
Role of phosphatidylglycerol in the function and assembly of photosystem II reaction center, studied...
11
Role of phosphatidylglycerol in the function and assembly of Photosystem II reaction center, studied in a cdsA-inactivated PAL mutant strain of Synechocystis sp. PCC6803 that lacks phycobilisomes Hajnalka Laczkó-Dobos a, 1 , Bettina Ughy a, 1 , Szilvia Z. Tóth a , Josef Komenda b,c , Ottó Zsiros a , Ildikó Domonkos a , Árpád Párducz d , Balázs Bogos a , Masayuki Komura e , Shigeru Itoh e , Zoltán Gombos a, ⁎ a Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H–6701 Szeged, Hungary b Institute of Microbiology, Academy of Sciences, Opatovický mlýn, 37981, Třeboň, Czech Republic c Institute of Physical Biology, University of South Bohemia, Zámek 136, 37333 Nové Hrady, Czech Republic d Institute of Biophysics, Biological Research Center, Hungarian Academy of Sciences, H–6701 Szeged, Hungary e Division of Material Science (Physics), Nagoya University, Japan ABSTRACT ARTICLE INFO Article history: Received 14 February 2008 Received in revised form 4 June 2008 Accepted 4 June 2008 Available online 10 June 2008 Keywords: Synechocystis sp. PCC6803 Phosphatidylglycerol Photosystem II Thermoluminescence OJIP transient Cell division To analyze the role of phosphatidylglycerol (PG) in photosynthetic membranes of cyanobacteria we used two mutants of Synechocystis sp. PCC6803: the PAL mutant which has no phycobilisomes and shows a high PSII/ PSI ratio, and a mutant derived from it by inactivating its cdsA gene encoding cytidine 5'-diphosphate diacylglycerol synthase, a key enzyme in PG synthesis. In a medium supplemented with PG the PAL/ΔcdsA mutant cells grew photoautotrophically. Depletion of PG in the medium resulted (a) in an arrest of cell growth and division, (b) in a slowdown of electron transfer from the acceptor Q A to Q B in PSII and (c) in a modification of chlorophyll fluorescence curve. The depletion of PG affected neither the redox levels of Q A nor the S 2 state of the oxygen-evolving manganese complex, as indicated by thermoluminescence studies. Two-dimensional PAGE showed that in the absence of PG (a) the PSII dimer was decomposed into monomers, and (b) the CP43 protein was detached from a major part of the PSII core complex. [ 35 S]-methionine labeling confirmed that PG depletion did not block de novo synthesis of the PSII proteins. We conclude that PG is required for the binding of CP43 within the PSII core complex. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Phosphatidylglycerol (PG) is an anionic phospholipid that is required for the formation and function of thylakoid membranes in cyanobacteria and plants, see a review by Wada and Murata [1]. Depletion of PG modifies the function of both photosystem (PS) I and PSII reaction centers (RCs) [2–7]. The structure of PSI reaction center core complex (RCC) of Ther- mosynechococcus elongatus, as revealed by X-ray crystallography [8– 10] showed four lipid molecules inserted into the lumenal side of the PsaA and PsaB polypeptides. Two PG molecules are bound to proteins at the peripheral moiety of the RC. One PG and one monogalactosyldiacylglycerol (MGDG) were found in the central core moiety in the vicinity of two electron acceptor phylloquinone molecules (A 1 and A 1 ′) [11]. PG molecules were also assumed to exist at the center of the PSI trimer in association with the small PsaL subunits. This suggestion was based on the observation that in a PG- depleted mutant of Synechocystis sp. PCC6803, in which a pgsA gene required for PG biosynthesis was inactivated [4] the amount of PSI trimers decreased. A recent study of the structure of PSII RCC in T. elongatus [12] reported the presence of several lipid molecules at the interface between the subunit proteins. One of these was a PG molecule located between the D1 and CP43 proteins in the postulated pathway of Q B - quinone exchange. Recent biochemical studies have indicated the binding of a large number of lipids to purified PSII RC [5]. For instance, 6–7 molecules of PG were found to bind to T. elongatus PSII and a smaller number of PG to higher plant PSII RCC. However, both the function and exact location of the individual X-ray–visible or invisible PG molecules still remain to be elucidated. Biochimica et Biophysica Acta 1777 (2008) 1184–1194 Abbreviations: ΔpgsA, Synechocystis sp. PCC6803 ΔpgsA; 2D-BN/SDS-PAGE, Two- dimensional blue native SDS-PAGE; Chl, Chlorophyll; CP43, Chlorophyll–protein complex 43; CP47, Chlorophyll–protein complex 47; F685, Fluorescence emission peak at 685 nm (CP43); PBS, Phycobilisome; PG, Phosphatidylglycerol; PSI, Photosystem I; PSII, Photosystem II; RC, Reaction center; RC47, PSII core complex lacking CP43; RCC, Reaction center core complex; RCCI(3) and RCCI(1), trimeric and monomeric PSI core complexes; RCCII(2) and RCCII(1), dimeric and monomeric PSII core complexes; TL, Thermoluminescence ⁎ Corresponding author. Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H–6701 Szeged, P. O. Box 521, Hungary. Fax: +36 62 433 434. E-mail address: [email protected] (Z. Gombos). 1 These authors contributed equally to the article. 0005-2728/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.bbabio.2008.06.003 Contents lists available at ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbabio
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Role of phosphatidylglycerol in the function and assembly of photosystem II reaction center, studied...

Biochimica et Biophysica Acta 1777 (2008) 1184–1194
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbab io
Role of phosphatidylglycerol in the function and assembly of Photosystem II reactioncenter, studied in a cdsA-inactivated PAL mutant strain of Synechocystis sp. PCC6803that lacks phycobilisomes
Hajnalka Laczkó-Dobos a,1, Bettina Ughy a,1, Szilvia Z. Tóth a, Josef Komenda b,c, Ottó Zsiros a,Ildikó Domonkos a, Árpád Párducz d, Balázs Bogos a, Masayuki Komura e, Shigeru Itoh e, Zoltán Gombos a,⁎a Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H–6701 Szeged, Hungaryb Institute of Microbiology, Academy of Sciences, Opatovický mlýn, 37981, Třeboň, Czech Republicc Institute of Physical Biology, University of South Bohemia, Zámek 136, 37333 Nové Hrady, Czech Republicd Institute of Biophysics, Biological Research Center, Hungarian Academy of Sciences, H–6701 Szeged, Hungarye Division of Material Science (Physics), Nagoya University, Japan
Abbreviations: ΔpgsA, Synechocystis sp. PCC6803 Δdimensional blue native SDS-PAGE; Chl, Chlorophycomplex 43; CP47, Chlorophyll–protein complex 47;peak at 685 nm (CP43); PBS, Phycobilisome; PG, PhosphaI; PSII, Photosystem II; RC, Reaction center; RC47, PSII coReaction center core complex; RCCI(3) and RCCI(1), trimcomplexes; RCCII(2) and RCCII(1), dimeric and monomThermoluminescence⁎ Corresponding author. Institute of Plant Biology
Hungarian Academy of Sciences, H–6701 Szeged, P. O.433 434.
E-mail address: [email protected] (Z. Gombos).1 These authors contributed equally to the article.
0005-2728/$ – see front matter © 2008 Elsevier B.V. Adoi:10.1016/j.bbabio.2008.06.003
A B S T R A C T
A R T I C L E I N F OArticle history:
To analyze the role of phosp Received 14 February 2008Received in revised form 4 June 2008Accepted 4 June 2008Available online 10 June 2008Keywords:Synechocystis sp. PCC6803PhosphatidylglycerolPhotosystem IIThermoluminescenceOJIP transientCell division
hatidylglycerol (PG) in photosynthetic membranes of cyanobacteria we used twomutants of Synechocystis sp. PCC6803: the PAL mutant which has no phycobilisomes and shows a high PSII/PSI ratio, and a mutant derived from it by inactivating its cdsA gene encoding cytidine 5'-diphosphatediacylglycerol synthase, a key enzyme in PG synthesis. In a medium supplemented with PG the PAL/ΔcdsAmutant cells grew photoautotrophically. Depletion of PG in the medium resulted (a) in an arrest of cellgrowth and division, (b) in a slowdown of electron transfer from the acceptor QA to QB in PSII and (c) in amodification of chlorophyll fluorescence curve. The depletion of PG affected neither the redox levels of QA
nor the S2 state of the oxygen-evolving manganese complex, as indicated by thermoluminescence studies.Two-dimensional PAGE showed that in the absence of PG (a) the PSII dimer was decomposed into monomers,and (b) the CP43 protein was detached from a major part of the PSII core complex. [35S]-methionine labelingconfirmed that PG depletion did not block de novo synthesis of the PSII proteins. We conclude that PG isrequired for the binding of CP43 within the PSII core complex.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Phosphatidylglycerol (PG) is an anionic phospholipid that isrequired for the formation and function of thylakoid membranes incyanobacteria and plants, see a review by Wada and Murata [1].Depletion of PG modifies the function of both photosystem (PS) I andPSII reaction centers (RCs) [2–7].
