plasmid was a gift from Dr. Philip Gottlieb
(SUNY, Buffalo), corresponding to a fluorescence-tagged Piezo1 protein with GFP inserted at
amino acid position 1591 (Cox et al., 2016). The plasmid was expanded in E. coli (HST08
strain) with colonies selected by kanamycin, and plasmid DNA was isolated by midi prep
(Qiagen) according to manufacturer’s instructions. Cultures were transfected with the plasmid
(0.125 µg/mL) using Effectene (Qiagen) according to the manufacturer’s instructions, and cells
were used for staining experiments after 48 hours post-transfection.
In experiments with forskolin treatment, cultures were incubated with serum-free media for 1
hour followed by 1-hour incubation with forskolin (1 µM, diluted from 1 mM stock in DMSO;
Sigma) prior to imaging.

68
3.2.3 qPCR for Neuro2A cells
Quantitative polymerase chain reaction (qPCR) was performed on Neuro2A cells as described in
Section 2.2.8. Probes from Integrated DNA Technologies were used to amplify PIEZO1
(mPiezo1-1 Mm.PT.58.11048868; mPiezo1-2 Mm.PT.58.10591550), PIEZO2 (mPiezo2-1
Mm.PT.58.6238757; mPiezo2-2 Mm.PT.58.30174298), and tyrosine hydroxylase (mTh
Mm.PT.58.33106186), with GAPDH (mGAPDH Mm.PT.39a.1) as the internal control.
3.2.4 Immunoblotting
Astrocyte and Neuro2A cultures were collected using 1X Laemmli buffer with cOmplete
protease inhibitor cocktail (Roche). Samples were heated for 10 minutes at 65°C, homogenized
with an insulin syringe, and heated for another 5 minutes at 65°C. Wild-type C57BL/6 mice
(P30) were anesthetized with isoflurane and decapitated according to protocols approved by the
University of British Columbia Animal Care Committee. Lung tissue was flash-frozen in liquid
nitrogen and homogenized with 2X Laemmli buffer supplemented with protease inhibitor
cocktail. Samples were heated for 10 minutes at 65°C, homogenized with a 23G syringe
followed by an insulin syringe, and heated for another 5 minutes at 65 °C. All samples were
then centrifuged for 10 minutes at 10,000 rpm at 4°C to remove debris. Protein concentration
was determined with the BCA protein assay (Pierce) measured at 660 nm with a microplate
reader. Samples were resolved on 3-8% Tris-acetate gradient gels (Invitrogen), and transferred
overnight to HyBond membranes (GE Life Sciences). Membranes were then equilibrated in
TBS-Tween (TBST) for 10 minutes and blocked with TBST + 2% milk for one hour at room
temperature. Primary antibodies used were: 1/1000 Piezo1 (Proteintech); 1/20,000 vinculin
(Sigma). Secondary antibodies used were: 1/5000 horseradish peroxidase (HRP)-conjugated

69
goat anti-rabbit; 1/20,000 HRP-conjugated goat anti-mouse (both from Pierce). Blots were
incubated using SuperSignal West Pico Chemiluminescent Substrate (Pierce), and developed on
film.
3.2.5 Immunocytochemistry
Based on the previously reported distribution of Piezo1 in different cell types, we used different
staining protocols that have been optimized to target those subcellular compartments to study
Piezo1 localization in astrocytes, as depicted in Fig. 3-1. For example, strong detergents such as
Triton-X allow antibodies to access intracellular cytoskeletal epitopes, but may also extract
membrane proteins along with lipids, suggesting that a single staining protocol is insufficient for
achieving optimal staining for all cellular structures (Jamur and Oliver, 2010; Bocksteins et al.,
2012; Maity et al., 2013). We therefore modified the duration of permeabilization and compared
the staining pattern of Piezo1 with organelle-specific markers to assess Piezo1 localization to the
corresponding subcellular compartments. To confirm the Piezo1 staining pattern, we compared
experiments performed using two commercial antibodies raised against different epitopes of the
rat Piezo1 protein (Fig. 3-2).
Cells were fixed for 10 minutes with 4% paraformaldehyde (PFA; Ted Pella), followed by two 5-
minute washes with phosphate buffered saline (PBS; NaCl 110 mM, K2HPO4 16 mM, KH2PO4 3
mM, pH 7.4). In the ‘with permeabilization’ conditions, cells were treated with either 0.1%
Triton X-100 (Thermo Scientific) or 0.01% saponin (Sigma), followed by two PBS washes.
Cells were then blocked with 4% normal goat serum (Abcam) for 30 minutes, and incubated
overnight at 4°C with primary antibodies diluted in 0.1% bovine serum albumin (BSA; Sigma).

70
Samples were washed twice with PBS and incubated with secondary antibodies diluted in 0.1%
BSA for 1 hour, followed by two PBS washes. Cells were incubated with Hoechst dye for 10
minutes to label the nuclei. After washing, coverslips were mounted using Vectashield mounting
media (Vector Labs) and sealed with nail polish. As negative controls, Piezo1 antibody was
omitted or replaced with rabbit IgG (Santa Cruz Biotechnology). Primary antibodies and probes
used were: 1/300 Piezo1 (Alomone and Proteintech); 1/200 vinculin (Sigma); 1/400 calnexin
(Abcam); 1/2000 GFP (Abcam); 1/200 phalloidin-633; 1/200 wheat germ agglutinin-555; 1/1000
Hoechst 33342 (all from Invitrogen). Secondary antibodies used were: 1/200 goat anti-rabbit
Alexa Fluor 488 and 546; 1/200 goat anti-chicken Alexa Fluor 488; 1/200 goat anti-mouse Alexa
Fluor 546 (all from Invitrogen).
Figure 3-1. Flowchart of immunocytochemistry protocols for optimization of Piezo1 staining in different
subcellular regions.

71
Figure 3-2. Schematic illustrating the epitopes targeted by the Alomone and Proteintech Piezo1 antibodies.
The Piezo1 topology is adapted from Cox et al. (2016) and Zhao et al. (2016). According to information from the
commercially supplied technical sheets, the Alomone and Proteintech antibodies were raised against epitopes
corresponding to amino acid sequences 1844 – 1888 and 2199 – 2437, respectively that we mapped to the rat Piezo1
protein sequence (NCBI reference sequence NP_001070668.2).
3.2.6 Immunohistochemistry
Wild-type C57BL/6 mice (P30) were anesthetized with halothane and given an intraperitoneal
injection of urethane (0.1 mL of 30% urethane per 10 g body weight). Mice were then
transcardially perfused with 0.1 M PBS (Oxoid) followed by 4% PFA (FD Neuro Technologies).
Brains were extracted and immersed in post-fix (10% sucrose in 4% PFA) at 4°C overnight,
followed by cryoprotection in 30% sucrose in 0.1 M PBS overnight. Using a cryostat (Leica), 40
µm coronal sections were collected and stored in cryoprotectant (30% sucrose, 30% ethylene
glycol, and 1% polyvinyl-pyrrolidone in 0.1 M PBS) at -20°C.
Free-floating sections were rinsed in PBS and blocked in 10% normal donkey serum (Jackson
Immunoresearch Laboratories) and 0.4% Triton-X (Sigma) for one hour, followed by incubation
with primary antibodies diluted in buffer (2.5% donkey serum + 0.04% Triton-X) overnight at
4°C. Sections were then rinsed in PBS and incubated with secondary antibodies for 1.5 hours at

72
room temperature. Sections were rinsed with PBS, mounted onto glass slides, and coverslipped
using FluorSave Reagent (Calbiochem). Primary antibodies used were: 1/100 Piezo1 (Alomone
and Proteintech); 1/250 GFAP (Invitrogen); 1/250 Aquaporin 4 (Santa Cruz Biotechnology).
Secondary antibodies used were: 1/500 donkey anti-rabbit Alexa Fluor 488; 1/500 donkey anti-
rat Alexa Fluor 546; 1/500 donkey anti-goat Alexa Fluor 546 (all from Invitrogen).
3.2.7 Confocal image acquisition and analysis
Cells and tissue sections were imaged using a TCS SP8 confocal laser scanning microscope
(Leica) with a 25x-W/0.95 NA and a 63x-O/1.4 NA objective lens. Images were acquired as 512
x 512-pixel frames using 4-line averaging. For siRNA-treated coverslips, 12 random fields were
acquired per coverslip.
We observed that non-permeabilized astrocytes displayed a heterogeneous staining pattern, and
decided to classify the cells into different categories based on the characteristics of their Piezo1
immunoreactivity, as described in Fig. 3-4. Quantitative analysis was focused on astrocytes
exhibiting Type 1 staining pattern (cells with multiple discrete Piezo1-positive clusters) as these
cells had the clearest signal-to-noise ratio, and the staining pattern resembled the Yoda1-
activated Ca2+ microdomain signals imaged in Chapter 2. Statistics were performed as described
in section 2.2.15, where unpaired t-tests were used to compare the significance of difference in
means between two groups. Data are reported as mean ± 95% confidence interval, and p < 0.05
was considered statistically significant.

73
3.3 Results
3.3.1 Antibody testing in Neuro2A cells that endogenously express Piezo1
We initially decided to test the Piezo1 staining procedure and antibody using Neuro2A mouse
neuroblastoma cells as a positive control, since Piezo1 was initially cloned from this cell line
(Coste et al., 2010). Neuro2A cells contained mRNA for Piezo1, but not the homolog Piezo2, as
quantified by qPCR with two sets of probes against independent regions of each gene (Fig. 3-
3A). In addition, tyrosine hydroxylase was probed as a phenotypic marker for Neuro2A cells,
which can be induced to differentiate into dopaminergic-like neurons (Tremblay et al., 2010).
Although a variety of commercially available antibodies against Piezo1 exist, the Proteintech
antibody was selected as it had been shown to stain bladder urothelial cells (Miyamoto et al.,
2014), a tissue known to express Piezo1 mRNA (Coste et al., 2010). Immunoblot analysis
demonstrated that the antibody recognized the Piezo1 protein with a single band from cultured
astrocyte lysate (Fig. 3-3B), albeit with a higher molecular weight, compared to the Neuro2A
cell lysate and mouse lung tissue positive controls (Coste et al., 2010). Interestingly, this
predominant band has a higher molecular weight than the previously observed band pattern at
~280 kDa (Fig. 2-2E), and may represent Piezo1 dimers that have been described by other
groups (Coste et al., 2012). Immunocytochemistry performed in the absence of membrane
permeabilization revealed that Piezo1 immunoreactivity is located in clusters at the surface of
Neuro2A cells (Fig. 3-3C). The ability of the Proteintech antibody to detect endogenous Piezo1
protein by immunoblotting and staining in Neuro2A cells suggests that it may also be used for
Piezo1 immunostaining in astrocytes.
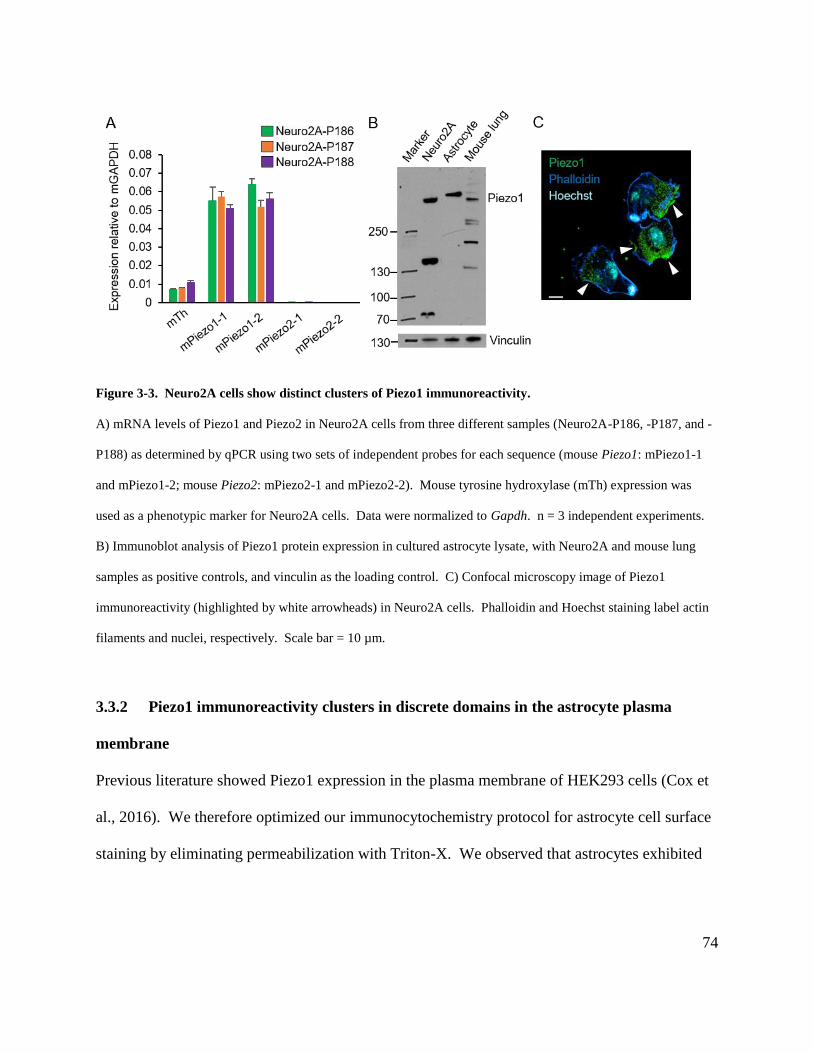
74
Figure 3-3. Neuro2A cells show distinct clusters of Piezo1 immunoreactivity.
A) mRNA levels of Piezo1 and Piezo2 in Neuro2A cells from three different samples (Neuro2A-P186, -P187, and -
P188) as determined by qPCR using two sets of independent probes for each sequence (mouse Piezo1: mPiezo1-1
and mPiezo1-2; mouse Piezo2: mPiezo2-1 and mPiezo2-2). Mouse tyrosine hydroxylase (mTh) expression was
used as a phenotypic marker for Neuro2A cells. Data were normalized to Gapdh. n = 3 independent experiments.
B) Immunoblot analysis of Piezo1 protein expression in cultured astrocyte lysate, with Neuro2A and mouse lung
samples as positive controls, and vinculin as the loading control. C) Confocal microscopy image of Piezo1
immunoreactivity (highlighted by white arrowheads) in Neuro2A cells. Phalloidin and Hoechst staining label actin
filaments and nuclei, respectively. Scale bar = 10 µm.
3.3.2 Piezo1 immunoreactivity clusters in discrete domains in the astrocyte plasma
membrane
Previous literature showed Piezo1 expression in the plasma membrane of HEK293 cells (Cox et
al., 2016). We therefore optimized our immunocytochemistry protocol for astrocyte cell surface
staining by eliminating permeabilization with Triton-X. We observed that astrocytes exhibited
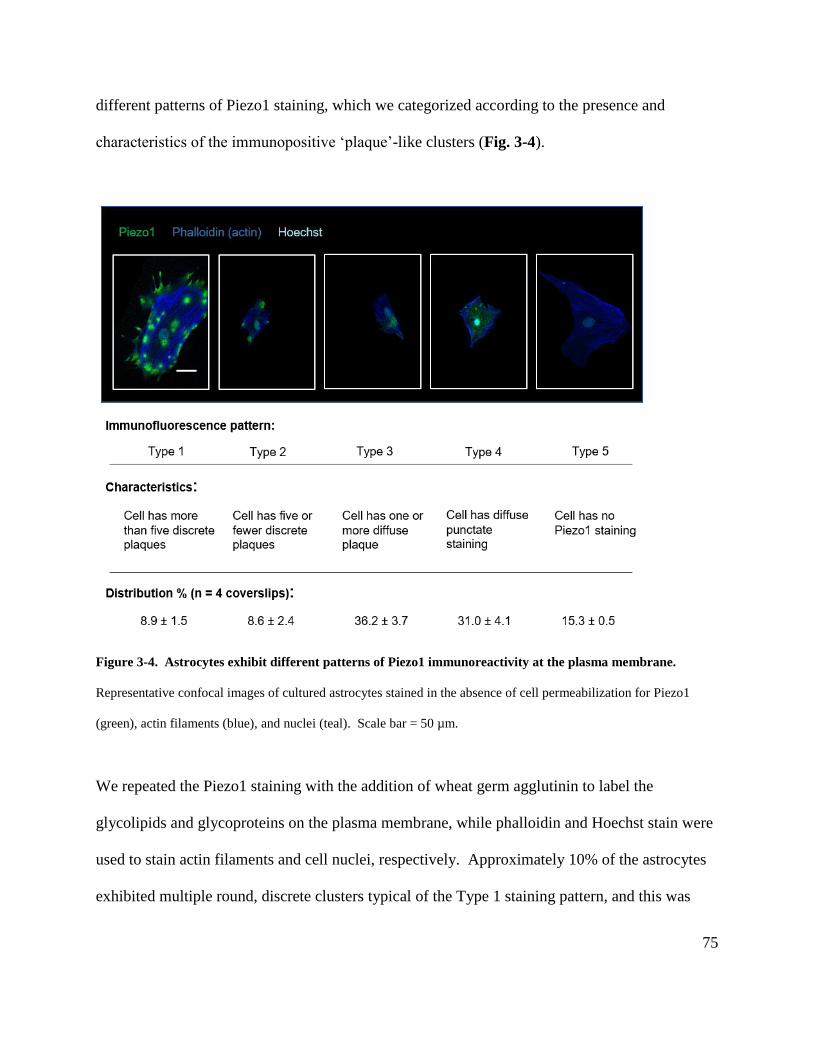
75
different patterns of Piezo1 staining, which we categorized according to the presence and
characteristics of the immunopositive ‘plaque’-like clusters (Fig. 3-4).
Figure 3-4. Astrocytes exhibit different patterns of Piezo1 immunoreactivity at the plasma membrane.
Representative confocal images of cultured astrocytes stained in the absence of cell permeabilization for Piezo1
(green), actin filaments (blue), and nuclei (teal). Scale bar = 50 µm.
We repeated the Piezo1 staining with the addition of wheat germ agglutinin to label the
glycolipids and glycoproteins on the plasma membrane, while phalloidin and Hoechst stain were
used to stain actin filaments and cell nuclei, respectively. Approximately 10% of the astrocytes
exhibited multiple round, discrete clusters typical of the Type 1 staining pattern, and this was

76
reminiscent of both spontaneous and Yoda1-induced Ca2+ microdomains observed in Chapter 2
(Fig. 3-5A). Similarly, astrocytes transfected with Piezo1-GFP showed a clustered
immunofluorescence pattern that co-localized with Piezo1 antibody staining (Fig. 3-5B). This
aggregate pattern was not observed in negative controls where the Piezo1 primary antibody had
been omitted (Fig. 3-5C) or replaced with an isotype control (Fig. 3-5D). The clustered staining
pattern of Piezo1 was replicated using an alternate Piezo1 antibody from Alomone Labs that was
raised against a different immunogen (Fig. 3-5E).
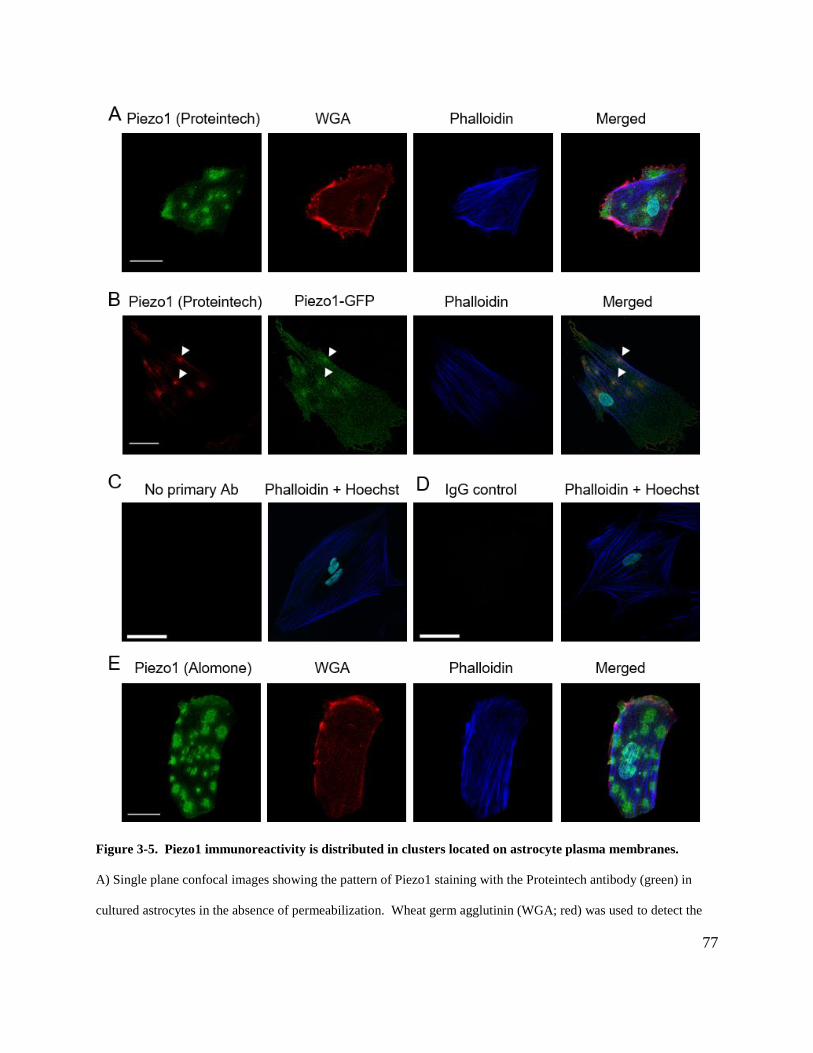
77
Figure 3-5. Piezo1 immunoreactivity is distributed in clusters located on astrocyte plasma membranes.
A) Single plane confocal images showing the pattern of Piezo1 staining with the Proteintech antibody (green) in
cultured astrocytes in the absence of permeabilization. Wheat germ agglutinin (WGA; red) was used to detect the

