
การดูแลสุขภาพทารกในครรภ ในระยะคลอด Intrapartum Fetal Monitor
Upload
independentCategory
view
1download
0
RP
TAa
tb
nc
D
AG(itiegfiw3fimBdhbiplfibtcn(dacprI
Kn
*UtEACsGpaPa
Neuroscience 163 (2009) 838–847
0d
OLE OF NEUROSTEROIDS IN REGULATING CELL DEATH AND
ROLIFERATION IN THE LATE GESTATION FETAL BRAINCtaio(e5uoha(tc(dttpG
ltsntw2cpacsr1
vCeaPntoisa
. YAWNO,a,b* J. J. HIRST,c M. CASTILLO-MELENDEZa
ND D. W. WALKERa
Department of Physiology, Monash University, Building 13F Welling-on Road, Clayton, Victoria 3800, Australia
Department of Obstetrics and Gynaecology, Monash University, Mo-ash Medical Centre, 246 Clayton Road, Victoria 3168, Australia
School of Biomedical Sciences, University of Newcastle, Universityrive, Callaghan, New South Wales 2308, Australia
bstract—The neurosteroid allopregnanolone (AP) is aABAergic agonist that suppresses central nervous system
CNS) activity in the adult brain, and by reducing excitotox-city is considered to be neuroprotective. A role for neuros-eroids in the developing brain, particularly in late gestation,s still debated. The aim of this study was to investigateffects on proliferation and cell death in the brain of lateestation fetal sheep after inhibition of AP synthesis usingnasteride, a 5�-reductase type 2 (5�-R2) inhibitor. Cathetersere implanted in fetal sheep at �125 days of gestation. At–4 days postsurgery, fetuses received infusions of eithernasteride (20 mg/kg/h; n�5), the AP analogue alfaxalone (5g/kg/h; n�5), or finasteride and alfaxalone together (n�5).rains were obtained at 24 h after infusion to determine celleath (apoptotic or necrotic) and cell proliferation in theippocampus and cerebellum, areas known to be suscepti-le to excitotoxic damage. Finasteride treatment significantly
ncreased apoptosis (activated caspase-3 expression) in hip-ocampal CA3 and CA1, and cerebellar molecular and granular
ayers, an effect abolished by co-infusion of alfaxalone andnasteride. Double-label immunohistochemistry showed thatoth neurons and astrocytes were caspase-3 positive. Finas-
eride treatment also increased the number of dead (pyknotic)ells in the hippocampus and cerebellum (Purkinje cells), butot when finasteride�alfaxalone was infused. Cell proliferation
Ki-67-immunoreactivity) increased after finasteride treatment;ouble-labeling showed the majority of Ki-67-positive cells werestrocytes. Thus, steroids such as AP appear to influence theonstitutive rate of apoptosis and proliferation in the hippocam-us and cerebellum of the fetal brain, and suggest an importantole for neurosteroids in the development of the brain. © 2009BRO. Published by Elsevier Ltd. All rights reserved.
ey words: allopregnanolone, neurosteroids, fetus, preg-ancy, brain injury.
Correspondence to: T. Yawno, Department of Physiology, Monashniversity, Building 13F Wellington Road, Clayton, Victoria 3800, Aus-
ralia. Tel: �61-3-9905-2356; fax: �61-3-9905-2547.-mail address: [email protected] (T. Yawno).bbreviations: ANOVA, analysis of variance; AP, allopregnanolone;A1, cornu ammonis 1; CA3, cornu ammonis 3; CNS, central nervousystem; DAB, diaminobenzidine; GABA, gamma-aminobutyric acid;FAP, glial fibrillary acidic protein; HR, heart rate; HRP, horseradisheroxidase; MAP, mean arterial pressure; MAP2, microtubule-associ-ted protein 2; NGS, normal goat serum; O2sat, oxygen saturation;
sBS, phosphate-buffered saline; PC, Purkinje cell; RIA, radioimmuno-ssay; TBS, Tris-buffered saline; 5�-R2, 5�-reductase type-2.
306-4522/09 $ - see front matter © 2009 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2009.07.009
838
onsiderable evidence now supports the use of proges-erone in reducing traumatic and ischemic injury to thedult brain (Stein, 2008). The protective effects of steroids
n the brain arise largely from the conversion of progester-ne to 5�-reduced metabolites such as allopregnanoloneAP, 5�-pregnane-3� ol, 20-one or 5�-tetrahydroprogest-rone). Progesterone is readily metabolized in the brain to�-dihydroprogesterone by 5�-reductase enzymes, partic-larly 5�-reductase type 2 (5�-R2); 5�-dihydroprogester-ne is then further reduced to AP by the enzyme 3�-ydroxysteroid oxido-reductase. Pregnane steroids suchs AP bind to a steroid-binding site on the GABA-type AGABAA) receptor complex, and by increasing chloride fluxhereby increase GABAergic receptor activity and de-rease the excitability of the central nervous system (CNS)Majewska et al., 1986). As a consequence, pregnaneerivatives of progesterone confer a degree of protectiono the brain from excess release of excitatory amino acidshat may occur following traumatic brain injury and hy-oxia-ischemia (Belelli and Lambert, 2005; Mellon andriffin, 2002).
In human pregnancy, it has been shown that highevels of AP are present in the maternal and fetal circula-ions at the time of birth (Bicikova et al., 2002). In pregnantheep the placental supply of precursors such as preg-enolone and progesterone contributes to the AP concen-rations found in the fetal brain (Crossley et al., 1997),hich increases as pregnancy progresses (Nguyen et al.,003). AP can also be synthesized either “de novo” fromholesterol by the fetal brain, or from pregnene steroidrecursors present in the fetal circulation that arise fromdrenal or placental synthesis. The relatively high AP con-entrations in the ovine fetal brain contribute to a globaluppression of fetal CNS activity and reduce the sensoryesponsiveness of the fetus to external stimuli (Nicol et al.,998, 2001).
The cerebellum and hippocampus are known to beulnerable to the effects of hypoxic–ischemic damage.erebellar abnormalities are recognized when they em-rge postnatally as motor coordination problems, includingbnormalities in gait and posture (Trouillas et al., 1997).urkinje cell (PC) damage has been linked to severaleuropathologies in the adult, such as epilepsy, Hunting-on’s disease, Alzheimer’s disease and mitochondrial dis-rders (Sarna and Hawkes, 2003). PCs receive excitatory
nputs from granule cells and this may make the PCsusceptible to glutamate excitotoxicity through excessivectivation of Ca2� permeable receptors.
Mitotic division of neurons in the human, monkey, and
heep CNS is thought to have largely ceased by the thirds reserved.

tatopstrbsaripctdco
A
Ttww(ptNhCee
S
AsNtsbcvecbbcatwtpmitfe(ev
E
FedWrd(5psci1Rwatfaatf2r
2i(Cc3bPesic0wiQSms
C
Cm
I
Idss
ctYnMpDa1
T. Yawno et al. / Neuroscience 163 (2009) 838–847 839
rimester of fetal development except in the cerebellumnd hippocampus where neurogenesis continues until af-
er birth and into adult life (Dobbing, 1974). The presencef pregnane steroids may be important to ensure appro-riate development of these brain regions, and the currenttudy was designed to determine if neurosteroid produc-ion had an effect on brain morphology and altered theates of cell proliferation and cell death (whether directly ory apoptosis) in the hippocampus and cerebellum of fetalheep in late gestation. We used the 5�-R2 inhibitor, fin-steride to block 5�-R2 activity, either without, or with,eplacement of an analogue of AP, alfaxalone. The find-ngs of the study suggest that neurosteroids have an im-ortant role in modulating cell death and constitutive rate ofell division even in the late gestation fetal brain, and thathe use of synthetic neurosteroids may influence brainevelopment when the steroidogenic capacity of the pla-enta or brain is compromised; for example, by fetal hyp-xia or when birth occurs prematurely.
