![Synthesis and anti-HCMV activity of 1-[ω-(phenoxy)alkyl]uracil derivatives and analogues thereof](https://static.fdokumen.com/doc/165x107/6343875247e02623e9066ff7/synthesis-and-anti-hcmv-activity-of-1-o-phenoxyalkyluracil-derivatives-and.jpg)
Synthesis and anti-HCMV activity of 1-[ω-(phenoxy)alkyl]uracil derivatives and analogues thereof
Upload
independentCategory
view
1download
0
DNA Repair 3 (2004) 1579–1590
Role of mismatch-specific uracil-DNA glycosylase inrepair of 3,N4-ethenocytosine in vivo
Juan Juradoa,1, Agnieszka Maciejewskab, Joanna Krwawiczb,Jacques Lavala, Murat K. Saparbaeva,∗
a Groupe “Reparation de l’AND”, CNRS UMR 8113, LBPA-ENS Cachan, Institut Gustave Roussy,39 rue Camille Desmoulins, 94805 Villejuif Cedex, France
b Institute of Biochemistry and Biophysics, Polish Academy of Sciences, 02-106 Warsaw, Poland
Received 30 March 2004Available online 29 July 2004
Abstract
4
m eneratedm lementst tatorpn in vitrow of thee .Ct tion aren hanem©
K
oBIgp
U
und,stic
ctsliancies
s
bo-d 2-
1d
The 3,N -ethenocytosine (�C) residue might have biological role in vivo since it is recognized and efficiently excised in vitro by theE. coliismatch-specific uracil-DNA glycosylase (MUG) and the human thymine-DNA glycosylase (hTDG). In the present work we have gugdefective mutant ofE. coliby insertion of a kanamycin cassette to assess the role of MUG in vivo. We show that human TDG comp
he enzymatic activity of MUG when expressed in amugmutant. The�C-DNA glycosylase defective strain did not exhibit spontaneous muhenotype and did not show unusual sensitivity to any of the following DNA damaging treatments: methylmethanesulfonate,N-methyl-N′-itro-N-nitrosoguanidine, ultraviolet light, H2O2, paraquat. However, plasmid DNA damaged by 2-chloroacetaldehyde treatmentas inactivated at a greater rate in amugmutant than in wild-type host, suggesting that MUG is required for the in vivo processingthenobases. In addition, 2-chloroacetaldehyde treatment induces preferentially G·C → C·G and A·T → T·A transversions inmugmutantomparison of the mutation frequencies induced by the site-specifically incorporated�C residue inE. coli wild-type versusmug indicates
hat MUG repairs more than 80% of�C residues in vivo. Furthermore, the results show that nucleotide excision repair and recombinaot involved in the processing of�C in E. coli. Based on the mutagenesis data we suggest that�C may be less toxic and less mutagenic txpected. The increased spontaneous mutation rate for G·C → A·T transition in theung mugdouble mutant as compared to the singleungutant suggest that MUG may be a back-up repair enzyme to the classic uracil-DNA glycosylase.2004 Elsevier B.V. All rights reserved.
eywords: Vinyl chloride; Lipid peroxidation; Exocyclic DNA adducts; Ethenobases; Ethenocytosine; Base excision repair; DNA glycosylase
Abbreviations:�, etheno; �C, 3,N4-ethenocytosine;�A, 1,N6-ethen-adenine;N2,3-�G, N2,3-ethenoguanine; 1,N2-�G, 1,N2-ethenoguanine;ER, base excision repair; AlkA,E. coli3-methyladenine-DNA glycosylase
I; MUG, mismatch-specific uracil-DNA glycosylase; UNG, uracil-N-DNAlycosylase; hTDG, human thymine-DNA glycosylase; M13G*1, bacterio-hage vector
∗ Corresponding author. Tel.: +33 1 42115404; fax: +33 1 42115276.E-mail address:[email protected] (M.K. Saparbaev).
1 Present address: Departamento de Bioquımica y Biologıa Molecular,niversidad de Cordoba, 14071 Cordoba, Spain.
1. Introduction
Ethyl carbamate, a widespread environmental compoand vinyl chloride, a chemical carcinogen used in plamanufacture[1] induce the formation of exocyclic adduin DNA [2,3]. These compounds are oxidized in mammacells by cytochrome P450 to reactive electrophilic spewhich react with bases in RNA and DNA to form�-bridgedadducts[4–6]. These modifications include�-adducts such a1,N6-ethenoadenine (�A), 3,N4-ethenocytosine (�C), N2,3-ethenoguanine (�G), and 1,N2-ethenoguanine (1,N2-�G) thatare produced in DNA by reaction with two oxidized metalites of vinyl chloride, namely 2-chloroacetaldehyde an
568-7864/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.dnarep.2004.06.012

1580 J. Jurado et al. / DNA Repair 3 (2004) 1579–1590
chloroethylene oxide[7,8]. Furthermore, the generation ofexocyclic DNA adducts by products of membrane lipidsperoxidation has been demonstrated in vivo[9,10]. The �-adducts are ubiquitous and highly variable background levelsof �A and�C were found in DNA from tissues under physi-ological conditions. It was shown that the level of�C presentin 10 different human liver DNA samples averaged 28± 9�Cper 108 bases[11] whereas in leukocytes, pancreas and colonDNA samples isolated from healthy volunteers it ranged from0.1 to 11�C per 108 parent bases[12]. The mutagenic speci-ficity of exocyclic DNA lesions has been studied. InE. coliand simian kidney cells,�C mostly produces�C·G → A·Ttransversions and�C·G→ T·A transitions[13,14]. For singlestranded shuttle vector containing a single�C residue the tar-geted mutation frequency yield is 2% inE. coli, 32% in SOS-inducedE. coli cells, and 81% in simian kidney cells[14].
Since,�-adducts are highly mutagenic they should be re-moved from DNA. An enzymatic activity repairing�C wasidentified and partially purified from human cells and it wasshown to be different from the ANPG protein[15] but itwas not identified at the molecular level, later, we showedthat the human mismatch-specific thymine-DNA glycosylase(hTDG), removes�C residues by a DNA glycosylase activity[16]. In addition to hTDG, two human enzymes that excise�C have been identified: methyl-CpG binding domain pro-t alut ch-si mo-l Gm ow-e rdsd esm tp[ n�e thv ner nt yde rtanti wth[ thatM ores UGi
gt e re-s NAdi
2. Materials and methods
2.1. Bacterial strains
All bacterial genetics procedures were performed as de-scribed[25]; these included preparation of media and P1transduction. The rich medium was the LB broth, supple-mented when required with either 100�g/ml ampicillin,or 50�g/ml kanamycine, or 20�g/ml streptomycin and20�g/ml spectinomycin. The M9 minimal medium was sup-plemented with thiamine.E. coli cells were transformed byDNA electroporation as described[26].
The E. coli K12 strains used in this study are listed inTable 1. Original Miller’s strain CC101–CC106 are deriva-tives of P90C[ara, �(lac proB)XIII ] from laboratory stock,each carrying a differentlacZ allele on an F′(lacI−, Z−proB+) plasmid and, therefore, exhibiting the Lac− pheno-type. These strains enable the detection of each of the sixpossible base substitutions (two transitions and four transver-sions) by measuring the number of Lac+ revertants[27]. Typeof the base substitutions in the F′ �-galactosidase gene lead-ing to the Lac+ phenotype are listed inTable 2.
Strain AM100 was obtained by curing episome fromCC111 with acridine orange[25]. AM200 is AM100 deriva-tive, butmug, and constructed by P1 transduction of MS50,a es ofs at-i ors
2
ris),i atp -Gd ,4e -G 157o ryo ase.T as�p E,S [3 A.,O . The[ sh site�s
inr ector[ .D NA
ein (MBD4/MED1) [17] and single-strand monofunctionracil-DNA glycosylase (SMUG1)[18]. TheE. colienzyme
hat excises�C was previously identified as a mismatpecific uracil-DNA glycosylase (MUG protein)[16]. MUG
s a small 18.6 kDa protein (168 aminoacids) and is the hoogue of the hTDG protein[19]. It was proposed that MU
ay act as a back-up enzyme for the UNG protein, hver, comparison of the catalytic efficiency of MUG towaifferent substrates shows that�C was excised 5–50 timore efficiently than uracil from G·U pair suggesting tharimary function of MUG could be the repair of�C in vivo
16,20]. MUG is able to remove various�-bases other thaC including: 1,N2-�G [21] and 8-(hydroxymethyl)-3,N4-thenocytosine[22]. Furthermore, MUG could excise wiery low efficiency�A, hypoxanthine and 5-hydroxycytosiesidues when present in DNA[20]. Previously, it was showhatE. coli strain deficient in�C-DNA glycosylase activitid not exhibit spontaneous mutator phenotype[23]. How-ver, later it has been suggested that MUG may be impo
n avoiding mutations during stationary phase of cell gro24]. In agreement with our previous data it was shownUG is a majorE. coli DNA glycosylase responsible f
xcision of�C [23]. However, neither the sensitivity ofmugtrain towards DNA damaging agents nor the role of Mn the repair of�C in vivo has been investigated.
