
On epicardial potential reconstruction using regularization schemes with the L1-norm data term

lable at ScienceDirect
Progress in Biophysics and Molecular Biology 115 (2014) 173e185
Contents lists avai
Progress in Biophysics and Molecular Biology
journal homepage: www.elsevier .com/locate/pbiomolbio
Review
Role of endo-epicardial dissociation of electrical activity andtransmural conduction in the development of persistent atrialfibrillation
Sander Verheule, Jens Eckstein, Dominik Linz, Bart Maesen, Elham Bidar, Ali Gharaviri,Ulrich Schotten*
Departments of Physiology and Cardiothoracic Surgery, Faculty of Health, Medicine and Life Sciences, Cardiovascular Research Institute Maastricht,Maastricht, The Netherlands
a r t i c l e i n f o
Article history:Available online 30 July 2014
Keywords:Atrial fibrillationFibrosisStructural remodelingDissociationComplexity
* Corresponding author. Department of Physiology,E-mail address: [email protected]
http://dx.doi.org/10.1016/j.pbiomolbio.2014.07.0070079-6107/© 2014 Published by Elsevier Ltd.
a b s t r a c t
Atrial fibrillation (AF) is an arrhythmia that occurs as a result of numerous pathophysiological processesin the atria. High rate, neurohumoral activation, aging and chronic stretch activate a variety of signalingpathways leading to electrical and structural remodeling. In particular, endomysial fibrosis within theepicardial layer, which also occurs as a result of AF itself, can disrupt electrical connections betweenmuscle bundles. This leads to electrical dissociation not only within the epicardial layer, but also betweenthe epicardial layer and the endocardial bundle network.
Although the normal, healthy atrium has a complex 3-dimensional shape, differences in activationtime between the epicardial layer and the underlying trabecular network in the atrial free walls aresmall, and the atrial walls essentially function as a 2-dimensional surface for propagating fibrillationwaves. However, progressive structural remodeling leads to increased dissociation of epicardial andendocardial activation patterns. Epi-endocardial dissociation allows fibrillation waves to propagate be-tween epicardium and endocardium, and become visible as ‘breakthrough waves’ that add to the overallcomplexity of fibrillatory conduction and thus to AF stability. This process greatly increases the effectivesurface area available to fibrillation waves and causes the atrial walls to behave as a 3-dimensionalsubstrate. Computer models support that this behavior can increase AF stability.
Under these conditions, ectopic activity originating from e.g. the pulmonary veins is likely to triggerlonger episodes of AF. Experiments using simultaneous endo-epicardial mapping of AF suggest thatdisseminated, irregular and non-repetitive ectopic focal discharges might also occur during AF. Theincreasingly 3-dimensional character of AF as a result of structural remodeling lowers the responsivenessto antiarrhythmic compounds and ablation therapy, thus advocating early rhythm control strategies.
© 2014 Published by Elsevier Ltd.
Contents
1. Epicardial mapping: information gleaned from the atrial surface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1741.1. Development of a substrate for AF . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1741.2. Atrial anatomy and structural remodeling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1741.3. Breakthrough waves as indicators for transmural conduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181
2. Direct assessment of endo-epicardial dissociation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1812.1. Animal models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1812.2. AF patients . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183
3. Computer models contribute to understanding the nature of endo-epicardial dissociation of electrical activity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1834. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
Universiteitssingel 50, P.O. Box 616, 6200 MD, Maastricht, The Netherlands. Tel.: þ31 433881076; fax: þ31 433884166.(U. Schotten).

S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185174
Funding . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184Editors’ note . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
Fig. 1. Anatomy of the atrial free walls. A) Endocardial, back-lit view of an explantedatria. The atria were opened up and stretched to expose the relation between theendocardial trabecular network and the thin epicardial layer. RAA and LAA: RA and LAappendages, CT: crista terminalis, SVC: superior vena cava, IAS: interatrial septum. B)Transverse section of the RA and LA walls (Toluidine Blue staining), showing the thinepicardial layer and underlying trabeculae. C) Epicardial and endocardial aspect of thepart the left atrial free wall (magnified view of the preparation in panel A, with colorsinverted to increase clarity). The endocardial trabeculae branch out into smallerepicardial bundles that are oriented at an angle to those trabeculae. Adapted from ref(Verheule et al., 2013).
1. Epicardial mapping: information gleaned from the atrialsurface
1.1. Development of a substrate for AF
Atrial fibrillation (AF) is the most common sustained arrhythmiain humans and its prevalence still increases as the population ages.Its impact on health care is substantial, as AF is associated with anincreased morbidity, in particular a 5-fold increase in risk of stroke,and a doubling of death rate (European Heart RhythmA et al., 2010).In the majority of patients, the arrhythmia progresses from short‘AF runs’ to more frequent and longer episodes of AF (paroxysmalAF), eventually degenerating into sustained forms of AF (persistentand longstanding persistent AF) (Schotten et al., 2011).
Paroxysms of AF are initiated by ectopic activity or rapid reen-trant circuits, often originating from muscular sleeves of the pul-monary veins (Haissaguerre et al., 1998). However, there is still adebate regarding electrophysiological mechanisms sustainingpersistent AF, centering on the question of whether AF at that stageis ‘hierarchical’ or ‘anarchical’. In a hierarchical interpretation, someparts of the atria may harbor 'AF drivers' that are essential formaintaining AF. Spiral reentry waves (rotors) and ectopic foci havebeen proposed as ‘AF drivers’ in canine, sheep andhumans (Narayan et al., 2012; Yamazaki et al., 2012; Lee et al., 2013;Skanes et al., 1998). Indeed, ablation strategies targeting culpritareas can terminate AF (Haissaguerre et al., 1998; Narayan et al.,2013). Alternatively, the nature of persistent AF may be moreanarchical, with multiple fibrillation waves propagating chaoticallywithin the atria, without clear culprit areas responsible for sus-taining AF (Moe and Abildskov, 1959; Allessie et al., 1985; Kumagaiet al., 1997). These mechanisms may transiently be present in anindividual patient. The notion of a local source for AF does notexclude an anarchical mechanism at anothermoment in time in thesame patient and vice versa.
The progressive, self-perpetuating nature of AF can be explainedby the fact that AF causes changes in the atria that increase thestability of the arrhythmia. These changes include a shortening ofthe action potential duration and effective refractory period (elec-trical remodeling) and alterations in tissue structure (structuralremodeling). The latter process includes altered connexin expres-sion, (Duffy and Wit, 2008) myocyte hypertrophy (Spach et al.,2004) and atrial fibrosis (Spach and Boineau, 1997). Whereaselectrical remodeling takes place rapidly, within 1e2 days, struc-tural remodeling progresses gradually over a time course ofmonths (Ausma et al., 2001, 2003). The stability of AF (measured asthe duration of AF episodes or as the amenability to pharmaco-logical cardioversion) increases over a comparable slow timecourse of months (Eijsbouts et al., 2006; Verheule et al., 2010).
Using high-density epicardial contact mapping of AF, our grouphas shown that progression of structural remodeling and AF stabilitygo hand in hand with an gradual increase in complexity of the AFconduction pattern, both in experimental and clinicalstudies (Verheule et al., 2010; de Groot et al., 2010; Allessie et al.,2010; Verheule et al., 2013; Eckstein et al., 2011, 2013; Maesenet al., 2013). With the duration of the arrhythmia, the number ofsimultaneous fibrillation waves and the occurrence of conductionblock increase, leading to a more dissociated pattern of fibrillatoryconduction (Verheule et al., 2010; Allessie et al., 2010). In these
studies, rotors were rare and unstable in the atrial free walls (deGroot et al., 2010; Allessie et al., 2010; Eckstein et al., 2013;Konings et al., 1994). The widespread process of structural remod-eling led to an increased complexity of fibrillatory conductionthroughout the atria. As a result, the contribution of the free walls tothe stability of AF increases, irrespective of the possible presence ofrotors or other types of drivers in other areas, representing a shiftfrom a hierarchical towards a more anarchical type of AF.
