
Rga2 is a Rho2 GAP that regulates morphogenesis and cell integrity in S. pombe
15
Rga2 is a Rho2 GAP that regulates morphogenesis and cell integrity in S. pombe M a Antonia Villar-Tajadura, 1 Pedro M. Coll, 1 Marisa Madrid, 2 Jose Cansado, 2 Beatriz Santos 1 and Pilar Pérez 1 * 1 Instituto de Microbiología Bioquímica, Consejo Superior de Investigaciones Científicas (CSIC)/Departamento de Microbiología y Genética, Universidad de Salamanca, Edificio Departamental, 37007 Salamanca, Spain. 2 Department of Genetics and Microbiology, University of Murcia, Murcia, Spain. Summary Schizosaccharomyces pombe Rho2 GTPase regu- lates a-D-glucan synthesis and acts upstream of Pck2 to activate the MAP kinase pathway for cell integrity. However, little is known about its regulation. Here we describe Rga2 as a Rho2 GTPase-activating protein (GAP) that regulates cell morphology. rga2 + gene is not essential for growth but its deletion causes longer and thinner cells whereas rga2 + overexpres- sion causes shorter and broader cells. rga2 + over- expression also causes abnormal accumulation of Calcofluor-stained material and cell lysis, suggesting that it also participates in cell wall integrity. Rga2 localizes to growth tips and septum region. The N-terminal region of the protein is required for its correct localization whereas the PH domain is neces- sary exclusively for Rga2 localization to the division area. Also, Rga2 localization depends on polarity markers and on actin polymerization. Rga2 interacts with Rho2 and possesses in vitro and in vivo GAP activity for this GTPase. Accordingly, rga2D cells contain more a-D-glucan and therefore partially sup- press the thermosensitivity of mok1–664 cells, which have a defective a-D-glucan synthase. Additionally, genetic interactions and biochemical analysis sug- gest that Rga2 regulates Rho2–Pck2 interaction and might participate in the regulation of the MAPK cell integrity pathway. Introduction Rho GTPases are key molecules in the modulation of a variety of morphological events in all eukaryotes (Etienne- Manneville and Hall, 2002; Ridley, 2006; Park and Bi, 2007). In yeasts, these GTPases provide the co-ordinated regulation of cell wall biosynthetic enzymes and actin organization that is required to maintain cell integrity and polarized growth (Cabib et al., 1998; Arellano et al., 1999a; García et al., 2006). The Schizosaccharomyces pombe genome contains six genes coding for putative Rho GTPases (Arellano et al., 1996). rho1 + is essential to maintain cell integrity and polarization of the actin cytosk- eleton (Arellano et al., 1996; 1997; Nakano et al., 1997). The Rho1 effects are mediated by its interaction with at least three targets: the (1,3)-b-D-glucan synthase (Arell- ano et al., 1996) and the kinases Pck1 and Pck2 (Arellano et al., 1999b; Sayers et al., 2000). Rho1 regulates both directly and indirectly the biosynthesis of (1–3)-b-D glucan, the major S. pombe cell wall structural component (54–60% of total polysaccharides). Rho2 also interacts with Pck1 and Pck2 (Arellano et al., 1999b) and regulates the a-D-glucan synthesis through Pck2 (Calonge et al., 2000). This kinase is required for the correct activity and localization of Mok1/Ags1, which is responsible for the biosynthesis of a-D glucan, the other major cell wall polymer (28–32% of total polysaccharides). Additionally, Rho2 also acts upstream of Pck2 in the activation of the cell integrity MAP kinase Pmk1/Spm1 (Ma et al., 2006). This kinase is structurally similar to Slt2/Mpk1 from Sac- charomyces cerevisiae (Toda et al., 1996; Zaitsevskaya- Carter and Cooper, 1997) and the mammalian extracellular signal-regulated kinases ERK/MAPK (p42/ p44), which are activated by phorbol esters, cytokines and osmotic stress (Roux and Blenis, 2004). Pmk1 is acti- vated by multiple stress signals (Madrid et al., 2006) and is involved in the maintenance of cell integrity, cytokinesis, ion homeostasis and vacuole fusion. Deletion of any gene coding for the components of the cell integrity cascade, namely mkh1 + (MAPKKK) (Sengar et al., 1997), pek1 + /skh1 + (MAPKK) (Sugiura et al., 1999; Loewith et al., 2000) or pmk1 + /spm1 + (MAPK) (Toda et al., 1996; Zaitsevskaya-Carter and Cooper, 1997) causes morpho- logical alterations, multiseptate phenotype, hypersensitiv- ity to potassium and to b-1,3 glucanases and defective Accepted 3 September, 2008. *For correspondence. E-mail [email protected]; Tel. (+34) 923 121644; Fax (+34) 923 224876. Molecular Microbiology (2008) 70(4), 867–881 doi:10.1111/j.1365-2958.2008.06447.x First published online 10 October 2008 © 2008 The Authors Journal compilation © 2008 Blackwell Publishing Ltd
Transcript of Rga2 is a Rho2 GAP that regulates morphogenesis and cell integrity in S. pombe

Rga2 is a Rho2 GAP that regulates morphogenesis and cellintegrity in S. pombe
Ma Antonia Villar-Tajadura,1 Pedro M. Coll,1
Marisa Madrid,2 Jose Cansado,2 Beatriz Santos1 andPilar Pérez1*1Instituto de Microbiología Bioquímica, Consejo Superiorde Investigaciones Científicas (CSIC)/Departamento deMicrobiología y Genética, Universidad de Salamanca,Edificio Departamental, 37007 Salamanca, Spain.2Department of Genetics and Microbiology, University ofMurcia, Murcia, Spain.
Summary
Schizosaccharomyces pombe Rho2 GTPase regu-lates a-D-glucan synthesis and acts upstream of Pck2to activate the MAP kinase pathway for cell integrity.However, little is known about its regulation. Here wedescribe Rga2 as a Rho2 GTPase-activating protein(GAP) that regulates cell morphology. rga2 + geneis not essential for growth but its deletion causeslonger and thinner cells whereas rga2 + overexpres-sion causes shorter and broader cells. rga2 + over-expression also causes abnormal accumulation ofCalcofluor-stained material and cell lysis, suggestingthat it also participates in cell wall integrity. Rga2localizes to growth tips and septum region. TheN-terminal region of the protein is required for itscorrect localization whereas the PH domain is neces-sary exclusively for Rga2 localization to the divisionarea. Also, Rga2 localization depends on polaritymarkers and on actin polymerization. Rga2 interactswith Rho2 and possesses in vitro and in vivo GAPactivity for this GTPase. Accordingly, rga2D cellscontain more a-D-glucan and therefore partially sup-press the thermosensitivity of mok1–664 cells, whichhave a defective a-D-glucan synthase. Additionally,genetic interactions and biochemical analysis sug-gest that Rga2 regulates Rho2–Pck2 interaction andmight participate in the regulation of the MAPK cellintegrity pathway.
Introduction
Rho GTPases are key molecules in the modulation of avariety of morphological events in all eukaryotes (Etienne-Manneville and Hall, 2002; Ridley, 2006; Park and Bi,2007). In yeasts, these GTPases provide the co-ordinatedregulation of cell wall biosynthetic enzymes and actinorganization that is required to maintain cell integrity andpolarized growth (Cabib et al., 1998; Arellano et al.,1999a; García et al., 2006). The Schizosaccharomycespombe genome contains six genes coding for putativeRho GTPases (Arellano et al., 1996). rho1 + is essential tomaintain cell integrity and polarization of the actin cytosk-eleton (Arellano et al., 1996; 1997; Nakano et al., 1997).The Rho1 effects are mediated by its interaction with atleast three targets: the (1,3)-b-D-glucan synthase (Arell-ano et al., 1996) and the kinases Pck1 and Pck2 (Arellanoet al., 1999b; Sayers et al., 2000). Rho1 regulates bothdirectly and indirectly the biosynthesis of (1–3)-b-Dglucan, the major S. pombe cell wall structural component(54–60% of total polysaccharides). Rho2 also interactswith Pck1 and Pck2 (Arellano et al., 1999b) and regulatesthe a-D-glucan synthesis through Pck2 (Calonge et al.,2000). This kinase is required for the correct activity andlocalization of Mok1/Ags1, which is responsible for thebiosynthesis of a-D glucan, the other major cell wallpolymer (28–32% of total polysaccharides). Additionally,Rho2 also acts upstream of Pck2 in the activation of thecell integrity MAP kinase Pmk1/Spm1 (Ma et al., 2006).This kinase is structurally similar to Slt2/Mpk1 from Sac-charomyces cerevisiae (Toda et al., 1996; Zaitsevskaya-Carter and Cooper, 1997) and the mammalianextracellular signal-regulated kinases ERK/MAPK (p42/p44), which are activated by phorbol esters, cytokines andosmotic stress (Roux and Blenis, 2004). Pmk1 is acti-vated by multiple stress signals (Madrid et al., 2006) andis involved in the maintenance of cell integrity, cytokinesis,ion homeostasis and vacuole fusion. Deletion of anygene coding for the components of the cell integritycascade, namely mkh1 + (MAPKKK) (Sengar et al., 1997),pek1 +/skh1 + (MAPKK) (Sugiura et al., 1999; Loewithet al., 2000) or pmk1 +/spm1 + (MAPK) (Toda et al., 1996;Zaitsevskaya-Carter and Cooper, 1997) causes morpho-logical alterations, multiseptate phenotype, hypersensitiv-ity to potassium and to b-1,3 glucanases and defective
Accepted 3 September, 2008. *For correspondence. [email protected]; Tel. (+34) 923 121644; Fax (+34) 923 224876.