In the present study we have constructed a set of�C-DNAlycosylase (mug) deficient mutants ofE. coli K12 allowing
he determination of all possible base substitutions. Thponse of the mutant strains with respect to a variety of Damaging agents and the role of MUG in repair of�C and U
n vivo are described.
nd selection for kanamycine resistance. The final seritrains carrying F′101–F′106 plasmids were obtained by mng their F− progenitors (AM200) with CC101–CC106 dontrains, followed by Pro+ selection on minimal medium.
.2. Oligonucleotides and plasmids
All oligonucleotides were purchased from Genset (Pancluding: 34 mer oligonucleotide containing thymineosition 19, T d(CGGTATCCACCAGGTCATTAATAACATGAAGCC), modified 34 mer oligonucleotide�C34(AAATACATCGTCACCTGGGXCATGTTGCAGATCC)1 mer oligonucleotide M13-�C with phosphate on the 5′-nd d(pCTATTACGCCAGCTGGXAAATGGGGGATGTCTGCAAGGCG) complementary to positions 117–f the LacZ sequence where X is�C and complementaligonucleotides, containing dG opposite a modified bhe resulting duplex oligonucleotides are referred toC·G. The oligonucleotides were labelled at 5′-end by T4olynucleotide kinase (New England Biolabs, OZYMaint Quentin Yvelines, France) in the presence of�-
2P]-ATP (4500 Ci/mmol, ICN Pharmaceuticals, S.rsay, France) as recommended by the manufacturers
32P]-labeled oligonucleotide containing the�C residue waybridized with the oligonucleotide containing dG oppoC at a 1:2 molar ratio by heating at 90◦C for 3 min thenlowly cooled down.
The 1.8 kbPvuII-DNA fragment containing kanamycesistance gene was obtained from pLG388 plasmid v28]. The pHP45� plasmid vector[29] was a gift from Drick d’Ari (Institute Jacques Monod, Paris). The 2.0 kb D

J. Jurado et al. / DNA Repair 3 (2004) 1579–1590 1581
Table 1Bacterial strains
Strain Genotype Source of derivation
AB1157 thr-1 leu-6 proA2 his-4 argE3 thi-1 lacYI galK2 ara-14 xyl-5 mtl-1 tsx-33rpsL31 supE44
Laboratory stock
GC4803 As AB1157 butalkA tagA Laboratory stockJC7623 thr-1 leu-6 proA2 his-4 argE3 thi-1 lacY1 galK2 ara-14 xyl-5 metl-1 tsx-33
rpsL31 supE44 recB21 rec22 sbcB15 sbcC201Laboratory stock
MS20 As JC7623 butmug::Knr This studyMS30 As JC7623 butung::Spcr This studyCC101-CC106 P90C [ara∆(lacproB)XIII ] carrying anF′lacI-Z–proB+ J.H. Miller University of California, USAAM100 As CC101–CC106 but without F′ This studyAM200 As AM100 butmug::Knr This studyAM201-AM206 As CC101–CC106 butmug::Knr This studyCC102 P90C [ara∆(lacproB)XIII ] carrying anF′lacI-Z–proB+ episome
(G·C → A·T)J.H. Miller University of California, USA
MS1020 As CC102 butmug-1::Knr P1(MS20)× CC102→ KnR, this studyMS1021 As CC102 butung-1::Spcr P1(MS30)× CC102→ SpcR, this studyMS1022 As CC102 butmug-1::Knr ung-1::Spcr P1(MS20)× MS1021→ KnR, this studyJM105 F′ traD36 lacIq∆(lacZ)M15 proA+B+/thi rpsL(Strr)endA sbcB15 sbcC201
hsdR4(rK− mK+)∆(lac-proAB)
Laboratory stock
MS50 As JM105 butmug-1::Knr P1(MS20)× JM105→ KnR, this studyMS51 As JM105 butmutL218::Tn10 P1(NR9161)× JM105→ Tetr, this studyMS52 As JM105 butmutS215::Tn10 P1(NR9162)× JM105→ Tetr, this studyMS53 As JM105 butsodA sodB This studyBH430 As JM105 butuvrA::Knr Laboratory stockJM109 F′ traD36 lacIq∆(lacZ)M15 proA+B+/e14−(McrA−)∆(lac-proAB)thi
gyrA96(Nalr)endA1 hsdR17(rK− mK+) relA1 supE44 recA1
Laboratory stock
NR9161 hsdR−hsdM+araD ∆(ara-leu)∆(lac-IZOPY)galU galK strAmutL218::Tn10
R.M. Schaaper National Institute of EnvironmentalHealth Sciences, USA
NR9162 hsdR− hsdM+araD ∆(ara-leu)∆(lac-IZOPY)galU galK strAmutS215::Tn10
R.M. Schaaper National Institute of EnvironmentalHealth Sciences, USA
fragment containing spectinomycin resistance gene (Spcr)was generated from pHP45� plasmid DNA after digestionby SmaI. The DNA fragments were isolated from agarosegel using the QIAquick Gel Extraction Kit (QIAGEN S.A.France, Courtaboeuf Cedex, France). Double-stranded DNAof the bacteriophage vector, M13G*1, was kindly providedby Dr. Thomas Reid (National Institute for OccupationalSafety and Health, Cincinnati, OH, USA). Single-strandedM13G*1 template was purified fromE. coli JM105 strainas described[30]. The plasmid DNA pT7-hTDG containing
Table 2Frequency of CAA-induced Lac+ revertants
Specificity forLac+ reversion
Mutation frequencya (×10−8) ± S.D. Ratio CAAinducedmug/wild-typeWild-type (CC101–CC106) mug(AM201–AM206)
SpontaneousLac+ reversion
CAA-inducedLac+ reversionb
Ratio induced/spontaneous
SpontaneousLac+ reversion
CAA-inducedLac+ reversionb
Ratio induced/spontaneous
AT → CG 0.29± 0.04 0.17± 0.24 0.59 ns 0.93± 1.25 4.43± 3.80 4.76 ns 26.06GC→ AT 0.29± 0.17 10.67± 3.00 36.79** 0.37± 0.38 67.20± 42.40 210.0*** 6 .30GC→ CG 0.10± 0.10 1.00± 1.86 10 ns 0.015± 0.017 76.22± 9.45 5081.33*** 76 .22GC→ TA 0.51± 0.52 28.88± 5.43 56.63*** 0.41 ± 0.12 74.61± 67.50 180.49* 2.58AT → TA 2.34± 2.15 2.42± 3.16 1.03 ns 0.13± 0.04 24.30± 17.30 186.92** 10.04AT → GC 0.013± 0.023 0.027± 0.055 2.08 ns 0.0062± 0.012 0.028± 0.025 4.52 ns 1.04
a Frequency numbers are the average± S.D. of at least three independent experiments. The frequencies of Rifr background mutations in untreated cultures(×108) were in the range of: CC101–106, 3.1–8.8; AM201–206, 1.8–8.9. Statistical significance of CAA-induced reversion was evaluated using two-tailedS
the TDG cDNA, coding for the human thymine-DNA gly-cosylase, was provided by Dr. Jiric¸ny (Institute for MedicalRadiobiology, Zurich, Switzerland)[31].
2.3. PCR amplification and recombinant plasmidconstruction
The DNA fragment containing the genuine MUG genewas obtained by PCR amplification usingE. coli wild-typegenomic DNA with the following oligonucleotide primers
tudent’s test (∗∗∗P < 0.001;∗∗P < 0.01;∗P < 0.05 andnsP > 0.05).
