
Roles of synorganisation, zygomorphy and heterotopy in floral evolution: the gynostemium and...
39
Biol. Rev. (2002), 77, pp. 403–441 " Cambridge Philosophical Society DOI : 10.1017}S1464793102005936 Printed in the United Kingdom 403 Roles of synorganisation, zygomorphy and heterotopy in floral evolution : the gynostemium and labellum of orchids and other lilioid monocots PAULA J. RUDALL" and RICHARD M. BATEMAN# " Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AB, UK # Department of Botany, Natural History Museum, Cromwell Road, London, SW7 5BD, UK (Received 30 July 2001 ; revised 21 February 2002 ; accepted 7 March 2002) ABSTRACT A gynostemium, comprising stamen filaments adnate to a syncarpous style, occurs in only three groups of monocots : the large family Orchidaceae (Asparagales) and two small genera Pauridia (Hypoxidaceae : Asparagales) and Corsia (Corsiaceae, probably in Liliales), all epigynous taxa. Pauridia has actinomorphic (polysymmetric) flowers, whereas those of Corsia and most orchids are strongly zygomorphic (monosym- metric) with a well-differentiated labellum. In Corsia the labellum is formed from the outer median tepal (sepal), whereas in orchids it is formed from the inner median tepal (petal) and is developmentally adaxial (but positionally abaxial in orchids with resupinate flowers). Furthermore, in orchids zygomorphy is also expressed in the stamen whorls, in contrast to Corsia. In Pauridia a complete stamen whorl is suppressed, but the ‘ lost ’ outer whorl is fused to the style. The evolution of adnation and zygomorphy are discussed in the context of the existing phylogenetic framework in monocotyledons. An arguably typological classification of floral terata is presented, focusing on three contrasting modes each of peloria and pseudopeloria. Dynamic evolutionary transitions in floral morphology are assigned to recently revised concepts of heterotopy (including homeosis) and heterochrony, seeking patterns that delimit developmental constraints and allow inferences regarding underlying genetic controls. Current evidence suggests that lateral heterotopy is more frequent than acropetal heterotopy, and that full basipetal heterotopy does not occur. Pseudopeloria is more likely to generate a radically altered yet functional perianth, but is also more likely to cause acropetal modification of the gynostemium. These comparisons indicate that there are at least two key genes or sets of genes controlling adnation, adaxial stamen suppression and labellum development in lilioid monocots ; at least one is responsible for stamen adnation to the style (i.e. gynostemium formation), and another controls adaxial stamen suppression and adaxial labellum formation in orchids. Stamen adnation to the style may be a product of over-expression of the genes related to epigyny (i.e. a form of hyper-epigyny). If, as seems likely, stamen–style adnation preceded zygomorphy in orchid evolution, then the flowers of Pauridia may closely resemble those of the immediate ancestors of Orchidaceae, although existing molecular phylogenetic data indicate that a sister-group relationship is unlikely. The initial radiation in Orchidaceae can be attributed to the combination of hyper-epigyny, zygomorphy and resupination, but later radiations at lower taxonomic levels that generated the remarkable species richness of subfamilies Orchidoideae and Epidendroideae are more likely to reflect more subtle innovations that directly influence pollinator specificity, such as the development of stalked pollinaria and heavily marked and}or spur-bearing labella. Key words : adnation, atavism, Corsia, evolutionary–developmental genetics, gynostemium, heterochrony, heterotopy, homeosis, Hypoxidaceae, key innovations, labellum, Orchidaceae, Pauridia, peloria, pseudo- peloria, resupination, synorganisation, terata, zygomorphy.
Transcript of Roles of synorganisation, zygomorphy and heterotopy in floral evolution: the gynostemium and...

Biol. Rev. (2002), 77, pp. 403–441 " Cambridge Philosophical SocietyDOI: 10.1017}S1464793102005936 Printed in the United Kingdom
403
Roles of synorganisation, zygomorphy and
heterotopy in floral evolution: the gynostemium
and labellum of orchids and other lilioid
monocots
PAULA J. RUDALL" and RICHARD M. BATEMAN#
"Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AB, UK
#Department of Botany, Natural History Museum, Cromwell Road, London, SW7 5BD, UK
(Received 30 July 2001; revised 21 February 2002; accepted 7 March 2002)
ABSTRACT
A gynostemium, comprising stamen filaments adnate to a syncarpous style, occurs in only three groups ofmonocots : the large family Orchidaceae (Asparagales) and two small genera Pauridia (Hypoxidaceae:Asparagales) and Corsia (Corsiaceae, probably in Liliales), all epigynous taxa. Pauridia has actinomorphic(polysymmetric) flowers, whereas those of Corsia and most orchids are strongly zygomorphic (monosym-metric) with a well-differentiated labellum. In Corsia the labellum is formed from the outer median tepal(sepal), whereas in orchids it is formed from the inner median tepal (petal) and is developmentally adaxial(but positionally abaxial in orchids with resupinate flowers). Furthermore, in orchids zygomorphy is alsoexpressed in the stamen whorls, in contrast to Corsia. In Pauridia a complete stamen whorl is suppressed, butthe ‘ lost ’ outer whorl is fused to the style. The evolution of adnation and zygomorphy are discussed in thecontext of the existing phylogenetic framework in monocotyledons. An arguably typological classification offloral terata is presented, focusing on three contrasting modes each of peloria and pseudopeloria. Dynamicevolutionary transitions in floral morphology are assigned to recently revised concepts of heterotopy(including homeosis) and heterochrony, seeking patterns that delimit developmental constraints and allowinferences regarding underlying genetic controls. Current evidence suggests that lateral heterotopy is morefrequent than acropetal heterotopy, and that full basipetal heterotopy does not occur. Pseudopeloria is morelikely to generate a radically altered yet functional perianth, but is also more likely to cause acropetalmodification of the gynostemium. These comparisons indicate that there are at least two key genes or setsof genes controlling adnation, adaxial stamen suppression and labellum development in lilioid monocots ; atleast one is responsible for stamen adnation to the style (i.e. gynostemium formation), and another controlsadaxial stamen suppression and adaxial labellum formation in orchids. Stamen adnation to the style maybe a product of over-expression of the genes related to epigyny (i.e. a form of hyper-epigyny). If, as seemslikely, stamen–style adnation preceded zygomorphy in orchid evolution, then the flowers of Pauridia mayclosely resemble those of the immediate ancestors of Orchidaceae, although existing molecular phylogeneticdata indicate that a sister-group relationship is unlikely. The initial radiation in Orchidaceae can beattributed to the combination of hyper-epigyny, zygomorphy and resupination, but later radiations at lowertaxonomic levels that generated the remarkable species richness of subfamilies Orchidoideae andEpidendroideae are more likely to reflect more subtle innovations that directly influence pollinatorspecificity, such as the development of stalked pollinaria and heavily marked and}or spur-bearing labella.
Key words : adnation, atavism, Corsia, evolutionary–developmental genetics, gynostemium, heterochrony,heterotopy, homeosis, Hypoxidaceae, key innovations, labellum, Orchidaceae, Pauridia, peloria, pseudo-peloria, resupination, synorganisation, terata, zygomorphy.

404 Paula J. Rudall and Richard M. Bateman
CONTENTS
I. Introduction ............................................................................................................................ 404II. Phylogenetic framework .......................................................................................................... 405
(1) Major groups within Orchidaceae..................................................................................... 405(2) Sister groups of Orchidaceae............................................................................................. 406
III. Terminology ............................................................................................................................ 407(1) Floral diagrams and floral formulae.................................................................................. 407(2) Gynostemium, gynostegium and column .......................................................................... 407(3) Floral symmetry ................................................................................................................ 407(4) Categories of floral terata.................................................................................................. 409(5) Evolutionary modification and translocation of pre-existing floral organs ........................ 410
IV. Nature and phylogenetic distribution of gynostemia and labella in lilioid monocots ............... 410(1) Pauridia (Hypoxidaceae).................................................................................................... 410(2) Corsia (Corsiaceae) ............................................................................................................ 412(3) Orchidaceae ...................................................................................................................... 414
V. Stamen suppression and androecial zygomorphy in Orchidaceae ........................................... 417VI. Possible compound origin of the labellum ............................................................................... 418
VII. Occurrence and taxonomic treatment of peloria, pseudopeloria and non-resupination withinOrchidaceae............................................................................................................................. 420(1) Peloria and pseudopeloria ................................................................................................. 420(2) Non-resupination .............................................................................................................. 423
VIII. Distinguishing floral plesiomorphy from atavism..................................................................... 423IX. Reconstructing the ‘ancestral orchid’...................................................................................... 424X. Describing evolutionary transitions in floral morphology: heterochrony, heterotopy and
homeosis .................................................................................................................................. 427XI. Interpreting evolutionary transitions in floral morphology: evolutionary–developmental
genetics of adnation, zygomorphy and resupination ................................................................ 430(1) Morphogenetics in monocots............................................................................................. 430(2) Morphogenetics in orchids : indirect evidence ................................................................... 431(3) Morphogenetics in orchids : direct evidence ...................................................................... 431
XII. Key innovations and radiation(s) of Orchidaceae .................................................................. 432XIII. Future directions...................................................................................................................... 435
(1) Static and dynamic descriptions of flowers ........................................................................ 435(2) Orchid evolution ............................................................................................................... 435
XIV. Conclusions ............................................................................................................................. 436XV. Acknowledgements .................................................................................................................. 436
XVI. References ............................................................................................................................... 437
I. INTRODUCTION
Orchidaceae constitute the second-largest extantangiosperm family. They represent an unusuallycoherent group among lilioid monocots, possessingseveral reliable floral morphological synapomor-phies, including (1) the presence of a gynostemium,which is a compound structure comprising the styleand at least part of the androecium, (2) strong floralzygomorphy, reflecting (a) reduction to staminodesand}or complete suppression of adaxial stamens and(b) the presence of a labellum, usually accompaniedby (3) resupination, most commonly caused by 180°torsion of the pedicel and}or ovary. Here, we assessfloral evolution of orchids in comparison with other
lilioid monocots that possess gynostemia and}orlabella ; both features have highly restricted distribu-tions among monocots.
Gynostemia are rare in flowering plants generally,although they are characteristic of a few groups,such as the magnoliid family Aristolochiaceae (e.g.Leins & Erbar, 1985; Gonza! lez & Stevenson, 2000)and the eudicot family Stylidiaceae (e.g. Erickson,1958). Within monocots, a gynostemium is normallyregarded as a structure peculiar to the orchids (e.g.Swamy, 1948; Dressler, 1993). However, someadnation of filaments to the style also occurs in a fewlilioid monocots other than orchids, although thesestructures have not hitherto been recognised asgynostemia (see Section IV). A comparative review

405Synorganisation, zygomorphy and heterotopy in floral evolution
of gynostemium-like structures and associated synor-ganisation and zygomorphy is especially timely inthe context of recent phylogenetic studies.
Zygomorphy with respect to both perianth andstamens is relatively infrequent in monocots otherthan Zingiberales and Commelinales (both comme-linids) and Orchidaceae (Asparagales). Other zygo-morphic lilioid monocots include the questionablymonophyletic family Corsiaceae (assignable to Li-liales or Dioscoreales), some Burmanniaceae (Dio-scoreales), and some other Asparagales, such as someIridaceae (e.g. Diplarrhena) and Tecophilaeaceae,which often have dimorphic stamens. Orchidsrepresent arguably the strongest expression of zygo-morphy among monocots, since in orchids zygo-morphy affects all three floral whorls and typicallyincludes both labellum formation and adaxialstamen suppression.
The distinction between angiosperm flowers thatare radially symmetrical and those that are bi-laterally symmetrical has long attracted discussionin the botanical literature (cf. Darwin, 1862).Interest in this debate was rekindled in the 1990sby the escalating number of morphological andespecially molecular phylogenies, which togetherprovided empirical support for previously expressedopinions that radial symmetry was plesiomorphic inangiosperms, and that evolutionary transitions fromradial to bilateral symmetry were more commonthan vice versa (e.g. Endress, 2001a). Further interesthas been prompted by the recent exponential ex-pansion of plant evolutionary–developmental gene-tics (e.g. Coen, 1996; Diggle & Endress, 1999;Cronk, 2001; Cronk, Bateman & Hawkins, 2002),led by the insights into floral morphogenesis providedby the ‘ABC’ model of genetic control that under-pins the development of the four classic floral whorls :sepals, petals, stamens and carpels (e.g. Coen &Meyerowitz, 1991; Theissen et al., 2002).
Here we review the evolutionary significance ofthe monocot gynostemium and labellum, placing ina phylogenetic context new evidence from bothfloral anatomy and teratology of orchids and fromtwo relatively poorly known lilioid monocots, Pauri-dia (Hypoxidaceae) and Corsia (Corsiaceae), thatalso possess a gynostemium. Questions that areaddressed include: Which evolved first in the orchidlineage, zygomorphy or stamen–style adnation? Iszygomorphy in orchid flowers controlled by rela-tively simple parameters, and strictly comparable tothat of other monocots and eudicots? Is the orchidlabellum a compound structure comprising differentorgan elements, as is the case with the gynostemium?
Is the orchid gynostemium homologous with those ofCorsia and}or Pauridia? Evolutionary transitions infloral morphology are related to specific concepts ofsynorganisation, heterotopy (including homeosis)and heterochrony; hypotheses are then erectedregarding the underlying genetic controls.
II. PHYLOGENETIC FRAMEWORK
(1) Major groups within Orchidaceae
Given their poor fossil record (e.g. Mehl, 1986),assessment of the phylogenetic position of orchidsinevitably relies heavily on morphological andespecially molecular studies of extant species. WithinOrchidaceae, the bigeneric subfamily Apostasioideae(Apostasia and Neuwiedia) is traditionally regarded asthe sister group to other orchids (reviewed byDressler, 1993). Although lack of internal statisticalsupport still leaves some room for doubt about boththe relative placement and monophyly of Apostasioi-deae, all of the most recent molecular and mor-phological analyses support this basal position(Cameron et al., 1999; Freudenstein & Rasmussen,1999; Cameron & Chase, 2000; Freudenstein, Senyo& Chase, 2000) (see Figs 13, 14). Apostasioids aretherefore critical in evolutionary assessments oforchid structures, especially Neuwiedia, which hasthree fertile stamens and hence is often thought torepresent the closest extant approximation to the‘primitive ’ orchid flower (cf. Dressler, 1993; Chase,2001). In contrast, the identity of the next-branchingsubfamily is ambiguous. Earlier molecular phylo-genetic studies indicated that it was the diandrousCypripedioideae, although bootstrap trees for recentmultigene studies tended to show a polytomy withthe monandrous subfamily Vanilloideae: both poss-ible topologies have been obtained using rbcL alone(cf. Cameron et al., 1999; Pridgeon et al., 2001)and 18S rDNA (cf. Cameron & Chase, 2000;Molvray, Kores & Chase, 2000). More recently,combined analysis of sequence data from rbcL, atpB
and psaBhave tentatively indicated thatVanilloideaediverged just before Cypripedioideae (K. Cameron,personal communication, 2001). Both subfamiliesVanilloideae and Cypripedioideae are relativelypoor in numbers of genera and species. Most of thetaxonomic diversity in orchids is encompassed bytwo recently expanded sister-subfamilies : Orchidoi-deae (including spiranthoids) and especially Epiden-droideae (including neottioids and vandoids).

406 Paula J. Rudall and Richard M. Bateman
Fig. 1. Diagram of monocot relationships, based on results of equally-weighted analyses in Fay et al. (2000), Rudallet al. (2000), Chase et al. (2000) and Neyland (2002), collapsing nodes that received less than 60% bootstrap support.Pauridia (not shown) is placed in Hypoxidaceae.
(2) Sister groups of Orchidaceae
Recent molecular analyses have shown Orchidaceaeas clearly monophyletic and placed in the orderAsparagales, either as the sole sister taxon to allother Asparagales (in the combined multigeneanalysis of Fay et al., 2000) or as sister to the first-branching clade of Asparagales (in analyses of rbcL
alone: e.g. Chase et al., 1995; Rudall et al., 1997),although they are at best weakly supported in thesepositions. In either case, close relatives of Orchi-daceae within Asparagales include Hypoxidaceae, asmall but widely distributed tropical family ofapproximately 100 species distributed among ninegenera, including Pauridia (discussed in Section IV).Synapomorphies for Asparagales include epigyny,simultaneous microsporogenesis (Rudall et al., 1997)and phytomelan in the seed coat (Huber, 1969), butthere are some reversals in these characters, es-pecially in the higher asparagoids. Orchids areindeed epigynous and most have simultaneousmicrosporogenesis, but of the two basal clades withinOrchidaceae, microsporogenesis is as yet undescri-bed in apostasioids and successive in cypripedioids
(Guignard, 1883; for review see Rudall et al., 1997).All orchids lack phytomelan in the seed coat, andshare several other morphological similarities withother monocot orders in a confusing mosaic ofpotential synapomorphies. Most notably, Dahlgren,Clifford & Yeo (1985) placed the orchids in Liliales,based on several similarities (e.g. absence of septalnectaries, presence of perigonal nectaries and pat-terned tepals). A further notable homoplasy is theshared occurrence of silica bodies in both orchidsand commelinids, otherwise absent from monocots(e.g. Rudall & Chase, 1996).
At higher taxonomic levels, even recent combinedmultigene analyses (e.g. Chase et al., 2000) yieldedan unresolved polytomy between six monocot clades :the four lilioid orders (Asparagales, Liliales, Dioscor-eales and Pandanales), the large commelinid clade,and the small bigeneric family Petrosaviaceae (Fig.1). This major polytomy may represent relativelydeep, rapid radiation that included the origin of theorchids (cf. Bateman, 1999). If so, orchids occupy acritical position in monocot systematics, and anassessment of orchid characters assumes greater

407Synorganisation, zygomorphy and heterotopy in floral evolution
importance in achieving a broader understanding ofmonocot evolution.
III. TERMINOLOGY
(1) Floral diagrams and floral formulae
Throughout this paper, the standard convention offloral diagrams is followed (e.g. Eichler, 1875).Floral diagrams (such as those in Fig. 4) are a usefuldevice that is employed in several evolutionary–developmental studies (e.g. Meyerowitz, Smyth &Bowman, 1989; Coen & Meyerowitz, 1991). Furtherclarification could be achieved by including a floralformula, a long-standing but perhaps under-utilisedtool (e.g. Weberling, 1989, p. 19) in which K, C, Aand G represent calyx, corolla, androecium andgynoecium, respectively. Hence, Arabidopsis wouldbe K4 P4 A6 G(2), where parentheses indicatefusion, and the underscore indicates hypogyny. Ifthere are two distinct whorls of stamens, the outerandroecial whorl (in the same sector as the outertepals}sepals) is denoted in upper case (A1, A2, A3)and the inner androecial whorl (in the same sector asthe inner tepals}petals) in lower case (a1, a2, a3)(see Fig. 4).
(2) Gynostemium, gynostegium and column
A gynostemium (plural gynostemia) is a compoundstructure formed by adnation of stamens and style.This is in contrast to the ‘ shelter ’-like gynostegiumfound in several monocots and eudicots. For ex-ample, gynostegia occur in the eudicot familyApocynaceae (e.g. Kunze, 1995), specifically intaxa ascribed to the former family Asclepiadaceae,which is phylogenetically nested within Apo-cynaceae (Endress & Bruyns, 2000). They are alsopresent in certain lilioid monocots such as someepigynous Dioscoreales (e.g. Burmannia, Avetra, Tri-chopus), in which the stamens develop apical antherprocesses that fuse postgenitally with the stigmabefore anthesis (e.g. Caddick, Rudall & Wilkin,2000). In Echeandia (Anthericaceae: Asparagales)the anthers fuse postgenitally to form an anther conesurrounding the style (Bernhardt & Montalvo,1977); such postgenital fusion also occurs in Curculigo
(Hypoxidaceae), apostasioid orchids and some othermonocots (Endress, 1996; Kocyan & Endress,2001b). A gynostegium is a structure formed bypostgenital union of the anthers (or anther exten-
sions) and the stigma (e.g. Zomlefer, 1994), andtherefore appears to be non-homologous with thegynostemium, in which the filaments (and sometimesalso the anthers) always show (at least proximal)congenital fusion with the style. Although syncarpyis normally regarded as (more or less) congenital,most syncarpous monocots are apocarpous at in-itiation; therefore, it could be argued that thedistinction between the gynostemium and gynoste-gium is due to heterochrony, specifically reflectinglater (and incomplete) fusion of the two inner floralwhorls in gynostegia, but this hypothesis requiresfurther testing.
In orchids, the gynostemium is commonly termeda ‘column’ (the latter being defined as a structureformed by union of stamens, style and stigma: e.g.Zomlefer, 1994), though increasing numbers ofauthors reject the use of the term ‘column’ in favourof ‘gynostemium’. However, the two structures arenot strictly fully homologous, as the gynostemiumdoes not include the stigma in all orchids (Rasmus-sen, 1981, 1982). A gynostemium is apparent (if notalways conspicuous) in all orchids, even during earlyontogenetic stages (e.g. Kurzweil, 1998). The term‘column’ is also ambiguous in comparative terms: inother angiosperm families it is used to describe aconnate staminal structure in which the stamens arefused to each other to form a tube, but not adnate tothe gynoecium, as in many Iridaceae. Indeed, asBernhardt & Goldblatt (2000) commented, thestaminal column in some Iridaceae (e.g. Sisyrinchium)resembles a gynostemium, since it closely encirclesthe style. Some authors further subdivide the orchidcolumn into regions such as the ‘column foot ’ and‘column part ’, only the latter being viewed asequivalent to the gynostemium (for more detailedreviews of terminology see Rasmussen, 1982, 1985;Kurzweil, 1998; Szlacheto & Rutkowski, 2000).
(3) Floral symmetry
The terms actinomorphy and zygomorphy (andtheir respective partial synonyms, regular andirregular) are often used without precise definition inorganismal botany studies. Moreover, as noted byEndress (2001a), in evolutionary–developmentalstudies they have been partly replaced by inap-propriate use of the terms symmetry and asymmetry,respectively. Neal et al. (1998) and Endress (1999,2001a) employed the presence of two or more‘ symmetry planes ’ to define polysymmetry (actino-morphy: Fig. 2A), the presence of a single symmetry