Molecular Microbiology (2008) 70(4), 867–881 � doi:10.1111/j.1365-2958.2008.06447.xFirst published online 10 October 2008
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd

vacuole fusion (Toda et al., 1996; Zaitsevskaya-Carterand Cooper, 1997; Bone et al., 1998; Sugiura et al., 1999;Loewith et al., 2000).
Rho GTPases are positively regulated by GDP-GTPexchange factors and negatively regulated by GTPase-activating proteins (GAPs) and GDP dissociation inhibi-tors (GDIs). These regulators play important roles in thespecificity of Rho functions, as they modify the signallingactivity of the GTPase and contain protein–protein inter-action domains that could be important for the GTPaselocalization and for the interaction with the effectorproteins. Nine genes coding for predicted proteins con-taining Rho GAP domains have been described in thefission yeast genome (Nakano et al., 2001). Three ofthese proteins, Rga1, Rga5 and Rga8, are Rho1 GAPs.Rga1 is involved in the F-actin patch localization and cellmorphogenesis (Nakano et al., 2001). Rga5 mainly regu-lates the Rho1–Pck1 interaction and cytokinesis (Calongeet al., 2003). Rga8 is regulated by Shk1 (Yang et al.,2003), a p21-actived kinase that in turn is regulated byCdc42 (Ottilie et al., 1995). Recently, another Rho GAP,Rga4, has been described as a target of the Orb6 kinaseand has been shown to be a Cdc42 GAP that regulatescell diameter (Das et al., 2007; Tatebe et al., 2008).
In this article, we report the characterization of Rga2, aRho2 GAP that plays a role in the regulation of the cellmorphogenesis and cell integrity pathway.
Results
Rga2 participates in the regulation of morphology andcell wall integrity of S. pombe cells
Schizosaccharomyces pombe genome contains nine openreading frames (ORF) coding for putative Rho GAPs. Oneof them, SPAC26A3.09c, encodes the protein Rga2with 1275 amino acid residues and a molecular mass of143.5 kDa. In silico structural analysis of Rga2 showed thepresence of a C-terminal GAP domain (amino acids 1083–1234), a central putative PH domain (amino acids 720–836) and the N-terminal region with no apparent motifs.
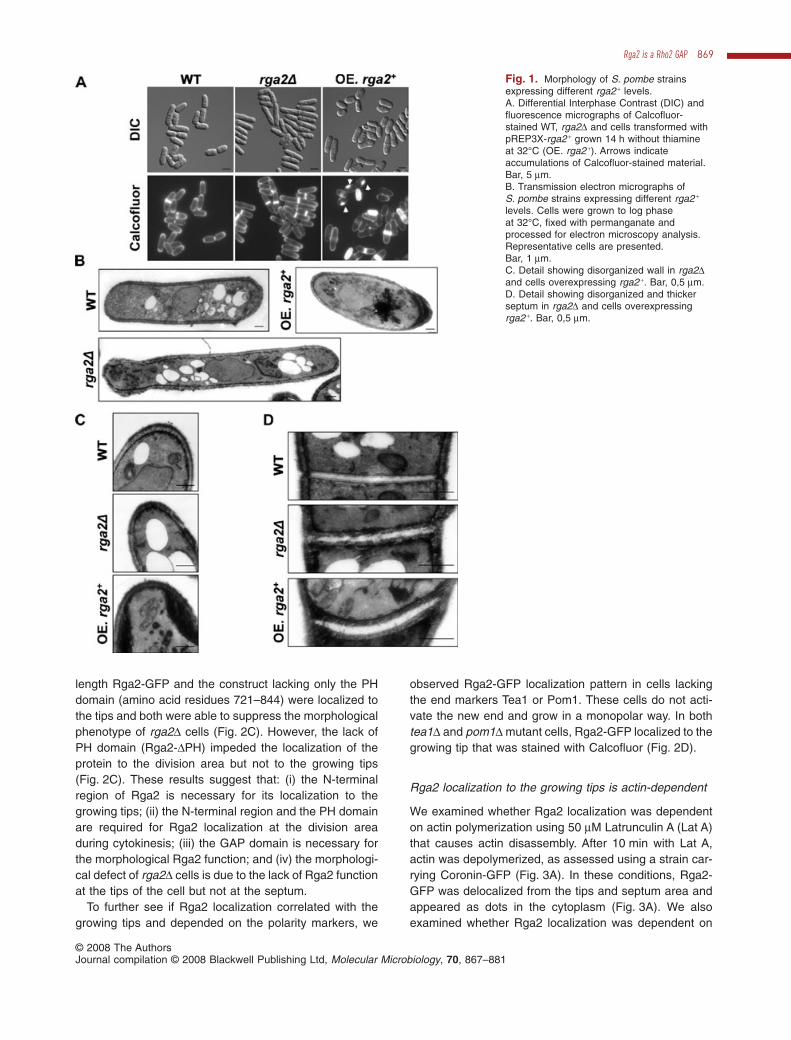
To determine Rga2 function, the rga2 + coding gene wasdeleted by replacing the ORF with KanMX6. As previouslyreported (Nakano et al., 2001), rga2 + is not essential.However, rga2D cells displayed morphological defects;they were 20% longer than wild type (WT) [16.50 �
2.02 mm (n = 100) versus 14.05 � 1.47 mm (n = 100) at thetime of division] and also thinner [3.55 � 0.4 mm (n = 120)versus 3.98 � 0.3 mm (n = 120)] (Fig. 1A and B). In con-trast, S. pombe rga2 +-overexpressing cells were shorterand broader than normal cells (Fig. 1A and B). Theseresults suggest that Rga2 has a role in the regulation of celldimensions, such as cell length and cell diameter that aredependent on the growth area. In addition, around 20% of
the cells overexpressing rga2 + shrank and died whengrown without thiamine for more than 20 h. The addition of1.2 M sorbitol to the medium only partially protected themfrom lysis. Moreover, 15% of the cells showed abnormalaccumulations of Calcofluor-stained material (see arrowsin Fig. 1A). By electron microscopy we can observe thatcells overexpressing rga2 + displayed a disorganized cellwall very thick in some areas. Those thick walls appearedas an electron-dense material that likely corresponds to theCalcofluor-stained accumulations (Fig. 1B and C). In sep-tating cells, the septum of cells overexpressing rga2 + wasalso considerably thicker than that in WT cells (Fig. 1D).The cell wall and septum of cells lacking Rga2 alsoappeared more disordered than in WT cells (Fig. 1C andD). Taken together, these results suggest that Rga2 isdispensable for growth but participates in the regulation ofcell morphology and cell wall integrity.
Rga2 localizes to areas of growth and cell division
We visualized Rga2 by replacing the chromosomal rga2 +
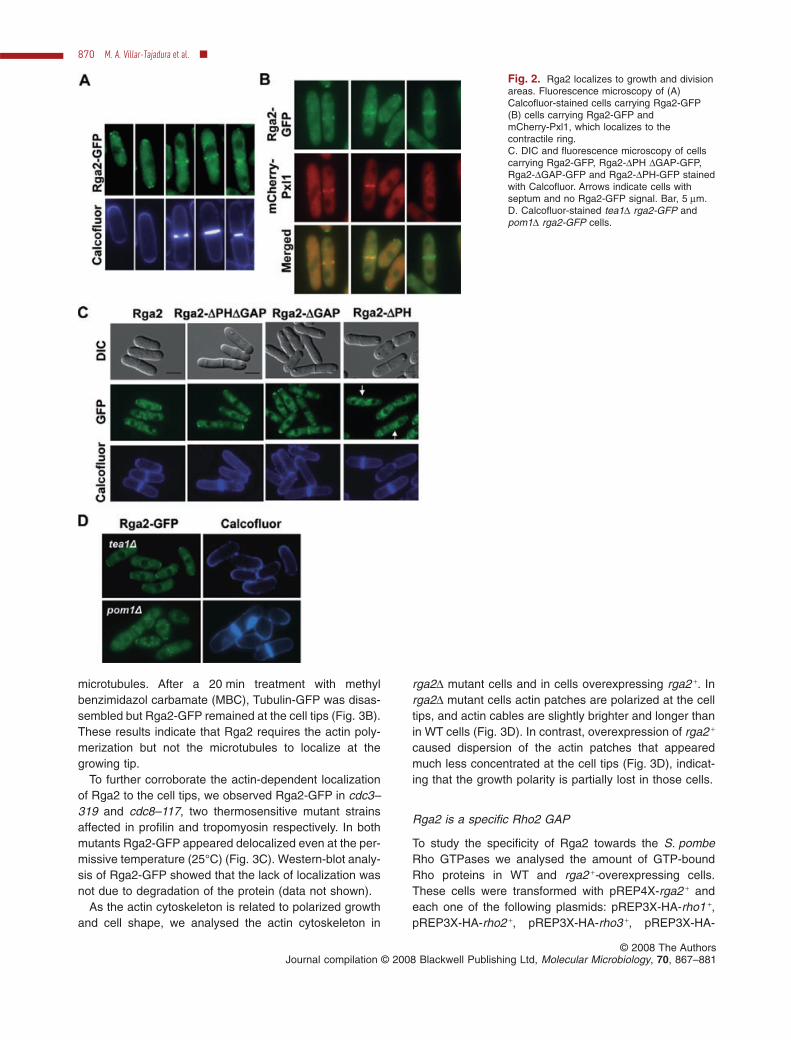
ORF with rga2 +-GFP (Fig. 2A). Cells carrying rga2 +-GFPwere morphologically normal and responded as WT cellsto different concentration of MgCl2 or FK506 (seeFig. 7C), indicating that the fusion protein is functional.The GFP-fluorescence pattern observed in these cellsindicated that Rga2 is dispersed in the cytoplasm andforms some bright dots restricted mainly to the cell tipsand to the medial region in septating cells. Calcofluorstaining showed that Rga2-GFP dots appeared in a singletip when cells were growing in a monopolar way and inboth tips when cells have activated NETO and growth isoccurring at both tips (Fig. 2A). These results suggest thatRga2-GFP could form part of a complex with other pro-teins at the growing regions.
