
The obesogenic effects of polyunsaturated fatty acids are ...
Upload
independentCategory
view
1download
0
Role of Essential Fatty Acids in the Function of the Developing Nervous System
Ricardo Uauy a, b,., Patricio Peirano a, Dennis Hoffman b, Patricia Mena a, David Birch b, and Eileen Birch b
alnstitute of Nutrition and Food Technology (INTA), University of Chile, Santiago, Chile, and bRetina Foundation of the Southwest, Dallas, Texas
ABSTRACT: The basis for n-3 fatty acid essentiality in humans includes not only biochemical evidence but functional mea- sures associated with n-3 deficiency in human and nonhuman primates. Functional development of the retina and the occipi- tal cortex are affected by (~-Iinolenic acid deficiency and by a lack of docosahexaenoic acid (DHA) in preterm infant formulas and, as reported more recently, in term diets. Functional effects of n-3 supply on sleep-wake cycles and heart rate rhythms sup- port the need for dietary n-3 fatty acids during early develop- ment. Our results indicate that n-3 long-chain polyunsaturated fatty acids should be considered provisionally essential for infant nutrition. DHA may also be required by individuals with inherited metabolic defects in elongation and desaturation activity, such as patients with peroxisomal disorders and some forms of retinitis pigmentosa.
Lipids 31, S-167-S-176 (1996).
Dietary essential fatty acids (EFA) have been considered part of the lipid supply necessary for energy, growth, cellular metabolism, and muscle activity. The fact that some EFA serve as indispensable dietary precursors for eicosanoid (prostaglandins, prostacyclins, thromboxanes, leukotrienes) and docosanoid formation has given greater significance to their study. During the past decade, increasing attention has focused on the role of n-3 and n-6 EFA in normal fetal and neonatal development (1-7).
Basis for nutritional essentiality. The concept that specific components of fat may be necessary for the proper growth
*To whom correspondence should be addressed at INTA, University of Chile, Casilla 138-11, Santiago, Chile. Abbreviations: AA, arachidonic acid (20:4n-6) ; ADRP, autosomal dominant retinitis pigmentosa; AS, active sleep; CNS, central nervous system; DHA, docosahexaenoic acid (22:6n-3); DPA (n-6) and DPA, docosapentaenoic acid (22:5n-6 and 22:5n-3); EFA, essential fatty acids; EPA, eicosapentaenoic acid (20:5n-3) ; ERG, electroretinogram; FA, fatty acids; HM, human milk; HRV, heart rate variability; LA, linoleic acid (18:2n-6) ; LCPUFA, long-chain polyun- saturated fatty acids; LNA, ot-linolenic acid (18:3n-3) ; MAR, minimal angle of resolution; QS, quiet sleep; RBC, red blood cells; REM, rapid eye movement; RP, retinitis pigmentosa; TPN, total parental nutrition; VEP, visual evoked potential; XLRP, X-linked retinitis pigmentosa.
and development of animals and possibly humans was intro- duced in the 1930s (8). Yet, EFA were considered of margin- al importance until the 1960s, when signs of clinical deficien- cy became apparent in infants fed skim milk-based formula and in those given lipid-free parental nutrition (9-12). During the past decade, it has become generally accepted that n-6 as well as n-3 fatty acids (FA) play a key role in perinatal nutri- tion, especially for the developing central nervous system (CNS).
Clinical EFA deficiency is associated with a set of specific findings (7,9-12). This syndrome has been related mostly to combined n-3 and n-6 deficits and includes poor growth; skin lesions; decreased skin pigmentation; loss of muscle tone; degenerative changes in kidney, lung, and liver; increased metabolic rate; impaired water balance; increased fragility and permeability of cell membranes; and increased suscepti- bility to infection (7,9-12).
Clinical manifestations disappear after administration of diets that provide 2% or more of the total calories as linoleic acid (LA, 18:2n-6) (9-14). More subtle clinical symptoms appear in n-3 EFA deficiency. They include skin changes unresponsive to LA supplementation, abnormal visual func- tion, and peripheral neuropathy (15). Nervous system mani- festations are likely caused by an insufficient supply of the metabolic derivative of 0t-linolenic acid (LNA, 18:3n-3), namely docosahexaenoic acid (DHA, 22:6n-3).
Structural role of EFA in the developing nervous system. Fetus and placenta are fully dependent on maternal EFA sup- ply for growth and development. The mother's diet before pregnancy plays an important role in determining maternal EFA status, since these essential nutrients are stored in adi- pose tissue and can be mobilized over time through lipolysis. Major EFA deposition in the human fetus occurs during the third trimester; phospholipids in placental vessels and uterine vasculature are dependent on EFA supplied by the mother for eicosanoid formation. Maternal preconceptional nutrition determines, in part, which specific fats accumulate in the con- ceptus and placental tissues (6,16,17).
Copyright �9 1996 by AOCS Press S-'t 67 Lipids, Vol. 31, Supplement (1996)

S-168 R. UAUY ETAL.
The dry weight of the human brain is predominantly lipid; 22% of the cerebral cortex and 24% of white matter consist of phospholipids. Studies of several animal species and recent evi- dence from humans established that brain phospholipid arachi- donic acid (AA, 20:4n-6) and DHA decrease, while n-9 and n-7 mono- and polyunsaturated FA increase when LA and LNA or only n-3 FA are deficient in the diet (18-20). Typically, n-3 FA-deficient cells have decreased DHA and increased levels of the end products of n-3 and n-6 metabolism, DHA and docos- apentaenoic acid (DPA, 22:5n-6), respectively. Within subcel- lular organelles, synaptosomes and mitochondria seem to be more sensitive to a low dietary n-3 supply (19-21).
High concentrations of long-chain derivatives of EFA in the cerebral cortex and retina suggests their participation in neural and visual function. The n-3 end product, DHA, reaches levels of up to 50% of total FA in the phospholipids of these tissues (22-24). A specific association of either DHA or AA with neur- al tissue function has not been fully defined. In the human, Clandinin and colleagues (24) showed significant increases in AA and DHA contents of brain tissue during the last trimester of gestation. More recently, Martinez (25) provided complemen- tary data on retinal and brain accretion of long-chain polyunsat- urated fatty acids (LCPUFA), using modem methodology.
The potential for dietary EFA deficiency has become a significant issue for the nutrition of preterm-bom infants, since they do not receive the third-trimester intrauterine sup- ply of DHA and AA. Even full-term infants are at risk of DHA deficiency since most formulas are devoid of this criti- cal EFA. Recent postmortem investigations in term infants suggest a dependency on diet in the FA composition of brain cortex. Infants who are breast fed have higher DHA and lower
DPA (n-6) and AA in their cortex phospholipids relative to infants receiving cow milk-based formulas (26, 27). DHA content increases with advancing age in breast-fed infants and is proportional to the duration of human-milk feeding, while AA content remains stable. Furthermore, the dietary ratio of n-6 to n-3 FA appears relevant, as the lowest DHA and high- est DPA (n-6) contents in brain cortex phospholipids were observed in preterm infants fed formulas with a high LA to LNA ratio (27).
Biochemical and functional effects of EFA in the develop- ing nervous system. Studies in nonhuman primates character- ized biochemical and functional effects of dietary n-3 deficiency. Animal models were developed using purified LA as the only dietary fat or using safflower or sunflower oil in which the n-6/n-3 FA ratio (approximately 250:1) was very high (28-31). High n-6/n-3 FA ratios are found in powdered infant formulas currently in use in some parts of the world (32, 33). If these diets are fed from conception and/or during early life, significant declines in DHA levels and elevations of DPA (n-6) contents of brain, retina, and other tissues occur in animals. Similar data was obtained from plasma and red blood cell (RBC) lipids in preterm infants fed corn oil-based formulas that have a high LA/LNA ratio (34).