The structure of PSI reaction center core complex (RCC) of Ther-mosynechococcus elongatus, as revealed by X-ray crystallography [8–
pgsA; 2D-BN/SDS-PAGE, Two-ll; CP43, Chlorophyll–proteinF685, Fluorescence emissiontidylglycerol; PSI, Photosystemre complex lacking CP43; RCC,eric and monomeric PSI coreeric PSII core complexes; TL,
, Biological Research Center,Box 521, Hungary. Fax: +36 62
ll rights reserved.
10] showed four lipid molecules inserted into the lumenal side of thePsaA and PsaB polypeptides. Two PG molecules are bound toproteins at the peripheral moiety of the RC. One PG and onemonogalactosyldiacylglycerol (MGDG) were found in the central coremoiety in the vicinity of two electron acceptor phylloquinonemolecules (A1 and A1′) [11]. PG molecules were also assumed toexist at the center of the PSI trimer in association with the small PsaLsubunits. This suggestion was based on the observation that in a PG-depleted mutant of Synechocystis sp. PCC6803, in which a pgsA generequired for PG biosynthesis was inactivated [4] the amount of PSItrimers decreased.
A recent study of the structure of PSII RCC in T. elongatus [12]reported the presence of several lipid molecules at the interfacebetween the subunit proteins. One of these was a PGmolecule locatedbetween the D1 and CP43 proteins in the postulated pathway of QB-quinone exchange. Recent biochemical studies have indicated thebinding of a large number of lipids to purified PSII RC [5]. For instance,6–7 molecules of PG were found to bind to T. elongatus PSII and asmaller number of PG to higher plant PSII RCC. However, both thefunction and exact location of the individual X-ray–visible or invisiblePG molecules still remain to be elucidated.

1185H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
The structural and functional roles of anionic lipids in photo-synthesis have raised scientific interest for a long time [1,13]. Earlyin vitro studies demonstrated the importance of PG by showing thesuppression of PSII activity [14] upon treatment of thylakoidmembranes with a phospholipase. Genomic analyses have openeda new experimental approach to the study of lipids. The role of PGin the assembly of PSI and PSII RCCs has been extensively studied ina PG-less mutant strain of Synechocystis sp. PCC6803 with aninactivated PG biosynthesis [4,6,15]. Satoh et al., [3] inactivated agene, cdsA, encoding cytidine 5′-diphosphate (CDP)-diacylglycerolsynthase that catalyzes the conversion of phosphatidic acid to CDP-diacylglycerol, and detected only a slight suppression of oxygenevolution. However, in another type of PG-less mutant cells inwhich the pgsA gene that encodes PG phosphate synthase, anenzyme known to catalyze the conversion of CDP-diacylglycerol toPG phosphate, was inactivated (ΔpgsA), Hagio et al. [2] found asignificant inhibition of PSII-driven oxygen evolution. In theseexperiments the mutant cells grew normally in a mediumsupplemented with PG, but after a few days of PG depletionshowed low oxygen-evolving activity with a concomitant decreaseof the content of chlorophyll a [2]. Gombos et al. [16] have shownthat inactivation of PSII by PG depletion in ΔpgsA cells was due tothe suppression of the secondary electron acceptor plastoquinoneQB reduction. This effect resembled that of a typical herbicide,DCMU, 3-(3, 4-dichlorophenyl)-1, 1′-dimethylurea, which binds atthe QB plastoquinone binding site and blocks re-oxidation of QA.Although these results clearly pointed out that PG depletion inducedinhibition of electron transfer at the PSII acceptor side, Sakurai et al.[6] using the same pgsA deletion mutant also suggested a functionfor PG on the donor side of PSII. Thus, the role of PG in thefunctioning of PSII needs further clarification, desirably by the use ofadditional PG mutants.
Previous results of Sakurai et al. [15] pointed to the destabiliza-tion of PSII RCC dimers upon PG depletion in a ΔpgsA mutant.These results and in vitro studies of plant PSII RCC indicated thatPG was involved in the dimerization of PSII [17]. The X-raycrystallographic structure showed only MGDG molecules but noPG in the lumenal region. On the other hand, a PG molecule wasdetected on the stromal side between CP43 and the D1 protein inthe neighborhood of the QB binding pocket [12]. This location is inaccordance with the previously mentioned effect of PG depletion onthe acceptor side of PSII. Moreover, location of a PG moleculebetween CP43 and D1 also implies a possible role of this lipidmolecule in the interaction between these two large PSII subunits.However, no data using PG-less mutants have suggested such a PGfunction so far.
The structure of the PSII RCC complex suggests that the psbC-encoded CP43 and psbB-encoded CP47 subunit proteins bind to theRCs around the D1 and D2 subunits, respectively, and provide thebinding site(s) for the smaller extrinsic proteins that cover the donorside [18]. Structural data have confirmed an interaction of CP47 withthe oxygen-evolving complex [19,20]. Although both the CP43 andCP47 proteins seem to be involved in the oxygen-evolving process,CP43 is known to be more easily detached from the RC than CP47[21,22]. Some RC complexes associated with CP47 may lack CP43(designated RC47 hereafter). RC47 exists but in a small fraction of RCunder normal cell growth conditions [23]. The results in the presentstudy indicated that the binding of CP43 to RC47 requires PG and thata loss of PG shifts the equilibrium between dimeric PSII core complex(RCCII(2)) and RC47 (+ free CP43).
In the present study we constructed and characterized a newmutant of Synechocystis sp. PCC6803with arrestedPGbiosynthesis.Weinactivated the cdsA gene in the PAL mutant, which compensates thephotosynthetic function of the cells for the missing phycobilisomes bya high cellular content of PSII [24]. The PAL/ΔcdsA mutant obtainedprovidedaunique experimental system foramoredetailed studyof the
function of PG in PSII.We analyzed the influence of PGdepletion on thefluorescence induction, thermoluminescence, biosynthesis andassem-bly of PSII protein subunits. Our results confirmed those obtainedearlier with the ΔpgsA mutant, i.e. the specific effect of PG molecules(a) on the acceptor side in PSII electron transfer and (b) on theoligomerization of PSII and PSI complexes. Additionally, we showedthat PG depletion affects the binding of the antenna CP43 to theperipheral regionwithin the PSII RCC. Themorphological studies of thePG-depleted PAL/ΔcdsA indicate an essential role of PG in cell fission.
2. Materials and methods
2.1. Organisms and growth condition
Cells of Synechocystis sp. PCC6803 wild-type, PAL and PAL/ΔcdsAmutant strains were grown photoautotrophically in BG11 medium[25] supplemented with 5 mMHEPES–NaOH (pH 7.5), and 20 μg mL−1
kanamycin. PAL/ΔcdsA mutant cells were cultivated in BG 11 mediumin the presence of 20 μMdioleoyl–PG (18:1/18:1 PG; P-9664; Sigma, StLouis). Cells were grown at 30 °C, under continuous illumination at theintensity of 30 μmol photons m−2 s−1 as described by Ajlani andVernotte [24] with minor modifications. Cultures were aerated on agyratory shaker operating at 100 rpm. PG depletion was achieved bywashing the cells twice with PG-free medium, followed by cultivationin PG-free medium.
2.2. Lipid analysis
Lipids were extracted from intact cells according to Bligh and Dyer[26]. Fatty acids were analyzed, by the use of a Supelco SP2330capillary column in Hewlett Packard HP6890 gas chromatographyequipment as described by Wada and Murata [27].
2.3. Construction of ΔcdsA and PAL/ΔcdsA mutant strains ofSynechocystis sp. PCC6803
A 1.4-kb genomic fragment of Synechocystis sp. PCC6803 containingcdsAwas amplified using PGF (5′-AGGTCCGCAACGTGGAGGTG-3′) andPGR (5′-CTGGAACGACTTGGGAAGTG-3′) primers. The kanamycin-resistance cassette from pUC4K was inserted between the MscI andNsiI sites of the cloned cdsA gene, yielding the inactivated construct.This cdsA-inactivated construct was used to transform the Synecho-cystis sp. PCC6803 wild-type and PAL mutant. Transformants wereselected on BG11 agar plates containing 20 μM PG and 50 μg/mLkanamycin. Primers were designed on the basis of the publishedcomplete genome sequence of Synechocystis sp. PCC6803 available at:http://www.kazusa.or.jp/. Complete segregation of the mutants wasconfirmed by PCR andby the fact that PGwas required for their growth.
2.4. Electron microscopic measurements
The harvested cells were fixed in 1% paraformaldehyde and 1%glutaraldehyde for 4 h at 4 °C and post-fixed in 1% osmium tetroxide.The samples were dehydrated in aqueous solutions of increasingethanol concentrations, and then embedded in Spurr resin. Followingpolymerization, 85—90 nm ultra thin sections were cut out by aReichert Ultracut E ultramicrotome. The sections were treated withuranyl acetate and lead citrate and subjected to electronmicroscopy ina Zeiss EM 902 electron microscope.
2.5. Radioactive labeling of the cells
Cells containing 75 μg of chlorophyll were labeled with [35S]methionine (at N1000 Ci mmol−1, Isotope Institute Ltd, Hungary)under irradiance of 60 μmol photons m−2 s−1 at 29 °C as described byKomenda et al. [28].