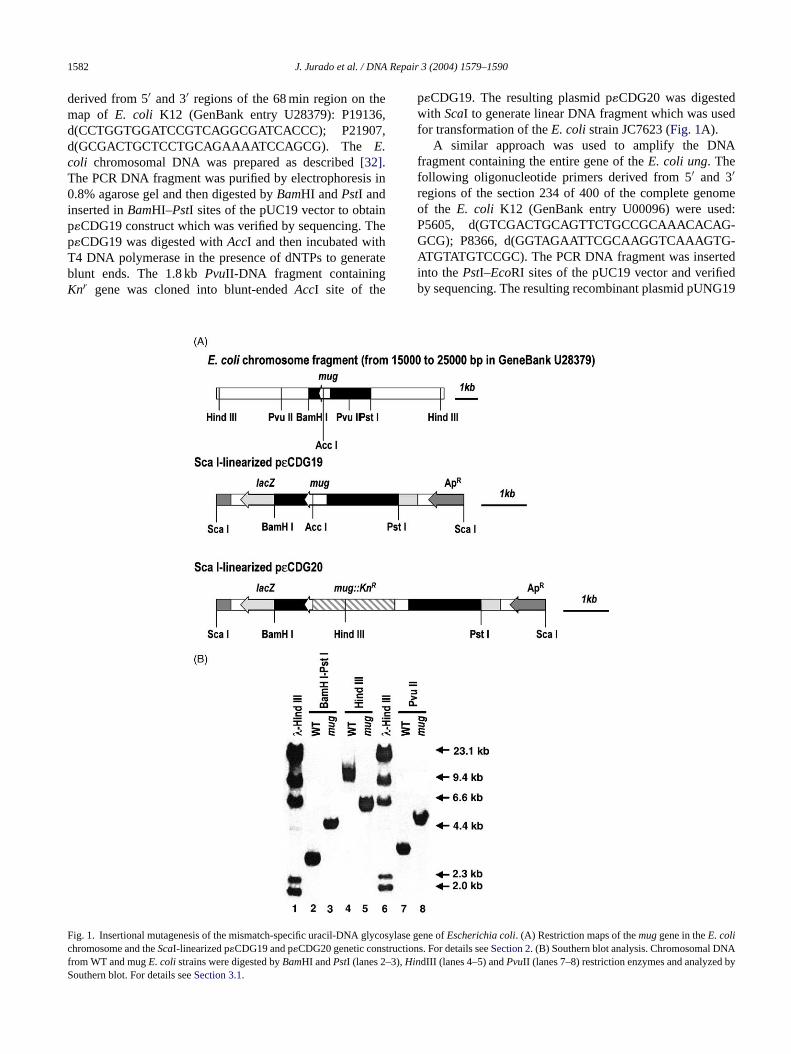
1582 J. Jurado et al. / DNA Repair 3 (2004) 1579–1590
derived from 5′ and 3′ regions of the 68 min region on themap of E. coli K12 (GenBank entry U28379): P19136,d(CCTGGTGGATCCGTCAGGCGATCACCC); P21907,d(GCGACTGCTCCTGCAGAAAATCCAGCG). The E.coli chromosomal DNA was prepared as described[32].The PCR DNA fragment was purified by electrophoresis in0.8% agarose gel and then digested byBamHI andPstI andinserted inBamHI–PstI sites of the pUC19 vector to obtainp�CDG19 construct which was verified by sequencing. Thep�CDG19 was digested withAccI and then incubated withT4 DNA polymerase in the presence of dNTPs to generateblunt ends. The 1.8 kbPvuII-DNA fragment containingKnr gene was cloned into blunt-endedAccI site of the
FcfS
p�CDG19. The resulting plasmid p�CDG20 was digestedwith ScaI to generate linear DNA fragment which was usedfor transformation of theE. coli strain JC7623 (Fig. 1A).
A similar approach was used to amplify the DNAfragment containing the entire gene of theE. coli ung. Thefollowing oligonucleotide primers derived from 5′ and 3′regions of the section 234 of 400 of the complete genomeof the E. coli K12 (GenBank entry U00096) were used:P5605, d(GTCGACTGCAGTTCTGCCGCAAACACAG-GCG); P8366, d(GGTAGAATTCGCAAGGTCAAAGTG-ATGTATGTCCGC). The PCR DNA fragment was insertedinto thePstI–EcoRI sites of the pUC19 vector and verifiedby sequencing. The resulting recombinant plasmid pUNG19
ig. 1. Insertional mutagenesis of the mismatch-specific uracil-DNA glycosyhromosome and theScaI-linearized p�CDG19 and p�CDG20 genetic constructirom WT and mugE. colistrains were digested byBamHI andPstI (lanes 2–3),Hinouthern blot. For details seeSection 3.1.
lase gene ofEscherichia coli. (A) Restriction maps of themuggene in theE. colions. For details seeSection 2. (B) Southern blot analysis. Chromosomal DNAdIII (lanes 4–5) andPvuII (lanes 7–8) restriction enzymes and analyzed by

J. Jurado et al. / DNA Repair 3 (2004) 1579–1590 1583
was digested withBamHI and incubated with T4 DNApolymerase in the presence of dNTPs to generate blunt ends.The 2.0 kbSmaI-DNA fragment containing theSpcr genewas cloned into blunt-endedBamHI site of the pUNG19.The resulting plasmid pUNG20 was digested withScaIto generate linear DNA fragment which was used fortransformation of theE. coli strain JC7623.
For cloning into pET19b plasmide (Novagen, Merck Eu-rolab S.A. France) the cDNAs encoding MUG was ampli-fied with the oligonucleotide primers containingNdeIand BamHI restriction sites: MUG5′-NdeI, d(GGAATT-CCATATGGTTGAGGATATTTTGGCTCC) and MUG3′-BamHI, d(GGGGATCCTTATCGCCCACGCACCACCA-GCGC). The product was then digested in the presence ofNdeI and BamHI and subcloned intoNdeI–BamHI sites ofpET19b yielding the plasmid pET19b-MUG.
2.4. Southern hybridization analyses
TheE. coli chromosomal DNA was isolated as describedabove. The DNA was digested by appropriate restrictionendonucleases prior to separation by electrophoresis inagarose gel. Transfer and hybridization were performed us-ing Gene-Screen hybridization transfer membrane accordingto the manufacturer’s instruction (New England Nuclear,C
2
i ics.T ndedi ,p ina ovedb
2
o-ta f a 34ma asee oft MH ol,1 oi rcf werea w/v)pb icsI ants
2.7. Sensitivity to DNA damage treatments
Alkylating agents treatments. Bacteria were grown in LBmedium with shaking (90 rpm) at 37◦C for 12 h. An aliquotof the bacterial suspension (∼5× 107 cells) and the mutagen(0–40 mM for MMS and 0–0.4 mM for MNNG) were pre-incubated with shaking (90 rpm) in 1 ml of LB medium at37◦C for 20 min. The mutagen was then washed by centrifu-gation and the bacteria were re-suspended in 1 ml of M9 salts.0.1 ml of different dilutions of the treated bacteria were com-bined with 2 ml of molten top agar and then poured onto LBplates. All data represent averages from at least two duplicateplates. Each assay was repeated at least twice using a widerange of mutagen concentrations. Plates were incubated at37◦C for 24 h and colonies were counted automatically us-ing an image analyzer (Analytical Measuring System Ltd.,UK, model 40-10).
Hydrogen peroxide and paraquat treatments: experimentswere performed under similar conditions described for mea-suring the sensitivity to alkylating agents.∼2× 105 bacterialcells were treated with 0–20 mM hydrogen peroxide and∼1× 104 cells were treated with 0–2 mM paraquat.
UV irradiation: bacterial cells were grown as described,harvested by centrifugation and re-suspended in M9 salts.Aliquots of 8 ml of the bacterial suspension (∼5 ×1 5 tedw of2
iledp rea mcr ol upt ltedo HCl( ns-f iesw
ngt lM ndt -t thent als at-m ionsp otso cu-lw resw nL .A t thet 7a mu-t e to
ourtaboeuf Cedex, France).
.5. Preparation of crude lysates
Individual clones were grown overnight at 37◦C for 16 hn 10 ml of LB broth containing the appropriate antibiothe culture was centrifuged, and the pellet was re-suspe
n 0.5 ml of lysis medium (300 mM Tris–HCl, 5 mM EDTAH 8.0) and stored at−20◦C. The frozen cells were thawedn ice-water bath, sonicated and the cell debris was remy centrifugation at 20,000× g, for 15 min, at 4◦C.
