
Review - J-Stage
21
῎ ῎ ῎ ῎ ῌ ῍ ῌ ῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍῍ Copyright , Japan Poultry Science Association. http://www.jstage.jst.go.jp/browse/jpsa doi: . /jpsa. . Received: April , , Accepted: April , Correspondence: Dr. M. Tsudzuki, Graduate School of Biosphere Science, Hiroshima University, Higashi-Hiroshima, Hiroshima , Japan. (E-mail: [email protected]) Graduate School of Biosphere Science, Hiroshima University, Higashi-Hiroshima , Japan Japanese quail ( ) is a valuable bird for both research and production. From a mainly biomedical research point of view, this article describes characteristics of mutations of Japanese quail that have been found in the last years, along with the recent ( or years) advances in molecular genetics of Japanese quail. In the years following the first report of a mutation in Japanese quail in , approximately mutations were found, and during the subsequent years, approximately mutations have been added to the list of mutations in Japanese quail to date. Based on advances in molecular genetics of Japanese quail, two main genetic linkage maps have been constructed, mostly using microsatellite DNA and amplified fragment length polymorphism (AFLP) markers, and several mutations have been solved at the molecular level for their causative genes. It is anticipated that molecular studies will further reveal the nature of Japanese quail in the near future and enhance its value as a laboratory research animal and agricultural (industrial) animal. : causative gene, gene mapping, genetic linkage map, Japanese quail, mutation other kinds of DNA markers were developed in the s, quail is also used as an experimental animal. The first report in which Japanese quail was genetically examined Wild Japanese quail ( ), belonging to was published by Shimakura ( ). Shimakura ( ) the order Galliformes and the family Phasianidae, inhabits reported the first plumage color mutation in Japanese the eastern part of the Eurasian continent, including quail, brown-splashed white, and recognized the advan- Hainan and Taiwan, and the islands of Japan. In each of tages of using Japanese quail as a laboratory research the continent and the islands of Japan, this species mi- animal. However, his proposal was not widely recognized grates from the northern area to the southern area in throughout the world, probably because the paper was winter season and from the southern area to the northern written in Japanese and submitted to a Japanese journal. area in summer season (Yamashina, ; Taka-Tsukasa, About years later, Padgett and Ivey ( ) and Wilson ; Kawahara, ; Wakasugi, ). It is generally ( ) again proposed the advantages of Japanese thought that Japanese quail was first domesticated in quail as a laboratory research animal. Since then, use of Japan around the th century as a pet song bird. Al- Japanese quail has become wide spread in the biomedical though the true origin and onset of domestication are research fields (Homma, ; Minvielle, ). The unclear (Kawahara, ; Wakasugi, ; Crawford, merits of this bird are found in its small body size, short ), it is apparently documented that domesticated generation turnover, and high egg production. Japanese quail were reared in Japan around the th In the genetic research fields employing Japanese quail, century as a pet bird for song and plumage colors a number of morphological, biochemical, and other muta- (Soseidoh-Syujin, ). Thereafter, at the beginning of tions have been discovered (Somes, ; Cheng and the s, the improvement and use of Japanese quail for Kimura, ). The studies on biochemical ( iso- egg production began in Japan (Oda, ; Ohmori, zymes) and immunological ( blood types) traits were ). Today, it is widely reared for egg and meat performed from the s to the s with the zenith in production not only in Japan, but also in several countries the s. Morphological mutants were also actively of Asia, Europe, and America (Minvielle, ). studied from the s to the s, but the number of Besides rearing for eggs/meat production, Japanese studies on classical mutants decreased after . Instead of these classical studies, modern molecular techniques began to be adapted to quail genomes from the s onwards. In chickens, a number of microsatellite and and a comprehensive genetic linkage map was constructed Masaoki Tsudzuki The Journal of Poultry Science, : , Coturnix japonica J. Poult. Sci., : , Coturnix japonica et al. e.g., e.g., Key words Review Introduction ,**2 +* ,+.+ ./ +/3 ,+ ,**2 ,2 ,**2 1-3 2/,2 1-3 2/,2 ,* +* +/ /* +3.* /* ,* ,/ +33* +3.* +3.* +30+ ,* +3/3 +301 +312 +32. +30+ +/ +31* ,**. +312 +32. +33* +1 +0.3 +322 +3** +33* +3+1 +3+2 +30* +32* +31* ,**. +30* +33* ,*** +33* ./ +/3 +13 ,**2 ./ +/3 +13 ,**2
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Review - J-Stage

�
�
�
�
� �
�
�����������������������
Copyright , Japan Poultry Science Association.
http://www.jstage.jst.go.jp/browse/jpsa
doi: . /jpsa. .
Received: April , , Accepted: April ,
Correspondence: Dr. M. Tsudzuki, Graduate School of Biosphere
Science, Hiroshima University, Higashi-Hiroshima, Hiroshima
, Japan. (E-mail: [email protected])
Graduate School of Biosphere Science, Hiroshima University, Higashi-Hiroshima , Japan
Japanese quail ( ) is a valuable bird for both research and production. From a mainly biomedical
research point of view, this article describes characteristics of mutations of Japanese quail that have been found in the last
years, along with the recent ( or years) advances in molecular genetics of Japanese quail. In the years following the
first report of a mutation in Japanese quail in , approximately mutations were found, and during the subsequent
years, approximately mutations have been added to the list of mutations in Japanese quail to date. Based on advances in
molecular genetics of Japanese quail, two main genetic linkage maps have been constructed, mostly using microsatellite DNA
and amplified fragment length polymorphism (AFLP) markers, and several mutations have been solved at the molecular level
for their causative genes. It is anticipated that molecular studies will further reveal the nature of Japanese quail in the near
future and enhance its value as a laboratory research animal and agricultural (industrial) animal.
: causative gene, gene mapping, genetic linkage map, Japanese quail, mutation
other kinds of DNA markers were developed in the s,
quail is also used as an experimental animal. The first
report in which Japanese quail was genetically examined
Wild Japanese quail ( ), belonging to was published by Shimakura ( ). Shimakura ( )
the order Galliformes and the family Phasianidae, inhabits reported the first plumage color mutation in Japanese
the eastern part of the Eurasian continent, including quail, brown-splashed white, and recognized the advan-
Hainan and Taiwan, and the islands of Japan. In each of tages of using Japanese quail as a laboratory research
the continent and the islands of Japan, this species mi- animal. However, his proposal was not widely recognized
grates from the northern area to the southern area in throughout the world, probably because the paper was
winter season and from the southern area to the northern written in Japanese and submitted to a Japanese journal.
area in summer season (Yamashina, ; Taka-Tsukasa, About years later, Padgett and Ivey ( ) and Wilson
; Kawahara, ; Wakasugi, ). It is generally ( ) again proposed the advantages of Japanese
thought that Japanese quail was first domesticated in quail as a laboratory research animal. Since then, use of
Japan around the th century as a pet song bird. Al- Japanese quail has become wide spread in the biomedical
though the true origin and onset of domestication are research fields (Homma, ; Minvielle, ). The
unclear (Kawahara, ; Wakasugi, ; Crawford, merits of this bird are found in its small body size, short
), it is apparently documented that domesticated generation turnover, and high egg production.