Fig. 1. Genetic inactivation of cdsA. (A) Physical map of the Synechocystis sp. PCC6803genomic fragment containing cdsA. In the inactivated strain the aphI gene cassettereplaces a 543-bp-long gene fragment. Small black arrows indicate positions of PCRprimers used to amplify the region and to verify gene replacement. (B) PCR analysis ofwild-type (lane 1) and PAL/ΔcdsA transformants (lanes 2–4), three independentsamples. Fragments containing wild-type and mutant alleles are indicated on the rightand size markers (lane M) on the left.
1186 H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
2.6. Preparation of membranes
Cyanobacterial membranes were prepared by breaking the cellswith glass beads (150–200 μm diameter) at 4 °C followed by dif-ferential centrifugation as described by Komenda and Barber [29].Breaking of cells and final resuspension were done in 25 mM Mes-NaOH, pH 6.5 containing 10mMCaCl2, 10 mMMgCl2 and 25% glycerol.
2.7. Protein analyses
The isolated membranes were solubilized with dodecyl-β-D-maltoside (at a dodecylmaltoside/chlorophyll ratio of 20:1, w/w)and analyzed by BN/SDS-PAGE at 4 °C in a 5–14% polyacrylamide gelaccording to Schägger and von Jagow [30]. Thewhole lane from the gelwas excised, incubated for 30 min in 25 mM Tris–HCl buffer (pH 7.5)containing 25% SDS and then layered onto the top of a denaturing gel.The protein composition of the complexes was then assayed by a 2ndelectrophoresis in a denaturing 12–20% linear gradient polyacrylamidegel containing 7 M urea [28]. In the gel the protein bands were stainedby Coomassie blue.
2.8. Measurement of cell density and pigment concentration
Absorption spectra of cell suspensions were recorded in aShimadzu UV-1601 spectrophotometer. Cell density in the cultureswas determined by measuring OD750. The chlorophyll concentrationwasmeasured by absorbance at 665 nm, using a 90%methanol extract.The A665 was multiplied by an extinction coefficient of 78.74 [31] tocalculate Chl a concentration (μg Chl a/mL).
2.9. Measurement of photosynthetic oxygen-evolving activity
Photosynthetic oxygen evolution in intact cells wasmeasured witha Clark-type oxygen electrode (Hansatech Instruments, Kings Lynn, U.K.) as described by Gombos et al. [16]. PSII oxygen evolving activitywas measured from H2O to exogenously added artificial quinones(500 μM p-benzoquinone). The cells were washed with BG11 mediumand suspended in fresh BG11 medium for the measurement of oxygenevolution. Light from an incandescent lamp equipped with a redoptical filter was used for all the oxygen evolution measurements at asaturating light intensity of 500 μmol photons m−2 s−1. The Chlconcentration of the cells was adjusted to 5 μg mL−1.
2.10. Thermoluminescence (TL) measurement
TLmeasurementswere carried out according to Vass et al. [32]withminor modifications. Intact cells were centrifuged and resuspended infresh BG11 medium. The cell suspensions were diluted to a Chlconcentration of Chl in 10 µg mL−1. The measurement of glow curvewas carried out in a temperature range from −10 °C to +80 °C using anapparatus similar to that described by Vass et al. [32]. Light emission ofthe samples was measured by a red-sensitive photomultiplier(Hamamatsu R/2228) and the signal was amplified through a homemade differential amplifier and fed to PC. The temperature of thesample holder was monitored using a platinum resistor thermometerplaced under the samples, and coolingwas achieved by liquid nitrogen.Samples were illuminated with 2 flashes (5 μs) at 5 °C before freezing.
2.11. The fast Chl a fluorescence (OJIP transient)
The Chl contents of the samples were set at 20 μg Chl a/ml.Samples were dark-adapted for about 2 h before the start of themeasurements. Fluorescence measurements were carried out by aHandy-PEA instrument (Hansatech Instruments Ltd, UK). A 20 μl cellsuspension was placed on a Whatman glass microfiber filter (GF/C)inserted in a leaf clip. The samples were illuminated with continuous
red light (650 nm peak wavelength) at the light intensity of 3500 μmolphotons m−2 s−1. The light was provided by three LEDs focused on acircle of 5 mm diameter. The first reliable measurable point of thefluorescence transient was at 20 μs. This intensity was taken as the F0value for dark-adapted cells. The length of the measurements was 3 s.
2.12. 77K fluorescence emission spectra
Low temperature steady-state fluorescence emission spectra (600–780 nm) were recorded using an excitation wavelength of 437 nm. Aspectrofluorimeter (Fluorolog-3/Jobin Yvon-Spex Instrument S.A.,Inc.) equipped with an adapter for measurement in liquid nitrogenwas applied. The emission spectra were corrected for photomultipliersensitivity and normalized to the fluorescence intensity at 700 nm.
3. Results
3.1. Inactivation of the cdsA gene in a PAL mutant strain of Synechocystissp. PCC6803
For a better understanding of the function of PG in thephotosynthetic apparatus of cyanobacteria we inactivated the cdsAgene, in a mutant of Synechocystis sp. PCC6803 strain denoted PAL. Thecoding region of cdsA was substituted by the kanamycin-resistancecassette (Kmr), yielding a strain denoted PAL/ΔcdsA (Fig. 1). The wild-type Synechocystiswas also transformedwith the same gene constructto yield ΔcdsA. Both the growth rate and the photosynthetic activitiesof ΔcdsAwere indistinguishable from those of the ΔpgsAmutant (dataare not shown). Also PG depletion affected pigment composition anddecreased the photosynthetic oxygen-evolving activity in ΔcdsAsimilar to that of ΔpgsA. The PAL strain mutant lacks phycobilisomes(PBSs) and the cells compensate for the loss of this major PSII antennaby increasing the PSII/PSI ratio [24]. The physiological features of theresulting mutant designated as PAL/ΔcdsA resembled those of theΔpgsAmutant. However, the cultivation time needed for PG depletionwas longer with PAL/ΔcdsA thanwith the ΔpgsAmutant, probably dueto the reduced usage of PG, which, in turn, may be the result of a lowcontent of PSI known to be rich in PG molecules [11].
3.2. Effects of PG depletion on growth rate and pigment content
The photoautotrophic growth of themutant cells inmediawith andwithout exogenously added PGwas followed bymeasuring the optical

Fig. 2. Growth curves and absorption spectra. (A) Growth curves of PAL/ΔcdsA mutantcells after inoculation to growth media with (■) and without (●) PG. (B) Absorptionspectra of PAL/ΔcdsAmutant cells of Synechocystis sp. PCC6803 grown for 20 days in thepresence (—) and in the absence (- - -) of PG.
Fig. 3. Effect of PG depletion on the oxygen evolving activity of (a) ΔpgsAmutant cells ofSynechocystis sp. PCC6803 and (b) PAL/ΔcdsA cells from H2O to CO2. Hundred percentactivity in “a” and “b” corresponds to 380 and 430 μmol O2mg−1 Chl h1, respectively. TheChl content was 5 μg mL−1. Activity values are expressed as the averages of threeindependent experiments in each point.
1187H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
density at 750 nmof the cell suspension (Fig. 2A). The growth rate in thepresence of PG was almost constant, giving a doubling time of 6–7 daysunder the present conditions at relatively low light intensity. In the PG-depleted medium the growth rate decreased gradually from the 4th dayon, and almost stopped on the 14th day of the PG depletion period.Nevertheless, it is worth mentioning that the cells were still alive evenafter 18days of PG starvationand showed light-inducedoxygen-evolvingactivity.Whenwe re-addedPG to thegrowthmediumafter 18 days of PGdepletion, the cells started to growagain (not shown). These features arevery similar to those reported for the ΔpgsA mutant cells [4].
The Chl content of the PG-depleted PAL/ΔcdsA cells decreasedslightly during the depletion process (Fig. 2B). This situation wasdifferent from that experienced with ΔpgsA cells. These latter lostabout 50% of their Chl content after three weeks of PG depletion,mainly because of a decreased PSI content [4]. The absorption spectraof the PG-depleted PAL/ΔcdsA cells showed a significant increase incarotenoid content compared to that of the mutant cells grown in thepresence of PG (Fig. 2B)
3.3. Photosynthetic oxygen-evolving activity
3We measured the photosynthetic oxygen-evolving activity ofPAL/ΔcdsA cells grown in PG-supplemented or PG-depleted medium.
The rate of net photosynthesis measured fromwater to carbon dioxidevia PSII and PSI decreased gradually during 20 days of cultivationwithout PG with a half-time of about 10 days. We detected about 15%of the full activity even after 18 days. Inactivation of the photosyn-thetic activity of PAL/ΔcdsA cells was faster than that of ΔpgsA cells(Fig. 3). On the other hand, when the oxygen-evolving activity of PSIIwas measured in the presence of an artificial electron acceptor, 1,4-p-benzoquinone (water to quinone electron transfer via PSII), the ratedecreased much faster, giving an apparent half-time of about 4 daysfor the PAL/ΔcdsA cells (not shown). PSII activity measured in thesecells in the presence of p-benzoquinone could not be detected after11 days of PG depletion while in the ΔpgsA cells oxygen evolutioncompletely ceased after 3 days [2].