78
glycolipids and glycoproteins on the cell plasma membrane, and phalloidin (blue) was used as an actin filament
marker. B) Confocal images of astrocytes transfected with 0.25 µg Piezo1-GFP DNA and stained with anti-GFP
(green) and anti-Piezo1 antibody (red). Co-localization between the GFP-tagged Piezo1 and the Piezo1 antibody
signal is highlighted with white arrowheads. Negative controls where Piezo1 primary antibody was C) omitted, or
D) replaced with the IgG isotype control; phalloidin and Hoechst staining was imaged in the same fields to show the
actin filaments and nuclei, respectively. E) Confocal images as in part A), with Piezo1 being detected by an antibody
from Alomone Labs. Scale bar = 50 µm.
We further examined Piezo1 expression in the astrocyte membrane by conducting
immunostaining following protein knockdown with Piezo1 siRNA. We focused on astrocytes
showing Type 1 staining based on the high signal-to-noise ratio of the Piezo1 clusters that
allowed them to be quantified accurately, and because the staining pattern resembles the Ca2+
microdomains imaged in Chapter 2. Piezo1 siRNA reduced the number of Piezo1 clusters in
these astrocytes (luciferase siRNA: 55 cells analyzed from 4 coverslips, 14 ± 2.8 clusters/cell;
Piezo1 siRNA: 52 cells analyzed from 4 coverslips, 9.8 ± 2.9 clusters/cell; p < 0.01)(Fig. 3-6A
and B).
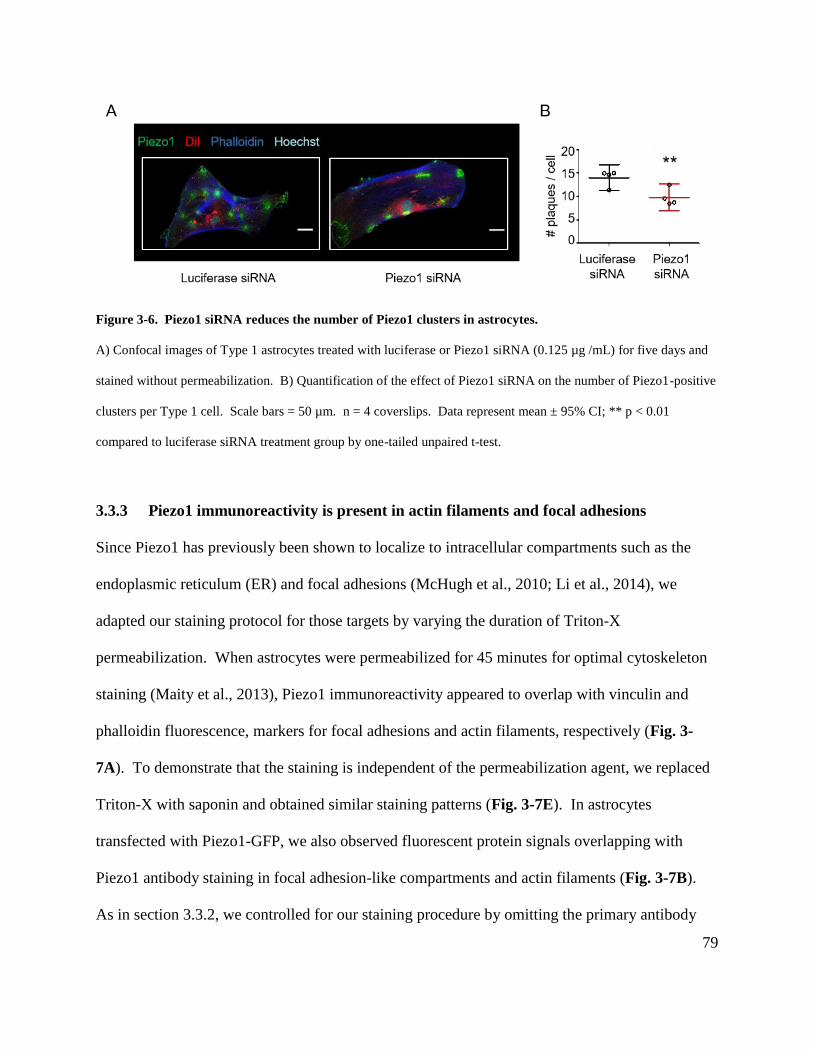
79
Figure 3-6. Piezo1 siRNA reduces the number of Piezo1 clusters in astrocytes.
A) Confocal images of Type 1 astrocytes treated with luciferase or Piezo1 siRNA (0.125 µg /mL) for five days and
stained without permeabilization. B) Quantification of the effect of Piezo1 siRNA on the number of Piezo1-positive
clusters per Type 1 cell. Scale bars = 50 µm. n = 4 coverslips. Data represent mean ± 95% CI; ** p < 0.01
compared to luciferase siRNA treatment group by one-tailed unpaired t-test.
3.3.3 Piezo1 immunoreactivity is present in actin filaments and focal adhesions
Since Piezo1 has previously been shown to localize to intracellular compartments such as the
endoplasmic reticulum (ER) and focal adhesions (McHugh et al., 2010; Li et al., 2014), we
adapted our staining protocol for those targets by varying the duration of Triton-X
permeabilization. When astrocytes were permeabilized for 45 minutes for optimal cytoskeleton
staining (Maity et al., 2013), Piezo1 immunoreactivity appeared to overlap with vinculin and
phalloidin fluorescence, markers for focal adhesions and actin filaments, respectively (Fig. 3-
7A). To demonstrate that the staining is independent of the permeabilization agent, we replaced
Triton-X with saponin and obtained similar staining patterns (Fig. 3-7E). In astrocytes
transfected with Piezo1-GFP, we also observed fluorescent protein signals overlapping with
Piezo1 antibody staining in focal adhesion-like compartments and actin filaments (Fig. 3-7B).
As in section 3.3.2, we controlled for our staining procedure by omitting the primary antibody

80
(Fig. 3-7C) or replacing it with rabbit IgG as the species isotype control (Fig. 3-7D); a faint
nuclear signal was detected in the isotype control, but neither controls showed staining in
cytoskeleton components. We repeated the experiments using the Alomone Piezo1 antibody and
achieved the same observation (data not shown).
Interestingly, Piezo1-GFP-transfected astrocytes also exhibited a perinuclear fluorescent signal
resembling ER staining (Fig. 3-7B). However, ectopic localization of fluorescently tagged
proteins is a known issue, as overexpressed proteins can be trapped in the endomembrane system
(Stadler et al., 2013). We therefore examined whether Piezo1 is expressed in the ER by
permeabilizing cells with Triton-X for 5 minutes and compared the staining pattern to the ER
marker calnexin. Since the antibodies are from the same host species, cells were independently
stained with Piezo1 or calnexin and the staining pattern was analyzed on separate coverslips.
Piezo1 immunoreactivity was more filamentous and peripheral, indicative of focal adhesions and
actin filaments, which is distinct from the perinuclear pattern of calnexin staining (Fig. 3-7F).
The difference between the staining patterns of Piezo1 and calnexin suggest that ER localization
of Piezo1-GFP might be an artifact.
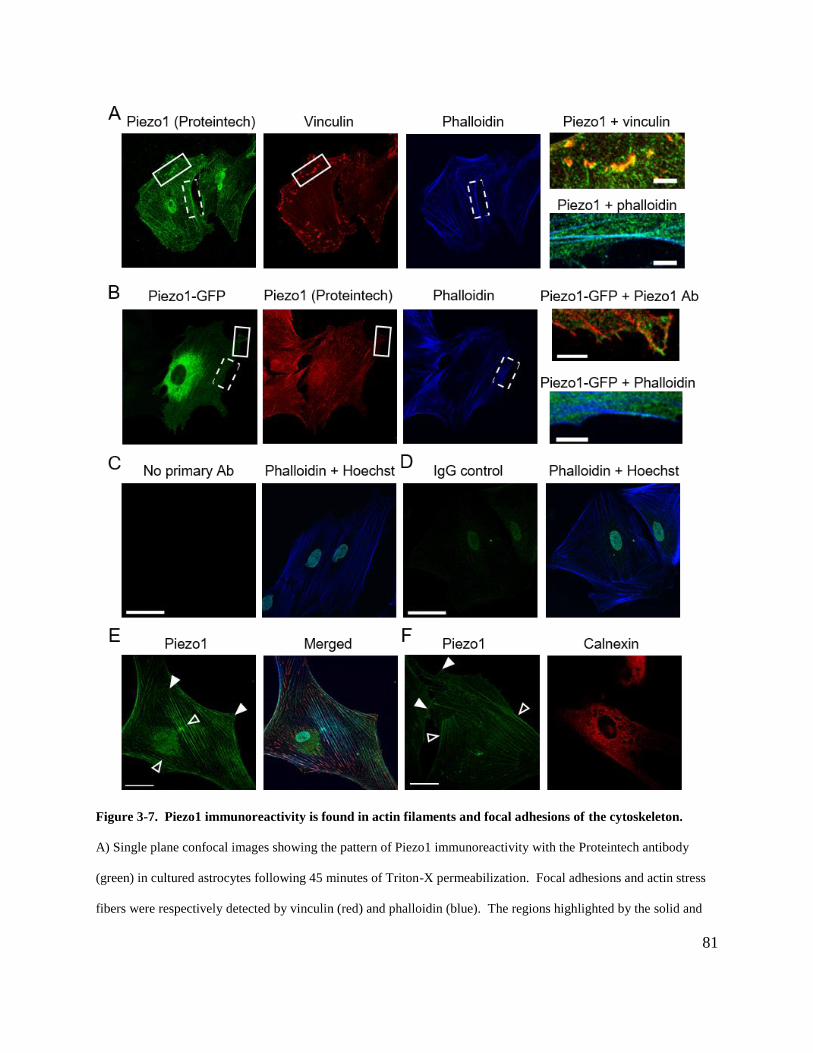
81
Figure 3-7. Piezo1 immunoreactivity is found in actin filaments and focal adhesions of the cytoskeleton.
A) Single plane confocal images showing the pattern of Piezo1 immunoreactivity with the Proteintech antibody
(green) in cultured astrocytes following 45 minutes of Triton-X permeabilization. Focal adhesions and actin stress
fibers were respectively detected by vinculin (red) and phalloidin (blue). The regions highlighted by the solid and

82
dashed rectangles are shown at higher magnification in the right-most panels, respectively representing the merged
images between Piezo1 and vinculin (top image) and between Piezo1 and phalloidin (bottom image). Scale bars =
10 µm. B) Confocal images of astrocytes transfected with 0.25 µg Piezo1-GFP DNA and stained with anti-GFP
antibody (green), anti-Piezo1 antibody (red), and phalloidin (blue). The regions highlighted by the solid and dashed
rectangles are shown at higher magnification in the right-most panels, respectively representing the merged images
between Piezo1-GFP and Piezo1 immunoreactivity with the antibody (top image) and between Piezo1-GFP and
phalloidin (bottom image). Scale bars = 10 µm. Negative controls where Piezo1 primary antibody was C) omitted,
or D) replaced with the IgG isotype control; phalloidin and Hoechst staining was imaged in the same fields to show
the actin filaments and nuclei, respectively. E) Confocal images as in part A), with permeabilization using saponin
instead of Triton-X. Piezo1 staining associated with focal adhesion structures (solid arrowheads) and actin stress
fibers (empty arrowheads) are highlighted. F) Confocal images showing Piezo1 and calnexin immunoreactivity
following 5 minutes of Triton-X permeabilization. Scale bars = 50 µm.
3.3.4 Forskolin-induced disorganization of the cytoskeleton alters Piezo1 distribution
Piezo1 is a mechanically activated channel, and our immunostaining data and the literature
suggest that it may be associated with cytoskeletal components that are involved in
mechanosensing. To further probe the interactions between Piezo1 and the cytoskeleton, we
assessed the effect of altering the cytoskeletal architecture with forskolin on the Piezo1 staining
pattern. Forskolin increases cAMP levels in astrocytes, leading to cell stellation characterized by
membrane retraction and loss of focal adhesions and actin stress fibers (Perez et al., 2005;
Rodnight and Gottfried, 2013; Vardjan et al., 2014). In this study, we tested different
concentrations of forskolin treatment and observed that 1-hour incubation with 1 µM forskolin
resulted in a typical stellation response. These morphological changes are shown in Fig. 3-8A,
in cells that were permeabilized with Triton-X for 45 minutes. Forskolin-induced stellation
(astrocyte highlighted by the dotted line in the ‘Piezo1’ panel) induced complete loss of Piezo1

83
immunoreactivity in focal adhesion and actin stress fiber-like compartments. This was
accompanied by stress fiber depolymerization, as seen by the loss of parallel structures detected
by phalloidin in the stellate astrocyte, although stress fibers remained present in the surrounding
non-stellate cells (marked by solid arrowheads). Stellation also induced loss of focal adhesions
in the periphery of the astrocyte cell body, in contrast to the distribution of vinculin
immunoreactivity displayed by non-stellate cells (marked by empty arrowheads, Fig. 3-8B). In
the absence of permeabilization, forskolin treatment resulted in redistribution of Piezo1
immunoreactivity to the membranes of astrocyte processes (highlighted by empty arrowheads);
phalloidin staining was also performed to show fragmentation of the actin stress fibers into
globular clumps (highlighted by solid arrowheads) (Fig. 3-8C). Taken together, our data
indicate that forskolin treatment alters intracellular and membrane Piezo1 redistribution,
suggesting an association between Piezo1 and the cytoskeleton.
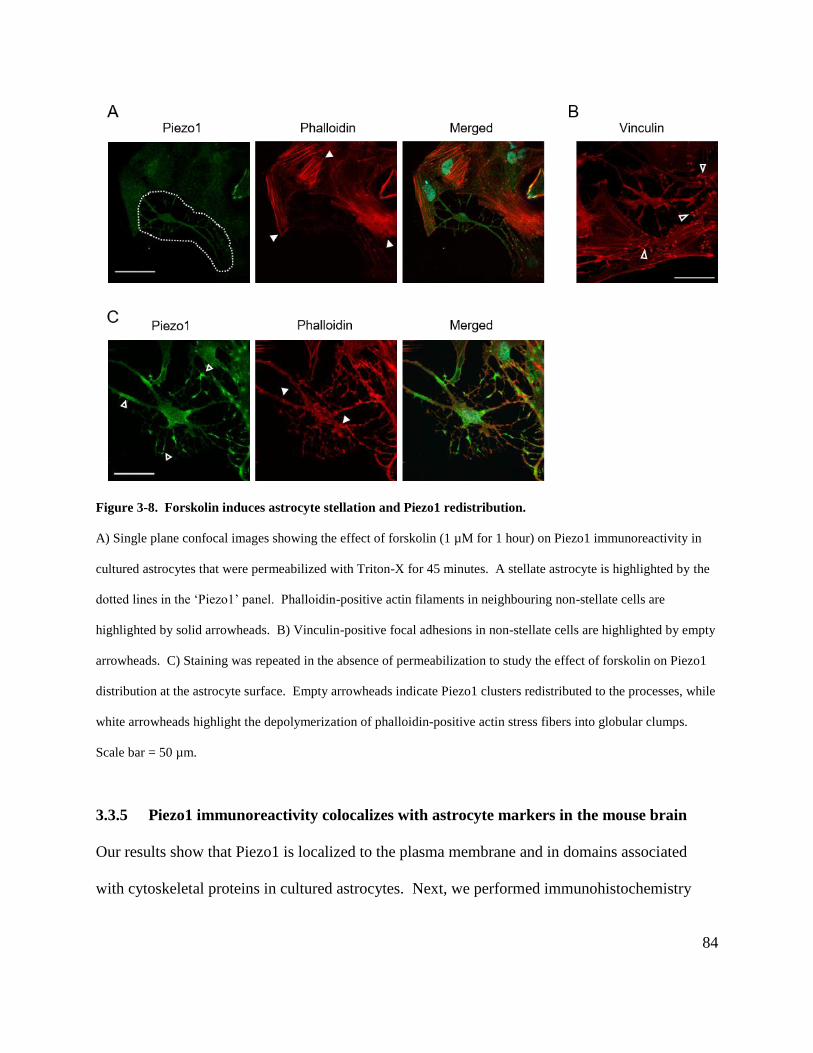
84
Figure 3-8. Forskolin induces astrocyte stellation and Piezo1 redistribution.
A) Single plane confocal images showing the effect of forskolin (1 µM for 1 hour) on Piezo1 immunoreactivity in
cultured astrocytes that were permeabilized with Triton-X for 45 minutes. A stellate astrocyte is highlighted by the
dotted lines in the ‘Piezo1’ panel. Phalloidin-positive actin filaments in neighbouring non-stellate cells are
highlighted by solid arrowheads. B) Vinculin-positive focal adhesions in non-stellate cells are highlighted by empty
arrowheads. C) Staining was repeated in the absence of permeabilization to study the effect of forskolin on Piezo1
distribution at the astrocyte surface. Empty arrowheads indicate Piezo1 clusters redistributed to the processes, while
white arrowheads highlight the depolymerization of phalloidin-positive actin stress fibers into globular clumps.
Scale bar = 50 µm.
3.3.5 Piezo1 immunoreactivity colocalizes with astrocyte markers in the mouse brain
Our results show that Piezo1 is localized to the plasma membrane and in domains associated
with cytoskeletal proteins in cultured astrocytes. Next, we performed immunohistochemistry

85
with fixed mouse coronal brain sections to determine whether Piezo1 is also present in astrocytes
in situ. Piezo1 immunoreactivity was observed to colocalize with the astrocyte markers glial
fibrillary acidic protein (GFAP) and aquaporin4 (Aqp4) in layer I of the cortex and in the CA1
region of the hippocampus (Fig. 3-9). In addition to astrocyte cell bodies, a diffuse ‘spongiform’
staining pattern was observed surrounding the large processes marked by GFAP, characteristic of
the morphology of fine processes present in mature astrocytes (Bushong et al., 2004).
Interestingly, Piezo1 immunoreactivity was also detected in the astrocyte endfeet, specialized
processes that contact the cerebral vasculature (highlighted by arrowheads). No staining was
observed in negative controls where the Piezo1 primary antibody had been omitted (data not
shown). Together the results indicate astrocytic expression of Piezo1 in the brain.
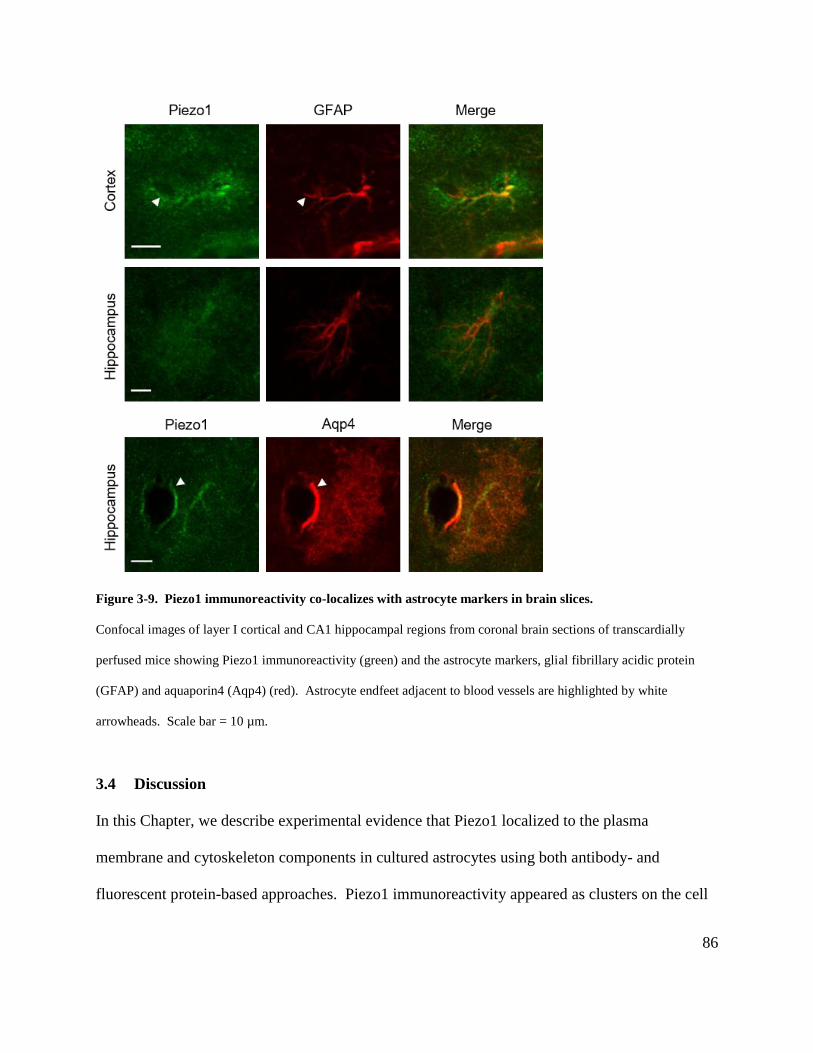
86
Figure 3-9. Piezo1 immunoreactivity co-localizes with astrocyte markers in brain slices.
Confocal images of layer I cortical and CA1 hippocampal regions from coronal brain sections of transcardially
perfused mice showing Piezo1 immunoreactivity (green) and the astrocyte markers, glial fibrillary acidic protein
(GFAP) and aquaporin4 (Aqp4) (red). Astrocyte endfeet adjacent to blood vessels are highlighted by white
arrowheads. Scale bar = 10 µm.
3.4 Discussion
In this Chapter, we describe experimental evidence that Piezo1 localized to the plasma
membrane and cytoskeleton components in cultured astrocytes using both antibody- and
fluorescent protein-based approaches. Piezo1 immunoreactivity appeared as clusters on the cell

