
Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Journal of Plant Physiology 168 (2011) 807–815
Contents lists available at ScienceDirect
Journal of Plant Physiology
journa l homepage: www.e lsev ier .de / jp lph
Salicylic acid alleviates decreases in photosynthesis under salt stress byenhancing nitrogen and sulfur assimilation and antioxidant metabolismdifferentially in two mungbean cultivars
Rahat Nazar, Noushina Iqbal, Shabina Syeed, Nafees A. Khan ∗
Department of Botany, Aligarh Muslim University, Aligarh 202 002, India
a r t i c l e i n f o
Article history:Received 23 September 2010Received in revised form 5 November 2010Accepted 5 November 2010
Keywords:Oxidative stressPhotosynthesisSalicylic acidSaltVigna radiata
a b s t r a c t
Salicylic acid (SA) is known to affect photosynthesis under normal conditions and induces tolerance inplants to biotic and abiotic stresses through influencing physiological processes. In this study, physiologi-cal processes were compared in salt-tolerant (Pusa Vishal) and salt-sensitive (T44) cultivars of mungbeanand examined how much these processes were induced by SA treatment to alleviate decrease in pho-tosynthesis under salt stress. Cultivar T44 accumulated higher leaf Na+ and Cl− content and exhibitedgreater oxidative stress than Pusa Vishal. Activity of antioxidant enzymes, ascorbate peroxidase (APX)and glutathione reductase (GR) was greater in Pusa Vishal than T44. Contrarily, activity of superox-ide dismutase (SOD) was greater in T44. The greater accumulation of leaf nitrogen and sulfur throughhigher activity of their assimilating enzymes, nitrate reductase (NR) and ATP-sulfurylase (ATPS) increasedreduced glutathione (GSH) content more conspicuously in Pusa Vishal than T44. Application of 0.5 mMSA increased nitrogen and sulfur assimilation, GSH content and activity of APX and GR. This resulted inthe increase in photosynthesis under non-saline condition and alleviated the decrease in photosynthesisunder salt stress. It also helped in restricting Na+ and Cl− content in leaf, and maintaining higher efficiencyof PSII, photosynthetic N-use efficiency (NUE) and water relations in Pusa Vishal. However, application of1.0 mM SA resulted in inhibitory effects. The effect of SA was more pronounced in Pusa Vishal than T44.These results indicate that SA application alleviates the salt-induced decrease in photosynthesis mainlythrough inducing the activity of NR and ATPS, and increasing antioxidant metabolism to a greater extentin Pusa Vishal than T44.
© 2010 Elsevier GmbH. All rights reserved.
Introduction
Salt stress is a major abiotic stress that causes detrimentaleffects on plant growth and productivity mainly through changesat physiological, biochemical and molecular level (Tester andDevenport, 2003; Khan et al., 2009; Syeed et al., 2010). The phys-iological processes that are primarily adversely affected by saltstress include ion toxicity, osmotic stress, nutrient deficiency andoxidative stress (Flowers, 2004). Salt stress may negatively affectphotosynthesis by causing excess accumulation of leaf Na+ andCl−, stomatal closure and oxidative stress resulting in the forma-
Abbreviations: APX, ascorbate peroxidase; ATPS, ATP-sulfurylase; DAS, days aftersowing; GR, glutathione reductase; GSH, reduced glutathione; GSSG, oxidized glu-tathione; NR, nitrate reductase; NUE, nitrogen-use efficiency; ROS, reactive oxygenspecies; SA, salicylic acid; SOD, superoxide dismutase; TBARS, thiobarbituric acidreactive substances.
∗ Corresponding author. Tel.: +91 571 2702016; fax: +91 571 2704225.E-mail address: [email protected] (N.A. Khan).
tion of reactive oxygen species (ROS). Excessive amounts of ROScan enhance membrane lipid peroxidation and electrolyte leakage(Gunes et al., 2007) and damage chloroplast, inhibit photochem-ical reactions and decrease photosynthesis (Steduto et al., 2000).ROS are toxic by-products of stress metabolism, but also play animportant role as important signal transduction molecules in stresssignaling and regulation of acclimation responses (Miller et al.,2010). As an adaptation response, plants activate several mech-anisms to counteract the adverse effects of salt-induced ROS inplants. These mechanisms include ion homeostasis, detoxificationof ROS and the use of available resources which directly influ-ence photosynthesis. One of the defense mechanisms of plantsincludes the up-regulation of reduced glutathione (GSH), a lowmolecular weight antioxidant. GSH plays essential roles withinplant metabolism and stress tolerance to ROS (Szalai et al., 2009).The potential of GSH as antioxidant is related to the activity of glu-tathione reductase (GR), which catalyzes the regeneration of GSHfrom oxidized glutathione (GSSG). Nitrogen (N) and sulfur (S) areconstituents of GSH. Therefore, the activity of nitrate reductase (NR)and ATP-sulfurylase (ATPS), the enzymes involved in N and S assim-
0176-1617/$ – see front matter © 2010 Elsevier GmbH. All rights reserved.doi:10.1016/j.jplph.2010.11.001

Author's personal copy
808 R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815
ilation may influence GSH content in plant cell. Plants containinghigh activities of antioxidant enzymes have shown considerableresistance to oxidative damage caused by ROS (Khan et al., 2007;Gapinska et al., 2008; Frary et al., 2010; Syeed et al., 2010). Thesemechanisms of ion homeostasis, GSH synthesis and antioxidantenzymes may be influenced by plant growth regulators under saltstress.
Salicylic acid (SA) is a naturally occurring plant hormone, influ-ences various physiological and biochemical functions in plants,acts as an important signaling molecule and has diverse effectson tolerance to biotic and abiotic stress (Arfan et al., 2007; Syeedet al., 2010; Wang et al., 2010). Its role in plant tolerance to abioticstresses such as ozone, UV-B, heat, heavy metal and osmotic stress(El-Tayeb, 2005; Wang et al., 2010) has been reported. In contrast, ithas also been shown that high concentration of SA increases oxida-tive damage generated by NaCl in Arabidopsis (Borsani et al., 2001)and decreases drought tolerance in Zea mays (Nemeth et al., 2002).These reports show that SA has contrasting effects in inducingstress tolerance that may depend upon the species or concentrationof SA applied. Therefore, it is critical to identify the physiologicalprocesses of plants differing in salt sensitivity, and examine howmuch these processes are induced by SA application in tolerant andsensitive types to alleviate the decrease in photosynthesis undersalt stress. One of the objectives of the reported research was tocompare the physiological processes in Pusa Vishal (salt-tolerant)and T44 (salt-sensitive) cultivars of mungbean. The other objec-tive was to study the effectiveness of SA application in modulatingthese processes and alleviating decrease in photosynthesis undersalt stress. The cultivars, Pusa Vishal and T44 have been shown todiffer in sensitivity to salt stress (Khan and Syeed, 2003).
