Reward- and Attention-related Biasing of Sensory Selection in Visual Cortex
17
Reward- and Attention-related Biasing of Sensory Selection in Visual Cortex Antje Buschschulte 1 , Carsten N. Boehler 2 , Hendrik Strumpf 1 , Christian Stoppel 1 , Hans-Jochen Heinze 1,3 , Mircea A. Schoenfeld 1,3 , and Jens-Max Hopf 1,3 Abstract ■ Attention to task-relevant features leads to a biasing of sensory selection in extrastriate cortex. Features signaling reward seem to produce a similar bias, but how modulatory effects due to reward and attention relate to each other is largely unexplored. To address this issue, it is critical to separate top–down settings defining reward relevance from those defining attention. To this end, we used a visual search paradigm in which the targetʼs definition (attention to color) was dissociated from reward rele- vance by delivering monetary reward on search frames where a certain task-irrelevant color was combined with the target- defining color to form the target object. We assessed the state of neural biasing for the attended and reward-relevant color by analyzing the neuromagnetic brain response to asynchro- nously presented irrelevant distractor probes drawn in the target-defining color, the reward-relevant color, and a com- pletely irrelevant color as a reference. We observed that for the prospect of moderate rewards, the target-defining color but not the reward-relevant color produced a selective enhance- ment of the neuromagnetic response between 180 and 280 msec in ventral extrastriate visual cortex. Increasing reward prospect caused a delayed attenuation (220–250 msec) of the response to reward probes, which followed a prior (160–180 msec) response enhancement in dorsal ACC. Notably, shorter latency responses in dorsal ACC were associated with stronger attenua- tion in extrastriate visual cortex. Finally, an analysis of the brain response to the search frames revealed that the presence of the reward-relevant color in search distractors elicited an enhanced response that was abolished after increasing reward size. The present data together indicate that when top–down definitions of reward relevance and attention are separated, the behavioral significance of reward-associated features is still rapidly coded in higher-level cortex areas, thereby commanding effective top– down inhibitory control to counter a selection bias for those features in extrastriate visual cortex. ■ INTRODUCTION Considerable experimental evidence suggests that the sudden onset of an unexpected visual stimulus can draw attention in an automatic bottom–up fashion—an effect referred to as attentional capture (Theeuwes, 2010; Theeuwes & Godljn, 2001; Yantis, 1996). Such capture, however, turns out to be not entirely automatic, as it has been shown to depend on the degree to which fea- tures of the capturing stimulus match the set of top– down defined descriptions of the task-relevant input (contingent involuntary orienting; Eimer & Kiss, 2008; Lamy, Leber, & Egeth, 2004; Gibson & Kelsey, 1998; Bacon & Egeth, 1994; Folk, Remington, & Wright, 1994; Folk, Remington, & Johnston, 1992). Top–down descrip- tions may entail a selection bias for one or a set of simple target-defining features (e.g., color, orientation) or refer to a selection bias contingent on the more general behav- ioral relevance of a target object, for example, its emo- tional significance, or association with reward. Attentional mechanisms of top – down biasing of feature selection in visual cortex have been extensively investigated, with data from different methodologies indicating that attention exerts its effect by modulating the gain of neural processing in sensory visual areas (Hopf, Heinze, Schoenfeld, & Hillyard, 2009; Maunsell & Treue, 2006; Treue, 2001; Kastner & Ungerleider, 2000; Hillyard, Vogel, & Luck, 1998). This may come about either by increasing the gain of neural represen- tations that match the attended feature (Maunsell & Treue, 2006; Treue & Martinez Trujillo, 1999; Motter, 1994) or by attenuating neural activity of irrelevant fea- ture representations (Lennert & Martinez-Trujillo, 2011; Chelazzi, Duncan, Miller, & Desimone, 1998; Chelazzi & Desimone, 1994). In addition, more recent research has mounted evidence suggesting that value-based factors like the reward relevance of stimuli may have an analo- gous modulatory influence on feature-selective processing in visual cortex (Hickey, Chelazzi, & Theeuwes, 2010; Franko, Seitz, & Vogels, 2009; Serences, 2008; Shuler & Bear, 2006) and recent behavioral and neurophysiological observations imply an influence on even earlier processing stages (Hickey & van Zoest, 2012; Ikeda & Hikosaka, 2003). 1 Otto-von-Guericke University Magdeburg, 2 Ghent University, 3 Leibniz Institute for Neurobiology, Magdeburg © 2014 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 26:5, pp. 1049–1065 doi:10.1162/jocn_a_00539
-
Upload
ifn-magdeburg -
Category
Documents
-
view
3 -
download
0
Transcript of Reward- and Attention-related Biasing of Sensory Selection in Visual Cortex

Reward- and Attention-related Biasing ofSensory Selection in Visual Cortex
Antje Buschschulte1, Carsten N. Boehler2, Hendrik Strumpf1,Christian Stoppel1, Hans-Jochen Heinze1,3, Mircea A. Schoenfeld1,3,
and Jens-Max Hopf1,3
Abstract
■ Attention to task-relevant features leads to a biasing of sensoryselection in extrastriate cortex. Features signaling reward seemto produce a similar bias, but how modulatory effects due toreward and attention relate to each other is largely unexplored.To address this issue, it is critical to separate top–down settingsdefining reward relevance from those defining attention. To thisend, we used a visual search paradigm in which the targetʼsdefinition (attention to color) was dissociated from reward rele-vance by delivering monetary reward on search frames wherea certain task-irrelevant color was combined with the target-defining color to form the target object. We assessed the stateof neural biasing for the attended and reward-relevant color byanalyzing the neuromagnetic brain response to asynchro-nously presented irrelevant distractor probes drawn in thetarget-defining color, the reward-relevant color, and a com-pletely irrelevant color as a reference. We observed that for theprospect of moderate rewards, the target-defining color but
not the reward-relevant color produced a selective enhance-ment of the neuromagnetic response between 180 and 280 msecin ventral extrastriate visual cortex. Increasing reward prospectcaused a delayed attenuation (220–250 msec) of the responseto reward probes, which followed a prior (160–180 msec)response enhancement in dorsal ACC. Notably, shorter latencyresponses in dorsal ACC were associated with stronger attenua-tion in extrastriate visual cortex. Finally, an analysis of the brainresponse to the search frames revealed that the presence of thereward-relevant color in search distractors elicited an enhancedresponse that was abolished after increasing reward size. Thepresent data together indicate that when top–down definitionsof reward relevance and attention are separated, the behavioralsignificance of reward-associated features is still rapidly codedin higher-level cortex areas, thereby commanding effective top–down inhibitory control to counter a selection bias for thosefeatures in extrastriate visual cortex. ■
INTRODUCTION
Considerable experimental evidence suggests that thesudden onset of an unexpected visual stimulus can drawattention in an automatic bottom–up fashion—an effectreferred to as attentional capture (Theeuwes, 2010;Theeuwes & Godljn, 2001; Yantis, 1996). Such capture,however, turns out to be not entirely automatic, as ithas been shown to depend on the degree to which fea-tures of the capturing stimulus match the set of top–down defined descriptions of the task-relevant input(contingent involuntary orienting; Eimer & Kiss, 2008;Lamy, Leber, & Egeth, 2004; Gibson & Kelsey, 1998;Bacon & Egeth, 1994; Folk, Remington, & Wright, 1994;Folk, Remington, & Johnston, 1992). Top–down descrip-tions may entail a selection bias for one or a set of simpletarget-defining features (e.g., color, orientation) or referto a selection bias contingent on the more general behav-ioral relevance of a target object, for example, its emo-tional significance, or association with reward.
Attentional mechanisms of top–down biasing offeature selection in visual cortex have been extensivelyinvestigated, with data from different methodologiesindicating that attention exerts its effect by modulatingthe gain of neural processing in sensory visual areas(Hopf, Heinze, Schoenfeld, & Hillyard, 2009; Maunsell& Treue, 2006; Treue, 2001; Kastner & Ungerleider,2000; Hillyard, Vogel, & Luck, 1998). This may comeabout either by increasing the gain of neural represen-tations that match the attended feature (Maunsell &Treue, 2006; Treue & Martinez Trujillo, 1999; Motter,1994) or by attenuating neural activity of irrelevant fea-ture representations (Lennert & Martinez-Trujillo, 2011;Chelazzi, Duncan, Miller, & Desimone, 1998; Chelazzi &Desimone, 1994). In addition, more recent research hasmounted evidence suggesting that value-based factorslike the reward relevance of stimuli may have an analo-gous modulatory influence on feature-selective processingin visual cortex (Hickey, Chelazzi, & Theeuwes, 2010;Franko, Seitz, & Vogels, 2009; Serences, 2008; Shuler &Bear, 2006) and recent behavioral and neurophysiologicalobservations imply an influence on even earlier processingstages (Hickey & van Zoest, 2012; Ikeda &Hikosaka, 2003).
1Otto-von-Guericke University Magdeburg, 2Ghent University,3Leibniz Institute for Neurobiology, Magdeburg
© 2014 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 26:5, pp. 1049–1065doi:10.1162/jocn_a_00539