1.2. Atrial anatomy and structural remodeling
The anatomy of atria is highly complex, with a large variability inlocal fiber architecture (Ho et al., 2002). Anatomical boundariessuch as the atrioventricular valves and venous ostia may act asanchors for reentry. Within the atria, there are regions where thefiber orientation is highly organized and uniform, such as the ter-minal crest, the area in between the pulmonary veins and

Fig. 2. Relation between tissue architecture and epicardial fibrillatory conduction in goats. A) Rose diagrams of angle differences between endocardial and epicardial bundles foracute and persistent AF. In persistent AF a significantly larger part of the angle differences is close to 90� (perpendicular to each other). Mean ¼ mean angle difference. B) Schematicrepresentation of endocardial (thick black lines) and epicardial bundles (thin blue lines) in acute AF (left) and persistent AF (right). In acute AF, good electrical connections betweenboth layers cause epicardial fibrillation waves to follow the thick endocardial bundles. In persistent AF, not only are endo- and epicardial bundles oriented more perpendicular toeach other, but endo-epicardial connections are lost due to endomysial fibrosis. As a consequence, epicardial fibrillation waves follow the thin epicardial bundles. C) Examples ofactivation maps illustrating the scheme in panel B. Left panels: epicardial ‘waves’ maps, in which each fibrillation wave has been assigned a different shade of gray. Right panels:tissue architecture for both maps, black regions represent the endocardial trabeculae, white arrows indicate the propagation of fibrillation waves. Adapted from ref (Maesen et al.,2013).
S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185 175
Bachmann's bundle, the major conduction pathway between RAand LA (Fig.1A) (Ho and Sanchez-Quintana, 2009). This architectureaffects local conduction and may thereby affect the role of theseregions in arrhythmogenesis. For example, Bachmann's bundle in
goats is characterized by a higher degree of fractionation comparedto other parts of the atrium. This is potentially a result of dispersionof refractoriness between Bachmann's bundle and the surroundingtissue, or of local reentry within Bachmann's bundle (Shan et al.,

Fig. 3. Development of endomysial fibrosis in goats. A) After 3 weeks of AF, myocytes within bundles are still tightly packed. After 6 months of AF, endomysial fibrosis has lead to anincrease transverse separation of myocytes within bundles. B) Intermyocyte distances in the LA free wall as a function of the distance to the epicardial side of the muscle layer. After6 months of AF, intermyocyte distances have increased markedly within the outer 1 mm of the atrial wall. C) Transmural view of a relatively thick part of the LA free wall, illustratingthe larger intermyocyte distances on the epicardial side. D) Optical mapping of epicardial wavefront expansion in 2 � 2 cm areas of the LA free walls, showing snapshots of theoptical action potential 5 ms after stimulation within the field of view in goats after 3 weeks (left panels) and 6 months of AF (right panels). Adapted from ref (Verheule et al., 2013).
S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185176
2004). In the muscular sleeves of the pulmonary veins, abruptchanges in fiber orientation correlatewith zones of activation delay,thereby facilitating (micro)-reentry and promotion of fibrillatoryconduction from a focal source (Hocini et al., 2002).
A number of other studies have correlated propagation patternsto the underlying fiber architecture. In experimental studies in theright atrium, the discordance between epicardial and endocardialpropagation was more pronounced in the thick trabeculated
parts (Eckstein et al., 2011; Schuessler et al., 1993). Furthermore, thedirection of pectinate muscle bundles corresponded to lines ofblocks (Gray et al., 1996; Wu et al., 1998; Eijsbouts et al., 2004). Inperfused sheep RA preparations, fast pacing led to breakdown of1:1 conduction at branching points of endocardial bundles andchaotic propagation in the trabecular network (Berenfeld et al.,2002). Simultaneous endo-epicardial optical mapping inLangendorff-perfused sheep LA identified 3-dimensional scroll

Fig. 4. Increasing complexity of fibrillatory conduction during the development of an AF substrate. A) Two successive maps recorded during AF in the LA of goats after 3 weeks(upper row) and 6 months of AF (lower row). These time points correspond to stages in the disease process with complete electrical remodeling without and with significantstructural remodeling. After 3 weeks, AF is persistent but can still be cardioverted pharmacologically. After 6 months of AF, the efficacy of all anti-arrhythmic drugs that have beentested has been lost completely. The wave maps of fibrillatory conduction show separate fibrillation waves in different shades of blue. Isochrones indicate conduction within thearea of a particular wave. Epicardial breakthroughs are indicated with asterisks. After 6 months of AF, the fibrillation pattern was more complex, with a higher number of smallerfibrillation waves and an increased incidence of epicardial breakthroughs. B) Distribution of epicardial breakthroughs, the size of the stars indicates the number of breakthroughsobserved at a particular electrode. In both goats and patients, the frequency of epicardial breakthroughs was higher in chronic AF, but the distribution was also less clustered.Adapted from refs (de Groot et al., 2010; Verheule et al., 2013).
S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185 177
waves that form and meander around regions of sharp transition inmyocardial thickness (Yamazaki et al., 2012). In the human rightatrial free wall, lines of block were predominantly found along thelong axis of the crista terminalis and large pectinatemuscles (Allessie et al., 2010; Wu et al., 2002). In human left atria,lines of conduction delay ran vertically between the pulmonary
veins, parallel to the fiber orientation (Roberts-Thomson et al.,2008; Markides et al., 2003). The atrial free walls (including ap-pendages) consist of a trabeculated endocardial bundle networkbranching out into a thin epicardial layer (Fig.1). The architecture ofthe large trabeculated regions of the atria is complex, with pecti-nate muscle bundles oriented obliquely to epicardial fibers

Fig. 5. Simultaneous epi- and endocardial mapping in goats. A) Clamp-like mapping tool with a lower and upper jaw used to record endocardial and epicardial electrograms,respectively. B) Endo-epicardial activation maps depicting a dissociated epicardial wave (left) and a non-dissociated epicardial wave (right). C) Endo-epicardial activation timedifferences at exactly opposed sites of the atrial wall in three groups of goats with increasing duration of AF. Curve fitting revealed a two-component Gaussian function with the firstcomponent representing synchronous activation (narrow distribution, small standard deviation) and the second component representing dissynchrony (broad distribution, largestandard deviation). The contribution of the second component increased with AF duration. D) Schematic representation of the transmural activation pattern in case of transmuralconduction (left panel) and in case of an ectopic focal discharge (right panel). E) Endo-epicardial activation maps illustrating different types of breakthrough events (marked withthe white star). The corresponding unipolar electrograms at the site of breakthrough at the opposing site are shown below. The red lines in the electrograms mark the moment ofbreakthrough. The left pair of maps depicts an endocardial activation preceding the epicardial breakthrough. The middle pair shows synchronous endo-epicardial breakthroughs,suggesting an ectopic focal discharge as the source. The right pair depicts a breakthrough as the earliest activation in the mapped region that cannot be explained by transmuralactivation. Most likely it represents a focal ectopic discharge that blocked intramurally. Adapted from refs (Eckstein et al., 2011; Eckstein et al., 2013).