Use of RBC lipids as a potential index of EFA status in retinal membranes is based on correlations between FA profiles in RBC and neural/retinal tissue from rats and non- human primates and in human infants (27, 35, 36).
Effects of n-3 deficiency, specifically DHA, were demon- strated on retinal photoreceptor function and maturation of corti- cal visual function in nonhuman primates and in preterm infants. Improved cognitive function measured by psychometric tests
FIG. 1. Docosahexaenoic acid (DHA) in total red blood cell (RBC) lipids and plasma phospholipids (PL) of preterm infants on study diets at 36 wk postconception. Arrows indicate significant differences (P< 0.05) between diet groups using Newman-Keuls multiple comparison analysis: inset; rod electroretinogram (ERG) thresholds (intensity units = log scotopic troland seconds) and visual evoked potential (VEP) visual acuity (Snellen equivalents) in study infants at 36 wk.
Lipids, Vol. 31, Supplement (1996)

ROLE OF ESSENTIAL FATTY ACIDS S-169
were also shown in DHA-supplemented infants at 12 mon of age.
Infants fed fat-free parental diets provide nutritional models to study the effect of EFA deficiency on the developing brain. Initially, total parental nutrition (TPN) did not include lipids in its formulation. This allowed Fagioli et al. (37) to charac- terize effects of long-term dietary EFA deficiency on sleep organization. Studies in children who were maintained on fat- free TPN for 2 to 6 mon had consistently reduced durations of slow-wave sleep compared with children who received TPN plus essential lipids. The main difference was a lower cumu- lative amount of slow-wave sleep time from the second hour of night-sleep onward. These results suggest that early lipid nutrition may modulate sleep regulation and may have impli- cations for whole-body energy expenditure during sleep, since slow-wave sleep is associated with a lower metabolic rate than are other stages of sleep (38). Interestingly, no significant difference was found in either total sleep time or intrasleep waking in the EFA-deficient group. Alterations in sleep-wake cycle development may become evident only if the methods used include evaluations that are more subtle than total sleep or waking time measures. The possibility that fat-free TPN affected structural brain lipids is suggested from these results (26, 27, 37).
Studies in preterm infants fed controlled EFA intake. Our studies (conducted in Dallas, Texas, and continued as a col- laborative work in both Dallas and Santiago, Chile) have served to document the essentiality of n-3 FA in humans. Initial studies were performed in very-low-birth-weight infants, since they are particularly vulnerable to deficiency, given the virtual absence of adipose tissue stores at birth. This
deficiency may be attributable to a possible immaturity of FA elongation/desaturation pathways and inadequate LNA and DHA intakes provided by formula. We and others conducted studies to evaluate the effect of n-3 in very low birth weight infants, examining effects of LNA or LNA plus DHA supple- mentation on plasma and tissue lipid composition, retinal elec- trophysiologic function, maturation of the visual cortex, and other measures of infant growth and development (39-45).
Before the 1990s, low levels of LNA were present in most infant formulas; virtually all infant formulas in developed countries are currently supplemented with LNA, and several manufacturers in Europe and Japan have added DHA or DHA plus AA to preterm formula (32, 33, 46). EFA composition of human milk (HM) and study formulas used in our studies were published previously (34). Briefly, corn oil-based for- mula corresponded to commercial, powdered premature for- mula, contained 24% LA, was extremely low in LNA (0.5%), and had no LCPUFA; soy oil-based formula was equivalent to present ready-to-feed premature formula used in the United States; and the third study formula was also soy oil-based (1.4% LNA) but was enriched with marine oil to give an n-3 LCPUFA content closer to that found in HM. At the time we started our study, n-3 LCPUFA could only be obtained from marine oil. We chose to include sufficient marine oil to mimic DHA content of HM, since we were uncertain as to how effectively preterm infants could convert eicosapentenoic acid (EPA, 20:5n-3) to DHA. The full nutritional content of the preterm milk formula and details of laboratory methods have been published (34, 39).
Composition of RBC lipids for all diet groups were similar on entry into the study (34). At the 36- and 57-wk postconception
FIG. 2. Docosahexaenoic acid (DHA) in total red blood cell (RBC) lipids and plasma phospho- lipids (PL) of preterm infants on study diets at 57 wk postconception. Arrows indicate significant differences (P < 0.05) between diet groups using Newman-Keuls multiple comparison analysis: inset; rod electroretinogram (ERG) thresholds (intensity units = log scotopic troland seconds) and visual evoked potential (VEP) visual acuity (Snelfen equivalents) in study infants at 57 wk.
Lipids, Vol. 31, Supplement (1996)

S-170
follow-up, marked differences in DHA contents were evident in RBC lipids of all diet groups. Infants fed the n-3 FA-deficient corn oil-formula had significant reductions in DHA as compared to other formula groups. The marine oil group presented DHA concentrations elevated above all other groups. DPA (n-6) rela- tive content found at 36 and 57 wk in RBC lipids was inversely related to DHA content. DPA (n-6) was significantly elevated in the corn oil group, in comparison to the HM and soy/marine groups. By 57 wk, differences in content of the endproducts of n- 3 and n-6 FA metabolism [i.e., DHA and DPA (n-6), respective- ly] were greatly accentuated, compared to 36-wk values. The soy oil group had intermediate DHA and DPA (n-6) values relative to the corn and soy/marine oil groups but differed significantly from the DHA-supplemented infants (34). Figures 1 and 2 sum- marize changes in DHA content of RBC and plasma phospho- lipids observed at 36 and 57 wk postconception, respectively.
Methods for assessment of visual function have been described elsewhere, and detailed results of our studies have been recently published (39-41,47-51). Electroretinographic responses demonstrated significantly higher threshold values from rod photoreceptors in the n-3-deficient group (corn oil- fed) at the 36-wk follow-up relative to groups receiving n-3 FA (26, 28), indicating that the sensitivity and maturity of rod photoreceptors in n-3-deficient infants were reduced significantly compared to infants fed soy/marine formula or HM. A group of 35-wk gestational age infants had their visu- al maturity tested a few days (3-5 d) after birth and served as a normal standard for comparison. This group of infants receiving transplacentally acquired EFA were nearly identical in all rod electroretinogram (ERG) functional indices to the equivalent conceptional age HM-fed or marine oil-fed pre- mature infants. The soy oil-fed group had higher threshold values than did the soy/marine oil-fed group. Cone photore- ceptor function was not significantly affected by diet, although trends were similar to results obtained from rods. At the 57-wk follow-up, corn oil-fed infants consistently had longer implicit times in light-adapted oscillatory potentials. The soy oil-fed infants had some response peaks differ significantly from the HM-fed infants, while the soy/marine and HM groups were similar in oscillatory potential respons- es. Oscillatory potentials are generated in the inner retina, mature later than distal retinal responses and most likely reflect ganglion cell-amacrine cell interactions (40).