.6. DNA glycosylase assays
Purification of theE. coliMUG, Fpg and human TDG preins was performed as described[21]. The release of�-basend mismatched thymine were measured by cleavage oer duplex oligonucleotide containing a single�C residues described[16]. The standard assay mixture for bxcision activity (50�l final volume) contained 0.5 pmolhe 5′-[32P]-end labeled oligonucleotide duplex in 70 mepes-KOH pH 7.8, 1 mM EDTA, 5 mM 2-mercaptoethan00�g/ml BSA, 5 ng ofE. coli Fpg protein (in order t
ncise DNA at abasic sites) and either purified enzyme oE.oli cell-free extract. Incubations were carried out at 37◦Cor 30 min unless otherwise stated. Reaction productsnalyzed by electrophoresis through denaturing 20% (olyacrylamide gels (7 M urea, 0.5× TBE), visualizedy PhosphorImager Storm 840 (Molecular Dynam
nc., Sunnyvale, CA) and quantified using ImageQuoftware.
0 cells/ml) in 9 cm in diameter Petri dish were irradiaith a germicidal UV source having a maximal output54 nm.
Chloroacetaldehyde (CAA) treatment of the supercolasmid DNA: 0.1 mg/mL of pUC19 plasmid DNA welkylated in buffer containing 0–0.5 M CAA, 50 mM sodiuacodylate (pH 7.0) and 1 mM EDTA at 23◦C for 10 min. Theeaction was stopped by the addition of 2-mercaptoethano 0.5 M, and the reaction mixture was immediately desan a Sephadex G50 column equilibrated in 10 mM Tris–pH 7.5) and 1 mM EDTA. CAA-treated pUC19 was traormed toE. coli strains and ampicillin resistant colonere counted.CAA-treatment ofE. coli cells was performed accordi
o Mroczkowska et al.[33]. An overnight culture in minima9 medium was diluted 1:100 with fresh LB medium a
he bacteria were grown to a density of 1× 108 cells/ml, cenrifuged and re-suspended in C salts. The bacteria werereated with 10 mM CAA for 50–60 min to obtain the finurvival level of 5–20%. Immediately after the CAA treent aliquots were withdrawn and after appropriate dilutlated on LB medium to estimate the cells survival. Aliquf 0.25 ml of CAA treated and control bacteria were ino
ated into 2.5 ml of fresh LB and grown overnight at 37◦Cith shaking. Samples of 0.1–0.5 ml of overnight cultuere spread on M9-Lac+ plates for Lac+ revertants and oB-rifampicin plates for rifampicin resistant (Rifr) mutantsppropriate dilutions were spread on LB plates to coun
otal number of viable cells. Plates were incubated at 3◦Cnd counted after 24 h for cell survival and after 48 h for
agenesis. Mutation frequency was calculated from thre

1584 J. Jurado et al. / DNA Repair 3 (2004) 1579–1590
four independent experiments as the number of mutants per108 viable cells.
2.8. Measurement of spontaneous rate of mutation
Five parallel cultures of 4 ml were inoculated from an in-dependent colony and grown at 37◦C for 16 h. The 3 ml ofthe overnight cultures were spun down (10,000× g for 15 s),washed in M9 medium and then re-suspended in 300�l of thesame medium. The appropriate dilutions were plated on LB,LB containing 100�g/ml rifampicin and M9 medium con-taining lactose to determine the survival, rifampicin-resistantmutants and Lac+ revertants, respectively. Mutation rateswere determined by the method of the median[34]. Unlessotherwise noted, rates given in the tables are the mean± S.D.of at least three independent experiments for each strain.
2.9. Site-specific incorporation of�C into M13G*1
Site-directed mutagenesis was performed as described[30]. Synthetic 41 mer oligonucleotide M13-�C was hy-bridized to single-stranded circular M13G*1 at a 25:1primer/template ratio in 20 mM Tris–HCl, 10 mM MgCl2,50 mM NaCl, pH 7.5 buffer by incubation for 5 min at 75◦C.The reaction (volume 30�l) was allowed to cool at a rateo ◦ ◦ ni ut bya oly-mt TP,1 c-taw np ium(c per-n edf es wthot n at3 ored.M bluep sis rea werec
3
3m
ifico .
As shown in Fig. 1A, the 2750 bp PCR-DNA fragmentcontaining theMUG gene was cloned into pUC19 plasmidvector resulting in p�CDG19 plasmid. The kanamycinresistance marker (1.8 kb fragment) was inserted into theuniqueAccI site located within theMUG gene in p�CDG19yielding the p�CDG20 plasmid (Fig. 1A). To transfer themutation into theE. coli chromosome theScaI-linearizedplasmid was transformed to kanamycin sensitiveE. colirecB recC sbcB sbcC, cells that acquired selectable markerby a double recombination event between the homologousregions flanking the marker were selected[35,36]. Threekanamycin resistant transformants were analyzed and foundto be sensitive to ampicillin and defective in�C-DNA gly-cosylase activity in cell-free extracts. Insertional mutationwas further confirmed by Southern analyses. As shown inFig. 1B BamHI–PstI digestion ofE. colichromosomal DNAfrom wild-type (WT) andmug::Knr clone (MS20) generates2.7 and 4.5 kb fragments, respectively, as predicted fromthe insertional mutagenesis (lanes 2 and 3). As expected,HindIII-digestion generates 5.6 kb fragment in MS20 and10 kb fragment in WT (Fig. 1B, lanes 4 and 5) andPvuIIdigestion generates 3 kb fragment in WT and 4.8 kb fragmentin MS20 (Fig. 1B, lanes 7 and 8). Cell-free extract frommug::Knr is deficient in excision of�C under conditiondescribed inSection 2(Fig. 2, lane 2). A P1 phage stockw andA
3g
com-bw atedw cityom l-f -c ee 9b,ng dw ro-t ly-c dw rmt de
3a
am-a -o , thes
f approximately 1C per min to 30 C and then placed oce. The synthesis of the second strand was carried odding to the annealing reaction 6 units of the T4 DNA perase, 3 units of the T4 DNA ligase and 3�l of 10× syn-
hesis buffer containing 5 mM of each dNTPs, 10 mM A00 mM Tris–HCl pH 7.5, 20 mM dithiotreithol. The rea
ion was incubated on ice for 5 min, then at 25◦C for 5 minnd finally at 37◦C for 90 min. CompetentE. coli strainsere transformed directly with 1�l of the synthesis reactioroduct via electroporation, then inoculated into LB med10 ml) and incubated overnight with shaking at 37◦C. Theultures were centrifuged twice and the remaining suatant (approximately 1011 phage particles per ml) was us
or transfection ofE. coli strain JM105. Dilutions of phagtock solution were mixed with fresh late-logarithmic grof JM105 and molten top agar containing X-gal (40�g/ml)
hen poured on LB agar plates. After overnight incubatio7◦C dark blue plaques representing revertants were scutation frequency was calculated as the ratio of darklaques to colorless plaques. Three independent synthections and five to seven independent transformationsarried out to determine mutation frequencies.
. Results
.1. Construction of a�C-DNA glycosylase defectiveutant of E. coli
The entireMUGgene was cloned using PCR with specligonucleotide primers andE. coli chromosomal DNA
-
as grown on MS20, and used to transduce JM105M101–AM106 strains to kanamycin resistance andmug.
.2. The hTDG protein can complement�C-DNAlycosylase in cell-free extracts of E. coli mug mutant
Previously, we have demonstrated that the purified reinant hTDG protein is able to excise�C [16]. However, itas possible that the hTDG preparation was contaminith MUG. To further substantiate the substrate specifif the human protein we expressed hTDG inE. coli mugutant and measured�C-DNA glycosylase activity in cel
ree extracts. As shown inFig. 2 when the duplex oligonuleotide containing an�C·G pair was incubated with cell-frextract frommugmutant containing parental vector, pET1o excision of�C was detected (lane 2). However,�C-DNAlycosylase activity was restored inmugmutant transformeith plasmid expressing the MUG (lane 3) and hTDG p
eins (lane 4). A weak mismatch-specific thymine-DNA gosylase activity was seen inE. coli mugmutant transformeith pT7-hTDG vector (Fig. 2, lane 9). These results confi
hat hTDG has an intrinsic�C-DNA glycosylase activity anliminate a possible MUG contamination.