By using a strain carrying Rga2-GFP and mCherry-Pxl1,a protein that localizes to the actomyosin ring (Pinar et al.,2008), we observed that Rga2 appears in the division arealater than Pxl1 and follows the actomyosin ring as it con-tracts during septation, remaining in the area after Pxl1disappearance (Fig. 2B). These results suggest that Rga2goes with the membrane formed upon ring contraction.
To analyse which portion of Rga2 is responsible fortargeting the protein to the growing areas, we expressedin an rga2D background different fragments of rga2 +
fused to EGFP under the endogenous rga2 + promoter.The lack of the N-terminal region (amino acid residues1–344) completely abolished Rga2 localization (data notshown). By contrast, the N-terminal region of Rga2(Rga2DPHDGAP) lacking both the PH and GAP domains(amino acid residues 721–1275) and Rga2DGAP-GFPlacking only the GAP domain (amino acid residues 980–1275) were localized to the growth areas but the cellswere thinner and longer than the WT (Fig. 2C). The full-
868 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881
length Rga2-GFP and the construct lacking only the PHdomain (amino acid residues 721–844) were localized tothe tips and both were able to suppress the morphologicalphenotype of rga2D cells (Fig. 2C). However, the lack ofPH domain (Rga2-DPH) impeded the localization of theprotein to the division area but not to the growing tips(Fig. 2C). These results suggest that: (i) the N-terminalregion of Rga2 is necessary for its localization to thegrowing tips; (ii) the N-terminal region and the PH domainare required for Rga2 localization at the division areaduring cytokinesis; (iii) the GAP domain is necessary forthe morphological Rga2 function; and (iv) the morphologi-cal defect of rga2D cells is due to the lack of Rga2 functionat the tips of the cell but not at the septum.
To further see if Rga2 localization correlated with thegrowing tips and depended on the polarity markers, we
observed Rga2-GFP localization pattern in cells lackingthe end markers Tea1 or Pom1. These cells do not acti-vate the new end and grow in a monopolar way. In bothtea1D and pom1D mutant cells, Rga2-GFP localized to thegrowing tip that was stained with Calcofluor (Fig. 2D).
Rga2 localization to the growing tips is actin-dependent
We examined whether Rga2 localization was dependenton actin polymerization using 50 mM Latrunculin A (Lat A)that causes actin disassembly. After 10 min with Lat A,actin was depolymerized, as assessed using a strain car-rying Coronin-GFP (Fig. 3A). In these conditions, Rga2-GFP was delocalized from the tips and septum area andappeared as dots in the cytoplasm (Fig. 3A). We alsoexamined whether Rga2 localization was dependent on
Fig. 1. Morphology of S. pombe strainsexpressing different rga2 + levels.A. Differential Interphase Contrast (DIC) andfluorescence micrographs of Calcofluor-stained WT, rga2D and cells transformed withpREP3X-rga2 + grown 14 h without thiamineat 32°C (OE. rga2 +). Arrows indicateaccumulations of Calcofluor-stained material.Bar, 5 mm.B. Transmission electron micrographs ofS. pombe strains expressing different rga2 +
levels. Cells were grown to log phaseat 32°C, fixed with permanganate andprocessed for electron microscopy analysis.Representative cells are presented.Bar, 1 mm.C. Detail showing disorganized wall in rga2Dand cells overexpressing rga2 +. Bar, 0,5 mm.D. Detail showing disorganized and thickerseptum in rga2D and cells overexpressingrga2 +. Bar, 0,5 mm.
Rga2 is a Rho2 GAP 869
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881
microtubules. After a 20 min treatment with methylbenzimidazol carbamate (MBC), Tubulin-GFP was disas-sembled but Rga2-GFP remained at the cell tips (Fig. 3B).These results indicate that Rga2 requires the actin poly-merization but not the microtubules to localize at thegrowing tip.
To further corroborate the actin-dependent localizationof Rga2 to the cell tips, we observed Rga2-GFP in cdc3–319 and cdc8–117, two thermosensitive mutant strainsaffected in profilin and tropomyosin respectively. In bothmutants Rga2-GFP appeared delocalized even at the per-missive temperature (25°C) (Fig. 3C). Western-blot analy-sis of Rga2-GFP showed that the lack of localization wasnot due to degradation of the protein (data not shown).
As the actin cytoskeleton is related to polarized growthand cell shape, we analysed the actin cytoskeleton in
rga2D mutant cells and in cells overexpressing rga2 +. Inrga2D mutant cells actin patches are polarized at the celltips, and actin cables are slightly brighter and longer thanin WT cells (Fig. 3D). In contrast, overexpression of rga2 +
caused dispersion of the actin patches that appearedmuch less concentrated at the cell tips (Fig. 3D), indicat-ing that the growth polarity is partially lost in those cells.
Rga2 is a specific Rho2 GAP
To study the specificity of Rga2 towards the S. pombeRho GTPases we analysed the amount of GTP-boundRho proteins in WT and rga2 +-overexpressing cells.These cells were transformed with pREP4X-rga2 + andeach one of the following plasmids: pREP3X-HA-rho1 +,pREP3X-HA-rho2 +, pREP3X-HA-rho3 +, pREP3X-HA-
Fig. 2. Rga2 localizes to growth and divisionareas. Fluorescence microscopy of (A)Calcofluor-stained cells carrying Rga2-GFP(B) cells carrying Rga2-GFP andmCherry-Pxl1, which localizes to thecontractile ring.C. DIC and fluorescence microscopy of cellscarrying Rga2-GFP, Rga2-DPH DGAP-GFP,Rga2-DGAP-GFP and Rga2-DPH-GFP stainedwith Calcofluor. Arrows indicate cells withseptum and no Rga2-GFP signal. Bar, 5 mm.D. Calcofluor-stained tea1D rga2-GFP andpom1D rga2-GFP cells.
870 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

rho4 +, pREP3X-HA-rho5 + and pREP3X-HA-cdc42 +. Thetotal amount of Rho proteins from the extracts was deter-mined by Western-blot using anti-HAmonoclonal antibody,and the GTP-bound Rho proteins were pulled down fromthe extracts with GST-C21RBD (Rho1–5) or GST-CRIB(Cdc42) (see Materials and methods). Cells overexpress-ing rga2 + repeatedly showed a decrease in GTP-boundRho2, whereas there was no change in the GTP-boundlevel of other Rho proteins (Fig. 4A). To corroborate theseresults we analysed the amount of endogenous GTP-bound HA-Rho2, HA-Cdc42 and HA-Rho1 in WT andrga2D cells. We observed that GTP-Rho2 was alwaysincreased in the mutant cells (Fig. 4B). This result, oppo-site to that obtained by overexpressing rga2 +, suggeststhat Rga2 negatively regulates Rho2 activity in vivo. Bycontrast, we also observed a decrease in the endogenousGTP-bound level of HA-Cdc42 in rga2D cells (Fig. 4B),suggesting that Rga2 increases the active Cdc42. GTP-Rho1 was not affected by the lack of Rga2. Interestingly, in
all the experiments performed we observed that the totalamount of Rho2 was lower in rga2D. We analysed the actinas a control of the total proteins loaded and corroboratedthat there was less Rho2 in rga2D cells (68% comparedwith WT). It might be possible that Rho2 is degraded uponactivation as an excess of this GTPase is toxic for the cells(Hirata et al., 1998).
We next tried to determine if there was a direct interac-tion between Rho2 and Rga2, by coimmunoprecipitationexperiments using extracts from cells carrying HA-rho2 +
and a myc-tagged version of rga2 +, both expressed attheir chromosomal loci. Cells carrying HA-cdc42 + andHA-rho1 + were used as control. A specific band corre-sponding to HA-Rho2 was detected in the immunoprecipi-tation of Rga2-Myc using anti-Myc antibody bound toprotein A-sepharose, indicating that both proteins interactin vivo (Fig. 4C). We also observed a very faint HA-Cdc42band in the anti-Myc immunoprecipitates of cells express-ing Rga2-Myc and HA-Cdc42, suggesting that these two
Fig. 3. Rga2 localization depends on actinpolimerization.A. Fluorescence microscopy of cellsexpressing Coronin-GFP or Rga2-GFP grownto mid-log phase and incubated for 10 min inthe presence of DMSO (top) or 50 mM Lat A(bottom).B. Fluorescence microscopy of cellsexpressing Tubulin-GFP or Rga2-GFP grownto mid-log phase and incubated for 20 min inthe presence of 1% DMSO (top) or 25 mg ml-1
MBC (bottom).C. Fluorescence microscopy of WT, cdc3–319and cdc8–117 cells expressing Rga2-GFPgrown to mid-log phase at 25°C andincubated for 3 h at either 25°C (top panels)or 37°C (bottom panels).D. WT, rga2D and cells transformed withpREP3X-rga2 + grown for 14 h withoutthiamine at 32°C (OE. rga2 +) were fixed andstained with Alexafluor 448–phalloidin toobserve actin localization.
Rga2 is a Rho2 GAP 871
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

proteins might interact. No HA-Rho1 band was observedin the corresponding immunoprecipitations.