Japanese quail were reared in Japan around the th In the genetic research fields employing Japanese quail,
century as a pet bird for song and plumage colors a number of morphological, biochemical, and other muta-
(Soseidoh-Syujin, ). Thereafter, at the beginning of tions have been discovered (Somes, ; Cheng and
the s, the improvement and use of Japanese quail for Kimura, ). The studies on biochemical ( iso-
egg production began in Japan (Oda, ; Ohmori, zymes) and immunological ( blood types) traits were
). Today, it is widely reared for egg and meat performed from the s to the s with the zenith in
production not only in Japan, but also in several countries the s. Morphological mutants were also actively
of Asia, Europe, and America (Minvielle, ). studied from the s to the s, but the number of
Besides rearing for eggs/meat production, Japanese studies on classical mutants decreased after . Instead
of these classical studies, modern molecular techniques
began to be adapted to quail genomes from the s
onwards. In chickens, a number of microsatellite and
and a comprehensive genetic linkage map was constructed
Masaoki Tsudzuki
The Journal of Poultry Science, : ,
Coturnix japonica
J. Poult. Sci., : ,
Coturnix japonica
et al.
e.g.,
e.g.,
Key words
Review
Introduction
,**2 +* ,+.+ ./ +/3
,+ ,**2 ,2 ,**2
1-3
2/,2
1-3 2/,2
,*+* +/ /*
+3.* /* ,*,/
+33*
+3.* +3.*
+30+ ,* +3/3+301 +312 +32. +30+
+/+31* ,**.
+312 +32.+33*
+1
+0.3 +322+3** +33*
+3+1+3+2 +30* +32*
+31*,**. +30* +33*
,***
+33*
./ +/3 +13 ,**2
./ +/3 +13 ,**2

�
�
(Groenen ; Schmid ). On the other feathers, the whole pigmentation is diluted and the wheat-
hand, over microsatellite markers and a number of straw colored shaft streak is narrower than the wild type.
other kinds of DNA markers were also developed in Furthermore, the transversal bars in individual feathers
Japanese quail from to , and later in chickens, are completely missing. The dotted white is a bottom
genetic linkage maps based on these markers were con- recessive allele at the panda ( ) locus (Mizutani
structed (Kayang ; Sasazaki a). ; Tsudzuki ). The panda is also a mutant
Biochemical and immunological variants of Japanese that shows white plumage with dark patches on the head,
quail have been well summarized by Kimura ( ), cheek, back, wing, and tail. In contrast with the dotted
Somes ( ), and Cheng and Kimura ( ), along with white, in the panda mutant the feathers from the dark
morphological, behavioral (neurological), and other kinds patches are similar to the wild type in both color and
of mutations by Somes ( ) and Cheng and Kimura markings (Tsudzuki ).
( ). It is now years since these reviews were done. ( ) (Tsudzuki ): This is an auto-
During this years however, there are very few biochem- somal incompletely dominant mutation that occurred at
ical and immunological variants that have been newly the yellow ( ) locus (Homma ). The neonatal
discovered. Therefore in this article, the author mostly plumage of the homozygote shows a creamy yellow color
reviews morphological, behavioral (neurological), and all over the body with three dark brownish obscure stripes
other kinds of mutants that have been discovered in on the back, and no stripes on the head. However, faint
Japanese quail during the past years, along with recent dark pigmentation is observed at the tip of some head
developments in molecular genetics for Japanese quail. plumules. In the chicks heterozygous for this mutation,
the creamy color is slightly deeper than that of the homo-
zygote. The three stripes on the back are more blackish in
Table is a list of morphological, behavioral, and some color and clearer than those of the homozygote. Sex
other mutations in Japanese quail that have so far been di erences are large in adult plumage of both the homozy-
discovered. Among the loci controlling these mutations, gote and heterozygote. The homozygous male shows
some are multiple-allelic loci, which are also summarized whitish light-brown plumage with a few small black speck-
in Table . The author herein describes features of the les on the back, while the homozygous female shows more
Japanese quail mutations that were found during the last creamy plumage than the male and has a relatively large
years, with those of some older mutations. Mutations number of black speckles on the back. Irrespective of the
have been categorized as morphological, behavioral or yellow mutant, the homozygote of the fawn- mutant is
muscular, and other mutations. Morphological mutations fully viable. The face of the homozygous male is rusty
have been further classified into plumage color, plumage with the crown composed of dark and creamy feathers,
system, eggshell color, and morphogenetic mutations. while the face of the homozygous female is creamy with
Gene symbols presented here are according to those sparsely dotted black pigmentation. In the heterozygote,
described in each original report, although there is some the primary plumage pattern is basically the same as tha
confusion in nomenclature of mutant genes.
( ) (Tsudzuki ): This
autosomal recessive plumage color mutation was found in
. The neonatal mutant chick has the same striped
plumage pattern as the wild type. However, the plumage
color is slightly lighter than the wild type. The brownish
stripes of the wild type are replaced by slightly lighter
brownish or yellowish colored stripes. In the wings and
thighs of the mutant chick, blurry black markings replace
the dense black markings of the wild type. Besides the
down color abnormality, of the mutant chicks have
bent digits. All the mutant chicks die within three days of
hatching.
( ) (Tsudzuki ): This is an
autosomal recessive plumage color mutation discovered in
. The mutant chick has creamy yellow down plu-
mules with a brown spot on the head and/or back.
of the mutant chicks have no colored spot. The creamy
yellow and brown plumules of the chick replace white and
dark feathers respectively in their adulthood. In the dark
f
dw
et al., et al.,
s et al.,
et al., et al., et al.,
et al.,
Y et al.,
Y et al.,
ddl et al.,
s et al.,
Morphological Mutations
Fawn-
Plumage Color Mutations
Dilute down lethal
Dotted white
Mutations in Japanese Quail
,*** ,***+**
,*** ,**/
,**0 ,**0 +31. +33-
+32,+322 +33*
+322 +33-+33* ,* +330
,*+301
,*
+#
,
,*,
+331
+33.
,/
+33,
+32,+,
,,
J. Poult. Sci., ( )
t
of the homozygote, but the brown color is deeper in the
heterozygote than in the homozygote in each sex. The
female plumage of the heterozygote has a much larger
amount of black pigmentation than the homozygous
female, somewhat similar to the yellow mutant plumage.
The phenotype and mode of inheritance of the fawn-
mutant are similar to those of the fawn mutant (Nichols
and Cheng, ) that is an allele at the yellow locus
(Cheng and Kimura, ). Furthermore, Minvielle
( ) described that the beige mutant is also similar to
the fawn mutant both in the phenotype and mode of
inheritance. Thus, there is a possibility that these three
(fawn, fawn- , and beige) are identical mutants, or alleles
at the yellow locus.
(formerly bleu) ( formerly, )
(Minvielle , ): This is one of the dilution
mutations that was renamed by Minvielle ( ,
). The previous mutant name was bleu (Cheng and
Kimura, ). The adult mutant shows bluish gray all
over the body with the same primary plumage pattern as
the wild type. The secondary plumage pattern (the mark-
ing in individual feathers) is also the same as that of the
wild type. However, in individual contour feathers, the
et al.
LAV*L, bl
et al.,
et al.
Lavender
160 ./ -
,
+322+33*
,**-
,
,**, ,**-,**,
,**-+33*

��
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�Table .