Materials and methods
Plant material and growth conditions
Healthy seeds of Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata L.) were surfacesterilized and sown in 15-cm-diameter earthen pots filledwith acid-washed sand purified according to Hewitt (1966).Plants grown in pots were kept in a greenhouse under natu-ral day/night conditions with photosynthetically active radiation(PAR) ∼900 ± 28 �mol m−2 s−1 and average day/night tempera-ture of 33/20 ± 2 ◦C. Plants (2 per pot) were subjected to either 0(control) or 50 mM NaCl. Pots were saturated daily with 300 mLof 50 mM NaCl in the form of modified full strength Hoagland’snutrient solution and the control group of plants were fed with300 mL nutrient solution. Salicylic acid (SA) was dissolved in abso-lute ethanol then added drop wise to water (ethanol/water: 1/1000v/v) (Williams et al., 2003). Fifteen days after sowing (DAS), SA wasapplied on the foliage of 0 and 50 mM NaCl grown plants at a con-centration of 0.1, 0.5 or 1.0 mM with a hand sprayer. A control groupof plants grown without NaCl and sprayed with ethanol/water(1/1000 v/v) was maintained. A surfactant teepol (0.5%) was addedwith the control and SA treatment solution. The volume of the spraywas 30 mL per pot. At 30 DAS (15 d after SA application), measure-ments were done and care was taken to select same age of leaves forthe determinations. The experiment followed a two-factor factorialrandomized complete block design and the number of replicates foreach treatment was three.
Content of ions
The content of Na+ and Cl− was determined in the digested plantsamples using Tri acid mixture (TAM), which is a mixture of nitricacid, sulfuric acid and perchloric acid in the ratio of 10:5:4. The
content of Na+ was estimated using flame photometer (Khera-391:Khera Instruments, New Delhi), whereas Cl− content was deter-mined by titration against 0.02 N silver nitrate solution using 5%K2CrO4 as indicator.
Activity of NR and N content
Activity of nitrate reductase (NR) in leaf was measured bypreparing enzyme extract using the method of Kuo et al. (1982).Leaves (1.0 g) were frozen in liquid N2, ground to a powder witha mortar and pestle, and then stored at −80 ◦C. The powder wasthawed for 10 min at 4 ◦C and homogenized in a blender in 250 mMTris–HCl buffer, pH 8.5, containing 10 mM cysteine, 1 mM EDTA,20 �M FAD, 1 mM DTT, and 10% (v/v) glycerol. The homogenatewas centrifuged at 10,000 × g for 30 min at 4 ◦C (REMI CPR24, NewDelhi, India). Activity of NR was assayed spectrophotometricallyas the rate of nitrite production at 28 ◦C adopting the procedureof Nakagawa et al. (1984). The assay mixture contained, KNO3(10 mM), HEPES (0.065 M; pH 7.0), NADH (0.5 mM) in phosphatebuffer (0.04 mM; pH 7.2) and enzyme in a final volume of 1.5 mL.The reaction was started by adding NADH. After 15 min the reac-tion was terminated by adding 1 mL of 1 N HCl solution containing1% sulfanilamide followed by adding 1 mL of 0.02% aqueous N-1-napthylethylene-di-amine-dihydrochloride (NED). The absorbancewas read at 540 nm after 10 min.
Leaf N content was determined in acid–peroxide digested mate-rial using the method of Lindner (1944).
Activity of ATPS and S content
Activity of ATP-sulfurylase (ATPS) was measured in leavesby the method of Lappartient and Touraine (1996). One gramfresh leaf tissue was ground at 4 ◦C in a buffer consisting of10 mM Na2EDTA, 20 mM Tris–HCl (pH 8.0), 2 mM dithiothre-itol (DTT), and 0.01 g mL−1 polyvinylpyrrolidone (PVP), using 1:4(w/v) tissue to buffer ratio. The homogenate was centrifuged at20,000 × g for 10 min at 4 ◦C. The supernatant was used for in vitroATP-sulfurylase assay. The enzyme activity was measured usingmolybdate-dependent formation of pyrophosphate. The reactionwas initiated by adding 0.1 mL of extract to 0.5 mL of the reactionmixture, which contained 7 mM MgCl2, 5 mM of Na2MoO4, 2 mM ofNa2ATP, and 0.032 units mL−1 of sulfate-free inorganic pyrophos-phate in 80 mM Tris–HCl buffer (pH 8.0). Another aliquot from thesame extract was added to the same reaction mixture except thatNa2MoO4 was absent. Incubations were carried out at 37 ◦C for15 min, after which phosphate was determined on a spectropho-tometer.
Content of S was determined in leaf samples digested in a mix-ture of concentrated HNO3 and 60% HClO4 (85:1,v/v) using theturbidimetric method of Chesnin and Yien (1950).
Content of H2O2 and TBARS and electrolyte leakage
The content of H2O2 was determined following the method ofOkuda et al. (1991). Fresh leaf tissues (50 mg) were ground in ice-cold 200 mM perchloric acid. After centrifugation at 1200 × g for10 min, perchloric acid of the supernatant was neutralized with4 M KOH. The insoluble potassium perchlorate was eliminated bycentrifugation at 500 × g for 3 min. The reaction was started by theaddition of peroxidase and the increase in absorbance was recordedat 590 nm for 3 min.
The level of lipid peroxidation in leaves was determined byestimating the content of thiobarbituric acid reactive substances(TBARS) as described by Dhindsa et al. (1981). Fresh leaf tis-sues (0.5 g) were ground in 0.25% 2-thiobarbituric acid in 10%trichloroacetic acid using mortar and pestle. After heating at

Author's personal copy
R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815 809
95 ◦C for 30 min, the mixture was quickly cooled on ice bathand centrifuged at 10,000 × g for 10 min. The absorbance of thesupernatant was read at 532 nm and corrected for non-specific tur-bidity by subtracting the absorbance of the same at 600 nm. Thecontent of TBARS was calculated using the extinction coefficient(155 mM−1 cm−1).
For measuring electrolyte leakage, samples were thoroughlywashed with sterile water, weighed and then kept in closed vialswith 10 mL of deionized water and were incubated at 25 ◦C for 6 husing shaker and electrical conductivity (EC) was determined (C1).Samples were then again kept at 90 ◦C for 2 h and EC was recordedafter attaining equilibrium at 25 ◦C (C2).