S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185178
(Fig. 1C) (Verheule et al., 2013; Schuessler et al., 1993; Markideset al., 2003; Ho et al., 1999). The thin epicardial layer, which isonly 1e4 mm thick, is highly anisotropic on a microscopic scale.However, because of the variability in fiber orientation, the
propagation during slow rhythms is quite isotropic on a macro-scopic scale (i.e. the conduction velocity is insensitive to the di-rection of propagation) (Hansson et al., 1998). Recently, we havecorrelated fibrillation patterns with epicardial and endocardial
S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185 179
bundle orientation using high-resolution magnetic resonanceimaging (Maesen et al., 2013). In the goat left atrium, epicardialfibers become oriented more perpendicularly to endocardial fibersduring persistent AF (Fig. 2A). Moreover, in normal, healthy atriaduring acute AF, the propagation of epicardial fibrillation wavescorresponds mainly with the orientation of thick endocardialbundles, whereas after 7 months of AF, epicardial propagationfollow the orientation of epicardial muscle bundles (Fig. 2B and C).We have demonstrated that transition of persistent to permanentAF in goats was histologically characterized by an increase inendomysial fibrosis (i.e. increased transverse separation of myo-cytes within bundles), particularly in the outer millimeter of the
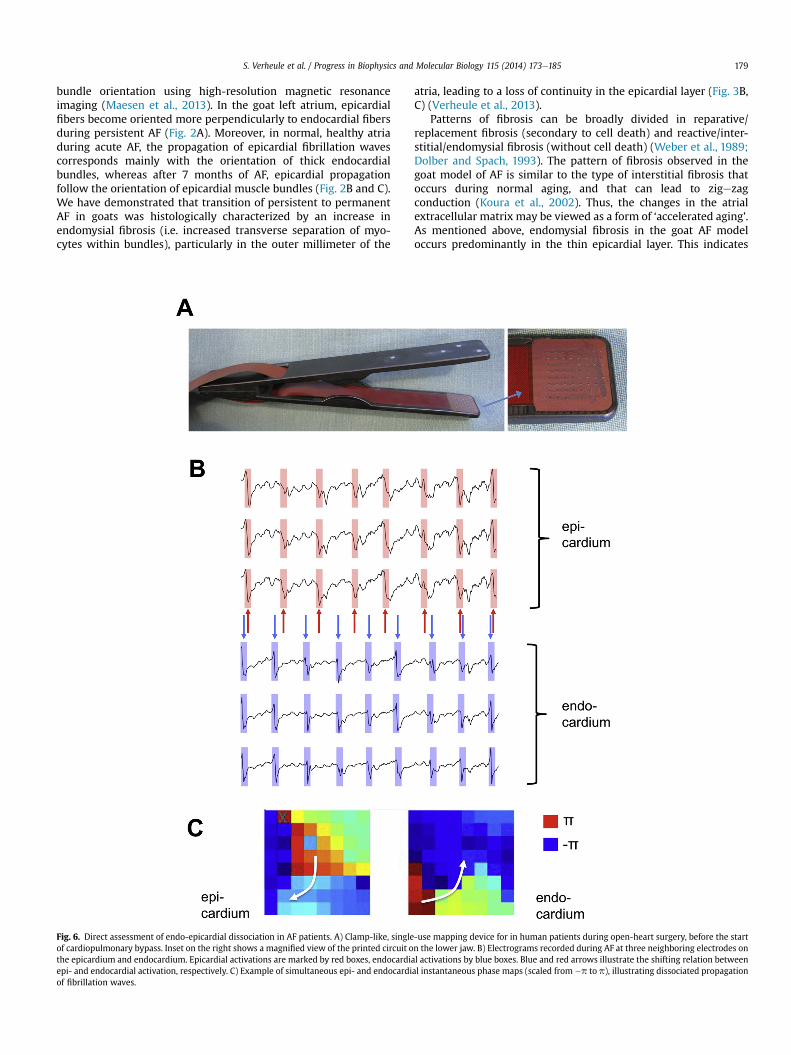
Fig. 6. Direct assessment of endo-epicardial dissociation in AF patients. A) Clamp-like, singleof cardiopulmonary bypass. Inset on the right shows a magnified view of the printed circuit othe epicardium and endocardium. Epicardial activations are marked by red boxes, endocardiaepi- and endocardial activation, respectively. C) Example of simultaneous epi- and endocardiof fibrillation waves.
atria, leading to a loss of continuity in the epicardial layer (Fig. 3B,C) (Verheule et al., 2013).
Patterns of fibrosis can be broadly divided in reparative/replacement fibrosis (secondary to cell death) and reactive/inter-stitial/endomysial fibrosis (without cell death) (Weber et al., 1989;Dolber and Spach, 1993). The pattern of fibrosis observed in thegoat model of AF is similar to the type of interstitial fibrosis thatoccurs during normal aging, and that can lead to zigezagconduction (Koura et al., 2002). Thus, the changes in the atrialextracellular matrix may be viewed as a form of ‘accelerated aging’.As mentioned above, endomysial fibrosis in the goat AF modeloccurs predominantly in the thin epicardial layer. This indicates
-use mapping device for in human patients during open-heart surgery, before the startn the lower jaw. B) Electrograms recorded during AF at three neighboring electrodes onl activations by blue boxes. Blue and red arrows illustrate the shifting relation betweenal instantaneous phase maps (scaled from �p to p), illustrating dissociated propagation

S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185180
that endomysial fibrosis is caused by a trigger (e.g. stretch/dilata-tion) that affects the thin epicardial layer more than the underlyingnetwork of trabeculae). By contrast, myocyte hypertrophy (whichmay also affect propagation ((Spach et al., 2004)) was homoge-neously distributed throughout the atrial wall from epicardium toendocardium (Verheule et al., 2013). This may indicate that myo-cyte hypertrophy is caused by a different trigger (e.g. neurohumoralsignaling) than endomysial fibrosis.
The increased transverse separation of myocytes is likely to leadto a loss of transverse electrical connections. Unfortunately, dif-ferences in the distribution of gap junction channel proteins(connexin, Cx) between the epi- and endocardium have not beenstudied in detail. In another model with increased vulnerability to
Fig. 7. Mathematical model of transmural conduction in the atrium. A) Model structure wElectrical activity is dissociated between the layers, leading to transmural propagation thatMembrane potential of a simulation at t ¼ 1 s (the time at which 6 connections were introdunumber of connections was changed. b) Continued simulation after removing all 6 connectiwaves and phase singularities (PS) in one layer for single-layer and double-layer simulatiodouble-layer simulations. Adapted from ref (Gharaviri et al., 2012).
AF, dogs with sterile pericarditis, expression of the gap junctionproteins Cx40 and 43 is strongly reduced in the epicardium, butrelatively normal in the endocardium (Ryu et al., 2007). In goats,the expression of Cx43 was not affected by 4 months of AF, but thedistribution of Cx40 became more heterogeneous, with small(<1 mm) areas showing low Cx40 expression (Ausma et al., 2003;van der Velden et al., 2000). The transmural distribution of theseareas is unknown. In addition, the impact of a selective reduction inCx40 is unclear. Cx40 and Cx43 are likely to form heteromericchannels in the atria (Elenes et al., 1999). Although genetic deletionof Cx40 in mice decreases atrial conduction velocity, (Verheuleet al., 1999) conduction velocity in the human right atrium wasinversely related to the amount of Cx40 labeling (Kanagaratnam
ith two layers (epi- and endocardium) with transmural connections (white arrows).is visible as a breakthrough wave in the upper layer, indicated by a black arrow. B) a)ced while only a spiral wave existed in upper layer) and at t ¼ 7 s (the time at which theons. c) Continued simulation without changing the connections. C) Average number ofns. D) KaplaneMeier curve showing the percentage of AF persistence for single- and

S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185 181
et al., 2002). In cultured strands of atrial myocytes, the Cx40/Cx43ratio was an important determinant of propagation, with Cx43increasing and Cx40 decreasing conduction velocity (Beauchampet al., 2006).