408 Paula J. Rudall and Richard M. Bateman
Fig. 2. Diagrams of floral symmetry. (A) Actinomorphic(polysymmetric) flower, with three planes of symmetry(i). (B) Disymmetric flower with two-fold axis of symmetryand two mirror planes that are mutually perpendicularbut differ in breadth, one being wider than the other (iand ii). (C) Zygomorphic (monosymmetric) flower, withone plane of symmetry (i), modelled on a typical orchidflower. L, labellum. (D) Asymmetric flower, lacking anyplanes of symmetry.
plane to define monosymmetry (zygomorphy: Fig.2C) and the absence of any symmetry planes todefine true asymmetry (Fig. 2D). True asymmetryoccurs in species with spirally-arranged floral organs(i.e. haplomorphy, as in many magnoliids : Neal et
al., 1998). It is also found in some species derivedfrom zygomorphic forms, such as the epidendroidorchid Tipularia discolor, in which one lateral petalis consistently oriented across the median sepal(Luer, 1975). The colloquial term ‘irregular ’generally encompasses both zygomorphy andasymmetry.
However, even this revised set of definitions isinsufficiently precise. Application of crystallographicconcepts of symmetry (e.g. Berry, 1959; Klein,Hurlbut & Dwight, 1998) prompts further modifica-tion of these definitions, taking into account theexistence of different types of symmetry planes aswell as their total number and also considering thepresence of axes of symmetry. Full actinomorphy,
involving all four floral whorls, requires a flower thatpossesses at least three perianth segments of equalsize and shape. When viewed perpendicular to thesubtending pedicel, it must therefore possess at leastthree identical mirror planes that precisely bisect theflower, and the pedicel must constitute at least athree-fold axis of symmetry. In other words, anactinomorphic flower can be rotated through a setangle (120°, reflecting a three-fold axis of symmetry,in Fig. 2A) yet its appearance and spatial orientationwill appear unchanged. However, zygomorphyencompasses two different categories of overallsymmetry. In addition to the traditional zygo-morphic flower with a single mirror plane and noaxes of symmetry (Fig. 2C), there is also anarrangement of organs that involves a two-fold axisof symmetry and two mirror planes that are mutuallyperpendicular but bisect portions of the flower thatdiffer in aggregate breadth, one being ‘wider’ thanthe other (i and ii in Fig. 2B). This arrangement,termed disymmetry by Endress (1992, 1999), occursin many Brassicaceae (including Arabidopsis), someBegonia species, Dicentra and occasional teratologicalorchids where the petal whorl has been lost and thetwo lateral sepals have become postgenitally fused.Most zygomorphic perianths do indeed possess onlyone mirror plane, which is typically (though notobligatorily) oriented dorsiventrally (Fig. 2C).
Fig. 2C is modelled on a typical orchid flower. Inmost orchid species, the labellum (L) is differentlysized (either larger or smaller), more complex and}ormore strongly marked than the remaining perianthsegments, commonly acting as a visual attractantand especially as a landing stage for pollinatinginsects. In order to fulfil these vital functions, in mostwell-differentiated orchid flowers the labellum (themedian petal), developmentally the uppermost te-pal, is spatially lowermost (Ernst & Arditti, 1994;Kurzweil, 1998). This phenomenon, termed re-supination, is a normal floral posture in pendentinflorescences, but in the more common erectmultiflowered inflorescences it reflects 180° torsion ofthe subtending pedicel and}or ovary (some interest-ing exceptions are discussed in Section VII). In erectinflorescences with terminal flowers, such as in mostcypripedioids, resupination occurs by reorientationof the junction of the pedicel. Because of resupi-nation, the terms abaxial and adaxial, often used todescribe floral structures, are ambiguous in com-parative terms (although still more useful than thehighly ambivalent terms dorsal and ventral, fre-quently used by developmental geneticists). Medianstructures in zygomorphic flowers are better defined

409Synorganisation, zygomorphy and heterotopy in floral evolution
as those which are bisected by the mirror plane ofsymmetry (Bateman, 1985; R. Bateman, unpub-lished observations) ; for example, in a trimerousflower there is only one median sepal, one medianpetal and one median carpel (see Fig. 4).
(4) Categories of floral terata
Terata are of evolutionary importance because theyoffer insights into the genetic control of floralzygomorphy if their causal mechanisms can bedetermined. Bateman & DiMichele (1994) defined ateratos (plural terata) as an individual showing aprofound phenotypic change from its parent(s),irrespective of whether the underlying cause isgenetic or non-genetic (i.e. ecophenotypic). Theyfurther defined a teratological individual that de-monstrably reflects a genetic modification, and thuscan in theory establish its own phenotypicallydistinct evolutionary lineage, as a hopeful monster,echoing the saltational evolutionary terminology ofGoldschmidt (1940).
Terata of greatest interest in the study of zygo-morphy are those that involve complete or partialreversions to actinomorphy of the perianth. Suchteratological flowers were first termed peloric byLinne! (1744). A literature survey by Bateman(unpublished observations) in 1990 indicated thatthe many subsequent definitions of the term differmainly in two criteria : (1) With regard to thepolarity of the symmetry transition, some authorsallowed only zygomorphy to actinomorphy, whereasothers also allowed actinomorphy to zygomorphy.(2) Some authors have invoked a specific require-ment for atavism, here defined as the expression inan organism of characters shown by progenitorsmore genealogically remote than its parents. Thesepoints reflect explicit differences of opinion, butother difficulties are caused by ambiguity. Inparticular, descriptions rarely clarify which of thefour whorls confers bilateralism on the flower.
Here, we allow a broad definition of peloria(termed ‘pelory’ by Endress, 1994) that encompassesall transitions between zygomorphy and actinomor-phy. Bateman (1985) noted that in orchids, peloricflowers occur in two strongly contrasting modes (Fig.3, top). In type A peloria, the labellum (and oftenthe associated spur, when present in the ‘normal ’morph) is duplicated and replaces the two lateralpetals. In type B peloria, the labellum is replaced bya third undifferentiated petal lacking a spur.Horsman (1990; see also Lang, 2001) added a thirdcategory to Bateman’s (1985) classifications of orchid
Fig. 3. Diagrams of orchid perianth whorls (sepal andpetal) illustrating ‘normal ’ morphology (centre), threecategories (types A–C) each of peloria and pseudopeloria,and the type of morphogenetic transition that eachcategory represents.
floral terata, recognising as type C peloria a rarermorph in which all three petals are replaced bysepal-like structures to generate sepal and petalwhorls that are not only actinomorphic but alsoidentical (Fig. 3, top). Obviously, type B and type Cpeloria can only be distinguished when there isstrong morphological divergence between sepals andpetals in the normal morph. Bateman (1985) notedthat orchid terata involving only partial simplifica-tion of the labellum to a ‘sepaloid’ condition (Fig. 3,bottom) have often been termed ‘semi-peloric ’, buttrue peloria is an absolute condition; either theflower is fully actinomorphic or it is not. Pseudo-peloric is therefore a more appropriate term todescribe individuals showing diminished, but none-theless incontrovertible, zygomorphy of the perianth

410 Paula J. Rudall and Richard M. Bateman
whorl. In originally defining pseudopeloria, Bate-man (1985) focused on de-differentiation of thelabellum, which may be partial (here termed type Apseudopeloria) or complete (type B pseudopeloria).However, in rare cases the labellum remains ‘nor-mal ’ and it is the lateral petals that approach asepaloid condition, a phenomenon described here astype C pseudopeloria.
(5) Evolutionary modification andtranslocation of pre-existing floral organs
After extensively reviewing the literature on majorphenotypic transitions occurring during evolution,Bateman (1994, table 1) coined the generalised term‘evolutionary–developmental change’ to encompassany phenotypically expressed, genetically basedchange in a lineage that profoundly alters the shapeand}or size of any organ in any developmental stagein the putative descendant relative to those of itsputative ancestor. He simplified the much debateddefinitions of the more widely used subsidiary termsheterochrony and heterotopy (cf. Gould, 1977;Alberch et al., 1979; Fink, 1982; Sattler, 1988), bothof which focus on phenotypic modifications of pre-existing structures. Thus, in morphological terms,heterochrony is a temporal change in the expressionof a homologous feature between putative ancestorand putative descendant, and heterotopy is a spatial(positional) change in the expression of a hom-ologous feature between putative ancestor andputative descendant (some authors allow under‘heterotopy’ the transfer of only part of the ancestralfeature).
Although some definitions of heterotopy requirethat the position of expression of the feature in thedescendant should previously have been occupied byanother, non-homologous trait in the ancestor, Baumand Donoghue (2002) argued that this more pre-cisely defined category of evolutionary–developmen-tal change is better termed homeoheterotopy. Wherethe replacement of one feature by the other iscomplete rather than partial, we have not onlyhomeoheterotopy but also homeosis : ‘ the assump-tion by one part of an organism the likeness ofanother part ’ (Sattler, 1988, p. 1606; see alsoLeavitt, 1909). In some but by no means all examplesof homeosis, at least one specific function is trans-ferred from the old structure to the new structure(Corner, 1958), a concept further developed in anevolutionary–developmental genetic context byBaum & Donoghue (2002) and adapted by Bateman& DiMichele (2002) (see Section X).
IV. NATURE AND PHYLOGENETIC
DISTRIBUTION OF GYNOSTEMIA AND
LABELLA IN LILIOID MONOCOTS
Comparative observations on Pauridia are derivedfrom Thompson (1978), on Corsia from Ru$ bsamen(1986) and Rudall & Eastman (2002), and on othermonocots from both personal observations byP. J. R. and an extensive literature review. Im-portant previous works on floral anatomy of orchidsinclude Swamy’s (1948) review of orchid vascularanatomy and Kurzweil’s well-documented series ofpapers on ontogeny (e.g. Kurzweil, 1987a, b, 1988,1996, 1998), together with papers by Vermeulen(1966), Rao (1969, 1973, 1974), and Kocyan &Endress (2001a).
A ‘true’ gynostemium, with filaments adnate tothe style, occurs in only three groups of lilioidmonocots : (a) all Orchidaceae, (b) Pauridia (Hypoxi-daceae) and (c) Corsia (Corsiaceae). All three possessinferior ovaries and therefore share a further degreeof synorganisation. Due to obvious functional con-straints, a true gynostemium never occurs in hy-pogynous taxa, which at best show a small degree ofproximal adnation; for example, Vaikos & Markan-deya (1976) reported fusion of the filament baseswith the base of the ovary (but not with the style) inAloe, Haworthia and Gasteria (Asphodelaceae: Aspara-gales), and (incorrectly) regarded the resultingstructure as an ‘ incipient gynostemium’. Further-more, perhaps as a consequence of spatial constraintsresulting from fusion of style and stamens, taxa thatpossess gynostemia invariably lack septal nectaries(Rudall, 2002). In addition to a gynostemium, Corsia
and most Orchidaceae possess a labellum, which isan enlarged (outer or inner) tepal. However, alabellum is absent from flowers of Pauridia, andindeed from almost all other lilioid monocots,although arguable exceptions include some Iri-daceae with a slightly zygomorphic perianth, such asDiplarrhena.
(1) Pauridia (Hypoxidaceae)
Pauridia is a small genus of two species of minutecormous geophytes endemic to the Cape region ofSouth Africa (Rand, 1980). The microscopic, white,stellate flowers are epigynous and actinomorphic;they are subtended by three bracts and bear onlythree fertile stamens. The three petals are fused intoa long, narrow perianth tube, to which the threestamen filaments are adnate for most of their length.The style possesses three characteristic appendages

411Synorganisation, zygomorphy and heterotopy in floral evolution
A B
C D
E F
L
L L
A1
a2 a1
A1
a2 a1
Fig. 4. Floral diagrams. (A) Hypoxis (Hypoxidaceae). (B) Pauridia (Hypoxidaceae). (C) Corsia (Corsiaceae). (D)Neuwiedia (Orchidaceae: Apostasioideae). (E) Cypripedium (Orchidaceae: Cypripedioideae). (F) Epipactis (Orchidac-eae: Epidendroideae). Perianth whorls : white¯ sepals, black¯petals. L¯ labellum, A¯ outer stamen whorl, a¯inner stamen whorl.
(Thompson, 1978, 1979). Pauridia was originallyplaced in Iridaceae because it has only three stamens(in contrast to all other Hypoxidaceae which havesix, as in the majority of monocots), but after beingvariously assigned to Amaryllidaceae and Haemo-doraceae it was finally transferred back to Hypoxi-daceae, where it had earlier been placed, due to itsclose similarity with another genus of Hypoxidaceae,Spiloxene (Thompson, 1979). In fact, the free fertilestamens of Pauridia represent the inner androecialwhorl (a1, a2, a3), in contrast to those of Iridaceae,
which represent the outer whorl (A1, A2, A3) (Fig.4B). Thompson (1978) convincingly postulated thatthe unique stylar appendages of Pauridia are stamino-dial in origin (i.e. they represent the ‘ lost ’ outerstamen whorl : A1, A2, A3). She cited as evidencethe fact that they have a vascular supply similar tothat of the outer stamens of the related genusSpiloxene, and also that occasional teratologicalflowers bear anthers on the stylar appendages.
Thompson’s hypothesis is confirmed by materialof Pauridia longituba examined here (Fig. 5). The style
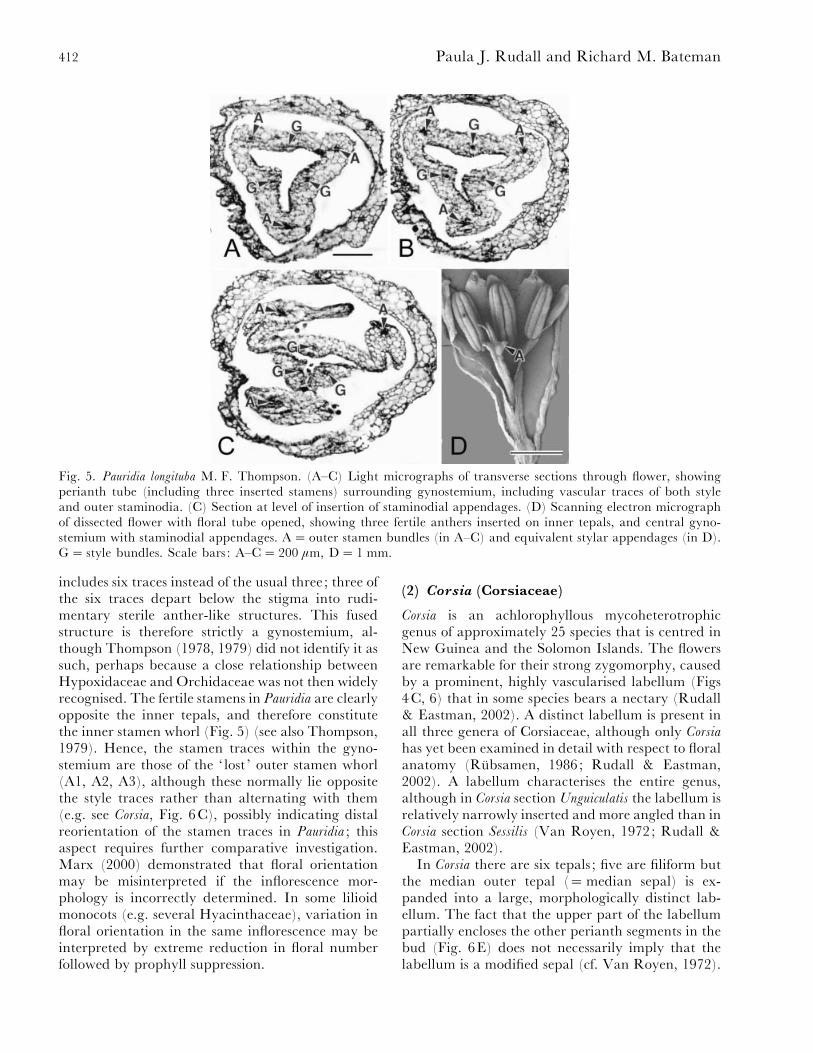
412 Paula J. Rudall and Richard M. Bateman
Fig. 5. Pauridia longituba M. F. Thompson. (A–C) Light micrographs of transverse sections through flower, showingperianth tube (including three inserted stamens) surrounding gynostemium, including vascular traces of both styleand outer staminodia. (C) Section at level of insertion of staminodial appendages. (D) Scanning electron micrographof dissected flower with floral tube opened, showing three fertile anthers inserted on inner tepals, and central gyno-stemium with staminodial appendages. A¯ outer stamen bundles (in A–C) and equivalent stylar appendages (in D).G¯ style bundles. Scale bars : A–C¯ 200 µm, D¯ 1 mm.
includes six traces instead of the usual three; three ofthe six traces depart below the stigma into rudi-mentary sterile anther-like structures. This fusedstructure is therefore strictly a gynostemium, al-though Thompson (1978, 1979) did not identify it assuch, perhaps because a close relationship betweenHypoxidaceae and Orchidaceae was not then widelyrecognised. The fertile stamens in Pauridia are clearlyopposite the inner tepals, and therefore constitutethe inner stamen whorl (Fig. 5) (see also Thompson,1979). Hence, the stamen traces within the gyno-stemium are those of the ‘ lost ’ outer stamen whorl(A1, A2, A3), although these normally lie oppositethe style traces rather than alternating with them(e.g. see Corsia, Fig. 6C), possibly indicating distalreorientation of the stamen traces in Pauridia ; thisaspect requires further comparative investigation.Marx (2000) demonstrated that floral orientationmay be misinterpreted if the inflorescence mor-phology is incorrectly determined. In some lilioidmonocots (e.g. several Hyacinthaceae), variation infloral orientation in the same inflorescence may beinterpreted by extreme reduction in floral numberfollowed by prophyll suppression.
(2) Corsia (Corsiaceae)
Corsia is an achlorophyllous mycoheterotrophicgenus of approximately 25 species that is centred inNew Guinea and the Solomon Islands. The flowersare remarkable for their strong zygomorphy, causedby a prominent, highly vascularised labellum (Figs4C, 6) that in some species bears a nectary (Rudall& Eastman, 2002). A distinct labellum is present inall three genera of Corsiaceae, although only Corsia
has yet been examined in detail with respect to floralanatomy (Ru$ bsamen, 1986; Rudall & Eastman,2002). A labellum characterises the entire genus,although in Corsia section Unguiculatis the labellum isrelatively narrowly inserted and more angled than inCorsia section Sessilis (Van Royen, 1972; Rudall &Eastman, 2002).
In Corsia there are six tepals ; five are filiform butthe median outer tepal (¯median sepal) is ex-panded into a large, morphologically distinct lab-ellum. The fact that the upper part of the labellumpartially encloses the other perianth segments in thebud (Fig. 6E) does not necessarily imply that thelabellum is a modified sepal (cf. Van Royen, 1972).