Assay of antioxidant enzymes
Fresh leaf tissues (200 mg) were homogenized with an extrac-tion buffer containing 0.05% (v/v) Triton X-100 and 1% (w/v)polyvinylpyrrolidone in potassium-phosphate buffer (100 mM, pH7.0) using chilled mortar and pestle. The homogenate was cen-trifuged at 15,000 × g for 20 min at 4 ◦C. The supernatant obtainedafter centrifugation was used for the assay of superoxide dismu-tase (SOD, EC 1.15.1.1) and glutathione reductase (GR, EC 1.6.4.2)enzymes. For the assay of ascorbate peroxidase (APX, EC 1.11.1.11),extraction buffer was supplemented with 2 mM ascorbate.
Activity of SOD was determined by monitoring the inhibition ofphotochemical reduction of nitro blue tetrazolium (NBT), accordingto the methods of Beyer and Fridovich (1987) and Giannopolitis andRies (1977). A 5 mL reaction mixture containing 5 mM HEPES (pH7.6), 0.1 mM EDTA, 50 mM Na2CO3 (pH 10.0), 13 mM methionine,0.025% (v/v) Triton X-100, 63 �mol NBT, 1.3 �mol riboflavin andthe enzyme extract was illuminated for 15 min (360 �mol m−2 s−1),and a control set was not illuminated to correct for backgroundabsorbance. A unit of SOD was defined as the amount of enzymerequired to cause 50% inhibition of the reaction of NBT at 560 nm.
Activity of APX was determined by adopting the method ofNakano and Asada (1981) by recording the decrease in absorbanceof ascorbate at 290 nm. The assay mixture contained phosphatebuffer (50 mM, pH 7.0), 0.1 mM EDTA, 0.5 mM ascorbate, 0.1 mMH2O2 and the enzyme extract. Activity of APX was calculated byusing the extinction coefficient 2.8 mM−1 cm−1. One unit of enzymewas the amount necessary to decompose 1 �mol of substrate permin at 25 ◦C.
Activity of GR was determined by the method of Foyer andHalliwell (1976) by monitoring the glutathione dependent oxida-tion of NADPH at 340 nm. The assay mixture contained phosphatebuffer (25 mM, pH 7.8), 0.5 mM oxidized glutathione (GSSG),0.2 mM NADPH and the enzyme extract. The activity of GR was cal-culated by using extinction coefficient 6.2 mM−1 cm−1. One unit ofenzyme was the amount necessary to decompose 1 �mol of NADPHper min at 25 ◦C.
Content of GSH and GSSG
Glutathione was assayed by an enzymic recycling procedure(Griffith, 1980) in which it was sequentially oxidized by 5,5′-dithiobis-2-nitrobenzoic acid (DTNB) and reduced by NADPH in thepresence of GR. For specific assay of GSSG, the reduced glutathione(GSH) was masked by derivatization with 2-vinylpyridine. Freshleaf tissues (0.5 g) were ground in liquid nitrogen using mortarand pestle. The ground tissue was suspended in 0.5 mL 5% sulfos-alicyclic acid and centrifuged at 12,000 × g for 10 min. A 300 �Laliquot of supernatant was removed and neutralized by additionof 18 �L 7.5 M triethanolamine. One 150 �L sample was then usedto determine concentrations of GSH plus GSSG. Another samplewas pretreated with 3 �L 2-vinylpyridine for 60 min at 20 ◦C tomask the GSH by derivatization, to allow the subsequent determi-
nation of GSSG alone. In each case, 50 �L aliquots of the two typesof sample were mixed with 700 �L 0.3 mM NADPH, 100 �L DTNBand 150 �L buffer containing 125 mM sodium phosphate, 6.3 mMEDTA (pH 6.5). A 10 �L aliquot of GR (5 U mL−1) was then addedand the change in absorbance at 412 nm monitored at 30 ◦C.
Leaf water potential, osmotic potential and photosynthetic traits
Leaf water potential was measured on second leaf from top(fully expanded youngest leaf) of the plant by using water potentialsystem (Psypro, WESCOR, USA). The leaf used for water potentialmeasurement was frozen in liquid nitrogen in sealed polythenebags which was thawed and cell sap was extracted with the helpof a disposable syringe. The extracted sap was used for the deter-mination of osmotic potential using a vapour pressure osmometer(5520, WESCOR, USA).
Net photosynthesis, stomatal conductance and intercellularCO2 concentration were measured in fully expanded uppermostleaves of plants in each treatment using infra red gas analyzer(LiCOR-6200, Portable Photosynthesis System, NA, USA). The mea-surements were done between 11.00 and 12.00 h at light saturatingintensity and at 370 ± 5 �mol mol−1 atmospheric CO2 concentra-tions. Chlorophyll fluorescence (Fv/Fm) of fully expanded leaf wasmeasured in vivo after 0.5 h dark adaptation of the leaves usingchlorophyll fluorometer (OS-30p, USA). Photosynthetic nitrogen-use efficiency (NUE) was calculated by the ratio of photosynthesisrate to N content per unit leaf area.
Statistical analysis
Data were statistically analyzed using analysis of variance(ANOVA) by SPSS 10.0 for Windows, and presented as treatmentmean ± SE of three measurements. Least significant difference(LSD) was calculated for the significant data at P < 0.05. Bars show-ing the same letter are not significantly different by LSD test atP < 0.05.
Results
Leaf and root Na+ and Cl− content
Salt treatment resulted in higher leaf Na+ and Cl− content incomparison to control in both the cultivars, but T44 exhibitedgreater accumulation than Pusa Vishal. Plants treated with 0.1 or0.5 mM SA exhibited leaf Na+ and Cl− content lesser than the con-trol in both the cultivars. Maximum reduction in the content of leafNa+ and Cl− was noted with 0.5 mM SA in Pusa Vishal. Applica-tion of 0.5 mM SA resulted in reduction of leaf Na+ and Cl− contentby 27.8% and 46.7% in Pusa Vishal and 10.0% and 36.5% in T44 incomparison to control (Fig. 1).
In contrast to greater accumulation of Na+ and Cl− in leaf in T44,Pusa Vishal accumulated greater content in root. SA at 0.1 mM and0.5 mM proved equally effective in reducing root Na+ content inPusa Vishal, but 0.5 mM was more effective in reducing root Na+
content in T44. Treatment of NaCl-grown plants with 0.5 mM SAresulted in lowering of root Na+ and Cl− by 27.2% and 25.6% in PusaVishal, and 37.5% and 29.9% in T44 in comparison to control.
High concentration of SA (1.0 mM) alone did not influence rootor leaf Na+ and Cl− content, but 1.0 mM SA plus NaCl increased thesecontents in comparison to control (Fig. 1).