.3. Sensitivity to DNA damaging agents, spontaneousnd CAA-induced mutagenesis of E. coli mug strains
To assess the role of the MUG protein towards DNA dge in vivo, the sensitivity of theE. coli mugmutant to varius DNA damaging agents was investigated. In additionpontaneous and induced mutation rates of themugmutant

J. Jurado et al. / DNA Repair 3 (2004) 1579–1590 1585
Fig. 2. Cleavage of the oligonucleotides containing�C·G and T·G base pairs by theE. coli cell-free extracts. 5′-[32P]-labeled�C·G and T·G oligonucleotideduplexes 10 nM were incubated with 20�g of cell-free extracts fromE. coliMS50 (DE3) at 30◦C for 30 min. Lanes: (1) control�C·G; (2) as (1), but incubatedwith extract from MS50 (DE3) containing pET19b; (3) MS50 (DE3) containing pET19b-MUG; (4) MS50 (DE3) containing pT7-hTDG; (5) 10 ng of thepurified MUG protein; (6) control T·G; (7) as (6), but incubated with extract from MS50 (DE3) containing pET19b; (8) MS50 (DE3) containing pET19b-MUG;(9) MS50 (DE3) containing pT7-hTDG; (10) 50 ng of the purified hTDG protein. The products of the reaction were analyzed as described inSection 2.
were compared to that of the WT strain. As controls we usedE. coli alkA tagA, uvrB andsodA sodBstrains known to besensitive to DNA damaging agents. As shown inFig. 3, E.coli alkA tagA, uvrBandsodA sodBstrains were sensitive toMNNG, UV and paraquat, respectively, whereas the sensi-tivity of mugstrain to alkylating agents MNNG and MMS(Fig. 3A and data not shown), to UV (Fig. 3B) and to theoxidating agents paraquat and H2O2 (Fig. 3C and data notshown) are similar to that of the WT. Furthermore, the re-sults show thatmugmutant is also not sensitive to CAA, anagent which is known to generate�-bases in DNA in vitro(data not shown).
Sincemug mutant was no more sensitive to genotoxicagents than the WT, and since MUG was presumed to beinvolved in the repair of ethenobases, we attempted to mea-sure surviving fraction of CAA-treated plasmid DNA afterbacteria transformation. As shown inFig. 3D, pUC19 DNAdamaged in vitro by CAA-treatment was inactivated at a two-to three-fold higher rate inmugmutant than in WT host.
To determine the spontaneous and CAA-induced mutationrates we used the set of strains, constructed as described inSection 2, carrying themugmutation that allows detection ofeach transition and transversion by monitoring the Lac− toLac+ reversion frequency[27]. Strains AM201–AM206 car-rying themugmutation were plated on selective medium tomn sw T( ainedw iciner t
was weakly mutagenic, although, significant increase in G·C→ A·T and G·C → T·A mutations was detected in WT. Asexpected,mugmutant exhibited higher frequency and widerspectra of mutations as compared to WT strains. Inmugmu-tant additionally an important increase of G·C → C·G andA·T → T·A transversions was found. ComparingmugandWT strains, it appeared that CAA inmugstrain induced sig-nificantly higher frequency of G·C → A·T, G·C → C·G andA·T → T·A base substitutions. Unexpectedly, only slightdifference was observed in the frequency of G·C → T·Atransversions.
3.4. Repair of the site-specifically incorporated�Cresidue in M13G*1 phage
The mug mutant is not sensitive to DNA damagingagents and CAA-treatment is weakly mutagenic (Fig. 3andTable 2). These may be due to inefficient formation of�C residues by CAA in theE. coli chromosomal DNA. Toassess the role of MUG, we measured mutation frequency ofthe site-specifically incorporated�C residue using a geneticreversion system. A single�C residue was incorporated intobacteriophage vector, M13G*1, opposite to C at position141 of thelacZα gene. All single base substitutions at thisposition can be detected by change in plaque color. As showniw con-s ver,i ldg→ tion
easure the spontaneous mutation rate. As shown inTable 2o increase in spontaneous Lac− to Lac+ reversion rateere detected inmug(AM201-AM206) as compared to W
CC101–CC106) strains. The same results were obthen measuring spontaneous mutation rate to rifamp
esistance (Table 2). As shown inTable 2, CAA-treatmen
n Fig. 4, excision of �C from �C·C base pair by MUGould lead to restoration of the missence mutation andequently to colourless “white” plaque phenotype. Howef not repaired, replication of�C-containing template wouenerate base substitutions�C → C, A, T, which, except�C
G, would lead to dark blue revertants due to restora

1586 J. Jurado et al. / DNA Repair 3 (2004) 1579–1590
Fig. 3. Sensitivity of theE. coli WT and mutant strains towards DNA damaging agents (A–C) and inactivation of the transforming DNA treated by CAA inWT andmughosts (D). (A–C) Bacterial strains were exposed to several doses of the chemical MNNG (A); paraquat (B); or physical UV (C) DNA damagingagents. After controlled exposure, bacteria were plated on LB plates. Surviving fraction was calculated using unexposed bacterial cultures as reference. (D)The pUC19 DNA was treated with several doses of CAA and transformed into competent cells. Survival fraction was determined on LB plates containingampicillin. Non-treated pUC19 DNA plasmid was used as a control. For details seeSection 2.
of �-galactosidase activity (Fig. 4). The M13G*1 double-stranded DNA containing�C·C base pair was transformedintoE. colistrains JM105 (WT), MS50 (mug), JM109 (recA),BH430 (uvrA), MS51 (mutL) and MS52 (mutS) and the�C-induced mutation frequency was measured. As shown
Fig. 4. Schematic representation of the proposed model for�C-directed mu-tagenesis.
in Table 3, �C-induced mutation frequency in MS50 (30.3± 12.0× 10−4) was about seven-fold higher as comparedto isogenic JM105 strain (4.6± 1.6× 10−4). In theE. colistrains deficient in nucleotide excision repair (BH430) andrecombination (JM109), the�C-induced reversion frequencywas similar to that of WT (Table 3). Interestingly,E. colimismatch repair mutants MS51 and MS52 showed slightlyhigher mutation frequency as compared to WT (Table 3).The phage DNA from 12 individual dark blue plaques wereisolated and subjected for sequencing. All of six revertantsfrom JM105, exhibited�C → C base substitution at thetarget site, whereas among six revertants from MS50, 4exhibited�C → C and two�C → T base substitution at thetarget site.
3.5. Role of the MUG protein in the prevention of themutagenic effect of cytosine deamination
Deamination of cytosine, if not repaired, gives rise to G·C→ A·T transition mutations. The majorE. coli uracil-DNAglycosylase (UNG) acts on single and double-stranded

J. Jurado et al. / DNA Repair 3 (2004) 1579–1590 1587
Table 3Mutation frequencies from in vivo replication of site-specifically incorporated�dC residue into M13G*1 phage DNA
Strain No. of plaques screened No. of blue plaques Mutation frequency Average± S.D. Ratio mutant/WT
JM1051Ex. 1 40470 20 4.9 × 10−4 4.6± 1.6× 10−4
Ex. 2 31080 15 4.8 × 10−4
Ex. 3 31600 6 1.9 × 10−4
Ex. 4 9500 5 5.3 × 10−4
Ex. 5 12820 8 6.2 × 10−4
MS50Ex. 1 21690 37 17.0 × 10−4 30.3± 12.0× 10−4 6.6**Ex. 2 22930 71 31.0 × 10−4
Ex. 3 1830 3 16.4 × 10−4
Ex. 4 22980 106 46.1 × 10−4
Ex. 5 960 3 31.2 × 10−4
Ex. 6 2980 12 40.3 × 10−4
BH430Ex. 1 120000 55 4.5 × 10−4 4.1± 0.6× 10−4 0.9 nsEx. 2 54500 24 4.4 × 10−4
Ex. 3 14800 5 3.4 × 10−4
JM109Ex. 1 52500 15 2.9 × 10−4 3.0± 1.7× 10−4 0.8 nsEx. 2 2073 1 4.8 × 10−4
Ex. 3 176000 23 1.3 × 10−4
MS51 50000 36 7.2 × 10−4 7.2× 10−4 1.5MS52 39600 23 6.2 × 10−4 6.2× 10−4 1.3
Mutation frequency was calculated as the ratio of dark blue plaques to colourless plaques. Five independent transformations were carried for JM105,six forMS50, three for BH430 and JM105 and only one for MS51 and MS52. Statistical significance in MS50, BH430 and JM109 as compared to JM105 was evaluatedusing two-tailed Student’s test (∗∗P < 0.01;nsP > 0.05).