EXPERIMENTAL PROCEDURES
nimals
wenty pregnant Merino-Border Leicester ewes of known gesta-ional age, carrying singleton or twin fetuses as indicated below,ere used for this study. The ewes were kept in individual cagesith free access to food and water under a 12-h light/dark cycle
lights on, 07:00 h). The use of these animals as well as therocedures performed was in accordance with the Code of Prac-
ice for the Care and Use of animals for Scientific Purposes of theational Health and Medical Research Council of Australia, andad received prior approval from the Monash University Standingommittee on Ethics and Animal Experimentation, in addition, allxperiments were conformed to international guidelines on thethical use of animals.
urgical preparation
t 125�2 days of gestation (term is approximately 147 days)urgery was performed under isoflurane (Isoflo; Abbott, Sydney,ew South Wales, Australasia) general anesthesia for implanta-
ion of fetal polyvinyl catheters. Using a mid-line abdominal inci-ion, the pregnant uterus was identified and the fetal head andoth forelegs exteriorized through a 6–8 cm incision. A polyvinylatheter was inserted into the fetal right brachial artery and ad-anced toward the heart until the tip was calculated to haventered the common carotid artery (Yawno et al., 2007). Thisatheter was used to infuse drugs or their vehicle (details givenelow) so that the solutions passed directly to the head and brainefore entering the systemic and placental circulations. A secondatheter was inserted approximately 3 cm into the left brachialrtery and was used to obtain arterial blood samples and to record
he arterial blood pressure; a catheter attached to the fetal skinas used to record the intra-amniotic pressure which was elec-
ronically subtracted from the fetal arterial pressure signal to com-ensate for pressure changes caused by uterine activity andaternal movement. The fetus was then returned to the uterus, all
ncisions were repaired, and the catheters were exteriorized fromhe maternal abdomen through a small flank incision. The ewe andetus were allowed 3–4 days of recovery before beginning thexperiment. Fetal mean arterial pressure (MAP) and heart rateHR) derived from the pulse pressure were recorded throughoutach experiment using appropriate amplifiers, as described pre-
iously (Yawno et al., 2007). (xperimental design
inasteride (20 mg/kg/h, Steraloids Inc., Newport, RI, USA), or anquivalent volume of the vehicle solution cyclodextrin (2-hy-roxypropyl-�-cyclodextrin [Sigma-Aldrich, Sydney, New Southales, Australia] in 40% w/vol distilled water) was infused at a
ate of 5 ml/h for 2 h via the right fetal brachial artery catheter. Thisose of finasteride (20 mg/kg/h) was based on a previous studyNicol et al., 2001) where it was shown to be sufficient to inhibit�-reductase enzyme activity in late gestation fetal sheep. Com-lete solubilization of finasteride and cyclodextrin was achieved byonication. The catheter was initially primed with 0.5 ml of 40%yclodextrin solution to prevent precipitation of the drug at thenterface with saline or plasma. Alfaxalone (5�-pregnane-3�-ol,1, 20-dione; obtained as the commercial formulation Alfaxan-CDTU; Jurox Pty. Ltd., Rutherford, New South Wales, Australia)as made up at a concentration of 10 mg/ml in 40% cyclodextrinnd infused at a rate of 5 mg/kg/h. Fetuses received either finas-eride (n�5) or alfaxalone (n�5, one twin) infusion for 2 h. Aurther five fetuses received both finasteride and alfaxalone for 2 ht the doses described above, and were then infused with andditional alfaxalone dose (5 mg/kg/h) for a further 3 h to counterhe fact that finasteride reduces AP concentrations in fetal bloodor many hours after cessation of a 2 h infusion (Yawno et al.,007). A further group of five ewes (two of which had twin fetuses)eceived infusion of the vehicle.
Fetal arterial blood samples were taken at �1, 0, 1, 2, 5, and4 h with respect to the time of starting each infusion, and used
mmediately for measurement of pH and oxygen saturationO2sat) using an ABL 510 blood gas analyzer (Radiometer,openhagen, Denmark). Fetal blood gas measurements wereorrected to the expected fetal temperature of approximately9 °C. The fetuses were sacrificed 24 h after the start of infusiony an i.v. injection of pentobarbitone sodium (Lethabarb, Virbacty. Ltd., Peakhurst, New South Wales, Australia) given to thewe. The fetal brain was immediately removed from the skull andagittally divided in the midline. The right hemisphere was placedn a custom-made mould shaped to fit the fetal sheep brain andoronal sections were obtained by passing a skin grafting knife at.5 cm intervals through the hemisphere. These slices, togetherith the ipsilateral hemi-section of the cerebellum, were fixed by
mmersion in 4% paraformaldehyde (ProSci Tech, Thuringowa,ueensland, Australia) for 24 h prior to embedding in paraffin.ubsequently, 10 �m sections were cut for examination of brainorphology using the immunohistochemical procedures de-
cribed below.
ortisol radioimmunoassay (RIA)
ortisol was extracted from plasma with dichloromethane andeasured by RIA as described previously (Bocking et al., 1986).
mmunohistochemistry
mmunohistochemistry was performed on 10 �m paraffin-embed-ed sections. All sections were placed on SuperFrost plus glasslides and were de-waxed in xylene and re-hydrated through aeries of dilute ethanol.
Activated caspase-3. Activated caspase-3 immunohisto-hemistry was used to identify cells undergoing apoptosis, usinghe protocol described previously (Castillo-Melendez et al., 2004;awno et al., 2007). In brief, sections were pre-blocked with 3%ormal goat serum (NGS) in phosphate-buffered saline (PBS; 0.1, pH 7.4) for 1 h and then incubated overnight with rabbitolyclonal anti-human/mouse activated caspase-3 antibody (R &
Systems, Minneapolis, MN, USA) diluted 1:1000 in 5% NGSnd 0.1% Triton X-100 in PBS. Sections were then incubated forh with a secondary biotinylated goat-anti-rabbit IgG antibody
1:200; Vector Laboratories, CA, USA). Color was visualized using
sszvTtNleta
dtahs3wnrmTitfDpci1igr
tcri
uPsPhficfh
bcfitwsat(mBBi
FarooFrttf
FaoCgba
T. Yawno et al. / Neuroscience 163 (2009) 838–847840
treptavidin horseradish peroxidase (HRP; 1:200; Amersham Bio-ciences, Buckinghamshire, England) conjugated to diaminoben-idine (DAB; Pierce Biotechnology Inc., IL, USA). Sections wereiewed at 400� magnification using a light microscope (Nikon,okyo, Japan), and activated caspase-3 positive cells were quan-
ified using Scion image software (version 4.0, Scion Corporation,ational Institutes of Health, USA). Four fields of view were ana-
yzed in each section, and two sections of each brain region werexamined. The average value for each fetus was obtained fromhese eight fields of view, and the results were averaged across allnimals (n�5) in each group.