The soy/marine oil- and the HM-fed groups had lower val- ues of log MAR (minimal angle of resolution) (i.e., better acu- ity) relative to infants fed formula devoid of DHA at the 57-wk follow-up using either visual evoked potentials (VEP) or forced-choice preferential looking response methods. A log MAR value of 0 corresponds to 20/20 Snellen equivalents, while a value of 1 corresponds to 20/200. A significant negative correlation was found between RBC DHA levels and log MAR values; that is, when erythrocyte DHA levels were high, log MAR values were correspondingly low (i.e., better visual acuities). HM- and soy/marine oil-fed infants had the highest DHA/DPA (n-6) ratio in RBC and also had the best visual acu- ity, i.e., lowest log MAR values. The group that received soy oil as a source of n-3 had poorer VEP acuity at the 57-wk follow-
Lipids, Vol. 31, Supplement (1996)
R. UAUY ETAL.
up relative to the soy/marine oil-fed group, indicating that, despite ample LNA, visual function was less mature than in the DHA-supplemented infants. A group of healthy full-term, breast-fed infants matched by conceptional age was used as a control. HM- and soy/marine oil-fed groups were virtually identical to the control group, while com and soy groups had poorer visual acuities (41, 48). Insets of Figures 1 and 2 sum- marize changes in ERG threshold and in VEP acuity observed in the four diet groups at 36 and 5"7 wk postconception, respec- tively. Visual acuity at 4 mon in full-term HM-fed infants was significantly better than in formula-fed infants and was virtual- ly identical to the HM- and marine oil-fed premature groups.
Reversibility of changes in visual function observed in preterm infants cannot be fully answered from our data since the study was terminated at 57 wk postconception. Most of the ERG alterations at 36 wk normalized with time, based on the 57-wk results, except for the ganglion cell-dependent, oscillatory potentials.
Other functional effects are being evaluated in our present studies in premature infants fed HM or one of three formulas (control, lacking LCPUFA; supplemented with AA and DHA; or supplemented solely with DHA). These measures include organization of sleep-wake cycles during early development, changes in heart rate patterns during sleep states, and sponta- neous activity detected by piezoelectric accelerometers dur- ing various sleep states and the wakeful period. At the time of this report, the study remains masked; thus results comparing LCPUFA-supplemented groups vs. the unsupplemented group have not been made, but comparisons between formu- la- and breast-fed groups are revealing.
Since intrauterine life is characterized by the virtual absence of wake periods (52) and since postnatal infants spend most of their time sleeping, study of sleep-wake cycles is an important component in evaluation of CNS develop- ment. Specific developmental patterns in infants have been described, and long-term effects of changes in sleep-wake cycles have been documented.
Sleep-wake cycle has been well characterized as a function of age during the first months of life (53-56). It is defined by a combination of physiological recordings, electroencephalo- gram, electrooculogram, electromyogram, cardiorespiratory pattern, and motor activity. Several striking and precise changes, with specific timing for each, occur during this peri- od, reflecting CNS maturation and adaptation to the extrauter- ine environment. Typically, the full-term newborn has significantly less indeterminate sleep, with well-differentiated active sleep (AS) and quiet sleep (QS), than do preterm infants whose indeterminate sleep may represent up to 30% of total sleep time. The architecture of the sleep-wake cycle in infants differs markedly from that of older age groups. Day-night dif- ferences in sleep-wake cycles, with an increase in the ability to remain asleep during the night and remain awake during the day for progressively longer periods, begin to arise, reflecting emergence of distinctive adult sleeping traits. These trends are relevant developmental indices of brain activity and CNS functional integrity. Information on the developmental pattern for these changes is limited by the scant data available

ROLE OF ESSENTIAL FATTY ACIDS
between 6 and 24 mort of age and by the short duration (<12 h) of recording periods in most published studies.
Following the initial observations of Fagioli et al. (37) on the effect of EFA deficiency on sleep-wake organization, we have included polygraphic evaluation of sleep organization during early development. Our initial findings (Uauy, R., and Peirano, P., unpublished observations) confirm previous reports (57) of less AS observed in HM-fed infants; in addi- tion, these infants showed longer duration of QS episodes, indicating a more mature pattern of CNS development. In addition, within the randomized formula groups, we find dif- ferences in AS and QS duration and in the percentage of sleep cycles within the sleep episode.
Changes during development in spontaneous heart rate variability (HRV) patterns have been associated with sympa- thetic and parasympathetic interactions and utilized as an index for normal brain maturation. In healthy neonates, the variability of the high-frequency component is higher in QS than in AS, while the low-frequency component is higher in AS. During development, there is a progressive increase in HRV both during AS and QS, with specific trends for each type of HRV. There is a progressive dominance of the sympa- thetic tone in AS, also called rapid eye movement (REM) sleep, and a predominance of the parasympathetic vagal tone in QS or non-REM sleep. Our preliminary findings (Uauy, R., and Peirano, P., unpublished observations) support published reports of a higher HRV during AS and QS in HM-fed infants, indicating a more mature pattern of CNS development (58, 59). The effect is observed in all frequency components but is more marked in the high-frequency component, suggesting a higher parasympathetic vagal tone in the HM-fed group. Within the randomized formula groups, we find differences in HRV. These differences are not observed during wake periods.
Measurement of spontaneous motor activity using piezo- electric transducers permits continuous recording of move- ments during sleep and wakefulness. Observations in healthy infants demonstrate a progressive decrease in motor activity with advancing age in both AS and QS (60). We have prelimi- nary evidence (Uauy, R., and Peirano, P., unpublished observa- tions) indicating that HM-fed infants tend to have more spon- taneous activity during sleep and that the duration of move- ments is longer, particularly during the early wake period.
Studies in term infants f e d EFA-controlled diets. We con- ducted a 3-yr follow-up of healthy, full-term infants to test if HM feeding, which contains significant amounts of DHA, had an effect on the development of visual function (42). The cohorts were breast fed from birth to at least 4 mon or fed for 12 mon formula containing ample LA and 0.5% of total fat as LNA. The breast-fed group was weaned to an oleic acid (18:1)-predominant formula and received egg yolk through 12 mon of age. The breast-fed group maintained higher plas- ma and RBC membrane phospholipid DHA concentrations throughout the first year of life. At 3 yr of age, stereo acuity, as measured by operant preferential looking techniques, was more mature in breast-fed infants relative to the formula-fed group: 92% of the breast-fed group had mature operant pref- erential looking stereo acuity, whereas 35% of infants in the
S-171
formula-fed group met the maturity criteria. Visual recogni- tion in the breast-fed group was also better: while only 61% of the formula-fed infants had perfect scores, 93% of the breast-fed group had perfect scores (42).
These and other data produced by Makrides et al. (61) are suggestive of a DHA effect on visual maturation in term infants. Definitive proof, however, must await the final results of randomized, blind, controlled clinical trials of DHA and DHA + AA supplementation in term infants. In addition, the possibility of optimizing LNA to DHA conversion by provid- ing sufficient LNA (>0.7% of total energy) and a lower LA to LNA ratio in formula (<10:1) may also provide for sufficient DHA for normal visual function. Initial results of Ponder et al. (62) indicate that this alternate strategy may be simpler and less expensive than addition of n-3 LCPUFA.
A recent study by Innis et al. (63) concluded that the need for DHA supplementation should not be based on the alleged improved visual acuity of breast-fed infants, since, despite differences in DHA blood contents, their study revealed no differences in preferential looking acuity between breast-fed and unsupplemented formula-fed infants. The formula used in this study contained 2.1% of total fat as LNA but no LCPU- FA. However, this study used a Teller acuity card protocol for acuity assessment, a method which is less sensitive than elec- trophysiologic techniques to subtle visual deficits.
Other controlled studies without special reference to EFA. The first studies proposing that human breast milk feeding might exert a beneficial effect on infant psychomotor devel- opment were published during the 1920s. Since then, several studies reported small but significant advantages in develop- mental indices for breast-fed infants. Although no study cate- gorically demonstrated the cause for this effect, most authors suggested that nutritional factors in human milk may account for this. Over the past decade, a renewed interest in this ques- tion has arisen, and several authors suggested that EFA may play a role in the improved developmental scores during infancy and preschool years observed in HM-fed infants. These studies were reviewed recently, but most are retrospec- tive in nature (64).