These observations raise the question how reward rele-vance imposes its effect on visual sensory processing.Despite the currently proliferating knowledge about braincircuits and functions mediating reward relevance in gen-eral (Haber & Knutson, 2010; Hikosaka, Bromberg-Martin,Hong, & Matsumoto, 2008; Schultz, 2000), the mechanismby which reward ultimately influences visual sensory selec-tion and whether they dissociate from modulatory effectsof attention is not well understood.
In fact, Maunsell (2004) emphasized the importance toclarify whether neural correlates of attention and rewarddissociate in their impact on early visual processing be-cause in animal experiments attention is typically tiedto the application of reward and therefore inherently atrisk to be confounded by reward (and vice versa). A fewrecent studies have explicitly aimed at a dissociation ofreward effects from attention in humans and monkeys,with mixed results ranging from modest (Weil et al.,2010) to clear-cut effects attributable to reward proper(Arsenault, Nelissen, Jarraya, & Vanduffel, 2013; Hickeyet al., 2010). Importantly, in humans, top–down settingsfor task-relevant stimulus properties can be set by plainverbal instruction without resorting to the direct applica-tion of reward, which makes it easier to separate task-and reward-relevant top–down settings (Boehler et al.,2011).
With the following two experiments, we aim at suchparadigmatic separation between reward relevance andfeature-based attention. Specifically, we ask how top–down settings that define reward relevance (a reward-defining feature) but not task relevance (a target-definingfeature) influence sensory processing in extrastriate cor-tex. To this end, we will use a simple visual search taskrequiring participants to search for a target defined bycolor (top–down settings for a task-relevant feature),whereas the combined occurrence of a specific task-irrelevant color with the target color will be associatedwith reward (top–down settings for reward relevance).To assess the degree of passive sensory biasing for theselection of those colors in visual cortex, we analyzedthe neuromagnetic brain response elicited by the suddenonset of a task-irrelevant color probe at performance-irrelevant time points between search frame presenta-tions. The probe was drawn either in the target color,the reward color, or a control color. Additionally, we alsoassessed the biasing effect of reward color during targetselection by analyzing the brain response elicited by thesearch frames as a function of the presence/absence ofthe reward color in the search distractor.
Electrophysiological signatures of attentional capturehave been documented to appear as amplitude modula-tions of early sensory visual components (Fukuda & Vogel,2009; Eimer &Kiss, 2008; Leblanc, Prime, & Jolicoeur, 2008;Lien, Ruthruff, Goodin, & Remington, 2008; Hickey,McDonald, & Theeuwes, 2006; Hopfinger & Ries, 2005;Arnott, Pratt, Shore, & Alain, 2001; Hopfinger & Mangun,1998), with the underlying current source activity originat-
ing from extrastriate visual cortex areas (Hopf, Boelmans,Schoenfeld, Luck, & Heinze, 2004; Hopf, Vogel, Woodman,Heinze, & Luck, 2002;Hopf et al., 2000;Martinez et al., 1999;Heinze et al., 1994).Moreover, several studies have reportedan N2pc component in response to attentional capturecontingent on distractors sharing task-relevant featureswith the target. The N2pc is known to index attentionalfocusing (Ansorge, Kiss, & Eimer, 2009; Eimer & Kiss,2008; Kiss, Jolicoeur, Dellʼacqua, & Eimer, 2008; Leblancet al., 2008) and has been shown to arise from sourceactivity in ventral extrastriate cortex based on event-relatedmagnetic field (ERMF) recordings (Hopf et al., 2000, 2002,2004). Here we will focus on the ERMF response elicited bytask-irrelevant color probes and analyze whether modu-lations in visual extrastriate visual cortex differs when com-paring target and reward probes relative to control probes.We will further investigate whether above reviewed sig-natures of attentional capture can be identified in theERMF response to search frames where the reward-relevant color is presented in the distractor.
METHODS
Participants (Experiment 1)
Twenty participants (mean age = 26.1 years, range = 21–32 years, 14 women) took part in Experiment 1. Theexperiment was undertaken with the understanding andwritten consent of the participants. All participants werestudents of the Otto-von-Guericke University Magdeburg,were right-handed, had normal color vision, and hadnormal or corrected-to-normal visual acuity. Participantswere paid for participation in addition to the payoff thatcould be gained on rewarded trials. All experiments wereapproved by the ethics review board of the Otto-von-Guericke University Magdeburg.
Stimuli and Procedure (Experiment 1)
Search arrays, probe stimuli, and the stimulation protocolare illustrated in Figure 1. Each search array containeda fixation cross and two double-colored 3-D spheres(diameter 2.7° visual angle), one in the left and one inthe right visual field (VF; 5° lateral, 2.7° downward fromfixation), presented on a homogenous gray background(luminance: 24 cd/m2). Each sphere was composed oftwo colors taken from a set of five colors (green, red,blue, yellow, gray), which were randomly assigned tothe left and right half of the spheres. On each trial, oneof the four half-spheres was drawn in the target color(green in Figure 1), with three of the other four colorsbeing assigned to the remaining half-spheres in thefollowing way. All half-spheres of a given search arraywere always drawn in a different color. On one third ofthe trials the target color was combined (randomly) withone of the three control colors (red in top search frameof Figure 1) with the distractor sphere containing a
1050 Journal of Cognitive Neuroscience Volume 26, Number 5

combination of the remaining control colors (yellow,gray). On another third of the trials, the target colorwas combined with one of the control colors (yellow inmiddle search frame of Figure 1), with one half-sphere ofthe distractor sphere being drawn in the reward color(blue). On the last third of the trials, the target colorwas combined with the reward color (green and blue inbottom search frame of Figure 1), and the distractorsphere contained a combination of two distractor colors(red and yellow). Importantly, reward was delivered onlyon the one third of the trials when the reward color wascombined with the target color. When the reward colorwas combined with a control color in the VF opposite tothe target (middle search frame in Figure 1), no rewardwas given. Note that the number of rewarded trials wasset to a comparably low proportion of one third of thetrials. This was done to de-emphasize the tendency thatwith an increasing proportion of rewarded trials partici-pants might take the specific combination of target andreward color as defining the target. Such combined target
definition would clearly undermine the present aim todissociate task and reward relevance. The participantsʼtask was to report whether the target color appearedon the left or right side of the target sphere with atwo-alternative button press of the right hand (indexfinger: left, middle finger: right) as fast and accurate aspossible.
Search arrays were presented for 700 msec. Onrewarded trials, a feedback frame was shown for 400 msec,1000–1500 msec after search frame offset, informing theparticipant whether reward was gained or not. The ISIbetween an unrewarded search frame or the feedbackframe and the next stimulus onset was randomizedbetween 1000 and 1500 msec (rectangular distribution).Randomly on 50% of the trials, this subsequent stimuluswas a single square (the probe, 1.8° × 1.8°, ISI 1000–1500 msec) that was flashed for 50 msec in the left or rightVF (centered at the location of the sphere). After suchtrials, the next search array appeared 600–900 msec afterthe probe. A probe was never directly followed by anotherprobe, that is, a probe appeared on average on everysecond trial of an experimental block. Probe color andthe VF of probe presentation were randomly assigned,such that the probe appeared as often in the VF of thesubsequent search target or in the opposite VF.
Experimental blocks were designed with target andreward color counterbalanced. That is, each color servedas the target in four blocks, with every other color servingas reward color on one of the four blocks. Throughout theentire experimental session, each color served four timesas reward color. During one experimental block (duration=5 min, one break), 72 search frames and 36 probes werepresented, yielding a session total of 60 trials per probecondition (target, reward, control) and VF (left, right) foreach participant. The luminance of the different probecolors was matched using heterochromatic flicker photom-etry (Lee, Martin, & Valberg, 1988) in three selected partic-ipants. Respective luminance values were used for allparticipants that participated in Experiments 1 and 2.
Reward Schedule
The payoff for a rewarded trial was 5 euro cents, whichamounted to a total bonus payment of approximatelyA10.70–12.00 per participant. Reward could be earnedupon correct target discrimination on trials where thetarget sphere also contained a reward color.
Participants (Experiment 2)
Sixteen participants (mean age = 26.5 years, range = 23–41 years, 14 women) participated in Experiment 2. Allparticipants were students of the Otto-von-Guericke Uni-versity Magdeburg, were right-handed, had normal colorvision, and had normal or corrected-to-normal visualacuity. Participants were paid for participation in additionto the payoff that could be gained on rewarded trials. All
Figure 1. Structure of an experimental trial block of Experiment 1.An instruction frame informed participants about the target-definingcolor (here green) and the color associated with reward (here blue)when combined with the target color. On each search frame (theframes containing two double-colored spheres), participants had todiscriminate the orientation of the convexity of the target half-sphere(left, right). Participants had to ignore the colored squares (colorprobes) flashed randomly between the search frames. The probesappeared randomly in the left and right VF and could be drawn in thetarget-defining color (target probe), the color associated with reward(reward probe), or one of the irrelevant colors (control probe(here gray, yellow, and red).
Buschschulte et al. 1051

experiments were approved by the ethics committee ofthe Otto-von-Guericke University Magdeburg.
Stimuli and Procedure (Experiment 2)
Stimuli and procedure were similar to Experiment 1,except for the following modifications. (1) As in Experi-ment 1, each color served as the target color with allother colors serving one time as the reward color, how-ever, under two different reward conditions (high-rewardblocks: 10 euro cents, low-reward blocks: 5 euro cents perrewarded trial). (2) As this doubled the number of experi-mental blocks per set and participant in comparison withExperiment 1, each participant performed two experi-mental sessions on different days (maximum delay be-tween sessions 40 days). During the first session, halfof the participants performed high-reward blocks onuneven block numbers and low-reward blocks on evenblocks numbers. The other half of the participants wasassigned to high- and low-reward blocks in the reverseorder. During the second session, the assignment ofreward magnitude to block number was reversed ineach participant, such that—with both sessions takentogether—participants performed every target–rewardcombination under high- and low-reward conditions.Again, across both sessions, each participant performed60 trials per probe condition (target, reward, control),VF (left, right), and reward magnitude (high, low). Par-ticipants were informed at the start of each block aboutwhether high or low reward will be delivered. The range ofpayoff gained from rewarded trials amounted to A15.85–18.00 per participant.
Data Recording and Analysis
The magnetoencephalogram (MEG) was recorded in amagnetically shielded chamber with a BTI Magnes 3600WH whole-head magnetometer system (4D Neuro-imaging, San Diego, CA) with 248 sensors. For control-ling eye movements, the EOG was recorded with ahorizontal and vertical montage using bipolar electrodeplacements at the temples (horizontal) as well as a uni-polar electrode below the left eye (vertical). Impedanceswere kept below 5 kΩ (EEG cap and Abralyt light gel,Easycap, Herrsching, Germany). EOG signals were ampli-fied with an EPA-6 amplifier (Sensorium, Inc., Charlotte,VT). The MEG and EOG signals were band-pass filtered(0.01–50 Hz) and digitized with a sampling rate of254.31 Hz. In addition to recording the EOG, fixation per-formance was continuously monitored with a custom-made zoom lens infrared camera system.
To coregister anatomical and functional data, anatomicallandmarks (nasion and left and right preauricular points)were digitized with a Polhemus 3Space Fastrak system(Polhemus Inc., Colchester, VT). These landmarks werethen brought into reference with magnetic marker fields
generated by five coils fixated on defined positions onthe EEG cap.Primary MEG data analysis included off-line artifact
rejection applied to epochs of interest with peak-to-peakamplitudes exceeding a threshold of 2–3 pT. For eachparticipant, epochs of interest ranging 200 msec before(baseline) to 750 msec after probe onset were extractedaccording to the relevant experimental conditions.Epochs containing peak-to-peak EOG amplitudes exceed-ing 150 μV were considered to contain eye motion arti-facts and were rejected from further data analysis. Thisresulted in an average rejection rate of 18.0% for Experi-ment 1 and 13.2% for Experiment 2, with no significantdifference between experimental conditions. For Experi-ment 1, averages were computed for the three differentprobe types (target, reward, control probes) in the leftand right VF, thereby collapsing data across the differentcolors. In addition, selective averages were computed forERMF responses elicited by the search frames (spheres)as a function of whether the reward color was present orabsent in the distractor opposite to the search target. Forthis analysis, only nonrewarded trials were considered.For Experiment 2, the same averages were computed,but separately for high- and low-reward blocks, and aftercollapsing the data from the two experimental sessions ofa given participant. For subsequent data analysis andvisualization, the ERP software ERPSS (Event-RelatedPotential Laboratory, University of California, San Diego,La Jolla, CA) was used. The statistical validation of wave-form differences was performed using a repeated-measuresANOVA (rANOVA) approach. If necessary, violations of datasphericity were corrected (Greenhouse–Geisser epsilon).Respective data will be reported with the original degreesof freedom, but with an adjusted level of significance( p value). The statistical validation of onset latency dif-ferences was performed using a sliding t-test approachapplied to subsequent time samples (window width of20 msec; Guthrie & Buchwald, 1991). The first sampleshowing a significant difference in a sequence of at leastthree subsequently significant time samples was taken tomark the onset latency.
Source Localization Analysis
Current source localization was performed by computingminimum norm least squares estimates (MNLS, sLORETA)as implemented in the CURRY V6.0 neuroimaging soft-ware (Compumedics Neuroscan, Charlotte, NC). Esti-mates for the grand averaged data were computed byusing realistic anatomical models of the source com-partment (3-D surface segmentation of the cortical graymatter space) and the volume conductor (3-D surfacesegmentation of the cerebrospinal fluid space) of thestandard MNI brain (Montreal Neurological Institute).Before averaging waveforms across participants, sensordata of each participant were coregistered to a referencesubject based on the participantʼs individual landmarks.
1052 Journal of Cognitive Neuroscience Volume 26, Number 5