Table 4Effect ofmugmutation on the rate of G·C → A·T transition
Strain Spontaneous Rifr
mutation per cellgeneration (×10−9)
Ratiomutant/WT
Ratioung,mug/ung
Spontaneous Lac+reversion per cellgeneration (×10−9)
Ratio mutant/WT Ratioung,mug/ung
CC102 WT 6.8± 1.1 1 0.66± 0.11 1MS1020mug:: Knr 3.8± 0.03 0.6* 0.50± 0.30 0.8 nsMS1021ung:: Spcr 13.3± 1.8 1.9** 1 4.91± 1.33 7.4** 1MS1022 ung:: Spcr ,
mug:: Knr13.6± 3.4 2.0* 1 ns 7.95± 0.94 12.0*** 1.6*
Rates in the tables are the mean± S.D. of three independent experiments for each strain. Statistical significance of the difference in mutation rates was evaluatedusing two-tailed Student’s test (∗∗∗P < 0.001;∗∗P < 0.01;∗P < 0.05 andnsP > 0.05).
uracil-containing DNA[37] consequently, the rate of sponta-neous C→ T transitions inE. coli ungmutant is elevated by30-fold[38]. It was speculated that MUG could be a back-uprepair enzyme for UNG inE. coli [19]. Themugmutant doesnot have mutator phenotype suggesting that inE. coli UNGremoves most of uracil residues generated through cytosinedeamination. However, the role of MUG in repair of uracilresidues in vivo may be masked by UNG, therefore, we mea-sured spontaneous mutation rates ofE. coli CC102 strainscarrying single and/or double mutations:mug, ungandmugung. For this purpose the cultures were grown at 30◦C tostationary phase and plated on selective media. As shown inTable 3, the frequencies of Rifr and Lac+ mutants formugwere slightly lower as compared to WT, although, the differ-ence for the latter was not statistically significant. In contrast,
the Rifs → Rifr mutation and Lac+ reversion rates inungwere two- and seven-fold higher, respectively, than in WT(Table 3). In the double mutantmug ung, the rate of Rifr mu-tants was similar, however, the frequency of Lac+ revertantswas 1.6-fold higher than in the singleungmutant (Table 4).The difference in Lac+ reversion frequencies betweenungandung mugmutants was small but statistically significant.
4. Discussion
To elucidate the physiological role of�C-DNA glycosy-lasesE. coli mug::Knr strain deficient in�C-excision activ-ity was generated. Our data confirmed previous ones showingthatE. colimugmutant lacks detectable�C-DNA glycosylase

1588 J. Jurado et al. / DNA Repair 3 (2004) 1579–1590
activity in cell-free extracts[23]. Previously, we have shownthat human homolog of MUG, hTDG is able to excise�C invitro [16,39]. Here, we show that expression of hTDG in theE. coli mugmutant complements the�C-activity in cell-freeextract indicating that the�C-DNA glycosylase activity is anintrinsic property of the human enzyme.
TheMUGgene product excises a broad spectrum of dam-aged purines and pyrimidines in duplex DNA suggesting thatE. coli mugmutant may have specific phenotype. The resultspresented above show thatmugmutant is not particularly sen-sitive to any of DNA damaging agents tested suggesting thatin vivo MUG is not involved in the repair of alkylated purines,UV-photoproducts and oxidized bases. The�C residue actsas a non-instructional lesion and although, less strongly thanabasic sites, inhibits translesion DNA synthesis[40]. Further-more, an�C residue incorporated in single-stranded genomereduced its survival by 70–90% indicating that this adductwhen present in DNA might be lethal to the bacterial cells[13,14,41]. Since so far�C is the main DNA substrate forMUG, we investigated whethermugmutant is sensitive toagents generating�-bases. Surprisingly,mugmutant was notsensitive to CAA treatment suggesting that theE. coli chro-mosomal DNA is not easily accessible to the reactive alde-hydes. Since, CAA generates�-adducts with good efficiencywith purified DNA, we examined the genetic requirement fort effi-cs asi s,iv
ownt ta-n ingti im-p ionsi ousm int doge-n thylc vinylc h nu-c ar-b iateda ic inS airs e-t des inv r-i 50( u-b ar-b waso nw
By using extra chromosomal vectors with a site-specifically incorporated�-base, it was shown that inE. coliN2,3-�G adduct induce G→ A transition[44], �C induce C→ T and C→ A mutations[13,14], �A induce A→ T and A→ C transversions[45] and deletions and 1,N2-�G leads to G→ A transitions and other types of mutation[46]. However,the genetic requirements for the�-base induced mutagenesiswere not investigated. Therefore, we used CAA-treatment ofthe E. coli cells to determine the role of MUG in inducedmutagenesis. The G·C → A·T transition was a major muta-tion in WT strain suggesting thatN2,3-�G and/or�C is themajor mutagenic adduct generated by CAA. As compared toknown mutagens such as alkylating agents and base analogstested on strains CC101–CC106[27], the CAA was a weakmutagen in both WT andmugstrains. However, themugmu-tant showed statistically significant increase in CAA-inducedmutations G·C→ C·G, G·C→ T·A and A·T → T·A as com-pared to WT strains. It has been shown that in vitro MUG canexcise 1,N2-�G and�A residues[20,21]. We propose that, inaddition to�C, MUG may participate in the removal of�Aand 1,N2-�G adducts in vivo.
Similar frequencies of CAA-induced C→ T and C→ Amutations in both WT andmugstrains and high standard de-viation values suggest inefficient formation of�C residues inDNA. High toxicity and reactivity of CAA may be one of ther to usesa entlyt ucedi int ions itht no sD s oftb n-foldi y in-c gt oft uta-t hem ovalo tly,s sc rn . Al-t sa
r, iteT cy-tamt t
he excision of�-bases by measuring the transformationiency of CAA-treated plasmid DNA inE. coli mugand WTtrains. We found that in vitro modified pUC19 vector wnactivated at a greater rate inmugmutant than in WT hostndicating that MUG is involved in the repair of�-bases inivo.
By using a plasmid-based reversion assay it was shhat inactivation ofmughas no effect on the rate of sponeous mutations[23]. We confirmed this observation show
hat no increase in Rifs → Rifr and Lac− → Lac+ mutationsn mugmutant were detected. These observations mayly that either�-bases are weakly mutagenic, or rare les
n E. coli, or both. We propose that the lack of spontaneutator phenotype ofmugmutant under conditions used
he present study might be due to the absence or low enous level of�C residues. In mammals the carcinogen, earbamate, is activated by P450-mediated oxidation toarbamate and then to an epoxide which can react witleic acids[3,4]. Although, in bacterial test system ethyl camate is not genotoxic, in the presence of P450-medctivation, ethyl carbamate was found to be mutagenalmonella typhimuriumstrain TA100, generating base-pubstitutions at G·C base pairs[42]. In present study, to mabolize ethyl carbamate and generate reactive aldehyivo, we constructedE. coli WT andmugstrains harboung plasmid pCW′/2E1/hNPR expressing cytochrome p4CYP2E1)[43]. Lac+ revertants were monitored after incation of theE. coli strains in media containing ethyl camate. No induction of revertants in the both strainsbserved suggesting that inE. coliP450-catalysed oxidatioas inefficient (data not shown).
easons of the low mutagenesis. Therefore, we decidedite-directed mutagenesis technique[44,47–49]to introducemodified base at defined sites in DNA and subsequ
o determine the frequency and types of mutations prodn vivo. In present study, to substantiate the role of MUGhe repair of�C residues in vivo we used a genetic reversystem[44] based on a bacteriophage vector, M13G*1, whe site-specifically incorporated�C residue. Transformatiof the phage DNA intoE. coli strains deficient in variouNA repair pathways allows comparing the frequencie
he�C-induced mutations at position 141 of thelacZα geney change in plaque color. Here, we demonstrate a seve
ncrease in the mutation frequency for the site-specificallorporated�C residue inmugas compared to WT indicatinhat MUG is responsible for the repair of more than 80%he�C residues present in DNA. A slight increase in mion frequencies inmutSandmutLstrains suggests that tismatch repair system may also participate in the remf �-adducts in newly synthesized DNA strand. Importanimilar mutation frequencies inuvrA and recA mutants aompared to WT strain indicate that�C is not a substrate foucleotide excision and recombination repair pathways
ogether, the results demonstrate that in vivo the�C residuere mainly repaired by MUG.