To corroborate the in vivo Rho2 GAP activity of Rga2,we performed in vitro assays of the Rga2 GAP activitytowards Rho2, Cdc42, Rho1, Rho3 and Rho4. GST fusionproteins of these GTPases, GST-Rga2 and GST werepurified by affinity chromatography from S. pombe cell
extracts, and the GTPase activity of GST-Rho proteinswas measured in the presence of GST-Rga2 or GST. Asshown in Fig. 5, Rga2 was able to increase the rate ofGTP hydrolysis caused by Rho2 but not by any of theother GTPases. Therefore, Rga2 is a specific in vitro GAPtowards Rho2 and negatively regulates the in vivo level ofactive Rho2 in fission yeast.
Fig. 4. Rga2 is a Rho2 GAP.A. Extracts from cells transformed withpREP4X-rga2 + and pREP3X-HA-rho1 +,pREP3X-HA-rho2 +, pREP3X-HA-rho3 +,pREP3X-HA-rho4 +, pREP3X-HA-rho5 + andpREP3X-HA-cdc42 + were precipitated withGST-C21RBD (HA-Rho1–5) or GST-CRIB(HA-Cdc42) and blotted against anti-HAantibody (lower panel). Total HA-Rhoproteins in the extracts were visualized byWestern-blot using anti-HA antibody(upper panel).B. WT and rga2D cells carrying eitherHA-rho2 +, HA-rho1 + or HA-cdc42 + expressedfrom their own promoters were precipitatedwith GST-C21RBD (HA-Rho2 and HA-Rho1)or GST-CRIB (HA-Cdc42) and blotted againstanti-HA antibody (middle panel). Total HA-Rhoproteins in the extracts were visualized byWestern-blot using anti-HA antibody (upperpanel). Actin was visualized in the sameextracts as loading control (lower panel).Quantification of GTP-bound versus total isshown for the corresponding GTPases.C. Coimmunoprecipitation of Rga2 and Rho2,Cdc42 and Rho1. Extracts from cells carryingrga2+-myc and either HA-rho2 +, HA-cdc42 + orHA-rho1 + were immunoprecipitated withpolyclonal anti-Myc antibody and blottedagainst anti-HA antibody or anti-Myc (lowerpanels). Total cell extracts were blotted withanti-Myc (upper panel) or anti-HA antibodies(middle panel) to visualize Rga2-Myc andHA-Rho2 levels respectively.
Fig. 5. Rga2 specifically stimulates Rho2GTPase activity in vitro. Purified GST-Rho2,GST-Cdc42, GST-Rho1, GST-Rho3 andGST-Rho4 were preloaded with [g-32P]GTP, asdescribed in Materials and methods, followedby addition of GST (�) or GST-Rga2 (�).Aliquots were removed at different times andthe amount of radioactivity bound to theprotein was determined. Values are the meanof three independent experiments and errorbars represent standard deviations.
872 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

Rga2 negatively regulates cell wall biosynthesis
Schizosaccharomyces pombe Rho2 GTPase actsupstream of the protein kinase C homologue Pck2 toregulate cell wall a-glucan biosynthesis (Calonge et al.,2000) and to activate the Pmk1 MAPK signalling cascade(Ma et al., 2006). To study the possible functional roleof Rga2 in the regulation of Rho2, we first analysedthe effect of Rga2 in cell wall integrity. rho2D cells arevery sensitive to treatment with zymolyase 100T, ab-glucanase complex that digests the cell wall b-glucan(Calonge et al., 2000). By contrast, rga2D cells were moreresistant than WT cells to b-glucanase treatment, aswould be expected of a negative regulator of Rho2(Fig. 6A). Surprisingly, the double mutant rho2D rga2Dbehaves as the rga2D cells therefore Rga2 is not onlyacting as a negative regulator of Rho2, but it is alsoaltering the cell wall by other means. To further examinethe role of Rga2 in the regulation of cell wall biosynthesis,we analysed the cell wall composition of WT, rga2D andcells transformed with pREP3X-rga2 + and grown withoutthiamine for 14 h at 32°C. As shown in Fig. 6B, there wasa significant increase in the amount of glucose incorpo-rated into the total cell wall of rga2D cells (from35.3 � 2.9% in the WT to 42.1 � 3.1% in rga2D cells),and the main increase was detected in the amount ofa-glucan (24% increase). By contrast, cells overexpress-ing rga2 + showed a decrease in the total cell wall (from35.3 � 2.9% in WT to 27.1 � 1.4% in cells overexpress-ing rga2 +) and there was a 30% decrease in the amountof a-glucan. These results indicate that Rga2 is negativelyregulating the biosynthesis of the cell wall, particularly thea-glucan.
To corroborate these results we looked for a functionalrelationship between Rga2 and Mok1/Ags1, which isresponsible for the biosynthesis of a-glucan in S. pombeand is regulated by Pck2 (Katayama et al., 1999) andRho2 (Calonge et al., 2000). We compared the ther-mosensitive growth of the strain mok1–664 with that of thedouble mutant mok1–664 rga2D. Single mok1–664 growspoorly and hardly forms colonies at 32°C (Fig. 6C and D),whereas the double mutant grew well under these condi-tions, and the morphology of the cells was partiallyrestored (Fig. 6C and D). However, at 36°C neither mok1–664 nor the double mutant mok1–664 rga2D was able togrow (Fig. 6C). This partial suppression might be due toan increase in Rho2 activity and in a-glucan synthesiscaused by the elimination of Rga2.
As mentioned previously in the paper, around 15% ofthe cells overexpressing rga2 + showed abnormal accu-mulations of Calcofluor-stained material, and there was15–20% lysis (Fig. 1A). However, when rga2 + was over-expressed in rho2D cells, we observed a reduction in theabnormal Calcofluor accumulations (from 14% in WT to
4% in rho2D cells, n = 400) and in the lysis (from 18% inWT to 5% in rho2D cells, n = 300). These results suggestthat the cell integrity defects caused by rga2 + overexpres-sion are at least partially mediated by Rho2.
GTP-bound Rho1 and Rho2 interact with Pck1 andPck2, and it has been demonstrated that Rho1 causesthe stabilization of these kinases (Arellano et al., 1999b;Sayers et al., 2000). Although it has not been proved thatGTP-bound Rho2 stabilizes either Pck1 or Pck2, most ofRho2 effects are mediated by Pck2 (Calonge et al., 2000;Ma et al., 2006), probably by stabilizing this kinase.
To explore if Rga2 regulates Rho2–Pck2 interaction weperformed coimmunoprecipitation of these proteins in WTor rga2D strains. As shown in Fig. 6E, the interactionbetween Rho2 and Pck2 was stronger in the absence ofRga2, in agreement with the fact that there is more GTP-Rho2 in the absence of the GAP. We next analysed ifRga2 could affect Pck2 stability, used WT or rga2D strainsin which the pck2 + promoter was replaced by the P41nmtpromoter, and three HA-epitopes were added to theN-terminus of Pck2. The strains were grown for 14 h in theabsence of thiamine to induce the HA-Pck2 expression.Then thiamine was added (time 0), and samples weretaken at 30 min intervals to analyse HA-Pck2 levels byWestern-blot (Fig. 6F). The results showed that Pck2 wasmore stable in a rga2D background, suggesting that Rga2negatively regulates the stability of this kinase.
Rga2 modulates the interaction of Rho2 with the MAPKcell integrity pathway
GTP-bound Rho2 interacts with Pck2 and activates theMAPK integrity pathway (Ma et al., 2006). To explore ifRga2 modulated this pathway, we analysed the geneticinteractions among rga2 + and genes coding for severalcomponents of the MAPK cascade. To this end, weanalysed the growth properties under different cell wallstresses of single- and double-mutant strains rho2D rga2Dor mkh1D rga2D, the last lacking the MAPKKK of theintegrity pathway. It has been described that knockout ofthe components of the Pmk1 MAPK signalling pathwaydisplays hypersensitivity to Micafungin, an echinocandininhibitor of (1,3)-b-D-glucan synthase (Ma et al., 2006).We found that rga2D cells were not hypersensitive to theechinocandin Caspofungin and grow as well as WT cells in0.2 mg ml-1 of this drug, whereas the double mutants,rho2D rga2D or mkh1D rga2D, were as sensitive as thesingle rho2D and mkh1D mutants (Fig. 7A). By contrast,both WT and rga2D cells could not grow in 0.1 mg ml-1
Calcofluor, while double mutants, rho2D rga2D or mkh1Drga2D, were resistant to this compound, as were the singlerho2D and mkh1D mutants (Fig. 7A). Therefore deletionsof genes coding components of the Pmk1 signallingpathway are epistatic to rga2D. Moreover rga2D cells were
Rga2 is a Rho2 GAP 873
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

Fig. 6. Rga2 negatively regulates the cell wall biosynthesis.A. Cells lacking Rga2 display Zymolyase resistance. Sensitivity to Zymolyase was measured in suspensions of log-phase WT, rga2D, rho2Dand rga2D rho2D cells at OD600 1.0. Cells were incubated at 37°C in water containing 25 mg ml-1 of Zymolyase 100T and OD600 was measuredevery 20 min.B. Cell wall composition of S. pombe strains with different levels of Rga2. WT, rga2D and cells transformed with pREP3X-rga2 + (OE. rga2 +)were grown for 14 h without thiamine at 32°C and 14C-glucose was added 6 h before harvesting the cells. Values are the mean of threeindependent experiments with duplicate samples. Error bars represent standard deviations for the total carbohydrate values.C. Partial suppression of mok1–664 mutant phenotype by rga2D. WT, rga2D, mok1–664 and two mok1–664 rga2D strains were grown in EMMmedium at 25°C, spotted on plates at OD600 4.0 and serial 1:4 dilutions. Plates were incubated for 2–4 days at 25°C, 32°C and 36°C.D. DIC micrographs of the early log-phase cells of the same strains grown in EMM liquid medium at 32°C. Bar, 5 mm.E. Coimmunoprecipitation of Rho2 and Pck2 in WT or rga2D strains. Extracts from WT or rga2D cells carrying GFP-rho2 + and HA-pck2 + wereimmunoprecipitated with anti-GFP antibody and blotted against anti-HA antibody (lower panel). Total cell extracts were blotted with anti-HA(upper panel) or anti-GFP antibodies (middle panel) to visualize HA-Pck2 and GFP-Rho2 levels respectively.F. Stability of Pck2 in WT or rga2D strains. Cell extracts from WT or rga2D strains carrying P41nmt-HA-pck2 were obtained after growing thecells for 14 h in EMM. Thiamine was then added to repress the nmt promoter and samples were taken at 30 min intervals. HA-Pck2 levelswere monitored by Western-blot using anti-HA monoclonal antibody (upper panel). Actin was visualized in the same extracts as loading control(lower panel).