Ki-67. Ki-67 immunohistochemistry was used to identify cellivision (proliferation). Antigen retrieval was performed by heating
he slides in citric acid buffer (0.01 M, pH 6.0) for 4�5 min usingmicrowave oven, followed by incubation for a further 30 min in
ot citric acid buffer. Sections were then rinsed in Tris-bufferedaline (TBS; 0.1 M, pH 7.6) with 0.1% Triton X-100, incubated in% hydrogen peroxide for 10 min at room temperature, incubatedith 5% NGS in TBS with 0.3% Triton X-100 for 30 min to blockonspecific binding, and then incubated overnight at 4 °C withabbit monoclonal anti-Ki-67 (1:2, Lab Vision NeoMarkers, Fre-ont, CA, USA) antibody diluted in 1:100 in 2% NGS and 0.1%riton X-100 in TBS Sections were then washed with TBS and
ncubated for 90 min at room temperature with a secondary bio-inylated goat anti-rabbit (1:500). Sections were washed with TBS,ollowed by the addition of streptavidin HRP and visualization withAB complex. Sections were viewed at 400� magnification (hip-ocampus) and 200� magnification (cerebellum) using light mi-roscopy. The number of cells that were immunoreactive for Ki-67n the cerebellum was counted from five folia on two slides to give0 fields of view, whereas in the hippocampus the number of Ki-67
mmunoreactive cells was counted in three fields of view within aiven region, on two slides per animal to give six fields of view per
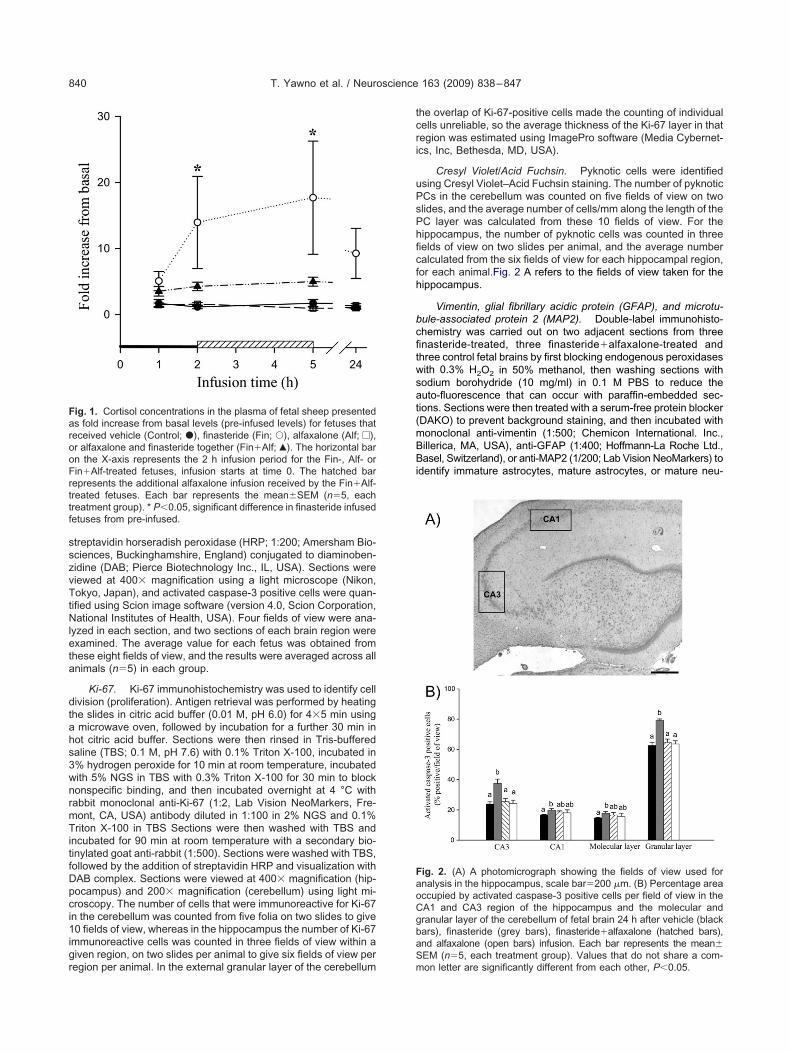
ig. 1. Cortisol concentrations in the plasma of fetal sheep presenteds fold increase from basal levels (pre-infused levels) for fetuses thateceived vehicle (Control; �), finasteride (Fin; Œ), alfaxalone (Alf; e),r alfaxalone and finasteride together (Fin�Alf; Œ). The horizontal barn the X-axis represents the 2 h infusion period for the Fin-, Alf- orin�Alf-treated fetuses, infusion starts at time 0. The hatched barepresents the additional alfaxalone infusion received by the Fin�Alf-reated fetuses. Each bar represents the mean�SEM (n�5, eachreatment group). * P�0.05, significant difference in finasteride infusedetuses from pre-infused.
egion per animal. In the external granular layer of the cerebellumSm
he overlap of Ki-67-positive cells made the counting of individualells unreliable, so the average thickness of the Ki-67 layer in thategion was estimated using ImagePro software (Media Cybernet-cs, Inc, Bethesda, MD, USA).
Cresyl Violet/Acid Fuchsin. Pyknotic cells were identifiedsing Cresyl Violet–Acid Fuchsin staining. The number of pyknoticCs in the cerebellum was counted on five fields of view on twolides, and the average number of cells/mm along the length of theC layer was calculated from these 10 fields of view. For theippocampus, the number of pyknotic cells was counted in threeelds of view on two slides per animal, and the average numberalculated from the six fields of view for each hippocampal region,or each animal.Fig. 2 A refers to the fields of view taken for theippocampus.