Mode ofCategory of traits Mutant names Gene symbols References
inheritance
Morphological mutations
Plumage color Beige AD Minvielle ( )
Black AD Chikamune and Kanai ( ), Somes ( )
Black at hatch AD Minezawa and Wakasugi ( )
Brown-splashed white AR Shimakura ( )
Bu AR Sittmann ( )
Cinnamon AR Fulton ( a)
Complete albino AR Homma ( )
Dilute down lethal AR Tsudzuki ( )
Dotted white AR Tsudzuki ( , )
Extended brown AD Somes ( ), Truax and Johnson ( )
Fawn AD Cheng and Kimura ( )
Fawn- AD Tsudzuki ( )
Imperfect albinism SR Lauber ( ), Homma ( )
Lavender (Bleu) AR Minvielle ( , )
Light down AD Ito and Tsudzuki ( )
Light down lethal AR Tsudzuki ( b)
Marbled plumage AR Yakovlev ( )
Orange AR Ito and Tsudzuki ( )
Panda AR Mizutani ( )
Pansy AR Tsudzuki ( )
Recessive black AR Fujiwara ( ), Hiragaki ( )
Recessive silver AR Homma ( )
Recessive white AR Roberts ( ), Somes ( )
Redhead AR Truax ( )
Roux SR Minvielle ( )
Rusty AR Minvielle ( a)
Sex-linked brown SR Wakasugi and Kondo ( )
Sex-linked cinnamon SR Wakasugi and Kondo ( )
Silver AD Homma ( )
White AD Wakasugi and Kondo ( )
White bib (white crescent) AR ( ) Roberts ( )
White breasted AR Roberts ( )
White primaries AR Somes ( )
White-feathered down AR Sittmann and Abplanalp ( )
Yellow AD Homma ( )
Plumage system Curly AR Minvielle ( a)
Defective feathers AD, AR Fulton ( )
Downless AR Savage and Collins ( a)
Fray AD, AR Tsudzuki ( c)
Partial featherlessness AD This article
Porcupine AR Fulton ( b)
Rough-textured AR Roberts and Fulton ( )
Ru e AR Somes ( )
Short barb AR Fulton ( c)
Eggshell color Celadon AR Ito ( )
Red eggshell AD Hardiman ( )
White eggshell AR Poole ( )
Tsudzuki: Mutations and Modern Genetics in Quail
brownish color area is replaced with bluish gray and the eter of the pupils is much larger than that of the heterozy-
wheat-straw color markings with a whitish appearance. gotes. Consequently, the iris area of the homozygotes
( ) (Ito and Tsudzuki, ): This plum- is extremely small. All of the homozygous chicks that
age color mutation is controlled by an incompletely dom- hatched show behavioral abnormalities with variable ex-
inant autosomal gene. The homozygotes, a third of which pressivity. Mildly a ected chicks slowly shake their heads
die before hatching, show creamy yellow down plumules. right and left, while severely a ected chicks rigidly bend
The eye of the homozygotes is not reddish, but the diam- the neck inward and subsequently, either draw backward
dw
F
f
ps
rh
c
FAW*BE et al.
D
Bh
p
pk et al.
cin et al.
a et al.
ddl et al.
s et al.
E
Y
Y et al.
al et al.
LAV*L et al.
L
ldl
ma et al.
or
s et al.
d et al.
Y*RB et al. et al.
rs et al.
wh et al.
e et al.
BR*R et al.
RU*R et al.
br
al
B et al.
W
bi cr et al.
wb et al.
wp
c
Y et al.
CU*C et al.
Df, mdf et al.
dl- , dl-
Fr, mod et al.
Pf
pc et al.
rt
rf
sb et al.
ce et al.
R et al.
we
L
Summary of mutations found in Japanese quail ( )Coturnix japonica
Light down
161
+
-,
.
/
0
0
1
/
.
.
0
2
1
2
3
+*
,**-+312 +32.
+311+3.*
# +300+32,
+302+331+33, +33-
+313 +313+33*
, +330+30. +302
,**, ,**-+33-
+33/+31/
+33.+31.+33*,**/ ,**2
+32/+312 +313
+313,**-,**/
+31-+31-
+303+31-
+312+312
+32.+30/
+301,**/
+32-+31+
+332
+32,+313
% +322+32,
+33-+31/
+30.
+33-
##
,
+ ,
+

��
�
�
�
�
�
�
�
�
�
�
�
� � �
�
�
Table .
Mode ofCategory of traits Mutant names Gene symbols References
inheritance
Morphogenesis Abnormal head AD Morse and Abplanalp ( ), Somes ( )
Arostroperocephaly AR Dodson and Coleman ( )
Chondrodystrophy AR Collins ( )
Chondrodystrophy- AR Hermes ( )
Crooked neck dwalf AR Sittmann and Craig ( )
Deformed beak AR Cheng and Kimura ( )
Ear tuft AR Tsudzuki and Wakasugi ( b)
Hereditary multiple malformation AR Tsudzuki ( a)
Long beak AR Cheng and Kimura ( )
Micromelia AR Hill ( )
Short beak AR Tsudzuki ( b)
Short lower beak AR Nakane and Tsudzuki ( c)
Stumpy limb AR Tsudzuki ( a)
Throat tuft AR Tsudzuki and Wakasugi ( )
Zazen AD, SR Nakane and Tsudzuki ( b)
Behavioral or Back drawer AR Tsudzuki and Wakasugi ( a)
muscular Circular AR Mizutani ( )
mutations Congenital loco AR Sittmann ( )
Crooked neck unknown Dodson and Coleman ( )
Dark feather nervous disorder AR Kawahara ( , ), Ueda ( )
Hypotrophic axonopathy AR Mizutani ( )
Muscular disorder AD Braga ( a, b)
Star gazing AR Savage and Collins ( )
Wry neck AR Savage and Collins ( b)
Other mutations Clench AR Nakane and Tsudzuki ( a)
Diabetes insipidus AR Minvielle ( a)
Glycogenosis type-II AR Nunoya ( )
Twinning AD Sittmann ( )
Biochemical and immunological variants are not included.
AD autosomal dominant over the wild-type allele, AR autosomal recessive to the wild-type allele, SR sex-linked recessive to the
wild-type allele. Almost all AD essentially are incompletely dominant.
Including some symbols that were not proposed in each original paper but thereafter proposed by Cheng and Kimura ( ) and this
article.
There is a possibility that the three mutations are identical or alleles at the same locus.
It is very likely that the two mutations are identical.
There is a possibility that the three mutations are identical or alleles at the same locus.
There is a possibility that the two mutations are identical or alleles at the same locus.
There is a possibility that the two mutations are identical or alleles at the same locus.
There are two synonyms for this mutation, such as red-eyed brown and dark-eyed dilute (Cheng and Kimura, ).
There are three synonyms for this mutation, such as dominant white, autosomal dilute, and dominant dilute (Cheng and Kimura,
).
J. Poult. Sci., ( )
or roll forward. Some severely a ected chicks draw the feathers, this mutant has four pairs of transversal bars,
head backward with the beak pointing upward. All homo- while the wild type has three pairs.
zygous chicks that could hatch die within a week of ( ) (Tsudzuki, b): This plum-
hatching. The neonatal plumage of the heterozygotes is age color mutant is an autosomal recessive with nearly
similar to that of the wild type. However, the black stripes total lethality. The neonatal plumage pattern of this
on the head and back are somewhat narrower than those mutant is the same as that of the wild type. However,
of the wild type, and their color is slightly diluted. Conse- black and tan stripes of the wild type are replaced with
quently, the brownish (yellowish) stripes are broader than blurry black and dilute tan stripes, respectively. Almost
those of the wild type, and the color is also lighter. The all of these mutants that could hatch die by three weeks of
whole plumage of the adult heterozygotes is also similar to age, with the majority dying within one week. Moreover,
that of the wild type, but its color is slightly diluted all prehatch mortality is high ( ) in this mutant. Among
over the body just like in the chicks (Fig. A). In the the light down lethal chicks, only two males survived
individual feathers of the adult heterozygotes, the number to adulthood and reproduced normally. The adult mutant
of transversal bars is increased. For example, in the back phenotype is very similar to that of the wild type.
Tt
ab
ar
ch et al.
ch- et al.
cn
db
hfd
hmm et al.
lb
m et al.
sbk et al.
slb- , slb-
sl
hfd
Zn, moz
bkd- , bkd-
cr
lo et al.
dn et al.
hax et al.
Md et al.
sg
wyn- , wyn-
cl
di et al.
gly II et al.
Tw et al.
ldl
(Continuation) Summary of mutations found in Japanese quail ( )Coturnix japonica
Light down lethal
162
+
-,
+
,
-
.