413Synorganisation, zygomorphy and heterotopy in floral evolution
Fig. 6. Corsia Becc. (A–C) C. merimantensis van Royen, light micrographs of proximal to distal serial transverse sectionsthrough a single flower. (A) Near top of ovary. (B) Point of departure of five tepals. (C) Proximal to insertion ofgynostemium on labellum. (D) Corsia ornata Becc., flower with gynostemium surrounded by labellum; three anthersfallen. (E) C. merimantensis, scanning electron micrograph of unopened bud showing labellum enclosing other tepals,and forming a ridge along one side of ovary. (F) C. merimantensis, junction of style and reflexed stamens. Gy¯ gyno-stemium, Lab¯ labellum. Vascular bundles : A¯ outer stamen bundle, a¯ inner stamen bundle, AG¯ fused(composite) stamen}style bundle, G¯ style bundle, T¯ outer tepal}sepal bundle, TAG¯ fused (composite) outerorgan bundle (sepal, stamen and style), ta¯ composite inner organ bundles (petal and stamen). Scale bars : A, B¯200 µm (also applies to C), D–F¯ 1 mm.
Indeed, this is a critical point, as in orchid flowers,which are mostly resupinate, the labellum is formedfrom the inner median tepal (¯median petal).However, despite the lack of material suitable for
developmental studies, examination of floral vascula-ture (Fig. 6A–C) indicates that in Corsia the labellumis indeed the outer median tepal (sepal). The centralvascular bundle of the labellum (T) and the vascular
414 Paula J. Rudall and Richard M. Bateman
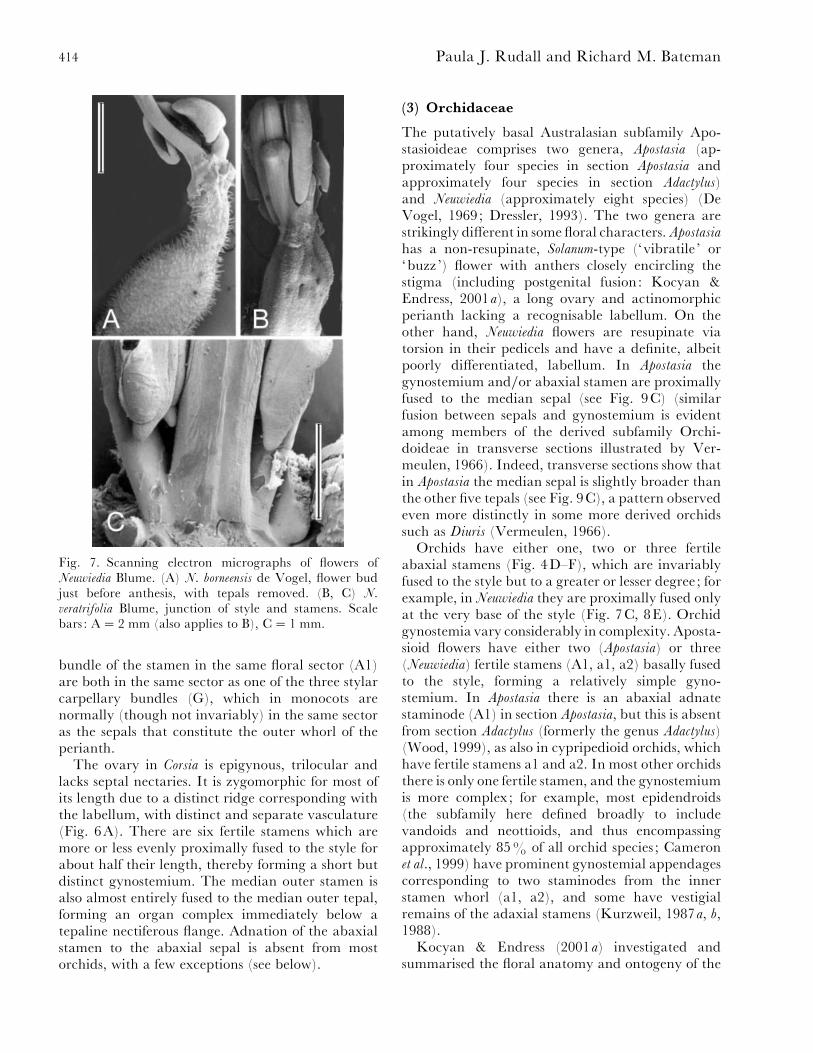
Fig. 7. Scanning electron micrographs of flowers ofNeuwiedia Blume. (A) N. borneensis de Vogel, flower budjust before anthesis, with tepals removed. (B, C) N.veratrifolia Blume, junction of style and stamens. Scalebars : A¯ 2 mm (also applies to B), C¯ 1 mm.
bundle of the stamen in the same floral sector (A1)are both in the same sector as one of the three stylarcarpellary bundles (G), which in monocots arenormally (though not invariably) in the same sectoras the sepals that constitute the outer whorl of theperianth.
The ovary in Corsia is epigynous, trilocular andlacks septal nectaries. It is zygomorphic for most ofits length due to a distinct ridge corresponding withthe labellum, with distinct and separate vasculature(Fig. 6A). There are six fertile stamens which aremore or less evenly proximally fused to the style forabout half their length, thereby forming a short butdistinct gynostemium. The median outer stamen isalso almost entirely fused to the median outer tepal,forming an organ complex immediately below atepaline nectiferous flange. Adnation of the abaxialstamen to the abaxial sepal is absent from mostorchids, with a few exceptions (see below).
(3) Orchidaceae
The putatively basal Australasian subfamily Apo-stasioideae comprises two genera, Apostasia (ap-proximately four species in section Apostasia andapproximately four species in section Adactylus)and Neuwiedia (approximately eight species) (DeVogel, 1969; Dressler, 1993). The two genera arestrikingly different in some floral characters. Apostasia
has a non-resupinate, Solanum-type (‘vibratile ’ or‘buzz’) flower with anthers closely encircling thestigma (including postgenital fusion: Kocyan &Endress, 2001a), a long ovary and actinomorphicperianth lacking a recognisable labellum. On theother hand, Neuwiedia flowers are resupinate viatorsion in their pedicels and have a definite, albeitpoorly differentiated, labellum. In Apostasia thegynostemium and}or abaxial stamen are proximallyfused to the median sepal (see Fig. 9C) (similarfusion between sepals and gynostemium is evidentamong members of the derived subfamily Orchi-doideae in transverse sections illustrated by Ver-meulen, 1966). Indeed, transverse sections show thatin Apostasia the median sepal is slightly broader thanthe other five tepals (see Fig. 9C), a pattern observedeven more distinctly in some more derived orchidssuch as Diuris (Vermeulen, 1966).
Orchids have either one, two or three fertileabaxial stamens (Fig. 4D–F), which are invariablyfused to the style but to a greater or lesser degree; forexample, in Neuwiedia they are proximally fused onlyat the very base of the style (Fig. 7C, 8E). Orchidgynostemia vary considerably in complexity. Aposta-sioid flowers have either two (Apostasia) or three(Neuwiedia) fertile stamens (A1, a1, a2) basally fusedto the style, forming a relatively simple gyno-stemium. In Apostasia there is an abaxial adnatestaminode (A1) in section Apostasia, but this is absentfrom section Adactylus (formerly the genus Adactylus)(Wood, 1999), as also in cypripedioid orchids, whichhave fertile stamens a1 and a2. In most other orchidsthere is only one fertile stamen, and the gynostemiumis more complex; for example, most epidendroids(the subfamily here defined broadly to includevandoids and neottioids, and thus encompassingapproximately 85% of all orchid species ; Cameronet al., 1999) have prominent gynostemial appendagescorresponding to two staminodes from the innerstamen whorl (a1, a2), and some have vestigialremains of the adaxial stamens (Kurzweil, 1987a, b,1988).
Kocyan & Endress (2001a) investigated andsummarised the floral anatomy and ontogeny of the

415Synorganisation, zygomorphy and heterotopy in floral evolution
Fig. 8. Neuwiedia veratrifolia, light micrographs of serial (proximal to distal) transverse sections through a single flower.(A) Trilocular ovary, median section. (B–D) Top of ovary, below point of insertion of floral parts. (E) Point oflabellum insertion. (F) Distal to tepal insertion, showing gynostemium. Lab¯ labellum, Gy¯ gynostemium. Vascularbundles : A¯ outer stamen bundles, a¯ inner stamen bundles, G¯ style bundles, T¯ outer tepal}sepal bundles, t¯inner tepal}petal bundles, TAG¯ composite outer organ bundles (sepal, stamen and style), ta¯ composite innerorgan bundles (inner tepal}petal and stamen). Scale bar (applies to all parts)¯ 200 µm.
416 Paula J. Rudall and Richard M. Bateman
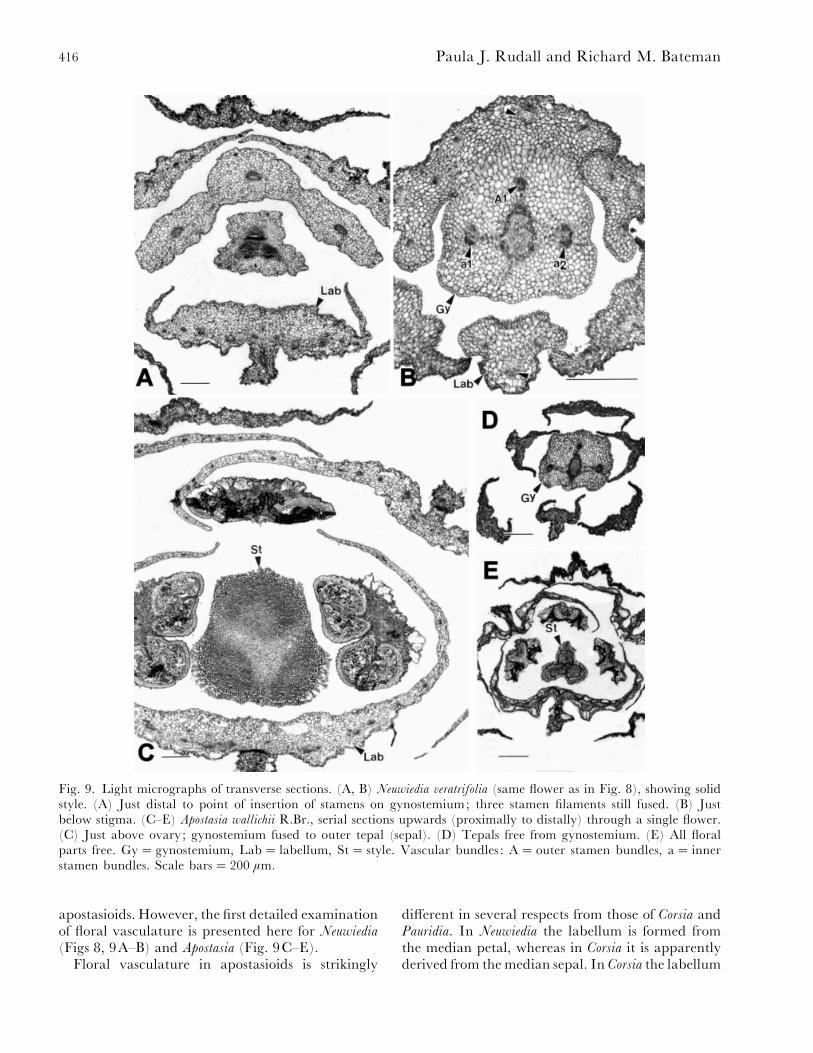
Fig. 9. Light micrographs of transverse sections. (A, B) Neuwiedia veratrifolia (same flower as in Fig. 8), showing solidstyle. (A) Just distal to point of insertion of stamens on gynostemium; three stamen filaments still fused. (B) Justbelow stigma. (C–E) Apostasia wallichii R.Br., serial sections upwards (proximally to distally) through a single flower.(C) Just above ovary; gynostemium fused to outer tepal (sepal). (D) Tepals free from gynostemium. (E) All floralparts free. Gy¯ gynostemium, Lab¯ labellum, St¯ style. Vascular bundles : A¯ outer stamen bundles, a¯ innerstamen bundles. Scale bars¯ 200 µm.
apostasioids. However, the first detailed examinationof floral vasculature is presented here for Neuwiedia
(Figs 8, 9A–B) and Apostasia (Fig. 9C–E).Floral vasculature in apostasioids is strikingly
different in several respects from those of Corsia andPauridia. In Neuwiedia the labellum is formed fromthe median petal, whereas in Corsia it is apparentlyderived from the median sepal. In Corsia the labellum

417Synorganisation, zygomorphy and heterotopy in floral evolution
is served only by sepal traces ; however, in Apostasia
and Neuwiedia (Figs 8, 9) the median trace of thelabellum is the median petal trace (t), but themarginal traces of the labellum (and the other twopetals) are always clearly supplied by the marginalveins of the adjacent sepals (T), as in many otherorchids (both diandrous and monandrous : Swamy,1948), and also in other Liliales and Asparagales (P.Rudall, unpublished observations). In Neuwiedia thecentral vascular bundle of the labellum (t) lacks acorresponding stamen bundle and alternates withtwo of the three stylar carpellary bundles (G); A1 issituated in the same floral sector as the sepal trace(T) rather than the median labellum trace (t) (Fig.9). Also, unlike most orchids, the flower of Corsia isnot resupinate, though the orientation of the sepalinelabellum (uppermost) is similar to that of thepetaline labellum of the minority of orchids wherethe flower is either non-resupinate or, less commonly,doubly resupinate (see Section VII). As the flower inCorsiaceae is solitary and terminal, the labellum canreadily be oriented in this manner.
V. STAMEN SUPPRESSION AND ANDROECIAL
ZYGOMORPHY IN ORCHIDACEAE
Much has been written about the systematic signifi-cance of adaxial stamen suppression in orchids.Several authors (e.g. Swamy, 1948; Vermeulen,1966; Chen, 1982; Burns-Balogh & Bernhardt,1985; Neyland & Urbatsch, 1996; Kurzweil, 1998;Freudenstein & Rasmussen, 1999) have noted thatOrchidaceae are defined by the loss or suppression of(developmentally) adaxial stamens from both outer(A) and inner (a) androecial whorls (i.e. A2, A3,a3), although Worsdell (1916) reported some tera-tological flowers where A2 or A3 and sometimeseven a3 were represented by staminodes or vasculartraces, Kocyan & Endress (2001a) found stamenprimordia on the adaxial side of the flower in someaspostasioids, and Kurzweil (1998) reported thatadaxial staminodes occur occasionally as teratologiesin most orchid subfamilies. Kurzweil (1987a, 1998)showed that in the epidendroid orchid Bletia thepresence of a ‘column foot ’ is developmentallylinked with the presence of adaxial staminodes.
In the apostasioid orchid Neuwiedia the functionalstamens are A1, a1 and a2 (Fig. 4D). Swamy (1948)regarded the triandrous Neuwiedia flower as repre-
senting the primitive orchid type, and postulatedthat a3 disappeared early in evolution, in correlationwith the differentiation of the labellum, followed bythe loss of A2 and A3. In the diandrous cypripe-dioids, a1 and a2 are normally present and func-tional, and A1 is a staminode (Fig. 4E). Inmonandrous orchids, encompassing 99% of allorchid species, the single functional stamen is A1(Fig. 4F). Freudenstein & Rasmussen (1999) codedandroecial characters with respect to individualstamens or groups of stamens, and demonstratedthat a reversal from diandry to triandry in theapostasioids, as suggested by some authors, isunlikely, since the apostasioids are the first-branch-ing clade in most orchid analyses (see Figs 13, 14).The most parsimonious optimisation of the stamencharacters across the orchid tree would involve twoseparate transitions from triandry to diandry, inApostasia and the cypripedioids (Freudenstein &Rasmussen, 1999), despite observations by Swamy(1948) and Rao (1969, 1973, 1974) that the floralvasculature of Apostasia closely resembles that of thecypripedioids. Freudenstein & Rasmussen (1999)based their polarisation of this character on thepresence of the dorsal median stamen (A1) in theirsingle chosen outgroup, Hypoxis (Fig. 4A). However,since Hypoxidaceae include another equally crediblesister group in Pauridia, which lacks the outer stamenwhorl that includes the dorsal median stamen (Fig.4B), this optimisation may require reconsideration(see later discussion: Section IX).
Within the monandrous orchids, in which A1 isthe functional stamen, there is a clear distinction invasculature between the Orchidoideae and Vanilloi-deae plus Epidendroideae s.l. In orchidoids only A1is present, and there are no staminodes or stamentraces. However, other monandrous orchids are farmore variable in this respect (Swamy, 1948). A1 isstill normally the functional stamen, but otherstamen traces are often present : a1 and a2, or A2and A3, or sometimes compound stamen tracesa1A2 and a2A3. Rarely, the a3 trace is alsoevident. Within the epidendroid genus Eulophia
(approximately 200 species), species differ in thereported origin of stamen traces (e.g. A1, A2, A3 inE. nuda but A1, a2, a3 in E. epidendraea). In therelated genus Geodorum there is infraspecific variationin G. densiflorum ; most commonly A1, a1A2 anda2A3 are evident, but unusual forms have onlyA1, A2 and A3 (i.e. the entire inner whorlsuppressed, as in Iridaceae), or A1, a2 and A2 (i.e.irregular), or even a1A2, a3 and A3 (i.e. only theadaxial stamens expressed) (Swamy, 1948).

418 Paula J. Rudall and Richard M. Bateman
VI. POSSIBLE COMPOUND ORIGIN OF THE
LABELLUM
The hypothesis of Brown (1833) and Darwin (1862)that the orchid labellum represents a compoundstructure that evolved by incorporation of stamensinto an ordinary petal was formerly widely held,based on evidence from both floral vasculature andterata. More recently, Nelson (1965, 1967) espouseda similar premise, emphasising the often trilobedstructure of the labellum. In normal flowers of manyorchid species the vascular strands of the non-expressed stamens A2 and A3 apparently pass intothe labellum, and labellum vasculature is otherwisecomplex, especially in epidendroids (Swamy, 1948).Furthermore, there are recorded cases of terata inwhich the labellum is suppressed or takes the form ofa lateral petal, and stamens A2 and A3 (bothsuppressed in normal flowers) are present, inaddition to A1. Conversely, there are cases in whichthe two lateral petals have been replaced byadditional labella and stamens a1 and a2 are absent(for summary see Worsdell, 1916), indicating adevelopmental link between labellum and stamens.Worsdell (1916) cited Magnus’ (1910) description ofan exceptional individual of Anacamptis (formerlyOrchis) papilionacea in which an anther occurred on‘either basal edge’ of the labellum.
However, Worsdell (1916, p. 143) emphatically(and influentially) asserted that ‘no special hy-pothesis ’ was required to explain the origin of theorchid labellum. He cited various teratological formsas evidence against the compound structure theory,making the rather draconian statement that ‘ theposition of vascular strands cannot be allowed to bea factor in solving morphological problems. ’ Teratareported by Worsdell (1916) which supposedlyindicated that stamens are not required for labellumformation included (1) a putative Neottia species inwhich the perianth was fully differentiated, yetstamens A2 and A3 were present, (2) similarly,flowers of Phaius and Odontoglossum in which A2and A3 were transformed into labella, and (3) aCypripedium possessing two fertile stamens A1 and a3,while the other four stamens were all replaced bylabella. Despite providing considerable relevant andpotentially contradictory data from floral vascula-ture, Swamy (1948) accepted Worsdell’s (1916)rejection of the hypothetical compound nature of theorchid labellum, and this view was further supportedby Kurzweil (1987a, b) and Kocyan & Endress(2001a), whose ontogenetic studies showed that thelabellum is initiated as a single primordium. Indeed,
the principle of vascular conservatism, which holdsthat vestigial (perhaps functionless) bundles maypersist when the organ that it formerly supplied hasdisappeared during evolution, was vigorously at-tacked by several authors, notably Carlquist (1969).However, this issue was subsequently put into amore balanced perspective by Wilson (1982), whodemonstrated that there is evidence that suchvascular ‘ stubs ’ do exist in some cases. Wilson(1982) endorsed Pennell’s (1948: p. 306) statementthat ‘The flower is both adapted to its presentmethod of functioning and may bear structuralevidence of its past. ’
We agree with Worsdell (1916) that the signifi-cance of these teratological and anatomical data isdifficult to assess in terms of orchid floral evolution,in part because floral vasculature in monocots is acomplex subject urgently requiring a fresh overview.As Endress (1994) noted, there is no simple answer tothe question of the origin(s) of the orchid labellum,which may vary among different orchid groups, as insome other monocots (for example, within Zingi-berales, the labellum corresponds to two unitedstaminodes in Zingiberaceae but five united stami-nodes in the putative sister family Costaceae:Endress, 1994; Kocyan & Endress, 2001a). Both thevascular traces and the organs they supply may becompound or fused. Orchid labella may credibly beregarded as compound structures in terms ofvasculature, because of the incorporation of the‘missing’ stamen traces into the labellum (at least insome cases). Furthermore, the marginal traces of thelabellum (and also those of the two lateral petals) arederived from the neighbouring sepal traces. Petal–sepal vasculature overlap occurs in many orchidtaxa, including the basally divergent apostasioids(Fig. 8). Such overlap was also well-documented bySwamy (1948) in both diandrous cypripedioids andmonandrous orchids, where the margins of thelabellum are supplied by the marginal veins of theadjacent sepals. Swamy (1948) further noted thatthe vasculature of the spur is normally derived fromthat of the inner tepals, but in a few orchidoid generawith two spurs per flower, such as Satyrium, the spurvasculature is derived from the marginal labellumtraces, which originate from the adjacent sepals.Furthermore, Kurzweil (1998) observed that in theepidendroid Bletia the median sepal and two lateralpetals originate from a common primordium.
However, although the plesiomorphic condition isbetween one and three traces per tepal, some overlapof petal–sepal vasculature also occurs in many othermonocots, and is also a common phenomenon in