87
surface that resembled the Ca2+ microdomains imaged in Chapter 2, and these clusters were
reduced by Piezo1 siRNA treatment. Forskolin-induced disorganization of focal adhesions and
actin filaments was associated with loss of intracellular Piezo1 staining and redistribution of the
surface membrane Piezo1 clusters to the processes of the stellate astrocytes, suggesting that the
cytoskeleton components may be involved in organizing Piezo1 domains. Finally,
immunohistochemistry using fixed brain slices revealed Piezo1 immunoreactivity in cortical and
hippocampal astrocytes, indicating that Piezo1 may also be expressed in astrocytes in situ.
Protein localization is commonly visualized by imaging of either immunofluorescence or
fluorescent-tagged proteins. Immunofluorescence relies on an antibody-antigen interaction to
label endogenous proteins of interest in fixed tissues or cells, but artifacts may arise from
antibody recognition of off-target proteins, or from the protocols used for fixation and
permeabilization (Schnell et al., 2012; Stadler et al., 2013). In contrast, fluorescent-tagged
proteins allow protein distribution to be visualized in live or fixed cells without the use of
antibodies. However, protein trafficking may be altered by overexpression of these proteins
above endogenous protein levels, or by the fluorescent tag itself (Stadler et al., 2013). As each
technique exhibits distinct advantages and disadvantages, we combined both approaches to
strengthen our findings.
Several commercially available and in-house antibodies have been used to examine Piezo1
distribution in different cell types (Coste et al., 2010; McHugh et al., 2010; Blumenthal et al.,
2014; Miyamoto et al., 2014; Gudipaty et al., 2017). To our knowledge, our immunostaining
using the Proteintech antibody is the first demonstration of surface Piezo1 immunofluorescence

88
in Neuro2A cells, where Piezo1 is functionally expressed (Coste et al., 2010). The antibody also
detected Piezo1 protein in astrocytes with both immunoblotting and immunostaining approaches,
consistent with previous transcriptome and proteome studies (Zhang et al., 2014; Sharma et al.,
2015; Srinivasan et al., 2016).
Our finding that Piezo1 is distributed as clusters on astrocyte plasma membranes is supported by
some examples from other cell types in the literature. For instance, Coste et al. (2010) showed
overlap between Piezo1 staining and the control surface marker TRPA1 in HEK293 cells
transfected with Piezo1. Other groups observed that fluorescent protein-tagged Piezo1 formed
clustered ‘domains’ at the surface of transfected HEK293 cells (Cox et al., 2016;
Gnanasambandam et al., 2017). The staining pattern observed is consistent with the organization
of discrete spatial domains described using electrophysiological techniques that suggest groups
of Piezo1 channels change kinetics simultaneously (Bae et al., 2013a). However, in contrast to
the irregularly-shaped clusters observed by Cox et al. (2016), astrocyte Piezo1 appeared to
aggregate as round ‘plaque’-like domains as detected by antibody and fluorescent protein
approaches. These Piezo1-immunopositive clusters bear a striking resemblance to the Ca2+
microdomains observed in Chapter 2, suggesting that the near-membrane Ca2+ signals detected
by Lck-GCaMP5 may accurately represent the spatial orientation of Piezo1 channels mediating
Ca2+ entry at the astrocyte plasma membrane.
We observed a variety of different Piezo1 staining patterns at the plasma membrane of cultured
astrocytes. Heterogeneity of Piezo1 expression has also been described by Gudipaty et al. (2017)
in canine kidney epithelial cells, where Piezo1 distribution and function depended on density-

89
induced mechanical forces experienced by the cells. Preliminary observations in our lab indicate
that increasing astrocyte culture density is associated with increased Piezo1 expression as
quantified by immunoblotting. However, we did not quantify cell density in the immunostaining
experiments, as the cells detach during the fixation procedure, and it remains to be determined if
Piezo1 localization in astrocytes is also density-dependent. Previous studies have demonstrated
that astrocyte density relies on factors including brain region, physiological development and
ageing, and pathological proliferation as seen in reactive gliosis in response to CNS injury such
as stroke or traumatic brain injury (Sofroniew and Vinters, 2010; Olude et al., 2015). Hence it
would be worthwhile to examine how astrocyte Piezo1 subcellular distribution and function are
affected by different physiological and pathological states. The range of staining patterns
observed in astrocytes may also be related to heterogeneity in astrocyte functional state. For
instance, the greatest number of discrete Piezo1 clusters was observed in large astrocytes,
suggesting that Piezo1 expression may also be dependent on progression through the cell cycle,
metabolic state, or interactions with the extracellular matrix and signaling pathways that regulate
cell morphology and growth (Lloyd, 2013; Levy et al., 2014).
Our experiments indicated that Piezo1 siRNA reduced the number of Piezo1-immunopositive
domains at the astrocyte plasma membrane, correlating with the result that the number of basal
Ca2+ microdomains are also sensitive to siRNA knockdown (Fig. 2-6C). However, we did not
observe an absolute absence of immunolabeled Piezo1 protein, supported by the Ca2+ imaging
experiments showing that Yoda1 could still evoke an increase in Ca2+ microdomain signals in
Piezo1 siRNA-treated astrocytes (Fig. 2-6B). Previous studies have suggested that the degree of
siRNA-mediated protein knockdown depends on factors such as the relation between siRNA

90
stability, mRNA transcription rate and half-life, and Piezo1 turnover rate (Stadler et al., 2013).
In addition, Thul et al. (2017) recently demonstrated that cells can exhibit variations in protein
expression levels and spatial distribution depending on their cell cycle stage. Thus, the
efficiency of siRNA-mediated knockdown may be improved by treating astrocytes with more
frequent, lower concentration applications of siRNA.
Piezo1 immunoreactivity in astrocytes was also found in the same cellular domains as focal
adhesions and actin stress fibers, and this expression pattern was lost in cells treated with
forskolin to induce cytoskeleton disorganization, supporting Piezo1 distribution in these
cytoskeletal compartments. This is an interesting observation given the importance of these
structures in the molecular mechanism for sensing extracellular matrix (ECM) rigidity. Previous
studies have indicated that ventral stress fibers are composed of actin filaments and myosin II
motor proteins that are anchored at both ends by large protein complexes known as focal
adhesions. These stress fibers exert traction forces that continually pull at focal adhesion
complexes. In addition, focal adhesions contain transmembrane adhesion molecules of the
integrin family that act as receptors for various ECM proteins. Therefore, focal adhesions
mediate cell detection of ECM stiffness by transmitting traction forces generated by the stress
fibers to the substrate (Geiger et al., 2009; Kobayashi and Sokabe, 2010; Tojkander et al., 2012).
The first demonstration that mechanosensitive channels may form a complex with focal
adhesions and actin stress fibers to transduce information about substrate stiffness into Ca2+
signals was performed by Hayakawa et al. (2008) in cultured human umbilical vein endothelial
cells. By using optical tweezers to stretch single stress fibers and performing simultaneous Ca2+

91
imaging, the authors found that mechanical stimulation of the actin cytoskeleton can generate
Ca2+ microdomains located near focal adhesions. These microdomain signals were inhibited by
Gd3+ or by removal of Ca2+ from the extracellular solution, suggesting the involvement of
putative mechanosensitive channels. Subsequent experiments by Pathak et al. (2014) found that
neural stem cells also exhibited microdomain Ca2+ transients that were sensitive to cytoskeletal
mechanosensing; for example, the Ca2+ signals increased in amplitude and frequency with
increasing substrate stiffness, while the signals were abolished by inhibition of myosin II-
mediated traction forces by blebbistatin. Strikingly, the Ca2+ transients were also blocked by
GsMTx4 and Piezo1 siRNA treatment, indicating that Piezo1 may be involved in sensing
mechanical properties of the substrate. Our findings are consistent with these studies, raising the
possibility that Piezo1 may associate with focal adhesions and stress fibers in astrocytes to
function as a mechanosensing unit that can also respond to different cell substrates.
Fluorescence-tagged Piezo1 has been found to localize to the ER by other researchers (Satoh et
al., 2006; McHugh et al., 2010). While we also observed a perinuclear fluorescent signal
resembling ER labeling in astrocytes that were transfected with Piezo1-GFP, subsequent
immunocytochemistry experiments showed that the Piezo1 labeling distribution differed from
the ER marker calnexin, suggesting that under our experimental conditions, the astrocyte ER
signal may represent ectopic Piezo1-GFP overexpression or a mislocalization artifact from
sequence differences between the fluorescent and native Piezo1 proteins (Nourse and Pathak,
2017).

92
To extend our findings to astrocytes in situ, Piezo1 colocalization with astrocyte markers was
assessed in the cortex and hippocampus of fixed brain slices. Interestingly, strong Piezo1
immunoreactivity was observed in the astrocyte endfeet compartment, where Ca2+ transients
were found to be sensitive to pharmacological manipulation of Piezo1 (section 2.3.7). The Ca2+
imaging and immunostaining experiments suggest that Piezo1 expression may be concentrated at
the endfeet of astrocytes in the brain. It is attractive to speculate that Piezo1 may be well-
positioned to sense mechanical forces exerted by dilating or constricting blood vessels,
modulating astrocyte Ca2+ microdomain activity that in turn can stimulate release of vasoactive
compounds to regulate brain blood flow.

93
Chapter 4: Role of Piezo1 in astrocyte swelling
4.1 Overview
Previous studies have shown that Piezo1 channels are involved in a variety of physiological
processes related to sensation and transduction of mechanical forces. For instance, Piezo1 has
been implicated in detecting shear stress and realigning vascular endothelial cells to the direction
of blood flow by mediating focal adhesion and actin stress fiber reorganization, a process that is
critical to normal vascular development (Li et al., 2014; Ranade et al., 2014). Piezo1 has also
been suggested to sense plasma membrane tension to regulate cell volume, as shown in red blood
cells where pharmacological activation and genetic knockdown of the channel were associated
with cell shrinkage and swelling, respectively (Faucherre et al., 2014; Cahalan et al., 2015). The
data presented in Chapter 3 indicate that Piezo1 is expressed in astrocyte plasma membrane and
is associated with cytoskeletal components such as cell adhesion structures, suggesting a
potential relevance for mechano-regulated responses that are functionally important for astrocyte
biology (section 1.2). We therefore examined the possibility of activating Piezo1-mediated Ca2+
signals in response to osmotic swelling as a model of mechanical stimulation.
We studied astrocyte Piezo1 activation using osmotic swelling because astrocytes are involved in
ion and volume homeostasis in the brain, and cell swelling has been observed under both
physiological and pathological conditions (Kimelberg et al., 1993; Florence et al., 2012; Xiang et
al., 2016). Previous studies have used hypotonicity-induced swelling as a strategy to
characterize the molecular components of mechanosensitive channels in astrocytes by measuring
their activation as swelling-activated currents or increases in intracellular Ca2+ (Suchyna et al.,

94
2000; Benfenati et al., 2007). Furthermore, Piezo1 channels reconstituted into lipid bilayers
have been activated by osmotic stress (Syeda et al., 2016). Hence we speculated that
hypotonicity-induced astrocyte swelling and subsequent plasma membrane stretch would
activate Piezo1 channels, leading to an increase in Ca2+ microdomain transients.
One common strategy to induce astrocyte swelling is the application of hypotonic solution,
where reduction in extracellular osmolarity triggers water movement into cells. In these
experiments, we used hyponatremia, defined as serum Na+ concentration below 135 mM
(Schober and Mongin, 2015), to induce swelling, as this model has been well-characterized in
previous studies examining the consequences of astrocyte swelling. For instance, hyponatremia
treatment has been shown to activate stretch-sensitive TRPV4 channels, induce release of
osmolytes such as glutathione and taurine, and alter glutamine synthesis in astrocytes (Clark et
al., 1996; Benfenati et al., 2011; Hyzinski-García et al., 2011; Schober and Mongin, 2015).
Interestingly, hyponatremia is of clinical relevance because it is associated with neuropathologies
involving reactive astrogliosis such as traumatic brain injury, subarachnoid hemorrhage, and
brain tumors (Kimelberg et al., 1993; Giuliani and Peri, 2014).
Since the data from Chapter 3 indicated that Piezo1 is also localized to components of the
astrocyte cytoskeleton, we wondered whether this association is also involved in mediating
Piezo1 responses to astrocyte swelling. Experimental evidence indicates that the sensitivity of
Piezo1 to mechanical forces is influenced by interactions with the cytoskeleton and extracellular
matrix (ECM) (Bavi et al., 2017). However, these interactions are complex; on one hand, the
cytoskeleton may act as a physical barrier to shield the channels from mechanical stress, but on

95
the other hand, cytoskeletal proteins may also activate Piezo1 by transducing mechanical forces
via direct physical coupling with the channel (Bavi et al., 2017; Nourse and Pathak, 2017). In
support of the idea that the cytoskeleton is mechanoprotective, Cox et al. (2016) patched Piezo1
channels in membrane blebs that lacked cytoskeleton, and showed that the pressure threshold for
Piezo1 activation was lowered. This result was consistent with the finding that deletion of
filamin A, a scaffold protein linking actin filaments to membrane proteins, increased pressure-
induced Piezo1 activation in smooth muscle cells (Retailleau et al., 2015). In contrast, other
studies have demonstrated mechanosensitive channel activation in endothelial cells in response
to direct actin cytoskeleton stimulation (Hayakawa et al., 2008), and mechanically-activated
Piezo1 currents were reduced by actin cytoskeleton disruption with cytochalasin D treatment
(Gottlieb et al., 2012). Therefore, it is likely that factors such as cell type and mode of
mechanical stimulus act in concert to determine the contribution of the cytoskeleton to
mechanosensing.
In this chapter, we aimed to test the hypothesis that hypotonicity-induced astrocyte swelling can
activate Piezo1 by combining Ca2+ imaging in Lck-GCaMP5-transfected astrocytes with Piezo1
siRNA treatment in response to hyponatremia. We also used forskolin-induced cytoskeleton
reorganization as a tool to explore the relevance of Piezo1 – cytoskeleton interactions in
hypoosmotic stress-mediated cell swelling and channel activation.

96
4.2 Methods
4.2.1 Cell culture and transfection
Astrocyte cultures were prepared as described in section 2.2.2. Cells were plated at a density of
~9 x 103 cells / mL onto sterile 15 mm No. 1.5 glass coverslips (MatTek) that were coated with
collagen IV (50 µg/mL; Corning). Previous studies have found that cells in vitro respond best
when grown in substrates that mimic their physiological extracellular matrix (ECM) (Kleinman
et al., 1987). Our data suggest that astrocyte endfeet processes may be a possible site of
concentrated Piezo1 expression in the brain (see Sections 2.3.7 and 3.3.5), and imaging studies
have shown that astrocyte endfeet are closely associated with the basal lamina of endothelial
cells surrounding cerebral capillaries (Kacem et al., 1998). Collagen IV, an abundant ECM
protein of the basement membrane, has been proposed to be a major component of the matrix at
the endfeet – endothelial cell interface (Benarroch, 2015). Therefore, astrocytes were plated on
collagen IV-coated coverslips to allow osmotic stress-induced Piezo1 activation to be studied in
a condition that better mimics the physiological ECM environment. Cells were grown for 3 to 5
days in a 37°C incubator with 5% CO2 in glia media. Cultures were transfected with LNP-
siRNA and LNP-plasmid complexes as described in section 2.2.6. In experiments with forskolin
treatment, cultures were incubated with serum-free media for 1 hour followed by 1-hour
incubation with forskolin (1 µM; Sigma) prior to imaging.
4.2.2 Culture imaging
Cells were imaged as described in section 2.2.10. Transmitted light images were collected using
infrared differential interference contrast optics. Experiments were performed at room

97
temperature with continuous perfusion (2.5 mL/min) of HEPES-aCSF (300 mOsm). Hypotonic
solution (150 mOsm) contained (in mM): NaCl, 59.9; HEPES, 10.8; KCl, 2.5; glucose, 10;
MgCl2, 2.0; CaCl2, 2.0.
4.2.3 Data analysis and statistics
Astrocyte Ca2+ microdomain signals were isolated and analyzed as described in section 2.2.14.
In contrast, Ca2+ transients in forskolin-treated cells were localized to segments of astrocyte
processes rather than presenting as circular ROIs. Discrete Ca2+ events that had connected pixels
in the x, y, and t axes were therefore identified by the 3D Connected Components function in
Matlab to allow the total number of events to be quantified per unit time. Hyponatremia-induced
astrocyte swelling was analyzed as the percent increase in cross-sectional area of the soma
relative to the baseline in isotonic solution. Statistics were performed as described in section
2.2.15, where unpaired t-tests were used to compare the significance of difference in means
between two groups, while paired t-tests were used to compare different treatments (i.e. isotonic
versus hypotonic) conducted on the same cell. Data are reported as mean ± 95% confidence
interval, and p < 0.05 was considered statistically significant.
4.3 Results
4.3.1 Piezo1 mediates Ca2+ microdomain signals in astrocytes under hypotonic stress
Astrocytes have been shown to respond to osmotic stress by elevating cytosolic Ca2+ as measured
by bulk-loaded dyes such as Fluo-4 AM and Fura-2 AM (Hua et al., 2010; Mola et al., 2016). To
determine whether Ca2+ microdomain transients are also affected by hypotonic stimulus, we

98
performed Ca2+ imaging in Lck-GCaMP5-transfected astrocytes that were exposed to
hyponatremia for 10 minutes. We found that perfusion of hypotonic solution increased the
frequency of Ca2+ microdomain signals by three-fold (baseline: 0.71 ± 0.27 events/ROI/5 min;
hypotonic: 2.17 ± 0.86 events/ROI/5 min; p < 0.01)(Fig. 4-1A and B). Furthermore, these
evoked Ca2+ signals were inhibited by Piezo1 siRNA treatment compared to the luciferase
control (luciferase baseline: 0.5 ± 0.5 events/ROI/5 min; luciferase hypotonic: 1.2 ± 0.8
events/ROI/5 min; p < 0.01; Piezo1 baseline: 0.7 ± 0.4 events/ROI/5 min; Piezo1 hypotonic: 0.8
± 0.4 events/ROI/5 min; p > 0.05)(Fig. 4-1A, C and D). Together, the data indicate that
hyponatremia-induced Ca2+ microdomain transients are mediated by Piezo1.
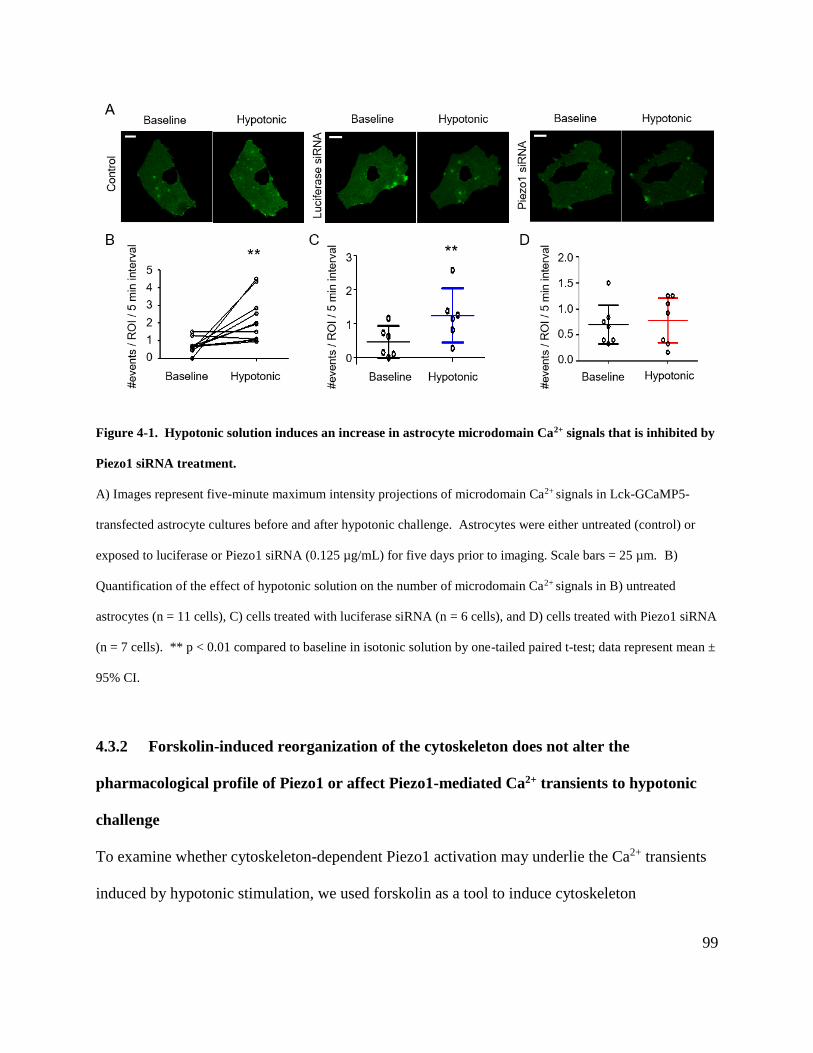
99
Figure 4-1. Hypotonic solution induces an increase in astrocyte microdomain Ca2+ signals that is inhibited by
Piezo1 siRNA treatment.
A) Images represent five-minute maximum intensity projections of microdomain Ca2+ signals in Lck-GCaMP5-
transfected astrocyte cultures before and after hypotonic challenge. Astrocytes were either untreated (control) or
exposed to luciferase or Piezo1 siRNA (0.125 µg/mL) for five days prior to imaging. Scale bars = 25 µm. B)
Quantification of the effect of hypotonic solution on the number of microdomain Ca2+ signals in B) untreated
astrocytes (n = 11 cells), C) cells treated with luciferase siRNA (n = 6 cells), and D) cells treated with Piezo1 siRNA
(n = 7 cells). ** p < 0.01 compared to baseline in isotonic solution by one-tailed paired t-test; data represent mean ±
95% CI.
4.3.2 Forskolin-induced reorganization of the cytoskeleton does not alter the
pharmacological profile of Piezo1 or affect Piezo1-mediated Ca2+ transients to hypotonic
challenge
To examine whether cytoskeleton-dependent Piezo1 activation may underlie the Ca2+ transients
induced by hypotonic stimulation, we used forskolin as a tool to induce cytoskeleton

100
reorganization. Previous immunostaining results indicated that forskolin treatment induces
remodeling of actin stress fibers and loss of focal adhesions in astrocytes, as well as Piezo1
redistribution at the plasma membrane (section 3.3.4). Here we observed that forskolin-treated
astrocytes exhibited spontaneous Ca2+ transients as detected by Lck-GCaMP5, but these
transients were localized along astrocyte processes instead of showing the circular microdomain
phenotype observed in control cells. Therefore, discrete Ca2+ events were quantified by 3D
connected components (Fig. 4-2A) rather than identifying microdomains based on fluorescence
intensity of 6 x 6-pixel ROIs (Fig. 2-3D). The frequency of Ca2+ transients was increased by
application of the Piezo1 agonist Yoda1 and blocked by subsequent addition of Gd3+ (baseline:
43 ± 66 events/5 min; Yoda1: 203 ± 78 events/5 min; Yoda1 + Gd3+: 63 ± 61 events/5 min; p <
0.01), suggesting that the channels are functional, independent of cytoskeleton organization (Fig.
4-2B and C).