Vimentin, glial fibrillary acidic protein (GFAP), and microtu-ule-associated protein 2 (MAP2). Double-label immunohisto-hemistry was carried out on two adjacent sections from threenasteride-treated, three finasteride�alfaxalone-treated andhree control fetal brains by first blocking endogenous peroxidasesith 0.3% H2O2 in 50% methanol, then washing sections withodium borohydride (10 mg/ml) in 0.1 M PBS to reduce theuto-fluorescence that can occur with paraffin-embedded sec-ions. Sections were then treated with a serum-free protein blockerDAKO) to prevent background staining, and then incubated withonoclonal anti-vimentin (1:500; Chemicon International. Inc.,illerica, MA, USA), anti-GFAP (1:400; Hoffmann-La Roche Ltd.,asel, Switzerland), or anti-MAP2 (1/200; Lab Vision NeoMarkers) to
dentify immature astrocytes, mature astrocytes, or mature neu-
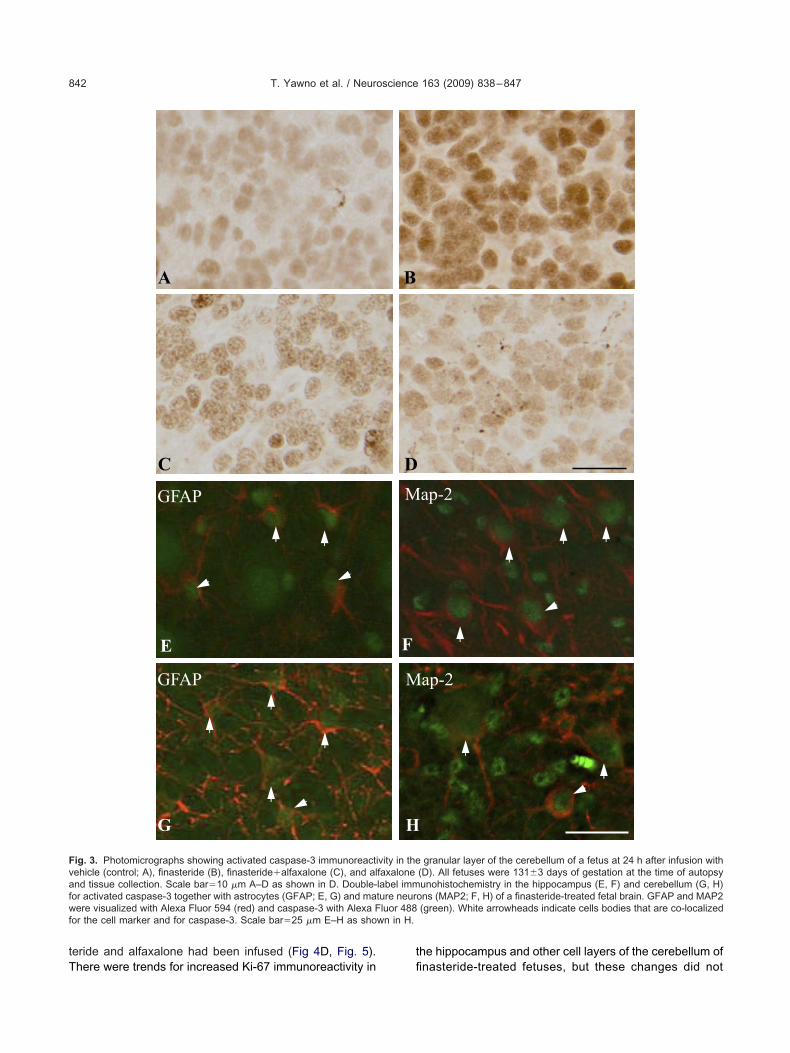
ig. 2. (A) A photomicrograph showing the fields of view used fornalysis in the hippocampus, scale bar�200 �m. (B) Percentage areaccupied by activated caspase-3 positive cells per field of view in theA1 and CA3 region of the hippocampus and the molecular andranular layer of the cerebellum of fetal brain 24 h after vehicle (blackars), finasteride (grey bars), finasteride�alfaxalone (hatched bars),nd alfaxalone (open bars) infusion. Each bar represents the mean�
EM (n�5, each treatment group). Values that do not share a com-on letter are significantly different from each other, P�0.05.
rr(AgMlZ
S
DSbprSeacia
F
Agatbffit1ict(f
atvt
E
ihlfFfapmig(cevtptrFwcAct
tlpi
Ta
p
O
M
H
T. Yawno et al. / Neuroscience 163 (2009) 838–847 841
ons, respectively. Sections were then exposed to the appropriateabbit polyclonal antibody to identify either activated caspase-31:1000) or Ki-67 (1:100). Immunoreactivity was visualized withlexa Fluor 594 goat anti-mouse (1:1000; Molecular Probes, Eu-ene, OR, USA) and Alexa Fluor 488 goat anti-rabbit (1:1000;olecular Probes, Invitrogen, Mount Waverley, Victoria, Austra-
ia), and viewed with a fluorescent microscope (Axioplan 2, Carleiss, Thornwood, NY, USA) at 400� magnification.
tatistical analysis
ata are shown as mean�SEM. All data were analyzed usingPSS software (version 9.0, SPSS Inc., Chicago, IL, USA). Fetallood gases, pH, MAP, and plasma cortisol values were com-ared between treatment groups and across time using a two-wayepeated measures analysis of variance (ANOVA) followed by atudent–Newman–Keuls post hoc test. Significance of the differ-nt counts for caspase-3 and Ki-67 immunoreactive cells wasnalyzed using one-way between groups ANOVA with post hocomparison using the least significant difference (LSD) test tondicate differences between each treatment. Significance was sett P�0.05.
RESULTS
etal systemic effects of the treatments
ll fetuses survived the treatments and were alive and inood physiological condition at the time of autopsy 24 hfter the start of each infusion. Finasteride and alfaxalone
reatments alone had no effect on fetal blood gases, acid–ase status, and MAP compared to the vehicle-treated
etuses (Table 1). In the finasteride�alfaxalone-treatedetuses the arterial pressure was slightly but significantlyncreased at 2 h after start of the combined infusion, al-hough it had returned to the normal range after 5 h (Table). HR remained unchanged throughout the entire record-
ng period for all treatment groups. Plasma cortisol con-entrations were increased at 2 and 5 h after the start ofhe finasteride infusion compared to pre-infused valuesFig. 1). A smaller, but similar trend was found in theetuses that received finasteride�alfaxalone. Infusion of
able 1. Fetal arterial blood pH, O2sat, MAP, and HR before, during anlone, or the infusion of finasteride and alfaxalone together (Fin�Alf,
Treatment –1 h �1
H Vehicle 7.35�0.01 7.3Fin 7.37�0.01 7.3Fin�Alf 7.37�0.01 7.3Alf 7.36�0.01 7.3
2sat (%) Vehicle 69.8�5.9 62Fin 66.7�9.3 61Fin�Alf 65.3�4.2 69Alf 65.5�4.2 70
AP (mm Hg) Vehicle 41.4�1.9 40Fin 39.9�1.7 42Fin�Alf 43.6�1.7 4Alf 41.4�1.2 40
R (beats/min) Vehicle 163.8�6.3 170Fin 144.8�6.7 168Fin�Alf 153.1�4.3 176Alf 154.9�2.9 146
Data shown as the mean�SEM* P�0.05, significant difference to preinfused
lfaxalone itself had no effect on plasma cortisol concen-rations (Fig. 1). The intra- and inter-assay coefficient ofariance was 9% (n�5) and 10% (n�5), respectively, andhe sensitivity of the assay was 0.42�0.02 ng/ml.
ffects of the treatments on the fetal brain
Apoptosis. The number of activated caspase-3 pos-tive cells present in the CA1 and CA3 regions of theippocampus and in the cerebellar molecular and granular
ayers was significantly increased in the finasteride-treatedetuses compared to the vehicle-infused fetuses (P�0.05;ig. 2B). When both finasteride and alfaxalone were in-
used, there was no significant difference in the number ofctivated caspase-3 expressing cells in either the hip-ocampus or the cerebellum compared to vehicle treat-ent (Fig. 2B). Examples from the cerebellum are shown
n Fig. 3, where the increase of the caspase-3 protein in theranular layer of a finasteride-treated fetus can be seenFig. 3B), and the less prominent expression produced byo-infusion of finasteride and alfaxalone (Fig. 3C), whilexpression with alfaxalone alone was no different fromehicle (Fig. 3D). Double-labeling revealed that many ofhe activated caspase-3-positive cells present in the hip-ocampus and cerebellum of the finasteride-treated fe-uses were astrocytes (GFAP-positive cells; Fig. 3E and G;espectively) and mature neurons (MAP2-positive cells;ig. 3F and H, respectively). Activated caspase-3 stainingas not present in oligodendrocytes (CNPase-positiveells) in either of these two brain regions (data not shown).lfaxalone treatment alone had no effect on activatedaspase-3 immunoreactivity compared to levels in vehiclereated animals (Figs. 2 and 3D).