In the Rodgers study (65), one of the few long-term fol- low-up studies of mental development comparing breast-fed vs. formula-fed children, where family, social, and economic variables were controlled, it was found that early breast feed- ing was associated with better picture intelligence at g yr of age and better scores in mathematics, nonverbal ability, and sentence completion at 15 yr of age. The study cohorts were selected from all births occurring during one week in 1946 in Great Britain. Based on confirmed records of early diet, 1, 133 individuals were considered entirely bottle fed, and 1, 291 individuals were never bottle fed. Functional benefits were proportional to the duration of breast feeding and remained statistically significant after controlling for social, cultural, and demographic variables by multivariate analysis. These observations could not be extrapolated to current for- mula-fed infants because, during the observation period, chil- dren were fed unmodified, diluted cow milk with the sole addition of sugar. Protein may have been excessive, and EFA
Lipids, Vol. 31, Supplement (1996)
S-172 R. UAUY ETAL.
may have been inadequate. In a recent study by Rogan et al. (66), 855 newborns receiving modern formula were enrolled from 1978-1982 and followed through school age. Cognitive development was assessed with appropriate methodology and confirmed the results of the earlier British study. Breast-fed infants had significantly higher scores in the Bayley scale of mental development at 2 yr, and in the McCarthy scale at 3 and 4 yr. In addition, slightly higher English grades on report cards were found in the breast-fed group after adjusting for relevant confounding variables. A recent publication from a study in Spain confirms the benefits in mental development and the lack of an effect on motor abilities (67).
A possible long-term effect of early HM feeding on brain development is also suggested by results from a controlled trial of preterm infants indicating that HM feeding by naso- gastric tube for 28 d was associated with a +8.3 point intelli- gence quotient difference at 8 yr of age relative to a formula- fed group after controlling for socioeconomic and other maternal variables (68). These controlled observations in preterm infants indicated that HM feeding may, in fact, offer unique advantages for brain development. More recently, a study of 526 full-term infants born in Groningen, The Netherlands, between 1975 and 1979 who were followed for 9 yr demonstrated a significantly lower occurrence of neuro- logical dysfunction in the 135 breast-fed infants (breast fed for more than 3 wk) after adjusting for obstetric, perinatal, neonatal neurological, and social differences (69).
Differences in the early functional organization of the CNS between breast- and bottle-fed infants have also been report- ed. Nocturnal sleep-wake state organization at 4 mo of age show that breast-fed infants spend a higher percentage of time in QS and a lower percentage of time in AS as compared to formula-fed infants (57). Furthermore, heart rate values were lower in all sleep-wake states in breast-fed infants (70). Even though total energy expenditure during AS and QS did not differ between feeding groups, the different organization of sleep states throughout the night might account for differ- ences in energy expenditure between feeding groups (57).
There are indications that breast-fed neonates even at the age of 2 d postpartum are better organized, physiologically, than are bottle-fed neonates (58,59). They exhibit a more orga- nized HRV pattern and a slower heart rate in both AS and QS. They also present higher incidence of low-frequency, slower heart rate, greater HRV, and elevated vagal tone, indicating more mature HRV patterns. Conversely, breast-fed neonates were also reported to be behaviorally more irritable, more difficult to test, and less able to be consoled once crying (59).
Evidence from patients with inborn errors of metabolism. There is yet no conclusive proof for an inherited metabolic defect of DHA and or AA synthesis causing altered brain or retinal function. There is increasing evidence, however, that peroxisomal diseases may be associated with defective for- mation of C22 LCPUFA. Recently, Bennett et al. (71) report- ed fascinating but uncontrolled observations on the improve-
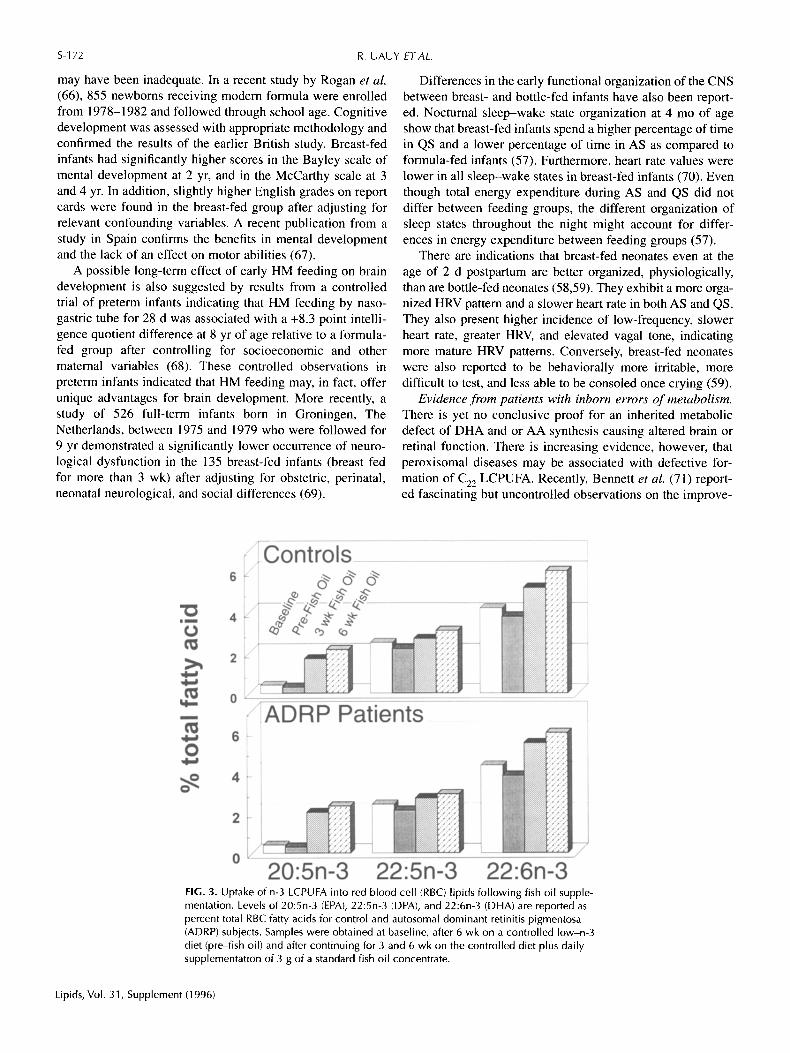
FIG. 3. Uptake of n-3 LCPUFA into red blood cell (RBC) lipids following fish oil supple- mentation. Levels of 20:5n-3 (EPA), 22:5n-3 (DPA), and 22:6n-3 (DHA) are reported as percent total RBC fatty acids for control and autosomal dominant retinitis pigmentosa (ADRP) subjects. Samples were obtained at baseline, after 6 wk on a controlled low-n-3 diet (pre-fish oil) and after continuing for 3 and 6 wk on the controlled diet plus daily supplementation of 3 g of a standard fish oil concentrate.
Lipids, Vol. 31, Supplement (1996)
ROLE OF ESSENTIAL FATTY ACIDS S-173
ment in neurological, retinal, and cortical electrophysiologic responses and in developmental tests in Batten's disease patients after n-3 supplementation. We studied lipid abnor- malities in patients with retinitis pigmentosa (RP) and have evidence suggesting that some autosomal dominant RP (ADRP) and X-linked RP (XLRP) patients may have defec- tive formation or incorporation of LCPUFA into membranes (72-74).