/
0
1
2
3
+*
+31* +322+31-
+302, +33*
+301+33*
+322+332
+33*+30-
+332+332
+33/+323
+332+322
,***+30/
+31,+313 +32* +313
+33,+33/
+31,+31+
+332,**1
+32-+31+
+33*
+33*
+33*
./ -
#
+33/
-1+ +*-
,
+ ,
+ ,
+ ,
+

� �
� �
��
Table .
Loci Alleles References
Black
Extended brown
Hyomandibular furrow closure defect
Imperfect a
( ) (Ito and Tsudzuki, ): This is an auto- small number of black spots. The pansy mutant is con-
somal recessive mutation. The plumage pattern of this trolled by a bottom recessive allele ( ) at the black ( )
mutant is the same as that of the wild type, but the locus (Chikamune and Kanai, ; Somes, ). The
pigmentation is obviously lightened in both chicks and pansy plumage resembles the redhead plumage which is a
adults. The mutant chicks have chocolate brown and light bottom recessive allele ( ) at the extended brown ( )
brownish-yellow stripes on the head and back. In the locus (Somes, ; Truax ). As described in
adult orange males, light reddish-brown is predominant on a later section, the black mutant will be identical to the
the whole body (Fig. B). The color is especially conspic- extended brown mutant. Thus, there is a possibility that
uous on the face and breast. In contrast, in the adult the pansy and redhead mutants are identical mutants, and
orange females, the light reddish-brown color is less that at least these two are alleles.
marked. The eye of the orange mutant is reddish when ( ) (Fujiwara ): This
held to the light. Hatchability of the orange mutant is mutant was first reported by Fujiwara ( ), and
significantly lower than that of the wild type because of the gene symbol was assigned. Later, Hiragaki
the increased prehatching mortality, but fertility in this ( ) found the allelism of this mutation at the yellow
mutant is normal. Furthermore, posthatch mortality ( ) locus (Homma ). This is a bottom reces-
within five weeks of age is greatly increased in the orange sive allele at the yellow locus. Both sexes of this mutant
mutant over that of the wild type ( ). There is exhibit blackish (or dark brownish) plumage throughout
a possibility that the orange mutant is identical to the bu the body that is very similar to the plumage of the black
(Sittmann ) or the cinnamon mutant (Fulton (Chikamune and Kanai, ; Somes, ) or the ex-
a). Ito and Tsudzuki ( ) did not propose a tended brown mutant (Somes, ; Truax ).
gene symbol for this mutation in their report. Here, for In individual feathers on the back of this mutant, the
the sake of convenience, the author proposes the gene wheat-straw colored shaft stream seen in the wild-type
symbol because this mutation was first reported in feathers is missing, although transversal bars are present
. The symbol may be when adopting the as in the wild type. However, the width of the transversal
new nomenclature proposed by Crittenden ( ). bars becomes narrower when compared with that of the
( ) (Tsudzuki and Wakasugi, ; Tsudzuki wild type, which gives it a blackish or dark brownish
): The mutant chicks have light yellow plumules appearance in this mutant. This is the first black colored
with three black stripes on the back. The black stripes are mutation occurring at the agouti signaling protein ( )
narrower and more obscure than those of the wild type. locus in poultry (Hiragaki ), and is also de-
The head plumules are brownish when compared to the scribed in a later section.
light yellow color seen on the back. The plumage pattern ( ) (Minvielle b, ): This is a
of the adults is totally di erent from that of the wild type. sex-linked mutation that belongs to the sex-linked brown
A shaft stream and transversal bars seen in the wild-type ( or recently ) locus (Wakasugi and Kondo, ;
feathers are absent in the pansy mutant. Adult plumage Cheng and Kimura, ) with a bottom recessive e ect.
shows a mixture of rust, black, and white (Fig. C). In The brown locus is linked to the imperfect albinism locus,
the male plumage, the face, chin, and throat are brown or and the recombination rate of was first reported by
heavy rust in color. There are large variations in neonatal Wakasugi and Kondo ( ) and of by Minvielle
and adult plumage. The neonatal plumage in some chicks ( b) between the two loci. The plumage color of
is somewhat similar to that of the fawn- (Tsudzuki the roux mutant is also brownish as is the brown mutant,
), and consequently some adult males resemble homo- but is paler than that of the brown mutant. Judging from
zygous fawn- males in plumage color. Such a type of the the color photo by Minvielle ( b), the plumage
pansy male shows fawn color all over the back with a color of the roux mutant is grayish light brown, rather
ps
rh
ps
or
d D
e E
et al.,
Y*RB et al.,
et al.
rb et al.
Y et al.,
vs.
et al., et
al., et al.,
or,
OR*OR,
et al.
d et
al.,
ASIP
et al.,
BR*R et al.,
br BR
et
al.
et al.,
et al.
A list of multiple-allelic loci on morphological characters
Orange
Recessive black
Pansy
Roux
,
+ ,
+33.
+312 +32.
+313 +313
+
,**/,**/
,**2+301
0* 2#
+300 +312 +32.+32, +33. +313 +313
+33.+330
+321+33*
,**2
,*** ,**-#
+31-+33* #
+-/
+31- -2,***
,+330
, ,***
�
�
�
�
�
�
�
�
�
� �� �� �� �� �� �
� � �� �
Tsudzuki ( )
Truax ( )
Tsudzuki and Wakasugi ( )
lbinism Wakasugi and Kondo ( ), Cheng and Kimura ( )
Panda Tsudzuki ( )
Sex-linked brown Minvielle ( b, )
Yellow Tsudzuki ( ), Hiragaki ( )
Cheng and Kimura ( )
The black locus is very likely to be identical to the extended brown locus.
Descriptions of the gene symbols are based on those that appeared in each original report.
Tsudzuki: Mutations and Modern Genetics in Quail
ps
rh
Tt
c
dw
f
F
D d d et al.
E e e et al.
Hfd hfd hfd
Al al al
S s s et al.
BR*N BR*B BR*R et al.
Y Y y Y*RB et al. et al.
Y Y y
163
+
,
+33*+313
+323+31- +33*
+33-,*** ,**-+330 ,**2
+33*
,

�
�
�
�
Fig. .
J. Poult. Sci., ( )
(A) Comparison of the wild-type (left)and light-down ( / ) mutant (right) quail. (B) A male of the orange ( / ) mutant quail. (C) A female (left)and male (right) of the pansy ( / ) mutant quail. (D) The homozygote for the white mutation ( / ). (E)The heterozygote for the white mutation ( / ). (F) Homozygous (middle) and heterozygous (left and right)chicks for the white ( ) mutation. (G) Wild-type (left) and mutant chicks (others) heterozygous for themutation. The neonatal mutant plumage is variable in color. (H) The fray mutant ( / , / ) with mildexpressivity. (I) The fray mutant with intermediate expressivity. (J) The fray mutant with severe expressivity.(K) A black-at-hatch mutant male. (L) The ventral surface of the partial featherlessness mutants. (M)Comparison of the normal (left) and stumpy-limb ( / ) mutant (right) quail. (N) The back-drawer ( /
/ ) mutant exhibiting severe abnormality. (O) The heterozygote (left) and homozygote (right) forthe silver ( ) mutation. (P) The red-eyed brown ( sex-linked cinnamon, ) mutant.
ps ps
c
L l or ord d W W
W wW W
Fr fr mod mod
sl sl bkd- bkd-, bkd- bkd-
B al
Several morphological and behavioral mutants in Japanese quail.
164
+
./ -
++ , ,

Fig. .