Activity of NR and N content
Salt stress decreased NR activity in both the cultivars, but T44showed higher decrease than Pusa Vishal. Under non-saline condi-tion, application of 0.1 and 0.5 mM SA increased NR activity in both

Author's personal copy
810 R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815
Fig. 1. Root and leaf Na+ and Cl− content in Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with 50 mM NaCl andtreated with foliar 0.1–1.0 mM SA at 30 days after sowing (DAS). Data are presentedas treatments mean ± SE. Data followed by same letter are not significantly differentby LSD test at P < 0.05.
the cultivars, but maximum increase was observed with 0.5 mMSA in Pusa Vishal. An increase of 46.9% in Pusa Vishal and 33.7% inT44 in NR activity in comparison to control was registered with theapplication of 0.5 mM SA. Application of 0.1 mM SA could only alle-viate the effect of salt stress on NR activity, but 0.5 mM SA promotedthe NR activity more conspicuously in Pusa Vishal. The increasein NR activity in Pusa Vishal and T44 with 0.5 mM SA under saltstress was 27.8% and 10.3%, respectively compared to control. SA at1.0 mM given alone or with NaCl inhibited the NR activity in boththe cultivars (Fig. 2).
Salt stress decreased N content by 23.3% in Pusa Vishal and46.8% in T44 in comparison to control. SA application (0.1–0.5 mM)to plants grown without NaCl enhanced N content, but the effectwas more pronounced with 0.5 mM SA (Fig. 2). The application of0.1 mM SA on plants grown with NaCl further reduced N contentcompared to control in both the cultivars, whereas application of
Fig. 2. Leaf NR activity and N content in Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with 50 mM NaCland treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are presented as treatmentsmean ± SE. Data followed by same letter are not significantly different by LSD testat P < 0.05.
0.5 mM SA promoted N content by 27.9% in Pusa Vishal and 16.7%in T44 in comparison to control. Treatment of 1.0 mM SA plus NaClinhibited N content in both the cultivars (Fig. 2).
Activity of ATPS and S content
Salt stress increased ATPS activity and S content to a greaterextent in Pusa Vishal than T44. Application of 0.5 mM SA resultedin the increase of ATPS activity by 88.9% in Pusa Vishal and 64.0%in T44 under non-saline condition, and 116.1% in Pusa Vishal and81.0% in T44 under salt stress. Plants grown with NaCl and treatedwith 0.1 or 0.5 mM SA showed further increase in ATPS activity.Maximum ATPS activity was recorded with 0.5 mM SA plus NaClin Pusa Vishal. The effect of 1.0 mM SA plus NaCl proved inhibitoryon ATPS activity in Pusa Vishal. However, the effect was equal withcontrol in T44 (Fig. 3). Similarly, maximum increase of 115.4% inPusa Vishal and 97.0% in T44 in S content in comparison to controlwas noted 0.5 mM SA under salt stress. The effect of 1.0 mM SAplus NaCl on S content was found inhibitory in both the cultivars(Fig. 3).
Content of H2O2 and TBARS and electrolyte leakage
Salt stress increased H2O2 and thiobarbituric acid reactive sub-stances (TBARS) content nearly by three-fold in both the cultivars,and electrolyte leakage by five-fold in Pusa Vishal and six-fold inT44 in comparison to control. Application of 0.5 mM SA under non-saline condition significantly and maximally reduced H2O2 andTBARS content in both the cultivars compared to control, but elec-trolyte leakage was significantly reduced in Pusa Vishal only. SA(0.1–1.0 mM) applied on plants grown with NaCl resulted in lessercontent of H2O2 and TBARS than the plants grown with NaCl alonein both the cultivars. SA application proved effective in reducing theeffect of salt stress on electrolyte leakage significantly only in PusaVishal. The effect of 1.0 mM SA under non-saline conditions did not

Author's personal copy
R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815 811
Fig. 3. Leaf ATPS activity and S content in Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with 50 mM NaCland treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are presented as treatmentsmean ± SE. Data followed by same letter are not significantly different by LSD testat P < 0.05.
significantly affect the content of H2O2 and TBARS and electrolyteleakage (Fig. 4).
Antioxidant enzymes
The activity of antioxidant enzymes increased under salt stressin both the cultivars and was higher in Pusa Vishal than T44, exceptfor superoxide dismutase (SOD) which was higher in T44. Salt stressincreased SOD, ascorbate peroxidase (APX) and GR by 71.4%, 62.8%and 7.1% in Pusa Vishal and 120.0%, 38.1%, and 4.1% in T44, respec-tively compared to control (Fig. 5). Application of 0.1 or 0.5 mMSA under non-saline condition decreased SOD activity in the cul-tivars, but SA plus salt increased the activity. Activity of APX andGR was induced with SA or SA plus salt. Application of 0.5 mM SAwithout NaCl resulted in increase in the activity of APX and GR by144.8% and 28.6% in Pusa Vishal and 96.6% and 25.0% in T44, respec-tively compared to control. Application of 0.5 mM SA plus NaClmaximally increased the activity of antioxidant enzymes, showingabout a three-fold increase in APX in Pusa Vishal and two-fold inT44, while GR activity was increased to a lesser extent compared tocontrol. The application of 1.0 mM SA on plants grown with or with-out NaCl proved inhibitory and decreased the activity of antioxidantenzymes in both the cultivars compared to their respective control(Fig. 5).
Content of GSH and GSSG
Salt treatment increased content of both GSH and GSSG in com-parison to control, and decreased the redox state of glutathione(GSH/GSSG) in both the cultivars, but to a greater extent in T44 thanPusa Vishal. SA application at 0.5 mM resulted in maximal increasein the redox state of glutathione under non-saline condition. Thisincrease was higher in Pusa Vishal than T44. Salt treatment plus0.5 mM SA decreased GSSG and increased GSH content to a greater
Fig. 4. Leaf H2O2 and TBARS content and electrolyte leakage Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grownwith 50 mM NaCl and treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are pre-sented as treatments mean ± SE. Data followed by same letter are not significantlydifferent by LSD test at P < 0.05.
extent in Pusa Vishal than T44. Plants grown with NaCl and treatedwith 1.0 mM SA exhibited lesser content of GSH than control,whereas GSSG content was higher than control (Fig. 6).
Leaf water potential and osmotic potential
Salt treatment decreased water potential and osmotic poten-tial by 68.7% and 51.7% in Pusa Vishal and 70.0% and 69.4% inT44, respectively compared to control. Under non-saline condi-tion, application of 0.1 and 0.5 mM SA significantly increased thewater and osmotic potential but the effect was more pronouncedwith 0.5 mM SA application. Supplementation of 0.5 mM SA to NaCltreated plants proved ameliorative and enhanced the water andosmotic potential by 53.7% and 33.6% in Pusa Vishal and 32.7% and15.3% in T44 in comparison to their respective control. The effect of1.0 mM SA application alone and/or in salt stressed plants provedineffective and further decreased water and osmotic potential inboth the cultivars (Fig. 7).