The grand averaged sensor data were then brought intoreference with the anatomical data of the MNI brain.
RESULTS
Experiment 1
Behavioral Performance as a Function ofProbe Color and Probe Location
Participants were to ignore the probes, so that no behav-ioral performance measure directly reflecting probe colorprocessing can be obtained. However, the effect of probecolor can be analyzed indirectly by determining theamount of exogenous spatial cuing (for the performance
on a subsequent search frame) created by the probes.That is, the amount of cuing produced by the differentprobe types will provide an index of the degree to whichspatial attention was captured. To assess the effect ofexogenous cuing, target discrimination performancewas analyzed as a function of whether a given probeappeared on the side of the subsequently appearingtarget sphere (valid probes) versus on the opposite side(invalid probe). Given that the probe target SOA variedbetween 600 and 900 msec, the expected cuing effectis one of inhibition of return (IOR; Klein, 2000; Maylor& Hockey, 1985; Posner & Cohen, 1984), that is, a rela-tive slowing of the response for valid probes, which wasin fact observed for all three probe conditions. As shownin Figure 2A, valid probes led to an average of 11 msecslower response than invalid probes. Importantly, IORdid not vary with probe type indicating that spatial atten-tion was captured by the probe to the same extent in allthree probe conditions. A two-way rANOVA with the fac-tors Probe Condition (target, reward, control) and ProbeValidity (valid, invalid) confirmed this impression. Therewas a significant main effect of Probe Validity, F(1, 19) =19.9, p < .0001, but no Probe Validity × Probe Conditioninteraction, F(2, 38) = 0.56. RTs were slightly slower fortargets following reward probes versus target or controlprobes, but the main effect of Probe Condition did notreach significance, F(2, 38) = 2.76, p = .09. Figure 2Bsummarizes response accuracy (RA; % correct responses).Consistent with IOR, accuracy was generally lower forvalid than for invalid trials, with this effect not beingdifferent for the three probe conditions. A respectiverANOVA yielded a significant main effect of Probe Valid-ity, F(1, 19) = 4.87, p < .05, but no interaction of ProbeValidity × Probe Condition, F(2, 38) = 0.03. Again, therewas a trend toward a main effect of Probe Condition, F(2,38) = 2.83, p = .08, reflecting that performance accuracywas slightly reduced after reward probes relative to targetand control probes. It should be noted that—althoughnot significant—the general pattern of responses hintsat a possible effect of reward on performance in form ofan increased IOR effect. However, given that the overallIOR effect is already small and not significant, strong inter-pretations may not be warranted.
MEG Responses to the Color Probes
Figure 3A shows ERMF waveforms and distributions elic-ited by the target probes, reward probes, and controlprobes at representative selected sensor sites (arrows)over the left and right occipito-temporal cortex contra-lateral to the VF of probe presentation. Shown are wave-forms and field distributions separately for right (toprow) and left VF probes (bottom row). As visible, targetprobes (red traces) led to a response enhancement rela-tive to control probes (black traces) between ∼180 and280 msec after probe onset. No such enhancement
Figure 2. Target discrimination performance as a function of probe-location in Experiments 1 and 2. (A) Mean RT and (B) percent correctvalues for targets following color probes in the same VF as the probe(valid) or in the opposite VF (invalid) in Experiment 1. Data are shownseparately for probes drawn in the target color (target), the rewardcolor (reward), and the control color (control). (C) Mean RT and (D)percent correct responses of Experiment 2. Data are shown separatelyfor the different probe types as well as the low- (low) and the high-reward condition (high). White bars index the standard error of mean.
Buschschulte et al. 1053
christianstoppel
Hervorheben

appeared for reward probes (green traces), for which theresponse remained almost indistinguishable from the re-sponse to control probes. This difference in responsestrength is further illustrated in the topographical maps(right) showing the average ERMF response between180 and 280 msec for the three probe conditions. Theresponse in visual cortex to the probe appears asefflux–influx configuration (encircled by black ellipses)over the lateral occipito-temporal cortex contralateral tothe VF of probe presentation. Note that the efflux andinflux component are shown in red and blue, respec-tively. Both together form the magnetic field effect thatis generated by source activity (see Figure 3B) locatedunder the transition zone (asterisks) between the effluxand influx component. The field effect is stronger fortarget probes than reward and control probes, with thelatter two showing effects of comparable size. For statis-tical validation, a three-way rANOVA with the factor
Probe Condition (target, reward, control) was performedon the mean ERMF response between 180 and 280 msec.Data from right and left VF and VF probes were analyzedseparately at corresponding contralateral sensor sitesover the left and right hemisphere, respectively (arrowsin Figure 3A). This analysis yielded a significant main effectof Probe Condition for both left and right VF probes (rightVF probes (sensor A196): F(2, 38) = 5.2, p < .01; left VFprobes (sensor A245): F(2, 38) = 4.66, p < .05). Sub-sequent pairwise comparisons (paired-samples t tests)showed that target probes differed significantly fromcontrol probes (right VF probes: p= .0048, left VF probes:p = .014) and from reward probes (right VF probes: p =.018, left VF probes: p= .041), whereas reward and controlprobes did not differ (right VF probes: p = .68, left VFprobes: p = .29).Figure 3B summarizes the results of the current source
density (CSD) analysis of the ERMF response to the three
Figure 3. MEG response to color probes of Experiment 1. (A) ERMF waveforms and distributions elicited by the different probe types in the right(top row) and left VF (bottom row). The shown waveforms were recorded at the sensor sites indicated by the small arrows. The black ellipseshighlight the efflux–influx field configuration representing the response elicited by the probe. (B) Source waveforms and CSD distributions estimatedfor the data shown in A. Source waveforms represent the time course of source density estimates in the cortical regions centered at the sourcedensity maximum highlighted by white small circles.
1054 Journal of Cognitive Neuroscience Volume 26, Number 5

probe types separately for probes appearing in the right(top row) and left VF (bottom row). The topographicalmaps show the CSD distribution (MNLS estimates, seeMethods) overlaid onto a 3-D surface segmentation ofthe cortical gray matter layer of the MNI brain (averageof 152 T1-weighted stereotaxic volumes of the ICBM proj-ect, ICBM152), which served as source compartment forthe computation. CSD maxima of the three probe typesappear in similar regions of the inferior occipito-temporalcortex contralateral to the VF of probe presentation. Inlinewith what is visible in the field distributions (Figure 3A),maximum CSD estimates are seen for target probes,whereas reward and control probes display comparablebut smaller effect sizes. This is further illustrated by thetime course of CSD estimates obtained from ROIs centeredat the CSD maximum of all three probe conditions (whiteellipses in Figure 3B). Starting at ∼180 msec after probeonset, the CSD of target probes (red) increases beyondthat of reward (green) and control probes (black) until∼280 msec.
Behavioral Performance as a Function of Reward ColorLocation in the Search Frames
Figure 4 (A, B) summarizes the RT and RA measures inExperiment 1 as a function of where the reward colorwas presented in the search frames (in the target, inthe distractor, absent). Apparently, RT is slowed whenthe reward color appeared with the distractor relative
to when it was presented with the target or was absent.A rANOVA with a three-level factor reward location vali-dates this impression by showing a significant effect, F(2,38) = 30.7, p < .0001. Post hoc pairwise comparisonsconfirm that the effect is because of increased RT for thereward color presented with the distractor relative to theother conditions (reward color in target vs. in distractor:p < .0001; reward color absent vs. in distractor: p <.0001). Notably, participants were not faster for trials wherethe reward color was combined with the target than forreward color absent trials ( p= .112). Reward color did alsoinfluence RA in a significant way, F(2, 38) = 11.3, p< .005.Although RA was generally high, the presence of rewardcolor caused a small decrease in performance relative toreward color absent trials. Post hoc pairwise comparisonsconfirm that performance on reward color absent trials wasbetter than when the reward color was with the target ( p<.005) or with the distractor ( p < .0001). Participants werealso more accurate when the reward color appeared in thetarget versus in the distractor, but this effect was not sig-nificant ( p = .102).
MEG Responses to the Search Frames (Spheres)
Figure 5A shows the response to the spheres of non-rewarded trials as a function of whether the reward colorwas presented in the nontarget VF (green traces) or not(black traces). Displayed are waveforms from a selectedsensor site showing the maximum modulatory effect over
Figure 4. Target discriminationperformance as a function ofreward color location in thesearch frames of Experiments 1and 2. (A) Mean RT and(B) percent correct responsesfor the reward color presentedin the target or in the distractoror not presented at all inExperiment 1. (C) Mean RTand (D) percent correctresponses of Experiment 2.Data are separately shownfor the low- (low) and thehigh-reward condition (high).White bars index the standarderror of mean.
Buschschulte et al. 1055