Endomysial fibrosis selectively hampers transverse propagation,largely without affecting longitudinal propagation (Spach andBoineau, 1997). On a microscopic scale, the structural alterationsin the epicardium of goats after 6 months of AF are indeed associ-ated with an increased anisotropy in epicardial wavefront expan-sion, consistent with a loss of transverse electrical connectionsbetween fibers (Fig. 3D). We propose that the thin epicardial layerhas a synchronizing effect during AF in healthy atria, i.e. it increasesthe synchroneity of fibrillatory activation between the epicardiumand the endocardial bundle network, as illustrated in Fig. 8 (upperhalf). Structural remodeling causes the epicardial layer to lose thissynchronizing effect, as schematically represented in Fig. 8 (lowerhalf). In this view, healthy atria walls behave as an essentially 2-dimensional sheet, where epicardial activation is synchronouswith the activation of the underlying trabeculae. Structuralremodeling causes the atrial walls to behave as a more 3-dimensional substrate, where synchroneity between endo- andepicardium is lost and propagation from the endocardium andepicardium (and vice versa) becomes much more frequent. Thisalteration in the character of fibrillation is evidenced by theincreased incidence of 'breakthrough waves' and increased disso-ciation between the epicardial layer and the underlying trabecularnetwork, as described in detail in the sections below.
1.3. Breakthrough waves as indicators for transmural conduction
With progressive structural remodeling caused by sustained AF,fibrillation patterns become increasingly complex (Fig. 4A) (Verheuleet al., 2010; Allessie et al., 2010). In addition to a decrease in the sizeof wavefronts and a concomitant increase in the number of waves, astriking feature is the increased incidence of breakthrough waves(Fig. 4B). Thesewaves appear suddenlywithin themapped epicardialarea and cannot be explained by propagation within the epicardialplane. Breakthrough waves add to the overall complexity of fibril-lation patterns, and thus, we propose, to AF stability. To detect theseevents in high-density epicardial mapping experiments, severalcriteria were used: the breakthrough site had to be activated earlierthan all surrounding electrodes, had to be located away from theperiphery of the mapped area, and should not occur at the border ofanother wavewithin a certain time delay, to distinguish these eventsfrom very slow or discontinuous epicardial conduction. In patients,the incidence of breakthroughs in the right atriumwas almost 4-foldhigher during persistent AF than during acute AF and the origin ofthese waves was non-repetitive in time and widespread throughoutthe atria (Fig. 4B, right) (Verheule et al., 2010; de Groot et al., 2010;Verheule et al., 2013). In a goat model with AF induced by rapidatrial pacing, the incidence of breakthroughs increased with AFduration from approximately 2% of all fibrillation waves in acute AFto approximately 14% of all fibrillation waves after 6 months of AF.The sites of origin of breakthrough waves were again widespread,and the degree of clustering of these sites was even lower after 6months of AF than after 3 weeks of AF (Fig. 4B, left) (de Groot et al.,2010; Verheule et al., 2013). In principle, these events may beexplained both by ectopic focal discharges, i.e. a wave originatingvery close to the recording electrode, or by transmural conduction,i.e. conduction of a fibrillation wave from the opposing layer of theatrial wall, in this case the endocardial trabecular network. Todistinguish between these two possible mechanisms, additionalcriteria were considered. Firstly, in the case of an ectopic focaldischarge, a unipolar electrogram recorded at the site of ectopyshould have an S-wave morphology, whereas in the case of endo-
epicardial transmural conduction, it is expected to exhibit a smallR-wave. Secondly, the degree of prematurity of breakthroughs wasdetermined by comparing their coupling interval with the averageAF cycle length (AFCL) at the site of origin. Focal activity should showprematurity, whereas in the case of endo-epicardial transmuralconduction, coupling interval should not differ from AFCL. Thirdly, toidentify a rapidly firing focus, the repetitivity of fibrillation waveswith a focal spread of activation was measured.
Interestingly, in humans as well as in goats the properties offibrillation waves did not support the existence of high-frequencyectopic focal discharges. Unipolar electrograms recorded at theepicardial sites of origin of these waves showed small but clear Rwaves (de Groot et al., 2010; Houben et al., 2004). In addition, themedian coupling interval of these waves was only 11 ms shorterthan expected (and in one third of the cases actually longer than theaverage AFCL) and were not preceded by a long interval (de Grootet al., 2010). Finally, a large majority (90%) were not repetitive butappeared as solitary events, with a non-clustered, widespreaddistribution (de Groot et al., 2010; Verheule et al., 2013). Thesefeatures of breakthrough waves suggest endo-epicardial conduc-tion, rather than focal activity as a predominant mechanism. Thisindicates that the increased incidence of breakthrough waves withprogressive structural remodeling indeed reflects an increasingly 3-dimensional character of AF, as schematically represented in Fig. 8.It should be noted, however, that experiments in dogs undergoingvagal nerve stimulation demonstrated that a large percentage ofwaves could be traced back to sites with radial spread of activationwith pure S-wave morphology at the site of earliest activation. Thissuggests that under these conditions ectopic focal discharges mayvery frequently occur and might even be the main mechanism forAF (Lee et al., 2013).
2. Direct assessment of endo-epicardial dissociation
2.1. Animal models
A fibrillation wave can only propagate transmurally, from theendocardium to the epicardium or vice versa, if the opposing layeris excitable, meaning that one layer is activated while the opposingside of the atrial wall is not. By implication therefore, one of theprerequisites for endo-epicardial conduction is endo-epicardialdissociation (EED). Endo-epicardial dissociation in the atrium wasfirst described in an ex-vivo model by Schuessler et al. (1993). Theyperformed simultaneous endo-epicardial mapping in isolatedcanine right atria and found significant activation time differencesbetween the epicardial and the endocardial surface during acetyl-choline induced acute atrial fibrillation. Yamazaki et al. establishedan ex vivo model of simultaneous endo-epicardial optical mappingin a Langendorff perfused sheep heart (Yamazaki et al., 2012). Theyfocused on scroll waves and pointed out the importance of theinhomogeneous atrial wall thickness. The presence of a transmuralscroll wave implies nearly synchronous endo-epicardial activation.Their data could therefore be interpreted in a way that there is lessEED in thinner parts of the atrial wall, thus serving as anchor forscroll waves. The correlation between local atrial wall thickness anddegree of EED has also been demonstrated in vivo endo-epicardialdirect contact mapping experiments (Eckstein et al., 2011).
Derakhchan et al. were the first to describe a method for simul-taneous endo- and epicardial contact mapping in vivo, showingpreferential pathways of atrial activation and dissociation of thosepathways during reentry (Derakhchan et al., 2001). Everett et al. havecompared endo- and epicardial activation in three different caninemodels of AF (heart failure, mitral regurgitation and rapid atrialpacing) using non-contact mapping endocardially and direct contactmapping epicardially (Everett et al., 2010). The models with strongly

Fig. 8. Schematic representation of the effect of structural remodeling on fibrillatory conduction. Normal atria consist of a thin, continuous epicardial layer that is well-connected tothe underlying endocardial network of trabeculae. An epicardial wavefront will rapidly breakthrough to the trabecular network, leading to synchronous endo- and epicardial waves.The thin epicardial layer thus has a synchronizing effect on fibrillation waves propagating in the epicardium and the underlying endocardial bundles. Under these conditions, theatria wall essentially behaves as a 2-dimensional substrate. In structurally remodeled atria, epicardial endomysial fibrosis reduces the continuity of the epicardial layer andconnectivity with the trabecular network. Spread of an epicardial wavefront is hampered. After eventually breaking through to the trabecular network, endocardial wavefrontspropagate more rapidly, leading to additional breakthroughs to the epicardium at distant sites. As a result, the paths of fibrillation waves become more tortuous and the effectivesurface available for propagation greatly increases. Note that conduction may be fairly normal at slow rates, and will show dramatic changes mainly with short coupling intervals(i.e. premature stimuli and fibrillation).