101
Figure 4-2. Forskolin treatment does not affect the pharmacological profile of Piezo1.
A) Astrocyte cultures were treated with forskolin (1 µM) for one hour before imaging. The left panel shows the
maximum intensity projection of a 20-minute recording of spontaneous Ca2+ signals as detected by Lck-GCaMP5
fluorescence. The signals were analyzed by 3D connected components as shown in the right panel to account for the
change in ROI morphology. Each colour represents a discrete Ca2+ transient signal, defined by pixels connected in
the x, y, and t axes. B) Images represent 10-minute maximum intensity projections of the microdomain Ca2+ signals
in Lck-GCaMP5-transfected astrocyte cultures before and after treatment with Yoda1 (1 µM) and Gd3+ (100 µM).
C) Quantification of the effect of Yoda1 and Gd3+ on the frequency of astrocyte Ca2+ signals. Scale bars = 50 µm. n
= 5 cells from independent coverslips. ** p < 0.01 by one-way ANOVA with Bonferroni’s multiple comparison
post hoc test.
Next, Lck-GCaMP5-transfected astrocytes were treated with forskolin and imaged during a 10-
minute application of hypotonic solution. Hypotonic stimulation increased the frequency of Ca2+
transients (baseline: 86 ± 56 events/5 min; hypotonic: 97 ± 58 events/5 min; p < 0.05)(Fig. 4-3A
and B), and these signals were sensitive to Piezo1 siRNA treatment compared to the luciferase
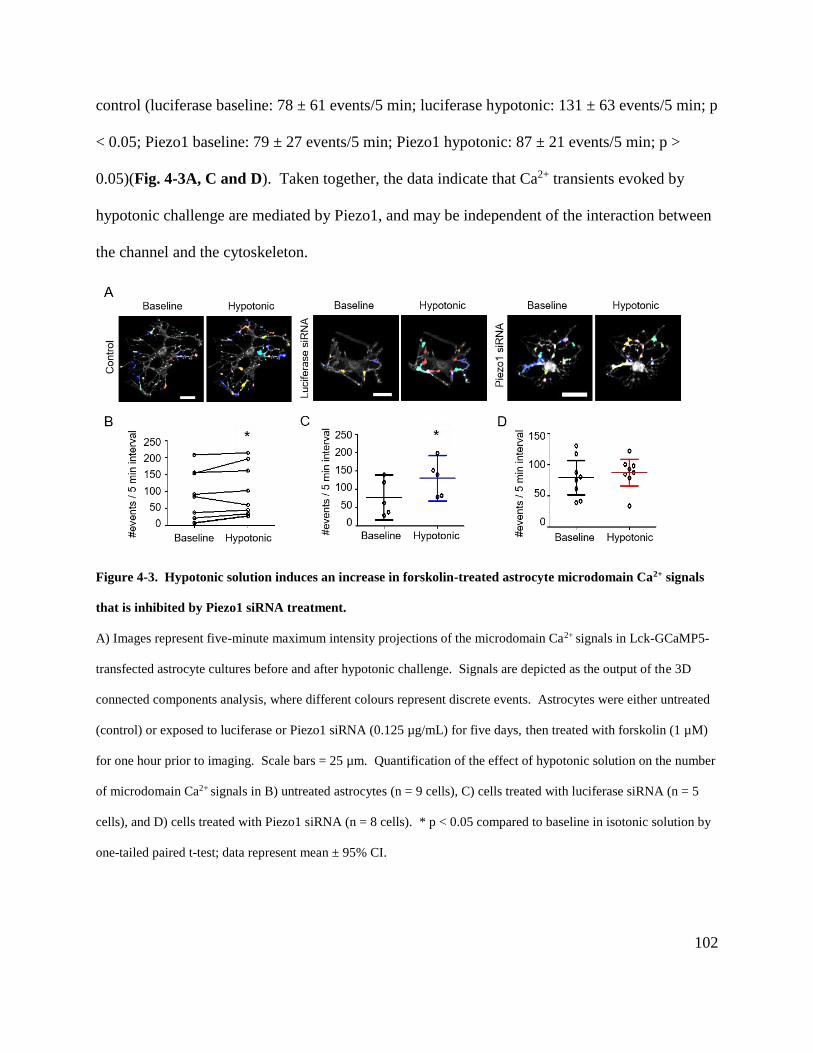
102
control (luciferase baseline: 78 ± 61 events/5 min; luciferase hypotonic: 131 ± 63 events/5 min; p
< 0.05; Piezo1 baseline: 79 ± 27 events/5 min; Piezo1 hypotonic: 87 ± 21 events/5 min; p >
0.05)(Fig. 4-3A, C and D). Taken together, the data indicate that Ca2+ transients evoked by
hypotonic challenge are mediated by Piezo1, and may be independent of the interaction between
the channel and the cytoskeleton.
Figure 4-3. Hypotonic solution induces an increase in forskolin-treated astrocyte microdomain Ca2+ signals
that is inhibited by Piezo1 siRNA treatment.
A) Images represent five-minute maximum intensity projections of the microdomain Ca2+ signals in Lck-GCaMP5-
transfected astrocyte cultures before and after hypotonic challenge. Signals are depicted as the output of the 3D
connected components analysis, where different colours represent discrete events. Astrocytes were either untreated
(control) or exposed to luciferase or Piezo1 siRNA (0.125 µg/mL) for five days, then treated with forskolin (1 µM)
for one hour prior to imaging. Scale bars = 25 µm. Quantification of the effect of hypotonic solution on the number
of microdomain Ca2+ signals in B) untreated astrocytes (n = 9 cells), C) cells treated with luciferase siRNA (n = 5
cells), and D) cells treated with Piezo1 siRNA (n = 8 cells). * p < 0.05 compared to baseline in isotonic solution by
one-tailed paired t-test; data represent mean ± 95% CI.

103
4.3.3 Piezo1 mediates hypotonicity-induced swelling in astrocytes
Our observation that hypotonic challenge can evoke astrocyte Ca2+ transients is intriguing, as
previous studies have suggested that these signals may act as a trigger for the homeostatic
mechanism known as regulatory volume decrease (RVD) (O’Connor and Kimelberg, 1993; Hua
et al., 2010). In RVD, ion channels or transporters are activated to decrease intracellular
osmolarity in response to external hypotonicity, allowing cells to revert to resting volume to
prevent cell lysis and death (Lang et al., 1998). Therefore, to assess whether Piezo1-mediated
Ca2+ transients are involved in regulation of cell volume in response to hypotonic challenge, the
cross-sectional areas of astrocyte soma were measured during baseline in isotonic solution and
following 10 minutes of perfusion with hyponatremia solution. Since control cells did not
appear to swell in the xy-plane, changes in cross-sectional area could not be measured. Imaging
was therefore performed in astrocytes that had been treated with forskolin (Fig. 4-4A). Piezo1
siRNA treatment inhibited hyponatremia-mediated astrocyte swelling relative to the luciferase
control (luciferase siRNA: 121 ± 16%; Piezo siRNA: 102 ± 9%; p < 0.05), indicating that Piezo1
may be required for astrocyte swelling in response to hypotonic stimulus (Fig. 4-4B).
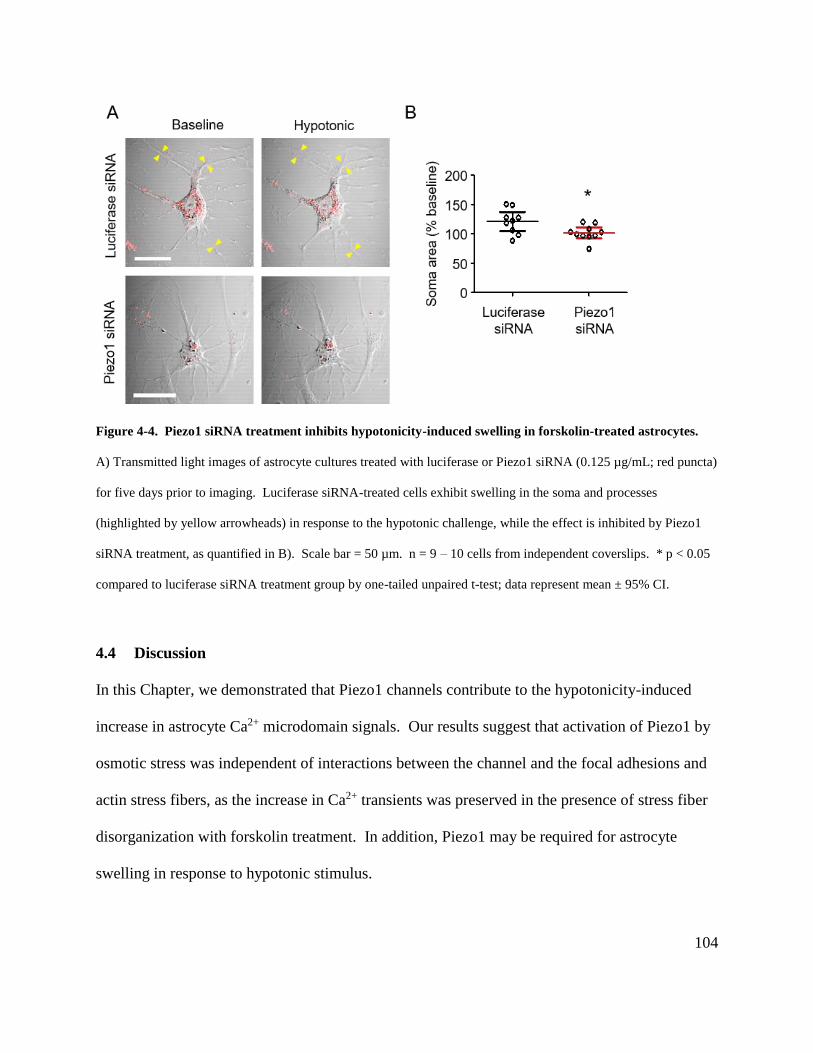
104
Figure 4-4. Piezo1 siRNA treatment inhibits hypotonicity-induced swelling in forskolin-treated astrocytes.
A) Transmitted light images of astrocyte cultures treated with luciferase or Piezo1 siRNA (0.125 µg/mL; red puncta)
for five days prior to imaging. Luciferase siRNA-treated cells exhibit swelling in the soma and processes
(highlighted by yellow arrowheads) in response to the hypotonic challenge, while the effect is inhibited by Piezo1
siRNA treatment, as quantified in B). Scale bar = 50 µm. n = 9 – 10 cells from independent coverslips. * p < 0.05
compared to luciferase siRNA treatment group by one-tailed unpaired t-test; data represent mean ± 95% CI.
4.4 Discussion
In this Chapter, we demonstrated that Piezo1 channels contribute to the hypotonicity-induced
increase in astrocyte Ca2+ microdomain signals. Our results suggest that activation of Piezo1 by
osmotic stress was independent of interactions between the channel and the focal adhesions and
actin stress fibers, as the increase in Ca2+ transients was preserved in the presence of stress fiber
disorganization with forskolin treatment. In addition, Piezo1 may be required for astrocyte
swelling in response to hypotonic stimulus.

105
Our data indicate that in vitro hyponatremia treatment increases the frequency of microdomain
Ca2+ signals in astrocytes via a Piezo1-dependent mechanism. This result is consistent with
earlier studies demonstrating that osmotic stress-induced cytosolic Ca2+ elevations, as detected
by bulk-loaded Ca2+ indicators in astrocytes, is mediated by a Ca2+ influx pathway that is
sensitive to the nonspecific mechanosensitive channel antagonists Gd3+ and ruthenium red (Liu
et al., 2006; Benfenati et al., 2011; Galizia et al., 2012; Mola et al., 2016). Interestingly,
Benfenati et al. (2011) showed that the intracellular Ca2+ increases can also be inhibited by
TRPV4 siRNA treatment, suggesting that TRPV4 is involved in mediating Ca2+ influx following
hypotonic challenge. While our previous data suggest that TRPV4 does not mediate spontaneous
Ca2+ transients (Section 2.3.6), it is possible that TRPV4, or other mechanosensitive channels,
are activated in addition to Piezo1 by astrocyte swelling.
To examine the contribution of the interaction between Piezo1 and the cytoskeleton to channel
activation by hypotonic stress, we studied the effect of cytoskeleton disorganization with
forskolin pre-treatment on the hypotonicity-induced Ca2+ microdomain signals. Despite Piezo1
redistribution to the processes of stellate astrocytes, its pharmacology and role in mediating Ca2+
signals in response to hypotonic stimulus were unaltered by forskolin, suggesting that Piezo1
basal function and activation by osmotic swelling can occur independently of interactions with
focal adhesion proteins or actin stress fibers. This result is supported by previous studies
showing Piezo1 activation by osmotic gradient when the channel was reconstituted in
cytoskeleton-free lipid bilayers (Syeda et al., 2016). However, our data do not rule out
cytoskeleton involvement in astrocyte Piezo1 activation by other types of mechanical stimuli in
vivo.

106
What is the functional significance of these hypotonicity-induced Ca2+ microdomain signals?
Previous studies have observed that astrocytes, when exposed to hypotonic solution, exhibit an
initial swelling phase followed by a return to original cell volume in a homeostatic process
termed regulatory volume decrease (RVD). Several groups have proposed that RVD may be
triggered by stretch-induced activation of mechanosensitive ion channels that then permit Ca2+
entry, either through the channels directly or by depolarizing the cell to activate voltage-gated
Ca2+ channels. Subsequent opening of Ca2+-activated channels and volume-regulated anion
channels leads to net efflux of K+ and Cl- to regulate osmolyte balance across the membrane,
allowing cells to revert to resting volume (Lang et al., 1993, 1998; O’Connor and Kimelberg,
1993). Consistent with this idea, removal of extracellular Ca2+ has been shown to abolish
hypotonicity-induced Ca2+ signals and RVD in astrocytes (O’Connor and Kimelberg, 1993; Hua
et al., 2010). However, other studies have indicated that RVD can occur independently of Ca2+
influx (Morales-Mulia et al., 1998; Mola et al., 2016). Therefore, the role of Ca2+ in mediating
RVD is still incompletely characterized.
We reasoned that if hypotonicity-induced Ca2+ microdomain transients were involved in
mediating RVD, then inhibition of the Ca2+ signals with Piezo1 siRNA treatment may be
associated with impaired volume homeostasis, leading to exacerbated cell swelling.
Surprisingly, our results indicated that swelling was inhibited by Piezo1 knockdown. While
Piezo1 activity may be required for astrocyte swelling, we did not observe any appreciable
increases in cell volume following application of the Piezo1 agonist Yoda1 (Section 4.3.2),
suggesting that Piezo1-mediated Ca2+ signaling alone does not induce cell swelling. It is
possible that Piezo1 may act synergistically with other molecules or cellular pathways as a

107
permissive signal for swelling. Another possible explanation is that Piezo1 siRNA treatment
may have altered aquaporin-4 expression, thereby inhibiting water influx and astrocyte swelling
in response to hypoosmotic stress.
One limitation of our experiments is that the soma cross-sectional area may not sensitive enough
to reflect true changes in cell volume, as swelling in the z-axis would not be measurable. Other
groups have loaded astrocytes with fluorescent dyes and quantified cell swelling as a function of
dye intensity decrease (Liu et al., 2006; Benfenati et al., 2011; Mola et al., 2016). However, the
relationship between dye intensity and cell volume is only linear for a specific range, and dye
efflux may occur due to active transport or leak when the cell is swollen (Kimelberg et al., 1993;
Liu et al., 2006). An alternate strategy relies on impedance-based methods to monitor cell
volume in real time, where the electrical resistance of the cells is inversely related to cell volume
(O’Connor et al., 1993; Hua et al., 2010). These techniques may provide a more accurate
measure of cell volume, particularly for flat polygonal astrocytes that do not appreciably swell in
the xy-plane.
Another complexity of measuring RVD is the heterogeneity of the phenomenon. For instance,
reports of the time it took for RVD to return astrocytes to baseline volume varied from ~1 minute
to ~30 minutes (Hua et al., 2010; Mola et al., 2016). Furthermore, RVD dependence on Ca2+
influx is controversial, as some groups still observed RVD in the absence of external Ca2+ (Mola
et al., 2016). While the parameters underlying these differences is unclear, presentation of RVD
may be affected by cell culture, experimental temperature, perfusion speed, magnitude of
hypotonic challenge, and technique used to monitor cell volume. Therefore, future studies can

108
be conducted to optimize the experimental parameters to examine the contribution of Piezo1 to
RVD.
Intriguingly, previous studies have found that RVD in keratinocytes and CHO cells is dependent
on interactions between TRPV4 and the actin cytoskeleton (Becker et al., 2009). This suggests
that RVD may be impaired in forskolin-treated astrocytes used in our experiments. It would be
interesting to study the contribution of Piezo1 to hypotonic-evoked RVD with astrocytes in vivo,
where putative interactions between the cytoskeleton and the ECM are preserved.

109
Chapter 5: Conclusion
5.1 Summary of research findings
In this thesis, I provide evidence that the activation of mechanosensitive cation channel Piezo1
contributes to spontaneous microdomain Ca2+ signals in rodent astrocytes in both cell culture and
acute brain slices. The Piezo1 agonist Yoda1 increased the frequency of microdomain Ca2+
transients, while these were inhibited by the mechanosensitive channel antagonists GsMTx4 and
Gd3+. The pharmacological data were supported by the reduction of Yoda1-induced Ca2+ signals
after siRNA-mediated Piezo1 knockdown. Immunostaining indicated that Piezo1 expression is
localized to subcellular compartments relevant to mechanosensation. Piezo1 immunoreactivity
was distributed as distinct plasma membrane clusters and associated with focal adhesion and
stress fiber compartments in cultured astrocytes. Piezo1 channels appeared to be concentrated
within the endfeet processes of brain slice astrocytes. Furthermore, Piezo1-mediated Ca2+
microdomain signals were activated by an osmotic stress model of astrocyte swelling. Taken
together, the data demonstrate that cultured and brain slice astrocytes express functional Piezo1
channels, suggesting that astrocyte Ca2+ signaling may play a role in integrating mechanical
stimuli to regulate physiological processes in the brain.
5.2 Research significance
Recent advances in techniques used to study astrocyte Ca2+ signaling have revealed a previously
unappreciated type of spontaneous Ca2+ signal localized to the microdomains of fine astrocytic
processes. Recent work from our lab and others has demonstrated that these signals depend on
transmembrane Ca2+ influx (Shigetomi et al., 2012; Rungta et al., 2016), but the underlying

110
molecular mechanisms have not been fully characterized. In this dissertation, I combined Ca2+
imaging with siRNA-mediated gene silencing and pharmacological approaches in an astrocyte
culture model, and identified the Piezo1 channel as a novel Ca2+ entry pathway that contributes
to the Ca2+ microdomain signals. Previous studies have shown that spontaneous Ca2+ transients
modulate basal astrocyte Ca2+ levels, with implications for regulation of synaptic activity and
arteriole tone (Shigetomi et al., 2012, 2013b; Rosenegger et al., 2015). As Piezo1 is a
mechanosensitive channel, this finding raises the intriguing possibility that astrocytes may serve
as a transducer of mechanical stimuli to affect neuronal and vascular function in the brain.
Our discovery of functional Piezo1 channels in astrocytes extends the repertoire of mechanically
activated channels expressed in these cells. Piezo1 appears to share several characteristics with
other astrocyte mechanosensitive channels; for instance, TRPV4 is also distributed in astrocyte
endfeet, and mediates Ca2+ signals in response to osmotic stress (Benfenati et al., 2007, 2011).
However, one caveat to interpreting these previous studies is their use of non-selective
pharmacological blockers such as Gd3+ and ruthenium red to assign function to a specific
channel (Benfenati et al., 2007). These tools block both TRPV4 and Piezo1 as well as other
channels, and therefore cannot distinguish between the contributions of Piezo1 and TRPV4. In
our experiments, we showed that astrocyte Ca2+ microdomain transients in cultured astrocytes
are sensitive to GsMTx4, but not to the selective TRPV4 antagonist HC067047, suggesting that
different mechanosensitive channels may have distinct functions in the same cell. This idea is
supported by studies comparing different modes of mechanical stimulation in chondrocytes,
which also express both TRPV4 and Piezo1; deflection of cell-substrate contact points activated
both Piezo1- and TRPV4-mediated currents, while membrane stretch only activated Piezo1