Cell proliferation. The number of cells immunoposi-ive for Ki-67 was significantly increased in the granularayer of the cerebellum of finasteride-treated fetuses com-ared to untreated control fetuses (Fig. 4A and B), but this
ncrease was less, and not significant when both finas-
fusion of finasteride (Fin, n�5), alfaxalone (Alf, n�5), or vehicle (n�5)
�2 h �5 h �24 h
7.37�0.01 7.36�0.01 7.35�0.017.34�0.02 7.32�0.02 7.38�0.017.36�0.01 7.34�0.01 7.36�0.0047.37�0.004 7.35�0.01 7.35�0.00266.1�5.4 67.4�4.6 67.6�5.664.8�7.9 54.8�7.8 59.5�8.870.1�4.1 54.4�2.9 63.2�4.270.2�3.0 62.2�4.9 65.1�3.040.3�2.1 39.9�1.8 42.2�2.544.2�1.8 41.8�2.3 41.1�2.256.9�2* 44.5�2.9 41.5�1.6
42�0.9 39.6�0.9 39.6�1.2163.1�9 158.3�8.6 161.4�4.9188.5�14.5 176.6�15 165.6�8.6164.7�10 166.5�13.7 159.8�9
149.36�6.9 149.2�5.6 159.5�6
d after inn�5)
h
6�0.016�0.026�0.018�0.002.7�4.8.3�8.3.3�3.4.4�3.3.1�1.1.8�3.67�1.9.5�0.4.6�5.4.9�16.3�17.7�4
values.
tT
t
Fvafwf wn in H.
T. Yawno et al. / Neuroscience 163 (2009) 838–847842
eride and alfaxalone had been infused (Fig 4D, Fig. 5).
A
C
GFAP
E
GFAP
G
ig. 3. Photomicrographs showing activated caspase-3 immunoreactiehicle (control; A), finasteride (B), finasteride�alfaxalone (C), and alnd tissue collection. Scale bar�10 �m A–D as shown in D. Double-or activated caspase-3 together with astrocytes (GFAP; E, G) and maere visualized with Alexa Fluor 594 (red) and caspase-3 with Alexa F
or the cell marker and for caspase-3. Scale bar�25 �m E–H as sho
here were trends for increased Ki-67 immunoreactivity in fi
he hippocampus and other cell layers of the cerebellum ofap-2
ap-2
granular layer of the cerebellum of a fetus at 24 h after infusion with(D). All fetuses were 131�3 days of gestation at the time of autopsyunohistochemistry in the hippocampus (E, F) and cerebellum (G, H)
ons (MAP2; F, H) of a finasteride-treated fetal brain. GFAP and MAP2(green). White arrowheads indicate cells bodies that are co-localized
B
DM
F
M
H
vity in thefaxalonelabel immture neurluor 488
nasteride-treated fetuses, but these changes did not

rlotGfinsi(lnccvt
co
tTaccacfpbs
Tpias
FAAiiaW ell marke
T. Yawno et al. / Neuroscience 163 (2009) 838–847 843
each significance (0.05�P�0.1; Fig. 5). When double-abel immunohistochemistry was used to identify the typef cell expressing the Ki-67 protein, it was evident that inhe granular layer of the cerebellum some of the cells wereFAP-positive and some were vimentin-positive after thenasteride treatment alone (Fig. 4G and H); the Ki-67 didot co-localize with either MAP2 or CNPase (data nothown). Alfaxalone treatment alone had no effect on Ki-67mmunoreactivity in the hippocampus or granular layerFig. 4E). At this stage of fetal development, the cerebel-um shows a high number of proliferating cells in an exter-al granular layer. Finasteride treatment tended to de-rease the thickness of cell layer displaying Ki-67 positiveells (0.12�0.01 mm) compared to fetuses infused withehicle solution (0.17�0.03 mm; Fig. 4C and F), but thisrend did not quite reach significance (P�0.076).
Pyknosis. Pyknotic cells were identified as shrunkenells with acidophilic, dark purple to bright pink cytoplasm,
VimentinGFAP
HG
BA
ED
ig. 4. Photomicrographs showing Ki-67 immunoreactivity in the granu), finasteride (B), finasteride�alfaxalone (D), and alfaxalone (E). All ferrowheads indicate cells positive for Ki-67 immunoreactivity. (C) Cont
n the external granular layer. Scale bars�20 �m, A, B, D, andmmunohistochemistry in the granular layer of the cerebellum for Ki-67strocytes (vimentin; H) of a finasteride-treated fetal brain. Cell types wehite arrowheads indicate cells bodies that are co-localized for the c
ften appearing to be without a nucleus. Examples from e
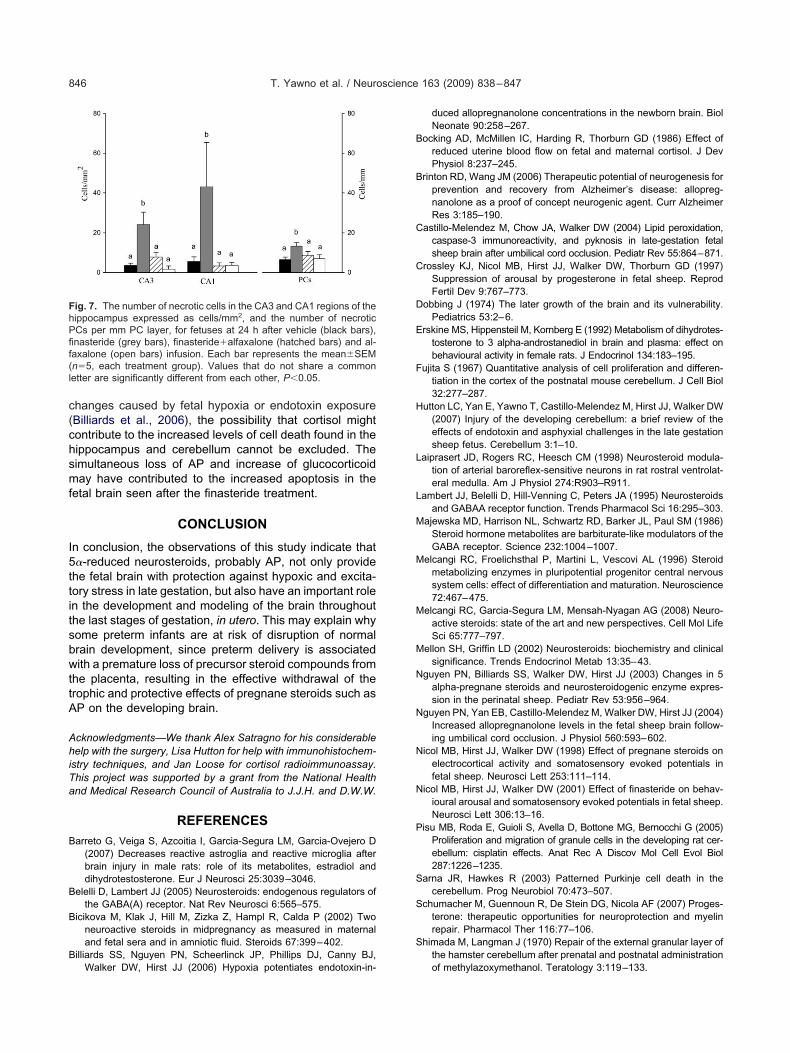
he hippocampus and cerebellum are shown in Fig. 6.here were increased numbers of pyknotic cells in the CA1nd CA3 of the hippocampus and the PC layer in theerebellum of finasteride-treated fetuses, compared toontrol fetuses (Figs. 6 and 7). When finasteride andlfaxalone were infused together, there was no signifi-ant increase in the number of pyknotic PCs. The al-axalone treatment alone did not change the number ofyknotic cells observed in the hippocampus and cere-ellum compared to the control, vehicle-infused fetuseshown in Fig. 7.