Photosynthetic traits
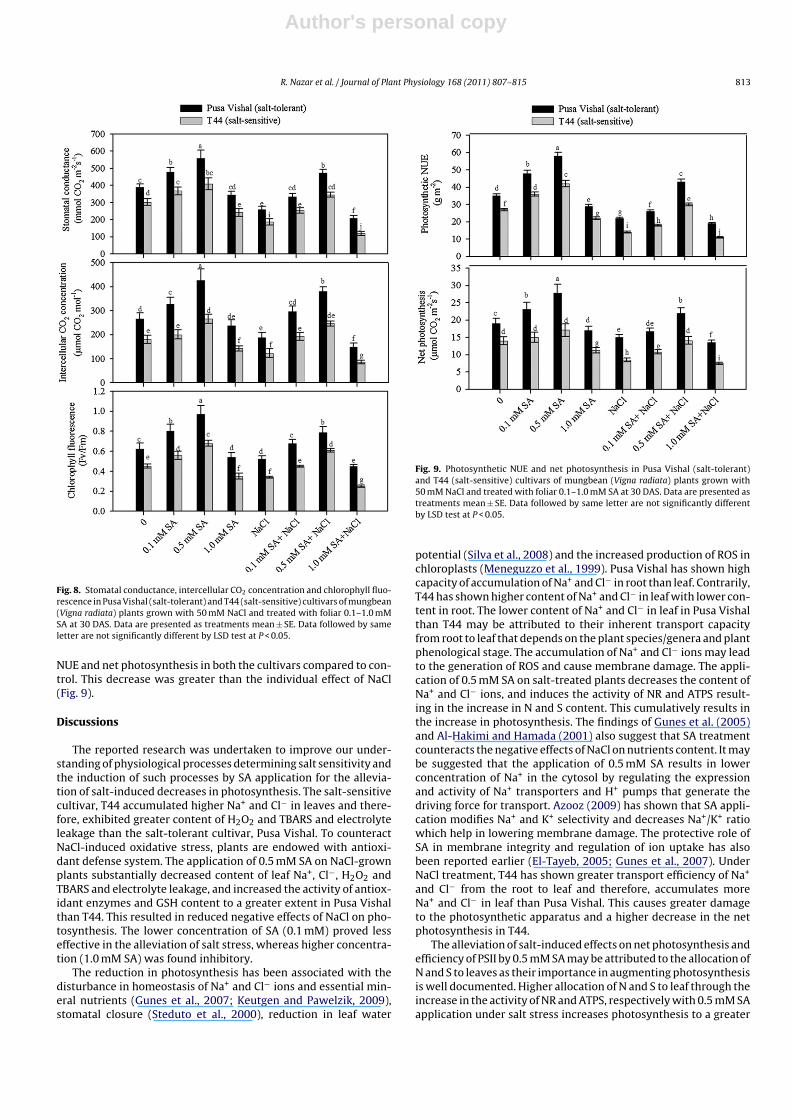
Salt stress significantly reduced stomatal conductance, inter-cellular CO2 concentration and chlorophyll fluorescence by 33.2%,29.4% and 16.1% in Pusa Vishal and 38.3%, 31.7% and 24.6% in T44compared to control. Under non-saline condition, SA at 0.1 and

Author's personal copy
812 R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815
Fig. 5. Activity of leaf SOD, APX and GR in Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with 50 mM NaCland treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are presented as treatmentsmean ± SE. Data followed by same letter are not significantly different by LSD testat P < 0.05.
0.5 mM increased these photosynthetic traits, but application of1.0 mM SA decreased these traits in both the cultivars compared tocontrol. SA application at 0.1 and 0.5 mM SA on plants grown withNaCl proved effective in alleviating salt stress in both the culti-vars. Pusa Vishal was more responsive to SA and exhibited greatercapacity for amelioration and promotion of photosynthetic traitsthan T44. Application of 1.0 mM SA proved inhibitory (Fig. 8).
Photosynthetic nitrogen-use efficiency (NUE) and net photosyn-thesis decreased in both the cultivars under salt stress by 31.5%and 21.0% in Pusa Vishal and 48.1% and 39.3% in T44, respec-tively compared to control. Under non-saline and saline conditions,plants showed differential response to SA treatment. SA at 0.1and 0.5 mM concentration under non-saline condition promotedphotosynthetic NUE in both the cultivars, but could promote netphotosynthesis only in Pusa Vishal compared to control. Applica-tion of 0.1 mM SA alleviated salt stress effects on photosyntheticNUE in both the cultivars, but could alleviate salt stress effects onphotosynthesis in T44 only. Application of SA at 0.5 mM under saltstress promoted photosynthetic NUE in both the cultivars com-pared to control, but could promote photosynthesis only in PusaVishal. In T44, the effect of salt stress was ameliorative with 0.5 mMSA. Application of 0.5 mM SA to plants grown with NaCl resulted inthe increase of photosynthetic NUE by 22.8% in Pusa Vishal and11.1% in T44, and photosynthesis by 15.8% in Pusa Vishal and 0.7%in T44. SA at 1.0 mM under salt stress decreased photosynthetic
Fig. 6. Leaf reduced GSH and GSSG content in Pusa Vishal (salt-tolerant) and T44(salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with 50 mM NaCland treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are presented as treatmentsmean ± SE. Data followed by same letter are not significantly different by LSD testat P < 0.05.
Fig. 7. Leaf water potential and osmotic potential in Pusa Vishal (salt-tolerant) andT44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with 50 mMNaCl and treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are presented as treat-ments mean ± SE. Data followed by same letter are not significantly different by LSDtest at P < 0.05.
Author's personal copy
R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815 813
Fig. 8. Stomatal conductance, intercellular CO2 concentration and chlorophyll fluo-rescence in Pusa Vishal (salt-tolerant) and T44 (salt-sensitive) cultivars of mungbean(Vigna radiata) plants grown with 50 mM NaCl and treated with foliar 0.1–1.0 mMSA at 30 DAS. Data are presented as treatments mean ± SE. Data followed by sameletter are not significantly different by LSD test at P < 0.05.
NUE and net photosynthesis in both the cultivars compared to con-trol. This decrease was greater than the individual effect of NaCl(Fig. 9).