the hemisphere contralateral to the nontarget VF. Appar-ently, the reward color elicits a response enhancementstarting around 200msec after search frame onset. rANOVAswith the factor Reward Color (present/absent in non-target VF) on mean amplitude measures between 200and 280 msec confirm this observation by yielding sig-nificant effects for the right VF, F(1, 19) = 10.09, p <.005, as well as the left VF, F(1, 19) = 8.3, p < .01.The topographical maps show the results of the currentsource localization analysis computed for the ERMF
difference reward color present minus absent in the un-attended VF. Current source maxima appear in ventrallateral extrastriate visual cortex, contralateral to the sideof reward color presentation. Hence, in contrast to thepresentation of task-irrelevant probes where rewardcolor had no effect on extrastriate processing, present-ing the reward-relevant color in a task-irrelevant distrac-tor during target selection in visual search producedenhanced responses. This indicates that reward rele-vance did in fact bias feature processing during the selec-tion of task-relevant features when actually performingthe visual search task.
Experiment 2
Experiment 1 failed to detect any difference in the ventralextrastriate response to color probes when they wereassociated with reward as compared with control probes.However, the amount of monetary reward to be gainedon a given trial of Experiment 1 was moderate (5 eurocents), and it is possible that the absence of modulatoryeffects reflects the fact that reward was not of adequatesignificance to the observer to generate the effect ofinterest. Experiment 2 addressed this possibility bydoubling the amount of money (10 euro cents) to begained on rewarded trials. Specifically, participantsperformed two types of trial blocks in which the amountof reward was either as high as in Experiment 1 (low-reward blocks) or doubled (high-reward blocks). Withrespect to stimulation, trial structure, and experimentaltask, Experiment 2 was identical to Experiment 1.
Behavioral Performance as a Function of Probe Colorand Probe Location
Figure 2C shows that, for all probe types and reward con-ditions, valid probes were associated with IOR, that is,with a slowing of RT relative to invalid probes. RT wasalso generally faster in high-reward blocks than in low-reward blocks. A three-way rANOVA with the factorsProbe Validity (valid, invalid), Probe Condition (target,reward, control), and Reward Size (low, high) confirmedthat there were significant main effects of Probe Validity,F(1, 15) = 43.2, p < .0001, and Reward Size, F(1, 15) =13.8, p< .005, but no main effect of Probe Condition, F(2,30) = 1.1, p < .4. There were no significant interactions(Reward Size × Probe Condition, F(2, 30) = 1.4, p < .3;Probe Validity × Probe Condition, F(2, 30) = 2.0, p <.15; and Reward Size × Probe Validity, F(1, 15) = 0.54,p < .5). Inspection of the RA in Figure 2D revealed aminimal variation because of the experimental conditionswithout any systematic effect of Probe Validity, ProbeCondition, or Reward Size. This is confirmed by a rANOVAshowing that neither of the respective main effects wassignificant (Probe Validity: F(1, 15) = 0.29, p < .6; ProbeCondition: F(2, 30) = 1.03, p < .4; reward size: F(1, 15) =0.49, p < .5).
Figure 5. MEG response to search frames of Experiments 1 and 2.(A) Experiment 1: ERMF waveforms elicited by nonrewarded searchframes, with the reward color being present (green) or absent (black)in the distractor sphere. The waveforms are shown for selected sensors(RH/LH sensor = right/ left hemisphere sensor) contralateral to theVF of distractor presentation. The topographical maps above displaythe corresponding CSD distributions estimated for the reward-minus-control difference. (B) Experiment 2: ERMF waveforms for reward colorpresent (green) and absent in the distractor VF (black), separatelyshown for the low- (top) and high-reward condition (bottom). Thetopographical maps display the CSD distributions estimated for thereward-minus-control difference of the low-reward condition.
1056 Journal of Cognitive Neuroscience Volume 26, Number 5
christianstoppel
Hervorheben

MEG Responses to the Color Probes
Figure 6 shows waveforms and ERMF distributions ofthe three different probe conditions of the low- (a) andhigh-reward condition (b) for probes in the right VF(qualitatively similar effects were observed for the leftVF). For the low-reward condition, the ERMF responseto the three probe types perfectly reproduces the patternseen in Experiment 1. The response to target probes isincreased in ventral extrastriate cortex between ∼180 and270 msec relative to reward and control probes, with thelatter being nearly indistinguishable. A similar picture isseen for the high-reward condition between 190 and220 msec (Figure 6B). Although the size of the responseto the probes is generally smaller than under low-rewardconditions, target probes, again, show an increased re-sponse relative to reward and control probes, whereasthe response elicited by reward probes does not differfrom control probes. In a subsequent time range (∼220–250 msec), however, the response to reward probes isreduced relative to target and control probes. This is alsovisible in the corresponding ERMF maps showing thedistribution of the mean probe response for the three
probe types of the low-reward condition at 200 msec(Figure 6A, right), as well as the high-reward condition at200 msec (top row maps in Figure 6B) and at 230 msec(bottom row maps in Figure 6B). At 200 msec after probeonset, the response to the three probe types shows apretty similar field distribution and amplitude pattern forthe low- and high-reward condition. That is, the efflux–influx configuration (ellipses) elicited by target probes overthe left lateral occipito-temporal cortex is stronger than theones elicited by reward and control probes. Notably, thepattern changes around 220 msec after probe onset, wheretarget and control probes show a field response of similarsize while the response to reward probes is significantlyreduced.
Figure 7 shows the results of the source localization anal-ysis (source density estimates, right VF probes only) per-formed on the mean target-minus-control differencebetween 190 and 220 msec of the low- (top) and high-reward condition (bottom). Apparently, current sourcemaxima (highlighted by white and green dots) appear overthe left ventral-lateral occipito-temporal cortex consistentwith the localization of current source maxima in Experi-ment 1 (cf. Figure 3B, top row). The reward-minus-control
Figure 6. MEG response to color probes of Experiment 2. Waveforms and distributions elicited by the three different probe types in the right VFunder low- (A) and high-reward conditions (B). The presented waveforms were recorded at the sensor sites indicated by the small arrow in A andcorrespond with the sensor site shown in Figure 3A. The topographical maps on the right side show the distribution of the ERMF response at thetime points indicated by the black arrowheads in the corresponding waveform displays. The black ellipses highlight the efflux–influx fieldconfiguration representing the probe-elicited response.
Buschschulte et al. 1057
christianstoppel
Hervorheben
christianstoppel
Hervorheben
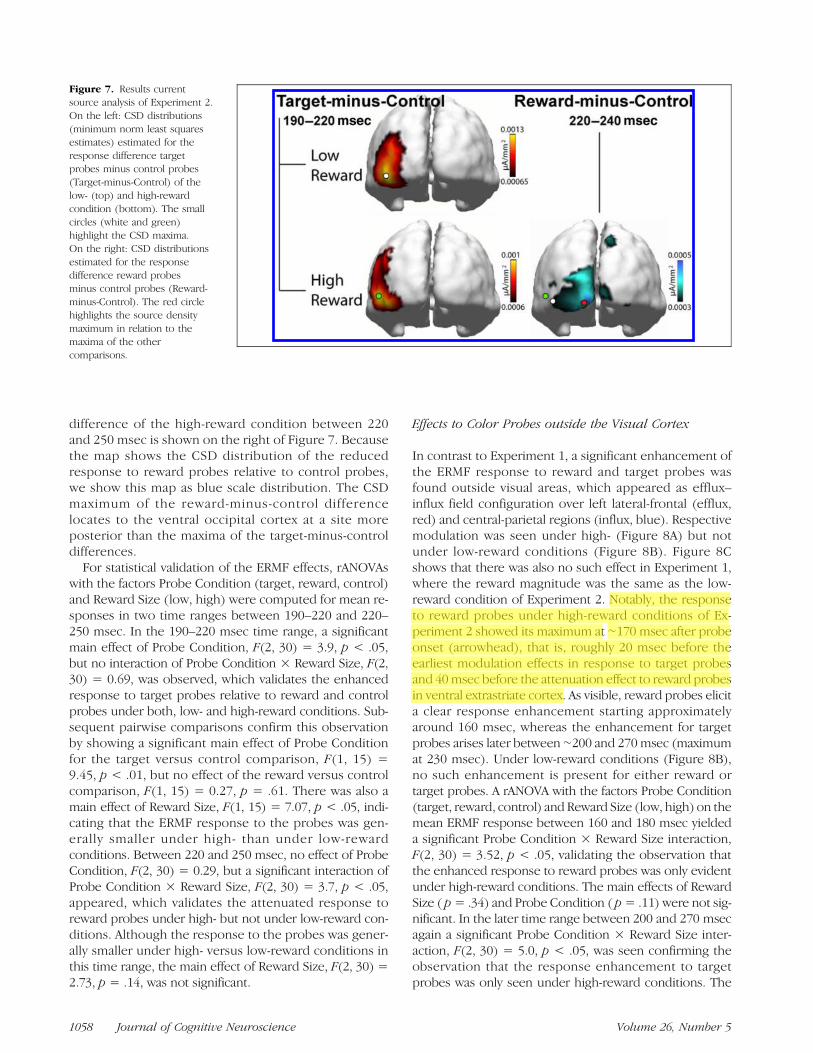
difference of the high-reward condition between 220and 250 msec is shown on the right of Figure 7. Becausethe map shows the CSD distribution of the reducedresponse to reward probes relative to control probes,we show this map as blue scale distribution. The CSDmaximum of the reward-minus-control differencelocates to the ventral occipital cortex at a site moreposterior than the maxima of the target-minus-controldifferences.
For statistical validation of the ERMF effects, rANOVAswith the factors Probe Condition (target, reward, control)and Reward Size (low, high) were computed for mean re-sponses in two time ranges between 190–220 and 220–250 msec. In the 190–220 msec time range, a significantmain effect of Probe Condition, F(2, 30) = 3.9, p < .05,but no interaction of Probe Condition × Reward Size, F(2,30) = 0.69, was observed, which validates the enhancedresponse to target probes relative to reward and controlprobes under both, low- and high-reward conditions. Sub-sequent pairwise comparisons confirm this observationby showing a significant main effect of Probe Conditionfor the target versus control comparison, F(1, 15) =9.45, p < .01, but no effect of the reward versus controlcomparison, F(1, 15) = 0.27, p = .61. There was also amain effect of Reward Size, F(1, 15) = 7.07, p < .05, indi-cating that the ERMF response to the probes was gen-erally smaller under high- than under low-rewardconditions. Between 220 and 250 msec, no effect of ProbeCondition, F(2, 30) = 0.29, but a significant interaction ofProbe Condition × Reward Size, F(2, 30) = 3.7, p < .05,appeared, which validates the attenuated response toreward probes under high- but not under low-reward con-ditions. Although the response to the probes was gener-ally smaller under high- versus low-reward conditions inthis time range, the main effect of Reward Size, F(2, 30) =2.73, p = .14, was not significant.
Effects to Color Probes outside the Visual Cortex
In contrast to Experiment 1, a significant enhancement ofthe ERMF response to reward and target probes wasfound outside visual areas, which appeared as efflux–influx field configuration over left lateral-frontal (efflux,red) and central-parietal regions (influx, blue). Respectivemodulation was seen under high- (Figure 8A) but notunder low-reward conditions (Figure 8B). Figure 8Cshows that there was also no such effect in Experiment 1,where the reward magnitude was the same as the low-reward condition of Experiment 2. Notably, the responseto reward probes under high-reward conditions of Ex-periment 2 showed its maximum at ∼170 msec after probeonset (arrowhead), that is, roughly 20 msec before theearliest modulation effects in response to target probesand 40msec before the attenuation effect to reward probesin ventral extrastriate cortex. As visible, reward probes elicita clear response enhancement starting approximatelyaround 160 msec, whereas the enhancement for targetprobes arises later between ∼200 and 270 msec (maximumat 230 msec). Under low-reward conditions (Figure 8B),no such enhancement is present for either reward ortarget probes. A rANOVA with the factors Probe Condition(target, reward, control) and Reward Size (low, high) on themean ERMF response between 160 and 180 msec yieldeda significant Probe Condition × Reward Size interaction,F(2, 30) = 3.52, p < .05, validating the observation thatthe enhanced response to reward probes was only evidentunder high-reward conditions. The main effects of RewardSize ( p= .34) and Probe Condition ( p= .11) were not sig-nificant. In the later time range between 200 and 270 msecagain a significant Probe Condition × Reward Size inter-action, F(2, 30) = 5.0, p < .05, was seen confirming theobservation that the response enhancement to targetprobes was only seen under high-reward conditions. The
Figure 7. Results currentsource analysis of Experiment 2.On the left: CSD distributions(minimum norm least squaresestimates) estimated for theresponse difference targetprobes minus control probes(Target-minus-Control) of thelow- (top) and high-rewardcondition (bottom). The smallcircles (white and green)highlight the CSD maxima.On the right: CSD distributionsestimated for the responsedifference reward probesminus control probes (Reward-minus-Control). The red circlehighlights the source densitymaximum in relation to themaxima of the othercomparisons.
1058 Journal of Cognitive Neuroscience Volume 26, Number 5
christianstoppel
Hervorheben