Zellweger disease. Martinez (75) provided data from sever- al patients, indicating that, in peroxisomal disorders, brain DHA content is extremely low, plasma and RBC DHA levels are below normal, and biochemical response of these patients to fish oil supplementation is only partial. Sprecher (76) indi- cated that the peroxisomal [3 oxidation step necessary for C22 PUFA biosynthesis is restricted in Zellweger disease patients, since they cannot convert 24:6n-3 to 22:6n-3. Preliminary results on a limited number of patients indicate that supple- mentation with pure DHA is capable of preventing and also of reverting abnormal motor and mental functions associated with these conditions (77). Concomitant evidence from fibroblast studies of these patients provides supportive evidence as to the relative importance of peroxisomal ]3 oxidation in determining biochemical and functional consequences of this disease (77).
Batten's disease. Recently, Bennett et al. (71) described alterations in LCPUFA metabolism in juvenile Neuronal Ceroid-lipofucsinosis, also known as Batten's disease. These patients present seizures, altered psychomotor development,
retinal degeneration of rods and cones leading to blindness, motor disturbances progressing to spastic tetraplegia, and death during the first decades of life. The disease has a variable rate of progression, but in all cases it leads to mental retarda- tion and severe motor impairment. RBC n-3 LCPUFA were found to be lower in patients with the disease relative to con- trols, and a significant rise was noted in these FA after marine oil supplementation (71,78). Follow-up of 6 patients after 5 to 7 yr of variable supplementation regimens demonstrated a possible amelioration in the rate of progression compared to what was predictable based on the natural history of the dis- ease (71). Given the uncontrolled nature of these observations and the variable natural course of progression in this condi- tion, caution should be exercised in interpreting these findings. A controlled multicenter trial is needed to clarify the clinical response to LCPUFA supplementation in these patients.
Retinitis pigmentosa. We and others have observed abnor- malities in DHA metabolism in patients with RP (72-74, 79-83). RP and associated retinal degenerations are character- ized by progressive night blindness and visual field constric- tion resulting from photoreceptor degeneration. Tissue levels of DHA and other LCPUFA may be involved in the patho- physiology of RE There is an accumulating body of evidence that selective reductions in the plasma lipid levels of DHA may be associated with various forms of hereditary retinal degeneration (79-83). However, there were lower 18:4n-3 and higher 22:6n-3 levels in the plasma lipids of ADRP patients.
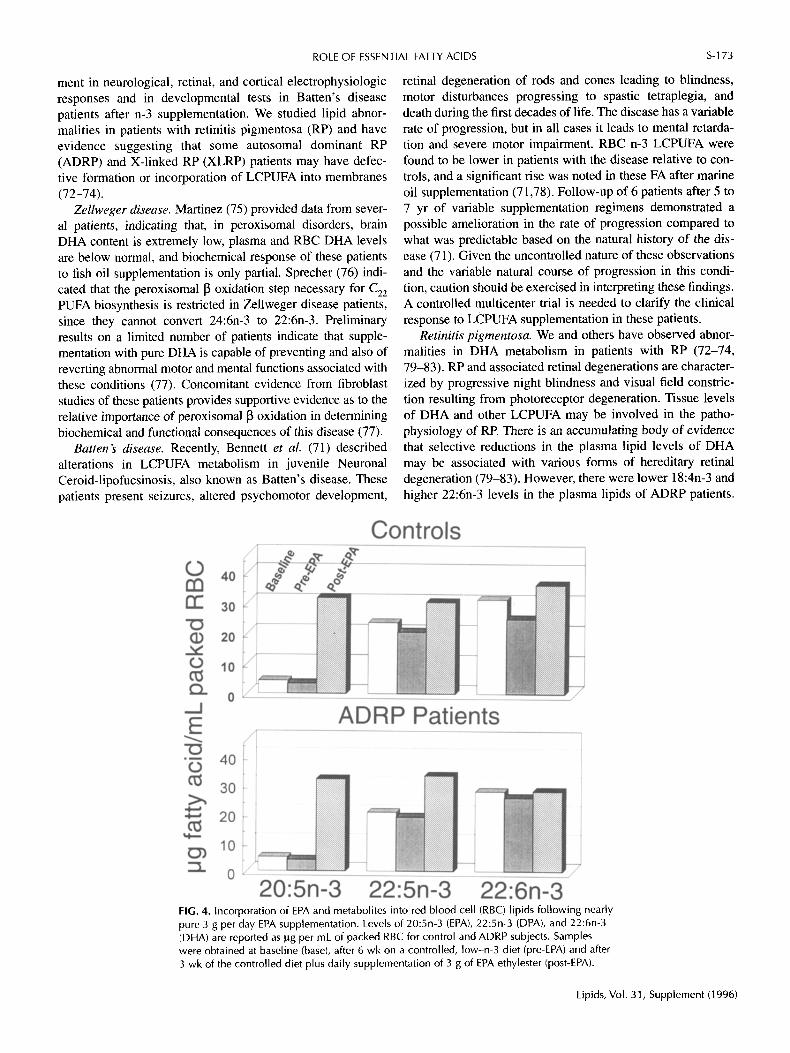
FIG. 4. Incorporation of EPA and metabolites into red blood cell (RBC) lipids following nearly pure 3 g per day EPA supplementation. Levels of 20:5n-3 (EPA), 22:5n-3 (DPA), and 22:6n-3 (DHA) are reported as ~g per mL of packed RBC for control and ADRP subjects. Samples were obtained at baseline (base), after 6 wk on a controlled, low-n-3 diet (pre-EPA) and after 3 wk of the controlled diet plus daily supplementation of 3 g of EPA ethylester (post-EPA).
Lipids, Vol. 31, Supplement (1996)

S-174
Segregation of patients into subgroups based on genetic inher- itance type revealed that the greatest deviation from normal in plasma DHA levels was found in X-linked patients, whereas those with autosomal dominant inheritance were not significantly different from controls (84). Furthermore, specific families affected with ADRP demonstrate FA abnor- malities, while other ADRP families have near normal FA profiles (80). Our results extend these findings by showing rel- atively similar levels of DHA in RBC among members within a family with ADRP. In ADRP, deficits in DHA levels have most frequently been identified in plasma lipids, which are rapidly modified by dietary fat intake. Abnormal lipid metab- olism may result in a subnormal availability of DHA for dis- tribution to peripheral target tissues. A limited supply of DHA for photoreceptor disc membrane structure may contribute to abnormal retinal function.
We have assessed the FA status in plasma and RBC of 50 subjects with ADRP as compared to that of 20 normal-sight- ed controls (72). In plasma lipids, patients with ADRP had significantly lower levels of total saturates than did the con- trols; derivatives of the n-6 series were significantly higher in patients with ADRP, in contrast to the lack of significant dif- ferences in n-3 FA between patients and controls. There were lower 18:4n-3 and higher 22:6n-3 in the plasma lipids of ADRP patients, however. Furthermore, a higher unsaturation index was found in the plasma lipids of patients with ADRP. In contrast to plasma, the RBC lipid FA profile of patients with ADRP compared to controls had significantly higher lev- els of monounsaturates and 18-carbon EFA of both the n-6 and n-3 series. Levels of n-6 LCPUFA levels were reduced in patients, particularly AA and 22:4n-6, which were markedly reduced. Overall, total n-3 LCPUFA were significantly lower than in controls. Consistent with this, the unsaturation index in RBC of patients with ADRP was reduced significantly. Full-field ERG were obtained from all patients under 50 yr of age to evaluate retinal function. Patients with lower RBC DHA levels (<3% of total FA) had significantly lower rod/cone amplitude ratios than did patients with higher RBC DHA levels (>4%) (72). Although FA changes found in ADRP patients were of moderate magnitude (<25%) com- pared to controls, these changes may have a significant impact on visual function if persistent over many years. This may be of particular importance for a patient who has visual function already compromised by a gene mutation.