Discussions
The reported research was undertaken to improve our under-standing of physiological processes determining salt sensitivity andthe induction of such processes by SA application for the allevia-tion of salt-induced decreases in photosynthesis. The salt-sensitivecultivar, T44 accumulated higher Na+ and Cl− in leaves and there-fore, exhibited greater content of H2O2 and TBARS and electrolyteleakage than the salt-tolerant cultivar, Pusa Vishal. To counteractNaCl-induced oxidative stress, plants are endowed with antioxi-dant defense system. The application of 0.5 mM SA on NaCl-grownplants substantially decreased content of leaf Na+, Cl−, H2O2 andTBARS and electrolyte leakage, and increased the activity of antiox-idant enzymes and GSH content to a greater extent in Pusa Vishalthan T44. This resulted in reduced negative effects of NaCl on pho-tosynthesis. The lower concentration of SA (0.1 mM) proved lesseffective in the alleviation of salt stress, whereas higher concentra-tion (1.0 mM SA) was found inhibitory.
The reduction in photosynthesis has been associated with thedisturbance in homeostasis of Na+ and Cl− ions and essential min-eral nutrients (Gunes et al., 2007; Keutgen and Pawelzik, 2009),stomatal closure (Steduto et al., 2000), reduction in leaf water
Fig. 9. Photosynthetic NUE and net photosynthesis in Pusa Vishal (salt-tolerant)and T44 (salt-sensitive) cultivars of mungbean (Vigna radiata) plants grown with50 mM NaCl and treated with foliar 0.1–1.0 mM SA at 30 DAS. Data are presented astreatments mean ± SE. Data followed by same letter are not significantly differentby LSD test at P < 0.05.
potential (Silva et al., 2008) and the increased production of ROS inchloroplasts (Meneguzzo et al., 1999). Pusa Vishal has shown highcapacity of accumulation of Na+ and Cl− in root than leaf. Contrarily,T44 has shown higher content of Na+ and Cl− in leaf with lower con-tent in root. The lower content of Na+ and Cl− in leaf in Pusa Vishalthan T44 may be attributed to their inherent transport capacityfrom root to leaf that depends on the plant species/genera and plantphenological stage. The accumulation of Na+ and Cl− ions may leadto the generation of ROS and cause membrane damage. The appli-cation of 0.5 mM SA on salt-treated plants decreases the content ofNa+ and Cl− ions, and induces the activity of NR and ATPS result-ing in the increase in N and S content. This cumulatively results inthe increase in photosynthesis. The findings of Gunes et al. (2005)and Al-Hakimi and Hamada (2001) also suggest that SA treatmentcounteracts the negative effects of NaCl on nutrients content. It maybe suggested that the application of 0.5 mM SA results in lowerconcentration of Na+ in the cytosol by regulating the expressionand activity of Na+ transporters and H+ pumps that generate thedriving force for transport. Azooz (2009) has shown that SA appli-cation modifies Na+ and K+ selectivity and decreases Na+/K+ ratiowhich help in lowering membrane damage. The protective role ofSA in membrane integrity and regulation of ion uptake has alsobeen reported earlier (El-Tayeb, 2005; Gunes et al., 2007). UnderNaCl treatment, T44 has shown greater transport efficiency of Na+
and Cl− from the root to leaf and therefore, accumulates moreNa+ and Cl− in leaf than Pusa Vishal. This causes greater damageto the photosynthetic apparatus and a higher decrease in the netphotosynthesis in T44.
The alleviation of salt-induced effects on net photosynthesis andefficiency of PSII by 0.5 mM SA may be attributed to the allocation ofN and S to leaves as their importance in augmenting photosynthesisis well documented. Higher allocation of N and S to leaf through theincrease in the activity of NR and ATPS, respectively with 0.5 mM SAapplication under salt stress increases photosynthesis to a greater

Author's personal copy
814 R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815
extent in Pusa Vishal than T44. Nitrogen influences the structureand composition of photosynthetic apparatus. In crops, ribulose 1,5,bisphosphate carboxylase (rubisco) content increases linearly withleaf N accumulation (Kumar et al., 2002). In the present study, N andS incorporate more efficiently in Pusa Vishal than T44 in thiol (–SH)groups of proteins (cysteine) and its immediate metabolite GSH. Ina cascade of enzymatic steps inorganic S is converted to the nutri-tionally important N and S-containing amino acid cysteine (Koprivaand Rennenberg, 2004; Kopriva, 2006). Cysteine is the essentialprecursor of all organic molecules containing reduced sulfur includ-ing GSH. GSH displays essential roles within plant metabolism andstress tolerance to ROS (Rausch et al., 2007). With the application of0.5 mM SA plants invests a larger fraction of leaf N and S into pho-tosynthetic apparatus under non-saline and salt stress conditions.A substantial increase in GSH content in Pusa Vishal with 0.5 mMSA application under salt stress helps in reversing the effects ofNaCl-induced ROS on photosynthesis. Thus, enhanced activity ofNR and ATPS together with GSH content resulted in lower con-tent of TBARS and H2O2 and electrolyte leakage in Pusa Vishal. Thiscumulatively resulted in greater alleviation of NaCl-induced salin-ity effects on photosynthesis in Pusa Vishal than T44. The reporton the effect of SA on the activity of NR and ATPS and concur-rent GSH content under salt stress is not available in literature.However, we have earlier reported that higher ATPS activity inmustard is necessary for the maintenance of optimal GSH levelsrequired for the proper functioning of ascorbate–glutathione path-way (Khan et al., 2009). Further, the increased content of GSH with Sapplication has been found to protect photosynthesis of cadmium-treated mustard (Anjum et al., 2008). Higher photosynthetic NUEwith 0.5 mM SA under salt stress provides further evidence thatN allocation to the photosynthetic apparatus is a major factor forthe increase in photosynthesis. Studies have suggested that plantswith lower photosynthetic NUE bears less ability to allocate N tothe photosynthetic apparatus (Takashima et al., 2004).
The increase in photosynthesis with 0.5 mM SA applicationunder salt stress in both the cultivars with concurrent increasein stomatal conductance and intercellular CO2 concentration sug-gests stomatal and non-stomatal limitation to photosynthesis, andshow that CO2 is utilized more efficiently by 0.5 mM SA applicationunder salt stress. Larque-Saavedra (1978), in contrast to this hasobserved that application of SA causes an antitranspiration effecton the leaves of Phaseolus vulgaris and decreases stomatal conduc-tance and intercellular CO2 concentration. The control of stomatalfunctioning under stress may also be related to changes in abscisicacid (ABA). In a study of Rai et al. (1986), SA was found to reverseABA-controlled stomatal closure, but Popova et al. (1995) haveshown that ABA alleviates the inhibitory effect of NaCl on growth,photosynthesis and translocation of assimilates.