corresponding main effects were not significant (probecondition: p = .28, reward size: p = .37). The prior re-sponse enhancement to reward probes relative to targetprobes visible in Figure 8A is notable and warrants fur-ther validation. To this end, we computed sliding windowt tests (see Methods) on the reward-minus-control andthe target-minus-control differences, which revealed re-sponse onsets for reward and target probes at 155 and190 msec, respectively.Figure 8D shows results of the source localization anal-
ysis (LORETA estimates, see Methods) computed for thereward-minus-control ERMF difference at 170 msec andoverlaid onto transsections of the MNI brain. The cor-responding source density maximum is located to a med-ial frontal cortex area of the dorsal ACC (dACC). Sourcewaveforms (Figure 8E) of the reward-minus-control (bluetrace) and the target-minus-control difference (red trace)taken from the CSD maximum (dACC) show that themodulation in response to reward probes appears earlierthan the response to target probes. Furthermore, in linewith the ERMF waveform effects, the response to rewardprobes in dACC arises prior (∼20 msec) to the activityenhancement because of target probes in ventral extra-striate cortex shown in Figure 8F (see difference betweendashed lines). In contrast, the anterior cingulate re-sponse enhancement to target probes (red trace in Fig-ure 8E) did not arise before the response in ventralextrastriate cortex (Figure 8F). Given that the responsein dACC to reward probes under high-reward conditionsappeared before the attenuation of the ERMF response in
ventral extrastriate visual cortex, it is reasonable to askwhether the latter is linked to activity changes in dACC.Such a direct modulatory influence of frontal lobe activityon processes of attentional selection in visual cortex hasbeen has been repeatedly documented (Noudoost &Moore, 2011; Cohen, Heitz, Schall, & Woodman, 2009;Moore & Armstrong, 2003; Moore, Armstrong, & Fallah,2003). To address this possibility, we analyzed the amountof ERMF attenuation as a function of the amplitude andlatency variation in dACC across participants. Specifically,we assessed the correlation between peak amplitude/peaklatency measures of the response (reward-minus-controlERMF difference) to reward probes in dACC and the meanresponse attenuation between 220 and 250 msec in ventralextrastriate cortex. This analysis revealed no correlationbetween amplitude measures (r = 0.213, t(16) = 0.816),but we found a significant correlation between responselatency in dACC and the amplitude reduction in ventralextrastriate cortex (r = 0.426, t(16) = 1.76, p < .05). Thescatterplot in Figure 9 illustrates this relation. It shows thatparticipants with progressively shorter dACC latenciesdisplayed an increasing effect of attenuation of the ERMFresponse to reward probes in extrastriate visual cortex.
Behavioral Performance as a Function of Reward ColorLocation in the Search Frames
Figure 4 (C, D) shows RT and RA measures as a functionof the location of the reward color in the search arrayseparately for the low- and high-reward condition of
Figure 8. MEG response tocolor probes outside the visualcortex (Experiments 1 and 2).ERMF waveforms elicited bythe three different probe typesoutside the visual cortex forthe (A) high-reward conditionof Experiment 2, (B) thelow-reward condition ofExperiment 2, and (C) ofExperiment 1. The smallarrow in A highlights thesensor over the parietal influxmaximum—the site wherethe shown waveforms wererecorded from. The coloredhorizontal bars index the timerange of significant amplitudedifferences between rewardand control probes (blue)and between target andcontrol probes (red).(D) CSD estimates (sLORETA)of the reward-minus-controldifference at the time pointhighlighted in (A; blackarrowhead). (E) Normalizedsource waveforms measured from the source density maximum shown in D of the reward-minus-control ERMF difference (blue) and thetarget-minus-control difference (red). (F) Normalized source waveform corresponding with the ventral extrastriate CSD maximum of thetarget-minus-control difference of the high-reward condition (green circle in Figure 7).
Buschschulte et al. 1059
christianstoppel
Hervorheben
christianstoppel
Hervorheben
christianstoppel
Hervorheben

Experiment 2. Consistent with the observation in Experi-ment 1, RTs are slowed when presenting the rewardcolor with the distractor relative to when reward is absentor presented with the target. In addition, RT is slightlyincreased when the reward feature is presented togetherwith target relative to the reward absent condition. Thispattern is seen for both the low- and high-reward condi-tion. A two-way rANOVA with the factors Reward Location(in target, in distractor, absent) and Reward Size (low,high) validates these observations by yielding a signifi-cant main effect of Reward Location, F(2, 30) = 23.2,p < .0001, but no interaction of Reward Location ×Reward Size, F(2, 30) = 0.49, p = .58. There was, how-ever, a significant main effect of Reward Size, F(1, 15) =12.3, p < .005, reflecting the fact that RT was generallyfaster on high-reward blocks. Pairwise post hoc compar-isons reveal that participants were significantly slower onthe reward in distractor condition relative to the rewardin target, F(1, 15) = 19.2, p < .005, and reward absentcondition, F(1, 15) = 27.9, p < .0001. The slight re-sponse slowing for the reward in target versus the rewardabsent condition was also significant, F(1, 15) = 13.6, p<.005, indicating that the presence of a reward color hadgenerally a distracting effect, which was largest whenappearing in the search distractor. This increased dis-tracting effect of reward color was also reflected by theRA pattern, as the reward in distractor condition showeda performance decrement relative to conditions wherethe reward color appeared in the target or was absent.This effect again did not vary with reward size. A respec-tive two-way rANOVA confirms this by showing a signifi-cant main effect of Reward Location, F(2, 30) = 7.6, p <.01, without showing a Reward Location × Reward Sizeinteraction, F(2, 30) = 0.86, p = .43. There was also nomain effect of Reward Size, F(1, 15) = 1.2, p = .29. Pair-wise post hoc comparisons revealed that RA was reduced
for the reward in distractor condition relative to the re-ward in target, F(1, 15) = 5.5, p < .05, and the rewardabsent condition, F(1, 15) = 9.5, p < .01, with the smalldecrement for the reward in target condition versus thereward absent condition being also significant, F(1, 15) =6.5, p < .05.
MEG Responses to the Search Frames (Spheres)
Figure 5B shows waveforms elicited by (nonrewarded)search frames with the reward color present (greentraces) or absent (black traces) in the nontarget VF.Results are separately shown for low- and high-rewardblocks. As in Experiment 1, on low-reward blocks thereward color elicited an enhanced response between200 and 260 msec after search frame onset. The cor-responding topographical map again reveals a currentsource maximum in ventral extrastriate cortex contra-lateral to the VF of reward color presentation. Most nota-bly, on high-reward blocks this modulatory effect isabolished, with the response to the reward color beingindistinguishable from the response to the control color.For statistical validation, a two-way rANOVA with the fac-tors Reward Color (present/absent in nontarget VF) andReward Size (low, high) was computed on mean ampli-tude measures between 200 and 260 msec after searchframe onset. This yielded a significant main effect ofReward Color, F(1, 15) = 6.09, p < .05, and a signifi-cant interaction between Reward Color and Reward Size,F(1, 15) = 4.62, p < .05, but no main effect of RewardSize, F(1, 15) = 0.08. Subsequent pairwise comparisonsrevealed a significant effect of Reward Color for low-reward trials, F(1, 15) = 7.49, p < .05, but no such effectfor high-reward trials, F(1, 15) = 0.11.In summary, after doubling the amount of money to be
gained on rewarded trials, reward probes did not elicit anincreased response in ventral extrastriate cortex as seenfor target probes. Instead, a delayed attenuation of theresponse to reward probes was observed in a more pos-terior ventral extrastriate region. In addition, reward andtarget probes elicited an increased response in medial-frontal/ACC under high-reward conditions, with the effectto reward probes arising before the effect to targetprobes as well as before the attenuation effect in ventralextrastriate cortex. A correlation analysis revealed that theamount of delayed attenuation to reward probes in pos-terior extrastriate visual cortex increased with shorterlatencies of the response in ACC, suggesting a functionallink between the speed of reward-representation in fron-tal cortex and the subsequent attenuation of sensory pro-cessing in visual areas. Finally, this observation dovetailswith the differential effect of reward size on the responseelicited by search arrays. The presentation of a rewardcolor in nontarget spheres led to a response enhance-ment under low-reward conditions as in Experiment 1.Doubling the amount of reward, however, eliminated thisenhancement effect.
Figure 9. Scatter diagram plotting the mean amplitude reductionbetween 220 and 250 msec in ventral extrastriate cortex against thepeak response latency in dACC of each participant (n = 16) observedfor the high-reward condition of Experiment 2. The line shows thelinear regression between measures.
1060 Journal of Cognitive Neuroscience Volume 26, Number 5