Based on these observations, we undertook supplementa- tion trials in a small group of patients with ADRP under con- trolled dietary conditions. The first trial assessed the capacity to absorb and incorporate n-3 LCPUFA as provided by a stan- dardized fish oil concentrate. Results of this study revealed that patients did as well as controls in incorporating n-3 LC- PUFA into RBC membranes, as shown in Figure 3. The sec- ond trial addressed the conversion of EPA to DHA, after oral administration of nearly pure EPA ethyl ester for 3 wk. Results of this study, as shown in Figure 4, demonstrated sim- ilar incorporation of 20:5n-3 and conversion to 22:5n-3 RBC for ADRP and control subjects after EPA supplementation. DHA in ADRP patients remained unchanged while controls
Lipids, Vol. 31, Supplement (1996)
R. UAUY ETAL.
exhibited a 27% higher incorporation into RBC after EPA was administered (73). These results suggest the presence of a metabolic defect in the final elongation/desaturation or [3 oxi- dation steps necessary for DHA formation from EPA.
We propose that low plasma and RBC DHA levels attrib- utable to impaired transport or metabolism of LCPUFA may coexist with certain gene mutations and contribute to the severity of visual impairment in those with ADRP.
Our studies provide clear evidence that dietary n-3 FA deficiency affects eye and brain functions of preterm infants as measured by ERG, cortical VEP, and behavioral testing of visual acuity. Preterm infants require DHA in their diet because they are unable to form them in sufficient quantity from LNA provided by soy oil-based formula products. Dietary n-3 and n-6 FA intake resulted in discernible differ- ences in the FA composition of plasma and RBC membrane lipids. Changes in membrane chemical structure are likely responsible for the observed functional effects. Definitive rec- ommendations for term infants await results of randomized trials of DHA supplementation.
Preliminary evidence indicates that peroxisomal disorders, autosomal dominant and X-linked forms of RP, and possibly Batten's disease are associated with altered n-3 metabolism and may represent defects in DHA biosynthesis or incorpora- tion into membranes. Clinical trials supplementing with pure DHA are required to evaluate this possibility further.
Our results indicate that n-3 FA are required for optimal nervous system development and for maturation of the retina in human infants and thus they should be considered essential nutrients. DHA is required by preterrn infants and possibly by individuals with genetic defects in LCPUFA metabolism, such as those with peroxisomal disorders and some forms of RP.
ACKNOWLEDGMENTS Work by the authors was supported in part by Fondecyt Chile grant 1930820 and by NIH grants HD 22380, EY 05235, EY 05236, and the Foundation Fighting Blindness.
REFERENCES
1. Sprecher, H. (1981) Biochemistry of Essential Fatty Acids, Prog. Lipid Res. 20, 13-22.
2. Bazan, N.G. (1989) The Metabolism of Omega-3 Polyunsaturated Fatty Acids in the Eye: The Possible Role of Docosahexaenoic Acid and Docosanoids in Retinal Physiology and Occular Pathology, Prog. Clin. Biol. Res. 312, 95-112.
3. Willis, A.L. (1984) Essential Fatty Acids, Prostaglandins, and Related Eicosanoids, in Present Knowledge in Nutrition (Olson, R.E. ed.) The Nutrition Foundation Inc., Washington, D.C., pp. 90-113.
4. Uauy, R., and Hoffman, D.R. (1991) Essential Fatty Acid Requirements for Normal Eye and Brain Development, Sernin Perinatol. 15, 449-455.
5. Simopoulos, A.P. (1991) Omega-3 Fatty Acids in Health and Disease and in Growth and Development, Am. J. Clin. Nutr. 54, 438-463.
6. Innis, S.M. (1991) Essential Fatty Acids in Growth and Development, Prog Lipid. Res 30, 39-103.
7. Uauy, R., Treen, M., and Hoffman, D.R. (1989) Essential Fatty Acid Metabolism and Requirements During Development, Semin. PerinatoL 13, 118-130.

ROLE OF ESSENTIAL FATTY ACIDS S-175
8. Burr, G.O., and Burr, M.M. (1929) A New Deficiency Disease Produced by Rigid Exclusion of Fat from the Diet, J. Biol. Chem. 82, 345-367.
9. Hansen, A.E., Wiese, H.E, Boelsche, A.N., Haggard, M.E., Adam, D.J.D., and Davis, H. (1963) Role of Linoleic Acid in Infant Nutrition: Clinical and Chemical Study of 428 Infants Fed on Milk Mixtures Varying in Kind and Amount of Fat, Pediatrics 31, 171-192.
10. Caldwell, M.D., Johnsson, H.T., and Othersen H.B. (1972) Essential Fatty Acid Deficiency in an Infant Receiving Prolonged Parental Alimentation, J. Pediatr. 81,894-898.
11. White, H.B., Turner, M.D., Turner, A.C., and Miller, R.C., (1973) Blood Lipid Alterations in Infants Receiving Intravenous Fat- Free Alimentation, J. Pediatr. 83, 305-313.
12. Paulsrud, J.R., Pensler, L., and Whitten, C.E (1972) Essential Fatty Acid Deficiency in Infants Induced by Fat-Free Intravenous Feeding, Am. J. Clin. Nutr. 25, 897-904.
13. Friedman, Z. (1980) Essential Fatty Acids Revisited, Am. J. Dis. Child. 134, 397-408.
14. Friedman, Z., Shocbat, S.J., and Maisels, M.J. (1976) Correction of Essential Fatty Acid Deficiency in Newborn Infants by Cutaneous Application of Sunflower Seed Oil, Pediatrics 58, 650--654.
15. Holman, R.T., Johnson, S.B., and Hatch, T.E (1982) A Case of Human Linolenic Acid Deficiency Involving Neurological Abnormalities, Am. J. Clin. Nutr. 35, 617-623.
16. Galli, C., and Socini, A. (1983) Dietary Lipids in Pre- and Post- Natal Development, in Dietary Fats and Health (Perkins, E.G., and Visek, W.J., eds.), American Oil Chemists' Society, Champaign, pp. 278-301
17. Ongary, M.A., Ritter, J.M., and Orchard, M.A. (1984) Correlation of Prostacyclin Synthesis by Human Umbilical Artery with Status of Essential Fatty Acid, Am. J. Obstet. Gynecol. 149, 455-460.
18. Galli, C., Trzeciak, H.I., and Paoletti, R. (1971) Effects of Dietary Fatty Acids on the Fatty Acid Composition of Brain Ethanolamine Phosphoglyceride: Reciprocal Replacement of n-6 and n-3 Polyunsaturated Fatty Acids, Biochim. Biophys. Acta. 248, 449-54.
19. Menon, N.K., and Dhopesbwarkar, G.A. (1982) Essential Fatty Acid Deficiency and Brain Development, Prog. Lipid Res. 21, 309-326.
20. Bourre, J.M., Durand, G., Pascal, G., and Youyou, A. (1989) Brain Cell and Tissue Recovery in Rats Made Deficient in n-3 Fatty Acids by Alteration of Dietary Fat, J. Nutr. 119, 15-22
21. Bourre, J.M., Francois, M., and Youyou, A. (1989) The Effects of Dietary ct-Linolenic Acid on the Composition of Nerve Membranes, Enzymatic Activity, Amplitude of Electro- physiological Parameters, Resistance to Poisons and Performance of Learning Tasks in Rats, J. Nutr. 119, 1880-1892.
22. Widdowson, E.M., and Dickerson, J.W. (1981) Composition of the Body in Geigy Scientific Tables (Letner, C., ed.) Ciba-Geigy, West Caldwell, pp. 217-225.