111
(Servin-Vences et al., 2017). Furthermore, Miyamoto et al. (2014) demonstrated that Piezo1 was
more sensitive than TRPV4 in detecting stretch stimulation of urothelial cells as measured by
intracellular Ca2+ elevations. Therefore, it is likely that Piezo1 acts in a complementary manner
with other mechanosensitive channels to refine astrocyte sensitivity to different types and
magnitudes of mechanical force.
In this thesis, we presented a novel method for quantitative analysis of microdomain Ca2+
signals. We observed that Lck-GCaMP5-transfected cultured astrocytes exhibited two types of
spontaneous Ca2+ signals: transient, spatially restricted Ca2+ microdomain signals, and long-
lasting Ca2+ waves that expanded and contracted from their point of origin. Since these signals
have different spatiotemporal properties and Ca2+ sources, we developed an algorithm to analyze
them separately. A key feature in our analysis algorithm is the use of the 3D connected
components function in Matlab, which identified discrete Ca2+ signals and allowed them to be
ranked according to total area. Thus Ca2+ waves could be selectively masked using a size
threshold while preserving the microdomain signals. In addition, up to 35 distinct microdomains
were observed in each cell, requiring the development of a strategy to objectively identify and
analyze the Ca2+ signals from these regions. An algorithm was written to automatically detect
these regions of interest based on fluorescence intensity, define Ca2+ transients, and calculate
their frequency, amplitude and duration. Interestingly, our independently developed algorithm
shares a number of similar features with recently published methods for quantifying
microdomain signals (Srinivasan et al., 2015; Agarwal et al., 2017), and the convergence of the
results further supports the reliability of our analysis approach. However, our algorithm has not
yet been tested on astrocytes in vivo, and it remains to be determined whether adaptations such as

112
masking the cell soma or large proximal processes are required to optimize the analysis for in
vivo applications.
The immunostaining data provide the first evidence of Piezo1 distribution to subcellular
localizations relevant to mechanosensing in cultured astrocytes. We observed that Piezo1
immunoreactivity formed discrete clusters at the plasma membrane, where the channels are well-
situated to detect membrane stretch. These clusters bore a striking resemblance to the
spontaneous and Yoda1-activated Ca2+ microdomain signals, and were sensitive to siRNA-
mediated Piezo1 knockdown. In addition, the data presented in Chapter 4 indicated that the
hypoosmotic stress model of cell swelling was associated with increased frequency of Ca2+
microdomain transients, and that was inhibited by Piezo1 siRNA treatment. Taken together, the
data suggest that Piezo1 channels expressed in the astrocyte plasma membrane may function as a
membrane stretch detector, where spontaneous Ca2+ signals may reflect basal Piezo1 activation
by resting membrane tension and with changes in Ca2+ signaling frequency encoding information
such as the rate, onset, or magnitude of cell swelling.
In cultured astrocytes, Piezo1 immunoreactivity was identified in subcellular compartments
associated with focal adhesions and actin stress fibers. We examined the relevance of this
putative channel – cytoskeleton interaction to hypotonic stress-induced Piezo1 activation by
treating the cells with forskolin. Hypotonic stimulation of forskolin-treated astrocytes still
increased microdomain Ca2+ signal frequency. However, we did not observe differences in
Piezo1-mediated Ca2+ microdomain signaling changes in response to hypotonic stimulation in
forskolin-treated versus untreated cells, despite striking changes in astrocyte morphology,

113
redistribution of plasma membrane Piezo1-positive clusters to the processes, and loss of Piezo1
immunoreactivity distributed to focal adhesion and actin stress fiber structures. As such, our
data indicate that at least in cultured astrocytes, the interactions with focal adhesions and actin
stress fibers are not a major contributor to Piezo1 activation by hypotonic stress, and suggest that
Piezo1 may mediate different functions depending on its specific subcellular location.
Piezo1 function may also depend on the developmental or proliferative state of the cell. This
interesting observation was recently described in Madin Darby canine kidney epithelial cells by
Gudipaty et al. (2017), where Piezo1 was shown to maintain epithelial cell number homeostasis
by matching the rate of cell division to the rate of cell death. Piezo1 was demonstrated to be
involved in sensing crowding in regions of high cell density and stretching in regions of low cell
density to stimulate cell extrusion and cell division, respectively (Gudipaty et al., 2017).
Therefore, Piezo1 may also mediate different functions at different timepoints of development,
or following damage where astrocytes become reactive and proliferate. These observations will
guide further experiments on Piezo1 function under various physiological and pathological states
in vivo. Preliminary experiments described in Section 5.3 indicate that these are important
considerations, as Piezo1 expression in astrocytes is upregulated during early postnatal
development and after stroke damage.
5.3 Future directions
The identification of Piezo1 as a mammalian mechanosensitive channel in 2010 has led to rapid
progress in the field of mammalian mechanotransduction. Increasingly higher resolution
structures of Piezo1 provided a better understanding of the structural basis underlying channel

114
function (Guo and MacKinnon, 2017; Saotome et al., 2017; Zhao et al., 2018), and studies have
begun to characterize protein partners that interact with Piezo1 to regulate channel activation
(Zhang et al., 2017). Recent studies have demonstrated novel roles for Piezo1 in mediating
mechanosensation in different organ systems (Liang et al., 2017; Rode et al., 2017), and exciting
possibilities exist for further research on the contributions of Piezo1 to physiological and
pathological conditions.
5.3.1 Transgenic mice for investigating Piezo1-mediated Ca2+ signaling in vivo
In this dissertation, we utilized an astrocyte culture model to test pharmacological and genetic
tools for modulating Piezo1 activity, and to develop an analysis algorithm for quantifying their
effects, culminating in the finding that Piezo1 contributes to spontaneous Ca2+ microdomain
signals in astrocytes in vitro. A next step to understanding the functional role of Piezo1 in
astrocyte physiology is to determine whether Piezo1 also contributes to astrocyte microdomain
Ca2+ signals in vivo. Preliminary observations from our lab indicate that endfoot Ca2+ signaling
is enhanced by the Piezo1 activator Yoda1 in brain slice astrocytes. Furthermore, preliminary
data from GCaMP7 mouse brain slices, where the GECI is expressed in both neurons and
astrocytes, have shown spontaneous Ca2+ signals that are inhibited by Gd3+ in astrocyte
compartments identified by SR101 staining. Recently, Srinivasan et al. (2016) generated a
transgenic mouse line in which the Lck-GCaMP6f GECI was expressed under the astrocyte-
specific Aldh1l1 promoter, resulting in a mouse model where spontaneous Ca2+ microdomain
transients could be monitored in fine astrocyte processes throughout the brain. This mouse
model is likely well-suited for examining whether these spontaneous signals are also sensitive to
pharmacological treatments or siRNA-mediated knockdown targeting Piezo1 in vivo.

115
Previous studies have demonstrated that constitutive Piezo1 knockouts are embryonically lethal
(Ranade et al., 2014). Therefore, another important tool to study the contribution of Piezo1 to
astrocyte function is the generation of transgenic mouse lines wherein conditional Piezo1
knockout can be achieved in astrocytes. For example, the Aldh1l1 promoter line contains a
Cre/ERT2 insertion at its start codon (Srinivasan et al., 2016) and this Aldh1l1-Cre/ERT2 mouse
line could be crossed with mice developed by the Patapoutian lab in which Piezo1 is flanked by
loxP sites, resulting in a mouse model where astrocyte-specific deletion of Piezo1 could induced
by tamoxifen administration. Thus, this model could be used to investigate the functional role of
astrocyte Piezo1 at different developmental stages, as well as during different pathological states.
5.3.2 Role of Piezo1 in regulation of cerebral vasculature tone
Our preliminary brain slice Ca2+ imaging and immunostaining suggest that astrocyte Piezo1
expression may be concentrated at endfeet processes that surround the cerebral vasculature. As
we consider the physiological role of Piezo1-mediated mechanotransduction in the context of the
brain, it is tempting to speculate that Piezo1 may be involved in sensing mechanical forces
exerted by dilating or constricting blood vessels, resulting in changes to astrocyte Ca2+
microdomain activity that can stimulate release of vasoactive compounds to regulate brain blood
flow. This idea is supported by studies demonstrating that disruption of astrocyte Ca2+ signals
with BAPTA is associated with alterations in prostaglandin release and arteriole tone
(Rosenegger et al., 2015). The role of Piezo1 in detecting vascular changes can be studied by
measuring the effect of agents, such as the α2 adrenoceptor agonist clonidine, that induce
vasoconstriction by acting on smooth muscle cells. The direct stimulation of vascular
constriction by this method should stretch the astrocyte endfeet, and thereby would be predicted

116
to cause Piezo1-dependent endfoot Ca2+ transients in transgenic mice expressing Lck-GCaMP6f
driven by the Aldh1l1 promoter. These vasomotion-induced responses in astrocyte endfeet can
then be compared to the responses of astrocytes where Piezo1 is pharmacologically blocked or
genetically deleted.
5.3.3 Role of Piezo1 in astrocyte differentiation
As described in Chapter 3, our observations of Piezo1 localization to focal adhesions and actin
stress fibers in cultured astrocytes suggest a role for the mechanosensitive channel in detecting
extracellular matrix (ECM) rigidity, and accumulating evidence indicates that this link to matrix
mechanics may have important consequences for cell differentiation. For instance, Pathak et al.
(2014) showed that neural stem cells exhibit Piezo1-mediated spontaneous Ca2+ microdomain
signals dependent on substrate stiffness, and pharmacological inhibition or siRNA-induced
knockdown of Piezo1 resulted in increased differentiation into astrocytes and decreased neuron
formation. In addition, Sugimoto et al. (2017) demonstrated that the hydrostatic pressure of
extracellular fluid induced differentiation of mesenchymal stem cells to osteoblasts, and this
process required Piezo1-mediated regulation of bone morphogenetic protein 2 (BMP2)
expression. Interestingly, BMP2 upregulation has also been shown to promote differentiation of
oligodendroglial-astroglial progenitor cells into astrocytes during development (Mabie et al.,
1997). Our lab has recently investigated Piezo1 expression in the brain using a transgenic mouse
line where Piezo1 is fused to the fluorescent reporter tdTomato (Ranade et al., 2014), and the
preliminary immunoblotting data indicate that Piezo1 expression levels are age-dependent.
Notably, Piezo1 expression was highest during early development (postnatal day 2), with levels
in the cortex and hippocampus declining to the lower levels found in the adult by postnatal day

117
14 – 21; importantly, this early period of high Piezo1 expression correlates with an active stage
of gliogenesis (Bushong et al., 2004; Freeman, 2010). It is therefore plausible that Piezo1-
mediated spontaneous Ca2+ transients may be relaying information about the mechanical
properties of the ECM to guide astrocyte differentiation. This hypothesis can be tested in the
conditional astrocyte Piezo1 knockout mice, where tamoxifen injections can be administered at
different timepoints throughout development to assess the contribution of Piezo1 to astrocyte
differentiation. Astrocytes from adult Aldh1l1-Lck-GCaMP6f mice can also be cultured on
substrates of varying stiffness, followed by Ca2+ imaging in combination with Piezo1
pharmacology, to examine whether Piezo1 is involved in sensing ECM rigidity in mature
astrocytes.
5.3.4 Role of Piezo1 in transmitter release
Previous studies have indicated that activation of mechanosensitive channels can lead to
downstream efflux of various transmitters. For example, stretch stimulation of cultured
astrocytes has been demonstrated to induce Ca2+ influx and endothelin-1 release, both of which
were inhibited by the mechanosensitive channel inhibitor GsMTx4 (Ostrow et al., 2000).
Similarly, Piezo1-dependent Ca2+ elevation and ATP efflux have been identified in red blood
cells (Cinar et al., 2015). Preliminary studies from our lab suggest that treatment with the Piezo1
agonist Yoda1 is associated with ATP release from cultured astrocytes, but its mechanism of
efflux and functional significance are currently unknown. ATP has been shown to contribute to
various types of astrocyte Ca2+ signaling in vivo, such as the spontaneous Ca2+ waves in
Bergmann glia of the cerebellum (Nimmerjahn et al., 2009), raising the possibility that Piezo1
activity may also be involved in mediating different Ca2+ signals in distinct brain regions.

118
However, the physiological stimuli that evoke Piezo1-mediated ATP release from in vivo
astrocytes remain to be determined.
5.3.5 Piezo1 and pathology
Piezo1 was initially observed in astrocytes associated with β-amyloid plaques (Satoh et al.,
2006). Although its function in Alzheimer’s disease pathology has not been characterized,
Piezo1 expression in astrocytes has since been found in other pathological conditions. For
example, Choi et al. (2015) performed quantitative PCR (qPCR) on astrocytes isolated from the
optic nerve head and observed an increase in Piezo1 mRNA three days following optic nerve
crush injury. Furthermore, using a photothrombotic stroke model, our lab has found elevated
levels of Piezo1 mRNA and protein in the glial scar astrocytes lining the borders of the infarct
tissue, as quantified by qPCR, immunoblotting, and immunohistochemistry seven days post-
stroke. Given the increase in Piezo1 expression in various pathological states, it is possible that
Piezo1 is upregulated in reactive astrocytes as a general response to CNS injury.
Reactive gliosis encompasses a broad spectrum of cellular responses that ranges from reversible
gene expression changes to formation of glial scars and permanent tissue reorganization
(Sofroniew and Vinters, 2010). While glial scars are considered to be an adaptive response for
creating a barrier around wounded brain tissue, it is unclear how reactive astrocytes determine
the boundaries of the scar (Burda et al., 2016). Interestingly, previous studies have identified
Piezo1 as an activator of integrin receptors; these molecules are involved in cell – extracellular
matrix adhesion, and have also been implicated in cell migration (McHugh et al., 2010;
Huttenlocher and Horwitz, 2011). Indeed, siRNA-mediated Piezo1 knockdown has been

119
associated with altered migration of cancer cells (McHugh et al., 2012; Yang et al., 2014).
Furthermore, Piezo1 has been shown to regulate retinal ganglion cell axonal pathfinding by
sensing brain stiffness and promoting axonal growth toward softer tissue (Koser et al., 2016).
Brain stiffness is altered in pathological conditions such as Alzheimer’s disease and stroke
(Murphy et al., 2011; Xu et al., 2013), and we speculate that Piezo1 may be involved in detecting
differences in the mechanical properties of brain tissue to guide reactive astrocyte migration,
ensuring that the resulting glial scar border is properly located to segregate potentially viable
tissue from damaged tissue. To address this possibility, glial scar formation can be compared
between wild-type and astrocyte-specific Piezo1 knockout mice in different CNS pathologies,
such as following stroke or traumatic brain injury.

120
References
Addepalli H, Meena, Peng CG, Wang G, Fan Y, Charisse K, Jayaprakash KN, Rajeev KG,
Pandey RK, Lavine G, Zhang L, Jahn-Hofmann K, Hadwiger P, Manoharan M, Maier MA
(2010) Modulation of thermal stability can enhance the potency of siRNA. Nucleic Acids
Res 38:7320–7331 Available at: http://www.ncbi.nlm.nih.gov/pubmed/20610434 [Accessed
November 12, 2017].
Agarwal A, Wu P-H, Hughes EG, Fukaya M, Tischfield MA, Langseth AJ, Wirtz D, Bergles DE
(2017) Transient Opening of the Mitochondrial Permeability Transition Pore Induces
Microdomain Calcium Transients in Astrocyte Processes. Neuron 93:587–605.e7 Available
at: http://www.ncbi.nlm.nih.gov/pubmed/28132831 [Accessed January 31, 2018].
Akerboom J et al. (2012) Optimization of a GCaMP Calcium Indicator for Neural Activity
Imaging. J Neurosci 32:13819–13840 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/23035093 [Accessed January 31, 2018].
Akinc A et al. (2008) A combinatorial library of lipid-like materials for delivery of RNAi
therapeutics. Nat Biotechnol 26:561–569 Available at:
http://www.nature.com/doifinder/10.1038/nbt1402 [Accessed November 13, 2017].
Albuisson J, Murthy SE, Bandell M, Coste B, Louis-Dit-Picard H, Mathur J, Fénéant-Thibault
M, Tertian G, de Jaureguiberry J-P, Syfuss P-Y, Cahalan S, Garçon L, Toutain F, Simon
Rohrlich P, Delaunay J, Picard V, Jeunemaitre X, Patapoutian A (2013) Dehydrated
hereditary stomatocytosis linked to gain-of-function mutations in mechanically activated
PIEZO1 ion channels. Nat Commun 4:1884 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/23695678 [Accessed December 27, 2017].
Alloui A, Zimmermann K, Mamet J, Duprat F, Noël J, Chemin J, Guy N, Blondeau N, Voilley

121
N, Rubat-Coudert C, Borsotto M, Romey G, Heurteaux C, Reeh P, Eschalier A, Lazdunski
M (2006) TREK-1, a K+ channel involved in polymodal pain perception. EMBO J
25:2368–2376 Available at: http://www.ncbi.nlm.nih.gov/pubmed/16675954 [Accessed
December 19, 2017].
Alper SL (2017) Genetic Diseases of PIEZO1 and PIEZO2 Dysfunction. Curr Top Membr
79:97–134 Available at: http://linkinghub.elsevier.com/retrieve/pii/S1063582317300017
[Accessed January 31, 2018].
Andolfo I et al. (2013) Multiple clinical forms of dehydrated hereditary stomatocytosis arise
from mutations in PIEZO1. Blood 121:3925–3935, S1-12 Available at:
http://www.bloodjournal.org/cgi/doi/10.1182/blood-2013-02-482489 [Accessed January 31,
2018].
Andriezen WL (1893) The Neuroglia Elements In The Human Brain. Br Med J 2:227–230
Available at: http://www.jstor.org.ezproxy.library.ubc.ca/stable/20225294 [Accessed
December 30, 2017].
Arani A, Murphy MC, Glaser KJ, Manduca A, Lake DS, Kruse SA, Jack CR, Ehman RL, Huston
J, Huston J, 3rd (2015) Measuring the effects of aging and sex on regional brain stiffness
with MR elastography in healthy older adults. Neuroimage 111:59–64 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25698157 [Accessed January 7, 2018].
Araque A, Navarrete M (2010) Glial cells in neuronal network function. Philos Trans R Soc
Lond B Biol Sci 365:2375–2381 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20603358 [Accessed December 31, 2017].
Attwell D, Buchan AM, Charpak S, Lauritzen M, MacVicar BA, Newman EA (2010) Glial and
neuronal control of brain blood flow. Nature 468:232–243 Available at:

122
http://www.ncbi.nlm.nih.gov/pubmed/21068832 [Accessed December 31, 2017].
Bae C, Gnanasambandam R, Nicolai C, Sachs F, Gottlieb PA (2013a) Xerocytosis is caused by
mutations that alter the kinetics of the mechanosensitive channel PIEZO1. Proc Natl Acad
Sci U S A 110:E1162-8 Available at: http://www.ncbi.nlm.nih.gov/pubmed/23487776
[Accessed December 24, 2017].
Bae C, Gottlieb PA, Sachs F (2013b) Human PIEZO1: Removing Inactivation. Biophys J
105:880–886 Available at: http://www.ncbi.nlm.nih.gov/pubmed/23972840 [Accessed
January 31, 2018].
Bae C, Sachs F, Gottlieb PA (2011) The mechanosensitive ion channel Piezo1 is inhibited by the
peptide GsMTx4. Biochemistry 50:6295–6300 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/21696149 [Accessed December 25, 2017].
Bautista DM, Jordt S-E, Nikai T, Tsuruda PR, Read AJ, Poblete J, Yamoah EN, Basbaum AI,
Julius D (2006) TRPA1 Mediates the Inflammatory Actions of Environmental Irritants and
Proalgesic Agents. Cell 124:1269–1282 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/16564016 [Accessed December 19, 2017].
Bavi N, Nikolaev YA, Bavi O, Ridone P, Martinac AD, Nakayama Y, Cox CD, Martinac B
(2017) Principles of Mechanosensing at the Membrane Interface. In, pp 85–119. Springer,
Singapore. Available at: http://link.springer.com/10.1007/978-981-10-6244-5_4 [Accessed
January 31, 2018].
Bazargani N, Attwell D (2016) Astrocyte calcium signaling: the third wave. Nat Neurosci
19:182–189 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26814587.
Becker D, Bereiter-Hahn J, Jendrach M (2009) Functional interaction of the cation channel
transient receptor potential vanilloid 4 (TRPV4) and actin in volume regulation. Eur J Cell

123
Biol 88:141–152 Available at:
http://www.sciencedirect.com.ezproxy.library.ubc.ca/science/article/pii/S017193350800144
1 [Accessed December 3, 2017].
Becker D, Blase C, Bereiter-Hahn J, Jendrach M (2005) TRPV4 exhibits a functional role in cell-
volume regulation. J Cell Sci 118:2435–2440 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/15923656 [Accessed January 8, 2018].
Benarroch EE (2015) Extracellular matrix in the CNS: Dynamic structure and clinical
correlations. Neurology 85:1417–1427 Available at:
http://www.neurology.org/lookup/doi/10.1212/WNL.0000000000002044 [Accessed
January 31, 2018].
Benfenati V, Amiry-Moghaddam M, Caprini M, Mylonakou MN, Rapisarda C, Ottersen OP,
Ferroni S (2007) Expression and functional characterization of transient receptor potential
vanilloid-related channel 4 (TRPV4) in rat cortical astrocytes. Neuroscience 148:876–892
Available at: http://linkinghub.elsevier.com/retrieve/pii/S0306452207007944 [Accessed
January 31, 2018].
Benfenati V, Caprini M, Dovizio M, Mylonakou MN, Ferroni S, Ottersen OP, Amiry-
Moghaddam M (2011) An aquaporin-4/transient receptor potential vanilloid 4
(AQP4/TRPV4) complex is essential for cell-volume control in astrocytes. Proc Natl Acad
Sci 108:2563–2568 Available at: http://www.ncbi.nlm.nih.gov/pubmed/21262839
[Accessed December 1, 2017].
Bezzi P, Volterra A (2001) A neuron-glia signalling network in the active brain. Curr Opin
Neurobiol 11:387–394 Available at: http://www.ncbi.nlm.nih.gov/pubmed/11399439
[Accessed January 31, 2018].