DISCUSSION
The reported experiments revealed that task-irrelevantcolor probes drawn in a target-defining color led to activ-ity enhancements between 180 and 280 msec in ventralextrastriate cortex contralateral to probe presentation.Probes drawn in a reward-relevant color, in contrast,did not show such response enhancement. Instead, in-creasing reward relevance by doubling the amount ofmoney to be gained on rewarded trials (Experiment 2)resulted in a delayed reduction (∼220-250 msec) of theresponse relative to control probes. Analyzing the effectof reward color presented during the visual search taskrevealed that presenting the reward-relevant color inthe distractor caused an activity enhancement startingaround 200 msec in ventral extrastriate cortex. This en-hancement was, however, abolished under high-rewardconditions of Experiment 2.These observations together suggest that reward-
contingent modulatory effects in extrastriate visual cortexare under effective top–down control even when thebehavioral significance of reward is separated from task-relevant feature descriptions defining the target. Underthe specific settings of the present experiments, reward-related biases of sensory processing in extrastriate visualcortex were eliminated or even suppressed by top–downinhibitory influences, with top–down inhibition becomingstronger with larger reward prospect. Such strong inhibi-tory control of distractor processing is consistent withrecent evidence from ERP recordings (Sawaki & Luck,2010, 2011). Respective studies showed that salient single-tons may cause an automatic (bottom–up) “attend-to-me”signal, but that the actual consequence of attentionalcapture may be counteracted by active suppression. Theexistence of such “attend-to-me” signal was inferred fromthe presence of a component called distractor positivity,an ERP modulation shown to reflect the suppression ofdistractors (Hickey, Di Lollo, & McDonald, 2009). Thegeneral attenuation of the probe-elicited response underhigh-reward conditions of Experiment 2 suggests that suchcounteracting suppression appeared to some extent for allprobe types. The delayed attenuation to reward probesunder high-reward conditions—presumably correspond-ing with an increased distractor positivity component—may then reflect extra suppression, which was of smalleramplitude under low-reward conditions, such that a posi-tive modulation bias, as seen for target probes, was justcancelled.Other observations in the present experiments as well
as in the literature lend support to this interpretation.First, the mere presence of the reward color led to aperformance decrement during target selection, indicat-ing that the reward color had a generally distractingeffect, even when combined with the target color. Sec-ond, RTs were overall faster under high- than low-rewardconditions of Experiment 2. Third, under high-rewardconditions an early response enhancement was seen in
dACC, which appeared before the response attenuationin ventral extrastriate cortex. A quantification of thiseffect revealed that the latency of the dACC response cor-related with the amount of attenuation in extrastriatecortex (Figure 9)—a relation consistent with dACC beinginvolved in the top–down (inhibitory) control of extra-striate sensory processing. In fact, fMRI research inhumans revealed that error-related activity modulationsin posterior medial frontal cortex correlate with the sup-pression of activity in visual cortex areas coding distract-ing features of the target input (Danielmeier, Eichele,Forstmann, Tittgemeyer, & Ullsperger, 2011). Further-more, it has been reported that reward-dependentchanges of stimulus saliency as well as the detectionand interpretation of reward are mediated in ACC (Hickeyet al., 2010; Hayden, Pearson, & Platt, 2009). Finally, directtop–down influences on extrastriate visual processinghave been documented in the monkey using differentmethodologies (Noudoost & Moore, 2011; Cohen et al.,2009; Moore & Fallah, 2004; Moore & Armstrong, 2003;Moore et al., 2003).
How then does the attenuation or absence of effects totask-irrelevant reward probes observed here reconcilewith previous studies showing that reward-dependentbiasing of sensory processing does appear in visual cortexand earlier stages even when the reward-defining featureis not task relevant (subject to discrimination), (Hickeyet al., 2010; Franko et al., 2009; Kiss, Driver, & Eimer,2009; Serences, 2008; Shuler & Bear, 2006; Ikeda &Hikosaka, 2003). Serences (2008) observed with fMRI inhuman observers that a history of color gratings beingassociated with reward that is independent of subjectivevaluation led to larger BOLD responses in early visual cor-tex areas (V1–V4). Franko et al. (2009) recorded localfield potentials from macaque V4 while the monkey pas-sively viewed gratings of different orientations. Orienta-tion was task-irrelevant, but the orientation consistentlypaired with the subsequent delivery of reward led toincreased LFP responses. Although these studies showpositive biasing effects of reward on visual selection fortask-irrelevant reward-defining features, they differ fromthe present experiments in a critical way, namely thatthe occurrence of a reward feature was consistently asso-ciated with subsequent reward delivery or the valuationof reward. In other words, the reward feature directedattention toward the subsequent event that deliveredor signaled the gain of reward without distracting perfor-mance in a negative way—a situation not giving explicitor implicit incentive to inhibit modulatory effects be-cause of the reward feature. In contrast, in the presentexperiments, the presentation of reward probes wascompletely nonpredictive of whether the subsequentsearch target would be combined with a reward color.Furthermore, the present experiments were designedto dissociate top–down definitions of reward and taskrelevance, with the biasing effect of reward being probedby distracting events during task-irrelevant phases of the
Buschschulte et al. 1061

experiment. These experimental conditions rendered anyoccurrence of the reward color a distracting event, evenwhen the reward color was combined with the targetcolor. In fact, the reward color impaired target discrimi-nation performance not only when presented in thesearch distractor but also when presented in the target.Hence, participants likely adopted a general set towarddeemphasizing reward feature selection, which is overallconsistent with the absence or the suppressive nature ofeffects elicited by reward probes.
Notable in this respect is recent research, which hasdocumented increased responses to task-irrelevant fea-tures even after dissociating their reward associationfrom endogenous attentional settings (Hickey et al.,2010). That is, positive neural biasing effects were foundunder conditions that should—according to the consid-erations above—deemphasize reward feature selection.For example, Hickey et al. (2010) had participants searchfor a shape singleton among similarly colored items andone color singleton distractor. Participants were givenhigh or low reward upon correct target discriminationon every trial. From trial to trial, color assignment tothe target and distractor changed or remained constant,such that, on a given trial, the target or the distractor waseither presented in the previously rewarded or the non-rewarded color. Importantly, color was completely taskirrelevant, and color discrimination was in fact counter-productive for performing the task. The critical observa-tion was that when color distractors appeared in thecolor that was combined with high-reward on a previoustrial, an enhanced P1 response was seen contralateral tothe location of the color distractor—an enhancement notseen when the same color was associated with low rewardon the previous trial. A P1 enhancement typically reflectsan effect of location selection associated with a gainamplification of neural processing in retinotopically cor-responding extrastriate visual areas (Hillyard et al.,1998). This suggests that reward relevance biased pro-cessing in extrastriate cortex, although color processingwas generally task-irrelevant and hindering target selec-tion (see Anderson, Laurent, & Yantis, 2011; Kristjansson,Sigurjonsdottir, & Driver, 2010, for converging observa-tions at the behavioral level). These results run counterto the present observations. Hickey et al. (2010) observedeffects of positive reward priming during target selectionin search arrays containing a distractor that carried thecolor associated with reward on the previous trial. Thatis, the priming effect in the neural response (P1 enhance-ment) emerged with increasing reward. We likewiseobserved an enhanced response to the reward-relevantcolor distractor in visual search, but the effect was insteadeliminated after increasing reward. We have no definitiveexplanation of the opposite response pattern, but thereare significant differences in experimental design pre-sumably causing different modes of top–down control.The strategic effects controlling the effect of rewardassignment in Hickey et al. (2010) were mainly reflecting
by trial-by-trial adjustments, because the color–rewardassociation as well as reward size varied constantly fromtrial to trial. In the present experiments, the color–rewardassociation and reward size were both fixed within trialblocks—conditions allowing for more consistent controlsettings, thereby permitting participants to adopt a stron-ger and more specific top–down inhibitory scheme tocounter the distracting effect of the reward color—hencethe general effect of response attenuation.Although the behavioral performance pattern is gen-
erally consistent with the conclusion that the effects ofreward color receive strong top–down inhibitory controlin extrastriate cortex, there is one observation in thebehavioral data that warrants further consideration.Increasing reward prospect had a generally inhibitoryimpact on the extrastriate modulations, reward size, how-ever, had no corresponding effect on search performance(in Experiment 2). The presence of the reward-relevantcolor caused a clear decrement of performance whenpresented with the distractor as well as when presentedwith the target. However, the amount of decrement interms of RT and accuracy was uninfluenced by rewardsize. Increased top–down inhibitory control under high-reward conditions would predict reward color to have aless distracting effect on target selection. We think thatthe unchanged performance level reflects the consequenceof increased top–down distractor attenuation under high-reward conditions, an interpretation that can of coursenot be ultimately decided based on the present data.One could object that defining reward relevance inde-
pendent of task relevance may have motivated partici-pants to ignore the reward feature completely andfocus exclusively on the target-defining feature. In otherwords, reward probes may simply not have capturedattention—a situation that would preempt top–downmodulatory influences to counter the distracting effectof the reward color in visual cortex areas. However, thisinterpretation is unlikely given that participants showed aclear performance decrement when presenting the rewardcolor in the search distractor under low- and high-rewardconditions. Furthermore, this possibility conflicts with theobservation that under high-reward conditions rewardprobes elicited a stronger and earlier response than targetprobes in dACC. ACC is believed to subserve the detectionand interpretation of reward and in particular to codereward-dependent changes of stimulus saliency (Hickeyet al., 2010; Hayden et al., 2009). Hence, the reward-relevant color was not only registered but also assigned ahigher significance and temporal priority than the target-defining feature.Although reward probes were subject to effective top–
down inhibition, the target probes showed a significantactivity enhancement relative to control probes underboth low- and high-reward conditions—an observationpredicted by the contingent involuntary orienting account(Remington, Folk, & McLean, 2001; Folk et al., 1992).According to this account, the attention-capturing effect
1062 Journal of Cognitive Neuroscience Volume 26, Number 5