874 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

hypersensitive to MgCl2 and slightly more sensitive thanWT cells to the immunosuppressant FK506 (Fig. 7B),whereas double mutants, rho2D rga2D or mkh1D rga2D,were not hypersensitive to MgCl2 and were more resistantto FK506, as were the single rho2D and mkh1D mutants. Ithas been demonstrated that inhibition of calcineurin activ-ity by FK506 results in MgCl2 hypersensitivity in fissionyeast and elimination of cell integrity MAPK pathway com-ponents, such as Mkh1, suppresses the hypersensitivity toCl- (Sugiura et al., 1998; 1999). This phenotype called vic(viable in the presence of immunosuppressant and chlo-ride ion) is also shown by rho2D cells (Ma et al., 2006).rga2D, like the WT cells, did not have vic phenotype andwas sensitive to MgCl2 + FK506, whereas the doublemutants, rho2D rga2D or mkh1D rga2D, were as resistantto those conditions as single rho2D or mkh1D mutants(Fig. 7B). All these experiments (performed at least threetimes with nearly identical results) strongly suggest thatRga2 negatively regulates Rho2 and the activity of Pmk1MAPK. Experiments performed with cells carrying the pre-viously described different fragments of rga2 + fused toEGFP (see Fig. 2C) demonstrated that Rga2 GAP domainis also necessary for the Rga2 function in Cl- homeostasis(Fig. 7C).
To further explore if Rga2 modulated the Pmk1pathway, we analysed the phosphorylation state of Pmk1in growing cells and in response to osmotic stress. Sur-prisingly, we did not observe any significant differencebetween WT and rga2D cells treated with 0.6 M KCl toactivate Pmk1 in response to a hyperosmotic stress, andthe basal phosphorylation of Pmk1 was not increasedeither (Fig. 7D). These results could be explained if theeffects of Rga2 on cell integrity and ion homeostasis weremediated by a signalling pathway not related to Rho2-dependent Pmk1 activation. They could also be explainedif Rga2 was not the only GAP for Rho2 and other redun-dant GAPs could negatively regulate the Rho2-dependentactivation of Pmk1 MAPK. If that were the case, Pmk1hyperactivation caused by the lack of Rga2 would beminor and difficult to detect biochemically. On the con-trary, overexpression of rga2 + should cause a decrease inthe Pmk1 phosphorylation level that might be more easilydetected. In agreement with this prediction, when phos-phorylation of the MAPK was analysed in cells over-expressing rga2 +, there was a clear decrease in Pmk1phosphorylation levels both in growing cells and inresponse to hyperosmotic stress (Fig. 7E). These resultssuggest that Rga2 negatively regulates the Rho2-dependent activation of the MAPK integrity pathway.
Effects of other S. pombe GAP proteins on Rho2
As we discussed above, the absence of an increasedPmk1 basal phosphorylation in rga2D cells could be
Fig. 7. Interaction of Rga2 and the Pmk1 MAPK pathway.A. Growth of WT, rga2D, rho2D, mkh1D, rho2D rga2D or mkh1Drga2D spotted at 1/4 dilution in YES medium supplemented with0.2 mg ml-1 Caspofungin or 0.1 mg ml-1 Calcofluor. Plates wereincubated at 32°C for 2–3 days.B. Growth of the same strains as is (A) in EMM supplemented with0.2 M MgCl2, 0,5 mg ml-1 FK506 or 0.2 M MgCl2 plus 0,5 mg ml-1
FK506. Plates were incubated at 32°C for 3 days.C. Growth of WT, rga2D, rho2D and strains carrying differentfragments of GFP-tagged Rga2 (rga2-GFP, rga2-DGAP-GFP,rga2-DPH DGAP-GFP and rga2-DPH-GFP) in EMM supplementedwith 0.2 M MgCl2 or 0,5 mg ml-1 FK506. Plates were incubated at32°C for 3 days.D. Activation of Pmk1 induced by osmotic stress in differentS. pombe strains expressing pmk1-HA6His. pmk1 +-HA6His (WT),pmk1 +-HA6His rga2D, pmk1 +-HA6His rho2D and pmk1 +-HA6Hisrho2D rga2D cells were grown in YES medium to mid log phaseand KCl was added to 0.6 M final concentration. At timed intervals,Pmk1-HA6H was purified by affinity chromatography under nativeconditions by its His tag. Activated and total Pmk1 were detectedby immunoblotting with antiphospho-p42/44 or anti-HA antibodiesrespectively.E. Activation of Pmk1 induced by osmotic stress in S. pombestrains: pmk1 +-HA6His (WT), pmk1 +-HA6His transformed withpREP3X-rga2 + and pmk1 +-HA6His rho2D. Cells were grown inEMM without thiamine for 14 h and supplemented with 0.6 M KCl.At timed intervals, activated and total Pmk1 was detected as in (D).
Rga2 is a Rho2 GAP 875
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

explained if other GAPs besides Rga2 were also nega-tively regulating Rho2 GTPase. Among S. pombe RhoGAPs, Rga1, Rga5 and Rga8 are Rho1 GAPs (Nakanoet al., 2001; Calonge et al., 2003; Yang et al., 2003), andRga4 is a Cdc42 GAP (Tatebe et al., 2008). We thereforeanalysed if other GAPs, such as Rga3, Rga6 and Rga7,were negative regulators of Rho2. Three independentGTP-Rho2 ‘pull down’ experiments were performed incells carrying endogenous HA-Rho2 and lacking oroverexpressing each of the different GAPs. The resultsobtained (Fig. 8) showed that Rga3 is not altering Rho2level or activation. Rga6 does not have a strong effect inthe level of GTP-Rho2; however, we observed that therewas always less amount of total Rho2 in rga6D cells (86%compared with WT cells). Remarkably, Rga7 has a clearnegative effect in Rho2 activation as detected when Rga7was overexpressed. Additionally there was a significantlower level of total Rho2 (59% compared with WT cells)but a relative increase in GTP-bound Rho2 in rga7D cells.Therefore Rga7 is acting as a Rho2 GAP. Rga7 is aprotein of 695 amino acids containing an N-terminal FCH(Fes/CIP4 homology) domain, with a potential role inregulating the actin cytoskeleton and a C-terminal GAPdomain. Rga7 primary structure is close to S. cerevisiaeRgd1, a GAP for Rho3 and Rho4 (Barthe et al., 1998).
Taking together, these results suggest that Rga2, Rga7and perhaps Rga6 might act as Rho2 GAPs. Moreover,we had observed that cells lacking Rga2, Rga7 or Rga6have a decrease in the amount of total Rho2. If theseRgas are Rho2 GAPs, a possible explanation to thisdecrease would be that Rho2 is being degraded onceactivated.
Discussion
Schizosaccharomyces pombe Rho GTPases interact withmany different effector proteins, but little is known abouthow these GTPases are regulated in a spatial and/ortemporal manner. Rho2 is not essential but participates inthe regulation of the biosynthesis of the a-glucan and thusthe cell wall in general. Rho2 also regulates actin poly-
merization (Hirata et al., 1998) and acts upstream of thePmk1 MAPK cell integrity signalling (Ma et al., 2006). Inthis report, we have provided genetic and molecular evi-dences for the functional interaction between the fissionyeast Rga2, a member of the Rho GAP protein family andthe GTPase Rho2. Some attempts to find the S. pombeorthologues of S. cerevisiae GAPs have been madebased on the primary structures of the proteins and theirGAP domains (Nakano et al., 2001). Rga1 has beenpaired to Lrg1, Rga2 to Bem3, Rga3 and Rga4 to Rga1,Rga5 and Rga6 to Bag7 and Sac7, Rga7 to Rgd1, andRga8 to Rgd2. However, although the primary structuremight reflect some conservation among the protein motifs,conclusions about functional conservation of the GAPs inthese two evolutionarily distant yeasts cannot be drawnbecause most GAPs display activity towards multiple Rhoproteins, and different GAPs might regulate the same Rhoprotein acting in different signalling pathways (Tcherke-zian and Lamarche-Vane, 2007). Moreover, the GAPdomain may act as a recognition module, with or withoutGAP activity. For instance, in animal cells it has beenshown that some proteins, which bind to Rho GTPasesthrough the GAP domain, such as TCGAP, which binds toCdc42 and TC10, are involved in signalling even if they donot display Cdc42 GAP activity in vivo (Chiang et al.,2003). Experimental results might support Rga1 andRga5, pairing to Rho1-GAPs respectively, although theydo not necessary play the same role in fission as inbudding yeast (Park and Bi, 2007). The results presentedin this paper indicate that Rga2, close to the Cdc42 GAPBem3 (Zheng et al., 1993), is in fact a Rho2 GAP inS. pombe, with no effect on the activity of Rho1, 3, 4 and5, and with a positive effect, direct or indirect, in Cdc42activation. Moreover, the morphological alteration causedby the rga2 + deletion consisting in cells becoming longerand thinner occurs even in rho2D cells (data not shown)and might be caused by the decrease of active Cdc42 inthese cells. Interestingly, deletion of Rga4, recentlydescribed as a Cdc42 GAP (Tatebe et al., 2008), causesthe opposite phenotype with wider and shorter cells (Daset al., 2007). Rga2 may play a role antagonistic to Rga4
Fig. 8. Effect of other Rho GAPs on the Rho2 GTPase.A. Extracts from WT and cells lacking or overexpressing the following GAPs: rga2 +, rga3 +, rga6 + and rga7 + and carrying HA-Rho2 wereprecipitated with GST-C21RBD and blotsted against anti-HA antibody (middle panel). Total HA-Rho2 proteins in the extracts were visualizedby Western-blot using anti-HA antibody (upper panel). Actin was visualized in the same extracts as loading control (lower panel).Quantification of GTP-bound versus total Rho2 is shown.