S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185182
increased fibrosis (mitral regurgitation and heart failure) had smalleractivation differences between the endo- and the epicardial surfaceand more repetitive activation patterns compared to the rapid atrialpacing model. AF was persistent in the rapid atrial pacing group only
and multiple wavelets and large endo-epicardial activation differ-ences were predominantly seen in this group.
To directly assess the origin of breakthroughs and degree of epi-endocardial dissociation in vivo, we developed a technique for

S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185 183
simultaneous endo-epicardial high-resolution direct contactmapping (Eckstein et al., 2008). We used a custom-made clamp-like mapping tool (Fig. 5A) with two multi-electrode arrays andinserted one array into the beating atrium through a small incisionnear the left atrial appendage. Unipolar electrograms were recor-ded during sinus rhythm, acute AF, after 3 weeks and 6 months AFand activation times of corresponding endocardial and epicardialelectrodes were analyzed. For the quantification of EED we used amethod comparable to that of Schuessler et al. (1993) (quantifica-tion of corresponding activation time differences) and a second,wave-based analysis allowing quantification of the surface that isactivated synchronously or dyssynchronously (Fig. 5B) (Ecksteinet al., 2013). We demonstrated that while there is almost no EEDduring sinus rhythm and little during acute AF, it had increasedmarkedly after 3 weeks of AF and further after 6 months of AF(Fig. 5C) (Eckstein et al., 2011). Mechanistically, the increase of EEDrepresents an increase of functional surface for fibrillation waves tocoexist and thereby contribute to stabilization of AF.
The increase of EED correlated very well with the incidence ofbreakthroughs, thus strengthening the argument that break-throughs are indicative of the 3-dimensional nature of propagationduring AF (Fig. 8). To further assess whether these activation pat-terns derive from truly focal, ectopic activity or whether theyrepresent transmural conduction from the opposing layer of theatrial wall, we analyzed breakthrough patterns and included theactivation times of the opposing site of the atrial wall.
If transmural conduction would be extremely slow, then anybreakthrough wave may be explained as propagation of an earlierwave on the opposite layer. However, although the conduction ve-locity during transmural propagation is not known exactly, there isno reason to assume that it would be very different from epicardialand endocardial conduction velocities. We tested the probabilitythat, assuming a normal physiological conduction velocity, thesebreakthroughs could derive from transmural activation(Fig. 5D) (Eckstein et al., 2013). We concluded that 86% of allbreakthrough events could be explained by transmural conductionwhereas only 13% were likely to originate from ectopic focal dis-charges, resulting in breakthrough pattern on both opposing sides ofthe wall (Fig. 5E). Thus, the quantitative contribution of focal acti-vations to observed activation patterns might be modest. However,their contribution to the maintenance of AF may still be important.The idea that ectopic focal discharges can contribute to AF perpet-uation is supported by the fact that already a single site with highpropensity to discharges (topical aconitine exposure) can largelyincrease AF stability (Scherf, 1947). By definition, each focal activa-tion reflects the generation of a new fibrillation wave. These newwaves, whether triggered by a previous action potential or not, maybe important both for the initiation (Burashnikov and Antzelevitch,2003) or maintenance of AF. In the ventricles, a regional synchro-nization of afterdepolarizations can lead to a diffuse, irregularpattern of focal activations (Sato et al., 2009). A comparable diffuse,irregular pattern of ectopy in the atria is consistent with theobserved distribution of breakthroughs. Their inhibition by the anti-arrhythmic drug flecainide (Hilliard et al., 2010) may in part beresponsible for the efficacy of this drug (Eijsbouts et al., 2006).
2.2. AF patients
Because there are no published quantitative data on endo-epicardial dissociation in humans, we cannot comment on therelative contributions of transmural conduction and focal activity inAF patients. However, as mentioned above, the incidence, distri-bution and other characteristics of epicardial breakthrough wavesare similar to those described in the goat model of AF. Thus,epicardial breakthrough may in a similar way reflect the
increasingly 3-dimensional character of fibrillatory propagationduring the progression of AF.
To directly assess endo-epicardial conduction and dissociationin humans, we have developed a mapping device suitable forclinical application during open-heart surgery. This clamp-like tool(Fig. 6A) has two arms ending in a square plaque (1.5�1.5 cm) of 64(8 � 8) unipolar electrodes that directly oppose each other whenthe device is closed. Using this device, we are conducting endo-epicardial mapping of the right atrial wall in patients with persis-tent AF. During surgery, prior to the insertion of the venous cannulafor the cardiopulmonary bypass in the right atrial appendage, theendocardial arm of the tongs-electrode was introduced into thepurse-string suture that was placed for the venous cannula. Bydesign, the electrode arrays were parallel to each with a 3 mmdistance between the arrays. However, the surgeon was able toadjust the distance guided by the quality and amplitude of theelectrograms. In the beating heart, a 30-second endo-epicardialrecording was made before the electrode was extracted and thevenous cannula was inserted for cardiopulmonary bypass. Pre-liminary results show endo-epicardial dissociation (Fig. 6B and C)that varies in degree with time, showing patterns similar to thoseobtained in goat atria.
3. Computer models contribute to understanding the natureof endo-epicardial dissociation of electrical activity
In an abstract sense, a piece of atrial muscle may be representedas a flat sheet. In such a homogeneous medium, sustained reentry ismostly based on stable rotors. When heterogeneity in for exampleaction potential duration or structural obstacles is introduced, rotorstend to become less stable or break up, leading to a more complexpattern. Some studies have used geometric models to represent partof the atrial anatomy, such as an orifice of a pulmonary vein. Anumber of studies have presented 3-dimensional models of theentire atria. The simplest models consist of a 2-dimensional sheet orfolded sheet into shape of the atrium including two main cavities,atrial appendages, and a septal ring (Haissaguerre et al., 2007;Ruchat et al., 2007; Uldry et al., 2012). More complex modelsinclude greater detail, and have taken variations in wall thickness orlocal fiber architecture into account (Vigmond et al., 2001; Aslanidiet al., 2011; Krueger et al., 2011; Zhao et al., 2012; Colman et al.,2013; Kuijpers et al., 2011). However, none of the modelsmentioned above show breakthrough waves that are such a strikingfeature of the fibrillation pattern in animal models and AFpatients (Verheule et al., 2010; de Groot et al., 2010; Eckstein et al.,2013; Konings et al., 1994). A sole exception in this respect is a 2-dimensional model of a transmural slice of the posterior LA, basedon which breakthroughs were proposed to result from intramuralrotors (Tanaka et al., 2007). To further study the characteristics of 3-dimensional/transmural conduction, we have recently developed asimple dual-layer model to help to elucidate the relation betweenthe number of connections between two atrial layers, endo-epicardial dissociation and breakthrough waves (Fig. 7A) (Gharaviriet al., 2012). Compared to a single layer model, the dual-layermodel showed a higher number of coexisting waves and a higherAF stability (Fig. 7BeD). Moreover, the fibrillation patterns in bothlayers of this symmetrical model included epicardial breakthroughwaves. This model was also used to simulate the effect of the patternof endomysial fibrosis that has been observed in goats after 6monthsof AF, represented as a loss of transverse connections betweenmyocytes in the epicardial layer (Verheule et al., 2013). Under theseconditions, both the number of simultaneous waves and the inci-dence of breakthrough waves on the epicardial side increased, par-alleling the experimentally observed changes in conduction pattern.A logical next step would be to implement transmural conduction

S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185184
between an epicardial layer and an underlying trabecular network ina more realistic atrial anatomy in a way that allows EED and break-through waves to occur.