Tsudzuki: Mutations and Modern Genetics in Quail
(continued)
165
+

�
��
�
similar to that of the sex-linked cinnamon. The roux plumules or feathers and are almost totally bare. This
plumage shows significant association with lower body mutant trait is controlled by the combination of an auto-
weight and less abdominal fat pad. Egg production somal dominant mutant gene ( ) and another autosomal
ability is normal, although egg weight is lower recessive modifier gene ( ). The gene has a com-
(Minvielle ). plete lethal e ect in the homozygous condition. The re-
( ) (Minvielle a): This auto- cessive modifier ( ) is epistatic to and the homo-
somal recessive plumage color mutant shows reddish- zygous state of the modifier allows the expression of the
brown (rusty) color all over the body. The individual gene; that is, only the / / genotype exhib-
contour feathers show rusty color in the pennaceous por- its the fray mutant phenotype. Furthermore, the and
tion. However, the plumulaceous portion shows a slate are closely linked on the same chromosome
like color as in the wild type, di ering from the roux (Tsudzuki c). The fray mutant is similar to the
mutant (Minvielle b, ) in which rusty defective feathers mutant in the phenotype and mode of
color is seen all over the contour feather. The rusty color inheritance of two-locus control (Fulton ).
is heavier in the rusty mutant than in the roux mutant. However, the two loci controlling the defective feathers
Hatchability of the rusty mutant is normal. mutant trait seem to be located at di erent chromosomes
( ) (Wakasugi and Kondo, ): This is an (Fulton ). Therefore, the two mutants are
autosomal dominant plumage color mutation. The mu- di erent.
tant name is assigned to the plumage appearing in the ( ): In , the author dis-
homozygotes (Fig. D). Very few homozygotes for this covered phenotypically unusual chicks among the black-
mutation survive to adulthood and show white plumage all at-hatch ( ) mutant (Minezawa and Wakasugi, )
over the body. However, according to the observations quail (Fig. K) flock kept by the Department of Labora-
made by the author, the white is not pure white but has tory Animal Science, Faculty of Agriculture, Osaka Pre-
extremely diluted gray tinges in the outer margin of indi- fecture University. In the mutant chicks, feather buds did
vidual contour feathers. The adult plumage color of the not extend in the limited areas of the hind head, back,
heterozygotes, which is fully viable, is brownish gray with abdomen, thigh, and chin (Fig. L). Although such
the wild-type plumage pattern (Fig E). The density of chicks showed high mortality, some survived to adulthood
the color is variable from individual to individual. Some and were able to reproduce. No feathers grew in the areas
are similar to that of the wild type and cannot be distin- that were bare at the neonatal chick days. From the
guishable from the wild-type plumage. In the most gray- matings with the wild-type colored normal birds and the
ish case, the wild-type plumage pattern becomes obscure. birds that have the black-at-hatch plumage and partial
The chicks homozygous for this mutation have dusty featherlessness, wild-type colored chicks that have partial
creamy yellow plumules accompanied by dark skin and featherlessness appeared with low incidence, although par-
crooked toes (Fig. F). Almost all homozygotes die tial featherlessness was nearly always exhibited ac-
around hatching. In the heterozygous chicks, the plumage companied by black-at-hatch plumage.
pattern is the same as that of the wild type, but black and
tan stripes seen in the wild type are replaced with grayish
and dusty yellowish stripes, respectively (Fig G). Ac-
cording to Cheng and Kimura ( ), the white mutant is
homologous to the autosomal dilute mutant (Somes,
) and the dominant dilute mutant (Truax and John-
son, ).
( ) (Minvielle a): This is an
autosomal recessive mutation. The adult plumage of the
curly mutant shows a flu er appearance than the wild-
type plumage. The curly phenotype results from the con-
nected calamus of the growing wing feathers at chick
days. The expressivity of the mutant plumage is variable
and the penetrance of the mutant gene is not complete.
Postnatal survivability of this mutant is normal.
( ) (Tsudzuki c): This mutant is
characterized by curly, short, and sparse plumules in
chicks and underdevelopment of barbs and barbules in
adults. There is a large variability in the degree of severity
of the mutant phenotype (Fig. H, I, J). In the most
severe case (Fig. J), both chicks and adults have no
Fr
mod Fr
et al.,
RU*R et al., mod Fr,
Fr Fr fr mod mod
Fr
mod
et al.,
et al.,
et al.,
W et al.,
Pf
Bh
CU*C et al.,
Fr, mod et al.,
Rusty
White
Partial featherlessness
Plumage System Mutations
Curly
Fray
--
,+333 #
,**/
# +332,*** ,**-
+32-
#+31- +32-
#+33/
++311
+
++
+
++33*
+32.+313
,**/
$
+332
++
��
J. Poult. Sci., ( )
This result strong-
ly suggests that the mutant locus controlling the partial
featherlessness is closely linked to the locus, and that
the gene is an autosomal dominant.
( ) (Ito ): The name was assigned
to this eggshell color mutation because of the similarity of
the eggshell color to that of the porcelain in the time of the
Sung, China. This mutation is controlled by an autosomal
recessive gene and is hypostatic to the white eggshell ( )
mutation (Poole, ). The celadon eggshell possesses
the pigments protoporphyrin and biliverdin, as does the
wild-type eggshell. However, the amount of the pro-
toporphyrin in the celadon eggshell is much smaller than
in the wild type ( . of the wild type). The biliverdin
content of the celadon eggshell is of that in the
wild-type eggshell.
( ) (Tsudzuki and Wakasugi, b): This is
a morphogenetic mutation controlled by an autosomal
recessive gene (hyomandibular furrow closure defect)
with incomplete penetrance. The is a bottom recessive
Bh
ce et al.,
we
hfd
hfd
hfd
Eggshell Color Mutations
Celadon
Morphogenetic Mutations
Ear tuft
166 ./ -
+33-
+30.
/ 0..
+322

�
� �
�
�
��
�
�
�
allele at the locus (Tsudzuki and Wakasugi, ). This mutant is not allelic to a similar mutant, stumpy limb
This mutation is characterized by ear-opening abnormali- ( ) (Tsudzuki, a)
ty and an ear tuft, which is composed of an epidermal ( ) (Nakane and Tsudzuki,
peduncle and covering feathers, protruding within or c): This mutant was found in . This morpho-
below the abnormal ear opening. Abnormal ear openings genetic mutant dies during late embryonic stages. The
are oval-shaped and accompanied by fissures of varying mutant embryos at days of incubation are first char-
size on the ventral margin. The incidence of the ear tuft acterized by a shortened lower beak. The severity of the
and ear-opening abnormality is and respectively shortening varies largely. In the most severe case, the
in the homozygotes. These abnormalities are attrib- length of the lower beak is approximately one third of the
uted to the hyomandibular furrow closure (or visceral upper beak, and in mild cases the lower beak is slightly
arch) defect occurring at five days of incubation. The shorter than the upper beak. Almost all mutant embryos
incidence of the hyomandibular furrow closure defect is keep the mouth wide open. In the skull of the mutant
in the homozygotes. However, the defect that oc- embryos, the Meckel’s cartilage of the mandible is abnor-
curred at five days of incubation is rapidly repaired by six mally bent downward in its proximal portion, and ossifica-
days of incubation in approximately a half of the a ected tion of the mandibular bone occurs around the abnormal
embryos, which results in the low ( ) incidence of the Meckel’s cartilage, which seems to be responsible for the
ear-opening abnormality at later stages (Tsudzuki and shortened lower beak and the opened mouth. In addition
Wakasugi, d). The early embryonic abnormalities to these abnormalities, this mutant shows hypoplasia or
occurred around the visceral arch region also give rise to partial lack of the hyoid apparatus, abnormal formation
head skeletal abnormalities; that is, partial deletion or and/or fusion of the sternal ribs, and increased number of
irregularity is seen in the mandible, basiparasphenoid, cervical vertebrae. This mutant trait seems to be con-
quadratojugal, squamosal, and quadrate bones. The inci- trolled by two autosomal recessive genes that are linked on
dence of the head skeletal abnormalities is in the the same chromosome, the recombination rate of which is
homozygotes (Tsudzuki and Wakasugi, c). approximately . However, further genetic studies will
( ) (Tsudzuki be needed to confirm the mode of inheritance of this
a): This autosomal recessive mutation is the first mutation.