DISCUSSION
he key finding of this study was that the incidences ofyknotic and apoptotic cell death, and cell division in two
mportant areas of the developing brain, the hippocampusnd cerebellum, were altered when the enzyme 5�-R2 wasuppressed by the systemic infusion of finasteride. These
F
C
of the cerebellum of a fetus at 24 h after infusion with vehicle (control;e 131�3 days of gestation at the time of autopsy and tissue collection.asteride (F) photomicrographs showing increased Ki-67 positive cellsown by bar in E; C and F as shown by bar in F. Double-labelating cells) together with mature astrocytes (GFAP; G) and immatureized with Alexa Fluor 594 (red) and Ki-67 with Alexa Fluor 488 (green).r and for Ki-67. Scale bar�25 �m in G and H as shown in H.
lar layertuses werrol and finE as sh(proliferre visual
ffects appear to be due to the loss of AP, because co-

ichasset
pttstdA2h2
tcbctiGbptptdsstmf
s
a(acfidrgmlicocamsavidppo
sp(as(pAeac(gtpoc(tdn2ioadwi(cdcp
FCufia(l
T. Yawno et al. / Neuroscience 163 (2009) 838–847844
nfusion of the AP analogue alfaxalone prevented thesehanges. The observation that alfaxalone treatment alonead no effect on the constitutive expression of caspase-3nd Ki-67 proteins in the hippocampus and cerebellumuggests that the effect exerted by steroids is alreadyufficient, and perhaps maximal, given that the level of anndogenous neurosteroid such as AP is relatively high in
he fetal brain.During late fetal development in sheep, the hippocam-
us and cerebellum are vulnerable to asphyxia- and infec-ion-related damage (Castillo-Melendez et al., 2004; Hut-on et al., 2007). We have previously shown that pregnaneteroids have a role in protecting the fetal brain from exci-otoxic damage arising from the effects of fetal asphyxia,ue in part to rapid upregulation of 5�-R2 expression andP synthesis (Nguyen et al., 2003, 2004; Yawno et al.,007). Suppression of this AP response leads to greaterypoxic/ischemic damage in the fetal brain (Nguyen et al.,004).
The current study extends these findings by showinghat neurosteroids influence the processes that determineell fate in at least two important areas of the developingrain, the hippocampus and cerebellum. Thus, basal oronstitutive neurosteroid synthesis may be important forhe final stages of brain development in utero, particularlyn relation to neurons and astrocytes. It may be thatABAergic activity provides a trophic drive for late fetalrain development, at least for some aspects of hippocam-al and cerebellar development. The finding that finas-eride affected the CA3 (but not CA1) region of the hip-ocampus, and the granular (but not molecular) layer ofhe cerebellum, suggests that these brain regions are quiteifferent in their sensitivity to the effects of removing neuro-teroids. Differences in GABAA receptor sub-unit expres-ion and/or in the local synthesis of AP may exist betweenhe CA1 and CA3 regions, but this is speculation at theoment as we have been unable to find evidence, either
rom our own or other studies, to support this idea.Previous studies have shown that progesterone and
ig. 5. The number of Ki-67 positive cells (per mm2) in the CA1 andA3 region of the hippocampus and the white matter tract and gran-lar layer of the cerebellum of fetal brain 24 h after vehicle (black bars),nasteride (grey bars), finasteride�alfaxalone (hatched bars), andlfaxalone (open bars) infusion. Each bar represents the mean�SEMn�5, each treatment group). Values that do not share a commonetter are significantly different from each other, P�0.05.
ome of its metabolites exert protective effects in neurons c
nd glia following brain injury by preventing cell deathwhether necrotic or apoptotic), minimizing inflammationnd cerebral edema, and enhancing neurogenerative pro-esses (Brinton and Wang, 2006). Our observations thatnasteride treatment increased both neuronal and glial celleath are consistent with these studies, and support a keyole for neurosteroids in controlling the vulnerability of therowing brain to oxidative and metabolic stresses. Further-ore, we found that PCs in the cerebellum appear to die
argely as a result of necrosis, based on the increasedncidence of pyknotic cells. In contrast, cerebellar granuleells were frequently apoptotic but rarely necrotic, as alsobserved in the newborn piglet brain following transienterebral hypoxia–ischemia (Yue et al., 1997). In the CA1nd CA3 regions of the hippocampus, finasteride treat-ent increased both activated caspase-3 and pyknotic
taining of mature neurons, and there was also increasedpoptotic cell death of astrocytes. Together these obser-ations support the concept that neurosteroids may be
mportant for the regulation of normal, programmed celleath during late gestation, and raise the possibility thaterturbations of pregnane steroid concentrations duringregnancy could result in disturbances to fetal brain devel-pment in late gestation.