of an onset stimulus depends on the degree to whichproperties of that stimulus meet top–down defined targetdescriptions (but see recent debate; Theeuwes, 2010).Previous studies have documented ERP/ERMF correlates ofsuch contingent attentional capture in visual areas. Arnottet al. (2001) observed with ERPs that attentional cap-ture contingent on a relevant color led to an enhancedoccipital N1 response contralateral to the capturing probestimulus. Hopf et al. (2004)report that the presentation ofa target-defining orientation feature at a nontarget locationin visual search was associated with a retinotopically con-sistent enhancement of neural activity in ventral extrastriatecortex before spatial focusing onto the target. The presentdata show that top–down settings for a task-relevant colortranslate into a color-selective bias of neural processingin ventral extrastriate cortex that is attenuated but stillpresent under high-reward conditions where task settingsemphasize distractor attenuation.It should be noted that although target probes pro-
duced increased extrastriate activations, they were notassociated with an IOR effect different from control andreward probes—an observation that appears to conflictwith the notion that the stronger extrastriate modulationto target probes relates to attentional capture. However,there are data suggesting that the neural operations giv-ing rise to IOR do not depend on the extrastriate process-ing bias for target color observed here. IOR has beensuggested to be strictly location/object related, and re-search has typically failed to demonstrate to IOR in thecolor domain (Kwak & Egeth, 1992). Although Law, Pratt,and Abrams (1995) reported some effects compatiblewith a color-based IOR (Law et al., 1995), they were onlyobservable when a nontarget distractor color was pre-sented between the color cue and the color target—asituation not matching the present experimental condi-tions. Hence, IOR may have been a consequence ofspatial orienting in response to the mere onset of theprobes, without being further influenced by an additionalcolor-selective bias. Indeed, Busse, Katzner, and Treue(2006) investigated effects of exogenous cuing fromcombined feature cues (color/motion direction) andobserved that for long cue-target SOAs, location-validversus −invalid cues produced the typical IOR effect,whereas the validity of motion direction cues did not giverise to any IOR. Finally, it is possible that target probesmay have caused a stronger location bias with the con-sequence of facilitated target selection after valid targetprobes. However, such facilitating effect would beexpected to appear for SOAs much shorter than the onesused in the present experiments (600–900msec; Hopfinger& Mangun, 1998).Under high-reward conditions of Experiment 2, both
reward probes and target probes elicited an enhancedresponse in dACC, with the enhancement for rewardprobes appearing earlier (between 160 and 200 msec)than the enhancement for target probes (between 200and 260 msec). Under low-reward conditions, no dACC
effect appeared for either reward or target probes. Con-sistently, a modulation in the dACC was also not seen inExperiment 1 where the magnitude of reward deliveredon rewarded trials was the same as that deliverd on low-reward trials of Experiment 2. The fact that activity en-hancements in dACC were present for both reward andtarget probes is generally consistent with the role thedACC is supposed to play in executive control (Mansouri,Tanaka, & Buckley, 2009; Dosenbach et al., 2006; Botvinick,Cohen, & Carter, 2004; Schall, Stuphorn, & Brown, 2002).Our observation particularly aligns with the more recentlyadvanced proposal that the dACC is a key part of a corticalsystem maintaining task sets (Dosenbach et al., 2006),which operates upon cues that entail increased demandson attentional control to keep focused on relevant input(Weissman, Gopalakrishnan, Hazlett, & Woldorff, 2005;Weissman, Warner, & Woldorff, 2004). Doubling theamount of monetary reward may have increased the gen-eral significance of the task- and reward-associated color,thereby rendering corresponding probes more potent incapturing attention. The activity enhancements in dACCmay then serve to block the distracting effect of probesdrawn in these colors to maintain the performance focuson subsequent task-relevant search frames.
Conclusion
The reported experiments show that when top–downsettings defining task and reward relevance are disso-ciated, the presentation of reward-relevant probes pro-duced no (low reward) or an attenuated sensory response(high reward) in extrastriate visual cortex relative to con-trol probes. Furthermore, increasing reward prospectcaused reward and target probes to elicit response en-hancements in dACC—a frontal cortex structure knownto mediate the selection of stimuli with particular behav-ioral salience. Importantly, the dACC response to rewardprobes appeared significantly before that to targetprobes, with shorter response latencies to the formerbeing followed by stronger attenuation in extrastriatecortex. Finally, an analysis of the brain response to thesearch frames revealed a response enhancement to thereward color when presented in the distractor—anenhancement that disappeared when increasing thereward relevance of the reward color. These obser-vations together suggest that the selection of reward-relevant features is under effective top–down inhibitorycontrol in extrastriate visual cortex even when top–downdefinitions of reward relevance attention are separated.
Acknowledgments
J.-M. H. and M. A. S. were supported by grant SFB779-TPA1.
Reprint requests should be sent to Jens-Max Hopf, LeibnizInstitute for Neurobiology and Otto-von-Guericke-University ofMagdeburg, Leipziger Strasse 44, 39120 Magdeburg, Germany,or via e-mail: [email protected].
Buschschulte et al. 1063