4. Conclusion
This review demonstrates recent experimental evidence forthe existence of the 3-dimensional character of atrial fibrillation,characterized by dissociation of electrical activity not only withinthe epicardial layer but also between the epicardial layer and theendocardial bundle network. Endomysial fibrosis preferentiallyoccurring in the epicardial layer, rearrangement of atrial bundleanatomy and loss of electrical coupling between the epicardiallayer and the endocardial bundle network appear to be the mainstructural mechanisms for the 3-dimensional fibrillation patterns.In view of the recently renewed interest in the electrophysiolog-ical mechanisms of AF, these mechanisms might be of interest asthey add to our understanding of the wide variety of conductionpatterns present during AF. The challenge of current research inunraveling the mechanisms of AF is to quantitatively describe thetransition from organized, at times localized sources of AF to morecomplex patterns of atrial activation occurring at later stagesduring the natural course of the arrhythmia. While organized andlocalized conduction pattern might well be suited as targets for AFablation, disorganized activity without preferential activation ofsites with critical importance for AF maintenance may preventsuccess of AF ablation.
Funding
Supported by grants from the Dutch Research Organization(NWO, VIDI-grant 016.086.379) to US, Foundation Leducq(07CVD03) to US, Centre of Molecular Medicine (COHFAR) to USand European Union (FP7 Collaborative project EUTRAF, 261057) toSV and US.
Editors’ note
Please see also related communications in this issue by Rouillardet al. (2014) and Yu et al. (2014).
References
Allessie, M.A., Lammers, W.J.E.P., Bonke, F.I.M., Hollen, J., 1985. Experimental eval-uation of Moe's multiple wavelet hypothesis of atrial fibrillation. In: Zipes, D.P.,Jalife, J. (Eds.), Cardiac Arrhythmias.
Allessie, M.A., de Groot, N.M., Houben, R.P., Schotten, U., Boersma, E., Smeets, J.L.,Crijns, H.J., 2010. Electropathological substrate of long-standing persistent atrialfibrillation in patients with structural heart disease: longitudinal dissociation.Circ. Arrhythm. Electrophysiol. 3, 606e615.
Aslanidi, O.V., Colman, M.A., Stott, J., Dobrzynski, H., Boyett, M.R., Holden, A.V.,Zhang, H., 2011. 3D virtual human atria: a computational platform for studyingclinical atrial fibrillation. Prog. Biophys. Mol. Biol. 107, 156e168.
Ausma, J., Litjens, N., Lenders, M.H., Duimel, H., Mast, F., Wouters, L., Ramaekers, F.,Allessie, M., Borgers, M., 2001. Time course of atrial fibrillation-induced cellularstructural remodeling in atria of the goat. J. Mol. Cell. Cardiol. 33, 2083e2094.
Ausma, J., van der Velden, H.M., Lenders, M.H., van Ankeren, E.P., Jongsma, H.J.,Ramaekers, F.C., Borgers, M., Allessie, M.A., 2003. Reverse structural and gap-junctional remodeling after prolonged atrial fibrillation in the goat. Circula-tion 107, 2051e2058.
Beauchamp, P., Yamada, K.A., Baertschi, A.J., Green, K., Kanter, E.M., Saffitz, J.E.,Kleber, A.G., 2006. Relative contributions of connexins 40 and 43 to atrial im-pulse propagation in synthetic strands of neonatal and fetal murine car-diomyocytes. Cir. Res. 99, 1216e1224.
Berenfeld, O., Zaitsev, A.V., Mironov, S.F., Pertsov, A.M., Jalife, J., 2002. Frequency-dependent breakdown of wave propagation into fibrillatory conduction acrossthe pectinate muscle network in the isolated sheep right atrium. Circ. Res. 90,1173e1180.
Burashnikov, A., Antzelevitch, C., 2003. Reinduction of atrial fibrillation immedi-ately after termination of the arrhythmia is mediated by late phase 3 earlyafterdepolarization-induced triggered activity. Circulation 107, 2355e2360.
Colman, M.A., Aslanidi, O.V., Kharche, S., Boyett, M.R., Garratt, C.J., Hancox, J.C.,Zhang, H., 2013. Pro-arrhythmogenic effects of atrial fibrillation induced electricalremodelling e insights from 3D virtual human atria. J. Physiol. 591, 4249e4272.
de Groot, N.M., Houben, R.P., Smeets, J.L., Boersma, E., Schotten, U., Schalij, M.J.,Crijns, H., Allessie, M.A., 2010. Electropathological substrate of longstandingpersistent atrial fibrillation in patients with structural heart disease: epicardialbreakthrough. Circulation 122, 1674e1682.
Derakhchan, K., Li, D., Courtemanche, M., Smith, B., Brouillette, J., Page, P.L.,Nattel, S., 2001. Method for simultaneous epicardial and endocardial mappingof in vivo canine heart: application to atrial conduction properties andarrhythmia mechanisms. J. Cardiovasc. Electrophysiol. 12, 548e555.
Dolber, P.C., Spach, M.S., 1993. Conventional and confocal fluorescence microscopyof collagen fibers in the heart. J. Histochem. Cytochem. 41, 465e469.
Duffy, H.S., Wit, A.L., 2008. Is there a role for remodeled connexins in AF? No simpleanswers. J. Mol. Cell. Cardiol. 44, 4e13.
Eckstein, J., Verheule, S., de Groot, N.M., Allessie, M., Schotten, U., 2008. Mechanismsof perpetuation of atrial fibrillation in chronically dilated atria. Prog. Biophys.Mol. Biol. 97, 435e451.
Eckstein, J., Maesen, B., Linz, D., Zeemering, S., van Hunnik, A., Verheule, S.,Allessie, M., Schotten, U., 2011. Time course and mechanisms of endo-epicardialelectrical dissociation during atrial fibrillation in the goat. Cardiovasc Res. 89,816e824.
Eckstein, J., Zeemering, S., Linz, D., Maesen, B., Verheule, S., van Hunnik, A.,Crijns, H., Allessie, M.A., Schotten, U., 2013. Transmural conduction is the pre-dominant mechanism of breakthrough during atrial fibrillation: evidence fromsimultaneous endo-epicardial high-density activation mapping. Circ. Arrhythm.Electrophysiol. 6, 334e341.
Eijsbouts, S.C., Houben, R.P., Blaauw, Y., Schotten, U., Allessie, M.A., 2004. Synergisticaction of atrial dilation and sodium channel blockade on conduction in rabbitatria. J. Cardiovasc. Electrophysiol. 15, 1453e1461.
Eijsbouts, S., Ausma, J., Blaauw, Y., Schotten, U., Duytschaever, M., Allessie, M.A.,2006. Serial cardioversion by class IC Drugs during 4 months of persistent atrialfibrillation in the goat. J. Cardiovasc. Electrophysiol. 17, 648e654.
Elenes, S., Rubart, M., Moreno, A.P., 1999. Junctional communication between iso-lated pairs of canine atrial cells is mediated by homogeneous and heteroge-neous gap junction channels. J. Cardiovasc. Electrophysiol. 10, 990e1004.