419Synorganisation, zygomorphy and heterotopy in floral evolution
Fig. 10. (A, B) Ophrys insectifera L. (A) Typical flower well-adapted to pollination via pseudocopulation by malesolitary wasps. (B) Flower showing type A peloria; the ‘antennae’ have been replaced by additional ‘bodies. ’ (C, D)Platanthera chloranthera (Custer) Reichb. (C) Typical flower well-adapted to pollination via moths attracted by the longnectar-filled spur. (D) Type A pseudopeloric flower with sepal-like labellum and vestigial spur (photograph by R.Bush). Scales : A, C, D, ¬3±5; B, ¬3±2.
eudicots (P. Endress, personal communication,2001), so may be insignificant in terms of organidentity. Such complexity of vasculature often occursin the commelinid clade (e.g. Hardy & Stevenson,2000), and also in some lilioids, especially in
epigynous taxa. For example, in Hypoxis (Aspara-gales) and Bomarea (Liliales) the marginal outertepal (sepal) bundles supply the margins of the innertepals. In contrast, in the phylogenetically am-biguous genus Corsia petal and sepal vasculature

420 Paula J. Rudall and Richard M. Bateman
remain entirely separate, including that of the largehighly vascularised (sepaline) labellum (Fig. 6). Inhypogynous lilioids such as Burchardia (and also inbasal monocots), petal and sepal vasculature nor-mally remain wholly separate, though Anderson(1941) reported that inAloe (Asphodelaceae: Aspara-gales) the sepal traces supply the petal margins.Perhaps Nelson’s (1965, 1967) emphasis on thetrilobate structure of the labellum, which he cited asevidence for a partially staminodial origin, may beexplained by such vascular overlap. A putatively‘compound’ petal–sepal nature is also evidenced byteratological forms of Ophrys ferrum-equinum (see Fig.11D) and Ophrys mammosa (A. J. Hughes, personalcommunication, 2001) in which one-third of thelabellum is sepaline in appearance. However, con-verse terata exist in which labellum expression hasspread into the lower portions of the lateral sepals(see Fig. 12A), indicating a dorsalisation effect.Nonetheless, we know of no examples of terata inwhich one or more sepals have been wholly replacedby labella, in contrast to the many recorded terata inwhich the other two petals form labella (e.g. Fig.10B). There are also a few reported cases in whichstamens A2 and A3 were transformed into labella(e.g. Phaius and Odontoglossum : Worsdell, 1916), butno cases in which labella are wholly transformed intostamens, as occurs in some non-orchid taxa withzygomorphic flowers, such as the heptandra mutant ofDigitalis (Meyerowitz et al., 1989).
VII. OCCURRENCE AND TAXONOMIC
TREATMENT OF PELORIA, PSEUDOPELORIA
AND NON-RESUPINATION WITHIN
ORCHIDACEAE
(1) Peloria and pseudopeloria
Information on peloria and pseudopeloria in orchidswas taken from a literature review (notably Chen,1982; Bateman, 1985; Chen & Tsi, 1987; R. M.Bateman & L. McCook, unpublished observations)and from observations made or collated by R. M. B.during extensive European fieldwork over a periodof 25 years. The wealth of observations made by theorchidological community has demonstrated thatmany peloric and pseudopeloric morphs occursporadically as terata in natural populations ofotherwise typical orchids. These complete or partialreversions to actinomorphy of the perianth (Fig. 3)are of obvious interest in the study of zygomorphy.
Although most such terata pass unreported, atleast 12 out of the approximately 50 orchid speciesnative to the British Isles have demonstrated type Apeloria : we can now add Ophrys sphegodes andDactylorhiza praetermissa (G. Goodfellow, personalcommunication, 2001) to those listed by Bateman(1985). These type A morphs are distributed amongthe two largest orchid subfamilies recognised in themolecular phylogenetic analysis of Cameron et al.(1999): (a) higher orchidoids and (b) both lowerand higher epidendroids. In contrast, type B peloriais rarely recorded as infraspecific terata (notableexceptions are recent records for Ophrys sphegodes : G.Goodfellow, personal communication, 2001) andtype C peloria has rarely been recorded at all.Horsman (1990) coined the term for a six-sepalledflower of Dactylorhiza cf. incarnata depicted byRichard (1823), but the only examples known to usare single occurrences in Ophrys apifera (Lang, 2001,photo 7) and the Continental O. bremifera (Delforge,2001, p. 26).
Instances of pseudopeloria have similarly beendocumented in several Orchidoideae, includingOphrys apifera (Lang, 2001, photo 8, erroneouslyattributed to type B peloria), O. insectifera (A. J.Hughes, personal communication, 2001), Gymnadenia
conopsea (e.g. Lang, 1980), Dactylorhiza fuchsii (Bate-man, 1985; S. J. Longrigg, personal communication,2001), D. maculata (Horsman, 1990) and Anacamptis
pyramidalis, together with the epidendroid Epipactis
phyllanthes var. phyllanthes (Young, 1952; Bateman,1985). Detailed comparison of these phylogeneticallydisparate occurrences reveals considerable hetero-geneity of floral morphology (Fig. 3, bottom),suggesting that pseudopeloria may be an aggregateterm for a spectrum of subtly different phenotypicmodifications (see Sections X and XI; see alsoBateman & DiMichele, 2002).
In uncommon cases where peloric and pseudo-peloric terata naturally occurring in the UK havebeen monitored, many recur in successive years andoccasionally reproduce by seed, indicating that theyare mutants and therefore bona fide hopeful mon-sters – in other words, they are prospecies sensu
Bateman & DiMichele (1994, 2002), theoreticallycapable of establishing lineages that are sufficientlylong-lived to be legitimately recognised as taxonomicspecies.
Not surprisingly, taxonomic treatment of thesehopeful monsters has been highly inconsistent. Inmost cases, including probably all examples of typeA peloria (Figs 3, 10A, B), each mutant has beenrecognised as a forma or variety within a readily

421Synorganisation, zygomorphy and heterotopy in floral evolution
recognised (and far more abundant) ‘parent ’ spe-cies. Types B and C peloria and types A and Bpseudopeloria present more serious challenges toidentification, as the reduction or loss of their labellaand spurs can eliminate most of the morphologicalcharacters diagnostic of the ‘parental ’ species.Consequently, type A pseudopeloric morphs foundin Scotland of the orchidoids Platanthera chlorantha
(Fig. 10C, D) (McKean, 1982) and Dactylorhiza
maculata (D. K. Mardon & C. A. Stace, personalcommunication, 1996; G. Goodfellow, personalcommunication, 2001) were erroneously identifiedas bigeneric hybrids (cf. Bateman, 1985, 2001) aswere pseudopeloric morphs of Gymnadenia species inthe Alps (Schmid, 2001). Nonetheless, type C peloriaand type B pseudopeloria have been readily identi-fied even in the taxonomically challenging genusOphrys (cf. Delforge, 2001; Lang, 2001). Most suchmorphs occur within larger populations of ‘normal ’morphs. The most common exception in Britain isEpipactis phyllanthes var. phyllanthes, which can estab-lish uniform populations of type A pseudopeloricmorphs, presumably due to the characteristic auto-gamy of the species which allows rapid proliferationof genetically identical individuals (Young, 1952;Bateman, 1985).
On the other hand, when we broaden oursurvey to non-British orchids, it becomes clear thatthe mutant morph occasionally reaches a level ofphenotypic consistency and numerical abundancesufficient to become recognised as a full taxonomicspecies. In some cases the mutant species is assignedto a pre-existing genus, a good example being thetype B peloric cypripedioid Phragmipedium lindenii
from the neotropics (Nelson, 1965; Garay, 1979;Chen & Tsi, 1987; McCook, 1989; Pridgeon et al.,1999). However, genetic fixation of the mutation isnot quite perfect, as at least one population includesindividuals that bear ‘normal ’ pouch-like labella(Dressler & Williams, 1975). Moreover, the peloricmorph characteristically expresses a third stamen(A1) (Atwood, 1984; Freudenstein & Rasmussen,1999). Among Australian diurid orchidoids the‘ sporadic and rare’ Calochilus imberbis has a sepaloidlabellum and is probably a type B pseudopeloricmorph of the far more widespread and moreornamental C. robertsonii (Jones, 1988; see Fig. 12E).Stabilised terata among south-east Asian higherepidendroids have similarly been awarded speciesstatus. Coelogyne leungiana (Barretto, 1990) co-existswith, and is probably a type A pseudopeloric formof, the more widespread C. fimbriata (G. Barretto,pers. comm, 1990; Gravendeel, 2000, p. 150), and in
Tibet Eria medogensis appears to be a type B or typeC peloric derivative of the more widespread E.coronaria (Chen & Tsi, 1987).
In other, more controversial cases the florallysimplified mutant has been recognised not only as adistinct species but also as a monotypic genus that isassumed to be phylogenetically primitive. Five such‘genera’ native to south-east Asia have been estab-lished by Chen and colleagues (for reviews see Chen,1982; Chen & Tsi, 1987), whose (possibly mis-leading) illustrations of all five taxa suggest fullactinomorphy of the perianth, consistent with eitherfloral plesiomorphy or types B or C peloria. Of thesefive ‘genera’, four are lower epidendroids (¯ ‘neot-tioids ’) with terminal stigmas; each of these taxaappears confined to a single locality (Chen, 1982,Fig. 2). Of these four neottioid taxa, three arevegetatively reduced mycoheterotrophs (‘ sapro-phytes ’) ; this makes identification of potentialancestors based on morphology unusually difficult,as few vegetative characters are available forcomparison.
The exception is Tangtsinia nanchuanica (Chen,1965), which closely resembles Cephalanthera (Dress-ler, 1993; Cribb, 1999) but bears five staminodes inaddition to the single functional stamen (A1).Among the apparently peloric mycoheterotrophs,Sinorchis simplex (Chen, 1978) bears the singlefunctional stamen and two staminodes (A1 anda2a3, respectively) typical of neottioids, and hasbeen attributed to either Cephalanthera (Dressler,1993) or, more likely, the closely related mycohetero-troph Aphyllorchis (Cribb, 1999). Diplandrorchis sinica
(Chen, 1979) resembles the similarly mycohetero-trophic Neottia, but has two opposing functionalstamens (presumably A1 and a3), lacks staminodesand has a distended ovary. In contrast with theabove monotypic ‘genera’, Archineottia includesseveral supposed species. Archineottia pantlingii and A.smithiana have well-developed neottioid labella butelongate filaments subtending the single functionalstamen, whereas A. gaudisartii, A. microglottis and A.acuminata are depicted as having actinomorphicperianths (Chen, 1982; Chen & Tsi, 1987). Again,all these putative species otherwise closely resembleNeottia (see also McCook & Bateman, 1990; Dressler,1993; Cribb, 1999).
The fifth south-east Asian ‘genus ’ that super-ficially appears to be type B peloric is an orchidoid.In contrast with the above neottioids, Aceratorchis is awidely distributed, cross-pollinated taxon (Chen &Tsi, 1987; Chen, Tsi & Luo, 1999) that, especially invegetative features, closely resembles the co-existing

422 Paula J. Rudall and Richard M. Bateman
Fig. 11. (A) Gymnadenia densiflora Wahlb. Typical resupinate flower well-adapted to pollination via butterflies attractedby the long nectar-filled spur. (B) Nigritella austriaca (Teppner & E. Klein) Delforge [¯Gymnadenia austriaca (Teppner& E. Klein) Delforge] ; stabilised, predominantly apomictic ‘genus ’ derived from Gymnadenia possessing type A pseu-dopeloric, non-resupinate flowers. (C, D) Ophrys ferrum-equinum Desf. (C) Typical flower with well-developed labellumbearing symmetrical markings (speculum). (D). ‘Lusus ’ flower with asymmetric labellum; the right-hand third hasdeveloped in the form of a sepal rather than a labellum (photograph by D. M. Turner Ettlinger). Scales : all ¬2±75.
genus Galearis (Wood & Cribb, 2001; cf. Bateman et
al., in press). Sadly, none of these five ‘genera’ haveyet been sequenced for DNA.
A similar example is evident among South Africanorchidoids of the Satyrium group, where two ‘subac-tinomorphic ’ (type A pseudopeloric) species withreduced labella and elongate gynostemia have been
assigned to the genus Pachites. It is not clear whetherthese are sister species (Linder & Kurzweil, 1999),nor is it clear whether they are the sister group of(and thus potentially ancestral to) Satyrium s.s.(Linder, 1986) or nested within Satyrium as sec-ondarily reduced morphs (Kurzweil, 1996). Al-though molecular phylogenies have begun to be

423Synorganisation, zygomorphy and heterotopy in floral evolution
constructed for South African orchids (Douzery et
al., 1999; Bateman et al., in press), ‘Pachites ’ has notyet been sequenced.
In yet other cases, the prospecies not onlysuccessfully establishes a new taxonomic species andgenus but also subsequently radiates to generateseveral descendant species, often via polyploidyand}or a transition from allogamy to autogamy.The best known examples are in subfamily Orchidoi-deae. The Australasian genus Thelymitra (Fig. 12F)is sister to Calochilus (Fig. 12E), then Cryptostylis, thenOrthoceras plus Diuris (Kores et al., 2001), all of whichhave labella, lateral petals and sepals that are easilydifferentiated (Jones, 1988; Pridgeon et al., 2001). Incontrast, Thelymitra bears six near-identical perianthsegments (the labellum is slightly narrower in a fewspecies, e.g. T. crinita and T. cucullata : Fig. 12F),suggesting that the genus originated by type Cpeloria with three sepals replacing the labellum andlateral petals. The presumed prospecies has sinceradiated into approximately 50 species (Jones, 1988;Clements, 2001) that vary greatly in the size andcolour of the diurnal flowers and encompass adiversity of pollination syndromes, including autog-amy, floral mimesis and a wide range of insectvectors (Bernhardt & Burns-Balogh, 1986; Burns-Balogh & Bernhardt, 1988; Bower, 2001).
The pseudopeloric European genus Nigritella
contains approximately 15 putative species, mostreflecting relatively recent polyploidy events and}ortransitions from allogamy to autogamy (e.g. Tepp-ner & Klein, 1998; Hedre!n, Klein & Teppner,2000). Nuclear rDNA (ITS) sequence data indicatethat Nigritella (Fig. 11B) is a molecularly homo-geneous monophyletic group nested phylogeneticallywithin Gymnadenia (Fig. 11A), which is very similarvegetatively but has far more strongly zygomorphicand fully resupinate flowers that possess longnectiferous spurs and are pollinated by lepidop-terans. Nigritella (which was sunk taxonomically intoGymnadenia by Bateman, Pridgeon & Chase, 1997)lacks resupination and has only a simplified labellumand vestigial spur; these are classic characteristics oftype A pseudopeloria (Bateman et al., 1997, in press ;Pridgeon et al., 1997; Bateman, 1999; Bateman &DiMichele, 2002).
(2) Non-resupination
With the possible exception of Apostasia, all orchidspecies or genera with erect multi-flowered inflores-cences that have flowers characterised by lack ofresupination reflect secondary loss of this character.In most cases the 180° torsion of the ovary}pedicel is
simply lost ; examples include the orchidoid Nigritella
and lower epidendroids Epipogium and Cephalanthera
damasonium, of which the latter bears unusuallyupright, goblet-shaped flowers and is phylogene-tically nested within an otherwise almost uniformlyresupinate genus. In a few extraordinary instances,rotation of the ovary has been extended through360°, returning the labellum to its position asuppermost tepal (e.g. Hammarbya : Darwin, 1877;Coen, 1999). Many of these taxa either become atleast facultatively autogamous or switch to polli-nating insects that prefer to walk rather than fly;both are strategies capable of recurring in a lineagethat has lost the functionality of its labellum.
VIII. DISTINGUISHING FLORAL
PLESIOMORPHY FROM ATAVISM
There are many accounts of teratologies affectingthe gynostemium and especially the perianth docu-mented in orchids in general and in the species-richsubfamilies Orchidoideae and Epidendroideae inparticular. This extensive record could be assumedto indicate a high mutation rate and thus be apossible contributor to the species richness of orchids.However, it is more likely to reflect two otherfactors : (1) the existence of a large and com-municative cadre of orchidologists (both professionaland amateur) actively reporting their observations,and (2) the unusually showy nature and stronglyexpressed zygomorphy of the flowers, which im-proves the probability of recognition of an ‘ab-normal ’ flower.
Given that the perianth is generally far moreconspicuous to the human eye than the gynoste-mium, contrasts in symmetry between these twopairs of whorls (compressed in the case of the threesepals and three petals that constitute the perianth,and fused in the case of the variable number ofstamens and the style that constitute the gyno-stemium) are often in practice overlooked. Thus, therecord of perianthial terata is on average morecomplete and more reliable than the record ofgynostemial terata. Nonetheless, some key examplesof probable perianthial terata remain equivocal incategorisation, the most important being the south-east Asian ‘genera’ Tangtsinia, Sinorchis, Diplandror-chis, Archineottia and Aceratorchis illustrated by Chen(1982). These all appear actinomorphic (i.e. types Bor C peloric) in his Fig. 1, but comparison of hisillustration of Aceratorchis tschiliensis (his Fig. 1.7)with the more detailed rendition of the same speciesin Genera Orchidacearum (Pridgeon et al., 2001, fig.

424 Paula J. Rudall and Richard M. Bateman
56.1) suggests that it is in fact type A or Bpseudopeloric, with a labellum that, although poorlydifferentiated, is marginally larger and significantlymore vascularised than the lateral petals.
Even when they are precisely described, thephylogenetic position, evolutionary significance andmost appropriate taxonomic treatment of orchidswith actinomorphic or weakly zygomorphic peri-anths remain highly contentious issues. For example,some reports of putative intergeneric hybrids (e.g.McKean, 1982) are more convincingly ascribed topseudopeloric morphs of one of the supposed parents.More importantly, the above-mentioned south-eastAsian ‘genera’ were said by Chen (1982, p. 17) topossess ‘ several primitive characteristics ; [this], andthe primitive position of the subtribes that theybelong to, show that these genera do not seem to bederivative or peloric, but surviving ancient types.Today they are restricted to some narrow regionsand appear near extinction’. However, the co-existence of each ‘genus ’ with vegetatively similarspecies, and the apparent restriction of each to asingle locality, seem remarkably coincidental. Wehypothesise that they are in fact recently originatedtype C peloric or, more likely, type B peloric orpseudopeloric morphs of lower epidendroids (or, inthe case of Aceratorchis, of a relatively derivedorchidoid) rather than distinct species or genera.
Fortunately, these contentious identifications cannow be resolved using DNA sequencing (e.g. Soltis,Soltis & Doyle, 1997) and various electrophoreticand DNA fingerprinting techniques (e.g. Hollings-worth, Bateman & Gornall, 1999) that are increa-singly applied to Orchidaceae (Qamaruz-Zaman et
al., 1998; Chase, 1999; Bateman, 2001). For ex-ample, such techniques have demonstrated (1) thatNigritella evolved recently, apparently from withinthe widespread, derived orchidoid genus Gymnadenia
(Bateman et al., 1997; Pridgeon et al., 1997;Bateman, 2001) (Fig. 11A, B), and therefore prob-ably originated by type A pseudopeloria, and (2)that Thelymitra is monophyletic and nested withinwell-differentiated, derived diurid Orchidoideae(Kores et al., 2001), and therefore probably origi-nated by type C peloria. Similarly, most cases ofnon-resupination are atavistically reduced ratherthan plesiomorphic.
IX. RECONSTRUCTING THE ‘ANCESTRAL
ORCHID’
At one particular moment in phylogenetic history anancestral population (or individual) of a non-orchid
species gave rise to the first population (or in-dividual) that we would readily recognise as anorchid. If this first orchid arose by anagenesis(presumably caused by directional selection) itsorigin would be morphologically imperceptible tothe observer and any attempt to define that originwithin the lineage would thus be arbitrary. However,if the first orchid arose by saltation, and hence wasby definition marked by a profound phenotypicchange across a single generation (Bateman &DiMichele, 1994, 2002), its origin is in theory readilydemarcated.
Some of the difficulties of reconstructing theancestral species (or prospecies) of Orchidaceae werediscussed by Frohlich (1987). As discussed above(Section V), choice of outgroup is ultimately criticalin polarisation of characters in morphological analy-ses, and in evaluating character evolution byoptimisation onto a molecular phylogeny. The mostcommonly selected outgroup for Orchidaceae isHypoxis (e.g. Freudenstein & Rasmussen, 1999),which is indeed a credible candidate in terms ofbeing hexatepalous (i.e. trimerous, as in mostmonocots), possessing many small seeds per ovarylocule, and is unusual among the potential sister-groups shown in Fig. 1 in being epigynous (see alsoFig. 13A). However, we believe that the ancestor ofthe Orchidaceae may, unlike Hypoxis, have possesseda rudimentary gynostemium. Dressler (1993) in-cluded stamen}staminodial fusion to the style amongthe features that he would expect to find in theorchid ancestor. Pauridia, Corsia and Orchidaceae arethe only monocots that unequivocally possess agynostemium. However, even given the existingrelatively unresolved optimisation framework shownin Fig. 1, the shared presence of a gynostemium inthese three clades cannot be taken as a singlesynapomorphy, but instead represents one of at leasttwo and probably three iterative originations of thegynostemium over a relatively short time period,perhaps reflecting an underlying pre-adaptation to‘hyper-epigyny’ in the group (Figs 13, 14). Indeed,homology between the gynostemia of Pauridia andorchids can be rejected on the similarity criterion;they are analogous, but may be homologous at adeeper level of a tendency to undergo congenitalfusion.
Pauridia and Corsia, with different degrees ofadnation and floral symmetry, may both resemblethe immediate ancestors of Orchidaceae, but indifferent ways. One possible scenario is that stamen–style adnation (linked with at least partial sup-pression of the outer stamen whorl) preceded zygo-