124
Biagi BA, Enyeart JJ (1990) Gadolinium blocks low- and high-threshold calcium currents in
pituitary cells. Am J Physiol 259:C515-20 Available at:
http://www.physiology.org/doi/10.1152/ajpcell.1990.259.3.C515 [Accessed December 25,
2017].
Bindocci E, Savtchouk I, Liaudet N, Becker D, Carriero G, Volterra A (2017) Three-dimensional
Ca2+ imaging advances understanding of astrocyte biology. Science 356:eaai8185
Available at: http://www.sciencemag.org/lookup/doi/10.1126/science.aai8185 [Accessed
January 31, 2018].
Blumenthal NR, Hermanson O, Heimrich B, Shastri VP (2014) Stochastic nanoroughness
modulates neuron–astrocyte interactions and function via mechanosensing cation channels.
Proc Natl Acad Sci 111:16124–16129 Available at:
http://www.pnas.org/lookup/doi/10.1073/pnas.1412740111 [Accessed January 31, 2018].
Bocksteins E, J. A, P. D, J. D (2012) Immunostaining of Voltage-Gated Ion Channels in Cell
Lines and Neurons – Key Concepts and Potential Pitfalls. In: Applications of
Immunocytochemistry. InTech. Available at:
http://www.intechopen.com/books/applications-of-immunocytochemistry/immunostaining-
of-cell-lines-and-neurons-key-concepts-and-potential-pitfalls [Accessed December 9, 2017].
Bonder DE, McCarthy KD (2014) Astrocytic Gq-GPCR-linked IP3R-dependent Ca2+ signaling
does not mediate neurovascular coupling in mouse visual cortex in vivo. J Neurosci
34:13139–13150 Available at:
http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.2591-14.2014 [Accessed January
31, 2018].
Borbiro I, Badheka D, Rohacs T (2015) Activation of TRPV1 channels inhibits

125
mechanosensitive Piezo channel activity by depleting membrane phosphoinositides. Sci
Signal 8:ra15-ra15 Available at: http://www.ncbi.nlm.nih.gov/pubmed/25670203 [Accessed
December 20, 2017].
Borbiro I, Rohacs T (2017) Regulation of Piezo Channels by Cellular Signaling Pathways. In:
Current topics in membranes, pp 245–261 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/28728819 [Accessed December 20, 2017].
Bowman CL, Ding JP, Sachs F, Sokabe M (1992) Mechanotransducing ion channels in
astrocytes. Brain Res 584:272–286 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/1381266 [Accessed January 8, 2018].
Bowman CL, Gottlieb PA, Suchyna TM, Murphy YK, Sachs F (2007) Mechanosensitive ion
channels and the peptide inhibitor GsMTx-4: History, properties, mechanisms and
pharmacology. Toxicon 49:249–270 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/17157345 [Accessed January 31, 2018].
Burda JE, Bernstein AM, Sofroniew M V. (2016) Astrocyte roles in traumatic brain injury. Exp
Neurol 275:305–315 Available at: http://www.ncbi.nlm.nih.gov/pubmed/25828533
[Accessed February 1, 2018].
Bushong EA, Martone ME, Ellisman MH (2004) Maturation of astrocyte morphology and the
establishment of astrocyte domains during postnatal hippocampal development. Int J Dev
Neurosci 22:73–86 Available at: http://www.ncbi.nlm.nih.gov/pubmed/15036382
[Accessed January 31, 2018].
Cahalan SM, Lukacs V, Ranade SS, Chien S, Bandell M, Patapoutian A (2015) Piezo1 links
mechanical forces to red blood cell volume. Elife 4 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26001274 [Accessed January 31, 2018].

126
Campbell ID, Humphries MJ (2011) Integrin structure, activation, and interactions. Cold Spring
Harb Perspect Biol 3 Available at: http://www.ncbi.nlm.nih.gov/pubmed/21421922
[Accessed January 7, 2018].
Chalfie M (2009) Neurosensory mechanotransduction. Nat Rev Mol Cell Biol 10:44–52
Available at: http://www.ncbi.nlm.nih.gov/pubmed/19197331 [Accessed December 19,
2017].
Chalfie M, Sulston J (1981) Developmental genetics of the mechanosensory neurons of
Caenorhabditis elegans. Dev Biol 82:358–370 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/7227647 [Accessed December 28, 2017].
Charles AC, Merrill JE, Dirksen ER, Sanderson MJ (1991) Intercellular signaling in glial cells:
calcium waves and oscillations in response to mechanical stimulation and glutamate.
Neuron 6:983–992 Available at: http://www.ncbi.nlm.nih.gov/pubmed/1675864 [Accessed
January 31, 2018].
Chemin J, Patel AJ, Duprat F, Lauritzen I, Lazdunski M, Honoré E (2005) A phospholipid sensor
controls mechanogating of the K+ channel TREK-1. EMBO J 24:44–53 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/15577940 [Accessed December 24, 2017].
Cheng H, Song L-S, Shirokova N, González A, Lakatta EG, Ríos E, Stern MD (1999) Amplitude
Distribution of Calcium Sparks in Confocal Images: Theory and Studies with an Automatic
Detection Method. Biophys J 76:606–617 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/9929467 [Accessed November 13, 2017].
Choi HJ, Sun D, Jakobs TC (2015) Astrocytes in the optic nerve head express putative
mechanosensitive channels. Mol Vis 21:749–766 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26236150 [Accessed January 31, 2018].

127
Cinar E, Zhou S, DeCourcey J, Wang Y, Waugh RE, Wan J (2015) Piezo1 regulates
mechanotransductive release of ATP from human RBCs. Proc Natl Acad Sci U S A
112:11783–11788 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26351678 [Accessed
December 29, 2017].
Clark EC, Thomas D, Br J, Sterns RH (1996) Depletion of glutathione from brain cells in
hyponatremia. Kidney Int 49:470–476 Available at: http://www.kidney-
international.com/article/S0085-2538(15)59360-5/pdf [Accessed December 11, 2017].
Corey DP, Hudspeth AJ (1979) Response latency of vertebrate hair cells. Biophys J 26:499–506
Available at: http://www.ncbi.nlm.nih.gov/pubmed/318064 [Accessed January 31, 2018].
Cornell-Bell AH, Finkbeiner SM, Cooper MS, Smith SJ (1990) Glutamate induces calcium
waves in cultured astrocytes: long-range glial signaling. Science 247:470–473 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/1967852 [Accessed January 31, 2018].
Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, Dubin AE, Patapoutian A
(2010) Piezo1 and Piezo2 are essential components of distinct mechanically activated cation
channels. Science 330:55–60 Available at:
http://www.sciencemag.org/cgi/doi/10.1126/science.1193270 [Accessed January 31, 2018].
Coste B, Murthy SE, Mathur J, Schmidt M, Mechioukhi Y, Delmas P, Patapoutian A (2015)
Piezo1 ion channel pore properties are dictated by C-terminal region. Nat Commun 6:7223
Available at: http://www.ncbi.nlm.nih.gov/pubmed/26008989 [Accessed January 31, 2018].
Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS, Kim SE, Schmidt M, Mathur J,
Dubin AE, Montal M, Patapoutian A (2012) Piezo proteins are pore-forming subunits of
mechanically activated channels. Nature 483:176–181 Available at:
http://www.nature.com/doifinder/10.1038/nature10812 [Accessed December 20, 2017].

128
Cox CD, Bae C, Ziegler L, Hartley S, Nikolova-Krstevski V, Rohde PR, Ng C-A, Sachs F,
Gottlieb P a, Martinac B (2016) Removal of the mechanoprotective influence of the
cytoskeleton reveals PIEZO1 is gated by bilayer tension. Nat Commun 7:1–13 Available at:
http://dx.doi.org/10.1038/ncomms10366.
Cox CD, Bavi N, Martinac B (2017) Origin of the Force. In: Current topics in membranes, pp
59–96 Available at: http://www.ncbi.nlm.nih.gov/pubmed/28728824 [Accessed January 31,
2018].
Crépel V, Panenka W, Kelly ME, MacVicar BA (1998) Mitogen-activated protein and tyrosine
kinases in the activation of astrocyte volume-activated chloride current. J Neurosci
18:1196–1206 Available at: http://www.ncbi.nlm.nih.gov/pubmed/9454830 [Accessed
January 31, 2018].
Dani JW, Chernjavsky A, Smith SJ (1992) Neuronal activity triggers calcium waves in
hippocampal astrocyte networks. Neuron 8:429–440 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/1347996 [Accessed January 31, 2018].
Deleavey GF, Damha MJ (2012) Designing chemically modified oligonucleotides for targeted
gene silencing. Chem Biol 19:937–954 Available at:
http://www.sciencedirect.com/science/article/pii/S1074552112002645 [Accessed November
12, 2017].
Delekate A, Füchtemeier M, Schumacher T, Ulbrich C, Foddis M, Petzold GC (2014)
Metabotropic P2Y1 receptor signalling mediates astrocytic hyperactivity in vivo in an
Alzheimer’s disease mouse model. Nat Commun 5:5422 Available at:
http://www.nature.com/doifinder/10.1038/ncomms6422 [Accessed January 4, 2018].
Delmas P, Hao J, Rodat-Despoix L (2011) Molecular mechanisms of mechanotransduction in

129
mammalian sensory neurons. Nat Rev Neurosci 12:139–153 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/21304548 [Accessed January 31, 2018].
Di Castro MA, Chuquet J, Liaudet N, Bhaukaurally K, Santello M, Bouvier D, Tiret P, Volterra
A (2011) Local Ca2+ detection and modulation of synaptic release by astrocytes. Nat
Neurosci 14:1276–1284 Available at: http://www.nature.com/doifinder/10.1038/nn.2929
[Accessed November 13, 2017].
Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P, Wood JN (2004) Acid-
sensing ion channels ASIC2 and ASIC3 do not contribute to mechanically activated
currents in mammalian sensory neurones. J Physiol 556:691–710 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/14990679 [Accessed December 19, 2017].
Duffy S, MacVicar BA (1995) Adrenergic calcium signaling in astrocyte networks within the
hippocampal slice. J Neurosci 15:5535–5550 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/7643199 [Accessed February 12, 2018].
Dunn KM, Hill-Eubanks DC, Liedtke WB, Nelson MT (2013) TRPV4 channels stimulate Ca2+-
induced Ca2+ release in astrocytic endfeet and amplify neurovascular coupling responses.
Proc Natl Acad Sci U S A 110:6157–6162 Available at:
http://www.pnas.org/cgi/doi/10.1073/pnas.1216514110 [Accessed January 31, 2018].
Elinder F, Arhem P (1994) Effects of gadolinium on ion channels in the myelinated axon of
Xenopus laevis: four sites of action. Biophys J 67:71–83 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/7919027 [Accessed December 25, 2017].
Ermakov YA, Kamaraju K, Sengupta K, Sukharev S (2010) Gadolinium ions block
mechanosensitive channels by altering the packing and lateral pressure of anionic lipids.
Biophys J 98:1018–1027 Available at: http://www.ncbi.nlm.nih.gov/pubmed/20303859

130
[Accessed December 25, 2017].
Ernstrom GG, Chalfie M (2002) Genetics of Sensory Mechanotransduction. Annu Rev Genet
36:411–453 Available at:
http://www.annualreviews.org/doi/10.1146/annurev.genet.36.061802.101708 [Accessed
December 28, 2017].
Faissner A, Pyka M, Geissler M, Sobik T, Frischknecht R, Gundelfinger ED, Seidenbecher C
(2010) Contributions of astrocytes to synapse formation and maturation — Potential
functions of the perisynaptic extracellular matrix. Brain Res Rev 63:26–38 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20096729 [Accessed January 7, 2018].
Faucherre A, Kissa K, Nargeot J, Mangoni ME, Jopling C (2014) Piezo1 plays a role in
erythrocyte volume homeostasis. Haematologica 99:70–75 Available at:
http://www.haematologica.org/cgi/doi/10.3324/haematol.2013.086090 [Accessed January
31, 2018].
Feliciangeli S, Chatelain FC, Bichet D, Lesage F (2015) The family of K2P channels: salient
structural and functional properties. J Physiol 593:2587–2603 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25530075 [Accessed January 8, 2018].
Florence CM, Baillie LD, Mulligan SJ (2012) Dynamic Volume Changes in Astrocytes Are an
Intrinsic Phenomenon Mediated by Bicarbonate Ion Flux Mongin AA, ed. PLoS One
7:e51124 Available at: http://dx.plos.org/10.1371/journal.pone.0051124 [Accessed
November 29, 2017].
Fotiou E, Martin-Almedina S, Simpson MA, Lin S, Gordon K, Brice G, Atton G, Jeffery I, Rees
DC, Mignot C, Vogt J, Homfray T, Snyder MP, Rockson SG, Jeffery S, Mortimer PS,
Mansour S, Ostergaard P (2015) Novel mutations in PIEZO1 cause an autosomal recessive

131
generalized lymphatic dysplasia with non-immune hydrops fetalis. Nat Commun 6:8085
Available at: http://www.ncbi.nlm.nih.gov/pubmed/26333996 [Accessed December 27,
2017].
Freeman MR (2010) Specification and morphogenesis of astrocytes. Science 330:774–778
Available at: http://www.ncbi.nlm.nih.gov/pubmed/21051628 [Accessed January 27, 2018].
Fruman DA, Meyers RE, Cantley LC (1998) PHOSPHOINOSITIDE KINASES. Annu Rev
Biochem 67:481–507 Available at:
http://www.annualreviews.org/doi/10.1146/annurev.biochem.67.1.481 [Accessed January
31, 2018].
Galizia L, Pizzoni A, Fernandez J, Rivarola V, Capurro C, Ford P (2012) Functional interaction
between AQP2 and TRPV4 in renal cells. J Cell Biochem 113:580–589 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/21938744 [Accessed December 1, 2017].
García-Marín V, García-López P, Freire M (2007) Cajal’s contributions to glia research. Trends
Neurosci 30:479–487 Available at:
http://www.sciencedirect.com.ezproxy.library.ubc.ca/science/article/pii/S016622360700182
8?via%3Dihub#bib7 [Accessed December 30, 2017].
Gaub BM, Müller DJ (2017) Mechanical Stimulation of Piezo1 Receptors Depends on
Extracellular Matrix Proteins and Directionality of Force. Nano Lett 17:2064–2072
Available at: http://www.ncbi.nlm.nih.gov/pubmed/28164706 [Accessed January 31, 2018].
Ge J, Li W, Zhao Q, Li N, Chen M, Zhi P, Li R, Gao N, Xiao B, Yang M (2015) Architecture of
the mammalian mechanosensitive Piezo1 channel. Nature 527:64–69 Available at:
http://www.nature.com/doifinder/10.1038/nature15247 [Accessed December 20, 2017].
Geiger B, Spatz JP, Bershadsky AD (2009) Environmental sensing through focal adhesions. Nat

132
Rev Mol Cell Biol 10:21–33 Available at: http://www.ncbi.nlm.nih.gov/pubmed/19197329
[Accessed January 31, 2018].
Geng J, Zhao Q, Zhang T, Xiao B (2017) In Touch With the Mechanosensitive Piezo Channels.
In: Current topics in membranes, pp 159–195 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/28728816 [Accessed January 31, 2018].
Gillespie PG, Walker RG (2001) Molecular basis of mechanosensory transduction. Nature
413:194–202 Available at: http://www.nature.com/doifinder/10.1038/35093011 [Accessed
December 19, 2017].
Giuliani C, Peri A (2014) Effects of Hyponatremia on the Brain. J Clin Med 3:1163–1177
Available at: http://www.ncbi.nlm.nih.gov/pubmed/26237597 [Accessed December 1,
2017].
Gnanasambandam R, Bae C, Gottlieb PA, Sachs F (2015) Ionic Selectivity and Permeation
Properties of Human PIEZO1 Channels. Barnes S, ed. PLoS One 10:e0125503 Available at:
http://dx.plos.org/10.1371/journal.pone.0125503 [Accessed January 31, 2018].
Gnanasambandam R, Ghatak C, Yasmann A, Nishizawa K, Sachs F, Ladokhin AS, Sukharev SI,
Suchyna TM (2017) GsMTx4: Mechanism of Inhibiting Mechanosensitive Ion Channels.
Biophys J 112:31–45 Available at:
http://linkinghub.elsevier.com/retrieve/pii/S0006349516310414 [Accessed January 31,
2018].
Gnatenco C, Han J, Snyder AK, Kim D (2002) Functional expression of TREK-2 K+ channel in
cultured rat brain astrocytes. Brain Res 931:56–67 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/11897089 [Accessed January 31, 2018].
Gordon GRJ, Howarth C, MacVicar BA (2011) Bidirectional control of arteriole diameter by

133
astrocytes. Exp Physiol 96:393–399 Available at:
http://doi.wiley.com/10.1113/expphysiol.2010.053132 [Accessed January 7, 2018].
Gottlieb PA, Bae C, Sachs F (2012) Gating the mechanical channel Piezo1: a comparison
between whole-cell and patch recording. Channels (Austin) 6:282–289 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/22790451 [Accessed December 23, 2017].
Gresch O, Altrogge L (2012) Transfection of difficult-to-transfect primary mammalian cells.
Methods Mol Biol 801:65–74 Available at: http://link.springer.com/10.1007/978-1-61779-
352-3_5 [Accessed January 31, 2018].
Gudipaty SA, Lindblom J, Loftus PD, Redd MJ, Edes K, Davey CF, Krishnegowda V,
Rosenblatt J (2017) Mechanical stretch triggers rapid epithelial cell division through Piezo1.
Nature 543:118–121 Available at: http://www.nature.com/doifinder/10.1038/nature21407.
Guo YR, MacKinnon R (2017) Structure-based membrane dome mechanism for Piezo
mechanosensitivity. Elife 6 Available at: http://www.ncbi.nlm.nih.gov/pubmed/29231809
[Accessed February 9, 2018].
Haim L Ben, Rowitch DH (2017) Functional diversity of astrocytes in neural circuit regulation.
Nat Rev Neurosci 18:31–41 Available at: http://www.ncbi.nlm.nih.gov/pubmed/27904142
[Accessed January 31, 2018].
Hamill OP, McBride DW (1996) The pharmacology of mechanogated membrane ion channels.
Pharmacol Rev 48:231–252 Available at: http://www.ncbi.nlm.nih.gov/pubmed/8804105
[Accessed January 31, 2018].
Hayakawa K, Tatsumi H, Sokabe M (2008) Actin stress fibers transmit and focus force to
activate mechanosensitive channels. J Cell Sci 121:496–503 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/18230647 [Accessed January 16, 2018].

134
Heidenreich M, Zhang F (2016) Applications of CRISPR–Cas systems in neuroscience. Nat Rev
Neurosci 17:36–44 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26656253
[Accessed January 31, 2018].
Henneberger C, Papouin T, Oliet SHR, Rusakov DA (2010) Long-term potentiation depends on
release of d-serine from astrocytes. Nature 463:232–236 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20075918 [Accessed January 4, 2018].
Herculano-Houzel S (2014) The glia/neuron ratio: How it varies uniformly across brain
structures and species and what that means for brain physiology and evolution. Glia
62:1377–1391 Available at: http://www.ncbi.nlm.nih.gov/pubmed/24807023 [Accessed
December 30, 2017].
Hua SZ, Gottlieb PA, Heo J, Sachs F (2010) A mechanosensitive ion channel regulating cell
volume. Am J Physiol Physiol 298:C1424–C1430 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20457830 [Accessed January 31, 2018].
Huttenlocher A, Horwitz AR (2011) Integrins in cell migration. Cold Spring Harb Perspect Biol
3:a005074 Available at: http://www.ncbi.nlm.nih.gov/pubmed/21885598 [Accessed January
28, 2018].
Hyzinski-García MC, Rudkouskaya A, Mongin AA (2014) LRRC8A protein is indispensable for
swelling-activated and ATP-induced release of excitatory amino acids in rat astrocytes. J
Physiol 592:4855–4862 Available at: http://www.ncbi.nlm.nih.gov/pubmed/25172945
[Accessed January 9, 2018].
Hyzinski-García MC, Vincent MY, Haskew-Layton RE, Dohare P, Keller RW, Mongin AA,
Mongin AA (2011) Hypo-osmotic swelling modifies glutamate-glutamine cycle in the
cerebral cortex and in astrocyte cultures. J Neurochem 118:140–152 Available at:

135
http://www.ncbi.nlm.nih.gov/pubmed/21517854 [Accessed December 1, 2017].
Jamur MC, Oliver C (2010) Permeabilization of Cell Membranes. In: Methods in molecular
biology (Clifton, N.J.), pp 63–66 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20012820 [Accessed December 9, 2017].
Johnson KM, Milner R, Crocker SJ (2015) Extracellular matrix composition determines
astrocyte responses to mechanical and inflammatory stimuli. Neurosci Lett 600:104–109
Available at: http://www.ncbi.nlm.nih.gov/pubmed/26067407 [Accessed January 7, 2018].
Kacem K, Lacombe P, Seylaz J, Bonvento G (1998) Structural organization of the perivascular
astrocyte endfeet and their relationship with the endothelial glucose transporter: a confocal
microscopy study. Glia 23:1–10 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/9562180 [Accessed January 31, 2018].
Kaech S, Banker G (2006) Culturing hippocampal neurons. Nat Protoc 1:2406–2415 Available
at: http://www.nature.com/doifinder/10.1038/nprot.2006.356 [Accessed January 31, 2018].
Khakh BS, McCarthy KD (2015) Astrocyte calcium signaling: from observations to functions
and the challenges therein. Cold Spring Harb Perspect Biol 7:a020404 Available at:
http://cshperspectives.cshlp.org/lookup/doi/10.1101/cshperspect.a020404 [Accessed
January 31, 2018].
Khakh BS, Sofroniew M V (2015) Diversity of astrocyte functions and phenotypes in neural
circuits. Nat Neurosci 18:942–952 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26108722 [Accessed January 3, 2018].
Kim KJ, Iddings JA, Stern JE, Blanco VM, Croom D, Kirov SA, Filosa JA (2015) Astrocyte
contributions to flow/pressure-evoked parenchymal arteriole vasoconstriction. J Neurosci
35:8245–8257 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26019339 [Accessed