MUG can repair mismatched uracil residue, howevexcises�C more efficiently than other damaged bases[16,20].o elucidate the role of MUG in repair of the deaminatedosines we have introduced single mutationsmugandungnd double mutationmug, ung into E. coli CC102 strain toeasure G·C→A·T transitions by reversion to Lac+ pheno-
ype. The frequency of Lac+ reversion inmugwas somewha

J. Jurado et al. / DNA Repair 3 (2004) 1579–1590 1589
lower than in WT, at variance, inung mutant the rate was7.4-fold higher as compared to WT. However, in the doublemug ungmutant grown at 30◦C the rate of C→ T transi-tion was increased further to 12-fold as compared to WT and1.6-fold as compared to singleungmutant. At variance, nodifference in the Rifr mutations frequency was observed be-tweenung andmug ungstrains. It should be stressed that,in contrast to Lac+ reversion, Rifr mutations reflect the sumof the base substitutions occurring at different sites of therpoBgene, which encodes the�-subunit of RNA polymerase[50]. We found that the difference in spontaneous mutationrates betweenungandmug ungmutants was not statisticallysignificant when the cultures were grown at 37◦C (data notshown). Similar results were obtained when using the re-version to Knr arising exclusively from C→ T transitions,MUG reduced slightly these mutations but the effect was notstatistically significant[23]. The results obtained in presentstudy suggest that in vivo approximately 90% of G·U pairsare repaired by UNG and only 4% by MUG. The underlyingmechanism for the temperature-dependent increase in spon-taneous mutation rate in the double mutant remains unclearand needs further investigation. We propose that under certaincultivation conditions MUG could act as a back-up enzymefor UNG.
In the present study we have demonstrated that themugm , in-c aves inD af-f ck-upr
A
eidf idv hu-m ffreyH forh wass 00-0 surl cel-l .);J 4/T)a
R
ver
cidethylancer
[3] F.P. Guengerich, D.H. Kim, Enzymatic oxidation of ethyl carbamateto vinyl carbamate and its role as an intermediate in the formationof 1,N6-ethenoadenosine, Chem. Res. Toxicol. 4 (1991) 413–421.
[4] G.A. Dahl, J.A. Miller, E.C. Miller, Vinyl carbamate as a promutagenand a more carcinogenic analog of ethyl carbamate, Cancer Res. 38(1978) 3793–3804.
[5] M.T. Leithauser, A. Liem, B.C. Stewart, E.C. Miller, J.A. Miller,1,N6-Ethenoadenosine formation, mutagenicity and murine tumor in-duction as indicators of the generation of an electrophilic epoxidemetabolite of the closely related carcinogens ethyl carbamate (ure-thane) and vinyl carbamate, Carcinogenesis 11 (1990) 463–473.
[6] K.K. Park, Y.J. Surh, B.C. Stewart, J.A. Miller, Synthesis and prop-erties of vinyl carbamate epoxide, a possible ultimate electrophilicand carcinogenic metabolite of vinyl carbamate and ethyl carbamate,Biochem. Biophys. Res. Commun. 169 (1990) 1094–1098.
[7] H.M. Bolt, Roles of etheno-DNA adducts in tumorigenicity ofolefins, Crit. Rev. Toxicol. 18 (1988) 299–309.
[8] F.P. Guengerich, K.S. Min, M. Persmark, M.S. Kim, W.G.Humphreys, J.M. Cmarik, R. Thier, Dihaloalkanes and poly-haloalkenes, IARC Sci. Publ. (1994) 57–72.
[9] F.L. Chung, H.J. Chen, R.G. Nath, Lipid peroxidation as a potentialendogenous source for the formation of exocyclic DNA adducts,Carcinogenesis 17 (1996) 2105–2111.
[10] F. el Ghissassi, A. Barbin, J. Nair, H. Bartsch, Formation of 1,N6-ethenoadenine and 3,N4-ethenocytosine by lipid peroxidation prod-ucts and nucleic acid bases, Chem. Res. Toxicol. 8 (1995) 278–283.
[11] L.J. Marnett, P.C. Burcham, Endogenous DNA adducts: potentialand paradox, Chem. Res. Toxicol. 6 (1993) 771–785.
[12] J. Nair, Lipid peroxidation-induced etheno-DNA adducts in humans,IARC Sci. Publ. (1999) 55–61.
[ Es-ride-,93)
[ ncy
94)
[ n of
r874.
[ nicdNA3.
[ en-In-ecificat-
[ o-2 ises,
1 as936.
[ ses996)
[ ag-
Biol.
[ h, H.A
c
utant is not sensitive towards DNA damaging agentsluding CAA. Using the genetic reversion system we hhown that inE. coli the majority of�C residues presentNA are processed by MUG. Although MUG does not
ect the spontaneous mutation rate, it may serve as a baepair enzyme for UNG.
cknowledgements
We wish to thank Drs. Lawrence Loeb and Thomas Ror the bacteriophage vector, Dr. Dick d’Ari for the plasmector, Dr. Peter Guengerich for the plasmid expressingan cytochrome P450, Dr. Roland M. Schaaper and Je. Miller for the bacterial strains and Dr. Barbara Tudekelpful discussion and financial support. This researchupported by the European Community Grant QLK4-200286 (M.K.S. and B.T.), Association pour la Recherche
e Cancer (M.K.S.) and by the program of the Center of Exence in Molecular Biotechnology IBB PAS (WP10) (B.T.J. was supported by fellowships from ESF (RF/96/3nd Ministry of Education and Science of Spain.
eferences
[1] T. Green, D.E. Hathway, Interactions of vinyl chloride with rat-liDNA in vivo, Chem. Biol. Interact. 22 (1978) 211–224.
[2] J.A. Miller, E.C. Miller, The metabolic activation and nucleic aadducts of naturally-occurring carcinogens: recent results withcarbamate and the spice flavors safrole and estragole, Br. J. C48 (1983) 1–15.
13] A.K. Basu, M.L. Wood, L.J. Niedernhofer, L.A. Ramos, J.M.sigmann, Mutagenic and genotoxic effects of three vinyl chloinduced DNA lesions: 1,N6-ethenoadenine, 3,N4-ethenocytosineand 4-amino-5-(imidazol-2-yl)imidazole, Biochemistry 32 (1912793–12801.
14] M. Moriya, W. Zhang, F. Johnson, A.P. Grollman, Mutagenic poteof exocyclic DNA adducts: marked differences betweenEscherichiacoli and simian kidney cells, Proc. Natl. Acad. Sci. U.S.A. 91 (1911899–11903.
15] B. Hang, B. Singer, G.P. Margison, R.H. Elder, Targeted deletioalkylpurine-DNA-N-glycosylase in mice eliminates repair of 1,N6-ethenoadenine and hypoxanthine but not of 3,N4-ethenocytosine o8-oxoguanine, Proc. Natl. Acad. Sci. U.S.A. 94 (1997) 12869–12
16] M. Saparbaev, J. Laval, 3,N4-Ethenocytosine, a highly mutageadduct, is a primary substrate forEscherichia colidouble-strandeuracil-DNA glycosylase and human mismatch-specific thymine-Dglycosylase, Proc. Natl. Acad. Sci. U.S.A. 95 (1998) 8508–851
17] F. Petronzelli, A. Riccio, G.D. Markham, S.H. Seeholzer, M. Guardi, M. Karbowski, A.T. Yeung, Y. Matsumoto, A. Bellacosa,vestigation of the substrate spectrum of the human mismatch-spDNA N-glycosylase MED1 (MBD4): fundamental role of the calytic domain, J. Cell Physiol. 185 (2000) 473–480.
18] B. Kavli, O. Sundheim, M. Akbari, M. Otterlei, H. Nilsen, F. Skrpen, P.A. Aas, L. Hagen, H.E. Krokan, G. Slupphaug, hUNGthe major repair enzyme for removal of uracil from U:A matchU:G mismatches, and U in single-stranded DNA, with hSMUGa broad specificity backup, J. Biol. Chem. 277 (2002) 39926–39
19] P. Gallinari, J. Jiricny, A new class of uracil-DNA glycosylarelated to human thymine-DNA glycosylase, Nature 383 (1735–738.