3.4. Effect of PG depletion on the content and fatty acid composition ofPG in PAL/ΔcdsA cells
Since the synthetic PGmolecules added to the cells externally weredifferent from the natural ones, they had to be retailored to generatethe natural PG molecules (Table 1). Externally added PG contained18:1 fatty acids exclusively. Cells grown in the presence of PG for4 days contained more than 40 mol% of oleic acid, which changed toabout 10 mol% following an 8-day PG depletion period. Thisconcentration was close to that found in PAL cells. During this periodof depletion, the PG content decreased from 4.4 nM PG/mg protein to2.1 nM PG/mg protein. This decrease to 50% of the original amount ofcells measured on a protein basis corresponded to one cell divisionduring this period.
3.5. Effect of PG depletion on cell size and cell structure
Fig. 4A shows an electron micrograph of a PAL mutant cell. Thecells lacking PBSs exhibited a very tight stacking of the thylakoidmembranes that resembles the granal structures of higher plantthylakoids [33]. The EM picture of a PAL/ΔcdsA mutant cell grown inthe presence of PG also showed a tight packing of thylakoidmembranes, similar to those in the PAL cell. The size of mutant cellsgrown in the presence of PG was also similar to that of the PAL cells.However, the growth rate of themutant was lower than that of the PALcells. This might be explained by a slow passage of PG through the cellmembranes. The size of PG-depleted PAL/ΔcdsA cells was significantlylarger than that of the cells grown in the presence of PG. We can seelarge empty areas or dense phosphate granules among the thylakoidmembranes in this case. The packaging of membranes was less regularin the PG-depleted cells than in the PG-supplemented ones. As to
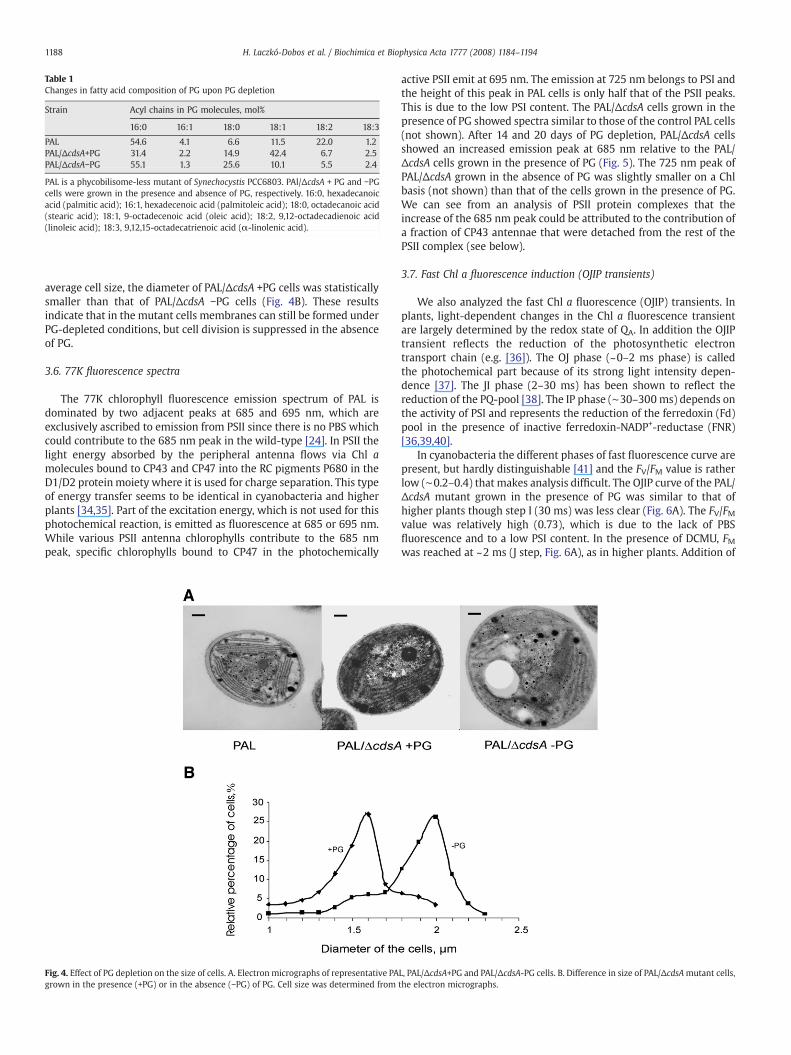
Table 1Changes in fatty acid composition of PG upon PG depletion
Strain Acyl chains in PG molecules, mol%
16:0 16:1 18:0 18:1 18:2 18:3
PAL 54.6 4.1 6.6 11.5 22.0 1.2PAL/ΔcdsA+PG 31.4 2.2 14.9 42.4 6.7 2.5PAL/ΔcdsA−PG 55.1 1.3 25.6 10.1 5.5 2.4
PAL is a phycobilisome-less mutant of Synechocystis PCC6803. PAl/ΔcdsA + PG and −PGcells were grown in the presence and absence of PG, respectively. 16:0, hexadecanoicacid (palmitic acid); 16:1, hexadecenoic acid (palmitoleic acid); 18:0, octadecanoic acid(stearic acid); 18:1, 9-octadecenoic acid (oleic acid); 18:2, 9,12-octadecadienoic acid(linoleic acid); 18:3, 9,12,15-octadecatrienoic acid (α-linolenic acid).
1188 H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
average cell size, the diameter of PAL/ΔcdsA +PG cells was statisticallysmaller than that of PAL/ΔcdsA −PG cells (Fig. 4B). These resultsindicate that in the mutant cells membranes can still be formed underPG-depleted conditions, but cell division is suppressed in the absenceof PG.
3.6. 77K fluorescence spectra
The 77K chlorophyll fluorescence emission spectrum of PAL isdominated by two adjacent peaks at 685 and 695 nm, which areexclusively ascribed to emission from PSII since there is no PBS whichcould contribute to the 685 nm peak in the wild-type [24]. In PSII thelight energy absorbed by the peripheral antenna flows via Chl amolecules bound to CP43 and CP47 into the RC pigments P680 in theD1/D2 protein moiety where it is used for charge separation. This typeof energy transfer seems to be identical in cyanobacteria and higherplants [34,35]. Part of the excitation energy, which is not used for thisphotochemical reaction, is emitted as fluorescence at 685 or 695 nm.While various PSII antenna chlorophylls contribute to the 685 nmpeak, specific chlorophylls bound to CP47 in the photochemically
Fig. 4. Effect of PG depletion on the size of cells. A. Electron micrographs of representative PAgrown in the presence (+PG) or in the absence (−PG) of PG. Cell size was determined from
active PSII emit at 695 nm. The emission at 725 nm belongs to PSI andthe height of this peak in PAL cells is only half that of the PSII peaks.This is due to the low PSI content. The PAL/ΔcdsA cells grown in thepresence of PG showed spectra similar to those of the control PAL cells(not shown). After 14 and 20 days of PG depletion, PAL/ΔcdsA cellsshowed an increased emission peak at 685 nm relative to the PAL/ΔcdsA cells grown in the presence of PG (Fig. 5). The 725 nm peak ofPAL/ΔcdsA grown in the absence of PG was slightly smaller on a Chlbasis (not shown) than that of the cells grown in the presence of PG.We can see from an analysis of PSII protein complexes that theincrease of the 685 nm peak could be attributed to the contribution ofa fraction of CP43 antennae that were detached from the rest of thePSII complex (see below).
3.7. Fast Chl a fluorescence induction (OJIP transients)
We also analyzed the fast Chl a fluorescence (OJIP) transients. Inplants, light-dependent changes in the Chl a fluorescence transientare largely determined by the redox state of QA. In addition the OJIPtransient reflects the reduction of the photosynthetic electrontransport chain (e.g. [36]). The OJ phase (~0–2 ms phase) is calledthe photochemical part because of its strong light intensity depen-dence [37]. The JI phase (2–30 ms) has been shown to reflect thereduction of the PQ-pool [38]. The IP phase (∼30–300 ms) depends onthe activity of PSI and represents the reduction of the ferredoxin (Fd)pool in the presence of inactive ferredoxin-NADP+-reductase (FNR)[36,39,40].
In cyanobacteria the different phases of fast fluorescence curve arepresent, but hardly distinguishable [41] and the FV/FM value is ratherlow (∼0.2–0.4) that makes analysis difficult. The OJIP curve of the PAL/ΔcdsA mutant grown in the presence of PG was similar to that ofhigher plants though step I (30 ms) was less clear (Fig. 6A). The FV/FMvalue was relatively high (0.73), which is due to the lack of PBSfluorescence and to a low PSI content. In the presence of DCMU, FMwas reached at ~2 ms (J step, Fig. 6A), as in higher plants. Addition of
L, PAL/ΔcdsA+PG and PAL/ΔcdsA-PG cells. B. Difference in size of PAL/ΔcdsAmutant cells,the electron micrographs.

Fig. 5. Fluorescence emission spectra. 77K fluorescence emission spectra of PAL/ΔcdsAmutant cells grown in the presence (—) and absence of PG for 14 (. . .) and for 20 (- - -)days, respectively.