136
December 17, 2017].
Kimelberg HK, Anderson E, Kettenmann H (1990) Swelling-induced changes in
electrophysiological properties of cultured astrocytes and oligodendrocytes. II. Whole-cell
currents. Brain Res 529:262–268 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/2282496 [Accessed January 8, 2018].
Kimelberg HK, Macvicar BA, Sontheimer H (2006) Anion channels in astrocytes: biophysics,
pharmacology, and function. Glia 54:747–757 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/17006903 [Accessed January 9, 2018].
Kimelberg HK, O’Connor E (1988) Swelling of astrocytes causes membrane potential
depolarization. Glia 1:219–224 Available at: http://doi.wiley.com/10.1002/glia.440010307
[Accessed January 8, 2018].
Kimelberg HK, O’Connor ER, Kettenmann H (1993) Effects of Swelling on Glial Cell Function.
In, pp 157–186. Springer, Berlin, Heidelberg. Available at:
http://www.springerlink.com/index/10.1007/978-3-642-77124-8_6 [Accessed November
29, 2017].
Kirischuk S (2009) Mechanosensitive Channels in Neuronal and Astroglial Cells in the Nervous
System. In: Mechanosensitivity of the Nervous System, pp 3–22. Dordrecht: Springer
Netherlands. Available at: http://link.springer.com/10.1007/978-1-4020-8716-5_1
[Accessed January 5, 2018].
Kleinman HK, Luckenbill-Edds L, Cannon FW, Sephel GC (1987) Use of extracellular matrix
components for cell culture. Anal Biochem 166:1–13 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/3314585 [Accessed January 31, 2018].
Kobayashi T, Sokabe M (2010) Sensing substrate rigidity by mechanosensitive ion channels with

137
stress fibers and focal adhesions. Curr Opin Cell Biol 22:669–676 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20850289 [Accessed January 31, 2018].
Koser DE, Thompson AJ, Foster SK, Dwivedy A, Pillai EK, Sheridan GK, Svoboda H, Viana M,
Costa L da F, Guck J, Holt CE, Franze K (2016) Mechanosensing is critical for axon growth
in the developing brain. Nat Neurosci 19:1592–1598 Available at:
http://www.nature.com/articles/nn.4394 [Accessed January 31, 2018].
Kuchibhotla K V, Lattarulo CR, Hyman BT, Bacskai BJ (2009) Synchronous hyperactivity and
intercellular calcium waves in astrocytes in Alzheimer mice. Science 323:1211–1215
Available at: http://www.sciencemag.org/cgi/doi/10.1126/science.1169096 [Accessed
January 31, 2018].
Kulkarni JA, Myhre JL, Chen S, Tam YYC, Danescu A, Richman JM, Cullis PR (2017) Design
of lipid nanoparticles for in vitro and in vivo delivery of plasmid DNA. Nanomedicine
Nanotechnology, Biol Med 13:1377–1387 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/28038954 [Accessed January 31, 2018].
Lang F, Busch GL, Ritter M, Lkl H V, Waldegger S, Gulbins E, Ussinger DH (1998) Functional
Significance of Cell Volume Regulatory Mechanisms. Physiol Rev 78:247–306 Available
at: http://www.ncbi.nlm.nih.gov/pubmed/9457175 [Accessed January 31, 2018].
Lang F, Ritter M, Volkl H, Haussinger D (1993) The biological significance of cell volume. Ren
Physiol Biochem 16:48–65 Available at: http://www.ncbi.nlm.nih.gov/pubmed/7684146
[Accessed January 31, 2018].
Lascola CD, Kraig RP (1996) Whole-cell chloride currents in rat astrocytes accompany changes
in cell morphology. J Neurosci 16:2532–2545 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/8786429 [Accessed January 31, 2018].

138
Lau LW, Cua R, Keough MB, Haylock-Jacobs S, Yong VW (2013) Pathophysiology of the brain
extracellular matrix: a new target for remyelination. Nat Rev Neurosci 14:722–729
Available at: http://www.nature.com/doifinder/10.1038/nrn3550 [Accessed January 7,
2018].
Lawrence KM, Jones RC, Jackson TR, Baylie RL, Abbott B, Bruhn-Olszewska B, Board TN,
Locke IC, Richardson SM, Townsend PA (2017) Chondroprotection by urocortin involves
blockade of the mechanosensitive ion channel Piezo1. Sci Rep 7:5147 Available at:
http://www.nature.com/articles/s41598-017-04367-4 [Accessed December 20, 2017].
Lee W, Leddy HA, Chen Y, Lee SH, Zelenski NA, McNulty AL, Wu J, Beicker KN, Coles J,
Zauscher S, Grandl J, Sachs F, Guilak F, Liedtke WB (2014) Synergy between Piezo1 and
Piezo2 channels confers high-strain mechanosensitivity to articular cartilage. Proc Natl
Acad Sci U S A 111:E5114-22 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25385580 [Accessed December 9, 2017].
Leung AKK, Hafez IM, Baoukina S, Belliveau NM, Zhigaltsev I V., Afshinmanesh E, Tieleman
DP, Hansen CL, Hope MJ, Cullis PR (2012) Lipid Nanoparticles Containing siRNA
Synthesized by Microfluidic Mixing Exhibit an Electron-Dense Nanostructured Core. J
Phys Chem C 116:18440–18450 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/22962627 [Accessed January 31, 2018].
Levy AF, Zayats M, Guerrero-Cazares H, Quiñones-Hinojosa A, Searson PC (2014) Influence of
Basement Membrane Proteins and Endothelial Cell-Derived Factors on the Morphology of
Human Fetal-Derived Astrocytes in 2D Deli MA, ed. PLoS One 9:e92165 Available at:
http://dx.plos.org/10.1371/journal.pone.0092165 [Accessed January 18, 2018].
Lewis AH, Cui AF, McDonald MF, Grandl J (2017) Transduction of Repetitive Mechanical

139
Stimuli by Piezo1 and Piezo2 Ion Channels. Cell Rep 19:2572–2585 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/28636944 [Accessed January 31, 2018].
Li J et al. (2014) Piezo1 integration of vascular architecture with physiological force. Nature
515:279–282 Available at:
http://dx.doi.org/10.1038/nature13701%5Cnpapers2://publication/doi/10.1038/nature13701.
Liang J, Huang B, Yuan G, Chen Y, Liang F, Zeng H, Zheng S, Cao L, Geng D, Zhou S (2017)
Stretch-activated channel Piezo1 is up-regulated in failure heart and cardiomyocyte
stimulated by AngII. Am J Transl Res 9:2945–2955 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/28670382 [Accessed February 9, 2018].
Liedtke W, Choe Y, Martí-Renom MA, Bell AM, Denis CS, Sali A, Hudspeth AJ, Friedman JM,
Heller S (2000) Vanilloid receptor-related osmotically activated channel (VR-OAC), a
candidate vertebrate osmoreceptor. Cell 103:525–535 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/11081638 [Accessed January 8, 2018].
Liedtke W, Kim C (2005) Functionality of the TRPV subfamily of TRP ion channels: add
mechano-TRP and osmo-TRP to the lexicon! Cell Mol Life Sci 62:2985–3001 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/16314934 [Accessed January 31, 2018].
Liu X, Bandyopadhyay B, Nakamoto T, Singh B, Liedtke W, Melvin JE, Ambudkar I,
Ambudkar I (2006) A Role for AQP5 in Activation of TRPV4 by Hypotonicity. J Biol
Chem 281:15485–15495 Available at: http://www.ncbi.nlm.nih.gov/pubmed/16571723
[Accessed December 1, 2017].
Lloyd AC (2013) The regulation of cell size. Cell 154:1194–1205 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/24034244 [Accessed January 31, 2018].
Lukacs V, Mathur J, Mao R, Bayrak-Toydemir P, Procter M, Cahalan SM, Kim HJ, Bandell M,

140
Longo N, Day RW, Stevenson DA, Patapoutian A, Krock BL (2015) Impaired PIEZO1
function in patients with a novel autosomal recessive congenital lymphatic dysplasia. Nat
Commun 6:8329 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26387913 [Accessed
January 31, 2018].
Mabie PC, Mehler MF, Marmur R, Papavasiliou A, Song Q, Kessler JA (1997) Bone
morphogenetic proteins induce astroglial differentiation of oligodendroglial-astroglial
progenitor cells. J Neurosci 17:4112–4120 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/9151728 [Accessed January 27, 2018].
Mailloux RJ, Harper M-E (2012) Mitochondrial proticity and ROS signaling: lessons from the
uncoupling proteins. Trends Endocrinol Metab 23:451–458 Available at:
http://www.sciencedirect.com/science/article/pii/S1043276012000653 [Accessed January 4,
2018].
Maity B, Sheff D, Fisher RA (2013) Immunostaining. In: Methods in cell biology, pp 81–105
Available at: http://www.ncbi.nlm.nih.gov/pubmed/23317899 [Accessed January 31, 2018].
Malécot CO, Bito V, Argibay JA (1998) Ruthenium red as an effective blocker of calcium and
sodium currents in guinea-pig isolated ventricular heart cells. Br J Pharmacol 124:465–472
Available at: http://www.ncbi.nlm.nih.gov/pubmed/9647469 [Accessed December 25,
2017].
McCormack JG, Halestrap AP, Denton RM (1990) Role of calcium ions in regulation of
mammalian intramitochondrial metabolism. Physiol Rev 70:391–425 Available at:
http://www.physiology.org/doi/10.1152/physrev.1990.70.2.391 [Accessed January 4, 2018].
McHugh BJ, Buttery R, Lad Y, Banks S, Haslett C, Sethi T (2010) Integrin activation by
Fam38A uses a novel mechanism of R-Ras targeting to the endoplasmic reticulum. J Cell

141
Sci 123:51–61 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20016066%5Cnhttp://jcs.biologists.org/content/123/1
/51.full.pdf.
McHugh BJ, Murdoch A, Haslett C, Sethi T (2012) Loss of the integrin-activating
transmembrane protein Fam38A (Piezo1) promotes a switch to a reduced integrin-
dependent mode of cell migration. Kyprianou N, ed. PLoS One 7:e40346 Available at:
http://dx.plos.org/10.1371/journal.pone.0040346 [Accessed February 1, 2018].
Milner R, Huang X, Wu J, Nishimura S, Pytela R, Sheppard D, ffrench-Constant C (1999)
Distinct roles for astrocyte alphavbeta5 and alphavbeta8 integrins in adhesion and
migration. J Cell Sci 112 ( Pt 23):4271–4279 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/10564645 [Accessed January 31, 2018].
Miyamoto T, Mochizuki T, Nakagomi H, Kira S, Watanabe M, Takayama Y, Suzuki Y, Koizumi
S, Takeda M, Tominaga M (2014) Functional role for Piezo1 in stretch-evoked Ca2+ influx
and ATP release in urothelial cell cultures. J Biol Chem 289:16565–16575 Available at:
http://www.jbc.org/lookup/doi/10.1074/jbc.M113.528638 [Accessed January 31, 2018].
Mola MG, Sparaneo A, Gargano CD, Spray DC, Svelto M, Frigeri A, Scemes E, Nicchia GP
(2016) The speed of swelling kinetics modulates cell volume regulation and calcium
signaling in astrocytes: A different point of view on the role of aquaporins. Glia 64:139–154
Available at: http://www.ncbi.nlm.nih.gov/pubmed/26413835 [Accessed January 31, 2018].
Morales-Mulia S, Vaca L, Hernandez-Cruz A, Pasantes-Morales H (1998) Osmotic swelling-
induced changes in cytosolic calcium do not affect regulatory volume decrease in rat
cultured suspended cerebellar astrocytes. J Neurochem 71:2330–2338 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/9832131 [Accessed January 31, 2018].

142
Moran MM, Xu H, Clapham DE (2004) TRP ion channels in the nervous system. Curr Opin
Neurobiol 14:362–369 Available at:
http://www.sciencedirect.com.ezproxy.library.ubc.ca/science/article/pii/S095943880400070
4?via%3Dihub [Accessed January 8, 2018].
Mulligan SJ, MacVicar BA (2004) Calcium transients in astrocyte endfeet cause cerebrovascular
constrictions. Nature 431:195–199 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/15356633 [Accessed January 31, 2018].
Murphy MC, Huston J, Jack CR, Glaser KJ, Manduca A, Felmlee JP, Ehman RL, Felmlee JP,
Ehman RL (2011) Decreased brain stiffness in Alzheimer’s disease determined by magnetic
resonance elastography. J Magn Reson Imaging 34:494–498 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/21751286 [Accessed January 28, 2018].
Murphy MC, Jones DT, Jack CR, Glaser KJ, Senjem ML, Manduca A, Felmlee JP, Carter RE,
Ehman RL, Huston J, Huston J, III (2016) Regional brain stiffness changes across the
Alzheimer’s disease spectrum. NeuroImage Clin 10:283–290 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26900568 [Accessed January 7, 2018].
Nagelhus EA, Ottersen OP (2013) Physiological roles of aquaporin-4 in brain. Physiol Rev
93:1543–1562 Available at: http://www.ncbi.nlm.nih.gov/pubmed/24137016 [Accessed
January 7, 2018].
Navarrete M, Araque A (2014) The Cajal school and the physiological role of astrocytes: a way
of thinking. Front Neuroanat 8:33 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/24904302 [Accessed January 31, 2018].
Nayerossadat N, Maedeh T, Ali PA (2012) Viral and nonviral delivery systems for gene
delivery. Adv Biomed Res 1:27 Available at:

143
http://www.ncbi.nlm.nih.gov/pubmed/23210086 [Accessed November 12, 2017].
Nett WJ, Oloff SH, McCarthy KD (2002) Hippocampal Astrocytes In Situ Exhibit Calcium
Oscillations That Occur Independent of Neuronal Activity. J Neurophysiol 87:528–537
Available at: http://www.ncbi.nlm.nih.gov/pubmed/11784768 [Accessed January 2, 2018].
Nielsen S, Nagelhus EA, Amiry-Moghaddam M, Bourque C, Agre P, Ottersen OP (1997)
Specialized membrane domains for water transport in glial cells: high-resolution
immunogold cytochemistry of aquaporin-4 in rat brain. J Neurosci 17:171–180 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/8987746 [Accessed January 7, 2018].
Niggel J, Hu H, Sigurdson W, Bowman C, Sachs F (1996) Grammostola spatulata venom blocks
mechanical transduction in GH3 neurons, Xenopus oocytes and chick heart cells. Biophys J
70:A347.
Nimmerjahn A, Mukamel EA, Schnitzer MJ (2009) Motor Behavior Activates Bergmann Glial
Networks. Neuron 62:400–412 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/19447095 [Accessed January 31, 2018].
Nizar K et al. (2013) In vivo Stimulus-Induced Vasodilation Occurs without IP3 Receptor
Activation and May Precede Astrocytic Calcium Increase. J Neurosci 33:8411–8422
Available at: http://www.ncbi.nlm.nih.gov/pubmed/23658179 [Accessed January 31, 2018].
Noël J, Sandoz G, Lesage F (2011) Molecular regulations governing TREK and TRAAK
channel functions. Channels (Austin) 5:402–409 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/21829087 [Accessed December 24, 2017].
Nonomura K, Woo S-H, Chang RB, Gillich A, Qiu Z, Francisco AG, Ranade SS, Liberles SD,
Patapoutian A (2016) Piezo2 senses airway stretch and mediates lung inflation-induced
apnoea. Nature 541:176–181 Available at:

144
http://www.nature.com/doifinder/10.1038/nature20793 [Accessed December 28, 2017].
Nourse JL, Pathak MM (2017) How cells channel their stress: Interplay between Piezo1 and the
cytoskeleton. Semin Cell Dev Biol 71:3–12 Available at:
http://www.sciencedirect.com.ezproxy.library.ubc.ca/science/article/pii/S108495211630300
7?via%3Dihub#bib0265 [Accessed November 19, 2017].
O’Connor ER, Kimelberg HK (1993) Role of calcium in astrocyte volume regulation and in the
release of ions and amino acids. J Neurosci 13:2638–2650 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/8501529 [Accessed January 31, 2018].
O’Connor ER, Kimelberg HK, Keese CR, Giaever I (1993) Electrical resistance method for
measuring volume changes in monolayer cultures applied to primary astrocyte cultures. Am
J Physiol Physiol 264:C471–C478 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/8447377 [Accessed January 31, 2018].
Oliet SHR, Bourque C (1993) Mechanosensitive channels transduce osmosensitivity in
supraoptic neurons. Nature 364:341–343 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/7687327 [Accessed January 7, 2018].
Olude MA, Mustapha OA, Aderounmu OA, Olopade JO, Ihunwo AO (2015) Astrocyte
morphology, heterogeneity, and density in the developing African giant rat (Cricetomys
gambianus). Front Neuroanat 9:67 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26074782 [Accessed December 9, 2017].
Ostrow LW, Langan TJ, Sachs F (2000) Stretch-induced endothelin-1 production by astrocytes. J
Cardiovasc Pharmacol 36:S274-7 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/11078397 [Accessed January 6, 2018].
Otsu Y, Couchman K, Lyons DG, Collot M, Agarwal A, Mallet J-M, Pfrieger FW, Bergles DE,

145
Charpak S (2015) Calcium dynamics in astrocyte processes during neurovascular coupling.
Nat Neurosci 18:210–218 Available at: http://www.nature.com/articles/nn.3906 [Accessed
January 31, 2018].
Panatier A, Vallée J, Haber M, Murai KK, Lacaille J-C, Robitaille R (2011) Astrocytes Are
Endogenous Regulators of Basal Transmission at Central Synapses. Cell 146:785–798
Available at: http://linkinghub.elsevier.com/retrieve/pii/S0092867411008208 [Accessed
January 31, 2018].
Parpura V, Basarsky TA, Liu F, Jeftinija K, Jeftinija S, Haydon PG (1994) Glutamate-mediated
astrocyte–neuron signalling. Nature 369:744–747 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/7911978 [Accessed January 31, 2018].
Parpura V, Verkhratsky A (2012) Homeostatic function of astrocytes: Ca(2+) and Na(+)
signalling. Transl Neurosci 3:334–344 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/23243501 [Accessed January 6, 2018].
Pasantes-Morales H, Murray RA, Lilja L, Moran J (1994) Regulatory volume decrease in
cultured astrocytes. I. Potassium- and chloride-activated permeability. Am J Physiol Physiol
266:C165–C171 Available at: http://www.ncbi.nlm.nih.gov/pubmed/8304413 [Accessed
January 31, 2018].
Pasti L, Volterra A, Pozzan T, Carmignoto G (1997) Intracellular calcium oscillations in
astrocytes: a highly plastic, bidirectional form of communication between neurons and
astrocytes in situ. J Neurosci 17:7817–7830 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/9315902 [Accessed January 31, 2018].
Patel AJ, Honoré E, Maingret F, Lesage F, Fink M, Duprat F, Lazdunski M (1998) A
mammalian two pore domain mechano-gated S-like K+ channel. EMBO J 17:4283–4290

146
Available at: http://www.ncbi.nlm.nih.gov/pubmed/9687497 [Accessed January 31, 2018].
Pathak MM, Nourse JL, Tran T, Hwe J, Arulmoli J, Le DTT, Bernardis E, Flanagan LA,
Tombola F (2014) Stretch-activated ion channel Piezo1 directs lineage choice in human
neural stem cells. Proc Natl Acad Sci U S A 111:16148–16153 Available at:
http://www.pnas.org/lookup/doi/10.1073/pnas.1409802111 [Accessed January 31, 2018].
Pedersen SF, Klausen TK, Nilius B (2015) The identification of a volume-regulated anion
channel: an amazing Odyssey. Acta Physiol 213:868–881 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25565132 [Accessed January 31, 2018].
Perea G, Navarrete M, Araque A (2009) Tripartite synapses: astrocytes process and control
synaptic information. Trends Neurosci 32:421–431 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/19615761 [Accessed January 31, 2018].
Perez V, Bouschet T, Fernandez C, Bockaert J, Journot L (2005) Dynamic reorganization of the
astrocyte actin cytoskeleton elicited by cAMP and PACAP: a role for phosphatidylInositol
3-kinase inhibition. Eur J Neurosci 21:26–32 Available at:
http://doi.wiley.com/10.1111/j.1460-9568.2004.03845.x [Accessed December 2, 2017].
Petravicz J, Fiacco TA, McCarthy KD (2008) Loss of IP3 Receptor-Dependent Ca2+ Increases
in Hippocampal Astrocytes Does Not Affect Baseline CA1 Pyramidal Neuron Synaptic
Activity. J Neurosci 28:4967–4973 Available at:
http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.5572-07.2008 [Accessed January
31, 2018].
Picht E, Zima A V., Blatter LA, Bers DM (2007) SparkMaster: automated calcium spark analysis
with ImageJ. AJP Cell Physiol 293:C1073–C1081 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/17376815 [Accessed November 13, 2017].