European Heart Rhythm A, European Association for Cardio-Thoracic S, Camm, A.J.,Kirchhof, P., Lip, G.Y., Schotten, U., Savelieva, I., Ernst, S., Van Gelder, I.C., Al-Attar, N., Hindricks, G., Prendergast, B., Heidbuchel, H., Alfieri, O., Angelini, A.,Atar, D., Colonna, P., De Caterina, R., De Sutter, J., Goette, A., Gorenek, B.,Heldal, M., Hohloser, S.H., Kolh, P., Le Heuzey, J.Y., Ponikowski, P., Rutten, F.H.,Guidelines ESCCfP, 2010. Guidelines for the management of atrial fibrillation:the task force for the management of atrial fibrillation of the European Societyof Cardiology (ESC). Europace 12, 1360e1420.
Everett, T.H., Wilson, E.E., Hulley, G.S., Olgin, J.E., 2010. Transmural characteristics ofatrial fibrillation in canine models of structural and electrical atrial remodelingassessed by simultaneous epicardial and endocardial mapping. Heart Rhythm 7,506e517.
Gharaviri, A., Verheule, S., Eckstein, J., Potse, M., Kuijpers, N.H., Schotten, U., 2012.A computer model of endo-epicardial electrical dissociation and transmuralconduction during atrial fibrillation. Europace 14 (Suppl. 5), v10ev16.
Gray, R.A., Pertsov, A.M., Jalife, J., 1996. Incomplete reentry and epicardial break-through patterns during atrial fibrillation in the sheep heart. Circulation 94,2649e2661.
Haissaguerre, M., Jais, P., Shah, D.C., Takahashi, A., Hocini, M., Quiniou, G.,Garrigue, S., Le Mouroux, A., Le Metayer, P., Clementy, J., 1998. Spontaneousinitiation of atrial fibrillation by ectopic beats originating in the pulmonaryveins. N Engl. J. Med. 339, 659e666.
Haissaguerre, M., Lim, K.T., Jacquemet, V., Rotter, M., Dang, L., Hocini, M.,Matsuo, S., Knecht, S., Jais, P., Virag, N., 2007. Atrial fibrillatory cycle length:computer simulation and potential clinical importance. Europace 9 (Suppl. 6),vi64e70.
Hansson, A., Holm, M., Blomstrom, P., Johansson, R., Luhrs, C., Brandt, J., Olsson, S.B.,1998. Right atrial free wall conduction velocity and degree of anisotropy inpatients with stable sinus rhythm studied during open heart surgery. Eur. HeartJ. 19, 293e300.
Hilliard, F.A., Steele, D.S., Laver, D., Yang, Z., Le Marchand, S.J., Chopra, N.,Piston, D.W., Huke, S., Knollmann, B.C., 2010. Flecainide inhibits arrhythmogenicCa2þ waves by open state block of ryanodine receptor Ca2þ release channelsand reduction of Ca2þ spark mass. J. Mol. Cell. Cardiol. 48, 293e301.
Ho, S.Y., Sanchez-Quintana, D., 2009. The importance of atrial structure and fibers.Clin. Anat. 22, 52e63.
Ho, S.Y., Sanchez-Quintana, D., Cabrera, J.A., Anderson, R.H., 1999. Anatomy of theleft atrium: implications for radiofrequency ablation of atrial fibrillation.J. Cardiovasc Electrophysiol. 10, 1525e1533.
Ho, S.Y., Anderson, R.H., Sanchez-Quintana, D., 2002. Atrial structure and fibres:morphologic bases of atrial conduction. Cardiovasc. Res. 54, 325e336.
Hocini, M., Ho, S.Y., Kawara, T., Linnenbank, A.C., Potse, M., Shah, D., Jais, P.,Janse, M.J., Haissaguerre, M., De Bakker, J.M., 2002. Electrical conduction incanine pulmonary veins: electrophysiological and anatomic correlation. Circu-lation 105, 2442e2448.
Houben, R.P., de Groot, N.M., Smeets, J.L., Becker, A.E., Lindemans, F.W.,Allessie, M.A., 2004. S-wave predominance of epicardial electrograms duringatrial fibrillation in humans: indirect evidence for a role of the thin sub-epicardial layer. Heart Rhythm 1, 639e647.

S. Verheule et al. / Progress in Biophysics and Molecular Biology 115 (2014) 173e185 185
Kanagaratnam, P., Rothery, S., Patel, P., Severs, N.J., Peters, N.S., 2002. Relativeexpression of immunolocalized connexins 40 and 43 correlates with humanatrial conduction properties. J. Am. Coll. Cardiol. 39, 116e123.
Konings, K.T., Kirchhof, C.J., Smeets, J.R., Wellens, H.J., Penn, O.C., Allessie, M.A., 1994.High-density mapping of electrically induced atrial fibrillation in humans.Circulation 89, 1665e1680.
Koura, T., Hara, M., Takeuchi, S., Ota, K., Okada, Y., Miyoshi, S., Watanabe, A.,Shiraiwa, K., Mitamura, H., Kodama, I., Ogawa, S., 2002. Anisotropic conductionproperties in canine atria analyzed by high-resolution optical mapping: pref-erential direction of conduction block changes from longitudinal to transversewith increasing age. Circulation 105, 2092e2098.
Krueger, M.W., Severi, S., Rhode, K., Genovesi, S., Weber, F.M., Vincenti, A.,Fabbrini, P., Seemann, G., Razavi, R., Dossel, O., 2011. Alterations of atrial elec-trophysiology related to hemodialysis session: insights from a multiscalecomputer model. J. Electrocardiol. 44, 176e183.
Kuijpers, N.H., Potse, M., van Dam, P.M., ten Eikelder, H.M., Verheule, S.,Prinzen, F.W., Schotten, U., 2011. Mechanoelectrical coupling enhances initiationand affects perpetuation of atrial fibrillation during acute atrial dilation. HeartRhythm 8, 429e436.
Kumagai, K., Khrestian, C., Waldo, A.L., 1997. Simultaneous multisite mappingstudies during induced atrial fibrillation in the sterile pericarditis model. In-sights into the mechanism of its maintenance. Circulation 95, 511e521.
Lee, S., Sahadevan, J., Khrestian, C.M., Durand, D.M., Waldo, A.L., 2013. Highdensity mapping of atrial fibrillation during vagal nerve stimulation in thecanine heart: restudying the Moe hypothesis. J. Cardiovasc. Electrophysiol.24, 328e335.
Maesen, B., Zeemering, S., Afonso, C., Eckstein, J., Burton, R.A., van Hunnik, A.,Stuckey, D.J., Tyler, D., Maessen, J., Grau, V., Verheule, S., Kohl, P., Schotten, U.,2013. Rearrangement of atrial bundle architecture and consequent changes inanisotropy of conduction constitute the 3-dimensional substrate for atrialfibrillation. Circ. Arrhythm. Electrophysiol. 6, 967e975.
Markides, V., Schilling, R.J., Ho, S.Y., Chow, A.W., Davies, D.W., Peters, N.S., 2003.Characterization of left atrial activation in the intact human heart. Circulation107, 733e739.
Moe, G.K., Abildskov, J.A., 1959. Atrial fibrillation as a self-sustaining arrhythmiaindependent of focal discharge. Am. Heart J. 58, 59e70.
Narayan, S.M., Krummen, D.E., Rappel, W.J., 2012. Clinical mapping approach todiagnose electrical rotors and focal impulse sources for human atrial fibrilla-tion. J. Cardiovasc. Electrophysiol. 23, 447e454.
Narayan, S.M., Shivkumar, K., Krummen, D.E., Miller, J.M., Rappel, W.J., 2013.Panoramic electrophysiological mapping but not electrogram morphologyidentifies stable sources for human atrial fibrillation: stable atrial fibrillationrotors and focal sources relate poorly to fractionated electrograms. Circ.Arrhythm. Electrophysiol. 6, 58e67.