REFERENCES
Anderson, B. A., Laurent, P. A., & Yantis, S. (2011). Value-drivenattentional capture. Proceedings of the National Academy ofSciences, U.S.A., 108, 10367–10371.
Ansorge, U., Kiss, M., & Eimer, M. (2009). Goal-driven attentionalcapture by invisible colors: Evidence from event-relatedpotentials. Psychonomic Bulletin & Review, 16, 648–653.
Arnott, S. R., Pratt, J., Shore, D. I., & Alain, C. (2001). Attentionalset modulates visual areas: An event-related potential study ofattentional capture.Brain Research, Cognitive Brain Research,12, 383–395.
Arsenault, J. T., Nelissen, K., Jarraya, B., & Vanduffel, W. (2013).Dopaminergic reward signals selectively decrease fMRIactivity in primate visual cortex. Neuron, 77, 1174–1186.
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-drivenattentional capture. Perception & Psychophysics, 55, 485–496.
Boehler, C. N., Hopf, J. M., Krebs, R. M., Stoppel, C. M.,Schoenfeld, M. A., Heinze, H. J., et al. (2011). Task-load-dependent activation of dopaminergic midbrain areas in theabsence of reward. Journal of Neuroscience, 31, 4955–4961.
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflictmonitoring and anterior cingulate cortex: An update. Trendsin Cognitive Sciences, 8, 539–546.
Busse, L., Katzner, S., & Treue, S. (2006). Spatial and feature-based effects of exogenous cueing on visual motionprocessing. Vision Research, 46, 2019–2027.
Chelazzi, L., & Desimone, R. (1994). Responses of V4 neuronsduring visual search. Society for Neuroscience Abstract,20, 1054.
Chelazzi, L., Duncan, J., Miller, E. K., & Desimone, R. (1998).Response of neurons in inferior temporal cortex duringmemory-guided visual search. Journal of Neurophysiology,80, 2918–2940.
Cohen, J. Y., Heitz, R. P., Schall, J. D., & Woodman, G. F. (2009).On the origin of event-related potentials indexing covertattentional selection during visual search. Journal ofNeurophysiology, 102, 2375–2386.
Danielmeier, C., Eichele, T., Forstmann, B. U., Tittgemeyer, M.,& Ullsperger, M. (2011). Posterior medial frontal cortexactivity predicts post-error adaptations in task-relatedvisual and motor areas. The Journal of Neuroscience, 31,1780–1789.
Dosenbach, N. U., Visscher, K. M., Palmer, E. D., Miezin, F. M.,Wenger, K. K., Kang, H. C., et al. (2006). A core system for theimplementation of task sets. Neuron, 50, 799–812.
Eimer, M., & Kiss, M. (2008). Involuntary attentional capture isdetermined by task set: Evidence from event-related brainpotentials. Journal of Cognitive Neuroscience, 20,1423–1433.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992).Involuntary covert orienting is contingent on attentionalcontrol settings. Journal of Experimental Psychology:Human Perception and Performance, 18, 1030–1044.
Folk, C. L., Remington, R. W., & Wright, J. H. (1994). Thestructure of attentional control: Contingent attentionalcapture by apparent motion, abrupt onset, and color.Journal of Experimental Psychology: Human Perceptionand Performance, 20, 317–329.
Franko, E., Seitz, A. R., & Vogels, R. (2009). Dissociable neuraleffects of long-term stimulus-reward pairing in macaquevisual cortex. Journal of Cognitive Neuroscience, 22,1425–1439.
Fukuda, K., & Vogel, E. K. (2009). Human variation in overridingattentional capture. Journal of Neuroscience, 29, 8726–8733.
Gibson, B. S., & Kelsey, E. M. (1998). Stimulus-drivenattentional capture is contingent on attentional set fordisplaywide visual features. Journal of Experimental
Psychology: Human Perception and Performance, 24,966–706.
Guthrie, D., & Buchwald, J. S. (1991). Significance testing ofdifference potentials. Psychophysiology, 28, 240–244.
Haber, S. N., & Knutson, B. (2010). The reward circuit:Linking primate anatomy and human imaging.Neuropsychopharmacology, 35, 4–26.
Hayden, B. Y., Pearson, J. M., & Platt, M. L. (2009). Fictivereward signals in the anterior cingulate cortex. Science,324, 948–950.
Heinze, H. J., Mangun, G. R., Burchert, W., Hinrichs, H., Scholz,M., Münte, T. F., et al. (1994). Combined spatial and temporalimaging of brain activity during visual selective attention inhumans. Nature, 372, 543–546.
Hickey, C., Chelazzi, L., & Theeuwes, J. (2010). Reward changessalience in human vision via the anterior cingulate. Journalof Neuroscience, 30, 11096–11103.
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009).Electrophysiological indices of target and distractorprocessing in visual search. Journal of CognitiveNeuroscience, 21, 760–775.
Hickey, C., McDonald, J. J., & Theeuwes, J. (2006).Electrophysiological evidence of the capture of visualattention. Journal of Cognitive Neuroscience, 18, 604–613.
Hickey, C., & van Zoest, W. (2012). Reward creates oculomotorsalience. Current Biology, 22, R219–R220.
Hikosaka, O., Bromberg-Martin, E., Hong, S., & Matsumoto, M.(2008). New insights on the subcortical representation ofreward. Current Opinion in Neurobiology, 18, 203–208.
Hillyard, S. A., Vogel, E. K., & Luck, S. J. (1998). Sensory gaincontrol (amplification) as a mechanism of selective attention:Electrophysiological and neuroimaging evidence.Philosophical Transactions of the Royal Society of London,Series B, 353, 1257–1270.
Hopf, J.-M., Boelmans, K., Schoenfeld, A., Luck, S. J., & Heinze,H.-J. (2004). Attention to features precedes attention tolocations in visual search: Evidence from electromagneticbrain responses in humans. Journal of Neuroscience, 24,1822–1832.
Hopf, J.-M., Heinze, H. J., Schoenfeld, M. A., & Hillyard, S. A.(2009). Spatio-temporal analysis of visual attention. In M. S.Gazzaniga (Ed.), The cognitive neurosciences IV(pp. 235–250). Cambridge, MA: MIT Press.
Hopf, J.-M., Luck, S. J., Girelli, M., Hagner, T., Mangun, G. R.,Scheich, H., et al. (2000). Neural sources of focused attentionin visual search. Cerebral Cortex, 10, 1233–1241.
Hopf, J.-M., Vogel, E., Woodman, G., Heinze, H. J., & Luck, S. J.(2002). Localizing visual discrimination processes in time andspace. Journal of Neurophysiology, 88, 2088–2095.
Hopfinger, J. B., & Mangun, G. R. (1998). Reflexive attentionmodulates processing of visual stimuli in human extrastriatecortex. Psychological Science, 9, 441–447.
Hopfinger, J. B., & Ries, A. J. (2005). Automatic versuscontingent mechanisms of sensory-driven neural biasing andreflexive attention. Journal of Cognitive Neuroscience, 17,1341–1352.
Ikeda, T., & Hikosaka, O. (2003). Reward-dependent gain andbias of visual responses in primate superior colliculus.Neuron, 39, 693–700.
Kastner, S., & Ungerleider, L. G. (2000). Mechanisms of visualattention in the human cortex. Annual Review ofNeuroscience, 23, 315–341.
Kiss, M., Driver, J., & Eimer, M. (2009). Reward priority ofvisual target singletons modulates event-related potentialsignatures of attentional selection. Psychological Science,20, 245–251.
Kiss, M., Jolicoeur, P., Dellʼacqua, R., & Eimer, M. (2008).Attentional capture by visual singletons is mediated by
1064 Journal of Cognitive Neuroscience Volume 26, Number 5

top–down task set: New evidence from the N2pc component.Psychophysiology, 45, 1013–1024.
Klein, R. M. (2000). Inhibition of return. Trends in CognitiveSciences, 4, 138–147.
Kristjansson, A., Sigurjonsdottir, O., & Driver, J. (2010). Fortuneand reversals of fortune in visual search: Reward contingenciesfor pop-out targets affect search efficiency and targetrepetition effects. Attention, Perception, & Psychophysics,72, 1229–1236.
Kwak, H. W., & Egeth, H. (1992). Consequences of allocatingattention to locations and to other attributes. Perception &Psychophysics, 51, 455–464.
Lamy, D., Leber, A., & Egeth, H. E. (2004). Effects of taskrelevance and stimulus-driven salience in feature-searchmode. Journal of Experimental Psychology: HumanPerception and Performance, 30, 1019–1031.
Law, M. B., Pratt, J., & Abrams, R. A. (1995). Color-based inhibitionof return. Perception & Psychophysics, 57, 402–408.
Leblanc, E., Prime, D. J., & Jolicoeur, P. (2008). Tracking thelocation of visuospatial attention in a contingent captureparadigm. Journal of Cognitive Neuroscience, 20, 657–671.
Lee, B. B., Martin, P. R., & Valberg, A. (1988). The physiologicalbasis of heterochromatic flicker photometry demonstrated inthe ganglion cells of the macaque retina. Journal ofPhysiology, 404, 323–347.
Lennert, T., & Martinez-Trujillo, J. (2011). Strength of responsesuppression to distracter stimuli determines attentional-filtering performance in primate prefrontal neurons. Neuron,70, 141–152.
Lien, M. C., Ruthruff, E., Goodin, Z., & Remington, R. W. (2008).Contingent attentional capture by top–down control settings:Converging evidence from event-related potentials. Journalof Experimental Psychology: Human Perception andPerformance, 34, 509–530.
Mansouri, F. A., Tanaka, K., & Buckley, M. J. (2009). Conflict-induced behavioural adjustment: A clue to the executivefunctions of the prefrontal cortex. Nature ReviewsNeuroscience, 10, 141–152.
Martinez, A., Anllo-Vento, L., Sereno, M. I., Frank, L. R., Buxton,R. B., Dubowitz, D. J., et al. (1999). Involvement of striate andextrastriate visual cortical areas in spatial attention. NatureNeuroscience, 2, 364–369.
Maunsell, J. H. (2004). Neuronal representations of cognitivestate: Reward or attention? Trends in Cognitive Sciences, 8,261–265.
Maunsell, J. H., & Treue, S. (2006). Feature-based attention invisual cortex. Trends in Neurosciences, 29, 317–322.
Maylor, E. A., & Hockey, R. (1985). Inhibitory component ofexternally controlled covert orienting in visual space. Journalof Experimental Psychology: Human Perception andPerformance, 11, 777–787.
Moore, T., & Armstrong, K. M. (2003). Selective gating of visualsignals by microstimulation of frontal cortex. Nature, 421,370–373.
Moore, T., Armstrong, K. M., & Fallah, M. (2003). Visuomotororigins of covert spatial attention. Neuron, 40, 671–683.
Moore, T., & Fallah, M. (2004). Microstimulation of the frontaleye field and its effects on covert spatial attention. Journal ofNeurophysiology, 91, 152–162.
Motter, B. C. (1994). Neural correlates of feature selectivememory and pop-out in extrastriate area V4. Journal ofNeuroscience, 14, 2190–2199.
Noudoost, B., & Moore, T. (2011). Control of visual corticalsignals by prefrontal dopamine. Nature, 474, 372–375.
Posner, M. I., & Cohen, Y. (1984). Components of visualorienting. In H. Bouma & D. Bowhuis (Eds.), Attentionand performance X (pp. 531–556). Hillsdale, NJ: Erlbaum.
Remington, R. W., Folk, C. L., & McLean, J. P. (2001).Contingent attentional capture or delayed allocation ofattention? Perception & Psychophysics, 63, 298–307.
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression ofattention by salient singletons: Electrophysiological evidencefor an automatic attend-to-me signal. Attention, Perception &Psychophysics, 72, 1455–1470.
Sawaki, R., & Luck, S. J. (2011). Active suppression of distractorsthat match the contents of visual working memory. VisualCognition, 19, 956–972.
Schall, J. D., Stuphorn, V., & Brown, J. W. (2002). Monitoring andcontrol of action by the frontal lobes. Neuron, 36, 309–322.
Schultz, W. (2000). Multiple reward signals in the brain. NatureReviews Neuroscience, 1, 199–207.
Serences, J. T. (2008). Value-based modulations in human visualcortex. Neuron, 60, 1169–1181.
Shuler, M. G., & Bear, M. F. (2006). Reward timing in theprimary visual cortex. Science, 311, 1606–1609.
Theeuwes, J. (2010). Top–down and bottom–up control ofvisual selection. Acta Psychologica (Amsterdam), 135, 77–99.
Theeuwes, J., & Godljn, R. (2001). Attentional and oculomotorcapture. In C. L. Folk & B. S. Gibson (Eds.), Attraction,distraction, and action: Multiple perspectives on attentionalcapture (pp. 121–150). Amsterdam: Elsevier.
Treue, S. (2001). Neural correlates of attention in primate visualcortex. Trends in Neurosciences, 24, 295–300.
Treue, S., & Martinez Trujillo, J. C. (1999). Feature-basedattention influences motion processing gain in macaquevisual cortex. Nature, 399, 575–579.
Weil, R. S., Furl, N., Ruff, C. C., Symmonds, M., Flandin, G.,Dolan, R. J., et al. (2010). Rewarding feedback aftercorrect visual discriminations has both general and specificinfluences on visual cortex. Journal of Neurophysiology, 104,1746–1757.
Weissman, D. H., Gopalakrishnan, A., Hazlett, C. J., & Woldorff,M. G. (2005). Dorsal anterior cingulate cortex resolves conflictfrom distracting stimuli by boosting attention toward relevantevents. Cerebral Cortex, 15, 229–237.
Weissman, D. H., Warner, L. M., & Woldorff, M. G. (2004).The neural mechanisms for minimizing cross-modaldistraction. Journal of Neuroscience, 24, 10941–10949.
Yantis, S. (1996). Attentional capture in vision. In A. Kramer,M. Coles, & G. Logan (Eds.), Converging operations in thestudy of selective visual attention (pp. 45–76). Washington,DC: American Psychological Association.
Buschschulte et al. 1065