425Synorganisation, zygomorphy and heterotopy in floral evolution
morphy in orchid evolution, in which case Pauridia,which lacks a complete stamen whorl (including A1)and possesses a gynostemium, may be the closeranalogue, even though Corsia, with both a gyno-stemium and labellum, superficially appears moresimilar to orchids in overall floral morphology (but,as noted above, the labellum is non-homologous).
Corsia is an unlikely candidate for an orchidoutgroup. As currently delimited, the small achloro-phyllous mycoheterotrophic family Corsiaceae com-prises three small, widely dispersed genera: Corsia
(mostly from New Guinea), Arachnitis (southernSouth America) and Corsiopsis (China) (Neinhuis &Ibisch, 1998; Zhang, Saunders & Hu, 1999).Phylogenetically, Corsiaceae have been variouslyplaced with several other facultatively or obligatorilymycoheterotrophic monocots, including orchids, butmost commonly with Thismia (Dioscoreales). Thismia
shares several morphological similarities with Corsia,including pollen aperture type and floral zygomor-phy (Rudall & Eastman, 2002). However, recentanalysis of molecular sequence data from 26S rDNA(Neyland, 2002) placed Corsia within Liliales, poss-ibly as sister to Campynema, a monotypic Tasmanianendemic in the taxonomically isolated, bigenericfamily Campynemataceae (Liliales). Preliminaryanalysis of other molecular data from three genes(atpA, cob and rbcL) have also placed Arachnitis withCampynema ; the two mitochondrial genes, atpA andcob, determined the position of Arachnitis (J. Davis,D. W. Stevenson, personal communication, 2001).Arachnitis and Campynema are the only two lilioidgenera with a combination of epigyny and threeseparate styluli, although this combination alsooccurs in a few Alismatales (Igersheim, Buzgo &Endress, 2001). Clearly, more work is needed tofirmly establish the relationships of Corsiaceae, buton current evidence a sister-group relationshipbetween Corsia and Orchidaceae seems unlikely.
Regarding the other taxon with a gynostemium,Pauridia, all recent molecular analyses (e.g. Chase et
al., 2000; Fay et al., 2000) indicate placement of bothHypoxidaceae and Orchidaceae within Asparagales,among a poorly resolved basal group of six families(Fig. 1). Hypoxidaceae are therefore potentiallysister family to Orchidaceae, a relationship inferredby some previous authors (e.g. Hutchinson, 1934;Garay, 1960, 1972) based on morphological features.Hence, the shared presence of a gynostemium inPauridia and Orchidaceae merits exploration, al-though a sister-group relationship seems unlikely(see below). Relationships within Hypoxidaceae arecurrently poorly resolved and require further investi-
gation. The most extensive molecular analysis ofHypoxidaceae, sampling representatives of eight outof the nine widely recognised genera includingPauridia (Rudall et al., 1998), found the large genusHypoxis to be polyphyletic (though this preliminaryconclusion was based on only two species sampledout of a potential 50). Pauridia occupied an unusuallylong branch, being placed as sister to Spiloxene plusHypoxis glabella. Neither Hypoxis nor Pauridia weresister to all other Hypoxidaceae, so neither repre-sented the most obvious outgroup for orchid analy-ses. However, Pauridia, a highly divergent ‘wildcard’taxon assigned to various families in earlier classifica-tions (Thompson, 1979), has characters in commonwith both Hypoxidaceae and Orchidaceae, and fellas tentative sister to Orchidaceae in a recentmorphological analysis of Asparagales (Rudall, inpress).
Both Pauridia and orchids have undergone stamenreduction and staminodial adnation to the style,although in Pauridia a complete whorl was lost (asalso in almost all orchids). Loss of a complete stamenwhorl is a relatively frequent event in floweringplants ; Walker-Larsen & Harder (2000) estimatedthat it has occurred at least once within 34% ofmonocot families. If Pauridia, which has fertilestamens a1, a2 and a3 (Fig. 4B), also lost the medianinner stamen (a3), it would resemble the floralarchitecture of the diandrous cypripedioid orchidsand Apostasia, but without a labellum. Comparablepatterns are evident in other lower asparagoids. Forexample, in Iridaceae only the outer stamen whorl ispresent (A1, A2 and A3), but in the Australiangenus Diplarrhena one of these stamens is sterile,leaving only two fertile stamens (Rudall & Gold-blatt, 2001). However, in Iridaceae there are novestiges of stamen traces corresponding to the ‘ lost ’inner whorl (P. Rudall, unpublished observations),in contrast to Pauridia.
A further similarity between Pauridia and theapostasioid Neuwiedia is that they have bisulcate(‘pontoperculate ’ sensu Erdtman, 1952) and opercu-late pollen, respectively (Newton & Williams, 1978;Thompson, 1979; Simpson, 1983); these two ap-erture conditions are closely similar and may behomologous, forming a credible transformationseries, although operculate pollen also occurs inseveral lower asparagoids such as Tecophilaeaceae(Simpson, 1985).
On the other hand, Pauridia lacks some of themajor synapomorphies for orchids, such as thepresence of silica bodies (although these are sec-ondarily lost in subfamilies Epidendroideae and

426 Paula J. Rudall and Richard M. Bateman
Orchidoideae: Møller & Rasmussen, 1984). Afurther major difference between Pauridia and Orchi-daceae is that Pauridia possesses actinomorphicflowers, in common with most other lilioid monocotsbut in contrast to the strongly zygomorphic flowersof most orchids, in which zygomorphy results fromadaxial stamen suppression and (except in Apostasia
and a few other orchids) by labellum development.Pauridia shares several characters with Hypoxi-daceae, including closely similar ovule and (ap-parently non-mycotrophic) seed morphology (deVos, 1949) and the presence of mucilage canals invegetative tissues, otherwise found only in Astelia-ceae among the monocots (Rudall et al., 1998). Oncurrent evidence it seems highly unlikely thatPauridia will ultimately prove to be the closest extantsister taxon to Orchidaceae. Rather, Pauridia pro-vides a credible morphological analogue for theorchid ancestor to compare with Hypoxis (Figs 13,14).
DNA sequencing has demonstrated that Apostasia
sections Apostasia and Adactylus are monophyleticsister taxa (Kocyan & Endress, 2001a) and Apostasia
s.l. is sister to Neuwiedia (Cameron et al., 1999;Kocyan & Endress, 2001a). Furthermore, Aposta-sioideae is indisputably the basally divergentsubfamily within Orchidaceae; on rbcL evidenceCypripedioideae diverge second (Cameron et al.,1999; but see Section II.1), and Selenipedium is thebasalmost genus within Cypripedioideae (Cox et al.,1997). Thus, Chen’s (1982, p. 18) intuitive statementthat ‘ the flowers of Neuwiedia, Selenipedium, Tangtsinia
and Aceratorchis [are] probably the most primitiverepresentatives of the four main groups [subfami-lies] ’ appears corroborated for the first two genera.
Clearly, supposedly ‘primitive ’ characteristicswithin Orchidaceae (at least, those relating togynostemium formation and labellum differentia-tion) can be both genuinely plesiomorphic in basallydivergent groups such as the apostasioids, andapomorphic, reflecting reversals (and thus by defini-tion atavistic), in more derived groups. In particular,trends towards reduction in the number of stamensand an increasingly well-differentiated labellum areboth frequently reversed, though it is notable that inneither case have phylogenies revealed furthertransitions (e.g. derived but peloric or pseudopeloricorchid genera such as Thelymitra and Nigritella havenot reverted to more strongly zygomorphic peri-anths).
The perception of the basal members of Orchi-daceae as a mosaic of plesiomorphic floral charactersthat was pioneered by Chen (1982) and elaborated
by Dressler (1993) and Chase (2001) appearsbroadly accurate. Of the ten ‘primitive ’ orchidcharacters listed by Chen (1982), the presence ofmultiple stamens (originally six), terminal positionof the stigma, absence of a rostellum, absence of aclinandrium separating the fertile stamen from thestigma, lack of viscid pollinia with pollen grains inmonads, at least basally trilocular inferior ovary,and small seeds within a sclerotic seed testa allclearly represent what might be termed ‘plesiomor-phic tendencies ’, as does the abscission of theperianth plus gynostemium specified by Dressler(1993). To these characters we add parasitism ofsymbiotic fungi by the seeds (‘germinatory mycohe-terotrophy’), which allowed the development of thecharacteristic ‘dust seeds ’, and emphasise the im-portance of epigyny and syncarpy, which are pre-requisites for gynostemium formation, as discussedearlier. It also seems likely that the hypothetical‘ancestral orchid’ had a gynostemium.
Two other features central to this paper, differen-tiation of the median petal into a labellum andresupination, are less clearly plesiomorphic. Asdiscussed in Section II(1), more recent phylogenetictrees of the orchid subfamilies contradict the rbcL
tree of Cameron et al. (1999) which placed cypripe-dioids as diverging below vanilloids, therebymaking the A1-monandrous orchids monophyletic(Fig. 13), although this topology lacked bootstrapsupport. More recent analyses using atpB andpsaB alone both placed the vanilloids below thecypripedioids, admittedly also without bootstrapsupport (K. Cameron, personal communication,2001), but a combination of the three plastid genesyielded the latter topology with significant bootstrapsupport. Thus, it is necessary to consider an alter-native set of morphological character optimisationsthat reflect this alternative molecular topology,wherein the A1-monandrous orchids are diphyletic(Fig. 14).
Part of the ambiguity in the origins of the labellumand resupination is caused by the fact that they areabsent from Apostasia but present in its sister genus,Neuwiedia. It is therefore equally parsimonious toview labellum differentiation and flower resupi-nation as together having separate origins in Neuwie-dia and the non-apostasioid orchids (and thus absentfrom the hypothetical ancestral orchid: Deltranoptimisation), or as having been lost from Apostasia
(and thus potentially being present in the hy-pothetical ancestral orchid: Acctran optimisation;Figs 13, 14). Floral atavism in Apostasia is a crediblehypothesis, as partial adnation of stamens to the style

427Synorganisation, zygomorphy and heterotopy in floral evolution
(and hence close contact of anthers) may havefunctionally established the ‘Solanum-type’, vibratilebuzz-pollination syndrome of the flowers. Moreover,the issue of resupination is made yet more complex ifthe morphological homologies are considered morecarefully, as the mechanism of resupination is poorlydescribed for Neuwiedia and apparently differs be-tween the cypripedioids, where it relies on inversionof the pedicel, and the monandrous subfamilies,where it occurs via torsion of the pedicel and}orovary. Thus, optimisation under Deltran wouldallow three separate origins of resupination bytorsion in the Orchidaceae.
With regard to expression and suppression ofstamens, if Pauridia were used as an orchid outgroupwith the rbcL topology, a secondary development oftriandry in Neuwiedia (in which A1 is present) wouldbe required, involving a secondary regain of stamenA1 in monandrous orchids and Neuwiedia, ratherthan loss of A1 in cypripedioids and Apostasia, asillustrated in Figs 13 and 14. Stamen regain seemsunlikely on first principles, but admittedly appearsto have at least occurred in the epidendroid ‘genus ’Diplandrorchis (Chen, 1979). If the atpB}psaB top-ology is correct (Fig. 14), stamen characters appeareven more homoplastic but the emphasis is placedmore credibly on losses, including independent lossesof A1 in Apostasia and Cypripedioideae, and in-dependent losses of a1 plus a2 in Vanilloideae andEpidendroideae plus Orchidoideae.
Overall, the difficulty of resolving the relativepositions of the five subfamilies using data fromeither DNA sequences (e.g. Cameron et al., 1999; K.Cameron, personal communication, 2001) or mor-phology (e.g. Freudenstein & Rasmussen, 1999;Johansen & Frederiksen, 2002), together with theretention of at least some plesiomorphic character-istics in the basal cypripedioid Selenipedium
(Albert, 1994; Cox et al., 1997) and to a lesser degreein lower Epidendroideae (Freudenstein & Ras-mussen, 1999), suggest a rapid initial radiation ofOrchidaceae (cf. Bateman, 1999). Recent molecularclock estimates indicate that this radiation may havebeen unexpectedly deep (dated to approximately100 million years before present by Chase, 2001).Initial evolutionary experimentation in the sup-pression or differentiation of floral organs, notablythe stamen and labellum, was superseded by moresubtle elaborations of the (usually single) fertilestamen (e.g. the pollinium, caudicle and viscidium),staminodes (including the column wings and theirputative fusion product, the clinandrium), stigmas(including the rostellum) and labellum (e.g. spur,
lobing, markings). We will return to these charactersin Section XII.
X. DESCRIBING EVOLUTIONARY
TRANSITIONS IN FLORAL MORPHOLOGY:
HETEROCHRONY, HETEROTOPY AND
HOMEOSIS
Section X sets inferred evolutionary transitions inthe context of heterochrony (evolutionary changesin the developmental timing of expression of specificfeatures), heterotopy (evolutionary changes in theposition of expression of specific features) andhomeosis (a subset of heterotopy), focusing initiallyon the potential application of these terms andconcepts to teratological orchid flowers. Here, weemploy the revised definitions of these terms thatwere developed by Baum & Donoghue (2002) andapplied to orchid flowers by Bateman & DiMichele(2002). Duplication of an existing structure and itsexpression in a position not previously occupied byanother structure is termed neoheterotopy (anexample would be the frequent orchid terata with asecond labellum, thus generating seven tepals ratherthan the usual six). Homeoheterotopy encompassescases where the duplicated structure replaces acontrasting pre-existing structure, either partly orcompletely (only the latter examples are strictlytermed homeosis).
Most cases of true peloria, which leads to perianthactinomorphy, appear simple to categorise using thisrevised terminology. In type A peloria, each of thetwo lateral petals is replaced by a labellum, and intype B peloria the labellum is replaced by a thirdlateral petal (Figs 3, 10, 12). Both type A and typeB peloria ostensibly represent homeoheterotopy; thetransfer of genetic identity from the donor structureof the ancestor to the recipient structure of theimmediate descendant. Moreover, in both cases thereplacement occurs between contrasting members ofthe same lateral appendage whorl (namely petals),and the heterotopy is thus termed lateral. However,Baum & Donoghue’s (2002) strict definition ofhomeosis requires the transfer of all aspects of thegenetic identity from donor to recipient. Thisrequirement is probably fulfilled in type A peloria,where the full, complex labellum is assembled atboth locations where the less complex lateral petalsare normally expressed. However, if the orchidlabellum indeed proves to be a developmentallycomplex structure (arguably compound; see SectionVI), the genetic apparatus underlying the devel-

428 Paula J. Rudall and Richard M. Bateman
Fig. 12. (A) Ophrys mammosa Desf. Teratological flower in which the lower portions of the expanded lateral sepalshave developed the pigmentation and reflective speculum typical of the labellum (photograph by A. J. Hughes).(B–D) Ophrys apifera Huds. (B) Typical flower with small, triangular, greenish lateral petals. (c) Flower with ex-panded, pink lateral petals intermediate between those of plants B and D. (D) Flower with strongly expanded, pink,

429Synorganisation, zygomorphy and heterotopy in floral evolution
opment of the lateral petals is likely to be a simplerversion of that controlling labellum development. Ittherefore seems more likely that, in type B peloria,the supplementary developmental–genetic appar-atus responsible for the differentiation of the la-bellum relative to the relatively poorly differentiatedlateral petals has been suppressed. In the parlance ofBaum & Donoghue (2002), this interpretation woulddefine type B peloria as homeoheterotopy but not ashomeosis s.s. In the rarer cases of type C peloria(Lang, 2001) three sepals replace the three petals.This transition also constitutes homeoheterotopy,but on this occasion the homeotic translocationoccurs from the basal to the adjacent whorl : this isacropetal homeoheterotopy.
Pseudopeloria, wherein the ancestral degree ofperianth zygomorphy is reduced but true actino-morphy is not achieved, is more troublesome tointerpret. Here, the labellum is simplified in shapeand}or altered in size so that it resembles the sepals(indeed, the derived morph will only be recognisableas pseudopeloric rather than types B or C peloric ifthe sepals and petals of the ancestral morph aredistinguishable from each other : Figs 3, 10, 11). Ifthe new structure occurring at the site normallyoccupied by the labellum is regarded as a bona fide
sepal, pseudopeloria is by definition heterotopic, liketrue peloria ; this is termed type B pseudopeloria inFig. 3 (bottom centre). Like type C peloric hetero-topy, this pseudopeloric heterotopy cannot be lateralbut rather should be viewed as acropetal ; theadditional sepal is expressed in the whorl of threepetals and therefore is inserted slightly closer to theapex of the floral axis than is the whorl of threesepals.
However, the majority of examples of pseudo-peloria (type A in Fig. 3) are open to an alternativeinterpretation. Rather than the spatial transferenceof a feature implicit in heterotopy, pseudopeloriacould equally be viewed as reflecting temporalchange in events during ontogeny; in other words, itcould represent heterochrony (cf. Bateman, 1994;Baum & Donoghue, 2002). More precisely, themorphologically simplified inner perianth segmentmay remain homologous with the ancestral labellum,but be paedomorphic, having suffered either slowedor precociously terminated development relative toits immediate progenitors (neoteny or progenesis,
sepaloid lateral petals showing type C pseudopeloria and attributable to ‘subsp. jurana’ (photographs B–D by D. M.Turner Ettlinger). (E) Calochilus robertsonii Benth. Highly differentiated flower typical of this sister-genus to Thelymitra.(F) Thelymitra cucullata Lindl. Flower typical of the genus, with labellum similar to the other perianth segments (inthis case slightly smaller). Scales: A–D, x3.3; E, F, x1.7.
respectively, sensu Alberch et al., 1979). This hypo-thesis could be tested by comparing the develop-mental trajectories of normal and pseudopeloricflower buds.
Closer examination suggests that in the mostcommon mode of pseudopeloria, type A, the la-bellum is indeed replaced by an organ that moreclosely resembles the sepals than the lateral petalsbut, in species that normally possess a labellar spur,the supposedly ‘ sepaloid’ labellum also usually bearsa partially developed or at least vestigial spur (e.g.Bateman, 1985), unlike the bona fide sepals. Also, ifthe labellum normally bears discrete markings, theseusually persist, albeit typically in a reduced form (seeFig. 3, bottom left). Together, these observationssuggest that the structure expressed in the locationnormally occupied by a well-differentiated labellumis in fact a heterochronically reduced labellum ratherthan a heterotopically expressed sepal.
Interestingly, the boundary between heterotopyand heterochrony itself is then challenged by a thirdcategory of pseudopeloria, type C (Fig. 3, bottomright). This is best illustrated by a widely anditeratively occurring range of morphs of Ophrys
apifera that are taxonomically lumped as subsp.jurana (the more commonly used descriptor ‘var.friburgensis ’ strictly refers to only a subset of thesemorphs). A typical Ophrys apifera flower has verysmall, greenish, triangular lateral petals (Fig. 12B),but in subsp. jurana these are pink and enlarged, thusmore closely resembling the sepals (Fig. 12C, D).This phenomenon reduces but does not eliminate thedegree of zygomorphy shown by the perianth, andhence is most appropriately termed pseudopeloria.However, the tepals replacing the lateral petals varygreatly in size in jurana ; those shown in Fig. 12C areunusually small, whereas those shown in Fig. 12Dare more frequent (Landwehr, 1977; Bourne! rias,1998; Ettlinger, 1998; Delforge, 2001), but onlyrarely do the replacement petals approach the size ofthe true sepals (Lang, 2001, his fig. 2). Moreover,those individuals with more expanded sepaloidlateral petals are more likely to show other floralabnormalities ; first the breakdown of the complexlabellum markings (Fig. 12D) and in more extremecases disruption of the oval outline of the labellum(Landwehr, 1977; Bourne! rias, 1998). Hence, theexpanded lateral petals of jurana do appear to be