876 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

respect to the regulation of cell dimensions mediated byCdc42. Consistent with this idea, Rga2 and Rga4 localizein opposite areas of the cell. Rga2 localize to the growingareas, similar to the pattern of many polarity moleculessuch as the GTPases Cdc42 (Merla and Johnson, 2000),Rho1 (Arellano et al., 1997) and Rho2 (Hirata et al., 1998)or kinases such as Pck2 (Sayers et al., 2000). By con-trast, Rga4 is excluded from the growing areas, and Pom1is essential for the growing tip exclusion of Rga4 (Tatebeet al., 2008). Pom1 is also necessary for the localizationof Rga2 in both cell tips as pom1D cells have a singlegrowth pole and Rga2 only localizes to this growing pole.However, it is not known if a direct interaction existsbetween both proteins, as in the case for Rga4 (Tatebeet al., 2008), or if other proteins are required to localizeRga2 to the growing area. Rga2 appears as small dots atthe tip membrane, suggesting that it may form part of aprotein complex. Our data suggest that the N-terminalpart of the protein is required and sufficient for its local-ization to the growing tips as deletion of the PH andGAP domain together does not alter the tip localization.Unfortunately, no canonical interaction domains exist inRga2 N-terminal sequence. Once at the tip, Rga2 GAPfunction is required for the regulation of cell dimensionsand ion homeostasis as cells carrying Rga2DGAP orRga2DPHDGAP behave like cells without Rga2. Interest-ingly, Rga2 PH domain is not required either for tip local-ization or for the Rga2 functions. However, it contributesto localize Rga2 to the division area during cytokinesis,suggesting that interaction with phospholipids is alsoimportant for this Rga2 localization.
The coimmunoprecipitation experiments of Rga2 andRho2 demonstrated an in vivo interaction between theseproteins, and the biochemical data strongly corroboratethat Rga2 acts in vitro as a negative regulator of Rho2 andmodulates the level of GTP-Rho2 in vivo, with no detect-able negative effect over other Rho GTPases. Surpris-ingly, a decrease in active GTP-Cdc42 was repeatedlyobserved in rga2D cells, as if Rga2 were a positive regu-lator of Cdc42, and a very weak coimmunoprecipitationwas observed between these two proteins. Although theRga2 effect on this GTPase is probably indirect, it istempting to propose that the opposite effects of Rga2 andRga4, a negative regulator of Cdc42 (Tatebe et al., 2008),are necessary to maintain the cell dimensions. Furtherstudies will be required to demonstrate this hypothesis.
Genetic and biochemical analyses shown in this studyrevealed the physiological relevance of the Rga2-mediated regulation of Rho2. Thus, we observed func-tional relationship between Rga2 and Mok1/Ags1, thea-glucan synthase which is activated by Rho2 via Pck2(Calonge et al., 2000). The spontaneous lysis of mok1–664 cells grown at 32°C was suppressed in the absenceof Rga2. Additionally, rga2 + deletion caused an increase
in cell wall and a-glucan content and reciprocally, overex-pression of rga2 + caused a decrease in cell wall biosyn-thesis, mainly in a-glucan. We previously described thatGTP-bound Rho1 interacts with Pck1 and Pck2 and thatactive Rho1 stabilizes Pck1 and Pck2 (Arellano et al.,1999b; Calonge et al., 2003). Similar experiments in thispaper showed that Rho2 interacts with Pck2 and thiskinase accumulates in rga2D as compared with WT cells.These results point to Rga2 acting as a negative regulatorof the Rho2 interaction and stabilization of Pck2.
Genetic evidence also unveiled the Rga2-mediatedregulation of Rho2 in the activation of the cell integrityMAPK pathway that is mediated by Rho2 via Pck2 (Maet al., 2006; Barba et al., 2008). Thus, cells lacking Rga2have opposite phenotypes than cells lacking Rho2 orMkh1, the MAPKKK. However, when double mutantswere made, rho2 + and mkh1 + were always epistatic torga2 +. In this context, the absence of Rho2 or Mkh1 isenough to suppress the hypersensitivity to MgCl2 orFK506 in rga2D cells, suggesting that they act down-stream in the same signalling pathway. It is worth notingthat we could not detect any increase in the Pmk1 phos-phorylation of rga2D mutant cells compared with that ofWT cells. Nevertheless, in cells overexpressing rga2 +, thePmk1 phosphorylation dropped to 30–40% that of the WTstrain and the kinase was not activated upon osmoticstress, suggesting that Rga2 is also acting as a negativeregulator of the Rho2 interaction with Pck2 and the MAPKpathway. An explanation to these conflicting results wouldbe that Rga2 mainly regulates Rho2 effect on cell wallbiosynthesis and only secondarily regulates the Rho2-mediated MAPK activation. Another possible explanationis that Rga2 regulates Rho2 interaction with Pck2 andmight also regulate Rho2 interaction with Pck1 that wouldexplain the lack of increase in Pmk1 phosphorylation inrga2D cells, as Pck1 and Pck2 play opposite roles in theregulation of this MAPK pathway (Barba et al., 2008). It isalso possible that Rga2 is not the only GAP regulating thesubset of Rho2 that activates the cell integrity pathwayand different Rho2 GAPs might act in regulating theRho2-mediated Pck2 stabilization. Analysis of the in vivoGTP-bound Rho2 levels in cells where other Rho GAPswere deleted or overexpressed showed that Rga7 andperhaps Rga6 might also be negative regulators of Rho2.This redundancy could explain why the lack of Rga2 doesnot cause hyperactivation of the Pmk1 signalling pathway.Furthermore, we had observed a decrease in the totalRho2 level of cells lacking Rga2, Rga7 and Rga6, sug-gesting that GTP-Rho2 is also regulated by degradation.Some examples of Rho regulation by protein degradationhave been described in animal cells (Wang et al., 2003;Schwamborn et al., 2007), and it has been recently pub-lished that Rdi, the only S. cerevisiae Rho GDI, regulatesRho4 degradation by a proteolytic pathway that includes
Rga2 is a Rho2 GAP 877
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

the proteasome, vacuolar proteases and the GSK-3bhomologue Ygk3 (Tiedje et al., 2008). Additional experi-ments would be required to study the hypothethical Rho2regulation by degradation. In summary, our results dem-onstrate that Rga2 is a Rho2 GAP regulating the Rho2signalling pathways and contributes to the maintenance ofthe cell shape and cell wall integrity.
Experimental procedures
Strains, growth conditions and genetic methods
Standard S. pombe media and genetic manipulations wereemployed (Moreno et al., 1991). All the strains used wereisogenic to WT strains 972 h- and 975 h+ and are described inTable S1. The strains were constructed by either tetrad dis-section or random spore germination method. Cells weregrown in rich medium (YES) or minimal medium (EMM)supplemented with the necessary requirements. Escherichiacoli DH5a was used as host for propagation of plasmids.Cells were grown in LB medium supplemented with50 mg ml-1 ampicillin when appropriate. Solid media con-tained 2% agar.
Recombinant DNA methods
All DNA manipulations were carried out by establishedmethods. Enzymes were used according to the recommen-dations of the suppliers. S. pombe was transformed by thelithium acetate method (Ito et al., 1983). The nmt1 + promoter-containing vectors pREP3X and pREP4X (Forsburg andSherman, 1997) were used for the overexpression of rga2 +,which was induced by growing the cells transformed withthese plasmids in the absence of thiamine for 14 h. To deleterga2 + from the S. pombe genome, 1000 bp of the 5′ rga2 +
ORF flanking sequence, the KanMX6 or ura4 + genes and1000 bp of the 3′ rga2 + ORF flanking sequence were clonedinto a Bluescript plasmid and used to transform S. pombecells. Stable transformants were selected and screened byPCR for the appropriate gene replacement. Genomic ver-sions of rga2 + with the GFP, Myc or HA epitope codingsequences fused to the 3′ end of the ORF were generated byPCR-base gene targeting as described (Bähler et al., 1998).Truncated rga2DGAP-GFP and rga2DPHDGAP-GFP werealso generated by PCR-base gene targeting. Truncatedrga2DN-GFP and rga2DPH-GFP with the KanMX6 gene atthe 3′ end of the deletion cassette were generated and clonedinto a Bluescript plasmid flanked by 1000 bp of the 5′ rga2 +
promoter and 1000 bp of the 3′ rga2 + terminator and usedto transform an S. pombe leu1–32 ura4-D18 rga2::ura4 +
strain (PPG4581). Stable transformants were selected andscreened by PCR for the appropriate gene replacement.