147
Plant TD (2014) TRPs in mechanosensing and volume regulation. Handb Exp Pharmacol
223:743–766 Available at: http://link.springer.com/10.1007/978-3-319-05161-1_2
[Accessed January 31, 2018].
Poole K, Herget R, Lapatsina L, Ngo H-D, Lewin GR (2014) Tuning Piezo ion channels to
detect molecular-scale movements relevant for fine touch. Nat Commun 5:3520 Available
at: http://www.nature.com/doifinder/10.1038/ncomms4520 [Accessed December 23, 2017].
Porter JT, McCarthy KD (1995) GFAP-positive hippocampal astrocytes in situ respond to
glutamatergic neuroligands with increases in [Ca2+]i. Glia 13:101–112 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/7544323 [Accessed January 31, 2018].
Porter JT, McCarthy KD (1996) Hippocampal astrocytes in situ respond to glutamate released
from synaptic terminals. J Neurosci 16:5073–5081 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/8756437 [Accessed January 31, 2018].
Qi Y, Andolfi L, Frattini F, Mayer F, Lazzarino M, Hu J (2015) Membrane stiffening by
STOML3 facilitates mechanosensation in sensory neurons. Nat Commun 6:8512 Available
at: http://www.nature.com/doifinder/10.1038/ncomms9512 [Accessed December 23, 2017].
Qiu Z, Dubin AE, Mathur J, Tu B, Reddy K, Miraglia LJ, Reinhardt J, Orth AP, Patapoutian A
(2014) SWELL1, a plasma membrane protein, is an essential component of volume-
regulated anion channel. Cell 157:447–458 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/24725410 [Accessed January 9, 2018].
Ranade SS, Qiu Z, Woo S-H, Hur SS, Murthy SE, Cahalan SM, Xu J, Mathur J, Bandell M,
Coste B, Li Y-SJ, Chien S, Patapoutian A (2014) Piezo1, a mechanically activated ion
channel, is required for vascular development in mice. Proc Natl Acad Sci U S A
111:10347–10352 Available at: http://www.pnas.org/cgi/doi/10.1073/pnas.1409233111

148
[Accessed January 31, 2018].
Ranade SS, Syeda R, Patapoutian A (2015) Mechanically Activated Ion Channels. Neuron
87:1162–1179 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26402601 [Accessed
December 24, 2017].
Reeves AMB, Shigetomi E, Khakh BS (2011) Bulk loading of calcium indicator dyes to study
astrocyte physiology: key limitations and improvements using morphological maps. J
Neurosci 31:9353–9358 Available at:
http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.0127-11.2011 [Accessed January
31, 2018].
Retailleau K, Duprat F, Arhatte M, Ranade SS, Peyronnet R, Martins JR, Jodar M, Moro C,
Offermanns S, Feng Y, Demolombe S, Patel A, Honoré E (2015) Piezo1 in Smooth Muscle
Cells Is Involved in Hypertension-Dependent Arterial Remodeling. Cell Rep 13:1161–1171
Available at:
http://www.sciencedirect.com.ezproxy.library.ubc.ca/science/article/pii/S221112471501114
6?via%3Dihub [Accessed January 20, 2018].
Rode B et al. (2017) Piezo1 channels sense whole body physical activity to reset cardiovascular
homeostasis and enhance performance. Nat Commun 8:350 Available at:
http://www.nature.com/articles/s41467-017-00429-3 [Accessed February 9, 2018].
Rodnight RB, Gottfried C (2013) Morphological plasticity of rodent astroglia. J Neurochem
124:263–275 Available at: http://doi.wiley.com/10.1111/jnc.12087 [Accessed December 2,
2017].
Rosenegger DG, Tran CHT, Wamsteeker Cusulin JI, Gordon GR (2015) Tonic Local Brain
Blood Flow Control by Astrocytes Independent of Phasic Neurovascular Coupling. J

149
Neurosci 35:13463–13474 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26424891
[Accessed January 31, 2018].
Rungta RL, Bernier L-P, Dissing-Olesen L, Groten CJ, LeDue JM, Ko R, Drissler S, MacVicar
BA (2016) Ca 2+ transients in astrocyte fine processes occur via Ca 2+ influx in the adult
mouse hippocampus. Glia 64:2093–2103 Available at:
http://doi.wiley.com/10.1002/glia.23042.
Rungta RL, Choi HB, Lin PJ, Ko RW, Ashby D, Nair J, Manoharan M, Cullis PR, Macvicar BA
(2013) Lipid Nanoparticle Delivery of siRNA to Silence Neuronal Gene Expression in the
Brain. Mol Ther Nucleic Acids 2:e136 Available at:
http://dx.doi.org/10.1038/mtna.2013.65.
Rungta RL, Choi HB, Tyson JR, Malik A, Dissing-Olesen L, Lin PJC, Cain SM, Cullis PR,
Snutch TP, MacVicar BA (2015) The cellular mechanisms of neuronal swelling underlying
cytotoxic edema. Cell 161:610–621 Available at:
http://linkinghub.elsevier.com/retrieve/pii/S0092867415003165 [Accessed January 31,
2018].
Ryoo K, Park J-Y (2016) Two-pore Domain Potassium Channels in Astrocytes. Exp Neurobiol
25:222–232 Available at:
https://synapse.koreamed.org/DOIx.php?id=10.5607/en.2016.25.5.222 [Accessed January
31, 2018].
Saotome K, Murthy SE, Kefauver JM, Whitwam T, Patapoutian A, Ward AB (2017) Structure of
the mechanically activated ion channel Piezo1. Nature Available at:
http://www.ncbi.nlm.nih.gov/pubmed/29261642 [Accessed February 9, 2018].
Satoh K, Hata M, Takahara S, Tsuzaki H, Yokota H, Akatsu H, Yamamoto T, Kosaka K,

150
Yamada T (2006) A novel membrane protein, encoded by the gene covering KIAA0233, is
transcriptionally induced in senile plaque-associated astrocytes. Brain Res 1108:19–27.
Schnell U, Dijk F, Sjollema KA, Giepmans BNG (2012) Immunolabeling artifacts and the need
for live-cell imaging. Nat Methods 9:152–158 Available at:
http://www.nature.com/doifinder/10.1038/nmeth.1855 [Accessed November 17, 2017].
Schober AL, Mongin AA (2015) Intracellular levels of glutamate in swollen astrocytes are
preserved via neurotransmitter reuptake and de novo synthesis: implications for
hyponatremia. J Neurochem 135:176–185 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26235094 [Accessed December 1, 2017].
Schober AL, Wilson CS, Mongin AA (2017) Molecular composition and heterogeneity of the
LRRC8-containing swelling-activated osmolyte channels in primary rat astrocytes. J
Physiol 595:6939–6951 Available at: http://doi.wiley.com/10.1113/JP275053 [Accessed
January 9, 2018].
Schwartz MA (2010) Integrins and extracellular matrix in mechanotransduction. Cold Spring
Harb Perspect Biol 2:a005066 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/21084386 [Accessed January 7, 2018].
Servin-Vences MR, Moroni M, Lewin GR, Poole K (2017) Direct measurement of TRPV4 and
PIEZO1 activity reveals multiple mechanotransduction pathways in chondrocytes. Elife 6
Available at: https://elifesciences.org/articles/21074 [Accessed January 31, 2018].
Sharma K, Schmitt S, Bergner CG, Tyanova S, Kannaiyan N, Manrique-Hoyos N, Kongi K,
Cantuti L, Hanisch U-K, Philips M-A, Rossner MJ, Mann M, Simons M (2015) Cell type -
and brain region - resolved mouse brain proteome. Nat Neurosci 18:1–16 Available at:
http://dx.doi.org/10.1038/nn.4160%5Cnpapers2://publication/doi/10.1038/nn.4160.

151
Shigetomi E, Bushong EA, Haustein MD, Tong X, Jackson-Weaver O, Kracun S, Xu J,
Sofroniew M V, Ellisman MH, Khakh BS (2013a) Imaging calcium microdomains within
entire astrocyte territories and endfeet with GCaMPs expressed using adeno-associated
viruses. J Gen Physiol 141:633–647 Available at:
http://www.jgp.org/lookup/doi/10.1085/jgp.201210949 [Accessed January 31, 2018].
Shigetomi E, Jackson-Weaver O, Huckstepp RT, O’Dell TJ, Khakh BS (2013b) TRPA1 channels
are regulators of astrocyte basal calcium levels and long-term potentiation via constitutive
D-serine release. J Neurosci 33:10143–10153 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/23761909 [Accessed January 31, 2018].
Shigetomi E, Kracun S, Sofroniew M V, Khakh BS (2010) A genetically targeted optical sensor
to monitor calcium signals in astrocyte processes. Nat Neurosci 13:759–766 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20495558 [Accessed January 31, 2018].
Shigetomi E, Patel S, Khakh BS (2016) Probing the Complexities of Astrocyte Calcium
Signaling. Trends Cell Biol 26.
Shigetomi E, Tong X, Kwan KY, Corey DP, Khakh BS (2012) TRPA1 channels regulate
astrocyte resting calcium and inhibitory synapse efficacy through GAT-3. Nat Neurosci
15:70–80 Available at: http://www.ncbi.nlm.nih.gov/pubmed/22158513 [Accessed January
31, 2018].
Simard M, Nedergaard M (2004) The neurobiology of glia in the context of water and ion
homeostasis. Neuroscience 129:877–896 Available at:
http://linkinghub.elsevier.com/retrieve/pii/S0306452204008462 [Accessed January 31,
2018].
Sofroniew M V, Vinters H V (2010) Astrocytes: biology and pathology. Acta Neuropathol

152
119:7–35 Available at: http://www.ncbi.nlm.nih.gov/pubmed/20012068 [Accessed
December 9, 2017].
Srinivasan R, Huang BS, Venugopal S, Johnston AD, Chai H, Zeng H, Golshani P, Khakh BS
(2015) Ca2+ signaling in astrocytes from Ip3r2−/− mice in brain slices and during startle
responses in vivo. Nat Neurosci 18:708–717 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25894291 [Accessed November 13, 2017].
Srinivasan R, Lu TY, Chai H, Xu J, Huang BS, Golshani P, Coppola G, Khakh BS (2016) New
Transgenic Mouse Lines for Selectively Targeting Astrocytes and Studying Calcium
Signals in Astrocyte Processes In Situ and In Vivo. Neuron 92:1181–1195.
Stadler C, Rexhepaj E, Singan VR, Murphy RF, Pepperkok R, Uhlén M, Simpson JC, Lundberg
E (2013) Immunofluorescence and fluorescent-protein tagging show high correlation for
protein localization in mammalian cells. Nat Methods 10:315–323 Available at:
http://www.nature.com/doifinder/10.1038/nmeth.2377 [Accessed November 15, 2017].
Strotmann R, Harteneck C, Nunnenmacher K, Schultz G, Plant TD (2000) OTRPC4, a
nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat Cell Biol
2:695–702 Available at: http://www.nature.com/doifinder/10.1038/35036318 [Accessed
January 8, 2018].
Suchyna TM, Johnson JH, Hamer K, Leykam JF, Gage DA, Clemo HF, Baumgarten CM, Sachs
F (2000) Identification of a peptide toxin from Grammostola spatulata spider venom that
blocks cation-selective stretch-activated channels. J Gen Physiol 115:583–598 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/10779316 [Accessed January 8, 2018].
Sugimoto A, Miyazaki A, Kawarabayashi K, Shono M, Akazawa Y, Hasegawa T, Ueda-
Yamaguchi K, Kitamura T, Yoshizaki K, Fukumoto S, Iwamoto T (2017) Piezo type

153
mechanosensitive ion channel component 1 functions as a regulator of the cell fate
determination of mesenchymal stem cells. Sci Rep 7:17696 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/29255201 [Accessed January 27, 2018].
Summers L, Kangwantas K, Nguyen L, Kielty C, Pinteaux E (2010) Adhesion to the
extracellular matrix is required for interleukin-1 beta actions leading to reactive phenotype
in rat astrocytes. Mol Cell Neurosci 44:272–281 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/20380881 [Accessed December 9, 2017].
Sun W, McConnell E, Pare J-F, Xu Q, Chen M, Peng W, Lovatt D, Han X, Smith Y, Nedergaard
M (2013) Glutamate-Dependent Neuroglial Calcium Signaling Differs Between Young and
Adult Brain. Science (80- ) 339:197–200 Available at:
http://www.sciencemag.org/cgi/doi/10.1126/science.1226740 [Accessed January 31, 2018].
Syeda R, Florendo MN, Cox CD, Kefauver JM, Santos JS, Martinac B, Patapoutian A (2016)
Piezo1 Channels Are Inherently Mechanosensitive. Cell Rep 17:1739–1746 Available at:
http://linkinghub.elsevier.com/retrieve/pii/S2211124716314383 [Accessed January 31,
2018].
Syeda R, Xu J, Dubin AE, Coste B, Mathur J, Huynh T, Matzen J, Lao J, Tully DC, Engels IH,
Petrassi HM, Schumacher AM, Montal M, Bandell M, Patapoutian A (2015) Chemical
activation of the mechanotransduction channel Piezo1. Elife 4 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/26001275 [Accessed November 13, 2017].
Tanigami H, Okamoto T, Yasue Y, Shimaoka M (2012) Astroglial integrins in the development
and regulation of neurovascular units. Pain Res Treat 2012:964652 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/23304493 [Accessed November 28, 2017].
Thrane AS, Rangroo Thrane V, Nedergaard M (2014) Drowning stars: reassessing the role of

154
astrocytes in brain edema. Trends Neurosci 37:620–628 Available at:
http://linkinghub.elsevier.com/retrieve/pii/S0166223614001532 [Accessed January 31,
2018].
Thul PJ et al. (2017) A subcellular map of the human proteome. Science 356:eaal3321 Available
at: http://www.sciencemag.org/lookup/doi/10.1126/science.aal3321 [Accessed January 31,
2018].
Tojkander S, Gateva G, Lappalainen P (2012) Actin stress fibers - assembly, dynamics and
biological roles. J Cell Sci 125:1855–1864 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/22544950 [Accessed January 31, 2018].
Tremblay RG, Sikorska M, Sandhu JK, Lanthier P, Ribecco-Lutkiewicz M, Bani-Yaghoub M
(2010) Differentiation of mouse Neuro 2A cells into dopamine neurons. J Neurosci
Methods 186:60–67 Available at:
http://linkinghub.elsevier.com/retrieve/pii/S0165027009006025 [Accessed January 31,
2018].
Tyler WJ (2012) The mechanobiology of brain function. Nat Rev Neurosci 13:867–878
Available at: http://www.nature.com/articles/nrn3383 [Accessed January 31, 2018].
Vardjan N, Kreft M, Zorec R (2014) Dynamics of β-adrenergic/cAMP signaling and
morphological changes in cultured astrocytes. Glia 62:566–579 Available at:
http://doi.wiley.com/10.1002/glia.22626 [Accessed December 2, 2017].
Vargova L, Sykova E (2014) Astrocytes and extracellular matrix in extrasynaptic volume
transmission. Philos Trans R Soc B Biol Sci 369:20130608–20130608 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25225101 [Accessed January 31, 2018].
Verkhratsky A, Nedergaard M (2014) Astroglial cradle in the life of the synapse. Philos Trans R

155
Soc Lond B Biol Sci 369:20130595 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/25225089 [Accessed January 13, 2018].
Verkhratsky A, Rodríguez JJ, Parpura V (2012) Calcium signalling in astroglia. Mol Cell
Endocrinol 353:45–56 Available at:
http://www.sciencedirect.com/science/article/pii/S0303720711005314?via%3Dihub
[Accessed January 11, 2018].
Ververken D, Veldhoven P, Proost C, Carton H, Wulf H (1982) On the Role of Calcium Ions in
the Regulation of Glycogenolysis in Mouse Brain Cortical Slices. J Neurochem 38:1286–
1295 Available at: http://doi.wiley.com/10.1111/j.1471-4159.1982.tb07903.x [Accessed
January 4, 2018].
Volterra A, Liaudet N, Savtchouk I (2014) Astrocyte Ca2+ signalling: an unexpected
complexity. Nat Rev Neurosci 15:327–335 Available at:
http://www.nature.com/doifinder/10.1038/nrn3725 [Accessed November 13, 2017].
Voss FK, Ullrich F, Munch J, Lazarow K, Lutter D, Mah N, Andrade-Navarro MA, von Kries
JP, Stauber T, Jentsch TJ (2014) Identification of LRRC8 Heteromers as an Essential
Component of the Volume-Regulated Anion Channel VRAC. Science (80- ) 344:634–638
Available at: http://www.ncbi.nlm.nih.gov/pubmed/24790029 [Accessed January 31, 2018].
Wang X, Lou N, Xu Q, Tian G-F, Peng WG, Han X, Kang J, Takano T, Nedergaard M (2006)
Astrocytic Ca2+ signaling evoked by sensory stimulation in vivo. Nat Neurosci 9:816–823
Available at: http://www.nature.com/articles/nn1703 [Accessed January 3, 2018].
Wang Y, Xiao B (2017) The mechanosensitive Piezo1 channel: structural features and molecular
bases underlying its ion permeation and mechanotransduction. J Physiol Available at:
http://doi.wiley.com/10.1113/JP274404 [Accessed January 31, 2018].

156
Wasterlain CG, Torack RM (1968) Cerebral Edema in Water Intoxication. Arch Neurol 19:79
Available at:
http://archneur.jamanetwork.com/article.aspx?doi=10.1001/archneur.1968.00480010097008
[Accessed December 11, 2017].
Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A, Moshourab R, Kozlenkov
A, Labuz D, Caspani O, Erdmann B, Machelska H, Heppenstall PA, Lewin GR (2007) A
stomatin-domain protein essential for touch sensation in the mouse. Nature 445:206–209
Available at: http://www.ncbi.nlm.nih.gov/pubmed/17167420 [Accessed December 23,
2017].
Wilson CL, Hayward SL, Kidambi S (2016) Astrogliosis in a dish: substrate stiffness induces
astrogliosis in primary rat astrocytes. RSC Adv 6:34447–34457 Available at:
http://digitalcommons.unl.edu/chemengall [Accessed December 9, 2017].
Wu BY, Liu RY, So KL, Yu ACH (2000) Multi-lipofection efficiently transfected genes into
astrocytes in primary culture. J Neurosci Methods 102:133–141 Available at:
http://www.sciencedirect.com.ezproxy.library.ubc.ca/science/article/pii/S016502700000285
5 [Accessed November 12, 2017].
Wu J, Young M, Lewis AH, Martfeld AN, Kalmeta B, Grandl J (2017) Inactivation of
Mechanically Activated Piezo1 Ion Channels Is Determined by the C-Terminal
Extracellular Domain and the Inner Pore Helix. Cell Rep 21:2357–2366 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/29186675 [Accessed January 31, 2018].
Xiang J, Tang Y, Li C, Su EJ, Lawrence DA, Keep RF (2016) Mechanisms Underlying
Astrocyte Endfeet Swelling in Stroke. In: Acta neurochirurgica. Supplement, pp 19–22
Available at: http://www.ncbi.nlm.nih.gov/pubmed/26463917 [Accessed January 20, 2018].

157
Xu ZS, Lee RJ, Chu SS, Yao A, Paun MK, Murphy SP, Mourad PD (2013) Evidence of changes
in brain tissue stiffness after ischemic stroke derived from ultrasound-based elastography. J
Ultrasound Med 32:485–494 Available at: http://www.ncbi.nlm.nih.gov/pubmed/23443189
[Accessed January 28, 2018].
Yang X-N, Lu Y-P, Liu J-J, Huang J-K, Liu Y-P, Xiao C-X, Jazag A, Ren J-L, Guleng B (2014)
Piezo1 is as a novel trefoil factor family 1 binding protein that promotes gastric cancer cell
mobility in vitro. Dig Dis Sci 59:1428–1435 Available at:
http://link.springer.com/10.1007/s10620-014-3044-3 [Accessed February 1, 2018].
Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, Rinehart J,
Gallagher PG (2012) Mutations in the mechanotransduction protein PIEZO1 are associated
with hereditary xerocytosis. Blood 120:1908–1915 Available at:
http://www.bloodjournal.org/cgi/doi/10.1182/blood-2012-04-422253 [Accessed January 31,
2018].
Zatsepin TS, Kotelevtsev Y V, Koteliansky V (2016) Lipid nanoparticles for targeted siRNA
delivery - going from bench to bedside. Int J Nanomedicine 11:3077–3086 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/27462152 [Accessed November 12, 2017].
Zhang T, Chi S, Jiang F, Zhao Q, Xiao B (2017) A protein interaction mechanism for
suppressing the mechanosensitive Piezo channels. Nat Commun 8:1797 Available at:
http://www.nature.com/articles/s41467-017-01712-z [Accessed February 9, 2018].
Zhang Y, Chen K, Sloan SA, Bennett ML, Scholze AR, O’Keeffe S, Phatnani HP, Guarnieri P,
Caneda C, Ruderisch N, Deng S, Liddelow SA, Zhang C, Daneman R, Maniatis T, Barres
BA, Wu JQ (2014) An RNA-sequencing transcriptome and splicing database of glia,
neurons, and vascular cells of the cerebral cortex. J Neurosci 34:11929–11947 Available at:

158
http://www.ncbi.nlm.nih.gov/pubmed/25186741.
Zhang Y, Gao F, Popov VL, Wen JW, Hamill OP (2000) Mechanically gated channel activity in
cytoskeleton-deficient plasma membrane blebs and vesicles from Xenopus oocytes. J
Physiol 523 Pt 1:117–130 Available at: http://www.ncbi.nlm.nih.gov/pubmed/10673548
[Accessed December 29, 2017].
Zhao Q, Wu K, Geng J, Chi S, Wang Y, Zhi P, Zhang M, Xiao B (2016) Ion Permeation and
Mechanotransduction Mechanisms of Mechanosensitive Piezo Channels. Neuron 89:1248–
1263 Available at: http://www.ncbi.nlm.nih.gov/pubmed/26924440 [Accessed January 31,
2018].
Zhao Q, Zhou H, Chi S, Wang Y, Wang J, Geng J, Wu K, Liu W, Zhang T, Dong M-Q, Wang J,
Li X, Xiao B (2018) Structure and mechanogating mechanism of the Piezo1 channel. Nat
2018 Available at: https://www.nature.com/articles/nature25743 [Accessed February 9,
2018].
Zhou M, Xu G, Xie M, Zhang X, Schools GP, Ma L, Kimelberg HK, Chen H (2009) TWIK-1
and TREK-1 are potassium channels contributing significantly to astrocyte passive
conductance in rat hippocampal slices. J Neurosci 29:8551–8564 Available at:
http://www.ncbi.nlm.nih.gov/pubmed/19571146 [Accessed January 8, 2018].




