Salt stress induces water deficit and increases ionic and osmoticeffects leading to the generation of ROS and oxidative stress (Paridaand Das, 2005). ROS acts as a signal molecule and plants ini-tiate antioxidant mechanism for protection against ROS (Khanet al., 2003). In the present study, salt stress decreases waterpotential and osmotic potential maximally in T44. This inducesmaximum oxidative stress and greatest content of H2O2 and TBARSand electrolyte leakage in T44 under salt stress. The applicationof SA (0.1–1.0 mM) increases SOD activity under salt stress inboth the cultivars, but to a greater extent in T44. This results inhigher H2O2 production in T44 than Pusa Vishal. Efficient destruc-tion of H2O2 in chloroplast requires the induction of APX viaascorbate–glutathione pathway. The application of 0.5 mM SA tosalt-stressed plants maximally induces APX activity in Pusa Vishal,exhibiting greater potential in this cultivar to detoxify H2O2. PusaVishal also shows higher constitutive and 0.1–0.5 mM SA-inducedGR, which enables this cultivar to detoxify ROS more efficiently thanT44. The high ratio of GSH to GSSG, occurring under optimal growth
conditions can be restored by means of higher GR activity (Szalaiet al., 2009). Moreover, the increased GSH content maintains theproper functioning of enzymes of ascorbate–glutathione pathway,and results in the increased activity of GR and APX with the appli-cation of 0.5 mM SA under salt stress. The increase in antioxidantenzymes with SA has been reported by Khan et al. (2003), Wang andLi (2006), Palma et al. (2009) and Syeed et al. (2010) under differentstress conditions.
The activity of APX and GR and the GSH content were inhib-ited by 1.0 mM SA application leading to maximum production ofH2O2, TBARS and electrolyte leakage. Durner and Klessig (1995)provided the evidence that inhibition of APX with higher concen-tration of SA blocked the H2O2 removal in the plant cell leadingto the increased levels of endogenous H2O2. Treatment of tobaccoplants with 1.0 mM SA induced 50–60% increase in endogenousH2O2 levels. Nemeth et al. (2002) also reported inhibitory effectof high concentration of exogenously applied SA on maize. Borsaniet al. (2001) demonstrated that transgenic Arabidopsis plants withlower endogenous SA were better able to resist the oxidative dam-age caused by salt stress than the wild type plants.
Conclusion
It is concluded that the low sensitivity of Pusa Vishal to saltstress was due to its capacity to accumulate less content of Na+ andCl− in leaf. This cultivar also exhibited lower activity of SOD andhigher activity of APX and GR and GSH content. These traits helpedto remove salt-induced ROS in this cultivar more efficiently thanT44. In contrast, Na+ and Cl− accumulation in leaf was higher thanroot in T44, and this cultivar exhibited higher SOD activity than PusaVishal showing greater oxidative stress. SA-treated plants exhib-ited changes in all physiological processes to maximize the useefficiency of N and S through higher activity of NR and ATPS andsynthesis of GSH. The higher synthesis of GSH in turn helped toincrease the antioxidant enzymes GR and APX. The effect of SAapplication in inducing physiological processes was more conspic-uous in Pusa Vishal than T44. SA alleviates the negative effects ofsalt stress on photosynthesis depending on the concentration of SAused. Maximum alleviation was found with 0.5 mM SA application.
References
Al-Hakimi AMA, Hamada AM. Counteraction of salinity stress on wheat plantsby grain soaking in ascorbic acid, thiamin or sodium salicylate. Biol Plant2001;44:253–61.
Anjum NA, Umar S, Ahmad A, Iqbal M, Khan NA. Sulphur protects mustard (Bras-sica campestris L.) from cadmium toxicity by improving leaf ascorbate andglutathione. Plant Growth Regul 2008;54:271–9.
Arfan M, Athar HR, Ashraf M. Does exogenous application of salicylic acid throughthe rooting medium modulate growth and photosynthetic capacity in twodifferently adapted spring wheat cultivars under salt stress? J Plant Physiol2007;164:685–94.
Azooz MM. Salt stress mitigation by seed priming with salicylic acid in two fababean genotypes differing in salt tolerance. Int J Agric Biol 2009;11:343–50.
Beyer WF, Fridovich I. Assaying for superoxide dismutase activity: some large con-sequences of minor changes in conditions. Anal Biochem 1987;161:559–66.
Borsani O, Valpuesta V, Botella MA. Evidence for a role of salicylic acid in the oxida-tive damage generated by NaCl and osmotic stress in Arabidopsis seedlings. PlantPhysiol 2001;126:1024–34.
Chesnin L, Yien CH. Turbidimetric determination of available sulphates. Soil Sci SocAm Proc 1950;15:149–51.
Dhindsa RH, Plumb-Dhindsa P, Thorpe TA. Leaf senescence correlated with increasedlevel of membrane permeability, lipid peroxidation and decreased level of SODand CAT. J Exp Bot 1981;32:93–101.
Durner J, Klessig DF. Inhibition of ascorbate peroxidase by salicylic acid and 2,6-dichloro-isonicotinic acid, two inducers of plant defense responses. Proc NatlAcad Sci USA 1995;92:11312–6.
El-Tayeb MA. Response of barley grains to the interactive effect of salinity andsalicylic acid. Plant Growth Regul 2005;45:215–24.
Flowers T. Improving crop salt tolerance. J Exp Bot 2004;55:307–19.Foyer CH, Halliwell B. The presence of glutathione and glutathione reduc-
tase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta1976;133:21–5.

Author's personal copy
R. Nazar et al. / Journal of Plant Physiology 168 (2011) 807–815 815
Frary A, Gol D, Keles D, Okmen B, Pinar H, Sigva HO, et al. Salt tolerance in Solanumpennellii: antioxidant response and related QTL. BMC Plant Biol 2010;10:58.
Gapinska M, Sklodowska M, Gabara B. Effect of short- and long-term salinity on theactivities of antioxidative enzymes and lipid peroxidation in tomato roots. ActaPhysiol Plant 2008;30:11–8.
Giannopolitis CN, Ries SK. Superoxide dismutase. I. Occurrence in higher plants.Plant Physiol 1977;59:309–14.
Griffith OW. Determination of glutathione disulphide using glutathione reductaseand 2-vinylpyridine. Anal Biochem 1980;106:207–12.
Gunes A, Inal A, Alpaslan M, Cicek N, Guneri E, Eraslan F, et al. Effects of exogenouslyapplied salicylic acid on the induction of multiple stress tolerance and mineralnutrition in maize (Zea mays L.). Arch Agron Soil Sci 2005;51:687–95.