Pull down and immunoprecipitation
Extracts from 5 ¥ 108 cells expressing the different taggedproteins were obtained as described (Arellano et al., 1997),using 200 ml of lysis buffer (20 mM Tris/HCl pH 8.0, 2 mMEDTA, 100 mM NaCl, 0.5% NP-40 and 10% Glycerol con-
taining 100 mM p-aminophenyl methanesulphonyl fluoride,2 mg ml-1 leupeptin and 2 mg ml-1 aprotinin). Cell extracts(2–3 mg total protein) were incubated with glutathionesepharose (GS) beads or with the corresponding antibodyand protein A-sepharose beads for 2–4 h at 4°C. The beadswere washed four times with lysis buffer and resuspended insample buffer. Proteins were separated by SDS-PAGE, trans-ferred to Immobilon-P membranes (Millipore), and blotted todetect the GST-, HA- or Myc-fused epitopes with the corre-sponding antibodies and the ECL detection kit (Amersham).Some 40 mg of total protein was used for Western-blot ascontrol of total amount of tagged protein.
In vivo analysis of Rho GAP activity
The amount of GTP-bound Rho proteins were analysed asdescribed (Calonge et al., 2003) using a Rho-GTP pull downassay. Briefly, WT or rga2 +-overexpressing cells weretransformed with the plasmids pREP3X-HA-rho1 +, pREP3X-HA-rho2 +, pREP3X-HA-rho3 +, pREP3X-HA-rho4 +, pREP3X-HA-rho5 + and pREP3X-HA-cdc42 +. The assay was alsoperformed with extracts from WT and rga2D cells containingHA-rho1 +, HA-rho2 + or HA-cdc42 + expressed from their ownpromoter. Cell extracts were obtained using 500 ml of lysisbuffer (50 mM Tris-ClH pH 7.5, 20 mM NaCl, 0.5% NP-40,10% Glycerol; 0,1 mM DTT, 1 mM NaF and 2 mM MgCl2containing 100 mM p-aminophenyl methanesulphonyl fluo-ride, leupeptin, and aprotinin). Some 10 mg of either GST-RBD (Rhotekin Rho-binding domain) or GST-CRIB (Pak2Cdc42-binding domain), previously obtained from E. coliDNA expression, purified and coupled to GS beads, wereused to precipitate the GTP-bound Rho GTPases from 2 mgof the total cell lysates. The extracts were incubated with thebeads for 2 h at 4°C, washed four times and blotted againstanti-HA monoclonal antibody (12CA5, Roche Molecular Bio-chemicals) to detect the corresponding GTP-bound HA-Rhoprotein. The total amounts of Rho proteins from the extractswere determined by Western-blot using the anti-HA mono-clonal antibody. The same kind of assay was performed oncells containing HA-rho2 + expressed from their own promoterand lacking or overexpressing the following Rho GAPs:rga3 +, rga6 + and rga7 +.
In vitro analysis of Rho GAP activity
The entire coding sequences for Rga2, Rho1, Cdc42, Rho2,Rho3 and Rho4 were fused in frame to the C-terminus of theGST sequence using the NdeI-NotI sites in the pREP-KZvector (Craven et al., 1998). These constructs were used totransform S. pombe and expression of the proteins wasinduced by growing the cells in the absence of thiaminefor 14 h. GST-tagged proteins were purified by affinitychromatography. Briefly, cell extracts from 100 ml cultures(0.8 OD600) were obtained as described above using as lysisbuffer (50 mM Tris-HCl, pH 7.6, 0.5% NP-40, 20 mM NaCl,0,1 mM DTT, 1 mM EDTA, 100 mM p-aminophenyl methane-sulphonyl fluoride and protease inhibitors). Cell extracts wereincubated with GS beads at 4°C for 2 h. After centrifugationfor 10 min at 5000 g, the beads were washed three times withlysis buffer and resuspended in 100 ml of the same buffer.
878 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

The GTP hydrolysis by Rho proteins in response to Rga2activation was performed as described by Self and Hall (1995).Briefly, 30 ml of purified GST-Rho proteins on GS beads werefirst incubated for 10 min at 30°C with 32gP-GTP (5 mCi) in 30 mlof 50 mM Tris-HCl, pH 7.6, 0.5% NP-40, 20 mM NaCl, 0,1 mMDTT, 1 mM EDTA to allow loading of the GTPase. The mixturewas then placed on ice and 10 ml of 25 mM MgCl2 were added.Aliquots of 10 ml were mixed with 40 ml of the reaction buffer(20 mM Tris-HCl, pH 7.6, 20 mM NaCl, 0.1 mM DTT, 1 mMGTP, 1 mg ml-1 BSA, 5 mM MgCl2) and 30 ml of either purifiedGST or GST-Rga2. The reactions were placed at 20°C and10 ml aliquots were removed at 2 min intervals and diluted withcold reaction buffer (1 ml).After filtration through nitrocellulosefilters, the amount of radioactivity bound to the protein wasdetermined by scintillation counting.
Purification and detection of activated Pmk1
Cell homogenates were prepared under native conditionsemploying chilled acid-washed glass beads and lysis buffer(10% glycerol, 50 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.1%Nonidet NP-40, plus specific protease and phosphataseinhibitor from Sigma Chemical). The lysates were cleared bycentrifugation at 13000 r.p.m. for 15 min Pmk1-HA6H waspurified by using Ni2 � NTA-agarose beads (Qiagen Inc.).The purified proteins were resolved in 10% SDS-PAGE gels,transferred to nitrocellulose filters and incubated with eithermonoclonal mouse anti-HA or polyclonal rabbit antiphospho-p42/44 antibodies (Cell Signalling Technologies). The immu-noreactive bands were revealed with anti-mouse or anti-rabbit HRP-conjugated secondary antibodies (SigmaChemical) and the the ECL detection kit.
Microscopy techniques
For Calcofluor-white staining, exponentially growing cellswere harvested, washed and resuspended in a Calcofluorsolution (0.1 mg ml-1) for 5 min at room temperature. Afterwashing with water, cells were observed in a microscope withthe corresponding UV filter. Actin staining was performed withphalloidin-Alexafluor 488. To depolymerize F-actin, Lat A dis-solved in dimethyl sulfoxide (DMSO) at 50 mM was added toS. pombe cultures to a final concentration of 50 mM. TheMBC (dissolved in DMSO at 1 mg ml-1) was used at a finalconcentration of 25 mg ml-1.
Cell samples were observed using a Leica DMXRA micro-scope equipped for Nomarski optics and epifluorescence andphotographed with a Photometrics Sensys camera. Electronmicroscopy was performed on permanganate-stained cellsas described (Wang et al., 2002).
Other methods
Sensitivity to b-glucanases complex was evaluated followingthe procedure previously described (Calonge et al., 2000).WT and rga2D cells were grown to mid-logarithmic phase,harvested, washed and resuspended at an OD600 of 1.0 inwater containing 25 mg ml-1 of Zymolyase-100T. Cell suspen-sions were incubated at 37°C with shaking, and cell lysiswas monitored by measuring the OD600. Labelling and frac-
tionation of cell wall polysaccharides were performed asdescribed (Arellano et al., 1997). Plate assays for growthsensitivity to different compounds were performed by spottingthe appropriate dilutions of log phase-growing cells on YES orEMM solid media containing 2% (w/v) bacto-agar supple-mented with the corresponding compounds. Incubation waseither at 28°C for 3 days or at 37°C for 2–3 days. All experi-ments were repeated at least three times.
Acknowledgements
We thank J.C. Ribas, Y. Sánchez and H. Valdivieso for usefulcomments. Thanks to D. Posner for language revision. Weare very grateful to M. Balasubramanian, K. Nakano andP. Nurse, for generous gifts of strains and plasmids. Thanksto C. Castro and C. Belinchón for technical help with themicroscopy. M.A. Villar-Tajadura was supported by a fellow-ship from the Spanish Ministerio de Educación. This workwas supported by grants BIO2004-0834, BFU2005-01401and BFU2007-60675 from the Comisión Interministerial deCiencia y Tecnología, Spain and by grant SA078A07 fromJunta de Castilla y León, Spain.
References
Arellano, M., Durán, A., and Pérez, P. (1996) Rho1 GTPaseactivates the (1–3) b-D-glucan synthase and is involved inSchizosaccharomyces pombe morphogenesis. EMBO J15: 4584–4591.
Arellano, M., Durán, A., and Pérez, P. (1997) Localization ofthe Schizosaccharomyces pombe rho1 GTPase and itsinvolvement in the organization of the actin cytoskeleton.J Cell Sci 110: 2547–2555.
Arellano, M., Coll, P.M., and Pérez, P. (1999a) Rho GTPasesin the control of cell morphology, cell polarity, and actinlocalization in fission yeast. Microsc Res Tech 47: 51–60.
Arellano, M., Valdivieso, M.H., Calonge, T.M., Coll, P.M.,Durán, A., and Pérez, P. (1999b) Schizosaccharomycespombe protein kinase C homologues, pck1p and pck2p,are targets of rho1p and rho2p and differentially regulatecell integrity. J Cell Sci 112: 3569–3578.
Bähler, J., Wu, J.-Q., Longtine, M.S., Shah, N.G., McKenzie,A., Steever, A.B., et al. (1998) Heterologous modulesfor efficient and versatile PCR-based gene targeting inSchizosaccharomyces pombe. Yeast 14: 943–951.
Barba, G., Soto, T., Madrid, M., Nunez, A., Vicente, J., Gacto,M., and Cansado, J. (2008) Activation of the cell integritypathway is channelled through diverse signalling elementsin fission yeast. Cell Signal 20: 748–757.
Barthe, C., de Bettignies, G., Louvet, O., Peypouquet, M.F.,Morel, C., Doignon, F., and Crouzet, M. (1998) First char-acterization of the gene RGD1 in the yeast Saccharomycescerevisiae. C R Acad Sci III 321: 453–462.
Bone, N., Millar, J.B.A., Toda, T., and Armstrong, J. (1998)Regulated vacuole fusion and fission in Schizosaccharo-myces pombe: an osmotic response dependent on MAPkinases. Curr Biol 8: 135–144.