Roberts-Thomson, K.C., Stevenson, I.H., Kistler, P.M., Haqqani, H.M., Goldblatt, J.C.,Sanders, P., Kalman, J.M., 2008. Anatomically determined functional conductiondelay in the posterior left atrium relationship to structural heart disease. J. Am.Coll. Cardiol. 51, 856e862.
Rouillard, A.D., Holmes, J.W., 2014. Coupled agent-based and finite-element modelsfor predicting scar structure followingmyocardial infarction. Prog. Bio. Mol. Biol.115 (2-3), 235e243. http://dx.doi.org/10.1016/j.pbiomolbio.2014.06.010.
Ruchat, P., Dang, L., Schlaepfer, J., Virag, N., von Segesser, L.K., Kappenberger, L.,2007. Use of a biophysical model of atrial fibrillation in the interpretation ofthe outcome of surgical ablation procedures. Eur. J. Cardiothorac. Surg. 32,90e95.
Ryu, K., Li, L., Khrestian, C.M., Matsumoto, N., Sahadevan, J., Ruehr, M.L., VanWagoner, D.R., Efimov, I.R., Waldo, A.L., 2007. Effects of sterile pericarditis onconnexins 40 and 43 in the atria: correlation with abnormal conduction andatrial arrhythmias. Am. J. Physiol. Heart Circ. Physiol. 293, H1231eH1241.
Sato, D., Xie, L.H., Sovari, A.A., Tran, D.X., Morita, N., Xie, F., Karagueuzian, H.,Garfinkel, A., Weiss, J.N., Qu, Z., 2009. Synchronization of chaotic early after-depolarizations in the genesis of cardiac arrhythmias. Proc. Natl. Acad. Sci. U. S.Am. 106, 2983e2988.
Scherf, D., 1947. Studies on auricular tachycardia caused by aconitine administra-tion. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol.Med. 64, 233e239.
Schotten, U., Verheule, S., Kirchhof, P., Goette, A., 2011. Pathophysiological mecha-nisms of atrial fibrillation: a translational appraisal. Physiol. Rev. 91, 265e325.
Schuessler, R.B., Kawamoto, T., Hand, D.E., Mitsuno, M., Bromberg, B.I., Cox, J.L.,Boineau, J.P., 1993. Simultaneous epicardial and endocardial activation sequencemapping in the isolated canine right atrium. Circulation 88, 250e263.
Shan, Z., Van Der Voort, P.H., Blaauw, Y., Duytschaever, M., Allessie, M.A., 2004.Fractionation of electrograms and linking of activation during pharmacologiccardioversion of persistent atrial fibrillation in the goat. J. Cardiovasc. Electro-physiol. 15, 572e580.
Skanes, A.C., Mandapati, R., Berenfeld, O., Davidenko, J.M., Jalife, J., 1998. Spatio-temporal periodicity during atrial fibrillation in the isolated sheep heart. Cir-culation 98, 1236e1248.
Spach, M.S., Boineau, J.P., 1997. Microfibrosis produces electrical load variations dueto loss of side-to-side cell connections: a major mechanism of structural heartdisease arrhythmias. Pacing Clin. Electrophysiol. 20, 397e413.
Spach, M.S., Heidlage, J.F., Barr, R.C., Dolber, P.C., 2004. Cell size and communication:role in structural and electrical development and remodeling of the heart. HeartRhythm 1, 500e515.
Tanaka, K., Zlochiver, S., Vikstrom,K.L., Yamazaki,M.,Moreno, J., Klos,M., Zaitsev, A.V.,Vaidyanathan, R., Auerbach, D.S., Landas, S., Guiraudon, G., Jalife, J., Berenfeld, O.,Kalifa, J., 2007. Spatial distribution of fibrosis governs fibrillationwave dynamicsin the posterior left atrium during heart failure. Circ. Res. 101, 839e847.
Uldry, L., Virag, N., Lindemans, F., Vesin, J.M., Kappenberger, L., 2012. Atrial septalpacing for the termination of atrial fibrillation: study in a biophysical model ofhuman atria. Europace 14 (Suppl. 5), v112ev120.
van der Velden, H.M., Ausma, J., Rook, M.B., Hellemons, A.J., van Veen, T.A.,Allessie, M.A., Jongsma, H.J., 2000. Gap junctional remodeling in relation tostabilization of atrial fibrillation in the goat. Cardiovasc. Res. 46, 476e486.
Verheule, S., van Batenburg, C.A., Coenjaerts, F.E., Kirchhoff, S., Willecke, K.,Jongsma, H.J., 1999. Cardiac conduction abnormalities in mice lacking the gapjunction protein connexin40. J. Cardiovasc. Electrophysiol. 10, 1380e1389.
Verheule, S., Tuyls, E., van Hunnik, A., Kuiper, M., Schotten, U., Allessie, M., 2010.Fibrillatory conduction in the atrial free walls of goats in persistent and per-manent atrial fibrillation. Circ. Arrhythm. Electrophysiol. 3, 590e599.
Verheule, S., Tuyls, E., Gharaviri, A., Hulsmans, S., van Hunnik, A., Kuiper, M.,Serroyen, J., Zeemering, S., Kuijpers, N.H., Schotten, U., 2013. Loss of continuityin the thin epicardial layer because of endomysial fibrosis increases thecomplexity of atrial fibrillatory conduction. Circ. Arrhythm. Electrophysiol. 6,202e211.
Vigmond, E.J., Ruckdeschel, R., Trayanova, N., 2001. Reentry in a morphologicallyrealistic atrial model. J. Cardiovasc Electrophysiol. 12, 1046e1054.
Weber, K.T., Pick, R., Jalil, J.E., Janicki, J.S., Carroll, E.P., 1989. Patterns of myocardialfibrosis. J. Mol. Cell. Cardiol. 21 (Suppl. 5), 121e131.
Wu, T.J., Yashima, M., Xie, F., Athill, C.A., Kim, Y.H., Fishbein, M.C., Qu, Z.,Garfinkel, A., Weiss, J.N., Karagueuzian, H.S., Chen, P.S., 1998. Role of pectinatemuscle bundles in the generation and maintenance of intra-atrial reentry:potential implications for the mechanism of conversion between atrial fibril-lation and atrial flutter. Circ. Res. 83, 448e462.
Wu, T.J., Doshi, R.N., Huang, H.L., Blanche, C., Kass, R.M., Trento, A., Cheng, W.,Karagueuzian, H.S., Peter, C.T., Chen, P.S., 2002. Simultaneous biatrial comput-erized mapping during permanent atrial fibrillation in patients with organicheart disease. J. Cardiovasc. Electrophysiol. 13, 571e577.
Yamazaki, M., Mironov, S., Taravant, C., Brec, J., Vaquero, L.M., Bandaru, K.,Avula, U.M., Honjo, H., Kodama, I., Berenfeld, O., Kalifa, J., 2012. Heterogeneousatrial wall thickness and stretch promote scroll waves anchoring during atrialfibrillation. Cardiovasc Res. 94, 48e57.
Yue Yu, T., Syeda, F., Holmes, A., Osborne, B., Dehghani, H., Brain, K.L., Kirchhof, P.,Fabritz, L., 2014. An automated system using spatial oversampling for opticalmapping in murine atria. Development and validation with monophasic andtransmembrane action potentials. Prog. Bio. Mol. Biol.115 (2-3), 340e348. http://dx.doi.org/10.1016/j.pbiomolbio.2014.07.012.
Zhao, J., Butters, T.D., Zhang, H., Pullan, A.J., LeGrice, I.J., Sands, G.B., Smaill, B.H.,2012. An image-based model of atrial muscular architecture: effects of struc-tural anisotropy on electrical activation. Circ. Arrhythm. Electrophysiol. 5,361e370.
Copyright © 2022 FDOKUMEN




