430 Paula J. Rudall and Richard M. Bateman
sepaline, and thus analogous to the acropetalheterotopy of type B pseudopeloria, but they are alsoin most cases less well developed than the true sepals,and thus could be viewed as showing paedomorphicheterochrony analogous to type A pseudopeloria.Moreover, when the recorded cases of type A peloriaare carefully re-examined, the additional labellasubstituting for the lateral petals also prove to besmaller than the normal labellum, even in examplesthat approach equality of size such as that in Fig.10B.
XI. INTERPRETING EVOLUTIONARY
TRANSITIONS IN FLORAL MORPHOLOGY:
EVOLUTIONARY–DEVELOPMENTAL
GENETICS OF ADNATION, ZYGOMORPHY AND
RESUPINATION
When fully integrated with ongoing ontogenetic andphylogenetic studies, evolutionary–developmentalgenetics should allow us to move from the descrip-tions of specific morphologies in Sections III and IVand the interpretations of specific evolutionarytransitions in Section X to the ‘New Morphology’ :genuinely causal explanations that relate profoundchanges in morphogenetic expression to modifica-tions in key developmental genes. However, despitemany recent advances in this field, the explanatorymodels remain simplistic and relevant empiricaldata remain sparse.
(1) Morphogenetics in monocots
Much of this work has relied on studies of mutationsin cultivated populations of a few ‘model ’ eudicotspecies, notably Arabidopsis and Antirrhinum, morerecently augmented by evolutionary studies span-ning a wider (yet still modest) range of higher taxa;typically, monocots are represented by economicallyimportant grasses (cf. Kramer & Irish, 1999; Cronk,2001; Cronk, Bateman & Hawkins, 2002). In floralstudies, MADS-box genes contributing to the nowfamous ABC model of floral whorl control (e.g. Coen& Meyerowitz, 1991; Theissen et al., 2002) havedominated conceptual thinking.
Endress (1999) postulated that in some largegroups with zygomorphic flowers, such as orchids,complex zygomorphy (‘monosymmetry’) is main-tained by a high degree of synorganisation (strictly,repositioning multiple organs in a single plane) andorgan fusion. However, it is becoming increasinglyevident that the two conditions are controlled bydifferent sets of genes. The recent cloning of the
fiddlehead gene (Yephremov et al., 1999; Pruitt et al.,2000), which is responsible for regulating contact-mediated epidermal cell adhesion (i.e. postgenitalfusion, a phenomenon that is primarily restricted tohigher plants : Endress, 1997), has prompted dis-cussion of its implications for vegetative and re-productive organ fusion (Sinha, 2000) and evensyncarpy in angiosperm evolution (Raven & Weyers,2001). However, as fiddlehead primarily affects post-genital fusion, syncarpy is probably controlled by adifferent set of genes ; genetic systems controllingcongenital fusion are apparently highly complex andcurrently poorly understood (Gustafsson & Albert,1999). Clearer knowledge of the homology andphylogenetic distribution of epigyny in monocotyle-dons is crucial in this context (Rudall, 2002).Specifically, stamen adnation to the style may be aproduct of over-expression of the genes related toepigyny, resulting in a form of hyper-epigyny.
On the other hand, expression of zygomorphy inorchids may be controlled by a relatively simplegenetic system. In some asterid eudicots, zygo-morphy is dependent on early expression of theCYCLOIDEA (CYC) gene on the adaxial side of theflower, accentuated by the DICHOTOMA (DICH)gene, resulting in abortion of the adaxial stamen(s)(e.g. Luo et al., 1996, 1999; Coen, 1996; Cronk &Mo$ ller, 1997; Cubas, Vincent & Coen, 1999;Citerne, Mo$ ller & Cronk, 2000; Theissen, 2000;Cronk, 2001; Endress, 2001a ; Cubas, 2002; Froh-lich, 2002; Gillies et al., 2002; Theissen et al., 2002).A similar situation therefore seems likely to charac-terise orchids, although recent unpublished exper-imental work has so far failed to prove this hypothesis(Q. Cronk, personal communication, 2001).
Among lilioid monocots, although a few otherfamilies have zygomorphic stamen reduction (e.g.Tecophilaeaceae and Iridaceae), the combination ofadaxial stamen suppression and adaxial labellumformation is apparently restricted to Orchidaceae.Therefore, this adaxial expression of zygomorphy inboth perianth and gynostemium is unique to orchidswithin lilioid monocots. Similar developmental pat-terns occur in some commelinid monocots in theorders Zingiberales and Commelinales. Within Pon-tederiaceae, Heteranthera has A1, a1 and a2 fullydeveloped, thus closely resembling Neuwiedia, where-as Hydrothrix (Pontederiaceae) possesses only A1, asin monandrous orchids (e.g. Vermeulen, 1966).
The contrasting expression of zygomorphy inCorsia (i.e. labellum}nectary formation from themedian sepal, rather than the median petal oforchids) may either reflect a novel genetic system or

431Synorganisation, zygomorphy and heterotopy in floral evolution
a heterotopic expression of orthologous genes. Again,similar patterns occur in commelinid monocots. Insome Commelinaceae (e.g. Commelina) the abaxialstamens are sterile (Vermeulen, 1966); these are theopposite stamens to those suppressed in the closelyallied family Pontederiaceae (e.g. Chase et al., 2000).Furthermore, in a few orchids such as some Apostasia
species, the median sepal is marginally the broadestperianth segment, at least toward the proximal end(Fig. 9C, D). In most orchids the spur is formed onthe adaxial median petal (labellum), but in contrast,in the orchidoid Disa and a few other Diseae, spursdevelop on the abaxial median sepal (e.g. Vermeu-len, 1966; Kurzweil, 1998).
(2) Morphogenetics in orchids: indirectevidence
To continue a recurring theme of this paper, terataoffer valuable insights into the evolutionary im-portance and, if their causal mechanism can bedetermined, into the genetic control of floral zygo-morphy across flowering plants (cf. Meyerowitz et
al., 1989; Cubas et al., 1999; Theissen, 2000; Cubas,2002). Probably the best-known example is An-tirrhinum, where several different cycloidea and dicho-toma mutations have been identified that producepeloric flowers (e.g. Coen, 1991, 1996). Moreprecisely, type A peloria in orchids can be analogisedwith the ‘dorsalised’ phenotype of the backpetals
mutant in Antirrhinum, whereas type B peloria rep-resents ‘ventralised’ phenotypes resembling the well-known cycloidea mutant.
The most obvious (though not the only) geneticexplanation of pseudopeloria s.l. is that one or moregenes of relatively minor morphogenetic effect,operating downstream of the few developmentalgenes of more profound effect that determine thepositions and gross morphology of the floral organs,have been under-expressed or suppressed. If so, thewide range of morphological transitions encom-passed under the catch-all term of pseudopeloria canbe explained by differences in the types of down-stream genes affected. This hypothesis can, of course,only be tested by accurately identifying the genes(and resulting biosynthetic pathways) responsible.
Further understanding of orchid floral evolutionshould result if we can identify morphogenetic linksbetween gynostemium, perianth and ovary}pedicel(notably resupination), in order to better focus time-consuming evolutionary–developmental investiga-tions of probable controlling genes.
Unfortunately, in many cases, careful descriptions
of perianth teratologies are unaccompanied bydescriptions of the gynostemia of the self-sameflowers, reinforcing the need to state explicitly thesymmetry of each floral whorl. Available dataindicate that most peloric modifications of the tepalsare concomitant with little if any modification of thestrongly zygomorphic gynostemium. In some casesof orchid pseudopeloria, such as the well-known‘sepaloid’ morph of Ophrys apifera (e.g. Lang, 1980,2001), the remaining perianth segments and thegynostemium appear normal. However, in manyother examples, including the jurana morph of O.apifera (Fig. 12D) and the type A pseudopeloricPlatanthera (Fig. 10D) the remaining perianth seg-ments are abnormal, and the gynostemium alsoappears to be modified (in some cases resulting insterility : Bateman, 1985). This suggests multiplegene control of perianth morphology in orchids, butalso some linkage of control of zygomorphy in theperianth and gynostemium.
In particular, true peloria, which involves genu-inely homeotic translocation of structures within asingle floral whorl in the more frequent type A andtype B peloria, appears less likely to disrupt otherwhorls than does pseudopeloria, in which sepalinecharacteristics are apparently transferred to thepetaline whorl (Bateman, 1985). As discussed above(Section VI), orchid petals are in any case partiallysepaline in their vasculature. The true evolutionarystory must be even more complex, given that weinfer that the apparently sepaline floral organ of typeA pseudopeloric orchids is actually a morpholo-gically reduced labellum. In addition, although wehave demonstrated partial replacement of the lateralsepals by labella (Fig. 12A), we have not encoun-tered any terata where fully differentiated labellawere expressed in the sepal whorl. This indicates thatstrong developmental constraints inhibit the basi-petal translocation of organs from the petal to thesepal whorl.
One more inference is possible. When the evidencefor reduced size of heterotopic structures in theabaxial (i.e. the non-labellar) half of the flower (Figs10B, 12D) is considered in tandem with the partiallylabelloid lateral sepals of the flower shown in Fig.12A, it is clear that there is a dorsiventral cline ingene expression within the orchid flower.
(3) Morphogenetics in orchids: directevidence
Sadly, experimental evidence has yet to catch upwith the above inferences. It was obvious during the

432 Paula J. Rudall and Richard M. Bateman
earliest phases of the evolutionary–developmentalgenetic revolution that the strong morphogeneticdifferentiation among orchids makes them poten-tially excellent models (McCook & Bateman, 1990),but their numerous chromosomes and relatively longlife histories are less conducive. Thus far, all researchon homeotic genes in orchids has focused on typicalhigher plant (‘MIK’) MADS-box genes in higherepidendroids (for review see Johansen & Frederik-sen, 2002). Of the five genes isolated to date, four (atleast two of them orthologues) belong to the poorlyunderstood AGL2 clade and appear to play asecondary role in inflorescence initiation and}orfloral ontogeny, and the fifth, DgMADS2, is an A-class gene (Yu & Goh, 2000). One member of theAGL2 gene family, Adom1, has been investigated ingreater detail using both Northern blots (Lu et al.,1993) and in situ hybridisation. These studies showthat Adom1 is implicated in the development of thestigma and pollinia, apparently by cell-wall modifi-cation, probably in tandem with at least one otherheterodimer. This gives some hints regarding thepossible control of the more subtle aspects of orchidfloral morphology, but it seems likely that the genesof most profound morphogenetic effect will prove tobe the usual suspects, such as SQUAMOSA (A-class),PISTILLATA}APETALA (B), AGAMOUS (C) andespecially CYCLOIDEA. Although CYCLOIDEA hasthus far never been isolated in orchids or indeed mostother monocots, Doebley, Stec & Hubbard (1997)reported that the teosinte branched1 (tb1) locus inteosinte (Zea mays subsp. parviglumis) shares tworegions of homology with the CYCLOIDEA gene ofAntirrhinum. They drew several parallels between tb1
and CYC, including arrested growth of the petalsand stamens in the (secondary axillary) ears inwhich tb1 is expressed in teosinte.
The potential reversibility of some of thesemutations also merits closer scrutiny (Bateman &DiMichele, 2002). Current phylogenetic evidencesuggests that the transition to fully developed(achlorophyllous) mycoheterotrophy is irreversible,despite occurring at least eight times within Orchi-daceae, in Orchidoideae, Vanilloideae and especiallyEpidendroideae (Molvray et al., 2000; Hollings-worth et al., 2001). This is easily explained by themutational free-fall (indels and point mutationsleading to frameshifts) experienced by genes associ-ated with photosynthesis once their functionality islost. In contrast, evidence from terata demonstratesthat the relative morphological complexity of thelabellum and the suppression of all but one func-tional stamen remain genetically reversible, perhaps
because many key developmental genes have mul-tiple functions. At least some terata may reflectchanges in regulation rather than loss of genefunction. Admittedly, in almost all cases the resultingterata probably fail to establish evolutionarily viablelineages (Bateman & DiMichele, 1994, 2002).
XII. KEY INNOVATIONS AND RADIATION(S)
OF ORCHIDACEAE
Endress (1999, 2001a, b) regarded the occurrence ofstrong zygomorphy in certain major plant groupssuch as orchids as a key innovation that promptedsubsequent adaptive radiations, an opinion thatwould probably attract widespread support amongevolutionary botanists. However, both key inno-vation and adaptation are notoriously difficultconcepts to prove (Erwin, 1992; Bateman et al.,1998; Bateman, 1999), especially in the near-absenceof a fossil record for the family (cf. Mehl, 1986).
Among orchids, the labellum and gynostemium(together conferring strong floral zygomorphy) arecommonly invoked as a vital pairing for attracting,and interacting with, the airborne pollinators thatare widely regarded as the primary cause of theextraordinary species richness of the family (e.g.Burns-Balogh & Bernhardt, 1985; Kocyan & End-ress, 2001a). The labellum has two likely functions inrelation to insect pollination: firstly as a visualattractant and secondly as a landing stage. Tooptimise its effectiveness as a landing stage thelabellum must in most cases be lowermost. In erectrather than pendent inflorescences this posturerequires 180° resupination, which could also intheory be a key character.
Irrespective of whether they are sister groups or,as seems more likely, merely close relatives (Figs 13,14), the presence of a gynostemium in both Orchi-daceae and Pauridia (perhaps a pre-adaptation to‘hyper-epigyny’, as discussed in Sections IX andXI) indicates that the origin of the gynostemiumpreceded the origin of the labellum, althoughadmittedly, Acctran optimisation would allow sim-ultaneous origin of the two structures. It is equallyparsimonious to envisage the origin of both labellumand resupination s.l. at the base of Orchidaceae,subsequently lost from Apostasia (Acctran), or in-dependent origins of this pair of characters inNeuwiedia and the four derived subfamilies (Deltran:Fig. 13). The atpB}psaB topology requires anadditional origin of resupination by torsion invanilloids (Fig. 14). As Pauridia (two species) andApostasia (eight species) are both species-poor and

433Synorganisation, zygomorphy and heterotopy in floral evolution
Fig. 13. Diagram of orchid subfamily relationships based on molecular topology achieved by analysis of rbcL
(Cameron et al., 1999), but assuming only two immediate outgroups: Pauridia Harvey and Hypoxis L. (both Hypo-xidaceae), with several non-molecular characters optimised as indicated. 1¯ suppression of adaxial stamens (A2, A3,a3), 2¯ (resupination), 3¯ (labellum), 4¯dust seeds, 5¯ germinatory mycoheterotrophy Parentheses indicatecharacter state not present in all taxa. Arrowed text in italics indicates either syndromes (suits of linked characters,e.g. zygomorphy) or apomorphic tendencies. Asterisk indicates character present in derived taxa only.
ecologically unsuccessful, the gynostemium alone (atleast, in its most plesiomorphic form) cannot beviewed as a key character. The combination ofgynostemium plus labellum (made fully functionalin most erect inflorescences by resupination) appearsfrom first principles to be a more credible candidatefor a key combination of characters, but Neuwiedia
(eight species) possesses this triple combination andyet has the same number of species as its sister-genus,Apostasia.
A well-differentiated labellum and adhesive pollengrains allowed the cypripedioids only a modestincrease in species richness (approximately 120species), as did reduction to a single functionalstamen and evolution of cohesive pollinia in themajority of vanilloids (approximately 200 species).Reproductive biologists place particular emphasison the sequence of acquisition of minute wind-blownseed and pollen grain aggregation as pollinia, sincethere is a genetic trade-off between (a) damagingcompetition among genetically similar ovules andpollen grains originating from the same individual,and (b) the need for a larger number of pollen grainsto fertilise an ovule-rich ovary (cf. Van der Pijl &
Dodson, 1966; Neiland & Wilcock, 1995, 1998;Johnson, Linder & Steiner, 1998; Johnson &Edwards, 2000). We agree with Johnson & Edwards(2000) that current evidence suggests that the minuteand (most importantly) mycotrophic seeds, probablypresent in the ancestor of Orchidaceae, were thedriving force behind the subsequent evolution ofcohesive pollinia (a characteristic that reaches itsmost extreme form in the hard pollinia of the morederived, species-rich members of the Epidendroi-deae: Dressler, 1993). Under these circumstancesthe considerable gene-flow that occurs during seeddispersal minimises the need for gene-flow throughdispersal of individual pollen grains (Squirrell et al.,2001). Furthermore, the presence of a gynostemiumwith intimate synorganisation of stamens and style,as in most orchids (in contrast to merely basal fusion,as in apostasioids), was a precondition for theevolution of pollinaria. Apostasia has apparentlysecondarily achieved such intimate synorganisationof stamens and style by postgenital fusion of theanthers to the stigma (Kocyan & Endress, 2001a).
It seems likely that even this impressive, graduallyaccrued suite of morphological innovations provided

434 Paula J. Rudall and Richard M. Bateman
Fig. 14. Diagram of orchid subfamily relationships based on molecular topology achieved by combined analysis ofthe plastid genes rbcL, atpB and psaB (K. Cameron, personal communication, 2001), in which the A1-monandrousorchids are diphyletic (i.e. with Cypripedioideae nested within them), assuming the same outgroups as in Fig. 13.Non-molecular characters are optimised as indicated. 1¯ suppression of adaxial stamens (A2, A3, a3), 2¯ (resu-pination), 3¯ (labellum), 4¯dust seeds, 5¯ germinatory mycoheterotrophy, 6¯ loss of stamens a1 and a2, 7¯(pollinaria), 8¯ (spur). Parentheses indicate character state not present in all taxa. Arrowed text in italics indicateseither syndromes (suites of linked characters, e.g. zygomorphy) or apomorphic tendencies. Asterisk indicates characterpresent in derived taxa only.
a useful foundation, but not a complete causalexplanation, for the primary orchid radiation; theyare at best pre-adaptations for the remarkable speciesrichness reflecting secondary radiations in the de-rived subfamilies Orchidoideae and Epidendroideae.Most members of these families possess pollinaria : amore sophisticated pollination package consisting ofone or more pollinia subtended by a caudicle and}orstipe, often terminating in an adhesive viscidial disc.Interestingly, these families are also dominated byspecies possessing labella that are often heavilymarked and}or bear spurs, perhaps prompting amyriad of co-evolutionary relationships as animalstransported pollinaria in response to a potentadmixture of various forms of genuine reward ordeceit. The best potential test of this hypothesis liesin the fossil record, but evidence of dominantpollinators such as hymenopterans is scanty, andthat of Orchidaceae almost non-existent (e.g. Mehl,1986).
Secondary morphological reversions (‘atavisms’),of the kind documented above as terata, allow some
further insights into function. Endress (1992, 1994)and Donoghue, Richard & Baum (1998) concludedthat in Lamiales (formerly called Scrophulariales),which have predominantly zygomorphic flowerswith only four or fewer stamens expressed, occasional‘peloric ’ species with five-staminate flowers arosefrom zygomorphic ancestors. In orchids, there is noevidence that type A peloria has ever led to bona fide
speciation, presumably because having three well-differentiated labella strongly reduces the attractionof the flower to pollinators. In contrast, types B andC peloria and pseudopeloria have on several occa-sions led to speciation, perhaps because they offer atleast some prospect of pollinator switching, as seemslikely in the origin via suspected type C peloria of therelatively species-rich genus Thelymitra. However,studies of other taxa originating by floral simplifica-tion, such as the pseudopeloric Nigritella, suggestthat polyploidy and}or autogamy are often neededto maintain such lineages. This is not surprising,given the presumed loss of landing (but notnecessarily attractant) function of the labellum of

435Synorganisation, zygomorphy and heterotopy in floral evolution
Nigritella following its loss of resupination. However,the loss of resupination in the erect Australian diuridgenus Prasophyllum does not impair either theattractant or landing roles of the labellum; cephalicpseudocopulation merely proceeds with the malethynnine wasp oriented head-downward. Indeed,allogamous orchids with erect inflorescences offlowers that possess labella but have either no torsionof the ovary or 360° torsion, and thus lackresupination, offer a viable model system to test therelative importance of the attractant and landingroles of the labellum, and the viability of pollinatorswitching.
XIII. FUTURE DIRECTIONS
(1) Static and dynamic descriptions offlowers
We conclude that the terminology used to describeangiosperm flowers urgently requires clarification.For example, as Endress (1992) pointed out, theterm ‘whorl ’ is in itself misleading, since it is oftentaken to mean all the organs of a particular type (e.g.stamens or petals), but sometimes there are twodistinct whorls of an organ (e.g. many monocotshave two whorls of stamens). In particular, asdiscussed in Section III 3, the long-established usesof terms relating to floral symmetry by plantmorphologists need reconciling with more recentadaptations of those terms by developmental geneti-cists. Floral symmetry planes should relate to allfloral whorls (including stamens and carpels) ratherthan just the calyx and corolla as in many currentfloral descriptions. Floral diagrams and floral for-mulae (Section III 1; Fig. 4) are useful in thisrespect. A possible solution would be to include floralformulae with the number of symmetry planes ineach whorl as superscripts. For example, Cypripedium
would then be K3$ C3" A0 [a2" G(−$)$], in which the
calyx whorl (K) has three planes of symmetry andthe perianth whorl (C) has one plane of symmetry.However, even this more precise notation is com-plicated by several factors ; for example, differentaestivation patterns may confer different patterns ofasymmetry (Endress, 2001a), and floral organarrangement may be interpreted differently bydifferent authors (e.g. for a summary of differentinterpretations of the origin of the stamens inArabidopsis see Endress, 1992).
With regard to describing morphogenetic transi-tions between putative ancestors and descendants,valuable upgraded terminologies are available for
heterotopy (Baum & Donoghue, 2002) and hetero-chrony (e.g. Zelditch & Fink, 1996). Continuedstudy of peloria s.s. in angiosperms will undoubtedlyyield much of value, but greater attention couldcertainly be paid to the more subtle, yet perhapsmore evolutionarily viable, transitions here aggre-gated under the term pseudopeloria (Bateman &DiMichele, 2002). These have not yet been meaning-fully explored in groups outside the orchid familywith zygomorphic flowers. Further detailed surveysof the frequency of specific types of profoundmorphogenetic transition that occur in nature indifferent plant groups could usefully delineateevolutionary–developmental constraints on the po-tentially vast array of permutations of organs, andthus valuably focus expensive, time-consuming lab-oratory investigations.
We recognise that the tripartite classifications ofboth peloria and pseudopeloria used here, and themost recent manifestations of the terminologiesassociated with heterotopy and heterochrony, retaina degree of typology. As befits a rapidly developingdiscipline, they will require further revision as moreis learned of the underlying causes of morphogenesis.
(2) Orchid evolution
In attempting to summarise current knowledge ofthe floral evolution of orchids and their closerelatives, it has become clear that much remains tobe learned. The phylogenetic ambiguities among thesubfamilies within Orchidaceae, and among thedozen or so families widely viewed as most closelyrelated to Orchidaceae, may reflect genuinely rapid,deep radiations that are by definition difficult toresolve, but greater phylogenetic resolution will beneeded if the morphology of the hypothetical‘ancestral orchid’ is to be reconstructed with greaterconviction. Given that morphological data areespecially effective in cases of deep, rapid radiations(Bateman, 1999), a combined morphological andmolecular programme is clearly desirable, encompas-sing not only the Orchidaceae but also some lessthoroughly investigated allied taxa (cf. Rudall, 2002,in press). More comparative data are required onfloral anatomy in the small but apparently highlyvariable genus Pauridia (Hypoxidaceae) and othergenera of Corsiaceae (Corsiopsis and Arachnitis).
Intensified studies of the evolutionary–develop-mental genetics of Orchidaceae are also needed,preferably focusing on carefully selected, phylogene-tically relevant ‘model ’ taxa. Species known tofrequently generate terata would also be advan-