1189H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
methylviologen, which accepts electrons from PSI led to a lower FMvalue (data not shown). Dibromothymoquinone, which inhibits the re-oxidation of plastoquinol by binding to the cyt b6f complex causedhigher fluorescence intensity at step I (data not shown). These featuresare typical of the OJIP transient of higher plants [36]. Thus it seemsthat the OJIP curve of the PAL/ΔcdsA mutant can be interpreted in asimilar manner.
Depletion of PG resulted in a gradual increase of the fluorescenceintensity at step J and after 14 days of PG depletion, the fluorescence
Fig. 6. A, Effect of PG depletion on Chl a fluorescence (OJIP) transients of PAL/ΔcdsAmutant c(called Ft/FM curves). B, Effect of DCMU on fluorescence transients of cells grown in the abseheat-treated cells (50 °C, 10 min) grown in the presence of PG, with (▴) and without (Δ) Drepresented on a logarithmic time scale. For additional details see Materials and methods.
curve became similar to that of DCMU-treated cells (Fig. 6A). Thissuggests that PG depletion inhibited or slowed down electrontransport at the acceptor side of PSII.
We also detected an approximately 10 decrease of the FM valuein PG-depleted cells (for a better comparison of the OJIP curves,they were normalized to FM). The FM values of cells depleted of PGfor 11 days were found to be identical both in the presence and inthe absence of DCMU (Fig. 6B, no normalization) indicating that allPSII reaction centers were closed at FM. Thus the 10% decrease ofFM may reflect a slightly reduced amount of photochemically activePSII. The value of F0 gradually increased upon PG depletion andafter 14 days it was ~20% higher than in the PG-supplementedcells; this might have arisen from a partial detachment of theCP43 unit (see below).
We also tested the possible impairment of the PSII donor sidethat has recently been proposed to be the cause of PSII inactivationunder PG deficiency [6]. For comparison, we partially inactivatedoxygen evolution by heat treatment (10 min at 50 °C). As a result,the fluorescence intensity at step J decreased, the fluorescenceincrease from J to P was delayed and a much lower FM value wasreached (Fig. 6C). The FM value could be largely restored by theaddition of DCMU but the increase in fluorescence intensity wasslower (compare Fig. 6B and C) similar to heat-treated leaves [42].In this case, the decrease of FM was due to a lack of electronscoming from the donor side of PSII. Heat treatment caused only anapproximately 15% increase in F0, indicating that there was neithermassive detachment of the antenna complexes nor degradation ofPSII RCs. The above characteristics of the heat-treated cells wereclearly different from those of the −PG cells. Thus, it appeared thatthe inhibition of PSII electron transport could be primarily ascribedto the slowdown or inhibition of QA
− re-oxidation and the inhibitionof the PSII donor side was probably negligible or only secondary.
ells and the effect of DCMU on +PG samples. Fluorescence curves were normalized to FMnce of PG for 11 days. The curves are presented without normalization. C, OJIP curves ofCMU. The curves are presented without normalization. All fluorescence transients are

Fig. 7. Effect of PG depletion on the TL characteristics of PAL/ΔcdsAmutant cells. (A) Cellsgrown in the presence of PG (a) were transferred for 7 (b) or 14 (c) days to a similarmedium without PG. To achieve cell recovery (d) the cells were grown without PG for14 days and thenwith PG. Q and B indicate the recombination steps between QA
− and S2,QB− and S2, respectively. (B) The TL curves of +PG cells (e) and PG-depleted cells for
14 days (f) measured after treatment with 10 μM DCMU.
Fig. 8. 2D-BN/SDS-PAGE of radioactive labeled membrane proteins from PAL/ΔcdsA cellseither supplemented with PG (+PG) or depleted of PG for 20 days (−PG 20 d). Cells wereradioactively labeled at 500 μmol photons m−2 s−1 and 29 °C with [35S]Met for 20 minand then used for the isolation and analysis by 2D-BN/SDS-PAGE of thylakoids. Theupper panels (A and B) show Coomassie stained gels, the lower ones (C and D) thecorresponding autoradiograms. Designations: RCCI(3) and RCCI(1), trimeric andmonomeric PSI core complexes, respectively; RCCII(2) and RCCII(1), dimeric andmonomeric PSII core complexes, respectively; RC47, PSII core complex lacking CP43.Sixty µg Chl was loaded for each sample.
1190 H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
3.8. Effect of PG depletion on thermoluminescence (TL)
To study electron transfer activity on the acceptor side of PSII inmore detail, we measured the thermoluminescence deriving fromPSII. Sample cells were illuminated at 5 °C with 2 flashes (5 μs) ofalmost saturating intensity, cooled down to −10 °C, and then warmedup by a heater to activate light emission. The TL curve of the PG-supplemented cells showed a peak at 27 °C. This TL band correspondsto band B that originates from charge recombination between QB
− andS2 states (Fig. 7A). A similar TL band was obtained following 7 days ofPG depletion, but after depletion of PG for 14 days the B band peaktemperature shifted down to 19 °C, towards band Q. Band Q representscharge recombination between QA
− and S2 states. Re-addition of PG tothe growthmedium of the cells depleted of PG for 14 days restored thepeak to 27 °C with the disappearance of the peak at 19 °C.
The band B (S2QB−) formed in the cells depleted of PG for 7 days
was similar to that in the +PG cells. This indicates that there wasalmost no change in the energy levels (redox potentials) of either S2or QB. The band that was detected in the 14-day-PG-depleted cellsresembled the one seen with DCMU in the wild-type cells [16] orwith DCMU in the + or −PG PAL/ΔcdsA mutant cells (Fig 7B). Thisresult indicates that PG depletion for a prolonged time results in thesuppression of electron transfer from QA
− to QB and confirms theconclusions drawn from other experiments described in previoussections of this paper. We conclude that the redox energy levels ofthe S2 and QA states are almost the same in PAL/ΔcdsA as in the wildtype cells.
3.9. Effect of PG depletion on the synthesis and assembly of PSII proteinsubunits
In order to reveal eventual structural changes induced by PGdepletion in PAL/ΔcdsA cells we performed two-dimensional gelelectrophoresis of the thylakoid proteins. We used blue native (BN)
electrophoresis for the separation of protein complexes and SDSelectrophoresis for the separation of protein subunits of the individualcomplexes (2D-BN/SDS-PAGE). Similar to the membrane proteincomplexes of the wild-type Synechocystis sp. PCC6803 cells [43] themost abundant complexes in the thylakoid membranes of the PG-containing PAL/ΔcdsA mutant cells (Fig. 8A) were monomeric andtrimeric PSI RC core complexes [RCCI(1) and RCCI(3)] as well asmonomeric and dimeric PSII RC core complexes [RCCII(1) and RCCII(2)]. Both PSII complexes contained all the four PSII large subunits, D1,D2, CP47 and CP43 (Fig. 8A). A small amount of PSII proteins was alsofound in the monomeric core complex lacking CP43 (RC47) as well asin the fraction of unassembled proteins CP47 and CP43. PAL/ΔcdsAcells depleted of PG for 20 days showed a significant decrease in PSIIdimer content (Fig. 8B), a phenomenon previously reported also forPG-depleted cells of ΔpgsA [15]. Trimeric PSI complexes weredestabilized and converted into monomers in these cells as reportedpreviously [4]. However, the two-dimensional gel also revealed apreviously unidentified increase in the content of RC47. This suggests alower efficiency of CP43 binding to the PSII core complexes, probably aconsequence of PG depletion (Fig. 8B), in agreement with thefluorescence emission experiments.
Radioactive labeling with [35S]Met of proteins of PG-containingPAL/ΔcdsA (Fig. 8C) cells also exhibited a protein labeling patterntypical of PSII inwild-type, with preferential incorporation of the labelinto protein D1 [43]. This pattern did not change significantly in thePAL/ΔcdsA cells even after 20 days of PG starvation (Fig. 8D),suggesting that de novo synthesis of large PSII subunits was stillgoing on, even though the formation of RCCI(3) and RCCII(2) wasrather suppressed. We observed a significant increase in the amountof newly synthesized RC47 lacking CP43.

1191H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
4. Discussion
Anionic lipids suchasPGorcardiolipin are important cell constituentsin the structure and function of protein complexes involved in thephotosynthetic and respiratory electron transport chains [44]. They canact as co-chaperones for trafficking of proteins in the assembly of RCsthrough or inside membranes [45] and they can also affect translationof proteins involved in mitochondrial electron transport [46–48].
4.1. Effect of PG depletion on the growth and structure of PAL/ΔcdsA cells
Thylakoid membranes in chloroplasts and cyanobacteria have alipid matrix with a highly conserved composition. PG is an indis-pensable component of these membranes. To get a better insight intoits role in the structure and function of PSII, we produced a PG-lessmutant cell line by inactivating the cdsA gene, which encodes a keyenzyme in PG biosynthesis in the phycobilisome-less mutant strain(PAL) of Synechocystis sp. PCC6803 [24]. As a consequence the mutantcells exhibit a high PSII content to compensate for the loss ofphycobilisomes. This strain allows a much easier interpretation ofPSII fluorescence data without the contribution of PBS. Also, the highFV/FM values resembling those found in higher plants make it possibleto interpret the OJIP curves in a similar manner as in higher plants.