23. Bazan, N.G., Reddy, T.S., Bazan, H.E.E, and Birkle, D.L. (1986) Metabolism of Arachidonic and Docosahexanoic Acids in the Retina, Prog. Lipid Res. 25, 595-606.
24. Clandinin, M.T., Chappell, J.E., and Leong, S. (1980) Intrauterine Fatty Acid Accretion Rates in Human Brain: Implication for Fatty Acid Requirements, Early Hum. Dev. 4, 121-130.
25. Martinez, M. (1992) Tissue Levels of Polyunsaturated Fatty Acids During Early Human Development, J. Pediatr. 120, S129-S138.
26. Farquharson, J., Cockburn, E, and Ainslie, RW. (1992) Infant Cerebral Cortex Phospholipid Fatty-Acid Composition and Diet, Lancet 340, 810-813.
27. Makrides, M., Neumann, M.A., Byard, R.W. and Gibson, R.A. (1994) Fatty Acid Composition of Brain Retina and Erythrocytes in Breast and Formula Fed Infants, Am. J. Clin. Nutr. 60, 189-194
28. Wheeler, T.G., Benolken, R.M., and Anderson, R.E. (1975)
Visual Membranes: Specificity of Fatty Acid Precursors for the Electrical Response to Illumination, Science 188, 1312-1314.
29. Watanabe, I., Kato, M., and Aonuma, H. (1987) Effect of Dietary Alphalinolenate/Linoleate Balance on the Lipid Composition and Electroretinographic Responses in Rats, Adv. Biosci. 62, 563-568.
30. Neuringer, M., Connor, W.E., Van Petten, C., and Barstad, L. (1984) Dietary Omega-3 Fatty Acid Deficiency and Visual Loss in Infant Rhesus Monkeys, J. Clin. Invest. 73, 272-276.
31. Neuringer, M., Connor, W.E., Lin, D.S., Anderson G.J., and Barstad, L. (1991) Dietary 0)-3 Fatty Acids: Effect on Retinal Lipid Composition and Function in Primates, in Retinal Degenerations (Anderson, R.E., Hollyfield, J.G., and LaVail M.M., eds.) CRC Press, New York, pp. 1-13:
32. Koletzko, B., and Bremer H.J. (1989) Fat Content and Fatty Acid Composition of Infant Formulas, Acta. Paediatr. Stand. 78, 513-521.
33. Hansen, J. (1993) Commercial Formulas for Preterm Infants, in Nutritional Needs of Preterm Infants: Scientific Basis and Practical Guidelines (Tsang, R.C., Lucas, A., Uauy, R., and Zlotkin, S., eds.), Williams and Wilkins Caduceus Medical Publishers, Inc., Pawling, pp. 297-301.
34. Hoffman, D.R., and Uauy, R. (1992) Essentiality of Dietary Omega-3 Fatty Acids for Premature Infants: Plasma and Red Blood Cell Fatty Acid Composition, Lipids 27, 886-895.
35. Carlson, S.E., Carver, J.D., and House, S.G. (1986) High Fat Diets Varying in Ratios of Polyunsaturated to Saturated Fatty Acid and Linoleic to Linolenic Acid: A Comparison of Rat Neural and Red Cell Membrane Phospholipids, J. Nutr. 116, 718-726.
36. Connor, W.E., Lin, D.S., and Neuringer, M. (1993) Is the Docosahexaenoic Acid (DHA, 22:6n-3) Content of Erythrocytes a Marker for the DHA Content of Brain Phospholipids? FASEB J. 7, A152.
37. Fagioli, I., Baroncini, P., Ricour, C., and Salzarulo, R (1989) Decrease of Slow-Wave Sleep in Children with Prolonged Absence of Essential Lipids Intake, Sleep 12, 495-499.
38. Stothers, J.K., and Warner, R.M. (1977) Oxygen Consumption and Sleep State in the Newborn, J. Physiol. (Lond.) 269, 57-58.
39. Uauy, R.D., Birch, D.G., Birch, E.E., and Uauy, R. (1990) Effect of Dietary Omega-3 Fatty Acids on Retinal Function of Very- Low-Birth-Weight Neonates, Pediatr. Res. 28, 485-492.
40. Birch, D.G., Birch, E.E., Hoffman, D.R., and Uauy, R. (1992) Retinal Development in Very-Low-Birth-Weight Infants Fed Diets Differing in Omega-3 Fatty Acids, Invest. Ophthalmol. Vis. Sci. 33, 2365-2376.
41. Birch, E.E., Birch, D.G., Hoffman, D.R., and Uauy, R. (1992) Dietary Essential Fatty Acid Supply and Visual Acuity Development, Invest. Ophthalmol. Vis. Sci. 33, 3242-3253.
42. Birch, E.E., Birch, D.G., Hoffman, D.R., Hale, L., and Uauy, R. (1993) Breast-Feeding and Optimal Visual Development, J. Pediatr Ophtalmol Strabismus 30, 33-38.
43. Koletzko, B., Schmidt, E., and Bremer, H.J. (1989) Effects of Dietary Long-Chain Polyunsaturated Fatty Acids on the Essential Fatty Acid Status of Premature Infants, Eur J. Pediatr 148, 669-675.
44. ~nnis, S.M., Foote, K.D., and MacKinnon, M.J. (1990) Plasma and Red Blood Cell Fatty Acids of Low-Birth-Weight Infants Fed Their Mother's Expressed Breast Milk or Preterm Infant Formula, Am. J. Clin. Nutr. 51,994-1000.
45. Carlson, S.E., Cooke, R.S., and Rhodes, RG. (1991) Effect of Vegetable and Marine Oils in Preterm Infant Formulas on Blood Arachidonic and Docosahexaenoic Acids, J. Pediatr. 120, S159-S167.
46. Koletzko, B. (1995) Long Chain Polyunsaturated Fatty Acids in Infant Formulae in Europe, ISSFAL Newsletter 2, 3-5.
47. Birch, E.E., Birch, D.G., Petrig, B., and Uauy, R. (1990) Retinal and Cortical Function of Infants at 36 and 57 weeks Postconception, Clin. Vision Sci. 5, 363-373.
Lipids, Vol. 31, Supplement (1996)

S-176 R. UAUY ETAL.
48. Uauy, R., Birch, E.E., Birch, D.G., and Peirano, P. (1992) Visual and Brain Function Measurements in Studies on n-3 Fatty Acid Requirements of Infants, J. Pediatr. 120, S 168-S 180.
49. Birch, D.G. (1989) Clinical Electroretinography, Ophthalmol Clin. N. Amer 2, 469-497.
50. Birch, E.E. (1989) Visual Acuity Testing in Infants and Young Children, Ophthalmol Clin. N. Am. 2, 369-389.
51. Sokol, S. (1976) Visually Evoked Potentials: Theory, Techniques, and Clinical Applications, Surv. Ophthalmol. 21, 18-44.
52. Rigatto, H. (1992) Maturation of Breathing, Clin. Perinat. 19, 739-756.
53. Coons, S., and Guilleminault, C. (1982) The Development of Sleep-Wake Patterns and Non-Rapid Eye Movement Sleep Stages During the First Six Months of Life in Normal Infants, Pediatrics 69, 793-798.
54. Fagioli, I., and Salzarulo, P. (1982) Sleep States Development in the First Year of Life Assessed Through 24 Hour Recording, Early Hum. Dev. 6, 215-228.
55, Anders, T.E, and Keener, M. (1985) Developmental Course of Nighttime Sleep--Wake Patterns in Full Term and Premature Infants During the First Year of Life, Sleep 8, 173-192.