436 Paula J. Rudall and Richard M. Bateman
tageous, allowing both genetic experimentation andmore careful description encompassing all four floralwhorls with equal rigour. Such studies will elucidatethe genetic underpinnings of the evolutionarilypivotal phenomena of adnation, heterochrony andheterotopy (including homeosis), and their expres-sion in synorganisation and floral symmetry.
The function of the morphological products ofthese processes also merits more careful investigation.The probable independent origins of the gyno-stemium, the labellum and resupination in severalnon-orchid lineages, including those discussed here,increase the chances of reaching justifiable generali-sations.
Lastly, we need to monitor the products ofevolutionary–developmental transitions in the field.Thus far, ‘experimental speciation’ has receivedwoefully little attention (cf. Glover & Martin, 2002).It is now clear that profound phenotypic changes,each reflecting a minute genotypic change, canoccur over a single generation (Bateman, 1994;Bateman & DiMichele, 1994, 2002), but the long-term viability of those ‘prospecies ’ in the wild (andthe most appropriate taxonomic treatment of them)remain highly controversial. We need a ‘NewEcology’ to monitor the progress of evolutionarynovelties in the real world. This can then be relatedto the ‘New Morphology’, a genuinely integrativeapproach to understanding morphogenesis thatproceeds from explicit descriptions of static mor-phology through explicit descriptions of morpho-logical transitions during evolution to the underlyinggenetic}epigenetic causes of those transitions.
XIV. CONCLUSIONS
(1) A gynostemium, comprising stamen filamentsadnate to a syncarpous style, occurs in only threegroups of monocots, all of them epigynous lilioids :Orchidaceae (Asparagales), Pauridia (Hypoxi-daceae: Asparagales) and Corsia (Corsiaceae: Lili-ales). A new term, hyper-epigyny, is proposed todescribe stamen adnation to the style, which may bea product of over-expression of genes controllingepigyny.
(2) Current evidence indicates that there are atleast two key genes or sets of genes controlling floraldevelopment in lilioid monocots ; at least one isresponsible for hyper-epigyny, and another controlsadaxial stamen suppression and adaxial labellumformation in orchids. Flowers of Pauridia are actino-morphic in all three whorls, whereas those of Corsia
and most orchids have a bilaterally symmetrical
perianth with a well-differentiated labellum. InCorsia the labellum is formed from the outer mediantepal (sepal), whereas in orchids it is formed from theinner median tepal (petal) and is developmentallyadaxial (but positionally abaxial in orchids withresupinate flowers). In orchids zygomorphy is alsoexpressed in the stamen whorls, in contrast to Corsia.In Pauridia a complete stamen whorl is suppressed,but the ‘ lost ’ outer whorl is fused to the style. If, asseems likely, hyper-epigyny preceded zygomorphyin orchid evolution, then the flowers of Pauridia mayclosely resemble those of the immediate ancestors ofOrchidaceae.
(3) Naturally occurring floral terata in orchidsare classified into three contrasting modes of peloriaand of pseudopeloria, respectively. Dynamic evolu-tionary transitions between floral morphs are assign-ed to recently revised concepts of heterotopy(including homeosis) and heterochrony. Lateralheterotopy is apparently more frequent than acro-petal heterotopy, and full basipetal heterotopy doesnot occur. Pseudopeloria is more likely to generate aradically altered yet functional perianth, but is alsomore likely to cause acropetal modification of thegynostemium. Terata usefully delimit the boundariesof developmental potential in specific lineages, anddemonstrate the potential for profound, instan-taneous phenotypic shifts during evolution.
(4) The initial radiation in Orchidaceae can beattributed to the combination of hyper-epigyny,floral zygomorphy and resupination, but the laterradiations at lower taxonomic levels that generatedthe remarkable species richness of subfamilies Orchi-doideae and Epidendroideae are more likely toreflect more subtle innovations that directly in-fluence pollinator specificity, such as the devel-opment of stalked pollinaria and heavily markedand}or spur-bearing labella.
XV. ACKNOWLEDGEMENTS
We are grateful to Alison Eastman (Imperial College,London) for technical assistance during her undergrad-uate placement year at RBG Kew, Lucille McCook(University of Mississippi, USA) for much helpfuldiscussion during an early precursor to this project, andQuentin Cronk (Royal Botanic Garden Edinburgh),Peter Endress (University of Zurich) and Mark Chase(Royal Botanic Gardens, Kew) for critically reviewing themanuscript. D. M. Turner Ettlinger, A. J. Hughes and R.Bush generously loaned photographs, and many nativeorchid enthusiasts, most recently the membership of theUK Hardy Orchid Society, kindly drew our attention tounusual floral morphs.

437Synorganisation, zygomorphy and heterotopy in floral evolution
We dedicate this paper to European orchid expertDerek Turner Ettlinger (1924–2001) in memory of hisconstant encouragement and considerable critical acu-men.
XVI. REFERENCES
A, P., G, S. J., O, G. F. & W, D. B. (1979).
Size and shape in ontogeny and phylogeny. Paleobiology 5,
296–317.
A, V. A. (1994). Cladistic relationships of the slipper
orchids (Cypripedioideae: Orchidaceae) from congruent
morphological and molecular data. Lindleyana 6, 115–132.
A, C. E. (1941). Some studies on the floral anatomy of
the Liliales. Unpublished thesis, Cornell University.
A, J. T. (1984). The relationships of the slipper orchids
(subfamily Cypripedioideae, Orchidaceae). Selbyana 7, 129–
247.
B, G. D.’A. (1990). A new species of Coelogyne Lindl.
from Hong Kong. Orchid Review 98 (1156), 37–41.
B, R. M. (1985). Peloria and pseudopeloria in British
orchids. Watsonia 15, 357–359.
B, R. M. (1994). Evolutionary–developmental change
in the growth architecture of fossil rhizomorphic lycopsids :
scenarios constructed on cladistic foundations. Biological
Reviews 69, 527–597.
B, R. M. (1999). Integrating molecular and morpho-
logical evidence for evolutionary radiations. In Molecular
Systematics and Plant Evolution (eds. P. M. Hollingsworth, R. M.
Bateman and R. J. Gornall), pp. 432–471. Taylor & Francis,
London.
B, R. M. (2001). Evolution and classification of Euro-
pean orchids : insights from molecular and morphological
characters. Journal EuropaX ischer Orchideen 33, 33–119.
B, R. M., C, P. R., DM, W. A., K,
P. R., R, N. P., S, T. & S, W. E. (1998). Early
evolution of land plants : phylogeny, physiology and ecology
of the primary terrestrial radiation. Annual Review of Ecology
and Systematics 29, 263–292.
B, R. M. & DM, W. A. (1994). Saltational
evolution of form in vascular plants : a neoGoldschmidtian
synthesis. In Shape and Form in Plants and Fungi (eds. D. S.
Ingram and A. Hudson), pp. 63–102. Linnean Society
Symposium Series 16. Academic Press, London.
B, R. M. & DM, W. A. (2002). Generating and
filtering major phenotypic novelties : neoGoldschmidtian
saltation revisited. In Developmental Genetics and Plant Evolution
(eds. Q. C. B. Cronk, R. M. Bateman and J. A. Hawkins), pp.
109–159. Taylor and Francis, London.
B, R. M., H, P. M., P, J., L,
Y. B., P, A. M. & C, M. W. (in press). Molecular
phylogenetics and evolution of the Orchidinae and selected
‘Habenariinae’ (Orchidaceae). Botanical Journal of the Linnean
Society.
B, R. M., P, A. M. & C, M. W. (1997).
Phylogenetics of subtribe Orchidinae (Orchidoideae, Orchi-
daceae) based on nuclear ITS sequences. 2. Infrageneric
relationships and taxonomic revision to achieve monophyly of
Orchis sensu stricto. Lindleyana 12, 113–141.
B, D. & D, M. J. (2002). Transference of function,
heterotopy and the evolution of plant development. In
Developmental Genetics and Plant Evolution (eds. Q. C. B. Cronk,
R. M. Batema and J. A. Hawkins), pp. 52–69. Taylor and
Francis, London.
B, P. & B B, P. (1986). Floral mimesis in
Thelymitra nuda (Orchidaceae). Plant Systematics and Evolution
151, 187–202.
B, P. & G, P. (2000). The diversity of
pollination mechanisms in the Iridaceae of southern Africa. In
Monocots : Systematics and Evolution (eds. K. L. Wilson and
D. A. Morrison), pp. 301–308. CSIRO, Collingwood,
Australia.
B, P. & M, E. A. (1977). The reproductive
phenology of Echeandia macrocarpa Greenm. (Liliaceae) with a
reexamination of the floral morphology. Bulletin of the Torrey
Botanical Club 104, 320–323.
B, L. G. (1959). Mineralogy: concepts, descriptions, determina-
tions. Freeman, San Francisco.
B! , M. (ed.) (1998). Les OrchideU es de France, Belgique et
Luxembourg. Parthe!nope, Paris.
B, C. C. (2001). Thelymitra : pollination. In Genera Orchi-
dacearum 2: Orchidoideae Part 1. (eds. A. M. Pridgeon, P. J.
Cribb, M. W. Chase and F. N. Rasmussen), pp. 208–213.
Oxford University Press, Oxford.
B, R. (1833). On the organs and mode of fecundation in
Orchideae and Asclepiadeae. Transactions of the Linnean Society
16, 685–745.
B-B, P. & B, P. (1985). Evolutionary
trends in the androecium of the Orchidaceae. Plant Systematics
and Evolution 149, 119–134.
B-B, P. & B, P. (1988). Floral evolution
and phylogeny in the tribe Thelymitreae (Orchidaceae:
Neottioideae). Plant Systematics and Evolution 159, 19–47.
C, L. R., R, P. J. & W, P. (2000). Floral
morphology and development in Dioscoreales. Feddes Reper-
torium 111, 189–230.
C, K. M. & C, M. W. (2000). Nuclear 18S rDNA
sequences of Orchidaceae confirm the subfamilial status and
circumscription of Vanilloideae. In Monocots : Systematics and
Evolution (eds. K. L. Wilson and D. A. Morrison), pp.
457–464. CSIRO, Collingwood, Australia.
C, K. M., C, M. W., W, W. M., K,
P. J., J, D. C., A, V. A., Y, T., H,
H. G. & G, D. H. (1999). A phylogenetic analysis of
the Orchidaceae: evidence from rbcL nucleotide sequences.
American Journal of Botany 86, 208–224.
C, S. (1969). Towards acceptable evolutionary inter-
pretations of floral anatomy. Phytomorphology 19, 332–363.
C, M. W. (1999). Molecular systematics, parsimony, and
orchid classification. In Genera Orchidacearum. 1. General
Introduction, Apostasioideae, Cypripedioideae (eds. A. M. Pridgeon,
P. J. Cribb, M. W. Chase and F. N. Rasmussen), pp. 81–88.
Oxford University Press, Oxford.
C, M. W. (2001). The origin and biogeography of
Orchidaceae. In Genera Orchidacearum 2. Orchidoideae, Part 1
(eds. A. M. Pridgeon, P. L. Cribb, M. W. Chase and F. N.
Rasmussen), pp. 1–5. Oxford University Press, Oxford.
C, M. W., D, M. R., H, H. G., C, J. G.,
C, A. V., E, L. E., H, J., F, M. F.,
C, L. R., C, K. M. & H, S. (1995).
Molecular systematics of Lilianae. In Monocotyledons: Systema-
tics and Evolution (eds. P. J. Rudall, P. J. Cribb, D. F. Cutler
and C. J. Humphries), pp. 109–137. Royal Botanic Gardens,
Kew.
C, M. W., S, D. S., S, P. S., R, P. J., F,

438 Paula J. Rudall and Richard M. Bateman
M. F., H, W. H., S, S., J, J., M, M.,
K, P. J., G, T. J., S, K. J. & P, J. C.
(2000). Higher-level systematics of the monocotyledons : an
assessment of current knowledge and a new classification. In
Monocots : Systematics and Evolution (eds. K. L. Wilson and
D. A. Morrison), pp. 3–16. CSIRO, Collingwood, Australia.
C, S.-C. (1965). A primitive new orchid genus Tangtsinia
and its meaning in phylogeny. Acta Phytotaxonomica Sinica 10,
193–207.
C, S.-C. (1978). Sinorchis – a primitive new genus of
Orchidaceae from China. Acta Phytotaxonomica Sinica 16, 82–85.
C, S.-C. (1979). On Diplandrorchis, a very primitive and
phylogenetically significant new genus of Orchidaceae. Acta
Phytotaxonomica Sinica 17, 1–6.
C, S.-C. (1982). The origin and early differentiation of the
Orchidaceae. Acta Phytotaxonomica Sinica 20, 1–20.
C, S.-C. & T, Z.-H. (1987). Eria medogensis, a probably
peloric form of Eria coronaria, with a discussion on peloria in
Orchidaceae. Acta Phytotaxonomica Sinica 25, 329–339.
C, S.-C., T, Z.-H. & L, Y.-B. (1999). Native Orchids of
China in Colour. Science Press, Beijing.
C, H. L., M$ , M. & C, Q. C. B. (2000).
Diversity of cycloidea-like genes in Gesneriaceae in relation to
floral symmetry. Annals of Botany 86, 167–176.
C, M. A. (2001). Diurideae: phylogeny. In Genera
Orchidacearum 2. Orchidoideae, Part 1 (eds. A. M. Pridgeon, P. L.
Cribb, M. W. Chase and F. N. Rasmussen), pp. 61–63.
Oxford University Press, Oxford.
C, E. S. (1991). The role of homeotic genes in flower
development and evolution. Annual Reviews of Plant Physiology
and Plant Molecular Biology 42, 241–279.
C, E. S. (1996). Floral symmetry. EMBO Journal 15,
6777–6788.
C, E. S. (1999). The Art of the Genes: How Organisms Make
Themselves. Oxford University Press, Oxford.
C, E. S. & M, E. M. (1991). The war of the
whorls : genetic interactions controlling flower development.
Nature 353, 31–37.
C, E. J. H. (1958). Transference of function. Botanical
Journal of the Linnean Society 56, 33–40.
C, A. V., P, A. M., A, V. A. & C, M. W.
(1997). Phylogenetics of the slipper orchids (Cypripedioideae:
Orchidaceae) : nuclear rDNA sequences. Plant Systematics and
Evolution 208, 197–223.
C, P. L. (1999). Morphology. In Genera Orchidacearum. 1.
General Introduction, Apostasioideae, Cypripedioideae (eds. A. M.
Pridgeon, P. L. Cribb, M. W. Chase and F. N. Rasmussen),
pp. 13–23. Oxford University Press, Oxford.
C, Q. C. B. (2001). Plant evolution and development in a
post-genomic context. Nature Reviews Genetics 2, 607–619.
C, Q. C. B., B, R. M. & H, J. A. .
(2002). Developmental Genetics and Plant Evolution. Taylor and
Francis, London.
C, Q. C. B. & M$ , M. (1997). Genetics of floral
symmetry revealed. Trends in Ecology and Evolution 12, 85–86.
C, P. (2002). Role of TCP genes in the evolution of
morphological characters in angiosperms. In Developmental
Genetics and Plant Evolution (eds. Q. C. B. Cronk, R. M.
Bateman and J. A. Hawkins), pp. 247–266. Taylor and
Francis, London.
C, P., V, C. & C, E. (1999). An epigenetic
mutation responsible for natural variation in floral symmetry.
Nature 401, 157–161.
D, R. M. T., C, H. T. & Y, P. F. (1985).
The Families of the Monocotyledons. Springer-Verlag, Berlin.
D, C. (1862). On the Various Contrivances by which British and
Foreign Orchids are Fertilised by Insects. Murray, London.
D, C. (1877). On the Various Contrivances by which British and
Foreign Orchids are Fertilised by Insects. (2nd edn). Murray,
London.
D V, E. F. (1969). Monograph of the tribe Apostasieae
(Orchidaceae). Blumea 17, 313–350.
D V, M. P. (1949). The development of the ovule and the
seed in the Hypoxideae. II. The genera Pauridia Harv. and
Forbesia Ecklon. Journal of South African Botany 15, 13–22.
D, P. (2001). Guide des OrchideU es d’Europe, d’Afrique du
Nord at du Proche-Orient (2nd edn). Delachaux & Niestle! , Paris.
D, P. K. & E, P. K. . (1999). Development,
function and evolution of symmetry in plants. International
Journal of Plant Science 160, S1–S166.
D, J., S, A. & H, L. (1997). The evolution of
apical dominance in maize. Nature 386, 485–488.
D, M. J., R, H. R. & B, D. (1998).
Phylogeny and the evolution of flower symmetry in the
Asteridae. Trends in Plant Science 3, 311–317.
D, E. J. P., P, A. M., K, P., L, H. P.,
K, H. & C, M. W. (1999). Molecular phylogene-
tics of Diseae (Orchidaceae) : a contribution from nuclear
ribosomal ITS sequences. American Journal of Botany 86,
887–899.
D, R. L. (1993). Phylogeny and Classification of the Orchid
Family. Cambridge University Press, Cambridge.
D, R. L. & W, L. O. (1975). Proposal for
conservation of the generic name Phragmipedium Rolfe. Taxon
24, 691.
E, A. W. (1875). BluX thendiagramme. Engelmann, Leipzig.
E, P. K. (1992). Evolution and floral diversity : the
phylogenetic surroundings of Arabidopsis and Antirrhinum.
International Journal of Plant Science 153, S106–S122.
E, P. K. (1994). Diversity and Evolutionary Biology of Tropical
Flowers. Cambridge University Press, Cambridge.
E, P. K. (1996). Diversity and Evolutionary Trends in
Angiosperm Anthers. In The Anther. Form, function and phylogeny
(eds. W. G. D’Arcy and R. C. Keating), pp. 92–110. Cam-
bridge University Press, Cambridge.
E, P. K. (1997). Evolutionary biology of flowers:
prospects for the next century. In Evolution and Diversification of
Land Plants (eds. K. Iwatsuki and P. H. Raven), pp. 99–119.
Springer-Verlag, Tokyo.
E, P. K. (1999). Symmetry in flowers : diversity and
evolution. International Journal of Plant Science 160, S3–S23.
E, P. K. (2001a). Evolution of floral symmetry. Current
Opinion in Plant Biology 4, 86–91.
E, P. K. (2001b). Origins of flower morphology. Journal
of Experimental Zoology 291, 105–115.
E, P. K. & B, P. (2000). A revised classification of
the Apocynaceae s.l.. Botanical Review 66, 1–56.
E, G. (1952). Pollen Morphology and Plant Taxonomy.
Angiosperms. Stockholm: Almqvist & Wiksell.
E, R. (1958). Triggerplants. P. Brokensha, Perth,
Australia.
E, R. & A, J. (1994). Resupination. In Orchid
Biology; Reviews and Perspectives VII (ed. J. Arditti), pp.
135–188. John Wiley & Sons, New York.
E, D. H. (1992). A preliminary classification of evol-
utionary radiations. Historical Biology 6, 133–147.