The PAL/ΔcdsA mutant required a continuous supply of PG toachieve photoautotrophic growth. The same experimental setup hasbeen used with the ΔpgsA mutant for studying the role of PG incyanobacterial photosynthesis [2,4,5,6,15,16]. The chlorophyll contentof PAL/ΔcdsA cells did not change significantly during PG depletion asevidenced by the absorption spectrum of cells even after a PGdepletion for 20 days. This result contrasts that obtained with a ΔpgsAmutant. This mutant showed a decrease of about 30% in Chl contentper cell upon prolonged PG depletion [2]. The less significant effect ofPG depletion detected in the present study may come from the highPSII/PSI ratio (i.e. high PSII content) of the PAL/ΔcdsA cells relative tothat of the ΔpgsA cells. In these latter, PG depletion has been shown toresult in a decreased PSI content after a prolonged PG depletion [4] for20 days. Since the PSI content of the PAL/ΔcdsA is initially low, theeffect of PG depletion on its further decrease would be hard to detect.Another possibility is that reduction of the severity of the effect of PGdepletion in the PAL cells relative to theΔpgsA cells even after a 20-daydepletion is due to a slower growth rate as discussed below.
Loss of PG resulted in inhibition of cell growth as reported for theΔpgsAmutant [2] or for Synechocystis sp. PCC6803 ΔcdsA cells [3]. Cellimages in Fig. 3 also indicate that PG depletion results in a 2 to 3-foldenlargement of cell volume. It seems that PG depletion inhibits celldivision although the biosynthesis of most of the proteins and otherlipids still continues under PG depletion [2,6].
The 77K fluorescence emission spectrum of PG-depleted PAL/ΔcdsA cells showed a remarkable increase of the 685-nm peak (F685)compared to the control +PG cells. In the absence of PBS, which mayalso contribute to this emission even when excited at 440 nm [6], thisincrease could unequivocally be ascribed to the emission fromantenna Chl a, mainly from the CP43 protein [49]. The increase ofthe F685 peak seems to indicate either an increased amount of theCP43 protein itself, or an increase in fluorescence yield owing todetachment of this antenna from RC. There is no indication of anabsolute increase in the cellular content of CP43 on two-dimensionalgels (Fig. 8), making the former possibility unlikely. On the other hand,protein analyses revealed that a significant fraction of CP43 wasdetached from the PSII core complex.We think that this detachment isthe most probable reason of the F685 increase and F0 increase.
4.2. Effect of PG depletion on PSII electron transfer in PAL/ΔcdsA cells
It has been reported that in ΔpgsA mutant cells PG depletionresults in a significant suppression of PSII electron transport measured
as the rate of oxygen evolution [2]. This kind of suppression has beenreported to occur in the ΔcdsA mutant cells as well [3] and confirmedin the present paper in the PAL/ΔcdsA cells. We assume that the slowerdecrease in the oxygen-evolving activity during PG depletion in thisstrain in comparison with ΔpgsA is related to a slower growth rate(Fig. 2A), which is probably due to an inefficient absorption of thephotosynthetically efficient radiation in the absence of PBS. Slowergrowth rate also means a slower consumption of remaining PGmolecules.
From a modification of flash-induced fluorescence kinetics aswell as from the TL results in the ΔpgsA mutant cells, Gombos et al.[16] suggested that PG loss resulted in an inhibition of electrontransfer between QA and QB. However, Sakurai et al. [6] haverecently suggested that in this case PSII was also damaged on thedonor side, a conclusion based on measuring variable fluorescenceby a PAM fluorimeter and PSII electron transport from water or froman artificial electron donor DPC to DCPIP of the PSII RC in the ΔpgsAcells.
The PAL/ΔcdsA mutant cells have high PSII/PSI ratios resulting inhigh FV/FM values. This allows a better assessment of Chl afluorescence (“Kautsky effect” [50]), especially of the fast (OJIP)transient. Fluorescence intensity increases with the reduction of QA
and during the reduction of electron transport chain [36,51–54]. UponPG depletion of PAL/ΔcdsA cells, a gradual increase in the fluorescenceintensity at step J was observed, similar to that seen in the presence ofincreasing DCMU or dibromothymoquinone concentrations [36].Therefore, the increase in the intensity at the J step may arise eitherfrom (i) a mixture of fully active reaction centers and reaction centerswhere QA
− to QB electron transfer is completely inhibited, or from (ii)slower electron transfer reactions at the acceptor side of PSII in all RCs.A shift in the TL band B towards TL band Q points to a slowdown andnot a complete inhibition of electron transfer reactions at the acceptorside of PSII RCs.
To check the possibility that upon PG depletion PSII inhibitionoccurs primarily on the donor side, as it was proposed by Sakurai etal. [6], we compared the OJIP fluorescence curves of PG-depletedcells of the PAL/ΔcdsA mutant and those of PG-supplemented cellswith partially inactivated (heat treatment at 50 °C for 10 min,[55,56]) oxygen evolution. Heat-treated cells displayed low J and FMvalues that could be largely restored by the addition of DCMU(Fig. 6C). Since these phenomena were clearly different from thatdetected in the −PG cells, inhibition of the oxygen-evolving complexseems to be negligible; this is corroborated by the fact that theintensities of TL bands B and Q did not decrease even after 14 days ofPG depletion (Fig. 7). The somewhat higher F0 value detected in thePG-depleted PAL/ΔcdsA cells (Fig. 6A) may be related to a highercontribution of F685 detected at 77K, i.e. a partial detachment of theCP43 unit.
4.3. Effect of PG depletion on the donor side: redox levels of QA and S2measured by thermoluminescence
The TL bands B and Q are known to reflect the difference betweenthe energy level of the P680+pheophytin− state and those of theradical pairs S2QB
− and S2QA− , respectively, accumulated in PSII [32].
The +PG PAL/ΔcdsA cells showed a TL band at 27 °C. The peaktemperature was similar to that of the B band measured undersimilar condition in wild-type cells [16]. This result suggests that inthis type of cells the energy level of the S2QB
− state is normal. UponPG depletion we detected a TL peak at 19 °C, a temperature similar tothat characteristic of the band Q in the presence of DCMU [32]. Theseresults suggest that the energy levels of QA, QB and S2 were notaffected by PG depletion.
During PG depletion for a short period of time (7 days), the peaktemperature (band B) of TL did not change. After PG depletion for14 days the band B shifted down towards the band Q. This indicates

Fig. 9. Scheme illustrating the tentative localization of PG molecules in the vicinity ofthe protein backbone of PSII reaction centers [RCCII(1) and RCCII(2)]. The symbol of PGindicates putative localization of PG molecules in the neighborhood of CP43 [RCCII(1)]and in the assembly of the dimer [RCCII(2)] attached to each other through CP47.Detachment of CP43 from the reaction center can be related to depletion of PG.
1192 H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
suppression of QB function. However, re-addition of PG fully restoredthe band B. These results were very similar to those reported for theΔpgsA cells [16]. Gombos et al., [16] have reported that the DCMU-induced TL band in the PG-depleted cells of ΔpgsA cells was sharperthan that obtained in cells without DCMU. Thus DCMU seems to bindto the modified QB site too [57].
Our data on the behavior of the TL band in PG-deprived cells(Fig. 7) were somewhat different from those obtained by treatmentof isolated thylakoid membranes with phospholipase C that digestsPG [14]. The latter treatment caused a rapid oxidation of QB
− andled to a decreased intensity of both the bands B and Q with aconcomitant increase of band C. This suggests a severe damage tothe structure of the PSII donor side. In the thermoluminescencepattern of PAL/ΔcdsA the band B shifted to the position of band Qbut the amplitude of the band B was comparable to that of bandQ. This suggests that the donor side of PSII was not severelyaffected.
4.4. Effect of PG depletion on the assembly of the PSII RC complex
Protein analysis by 2D-BN/SDS-PAGE confirmed earlier conclu-sions obtained from sucrose density gradient centrifugationanalyses of PSII complexes of the ΔpgsA strain: PG depletion leadsto the destabilization of both dimeric PSII and trimeric PSIcomplexes [4,15]. However, our data reported in this paper alsoshowed a new aspect of PG depletion, which was not identifiedbefore, namely, detachment/destabilization of the binding of CP43within PSII RCC. In the structure of the monomeric RCCII complexthe antenna subunits CP43 and CP47 are symmetrically located onthe opposite sides of the D1–D2 heterodimer, with CP43 on the sideof D1 and CP47 on the side of D2 [18]. CP43 is known to be moreeasily detached from RCCII than CP47 [21,22]. In cells of Synecho-cystis sp. PCC6803 there is always a certain fraction of PSII RCC thatlacks CP43 (designated RC47 hereafter). This reflects a selectiveturnover of D1, which requires detachment of CP43. We assume thatthe reason why detachment of CP43 has not been detectedpreviously is that the 2D-BN/SDS-PAGE has a higher resolvingpower than sucrose density gradient centrifugation, used earlier. Itis very probable that the fraction of monomeric PSII RCC obtained bythe centrifugation method contains not only the genuine RCCII(1)but also the complex lacking CP43 (RC47). To support this view wecan argue that Sakurai et al. [15] found Psb28 protein in this fraction,which should be a component of RC47 rather than a constituent ofRCCII(1) (Komenda, unpublished data). In other words, PG seems tobe required for the binding of CP43 to the PSII RC. It is not yet clearwhether RC47 has no CP43 or just has loosely bound CP43, becausemild detergent treatment in the first dimension might dissociateloosely bound CP43. Fluorescence data show that some CP43 isremoved in vivo.