Gunes A, Inal A, Alpaslan M, Eraslan F, Bagci EG, Cicek N. Salicylic acid inducedchanges on some physiological parameters symptomatic for oxidative stressand mineral nutrition in maize (Zea mays L.) grown under salinity. J Plant Physiol2007;164:728–36.
Hewitt EJ. Sand and water culture methods used in the study of plant nutrition.England: Commonwealth Agricultural Bureaux; 1966.
Keutgen AJ, Pawelzik E. Impact of NaCl stress on plant growth and mineral nutrientassimilation in two cultivars of strawberry. Environ Exp Bot 2009;65:170–6.
Khan NA, Syeed S. Low activity of plasma membrane transporters for sodium toler-ance in Vigna radiata to salinity stress. Indian J Trop Biodiv 2003;11:49–52.
Khan NA, Samiullah, Singh S, Nazar R. Activities of antioxidative enzymes, sulphurassimilation, photosynthetic activity and growth of wheat (Triticum aestivum)cultivars differing in yield potential under cadmium stress. J Agron Crop Sci2007;193:435–44.
Khan NA, Nazar R, Anjum NA. Growth, photosynthesis and antioxidant metabolismin mustard (Brassica juncea L.) cultivars differing in ATP-sulfurylase activityunder salinity stress. Sci Hort 2009;122:455–60.
Khan W, Prithviraj B, Smith DL. Photosynthetic responses of corn and soybean tofoliar application of salicylates. J Plant Physiol 2003;160:485–92.
Kopriva S. Regulation of sulfate assimilation in Arabidopsis and beyond. Ann Bot2006;97:479–95.
Kopriva S, Rennenberg H. Control of sulphate assimilation and glutathione synthesis:interaction with N and C metabolism. J Exp Bot 2004;55:1831–42.
Kumar PA, Parry MAJ, Mitchell RAC, Ahmad A, Abrol YP. Photosynthesis and nitro-gen use efficiency. In: Foyer CH, Noctor G, editors. Photosynthetic nitrogenassimilation and associated carbon and respiratory metabolism: advances inphotosynthesis and respiration. Dordrecht: Springer; 2002. p. 23–34.
Kuo TM, Warner RL, Kleinhofs A. In vitro stability of nitrate reductase from barleyleaves. Phytochemistry 1982;21:531–3.
Lappartient AG, Touraine B. Demand-driven control of root ATP sulfurylase activityand SO4
2− uptake in intact canola. Plant Physiol 1996;111:147–57.Larque-Saavedra A. The antitranspirant effect of acetylsalicylic acid on Phaseolus
vulgaris. Physiol Plant 1978;43:126–8.Lindner RC. Rapid analytical methods for some of the more common inorganic con-
stituents of plant tissues. Plant Physiol 1944;19:70–89.Meneguzzo S, Navarri-Izzo F, Izzo R. Antioxidative responses of shoots and
roots of wheat to increasing NaCl concentrations. J Plant Physiol 1999;155:274–80.
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R. Reactive oxygen species homoestasisand signaling during and salinity stresses. Plant Cell Environ 2010;33:453–67.
Nakagawa H, Poulle M, Oaks A. Characterization of nitrate reductase from corn leaves(Zea mays cv. W64A × W182E). Plant Physiol 1984;75:285–9.
Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate specific peroxidasein spinach chloroplasts. Plant Cell Physiol 1981;22:867–80.
Nemeth M, Janda T, Horvath E, Paldi E, Szalai G. Exogenous salicylic acid increasespolyamine content but may decrease drought tolerance in maize. Plant Sci2002;162:569–74.
Okuda T, Masuda Y, Yamanka A, Sagisaka S. Abrupt increase in the level of hydrogenperoxide in leaves of winter wheat is caused by cold treatment. Plant Physiol1991;97:1265–7.
Palma F, Lluch C, Iribarne C, Garcia-Garrida JM, Tejera Garcia NA. Combinedeffect of salicylic acid and salinity on some antioxidant activities, oxidativestress and metabolite accumulation in Phaseolus vulgaris. Plant Growth Regul2009;58:307–16.
Parida AK, Das AB. Salt tolerance and salinity effects on plants: a review. EcotoxicolEnviron Saf 2005;60:324–49.
Popova LP, Stoinova ZG, Maslenkova LT. Involvement of abscisic acid in photosyn-thetic process in Hordeum vulgare L. during salinity stress. J Plant Growth Regul1995;14:211–8.
Rai VK, Sharma SS, Sharma S. Reversal of ABA-induced stomatal closure by phenoliccompounds. J Exp Bot 1986;37:129–34.
Rausch T, Gromes R, Liedschulle V, Muller I, Bogs J, Galovic V, et al. Novel insight intothe regulation of GSH biosynthesis in higher plants. Plant Biol 2007;9:565–72.
Silva C, Martínez V, Carvajal M. Osmotic versus toxic effects of NaCl on pepper plants.Biol Plant 2008;52:72–9.
Steduto P, Albrizio R, Giorio P, Sorrentino G. Gas exchange response and stomataland non-stomatal limitations to carbon assimilation of sunflower under salinity.Environ Exp Bot 2000;44:243–55.
Syeed S, Anjum NA, Nazar R, Iqbal N, Masood A, Khan NA. Salicylic acid-mediatedchanges in photosynthesis, nutrients content and antioxidant metabolism intwo mustard (Brassica juncea L.) cultivars differing in salt tolerance. Acta PhysiolPlant 2010., doi:10.1007/s11738-010-12614-7.
Szalai G, Kellos T, Galiba G, Kocsy G. Glutathione as an antioxidant and regula-tory molecule in plants under abiotic stress conditions. J Plant Growth Regul2009;28:66–80.
Takashima T, Hikosaka K, Hirose T. Photosynthesis or persistence: nitrogen alloca-tion in leaves of evergreen and deciduous Quercus species. Plant Cell Environ2004;7:1047–54.
Tester M, Devenport R. Na+ tolerance and Na+ transport in higher plants. Ann Biol2003;91:503–7.
Wang LJ, Li SH. Salicylic acid-induced heat or cold tolerance in relation to Ca2+ home-ostasis and antioxidant system in young grape plants. Plant Sci 2006;170:454–9.
Wang LJ, Fan L, Loescher W, Duan W, Liu GJ, Cheng JS, et al. Salicylic acid allevi-ates decreases in photosynthesis under heat stress and accelerates recovery ingrapevine leaves. BMC Plant Biol 2010;10:34.
Williams M, Senaratna T, Dixon K, Sivasithamparam K. Benzoic acid induces toler-ance to biotic stress caused by Phytophthora cinnamomi in Banksai attenuate.Plant Growth Regul 2003;41:89–91.




