Alfaxalone is a highly selective agonist for the neuro-teroid binding site on the GABAA receptor with a similarotency to AP (Lambert et al., 1995). In contrast to APwhich can be converted back to 5�–dihydroprogesteronend potentially interact with the progesterone receptor—ee below), alfaxalone acts only at the GABAA receptorLambert et al., 1995). Thus, alfaxalone effectively re-laces the GABAergic activity arising from the presence ofP which was removed by the finasteride treatment. How-ver, effects mediated via progesterone and testosteronesre also possible. 5�-Reductases catalyze the irreversibleonversion of progesterone to 5�-dihydroprogesteronesee Melcangi et al., 2008 for review), and 5�-dihydropro-esterone is also likely to be reduced by the finasteridereatment. 5�-Dihydroprogesterone is an agonist at therogesterone receptor, but is less potent than progester-ne, and reduced progesterone receptor stimulation mayontribute to the action of finasteride on proliferationSchumacher et al., 2007). Inhibition of 5�-reductase ac-ivity would reduce the conversion of testosterone to 5�-ihydrotestosterone and, as 5�-dihydrotestosterone haseuroprotective effects in the adult brain (Barreto et al.,007); some effects may arise from changes in this steroid
n the fetal brain. This may explain the only partial reversalf the effect of finasteride on Ki-67 immunoreactivity bylfaxalone observed in this study. In addition, 5�-dihy-rotestosterone may be converted to 3�-androstanediolhich may modulate the GABAA receptor, although little
nformation is available on its efficacy compared to APErskine et al., 1992). It is difficult to determine the relativeontribution of these alternate steroid pathways in theeveloping brain, and further studies are required to elu-idate the potentially synergistic roles of AP, 5�-dihydro-rogesterone, and 5�-dihydrotestosterone in regulating
ell proliferation in the late gestation fetal brain.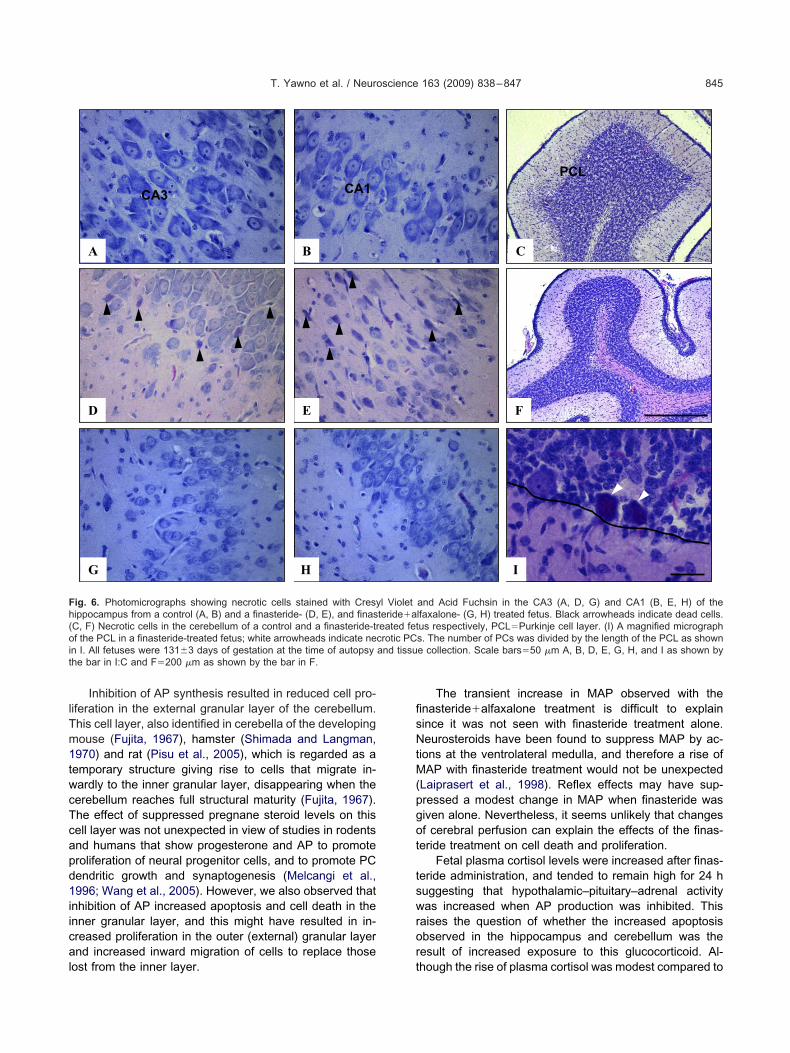
lTm1twcTcapd1iical
fisNtM(pgot
tswror
Fh(oit
T. Yawno et al. / Neuroscience 163 (2009) 838–847 845
Inhibition of AP synthesis resulted in reduced cell pro-iferation in the external granular layer of the cerebellum.his cell layer, also identified in cerebella of the developingouse (Fujita, 1967), hamster (Shimada and Langman,970) and rat (Pisu et al., 2005), which is regarded as aemporary structure giving rise to cells that migrate in-ardly to the inner granular layer, disappearing when theerebellum reaches full structural maturity (Fujita, 1967).he effect of suppressed pregnane steroid levels on thisell layer was not unexpected in view of studies in rodentsnd humans that show progesterone and AP to promoteroliferation of neural progenitor cells, and to promote PCendritic growth and synaptogenesis (Melcangi et al.,996; Wang et al., 2005). However, we also observed that
nhibition of AP increased apoptosis and cell death in thenner granular layer, and this might have resulted in in-reased proliferation in the outer (external) granular layernd increased inward migration of cells to replace those
BA
HG
ED
CA3 CA1
ig. 6. Photomicrographs showing necrotic cells stained with Cresyippocampus from a control (A, B) and a finasteride- (D, E), and finasC, F) Necrotic cells in the cerebellum of a control and a finasteride-trf the PCL in a finasteride-treated fetus; white arrowheads indicate ne
n I. All fetuses were 131�3 days of gestation at the time of autopsyhe bar in I:C and F�200 �m as shown by the bar in F.
ost from the inner layer. t
The transient increase in MAP observed with thenasteride�alfaxalone treatment is difficult to explainince it was not seen with finasteride treatment alone.eurosteroids have been found to suppress MAP by ac-
ions at the ventrolateral medulla, and therefore a rise ofAP with finasteride treatment would not be unexpected
Laiprasert et al., 1998). Reflex effects may have sup-ressed a modest change in MAP when finasteride wasiven alone. Nevertheless, it seems unlikely that changesf cerebral perfusion can explain the effects of the finas-eride treatment on cell death and proliferation.
Fetal plasma cortisol levels were increased after finas-eride administration, and tended to remain high for 24 huggesting that hypothalamic–pituitary–adrenal activityas increased when AP production was inhibited. This
aises the question of whether the increased apoptosisbserved in the hippocampus and cerebellum was theesult of increased exposure to this glucocorticoid. Al-
F
I
C
PCL
and Acid Fuchsin in the CA3 (A, D, G) and CA1 (B, E, H) of thelfaxalone- (G, H) treated fetus. Black arrowheads indicate dead cells.us respectively, PCL�Purkinje cell layer. (I) A magnified micrographs. The number of PCs was divided by the length of the PCL as showne collection. Scale bars�50 �m A, B, D, E, G, H, and I as shown by
l Violetteride�aeated fetcrotic PCand tissu
hough the rise of plasma cortisol was modest compared to
c(chsmf
I5ttitsbwttA
AhiTa
B
B
B
B
B
B
C
C
D
E
F
H
L
L
M
M
M
M
N
N
N
N
P
S
S
S
FhPfif(l
T. Yawno et al. / Neuroscience 163 (2009) 838–847846
hanges caused by fetal hypoxia or endotoxin exposureBilliards et al., 2006), the possibility that cortisol mightontribute to the increased levels of cell death found in theippocampus and cerebellum cannot be excluded. Theimultaneous loss of AP and increase of glucocorticoiday have contributed to the increased apoptosis in the
etal brain seen after the finasteride treatment.
CONCLUSION
n conclusion, the observations of this study indicate that�-reduced neurosteroids, probably AP, not only providehe fetal brain with protection against hypoxic and excita-ory stress in late gestation, but also have an important rolen the development and modeling of the brain throughouthe last stages of gestation, in utero. This may explain whyome preterm infants are at risk of disruption of normalrain development, since preterm delivery is associatedith a premature loss of precursor steroid compounds from
he placenta, resulting in the effective withdrawal of therophic and protective effects of pregnane steroids such asP on the developing brain.
cknowledgments—We thank Alex Satragno for his considerableelp with the surgery, Lisa Hutton for help with immunohistochem-
stry techniques, and Jan Loose for cortisol radioimmunoassay.his project was supported by a grant from the National Healthnd Medical Research Council of Australia to J.J.H. and D.W.W.