Dissociation of CP43 by PG depletion may also contribute to thedestabilization of the dimeric PSII RCC. CP43 participates in thebinding of the Mn4Ca cluster and the peripheral proteins [18,58] ofoxygen-evolving complex. It is especially the 33 kDa protein thatcontributes to the stability of the dimeric PSII RCC. In addition, takinginto account the involvement of CP43 in liganding Mn4Ca cluster ofthe oxygen-evolving complex, detachment of CP43 induced by PGdepletion might be responsible for the observed effects of PGdepletion on the donor side of PSII [6].
By using [35S]-labeled methionine we also followed thesynthesis of PSII proteins and their incorporation into PSIIcomplexes. We confirmed that PG depletion does not inhibit thesynthesis of large PSII subunits and that selective replacement ofD1 is not abolished, as has also been shown by Sakurai et al. [15].Localization of the PG molecules in PSII RCs has been suggested inboth earlier X-ray crystallographic studies [12] and in our presentprotein analyses by 2D gel electrophoresis (Fig. 9). Taken together
these data imply that one PG molecule is localized in the vicinityof D1 protein and QB plastoquinone and another one can stabilizethe attachment of CP43 to the PSII reaction center core.
5. Conclusions
We constructed a mutant strain PAL/ΔcdsA that contains largeamounts of PSII without phycobilisome and is therefore a uniqueexperimental object for the study of the role of PG in PSII function/assembly. The mutant cells grew only in a medium supplementedwith PG. Depletion of PG in the cells resulted (i) in an inhibition ofcell growth/division, (ii) in a small change in pigment composition,(iii) in the inactivation of oxygen evolution, (iv) in a modification ofthe fluorescence induction curve that pointed to some damage to QB,but not to the donor side, (v) in a modification of the TL glow curve togive only the shifted Q-band, owing to a suppression of electrontransfer between QA and QB, and leaving the redox levels of QA and S2normal, (vi) in changes of the protein pattern of the cells to give riseto decreased trimeric PSI and dimeric PSII RCC, (vii) in a destabiliza-tion of PSII RCC structure owing to a release of protein CP43 (increasein RC47 content), and (viii) in continuous incorporation of methio-nine [35S] label into proteins (i.e., continuous biosynthesis of PSI andPSII subunit proteins). PG depletion may also affect the donor side ofPSII at a later stage. This effect, however, seems to be small andmightbe associated with the release of CP43 from PSII RCC, as this antennachlorophyll protein is directly involved in liganding the Mn4Cacluster [12].

1193H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
Acknowledgements
This work was supported by grants from the Hungarian ScienceFoundation (OTKA; grant nos. T 60109 and T 68692) and co-financedby the French–Hungarian bilateral research project “Balaton”, by theMinistry of Education, Youth and Sports of the Czech Republic(project no. MSM6007665808) and by the Czech Academy ofSciences (Institutional Research Concept no. AV0Z50200510,Czech–Hungarian bilateral research priority project and projectIAA400200801). The authors thank Dr. Ghada Ajlani, CEA Saclay, Gif-sur-Yvette, France, for her advice in preparing the PAL mutant, aswell as for reading and correcting the manuscript. We are grateful toProf. Ferenc Solymosy for reading and correcting the manuscript.The authors are grateful to Anna Sallai for her technical assistanceand to Dr. Gert Schansker for the useful discussions.
References
[1] H. Wada, N. Murata, The essential role of phosphatidylglycerol in photosynthesis,Photosynth. Res 92 (2007) 205–215.
[2] M. Hagio, Z. Gombos, Z. Varkonyi, K. Masamoto, N. Sato, M. Tsuzuki, H. Wada,Direct evidence for requirement of phosphatidylglycerol in photosystem II ofphotosynthesis, Plant Physiol 124 (2000) 795–804.
[3] N. Sato, M. Hagio, H. Wada, M. Tsuzuki, Requirement of phosphatidylglycerol forphotosynthetic function in thylakoid membranes, Proc. Natl. Acad. Scie. USA 97(2000) 10655–10660.
[4] I. Domonkos, P. Malec, A. Sallai, L. Kovacs, K. Itoh, G.Z. Shen, B. Ughy, B. Bogos, I.Sakurai, M. Kis, K. Strzalka, H. Wada, S. Itoh, T. Farkas, Z. Gombos, Phosphati-dylglycerol is essential for oligomerization of photosystem I reaction center, PlantPhysiol 134 (2004) 1471–1478.
[5] I. Sakurai, J-R. Shen, J. Leng, S. Ohashi, M. Kobayashi, H. Wada, Lipids in oxygen-evolving photosystem II complexes of cyanobacteria and higher plants, J. Biochem40 (2006) 201–209.
[6] I. Sakurai, N. Mizusawa, S. Ohashi, M. Kobayashi, H. Wada, Effects of the lack ofphosphatidylglycerol on the donor side of photosystem II, Plant Physiol 144(2007) 1336–1346.
[7] C.C. Xu, H. Hartel, H. Wada, M. Hagio, B. Yu, C. Eakin, C. Benning, The pgp1 mutantlocus of Arabidopsis encodes a phosphatidylglycerolphosphate synthase withimpaired activity, Plant Physiol 129 (2002) 594–604.
[8] P. Fromme, Structure and function of photosystem I, Curr. Opin. Struct. Biol 6(1996) 473–484.
[9] P. Fromme, P. Jordan, N. Krauss, Structure of photosystem I, Biochim. Biophys. Acta1507 (2001) 5–31.
[10] W. Saenger, N. Krauss, Three-dimensional structure of cyanobacterial photo-system I at 2.5 ångström resolution, Nature 411 (2001) 909–917.
[11] P. Jordan, P. Fromme, H.T. Witt, O. Klukas, W. Saenger, N. Krauss, Three-dimensional structure of cyanobacterial photosystem I at 2.5 ångström resolution,Nature 411 (2001) 909–917.
[12] B. Loll, J. Kern, W. Saenger, A. Zouni, J. Biesiadka, Towards complete cofactor arrange-ment in the 3.0 Å resolution structure of photosystem II, Nature 438 (2005) 1040–1044.
[13] N. Sato, Roles of the acidic lipids sulfoquinovosyl diacylglycerol and phosphati-dylglycerol in photosynthesis: their specificity and evolution, J. Plant Res 117(2004) 495–505.
[14] M. Droppa, G. Horvath, E. Hideg, T. Farkas, The role of phospholipids in regulatingphotosynthetic electron transport activities: treatment of thylakoids withphospholipase C, Photosynth. Res 46 (1995) 287–293.
[15] I. Sakurai, M. Hagio, Z. Gombos, T. Tyystjarvi, V. Paakkarinen, E.M. Aro, H. Wada,Requirement of phosphatidylglycerol for maintenance of photosynthetic machin-ery, Plant Physiol 133 (2003) 1376–1384.
[16] Z. Gombos, Z. Varkonyi, M. Hagio, M. Iwaki, L. Kovacs, K. Masamoto, S. Itoh, H. Wada,Phosphatidylglycerol requirement for the function of electron acceptor plastoqui-none QB in the photosystem II reaction center, Biochemistry 41 (2002) 3796–3802.
[17] O. Kruse, B. Hankamer, C. Konczak, C. Gerle, E. Morris, A. Radunz, G.H. Schmid, J.Barber, Phosphatidylglycerol is involved in the dimerization of photosystem II, J.Biol. Chem. 275 (2000) 6509–6514.
[18] A. Zouni, H.T. Witt, J. Kern, P. Fromme, N. Krauss, W. Saenger, P. Orth, Crystalstructure of photosystem II from Synechococcus elongatus at 3.8 Å resolution,Nature 409 (2001) 739–743.
[19] T.M. Bricker, L.K. Frankel, Use of amonoclonal antibody in structural investigationsof the 49 kDa polypeptide of photosystem II, Arch. Biochem. Biophys 256 (1987)295–301.
[20] T.M. Bricker, L.K. Frankel, The structure and function of the 33 kDa extrinsicprotein of photosystem II. A critical review, Photosynth. Res 56 (1998) 157–173.
[21] K. Akabori, H. Tsukamoto, J. Tsukihara, T. Nagatsuka, O. Motokawa, Y. Toyoshima,Disintegration and reconstitution of photosystem II reaction center core complex.I. Preparation and characterization of three different types of subcomplexes,Biochim. BiophysActa 932 (1988) 345–357.
[22] N. Yamaguchi, Y. Takahashi, K. Satoh, Isolation and characterization of aphotosystem II core complex depleted in the 43 kDa chlorophyll binding subunit,Plant Cell Physiol 29 (1988) 123–129.