polydactyl mutation found in Japanese quail. No mutant ( ) (Tsudzuki ; Tsudzuki,
embryos survive beyond days of incubation, with the a): This mutation was discovered in as an embryonic
peak of death at the sixth day of incubation. Mutant completely lethal mutation. However, changing of genetic
embryos at late incubation stages ( days) show an backgrounds gave rise to viable adult mutants (Fig. M)
early embryo-like body shape, and have no plumules, al- (Tsudzuki, a). Late embryos an
though feather buds are seen in their body. In the mutant
head, the eyelid is left wide open. The upper and lower
beaks are greatly deformed and set apart. The wings and
legs are stumpy and showed syndactylous polydactyly. In
the abdomen, a part of the ventriculus, liver, and small
intestine protrudes out of the umbilicus region. The
skeleton of the mutant embryos shows underdevelopment
and/or morphogenetic abnormalities all over the body. In
the mutant skeleton, ossification is generally delayed, and
cartilage is predominant throughout the body except the
head. Delay of the ossification is most remarkable in the
fore and hind limbs.
( ) (Tsudzuki b): This mutant
was found in . The majority of the short beak mu-
tants die at the late embryonic stages and within three
days of hatching. In this mutant, the length of the beak,
shanks, and digits is shortened to approximately of
that of the wild type, although there is no significant
di erence in body weight between the mutant and wild
type. Irrespective of the shortening, the shank is
thickened to approximately of the wild type. The
shortened beak shows no parrot-like shape, contrasting
with other poultry condrodystrophic mutants (Hays,
; Collins ; Nestor, ; Hermes
; Tsudzuki ). Some of the mutants
( . ) reach sexual maturity and are able to reproduce.
hfd
sl
slb- , slb-
hfd
hfd
hmm et
al.,
sl et al.,
sbk et al.,
sbk
et al., et al.,
et al., sbk
Short lower beak
Hereditary multiple malformation
Stumpy limb
Short beak
+323+33/
+332 +330
+/
-- .,
3+
#.,
+322
21+322 -*
+332+33+ +33/
+/ +32/
+- +/ ++33/
+332+33.
3*
#
++*
+3.. +302 +312+33* +33+/ -
+ ,
�
�
��
Tsudzuki: Mutations and Modern Genetics in Quail
d chicks are charac-
terized by brachycephaly, short beak, thick neck, and
short and thick extremities. At the skeleton level, the
mutant exhibits globular skull, unusual curvature of the
of the upper beak, and
shortening and thickening of the appendicular bones.
Some embryos show a bending of the humerus, femur
and/or tibiotarsus. Abnormalities are more conspicuous
in the leg bones than in the wing bones. In the UOP-WT
genetic background (Ito and Tsudzuki, ), of the
a ected embryos could hatch out of the eggshell with no
assistance, and could walk in a relatively smooth manner.
of the chicks that could hatch died at various ages
before reaching maturity with a mortality peak at three
days of age. Of these, the birds that survived for relatively
long periods started showing di culty in walking around
two weeks of age and subsequently died at various ages.
The remaining ( ) survived maturity, but the birds
that sexually matured were only of the hatched mu-
tant chicks. Although reproductive ability of these sexual-
ly matured mutant birds varied from individual to individ-
ual due to the degree of the di culty in walking, sper-
matogenesis and oogenesis seemed to progress normally
in the gonads of the sexually developed mutants.
( ) (Tsudzuki and Wakasugi, ):
This mutant shows ear-opening abnormality and a feath-
ered tuft as seen in the ear-tufted ( ) mutant (Tsudzuki
Tt
Processus palatinus maxillaris
sl
hfd
hfd
Throat tuft
167
+33. /0#
1-
$
,1+,
$
+323

�
� �
�
��
�
and Wakasugi, b), and is autosomally recessive to the abnormalities, some mutants show shortening of the lower
wild type and dominant over the mutation. As seen in beak with upward bending of its tip, lack of the edges
the case of the the penetrance of the allele is also around the basal part of the upper beak, and lateral
not complete. The tuft originates in the throat region and curvature of the upper beak. In the legs, all the mutant
is composed of an epidermal peduncle and covering feath- embryos have full-length fibula from the stifle articulation
ers. Some birds possessing these throat tufts exhibit to the tibiotarsal-tarsometatarsal articulation. The femur
abnormal ear-openings, in which either a wide cleft or a of the mutant embryos is articulated with the proximal
short fissure is formed at the ventral margin. The inci- end of the fibula, whereas in the normal embryos the
dence of the throat tuft and ear-opening abnormality is femur is articulated with the tibia. In the mutant head, the
and , respectively. Throat tufts are usually ac- maxillary and the palatine processes of the premaxilla are
companied by normal ear openings, contrasting with the extremely reduced in length, which seems to result in the
case of the ear-tuft mutant. Furthermore, the position lack of the basal edges of the upper beak. Almost all zazen
from which peduncles protrude in the mutant is mutants show lack of the parietal and abnormally formed
di erent from that of the mutant. There is also a dif- ribs. The joint between the sternal and vertebral ribs is
ference in abnormal morphology of the ear opening be- fused and/or ossified. The vertebral ribs gain or lose one
tween the two. The ear-tuft mutants exhibit a fissure rib in number, and the ribs themselves are abnormally
below the ear opening, whereas the throat-tuft mutants shaped and/or deformed. This mutation seems to be
express a wide cleft, much wider than the fissure of the controlled by two genes; one is an autosomal dominant
ear-tuft mutants, at the ventral margin. Throat tufts and mutant gene ( ) and the other is a sex-linked recessive
ear-opening abnormalities are also attributed to visceral modifier (epistatic) gene ( ) that allows the expression
arch defects seen in the embryos at five days of age, as in of the in the / state. However, further genetic
the case of the ear-tuft mutants. The incidence of the studies will be necessary to confirm the mode of inherit-
defects is , greatly lower than that of the ear-tuft ance of this mutation.
mutant. Incomplete closure of the hyomandibular furrow
is also repaired in some a ected embryos. However,
abnormal morphology in the visceral arch regions is dif- ( ) (Tsudzuki and Wakasugi,
ferent between the two mutants, which leads to partial de- a): The onset of the expression of the abnormal be-
letion or irregularity in the head skeleton of the throat- havior varies from hatching to eight weeks of age. All
tuft mutant with di erent severity and incidence of each individuals that exhibit the abnormality at hatching die
bone from the case of the ear-tuft mutant (Tsudzuki and within five days. Severely a ected birds crouch and draw
Wakasugi, b). Irrespective of the di erent incidence backward wi
of each bone abnormality, the whole incidence of the head
skeletal abonormality of the throat-tuft mutant is ,
similar to that ( ) of the ear-tuft mutant (Tsudzuki
and Wakasugi, a, b). Comparative studies on the
head skeletal abnormalities in the ear-tuft and throat-tuft
mutants suggested that the gene influences the dorsal
region of the visceral archs with stronger intensity than its
allele In contrast, appears to have a greater
e ect than on developmental events associated with
the third arch and the ventral or median ventral regions of
the first and second arches (Tsudzuki and Wakasugi,
b). Interestingly, the incidence of the head skeletal ab-
normality is significantly lower in the / heterozy-
gotes than in the / and / homozygotes
(Tsudzuki and Wakasugi, ).