REFERENCES
arreto G, Veiga S, Azcoitia I, Garcia-Segura LM, Garcia-Ovejero D(2007) Decreases reactive astroglia and reactive microglia afterbrain injury in male rats: role of its metabolites, estradiol anddihydrotestosterone. Eur J Neurosci 25:3039–3046.
elelli D, Lambert JJ (2005) Neurosteroids: endogenous regulators ofthe GABA(A) receptor. Nat Rev Neurosci 6:565–575.
icikova M, Klak J, Hill M, Zizka Z, Hampl R, Calda P (2002) Twoneuroactive steroids in midpregnancy as measured in maternaland fetal sera and in amniotic fluid. Steroids 67:399–402.
illiards SS, Nguyen PN, Scheerlinck JP, Phillips DJ, Canny BJ,
ig. 7. The number of necrotic cells in the CA3 and CA1 regions of theippocampus expressed as cells/mm2, and the number of necroticCs per mm PC layer, for fetuses at 24 h after vehicle (black bars),nasteride (grey bars), finasteride�alfaxalone (hatched bars) and al-axalone (open bars) infusion. Each bar represents the mean�SEMn�5, each treatment group). Values that do not share a commonetter are significantly different from each other, P�0.05.
Walker DW, Hirst JJ (2006) Hypoxia potentiates endotoxin-in-
duced allopregnanolone concentrations in the newborn brain. BiolNeonate 90:258–267.
ocking AD, McMillen IC, Harding R, Thorburn GD (1986) Effect ofreduced uterine blood flow on fetal and maternal cortisol. J DevPhysiol 8:237–245.
rinton RD, Wang JM (2006) Therapeutic potential of neurogenesis forprevention and recovery from Alzheimer’s disease: allopreg-nanolone as a proof of concept neurogenic agent. Curr AlzheimerRes 3:185–190.
astillo-Melendez M, Chow JA, Walker DW (2004) Lipid peroxidation,caspase-3 immunoreactivity, and pyknosis in late-gestation fetalsheep brain after umbilical cord occlusion. Pediatr Rev 55:864–871.
rossley KJ, Nicol MB, Hirst JJ, Walker DW, Thorburn GD (1997)Suppression of arousal by progesterone in fetal sheep. ReprodFertil Dev 9:767–773.
obbing J (1974) The later growth of the brain and its vulnerability.Pediatrics 53:2–6.
rskine MS, Hippensteil M, Kornberg E (1992) Metabolism of dihydrotes-tosterone to 3 alpha-androstanediol in brain and plasma: effect onbehavioural activity in female rats. J Endocrinol 134:183–195.
ujita S (1967) Quantitative analysis of cell proliferation and differen-tiation in the cortex of the postnatal mouse cerebellum. J Cell Biol32:277–287.
utton LC, Yan E, Yawno T, Castillo-Melendez M, Hirst JJ, Walker DW(2007) Injury of the developing cerebellum: a brief review of theeffects of endotoxin and asphyxial challenges in the late gestationsheep fetus. Cerebellum 3:1–10.
aiprasert JD, Rogers RC, Heesch CM (1998) Neurosteroid modula-tion of arterial baroreflex-sensitive neurons in rat rostral ventrolat-eral medulla. Am J Physiol 274:R903–R911.
ambert JJ, Belelli D, Hill-Venning C, Peters JA (1995) Neurosteroidsand GABAA receptor function. Trends Pharmacol Sci 16:295–303.
ajewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM (1986)Steroid hormone metabolites are barbiturate-like modulators of theGABA receptor. Science 232:1004–1007.
elcangi RC, Froelichsthal P, Martini L, Vescovi AL (1996) Steroidmetabolizing enzymes in pluripotential progenitor central nervoussystem cells: effect of differentiation and maturation. Neuroscience72:467–475.
elcangi RC, Garcia-Segura LM, Mensah-Nyagan AG (2008) Neuro-active steroids: state of the art and new perspectives. Cell Mol LifeSci 65:777–797.
ellon SH, Griffin LD (2002) Neurosteroids: biochemistry and clinicalsignificance. Trends Endocrinol Metab 13:35–43.
guyen PN, Billiards SS, Walker DW, Hirst JJ (2003) Changes in 5alpha-pregnane steroids and neurosteroidogenic enzyme expres-sion in the perinatal sheep. Pediatr Rev 53:956–964.
guyen PN, Yan EB, Castillo-Melendez M, Walker DW, Hirst JJ (2004)Increased allopregnanolone levels in the fetal sheep brain follow-ing umbilical cord occlusion. J Physiol 560:593–602.
icol MB, Hirst JJ, Walker DW (1998) Effect of pregnane steroids onelectrocortical activity and somatosensory evoked potentials infetal sheep. Neurosci Lett 253:111–114.
icol MB, Hirst JJ, Walker DW (2001) Effect of finasteride on behav-ioural arousal and somatosensory evoked potentials in fetal sheep.Neurosci Lett 306:13–16.
isu MB, Roda E, Guioli S, Avella D, Bottone MG, Bernocchi G (2005)Proliferation and migration of granule cells in the developing rat cer-ebellum: cisplatin effects. Anat Rec A Discov Mol Cell Evol Biol287:1226–1235.
arna JR, Hawkes R (2003) Patterned Purkinje cell death in thecerebellum. Prog Neurobiol 70:473–507.
chumacher M, Guennoun R, De Stein DG, Nicola AF (2007) Proges-terone: therapeutic opportunities for neuroprotection and myelinrepair. Pharmacol Ther 116:77–106.
himada M, Langman J (1970) Repair of the external granular layer ofthe hamster cerebellum after prenatal and postnatal administration
of methylazoxymethanol. Teratology 3:119–133.
S
T
W
Y
Y
T. Yawno et al. / Neuroscience 163 (2009) 838–847 847
tein DG (2008) Progesterone exerts neuroprotective effects afterbrain injury. Brain Res Rev 57:386–397.
rouillas P, Takayanagi T, Hallett M, Currier RD, Subramony SH, WesselK, Bryer A, Diener HC, Massaquoi S, Gomez CM, Coutinho P, BenHamida M, Campanella G, Filla A, Schut L, Timann D, Honnorat J,Nighoghossian N, Manyam B (1997) International Cooperative AtaxiaRating Scale for pharmacological assessment of the cerebellar syn-drome. The Ataxia Neuropharmacology Committee of the World Fed-eration of Neurology. J Neurol Sci 145:205–211.
ang JM, Johnston PB, Ball BG, Brinton RD (2005) The neurosteroid
allopregnanolone promotes proliferation of rodent and human neu-ral progenitor cells and regulates cell-cycle gene and proteinexpression. J Neurosci 25:4706–4718.
awno T, Yan EB, Walker DW, Hirst JJ (2007) Inhibition of ne-urosteroid synthesis increases asphyxia-induced brain injuryin the late gestation fetal sheep. Neuroscience 146:1726 –1733.
ue X, Mehmet H, Penrice J, Cooper C, Cady E, Wyatt JS, Re-ynolds EO, Edwards AD, Squier MV (1997) Apoptosis andnecrosis in the newborn piglet brain following transient ce-rebral hypoxia-ischaemia. Neuropathol Appl Neurobiol 23:
16 –25.(Accepted 5 July 2009)(Available online 8 July 2009)
Copyright © 2022 FDOKUMEN




