[23] M. Rögner, D.A. Chisholm, B.A. Diner, Site-directed mutagenesis of the psbC geneof photosystem II: isolation and functional characterization of CP43-lessphotosystem II core complexes, Biochemistry 30 (1991) 5387–5395.
[24] G. Ajlani, C. Vernotte, Construction and characterization of a phycobiliprotein-lessmutant of Synechocystis PCC6803, Plant Mol. Biol 37 (1998) 577–580.
[25] M.M. Allen, Simple conditions for growth of unicellular blue-green algae on plates,J. Phycol 4 (1968) 1–4.
[26] E.G. Bligh, W.J. Dyer, A rapidmethod of total lipid extraction and purification, Can JBiochem Physiol 37 (1959) 911–917.
[27] H. Wada, N. Murata, Synechocystis PCC6803 mutants defective in desaturation offatty acids, Plant Cell Physiol 30 (1989) 971–978.
[28] J. Komenda, V. Reisinger, B. Müller, M. Dobáková, B. Granvogl, L.A. Eichacker,Accumulation of the D2 protein is a key regulatory step for assembly of thephotosystem II reaction center complex in Synechocystis PCC6803, J. Biol. Chem279 (2004) 48620–48629.
[29] J. Komenda, J. Barber, Comparison of psb0 and psbH deletion mutants of Syne-chocystis PCC6803 indicates that degradation of D1 protein is regulated by the QB
site and dependent on protein synthesis, Biochemistry 34 (1995) 9625–9631.[30] H. Schägger, G. van Jagow, Blue native electrophoresis for isolation of membrane
protein complexes in enzymatically active form, Anal. Biochem 199 (1991) 223–231.[31] J.C. Meeks, R.W. Castenholz, Growth and photosynthesis in an extreme
thermophile, Synechococcus lividus (Cyanophyta), Arch. Mikrobiol 78 (1971)25–41.
[32] I. Vass, G. Horváth, T. Herczeg, S. Demeter, Photosynthetic energy conservationinvestigated by thermoluminescence. Activation energies and half-lives ofthermoluminescence bands of chloroplasts determined by mathematical resolu-tion of glow curves, Biochim. Biophys. Acta 634 (1981) 140–152.
[33] J. Olive, G. Ajlani, C. Astier, M. Recouvreur, C. Vernotte, Ultrastructure and lightadaptation of phycobilisomemutants of Synechocystis PCC6803, Biochim. Biophys.Acta 1319 (1997) 275–282.
[34] G.Z. Shen, W.F.J. Vermaas, Chlorophyll in a Synechocystis sp. PCC-6803 mutantwithout photosystem-I and photosystem-II core complexes — evidence forperipheral antenna chlorophylls in cyanobacteria, J. Biol. Chem 269 (1994)13904–13910.
[35] T.M. Bricker, L.K. Frankel, The structure and function of CP47 and CP43 inphotosystem II, Photosynth. Res 72 (2002) 131–146.
[36] G. Schansker, SZ. Tóth, R.J. Strasser, Methylviologen and dibromothymoquinonetreatments of pea leaves reveal the role of photosystem I in the Chl a fluorescencerise OJIP, Biochim. Biophys. Acta 1706 (2005) 250–261.
[37] C. Neubauer, U. Schreiber, The polyphasic rise of chlorophyll fluorescence upononset of strong illumination; I. Saturation characteristics and partial control by thephotosystem II acceptor side, Z. Naturforsch 42c (1987) 1246–1254.
[38] U. Schreiber, C.Neubauer, C. Klughammer,Devices andmethods for room-temperaturefluorescence analysis, Philos. Trans. R. Soc. Lond., B 323 1216 (1989) 241–251.
[39] J.C. Munday, Govindjee, Light-induced changes in the fluorescence yield ofChlorophyll a in vivo. III. The dip and the peak in the fluorescence transient ofChlorella pyrenoidosa,, Biophys. J 9 (1969) 1–21.
[40] G. Schansker, SZ. Tóth, R.J. Strasser, Dark recovery of the Chl a fluorescencetransient (OJIP) after light adaptation: the qT-component of non-photochemicalquenching is related to an activated photosystem I acceptor side, Biochim.Biophys. Acta 1757 (2006) 787–797.
[41] K. Stamatakis, M. Tsimilli-Michael, G.C. Papageorgiou, Fluorescence induction inthe phycobilisome-containing cyanobacterium Synechococcus sp PCC 7942:analysis of the slow fluorescence transient, Biochim. Biophys. Acta 1767 (2007)766–772.
[42] S.Z. Tóth, G. Schansker, G. Garab, R.J. Strasser, Photosynthetic electron transportactivity in heat-treated barley leaves: the role of internal alternative electrondonors to photosystem II, Biochim. Biophys. Acta 1767 (2007) 295–305.
[43] M. Dobakova, M. Tichy, J. Komenda, Role of the PsbI protein in photosystem IIassembly and repair in the cyanobacterium Synechocystis sp. PCC6803, PlantPhysiol 145 (2007) 1681–1691.
[44] M.C. Wakeham, R.B. Sessions, M.R. Jones, P.K. Fyfe, Is there a conserved interactionbetween cardiolipin and the type II bacterial reaction center? Biophys. J 80 (2001)1395–1405.
[45] K.E. McAuley, P.K. Fyfe, J.P. Ridge, N.W. Isaacs, R.J. Cogdell, M.R. Jones, Structuraldetails of an interaction between cardiolipin and an integral membrane protein,Proc. Natl. Acad. Sci. U.S.A 96 (1999) 14706–14711.
[46] C. Weiss, W. Oppliger, G. Vergeres, R. Demel, P. Jeno, M. Horst, B. de Kruijff, G.Schatz, A. Azem, Domain structure and lipid interaction of recombinant yeastTim44, Proc. Natl. Acad. Sci. U.S.A. 96 (1999) 8890–8894.
[47] F. Jiang, M.T. Ryan, M. Schlame, M. Zhao, Z. Gu, M. Klingenberg, N. Pfanner, M.L.Greenberg, Absence of cardiolipin in the crd1 null mutant results in decreasedmitochondrial membrane potential and reduced mitochondrial function, J. Biol.Chem 275 (2000) 22387–22394.
[48] D.B.Ostrander,M. Zhang,E.Mileykovskaya,M.Rho,W.Dowhan, Lackofmitochondrialanionicphospholipids causesan inhibitionof translationof protein components of theelectron transport chain: a yeast genetic model system for the study of anionicphospholipid function in mitochondria, J. Biol. Chem 276 (2001) 25262–25272.
[49] E.G. Andrizhiyevskaya, A. Chojnicka, J.A. Bautista, B.A. Diner, R. van Grondelle1, J.P.Dekker, Origin of the F685 and F695 fluorescence in photosystem II, Photosynth.Res 84 (2005) 173–180.
[50] H. Kautsky, W. Appel, H. Amann, Chlorophyllfluorescenz und Kohlensaureassimi-lation, Biochem. Z 332 (1960) 277–292.
[51] L.N.M. Duysens, H.E. Sweers, Mechanism of the two photochemical reactions inalgae as studied by means of fluorescence, Studies on Microalgae andPhotosynthetic Bacteria, University of Tokyo Press Ed, Tokyo, 1963, pp. 353–372.

1194 H. Laczkó-Dobos et al. / Biochimica et Biophysica Acta 1777 (2008) 1184–1194
[52] N. Murata, M. Nishimura, A. Takamiya, Fluorescence of chlorophyll in photo-synthetic systems. Emission and action spectra of fluorescence-three emissionbands of chlorophyll a and the energy transfer between two pigment systems,Biochim. Biophys. Acta 126 (1966) 234–243.
[53] S. Malkin, B. Kok, Fluorescence induction studies in isolated chloroplasts. I.Number of components involved in the reaction and quantum yields, Biochim.Biophys. Acta 126 (1966) 413–432.
[54] D. Lazar, The polyphasic chlorophyll a fluorescence rise measured under highintensity of exciting light, Functional Plant. Biol 33 (2006) 9–30.
[55] I. Enami, M. Kitamura, T. Tomo, Y. Isokawa, H. Ohta, S. Katoh, Is the primary causeof thermal inactivation of oxygen evolution in spinach PSII membranes release
of the extrinsic 33 kDa protein or of Mn? Biochim. Biophys. Acta 1186 (1994)52–58.
[56] Y. Yamane, Y. Kashino, H. Koike, K. Satoh, Effects of high temperatures on thephotosynthetic systems in spinach: oxygen-evolving activities, fluorescencecharacteristics and the denaturation process, Photosynth. Res 57 (1998) 51–59.
[57] S. Demeter, C. Goussias, G. Bernat, L. Kovacs, V. Petrouleas, Participation of theG=1.9 and G=1.82 EPR forms of the semiquinone-iron complex QA−. Fe2+ ofphotosystem-II in the generation of the Q and C thermoluminescence bands,respectively, FEBS Lett 336 (1993) 352–356.
[58] Y. Isogai, Y. Yamamoto, M. Nishimura, Association of the 33 kDa polypeptide withthe 43 kDa component in photosystem II particles, FEBS Lett 187 (1985) 240–244.




