( ) (Nakane and Tsudzuki, b): This
mutation was found in . The mutant name is based
on the leg condition that is similar to the figure of religious
meditation ( zazen) in the Zen sect of Buddhism. This
mutant shows complete lethality at the late embryonic
stages. The -day mutant embryos cross their legs in the
front of the abdomen around the yolk sack. In about half
the mutant embryos, the crus is joined to the shank at an
abnormal angle, which results in the shank turning toward
the backside. In almost all ( ) of the mutant embryos,
the eyelid is left wide open. In addition to the leg and eye
Tt
Tt
Tt Tt
Tt
Tt Tt
hfd
hfd, hfd
hfd
hfd
Zn
moz
Zn moz moz
bkd- , bkd-
hfd
hfd . hfd
hfd
hfd hfd
hfd hfd hfd hfd
Zn, moz
Behavioral or Muscular Mutations
Back drawer
Zazen
+322
/* +1
#
-2
#
+322
##
+33* #
2221+33*
#
+33*
+33++332
+330
+/
3.
+ ,
J. Poult. Sci., ( )
th the head pressing on the floor and the beak
pointing toward the abdomen (Fig. N). Such birds
occasionally roll forward. Intermediately a ected birds
usually walk forward with neck bending downward and
occasionally draw backward or roll forward. Mildly af-
fected birds direct their heads downward slightly and
neither draw backward nor roll forward. The mild and
intermediate abnormalities appear before and/or after the
period of the severe abnormality. Some severely a ected
birds survive for a long period comparable to normal
birds, with gradual recovery from the abnormality. The
reproductive ability of the mutants is reduced in both
sexes, particularly the laying ability of females. Although
this mutation is controlled by two independent autosomal
recessive genes, Tsudzuki and Wakasugi ( a) did not
propose a gene symbol for this mutation. Later, Cheng
and Kimura ( ) assigned the symbols and
to the genes.
( ) (Mizutani, ): This behavioral mutant
shakes its neck and subsequently shows a turning move-
ment right or left. Although the cause of the abnormal
behavior is unknown, it has been revealed that in the
mutant brain the total amount of proteins is less than that
of the normal quail. This mutation is controlled by an
autosomal recessive gene. So far, no gene symbol has been
assigned to this mutation. Here, the author tentatively
assigns the symbol to this mutation.
bkd- bkd-
cr
cr
Circular
168 ./ -
+#
#
+322
+33*
,***
+,

� ��
have
been frequently utilized as biomedical animal models,
( ) (Mizutani ): the first nephrogenic diabetes insipidus mutation reported
This is an autosomal recessive mutation characterized by in Japanese quail. This mutation is inherited as an auto-
neurofilament deficiency. The quail that shows the neuro- somal recessive trait. Minvielle ( a) found the
filament deficiency (hypotrophic axonopathy) is usually quail showed excessive drinking and excessive urination,
known as quiver (Quv) quail (Yamasaki ; and carried out experiments on plasma levels and brain
Mizutani, ). The mutants exhibit quivering at the mRNA contents for avian Arg vasotocin. As a result,
neonatal stage. In mildly a ected birds, the quivering is they found that these were a ected little by the mutation,
observed only in the head and neck, whereas in severely however plasma avian Arg vasotocin was -fold higher
a ected birds it is manifested in the entire body. This and brain mRNA contents were significantly increased in
symptom is more marked when the a ected birds are both normal and mutant quail following a -hour water
frightened by some visual or auditory stimuli. Fertility, deprivation. The mutant quail shows similar performance
hatchability, and posthatch mortality are . , . , to the normal quail in the traits such as egg production
and . , respectively. Light microscopic morphometry and quality, feed intake, and gross carcass traits. Howev-
revealed that the mutant has significantly smaller cross- er, residual feed consumption is higher in the mutant.
sectional areas of the cervical spinal cord and the optic
and sciatic nerves. Furthermore, electron microscopic
morphometry indicated that in the cervical spinal cord, Among the Japanese quail mutants that are listed in
myelinated axons in the a ected birds are significantly Table or described in this article, black-at-hatch (Fig.
smaller in size than normal, though greater in density. K), glycogenesis type-II, hypotrophic axonopathy, imper-
Electron microscopically and immunohistochemically, fect albinism, muscular disorder, and silver (Fig. O)
neurofilaments are not detected in the axons or neuronal as
cell bodies (Yamasaki ). Later on, through reported in Ono and Wakasugi ( a, b), Kubota
electrophoresis and Western blot analysis, Yamasaki ( ), Satoh ( ), Shiojiri ( ), and
( ) revealed that low, middle, and high molecular Niwa ( ) for the black-at-hatch; Matsui
mass neurofilament subunits are markedly deficient in the ( ), Tsujino ( , ), Kikuchi ( ),
brain, cervical spinal cord, and sciatic nerves of the a ect- Yang ( ), Lin ( ), and McVie-Wylie
ed quail. Immunohistochemically, the spinal cord of the ( ) for the glycogenesis type-II; Zhao ( ,
mutant has no immunoreactive products corresponding to , ), Takahashi ( , ), Hasegawa
low molecular mass neurofilament. ( , ), Hirai ( a, b), Toyoshima
( ) (Braga a, b): The ( ) and Sheykholeslami ( ) for the hypotro-
mutant quail with this disorder is usually known as LWC phic axnopathy; Dkhissi ( , ) a
quail (Mizutani, ). This mutation is characterized by
generalized myotonia, muscle sti ness, and muscle weak-
ness, and is classifiable as a type of progressive muscular
disorder. A ected birds are externally identified by their
inability to lift their wings upward and by their inability to
stand upright when placed on their dorsum. These symp-
toms are clinically observed as early as days of age.
The mutant skeletal muscles include characteristic histo-
logical lesions found in sarcoplasmic masses, ringed fibers,
internal migration of nuclei, and fiber size variation.
These lesions appear first in the pectoral region and later
in the muscles of the thoracic and pelvic limbs. This
mutation is an autosomal dominant with homozygous
lethality, but no gene symbol has been assigned to this
mutation. Here, the author tentatively assigns the symbol
to this mutation.
( ) (Nakane and Tsudzuki, a): This muta-
tion was discovered in . The abnormality exhibited
by this mutant is limited to the toes. The mutants have
rigidly clenched toes in both legs at hatching, which gives
rise to a human fist-like appearance in the distal end of the
legs. Both chicks and adults can stand and walk. In spite
of such abnormality, this mutant can reproduce normally.
( ) (Minvielle a): This is
hax et al.,
et al.
et al.,
et al., et al.
et al. et al. et al.
et al. et al.
et al. et al.
et al. et al. et
al. et al.
et al. et
al. et al. et al.
Md et al., et al.
et al.
Md
cl
di et al.,
Practical Utilization of Quail Mutants
Other Mutations
Hypotrophic axonopathy
Muscular disorder
Clench
Diabetes insipidus
+33,
,**1
+33+,**,
# #+-
## ,.
,. / .- /+3 2
# + +
+
+33+ +32-+33/ +331 +333
+33, ,**,+32- +331 +332 +332
# +332 ,**,,**- +33-
+33. +33/ +33. +33/+33. +330 +333
+33/ ,*** ,**++33. +330
,**,#
#
,2
+332+33/
,**1
Tsudzuki: Mutations and Modern Genetics in Quail
nd Schrodl
( ) for the imperfect albinism; Braga ( a,
b) and Tanaka ( a, b) for the muscular disorder;
and Araki ( , ), Tsukamoto ( )
and Kawaguchi ( ) for the silver. Besides the
mutants mentioned above, other mutants are also poten-
tially powerful resources in biomedical fields to elucidate
the regulatory mechanisms for pigmentation, morpho-
genesis, nervous systems, and metabolism.