Cabib, E., Drgonova, J., and Drgon, T. (1998) Role of small Gproteins in yeast cell polarization and wall biosynthesis.Annu Rev Biochem 67: 307–333.
Rga2 is a Rho2 GAP 879
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

Calonge, T.M., Nakano, K., Arellano, M., Arai, R., Katayama,S., Toda, T., et al. (2000) Schizosaccharomyces pombeRho2 GTPase regulates the cell wall a-glucan biosynthe-sis, through the protein kinase Pck2p. Mol Biol Cell 11:4393–4401.
Calonge, T.M., Arellano, M., Coll, P.M., and Pérez, P. (2003)Rga5p is a specific Rho1p GTPase-activating protein thatregulates cell integrity in Schizosaccharomyces pombe.Mol Microbiol 47: 507–518.
Chiang, S.H., Hwang, J., Legendre, M., Zhang, M., Kimura,A., and Saltiel, A.R. (2003) TCGAP, a multidomain RhoGTPase-activating protein involved in insulin-stimulatedglucose transport. EMBO J 22: 2679–2691.
Craven, R.A., Griffiths, D.J., Sheldrick, K.S., Randall, R.E.,Hagan, I.M., and Carr, A.M. (1998) Vectors for the expres-sion of tagged proteins in Schizosaccharomyces pombe.Gene 221: 59–68.
Das, M., Wiley, D.J., Medina, S., Vincent, H.A., Larrea, M.,Oriolo, A., and Verde, F. (2007) Regulation of cell diameter,For3p localization, and cell symmetry by fission yeast Rho-GAP Rga4p. Mol Biol Cell 18: 2090–2101.
Etienne-Manneville, S., and Hall, A. (2002) Rho GTPases incell biology. Nature 420: 629–635.
Forsburg, S.L., and Sherman, D.A. (1997) General purposetagging vectors for fission yeast. Gene 191: 191–195.
García, P., Tajadura, V., García, I., and Sánchez, Y. (2006)Role of Rho GTPases and Rho-GEFs in the regulation ofcell shape and integrity in fission yeast. Yeast 23: 1031–1043.
Hirata, D., Nakano, K., Fukui, M., Takenaka, H., Miyakawa,T., and Mabuchi, I. (1998) Genes that cause aberrant cellmorphology by overexpression in fission yeast: a role for asmall GTP-binding protein Rho2 in cell morphogenesis.J Cell Sci 111: 149–159.
Ito, H., Fukuda, Y., Murata, K., and Kimura, A. (1983) Trans-formation of intact yeast cells treated with alkali cations.J Bacteriol 153: 163–168.
Katayama, S., Hirata, D., Arellano, M., Pérez, P., and Toda,T. (1999) Fission yeast a-glucan synthase Mok1 requiresthe actin cytoskeleton to localize the sites of growth andplays an essential role in cell morphogenesis downstreamof protein kinase C function. J Cell Biol 144: 1173–1186.
Loewith, R., Hubberstey, A., and Young, D. (2000) Skh1, theMEK component of the Mkh1 signalling pathway inSchizosaccharomyces pombe. J Cell Sci 113: 153–160.
Ma, Y., Kuno, T., Kita, A., Asayama, Y., and Sugiura, R. (2006)Rho2 is a target of the farnesyltransferase Cpp1 and actsupstream of Pmk1 mitogen-activated protein kinase signal-ling in fission yeast. Mol Biol Cell 17: 5028–5037.
Madrid, M., Soto, T., Khong, H.K., Franco, A., Vicente, J.,Pérez, P., et al. (2006) Stress-induced response, localiza-tion, and regulation of the Pmk1 cell integrity pathway inSchizosaccharomyces pombe. J Biol Chem 281: 2033–2043.
Merla, A., and Johnson, D.I. (2000) The Cdc42p GTPase istargeted to the site of cell division in the fission yeastSchizosaccharomyces pombe. Int J Cell Biol 79: 469–477.
Moreno, S., Klar, A., and Nurse, P. (1991) Molecular geneticanalysis of fission yeast Schizosaccharomyces pombe.Methods Enzymol 194: 795–823.
Nakano, K., Arai, R., and Mabuchi, I. (1997) The small GTP-
binding protein Rho1 is a multifunctional protein that regu-lates actin localization, cell polarity, and septum formationin the fission yeast Schizosaccharomyces pombe. GenesCells 2: 679–694.
Nakano, K., Mutoh, T., and Mabuchi, I. (2001) Characteriza-tion of GTPase-activating proteins for the function of theRho-family small GTPases in the fission yeast Schizosac-charomyces pombe. Genes Cells 6: 1031–1042.
Ottilie, S., Miller, P.J., Johnson, D.I., Creasy, C.L., Sells,M.A., Bagrodia, S., et al. (1995) Fission yeast pak1 +
encodes a protein kinase that interacts with Cdc42p and isinvolved in the control of cell polarity and mating. EMBO J14: 5908–5919.
Park, H.O., and Bi, E. (2007) Central roles of small GTPasesin the development of cell polarity in yeast and beyond.Microbiol Mol Biol Rev 71: 48–96.
Pinar, M., Coll, P.M., Rincon, S.A., and Pérez, P. (2008)Schizosaccharomyces pombe Pxl1 Is a Paxillin Homo-logue That Modulates Rho1 Activity and Participates inCytokinesis. Mol Biol Cell 19: 1727–1738.
Ridley, A.J. (2006) Rho GTPases and actin dynamics inmembrane protrusions and vesicle trafficking. Trends CellBiol 16: 522–529.
Roux, P.P., and Blenis, J. (2004) ERK and p38 MAPK-activated protein kinases: a family of protein kinases withdiverse biological functions. Microbiol Mol Biol Rev 68:320–344.
Sayers, L.G., Katayama, S., Nakano, K., Mellor, H., Mabuchi,I., Toda, T., and Parker, P.J. (2000) Rho-dependence ofSchizosaccharomyces pombe Pck2. Genes Cells 5: 17–27.
Schwamborn, J.C., Muller, M., Becker, A.H., and Puschel,A.W. (2007) Ubiquitination of the GTPase Rap1B by theubiquitin ligase Smurf2 is required for the establishment ofneuronal polarity. EMBO J 26: 1410–1422.
Self, A.J., and Hall, A. (1995) Measurement of intrinsicnucleotide exchange and GTP hydrolysis rates. MethodsEnzymol 256: 67–76.
Sengar, A.S., Markley, N.A., Marini, N.J., and Young, D.(1997) Mkh1, a MEK kinase required for cell wall integrityand proper response to osmotic and temperature stress inSchizosaccharomyces pombe. Mol Cell Biol 17: 3508–3519.
Sugiura, R., Toda, T., Shuntoh, H., Yanagida, M., and Kuno,T. (1998) pmp1 +, a suppressor of calcineurin deficiency,encodes a novel MAP kinase phosphatase in fission yeast.EMBO J 17: 140–148.
Sugiura, R., Toda, T., Dhut, S., Shuntoh, H., and Kuno, T.(1999) The MAPK kinase Pek1 acts as a phosphorylation-dependent molecular switch. Nature 399: 479–483.
Tatebe, H., Nakano, K., Maximo, R., and Shiozaki, K. (2008)Pom1 DYRK Regulates Localization of the Rga4 GAP toEnsure Bipolar Activation of Cdc42 in Fission Yeast. CurrBiol 18: 322–330.
Tcherkezian, J., and Lamarche-Vane, N. (2007) Currentknowledge of the large RhoGAP family of proteins. Biol Cell99: 67–86.
Tiedje, C., Sakwa, I., Just, U., and Höfken, T. (2008) The RhoGDI Rdi1 regulates Rho GTPases by distinct mechanisms.Mol Biol Cell 19: 2885–2896.
Toda, T., Dhut, S., Superti-Furga, G., Gotoh, Y., Nishida, E.,Sugiura, R., and Kuno, T. (1996) The fission yeast pmk1+
880 M. A. Villar-Tajadura et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881

gene encodes a novel mitogen-activated protein kinasehomologue which regulates cell integrity and functionsco-ordinately with the protein kinase C pathway. Mol CellBiol 16: 6752–6764.
Wang, H., Tang, X., Liu, J., Trautmann, S., Balasundaram, D.,McCollum, D., and Balasubramanian, M.K. (2002) TheMultiprotein Exocyst Complex Is Essential for Cell Separa-tion in Schizosaccharomyces pombe. Mol Biol Cell 13:515–529.
Wang, H.R., Zhang, Y., Ozdamar, B., Ogunjimi, A.A., Alex-androva, E., Thomsen, G.H., and Wrana, J.L. (2003) Regu-lation of cell polarity and protrusion formation by targetingRhoA for degradation. Science 302: 1775–1779.
Yang, P., Qyang, Y., Bartholomeusz, G., Zhou, X., andMarcus, S. (2003) The novel Rho GTPase-activatingprotein family protein, Rga8, provides a potential linkbetween Cdc42/p21-activated kinase and Rho signallingpathways in the fission yeast, Schizosaccharomycespombe. J Biol Chem 278: 48821–48830.
Zaitsevskaya-Carter, T., and Cooper, J.A. (1997) Spm1, astress-activated MAP kinase that regulates morphogenesisin S. pombe. EMBO J 16: 1318–1331.
Zheng, Y., Hart, M.J., Shinjo, K., Evans, T., Bender, A., andCerione, R.A. (1993) Biochemical comparisons of the Sac-charomyces cerevisiae Bem2 and Bem3 proteins. Delinea-tion of a limit Cdc42 GTPase-activating protein domain.J Biol Chem 268: 24629–24634.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
Rga2 is a Rho2 GAP 881
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 867–881




