20] R.J. O’Neill, O.V. Vorob’eva, H. Shahbakhti, E. Zmuda, A.S. Bhwat, G.S. Baldwin, Mismatch uracil glycosylase fromEscherichiacoli: a general mismatch or a specific DNA glycosylase? J.Chem. 278 (2003) 20526–20532.
21] M. Saparbaev, S. Langouet, C.V. Privezentzev, F.P. GuengericCai, R.H. Elder, J. Laval, 1,N(2)-Ethenoguanine, a mutagenic DNadduct, is a primary substrate ofEscherichia colimismatch-specifi

1590 J. Jurado et al. / DNA Repair 3 (2004) 1579–1590
uracil-DNA glycosylase and human alkylpurine-DNA-N-glycosylase,J. Biol. Chem. 277 (2002) 26987–26993.
[22] B. Hang, G. Downing, A.B. Guliaev, B. Singer, Novel activity ofEscherichia colimismatch uracil-DNA glycosylase (Mug) excising8-(hydroxymethyl)-3,N4-ethenocytosine, a potential product result-ing from glycidaldehyde reaction, Biochemistry 41 (2002) 2158–2165.
[23] E. Lutsenko, A.S. Bhagwat, The role of theEscherichia colimugprotein in the removal of uracil and 3,N(4)-ethenocytosine fromDNA, J. Biol. Chem. 274 (1999) 31034–31038.
[24] S.K. Mokkapati, A.R. Fernandez de Henestrosa, A.S. Bhagwat,Es-cherichia coli DNA glycosylase Mug: a growth-regulated enzymerequired for mutation avoidance in stationary-phase cells, Mol. Mi-crobiol. 41 (2001) 1101–1111.
[25] J.H. Miller, A Short Course in Bacterial Genetics. A LaboratoryManual and Handbook forEscherichia coliand Related Bacteria,Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY,1972.
[26] W.J. Dower, J.F. Miller, C.W. Ragsdale, High efficiency transforma-tion of E. coli by high voltage electroporation, Nucleic Acids Res.16 (1988) 6127–6145.
[27] C.G. Cupples, J.H. Miller, A set of lacZ mutations inEscherichiacoli that allow rapid detection of each of the six base substitutions,Proc. Natl. Acad. Sci. U.S.A. 86 (1989) 5345–5349.
[28] N.G. Stoker, N.F. Fairweather, B.G. Spratt, Versatile low-copy-number plasmid vectors for cloning inEscherichia coli, Gene 18(1982) 335–341.
[29] P. Prentki, H.M. Krisch, In vitro insertional mutagenesis with a se-lectable DNA fragment, Gene 29 (1984) 303–313.
[30] P.D. Yuckenberg, F. Witney, J. Geisselsoder, J. McClary, Mutagen-ord,
[ J.J.manem.
[ omal
[ k,
tive.
[ ants
[ recBcad.
[ cted
[ ns,
[38] B.K. Duncan, B. Weiss, Specific mutator effects of ung (uracil-DNAglycosylase) mutations inEscherichia coli, J. Bacteriol. 151 (1982)750–755.
[39] B. Hang, M. Medina, H. Fraenkel-Conrat, B. Singer, A 55-kDa pro-tein isolated from human cells shows DNA glycosylase activity to-ward 3,N4-ethenocytosine and the G/T mismatch, Proc. Natl. Acad.Sci. U.S.A. 95 (1998) 13561–13566.
[40] G. Wang, P.M. Dunman, M.Z. Humayun, Replication of M13 single-stranded DNA bearing a site-specific ethenocytosine lesion byEs-cherichia coilcell extracts, Cell Res. 7 (1997) 1–12.
[41] V.A. Palejwala, D. Simha, M.Z. Humayun, Mechanisms of mutage-nesis by exocyclic DNA adducts. Transfection of M13 viral DNAbearing a site-specific adduct shows that ethenocytosine is a highlyefficient RecA-independent mutagenic noninstructional lesion, Bio-chemistry 30 (1991) 8736–8743.
[42] P. Hubner, P.M. Groux, B. Weibel, C. Sengstag, J. Horlbeck, P.M.Leong-Morgenthaler, J. Luthy, Genotoxicity of ethyl carbamate (ure-thane) inSalmonella, yeast and human lymphoblastoid cells, Mutat.Res. 390 (1997) 11–19.
[43] Y. Oda, P. Aryal, T. Terashita, E.M. Gillam, F.P. Guengerich, T.Shimada, Metabolic activation of heterocyclic amines and otherprocarcinogens inSalmonella typhimurium umutester strains ex-pressing human cytochrome P4501A1, 1A2, 1B1, 2C9, 2D6, 2E1,and 3A4 and human NADPH-P450 reductase and bacterialO-acetyltransferase, Mutat. Res. 492 (2001) 81–90.
[44] K.C. Cheng, B.D. Preston, D.S. Cahill, M.K. Dosanjh, B. Singer,L.A. Loeb, The vinyl chloride DNA derivativeN2,3-ethenoguanineproduces G to A transitions inEscherichia coli, Proc. Natl. Acad.Sci. U.S.A. 88 (1991) 9974–9978.
[45] G.A. Pandya, M. Moriya, 1,N6-Ethenodeoxyadenosine, a DNA35
[ TPsy-9-imun-
beta,
[ enic,8-ar-
[ lo-oso-
[ is inRes.
[ theol.
esis using uracil-containing DNA, Oxford University Press, Oxf1991.
31] P. Neddermann, P. Gallinari, T. Lettieri, D. Schmid, O. Truong,Hsuan, K. Wiebauer, J. Jiricny, Cloning and expression of huG/T mismatch-specific thymine-DNA glycosylase, J. Biol. Ch271 (1996) 12767–12774.
32] J.W. Dale, P.J. Greenaway, T. Walker, Preparation of chromosDNA from E. coli, Humana Press, Clifton, NJ, 1984.
33] M.M. Mroczkowska, I.K. Kolasa, J.T. KusmiereChloroacetaldehyde-induced mutagenesis inEscherichia coli:specificity of mutations and modulation by induction of the adapresponse to alkylating agents, Mutagenesis 8 (1993) 341–348
34] D.E. Lea, C.A. Coulson, The distribution of the numbers of mutin bacterial populations, J. Genet. 49 (1949) 264–285.
35] S.R. Kushner, H. Nagaishi, A.J. Clark, Indirect suppression ofand recC mutations by exonuclease I deficiency, Proc. Natl. ASci. U.S.A. 69 (1972) 1366–1370.
36] S.C. Winans, S.J. Elledge, J.H. Krueger, G.C. Walker, Site-direinsertion and deletion mutagenesis with cloned fragments inEs-cherichia coli, J. Bacteriol. 161 (1985) 1219–1221.
37] T. Lindahl, S. Ljungquist, W. Siegert, B. Nyberg, B. SpereDNA N-glycosidases: properties of uracil-DNA glycosidase fromEs-cherichia coli, J. Biol. Chem. 252 (1977) 3286–3294.
adduct highly mutagenic in mammalian cells, Biochemistry(1996) 11487–11492.
46] S. Langouet, M. Muller, F.P. Guengerich, Misincorporation of dNopposite 1,N2-ethenoguanine and 5,6,7,9-tetrahydro-7-hydroxoxoimidazo[1,2-a]purine in oligonucleotides byEscherichia colpolymerases I exo- and II exo-, T7 polymerase exo-, human imodeficiency virus-1 reverse transcriptase, and rat polymeraseBiochemistry 36 (1997) 6069–6079.
47] X. Tan, A.P. Grollman, S. Shibutani, Comparison of the mutagproperties of 8-oxo-7,8-dihydro-2′-deoxyadenosine and 8-oxo-7dihydro-2′-deoxyguanosine DNA lesions in mammalian cells, Ccinogenesis 20 (1999) 2287–2292.
48] S. Akasaka, F.P. Guengerich, Mutagenicity of site-specificallycated 1,N2-ethenoguanine in Chinese hamster ovary cell chrommal DNA, Chem. Res. Toxicol. 12 (1999) 501–507.
49] M. Hill-Perkins, M.D. Jones, P. Karran, Site-specific mutagenesvivo by single methylated or deaminated purine bases, Mutat.162 (1986) 153–163.
50] D.J. Jin, C.A. Gross, Mapping and sequencing of mutations inEscherichia colirpoB gene that lead to rifampicin resistance, J. MBiol. 202 (1988) 45–58.
Copyright © 2022 FDOKUMEN




