56. Curzi-Dascalova, L., Peirano, E, and Morel-Kahn, E (1988) Development of Sleep States in Normal Premature and Full Term Newborns, Dev. Psychobiol. 21, 431-44.
57. Butte, N.C., Jensen, C.L., Moon, J.K., Glaze, D.G., and Frost, J.D. (1992) Sleep Organization and Energy Expenditure of Breast-Fed and Formula-Fed Infants, Pediatr. Res. 32, 514-519.
58. Zeskind, ES., Marshall, T.R., and Goff, D.M. (1992) Rhythmic Organization of Heart Rate in Breast-Fed and Bottle-Fed Newborn Infants, Early Develop. Parent 1, 79-87.
59. DiPietro, J.A., Larson, K.S., and Porges, S.W. (1987) Behavioral and Heart Rate Pattern Differences Between Breast-Fed and Bottle-Fed Neonates, Dev. Psychol. 23, 467-474.
60. Peirano, P., and Curzi-Dascalova, L. (1995) Modulation of Motor Activity Patterns and Sleep States in Low Risk Prematurely Bornlnfants Reaching Normal Term: A Comparison with Full Term Newborns, Neuropediatrics 26, 8-13.
61. Makrides, M., Simmer, K., Goggin, M., and Gibson, R.A. (1993) Erythrocyte Docosahexaenoic Acid Correlates with the Visual Response of Healthy, Term Infants, Pediatr. Res. 34, 425-427.
62. Ponder, D.L., Innis, S.M., and Benson, J.D. (1992) Docosahexanoic Acid Status of Term Infants Fed Breast Milk, or Soy Oil or Corn Oil Containing Infants Formula, Pediatr. Res. 32, 638-688.
63. Innis, S.M., Nelson, C.M., Rioux, M.E, and King, J. (1994) Development of Visual Acuity in Relation to Plasma and Erythrocyte n-6 and n-3 Fatty Acids in Healthy Term Gestation Infants, Am. J. Clin. Nutr. 60, 347-352.
64. De Andraca, I., and Uauy, R. (1995) Breast Feeding for Optimal Mental Development: The Alpha and the Omega in Human Milk, in Behavioral and Metabolic Aspects of Breastfeeding: International Trends in World Review of Nutrition Dietetics 78, (Simopoulos, A.E, Dutra de Oliveira, J.E., and Desai, I.D., eds.) pp. 1-27.
65. Rodgers, B. (1978) Feeding in Infancy and Later Ability and Attainment: A Longitudinal Study, Dev. Med. Child Neurol. 20, 421-426.
66. Rogan, J.W., and Gladen, B.C. (1993) Breast Feeding and Cognitive Development, Early Hum. Dev. 31, 181-93.
67. Temboury, M.C., Otero, A., Polanco, I., and Arribas, E. (1994) Influence of Breast-Feeding on the Infant's Intellectual Development, J. Ped. Gastr. Nutr. 18, 32-36.
68. Lucas, A., Morley, R., and Cole, T.J. (1992) Breastmilk and
Subsequent Intelligence Quotient in Children Born Preterm, Lancet 339, 261-264.
69. Lanting, C.I., Fidler, V., Huisman, M., Touwen, B.C.L., and Boersma, E.R. (1994) Neurological Differences Between 9-Year- Old Children Fed Breast-Milk or Formula-Milk as Babies, Lancet 344, 1319-1322
70. Butte, N.E, Smith, E.O., and Garza, C. (1991) Heart Rate of Breast-Fed and Formula-Fed Infants, J. Pediatr. Gastroenterol. Nutr. 13, 391-396.
71. Bennett, M.J., Gayton, A.R., Rittey, C.D.C., and Hosking, P.G. (1994) Juvenile Neuronal Ceroid-Lipofuscinosis: Developmental Progress After Supplementation with Polyunsaturated Fatty Acids, Dev. Med. Child. Neurol. 36, 630-638.
72. Hoffman, D.R., Uauy, R., and Birch, D.G. (1993) Red Blood Cell Fatty Acid Levels in Patients with Autosomal Dominant Retinitis Pigmentosa, Exp. Eye Res. 57, 359-368.
73. Hoffman, D.R., Uauy, R., and Birch, D.G. (1995) Metabolism of Omega-3 Fatty Acids in Patients with Autosomal Dominant Retinitis Pigmentosa, Exp. Eye Res. 60, 279-289.
74. Hoffman, D.R., and Birch, D.G. (1995) Docosahexaenoic Acid (DHA) Content in Red Blood Cells of Patients with X-Linked Retinitis Pigmentosa, Invest. Opthalmol. Vis. Sci. 36, 1009-1018.
75. Martinez, M. (1994) Polyunsaturated Fatty Acids in the Developing Human Brain, Red Cells and Plasma: Influence of Nutrition and Peroxisomal Disease, World Rev. Nutr. Diet 75, 70-78.
76. Sprecher, H. (1992) Interconversions Between 20- and 22- Carbon n-3 and n-6 Fatty Acids via 4-Desaturase Independent Pathways, in Essential Fatty Acids and Eicosanoids (Sinclair A., and Gibson, R, eds.) American Oil Chemists' Society, Champaign, pp. 18-22.
77. Moore, S.A. (1994) Local Synthesis and Targeting of Essential Fatty Acids at the Cellular Interface between Blood and Brain: A Role for Cerebral Endothelium and Astrocytes in the Accretion of CNS Docosahexaenoic Acid, World Rev. Nutr. Diet 75, 128-133
78. Bennett, M.J., Gwilym, P.H., Gayton, R., Thompson, G., Galloway, J.H., and Cartwright, I.J. (1988) Therapeutic Modification of Membrane Lipid Abnormalities in Juvenile Neuronal Ceroid-Lipofuscinosis (Batten Disease), Am. J. Med. Genet. Suppl. 5, 275-281.
79. Bazan, N.G., Scott, B.L., Reddy, T.S., and Pelias, M.Z. (1986) Decreased Content of Docosahexaenoate and Arachidonate in Plasma Phospholipids in Usher's Syndrome, Biochem. Biophys. Res. Comm. 141,600-604.
80. Converse, C.A., Hammer, H.M., Packard, C.J., and Shepherd, J. (1983) Plasma Lipid Abnormalities in Retinitis Pigmentosa and Related Conditions, Trans. Ophthalmol. Soc. UK 103, 508-512.
81. McLachlan, T., McColl, A.J., Collins, M.E, Converse, C.A., Packard, C.J., and Shepherd, J. (1990) A Longitudinal Study of Plasma n-3 Fatty Acid Levels in a Family with X-linked Retinitis Pigmentosa, Biochem. Soc. Trans. UK 18, 905-906.
82. Anderson, R.E., Maude, M.B., Lewis, R.A., Newsome, D.A., and Fishman, G.A. (1987) Abnormal Plasma Levels of Polyunsaturated Fatty Acid in Autosomal Dominant Retinitis Pigmentosa, Exp. Eye. Res. 44, 155-158.
83. Gong, J., Rosner, B., Rees, D.G., Berson, E.L., Weigel-DiFranco, C.A., and Schaefer, E.J. (1992) Plasma Docosahexaenoic Acid Levels in Various Genetic Forms of Retinitis Pigmentosa, Invest. Ophthalmol. Vis. Sci. 33, 2596-2602.
84. Berson, E.L., Rosner, B., Sandberg, M.A., and Dryja, T.E (1991) Occular Findings in Patients with Autosomal Dominant Retinitis Pigmentosa and a Rhodopsin Gene Defect (Pro-23-His), Arch. Ophthalmol. 109, 92-101.
Lipids, Vol. 31, Supplement (1996)
Copyright © 2022 FDOKUMEN




