439Synorganisation, zygomorphy and heterotopy in floral evolution
E, D. M. T. (1998). Illustrations of British and Irish
Orchids. Published by the author, Royden Cottage, Dorking,
Surrey, U.K.
F, M. F., R, P. J., S, S., S, K. L.,
B, A. Y., R, G., Q-Z, F., H,
W. P., J, J., H, W. J., C, J. G. & C,
M. W. (2000). Phylogenetic studies of Asparagales based on
four plastid DNA loci. In Monocots : Systematics and Evolution
(eds. K. L. Wilson and D. A. Morrison), pp. 360–371.
CSIRO, Collingwood, Australia.
F, W. L. (1982). The conceptual relationship between
ontogeny and phylogeny. Paleobiology 8, 254–264.
F, J. V. & R, F. (1999). What does
morphology tell us about orchid relationships? A cladistic
analysis. American Journal of Botany 86, 225–248.
F, J. V., S, D. M. & C, M. W. (2000).
Mitochondrial DNA and relationships in the Orchidaceae. In
Monocots : Systematics and Evolution (eds. K. L. Wilson and
D. A. Morrison), pp. 421–429. CSIRO, Collingwood,
Australia.
F, M. W. (1987). Common-is-primitive : a partial
validation by tree counting. Systematic Botany 12, 217–237.
F, M. W. (2002). The Mostly Male theory of flower
origins : summary and update regarding the Jurassic pterido-
sperm Pteroma. In Developmental Genetics and Plant Evolution (eds.
Q. C. B. Cronk, R. M. Bateman and J. A. Hawkins), pp.
85–108. Taylor and Francis, London.
G, L. A. (1960). On the origin of the Orchidaceae.
Botanical Museum Leaflets, Harvard University 19, 57–95.
G, L. A. (1972). On the origin of the Orchidaceae II.
Journal of the Arnold Arboretum 53, 202–215.
G, L. A. (1979). The genus Phragmipedium. Orchid Digest 43,
133–148.
G, A. C. M., C, P., C, E. S. & A, R. J.
(2002). Making rays in the Asteraceae: genetics and evolution
of radiate versus discoid flower heads. In Developmental Genetics
and Plant Evolution (eds. Q. C. B. Cronk, R. M. Bateman and
J. A. Hawkins), pp. 233–246. Taylor and Francis, London.
G, B. J. & M, C. (2002). Evolution of adaptive petal
cell morphology. In Developmental Genetics and Plant Evolution
(eds. Q. C. B. Cronk, R. M. Bateman and J. A. Hawkins), pp.
160–172. Taylor & Francis, London.
G, R. (1940). The Material Basis of Evolution. Yale
University Press, New Haven.
G! , F. & S, D. W. (2000). Gynostemium
development in Aristolochia (Aristolochiaceae). Botanische Jahr-
buX cher 122, 249–291.
G, S. J. (1977). Ontogeny and Phylogeny. Belknap, Cam-
bridge, Mass.
G, B. (2000). Reorganising the Orchid Genus Coelogyne:
A Phylogenetic Classification Based on Morphology and Molecules.
Nationaal Herbarium Nederland, Leiden.
G, L. (1883). Recherches sur le de! veloppement de
l’anthe' re et du pollen des Orchidace! es. Annales des Sciences
Naturelles, Botanique, 6e seU rie, 14, 26–44.
G, M. H. G. & A, V. A. (1999). Inferior ovaries
and angiosperm diversification. In Molecular Systematics and
Plant Evolution (eds. P. M. Hollingsworth, R. M. Bateman and
R. J. Gornall), pp. 403–431. Taylor and Francis, London.
H, C. R. & S, D. W. (2000). Development of the
gametophytes, flower, and floral vasculature in Cochliostema
odoratissimum (Commelinaceae). Botanical Journal of the Linnean
Society 134, 131–157.
H! , M., K, E. & T, H. (2000). Evolution of
polyploids in the European orchid genus Nigritella : evidence
from allozyme data. Phyton, 40, 239–275.
H, P. M., B, R. M. & G, R. J.
. (1999). Molecular Systematics and Plant Evolution. Taylor
& Francis, London.
H, P. M., B, R. M., D, J. H.,
S, J. & T, M. (2001). Population genetic
structure of Epipactis (Orchidaceae). First International Orchid
Conservation Congress (Perth) Abstracts : 62.
H, F. (1990). Peloria in Dactylorhiza. BSBI News 55,
16–18.
H, H. (1969). Die Samenmerkmale und Verwandtschafts-
verhaltnisse der Liliifloren. Mitteilungen der Botanischen Staats-
sammlung MuX nchen 8, 219–538.
H, J. (1934). The Families of Flowering Plants. II.
Monocotyledons. Clarendon Press, Oxford.
I, A., B, M. & E, P. K. (2001). Gynoecium
diversity and systematics in basal monocots. Botanical Journal
of the Linnean Society 136, 1–65.
J, B. & F, S. (2002). Orchid flowers :
evolution and molecular development. In Developmental
Genetics and Plant Evolution (eds. Q. C. B. Cronk, R. M.
Bateman and J. A. Hawkins), pp. 206–219. Taylor and
Francis, London.
J, S. D. & E, T. J. (2000). The structure and
function of orchid pollinaria. In Pollen and Pollination (eds. A.
Dafni, M. Hesse and E. Pacini), pp. 244–269. Springer, Wien.
J, S. D., L, H. P. & S, K. E. (1998).
Phylogeny and radiation of pollination systems in Disa
(Orchidaceae). American Journal of Botany 85, 402–411.
J, D. L. (1988). Native Orchids of Australia. Reed, Chatswood
NSW, Australia.
K, C., H, C. S. & D, J. (1998). Manual of
Mineralogy (21st edn). Wiley, New York.
K, A. & E, P. K. (2001a). Floral structure and
development of Apostasia and Neuwiedia (Apostasioideae) and
their relationships to other Orchidaceae. International Journal
of Plant Science 162, 847–867.
K, A. & E, P. K. (2001b). Floral structure and
development and systematic aspects of some ‘ lower’ Aspara-
gales. Plant Systematics and Evolution 229, 187–216.
K, P. J., M, M., W, P. H., H, S. D.,
B, A. P., C, K. C. & C, M. C. (2001). A
phylogenetic analysis of Duirideae (Orchidaceae) based on
plastid DNA serquence data. American Journal of Botany 88,
1903–1914.
K, E. M. & I, V. F. (1999). Evolution of genetic
mechanisms controlling petal development. Nature 399,
144–148.
K, H. (1995). Bau und Funktion der Asclepiadaceenblu$ te.Phyton 35, 1–24.
K, H. (1987a). Developmental studies in orchid flowers
I : Epidendroid and vandoid species. Nordic Journal of Botany 7,
427–442.
K, H. (1987b). Developmental studies in orchid flowers
II : Orchidoid species. Nordic Journal of Botany 7, 443–451.
K, H. (1988). Developmental studies in orchid flowers
III : Neottioid species. Nordic Journal of Botany 8, 271–282.
K, H. (1996). Floral morphology and ontogeny in
subtribe Satyriinae (Orchidaceae). Flora 191, 9–28.
K, H. (1998). Floral ontogeny of orchids : a review.
BeitraX ge zur Biologie der Pflanzen 71, 45–100.

440 Paula J. Rudall and Richard M. Bateman
L, J. (1977). Wilde OrchideeeX n van Europa. Amsterdam.
Publisher unknown.
L, D. C. (1980). Orchids of Britain. Oxford University Press,
Oxford.
L, D. C. (2001). A new variant of Ophrys apifera in Britain.
BSBI News 88, 40–46.
L, R. G. (1909). A vegetative mutant, and the principle
of homeosis in plants. Botanical Gazette 47, 30–68.
L, P. & E, C. (1985). Ein Beitrag zur Blu$ tenentwick-
lung der Aristolochiaceen, einer Vermittlergruppe zu den
Monokotylen. Botanische JahrbuX cher 107, 343–368.
L, H. P. (1986). Notes on the phylogeny of the Orchidoi-
deae, with particular reference to the Diseae. Lindleyana 1,
51–64.
L, H. P. & K, H. (1999). Orchids of Southern Africa.
Balkema, Rotterdam.
L! , C. (1744). Dissertatio botanica de Peloria. Amoenitates
Academica, Uppsala.
L, C. (1975). The Native Orchids of the United States and Canada,
Excluding Florida. New York Botanical Garden, New York.
L, Z. X., W, M., L, C. S., Y, C. Y. & G, C. J.
(1993). Nucleotide sequence of a flower-specific MADS box
cDNA clone from an orchid. Plant Molecular Biology 23,
901–904.
L, D., C, R., V, C., C, L. & C, E.
(1996). Origin of floral asymmetry in Antirrhinum. Nature 383,
794–799.
L, D., C, R., C, L., V, C., C, J. &
C, E. (1999). Control of organ asymmetry in flowers of
Antirrhinum. Cell 99, 367–376.
M, P. W. (1910). Anwachsungen der Sepalen an das
Gynostemium von Orchideenblu$ ten. Annales du Jardin Bot-
anique de Buitenzorg, 3rd suppl., 1, 61–68, pls. IV, V.
M, T. W. (2000). Stellung von Blu$ ten und Aufblau von
Blu$ ten sta$ nden als systematisches Merkmal zur Gliederung
ausgewa$ hlter Monokotyledonen. Diplomarbeit (unpub-
lished), Faculty of Biology, Ruhr-Universita$ t Bochum.
MC, L. M. (1989). Systematics of Phragmipedium (Cypri-
pedioideae: Orchidaceae). Doctoral thesis, Cornell University,
Ithaca, NY.
MC, L. M. & B, R. M. (1990). Homeosis in orchid
flowers : a potential mechanism for saltational evolution.
American Journal of Botany (Abstracts) 77, 145.
MK, D. R. (1982). x Pseudanthera breadalbanensis McKean:
a new integeneric hybrid from Scotland. Watsonia 14, 129–131.
M, J. (1986). Die fossile Dokumentation der Orchideen.
Journal Berlein Naturwissenschaften Vereins, Wuppertal 39, 121–
133.
M, E. M., S, D. R. & B, J. L. (1989).
Abnormal flowers and pattern formation in floral devel-
opment. Development 106, 209–217.
M, J. D. & R, H. (1984). Stegmata in Orchi-
dales : character state distribution and polarity. Botanical
Journal of the Linnean Society 89, 53–76.
M, M., K, P. J. & C, M. W. (2000). Polyphyly
of mycoheterotrophic orchids and functional influences on
floral and molecular characters. In Monocots : Systematics and
Evolution (eds. K. L. Wilson and D. A. Morrison), pp.
441–448. CSIRO, Melbourne.
N, P. R., D, A. & G, M. (1998). Floral symmetry
and its role in plant-pollinator systems: terminology, dis-
tribution and hypotheses. Annual Reviews of Ecology and
Systematics 29, 345–373.
N, M. R. M. & W, C. C. (1995). Maximisation of
reproductive success by European Orchidaceae under condi-
tions of infrequent pollination. Protoplasma 187, 37–48.
N, M. R. M. & W, C. C. (1998). Fruit set, nectar
reward, and rarity in the Orchidaceae. American Journal of
Botany 85, 1657–1671.
N, C. & I, P. L. (1998). Corsiaceae. In Families and
Genera of Vascular Plants. II. Flowering plants–Monocotyledons–
Lilianae (ed. K. Kubitzki), pp. 198–201. Springer-Verlag,
Berlin.
N, E. (1965). Zur organophyletischen Natur des Orchi-
deenlabellums. Botanische JahrbuX cher 84, 175–214.
N, E. (1967). Das Orchideenlabellum, ein Homologon des
einfachen Medianen Petalums der Apostasiaceen oder ein
zusammengesetztes Organ? Botanische JahrbuX cher 87, 22–35.
N, G. D. & W, N. H. (1978). Pollen morphology
of the Cypripedioideae and the Apostasioideae (Orchi-
daceae). Selbyana 2, 169–182.
N, R. (2002). A phylogeny inferred from large ribosomal
subunit 26S rDNA gene sequences suggests that Burmanniales
are polyphyletic. Australian Systematic Botany 15, 1–10.
N, R. & U, L. E. (1996). Evolution in the
number and position of fertile anthers in Orchidaceae inferred
from ndhF chloroplast gene sequences. Lindleyana 11, 47–53.
P, F. W. (1948). The taxonomic significance of under-
standing of floral evolution. Brittonia 6, 301–308.
P, A. M., B, R. M., C, A. V., H, J. R.
& C, M. W. (1997). Phylogenetics of subtribe Orchidinae
(Orchidoideae, Orchidaceae) based on nuclear ITS se-
quences. 1. Intergeneric relationships and polyphyly of Orchis
sensu lato. Lindleyana 12, 89–109.
P, A. M., C, P. J., C, M. W. & R,
F. N. eds. (1999). Genera Orchidacearum. 1. General Introduction,
Apostasioideae, Cypripedioideae. Oxford University Press, Oxford.
P, A. M., C, P. J., C, M. W. & R,
F. N. eds. (2001). Genera Orchidacearum 2: Orchidoideae Part 1.
Oxford University Press, Oxford.
P, R. E., V-C, J. P., P, S. E., G-
, U. & L, S. J. (2000). Fiddlehead, a gene
required to suppress epidermal cell interactions in Arabidopsis,
encodes a putative lipid biosynthetic enzyme. Proceedings of the
National Academy of Sciences, USA 97, 1311–1316.
Q-Z, F., F, M. F., P, J. S. & C,
M. W. (1998). Molecular techniques employed in the
assessment of genetic diversity : a review focusing on orchid
conservation. Lindleyana 13, 259–283.
R, M. (1980). A shower of brilliant little stars. Veld & Flora
(Kirstenbosch) 66, 81–82.
R, V. S. (1969). The floral anatomy and relationship of the
rare Apostasias. Journal of the Indian Botanical Society 48,
374–386.
R, V. S. (1973). A further contribution to the floral anatomy
of the rare Apostasias : genus Neuwiedia. Journal of the Indian
Botanical Society 52, 65–71.
R, V. S. (1974). The relationship of the Apostasiaceae on the
basis of floral anatomy. Botanical Journal of the Linnean Society
68, 319–327.
R, F. N. (1981). The orchidaceous gynostemium. I.
Neottioid orchids. Copenhagen.
R, F. N. (1982). The gynostemium of the neottioid
orchids. Opera Botanica 65, 1–96.
R, F. N. (1985). The gynostemium of Bulbophyllum

441Synorganisation, zygomorphy and heterotopy in floral evolution
ecornutum (J. J. Smith) J. J. Smith (Orchidaceae). Botanical
Journal of the Linnean Society 91, 447–456.
R, J. A. & W, J. D. B. (2001). Significance of
epidermal fusion and intercalary growth for angiosperm
evolution. Trends in Plant Science 6, 111–113.
R, A. (1823). Notice sur une monstrosite! remarquable
des fleurs de l’Orchis latifolia L. Memoires de la societeU d’Histoire
Naturelles de Paris 1, 202–209.
R$ , T. (1986). Morphologische, embryologische und
systematische Untersuchungen an Burmanniaceae und Corsi-
aceae. Dissertationes Botanicae 92. J. Cramer, Berlin.
R, P. J. (2002). Homologies of inferior ovaries and septal
nectaries in monocotyledons. International Journal of Plant
Sciences 163, 261–276.
R, P. J. (in press). Unique structures and iterative themes
in Asparagales : a revised morphological analysis considered
in relation to floral evolution. Botanical Review.
R, P. J. & C, M. W. (1996). Systematics of Xanthor-
rhoeaceae sensu lato : evidence for polyphyly. Telopea 6,
629–647.
R, P. J., C, M. W., C, D. F., R, J. &
B, A. (1998). Anatomical and molecular systematics of
Asteliaceae and Hypoxidaceae. Botanical Journal of the Linnean
Society 127, 1–42.
R, P. J. & E, A. (2002). The questionable affinities
of Corsia (Corsiaceae) : evidence from floral anatomy and
pollen morphology. Botanical Journal of the Linnean Society, 138,
315–324.
R, P. J., F, C. A., C, M. W. & F, M. F.
(1997). Microsporogenesis and pollen sulcus type in Aspara-
gales (Lilianae). Canadian Journal of Botany 75, 408–430.
R, P. J. & G, P. (2001). Floral anatomy and
systematic position of Diplarrhena (Iridaceae): a new tribe
Diplarrheneae. Annali di Botanica (Roma) n. s. 58, 59–66.
R, P. J., H, W. P., F, C. A., C, J. G.,
K, G., S, K. L. & C, M. W. (2000). Consider
the lilies – systematics of Liliales. In Monocots : Systematics and
Evolution (eds. K. L. Wilson and D. A. Morrison), pp.
347–357. CSIRO, Collingwood, Australia.
S, R. (1988). Homeosis in plants. American Journal of
Botany 75, 1606–1617.
S, W. (2001). U> ber x Gymnoglossum jacksonii (Dr.) Rolfe
und exklusive formen bei Gymnadenia (sowie Anacamptis und
Dactylorrhiza). Journal EuropaX ischer Orchideen 33, 1067–1078.
S, M. G. (1983). Pollen ultrastructure of the Haemodor-
aceae and its taxonomic significance. Grana 22, 79–103.
S, M. G. (1985). Pollen ultrastructure of the Tecophilaea-
ceae. Grana 24, 77–92.
S, N. (2000). The response of epidermal cells to contact.
Trends in Plant Science 5, 233–234.
S, D. E., S, P. M. & D, J. J. (1997). Molecular
Systematics of Plants (2nd edn.). Chapman & Hall, London.
S, J., H, P. M., B, R. M.,
D, J. H., L, M. H. S., MC, M. & T,
M. C. (2001). Partitioning and diversity of nuclear and
organelle markers in native and introduced populations of
Epipactis helleborine (Orchidaceae). American Journal of Botany,
88, 1409–1418.
S, B. G. L. (1948). Vascular anatomy of orchid flowers.
Botanical Museum Leaflets, Harvard University 13, 61–95.
S, D. L. & R, P. (2000). Gynostemia
Orchidalium 1. Acta Botanica Fennica 169, 1–380.
T, H. & K, E. (1998). Etiam atque etiam – Nigritella
versus Gymnadenia : Neukombinationen und Gymnadenia dolomi-
tensis spec. nova (Orchidaceae: Orchideae). Phyton (Horn) 38,
220–224.
T, G. (2000). Evolutionary developmental genetics of
floral symmetry: the revealing power of Linnaeus’ monstrous
flower. Bioessays 22, 209–213.
T, G., B, A., W, K. U., M$ , T.,
K, G. & S, H. (2002). How the land plants
learned their floral ABCs: the role of MADS-box genes in the
evolutionary origin of flowers. In Developmental Genetics and
Plant Evolution (eds. Q. C. B. Cronk, R. M. Bateman and J. A.
Hawkins), pp. 173–205. Taylor and Francis, London.
T, M. F. (1978). Studies in the Hypoxidaceae. II.
Floral morphology and anatomy. Bothalia 12, 429–435.
T, M. F. (1979). Studies in the Hypoxidaceae III. The
genus Pauridia. Bothalia 12, 621–625.
V, N. P. & M, S. K. (1976). An incipient
gynostemium in the Aloineae (Liliaceae). Current Science 45,
112–113.
V P, L. & D, C. H. (1966). Orchid Flowers: Their
Pollination and Evolution. University of Miami Press.
V R, P. (1972). Sertulum Papuanum 17. Corsiaceae of
New Guinea and surrounding areas. Webbia 27, 223–255.
V, P. (1966). The system of the Orchidales. Acta
Botanica Neerlandica 15, 224–253.
W-L, J. & H, L. D. (2000). The evolution of
staminodes in angiosperms: patterns of stamen reduction, loss,
and functional re-invention. American Journal of Botany 87,
1367–1384.
W, F. (1989). Morphology of Flowers and Inflorescences.
(Translated by R. J. Pankhurst). Cambridge University Press,
Cambridge.
W, C. L. (1982). Vestigial structures and the flower.
American Journal of Botany 69, 1356–1365.
W, J. J. (1999). Apostasioideae. In Genera Orchidacearum (eds.
A. M. Pridgeon, P. J. Cribb, M. W. Chase and F. N.
Rasmussen), pp. 94–104. Oxford University Press, Oxford.
W, J. J. & C, P. L. (2001). Aceratorchis. In Genera
Orchidacearum 2. Orchidoideae, Part 1 (eds. A. M. Pridgeon, P. J.
Cribb, M. W. Chase and F. N. Rasmussen), pp. 244–245.
Oxford University Press, Oxford.
W, W. C. (1916). Principles of Plant Teratology. The Ray
Society, London.
Y, A., W, E., H, P., H, C.,
W, K. & S, H. (1999). Characterization of the
FIDDLEHEAD gene of Arabidopsis reveals a link between
adhesion response and cell differentiation in the epidermis.
Plant Cell 11, 2187–2201.
Y, D. P. (1952). Studies in the British Epipactis, III, IV.
Watsonia 2, 253–276.
Y, H. & G, C. J. (2000). Identification and characterization
of three orchid MADS-box genes of the AP1}AGL9 subfamily
during floral transition. Plant Physiology 123, 1325–1336.
Z, M. L. & F, W. L. (1996). Heterochrony and
heterotopy: stability and innovation in the evolution of form.
Paleobiology 22, 247–250.
Z, D., S, R. M. K. & H, C. M. (1999). Corsiopsis
chinensis gen. et sp. nov. (Corsiaceae) : first record of the family
in Asia. Systematic Botany 24, 311–314.
Z, W. B. (1994). Guide to Flowering Plant Families.
University of North Carolina Press, Chapel Hill.




