In addition to these biomedical uses, the growth and
productive ability have also been investigated for the re-
cessive white (Petek ), roux (Minvielle
, a), and yellow (Minvielle b) from
the view point of possible use in agricultural (industrial)
fields.
In , an integrated comprehensive linkage map was
constructed in chickens (Groenen ; Schmid
) on the basis of modern molecular techniques,
including linkage groups. This generally resulted from
the development of DNA markers, especially with the
development of a large number of microsatellite DNA
markers. Today, over microsatellite loci have been
mapped in the integrated map with chromosomes and
et
al. et al.
et al.
et al. et al.
et al.
et al., et al.,
et al.,
et al., et
al.,
Development of DNA Markers and Genetic Linkage Maps
169
Modern Advances of Molecular Genetics
for Japanese Quail
,**/ +33/+330
+332 ,**, +333,**+
,**.+333 ,*** ,**1
,***,***
,***/+
3**-+

between loci of . cM. Furthermore, Kayang ( )
linkage groups (Schmid ). On the other ( ) and Sasazaki ( a) assigned some Japanese
hand, in , only linkage groups ( autosoms and Z quail AFLP linkage groups to chicken chromosomes. In
chromosome) were known in Japanese quail, with a few addition, with the same reference family used by Kikuchi
loci in each linkage group, due to the absence of DNA ( ), Mannen ( ), and Sasazaki
markers (Wakasugi and Kondo, ; Ito a, b; ( a), Sasazaki ( b) mapped the orthologous
Cheng and Kimura, ; Shibata and Abe, ). genes/ESTs of chickens on the Japanese quail genome,
To obtain microsatellite markers for Japanese quail, a and made direct comparisons between the Japanese quail
few trials were performed by adapting already developed linkage groups and chicken chromosomes. However,
chicken microsatellite markers to the genome of Japanese there are some discrepancies between the studies (Kikuchi
quail (Pang ; Inoue-Murayama ; ; Mannen ; Sasazaki a, b)
Kayang ). From these trials however, it was in assigning Japanese quail AFLP linkage groups to chick-
concluded that the chicken microsatellite marker primers en chromosomes. Apart from the correctness of the as-
for polymerase chain reaction (PCR) are not e cient in signment, the AFLP linkage map, which was first reported
amplifying Japanese quail microsatellites, therefore micro- by Kikuchi ( ), has become a map of cM in
satellite markers for Japanese quail should be generated total length, with an average marker interval of . cM, by
from Japanese quail genome itself. Kayang ( , the work of Sasazaki ( b). This map is composed
, ) developed microsatellite markers directly of markers, including AFLP markers, three
from the Japanese quail genome and constructed a first- phenotypic loci, and genes/ESTs. These are assigned
generation microsatellite linkage map that included to linkage groups including W chromosome (Sasazaki
microsatellite markers (Kayang ). The total b). As mentioned earlier, there are at present
length of the map was cM, with an average spacing of two main genetic linkage maps for Japanese quail in the
cM between loci. Besides the microsatellite linkage world. It is desired that the two maps should be integrated
map, Roussot ( ) developed a number of am- in the near future.
plified fragment length polymorphism (AFLP) markers
and constructed an AFLP-based linkage map with
linkage groups ( autosomal, and Z and W chromo-
somes), in which the number of AFLP markers included, Table summarizes linkage relationships between loci
the total length of the map, and the average spacing (markers) that were revealed on the basis of modern
between markers were , cM, and . cM, respec- molecular techniques, along with classical linkage rela-
tively. Later, Kayang ( ) integrated the micro- tionships. As mentioned earlier, in there were only
satellite map of Kayang ( ) and the AFLP map three autosomal linkage groups and a Z-chromosamal
to one linkage map of . cM, with an average spacing linkage group composed of a small number of classical
mutant phenotypic loci and bi
assigned the Japanese quail chromosomes (CJA) to chick-
en chromosomes (GGA) by a comparative mapping be-
tween the two species. As a result, CJA to , , ,
, , , , and Z have been assigned to GGA to
, , , , , , , and Z, respectively.
Besides the linkage map of Kayang ( ),
Kikuchi ( ) constructed a genetic linkage map
based on a large number of AFLP markers, together with
several orthologous chicken microsatellite markers to Jap-
anese quail and two phenotipic loci, and tried some com-
parisons between the Japanese quail AFLP linkage groups
and chicken chromosomes. The total length of the AFLP
linkage map was cM with an average marker inter-
val of . cM. Subsequently, Sasazaki ( a) im-
proved the AFLP map by increasing the number of AFLP
markers. Moreover, in addition to the AFLP markers,
Sasazaki ( a) developed expressed sequence tag
(EST) markers by using chicken gene sequence informa-
tion on the web site, and located the EST markers on the
AFLP map. Mannen ( ) also developed micro-
satellite markers on the basis of the sequence of cDNA,
and mapped them on the same AFLP map. With the
mapping information of the EST markers and the micro-
satellite markers derived from cDNA, Mannen
et al.
et al., et al.
et al. et al. et al.
et al., et al.
et al., et al., et al., et al., et al.,
et al.,
et al.
et al. et al.
et al., et al.,
et al.
et al.
et al.
et al.
et al.
et al.
et al.
et al.
et al.
Linkage Groups for Phenotypic Loci, Functional Genes,
and ESTs
3 1 ,**0
,, ,**/ ,**/ ,**0,*** . -
,**/ ,**/+31- +322 ,**0 ,**0
+33* +330
+333 ,**+ ,**/ ,**/ ,**0,**-
$,**/ -+33
/ *,*** ,**0
,**, ,**. +** +33/ +3--/3
1, 00,**. ,**0
/10+*
,**-
.+-3
-
,/2 +/+0 1 0,**0 ,***
,**.3*. -
*+ *1 *3 +*+- +. +2 ,* ,1 *+*1 *3 +* +- +. +2 ,* ,1
,**0,**/
,2+0/ / ,**0
,**0
,**/
J. Poult. Sci., ( )
ochemical trait loci, and the
locus order in each linkage group, and the chromosome
number, were unclear. The linkage relationships that had
been known in are: extended brown ( ) and phos-
phoglucose isomerase ( ) (Ito a); panda ( ),
serum albumin ( ), and vitamin D binding protein
( ) (Ito b; Shibata and Abe, ); Yellow
( ) and white breasted ( ) (Cheng and Kimura, );
and imperfect albinism ( ) and sex-linked brown ( )
(Wakasugi and Kondo, ). Today, the number of
linkage groups that include phenotypic traits, genes, and
ESTs is approximately , and the chromosome number
has already been assigned to the linkage groups in most
cases. However, it is di cult to declare the exact number
of linkage groups, because there are some discrepancies in
the assignment of linkage groups to chromosomes as
mentioned before. The classical phenotypic loci that have
been mapped with DNA markers are black-at-hatch plum-
age ( ), panda plumage ( ), yellow plumage ( ), hy-
potrophic axonopathy ( ), transferring ( ), haemo-
globin ( ), and prealbumin- ( ). They, respec-
tively, were mapped on chromosomes/linkage groups CJA
(Niwa ; Miwa ), CJA (Miwa
), CJA (Miwa ; Kayang
), JQG (Kikuchi ), CJA (Miwa
E
PGI et al., s
ALB
GC et al.,
Y wb
al br
Bh s Y
hax Tf
Hb- Pa-
et al., et al.,
et al., et al., et al.,
et al., et al.,
170 ./ -
,***+322
+322 +330+33*
+31-
,*
$
+
*+ ,**- ,**/ *.,**0 ,* ,**/
,**0 */ ,**/ *3
+ +
��
�
�
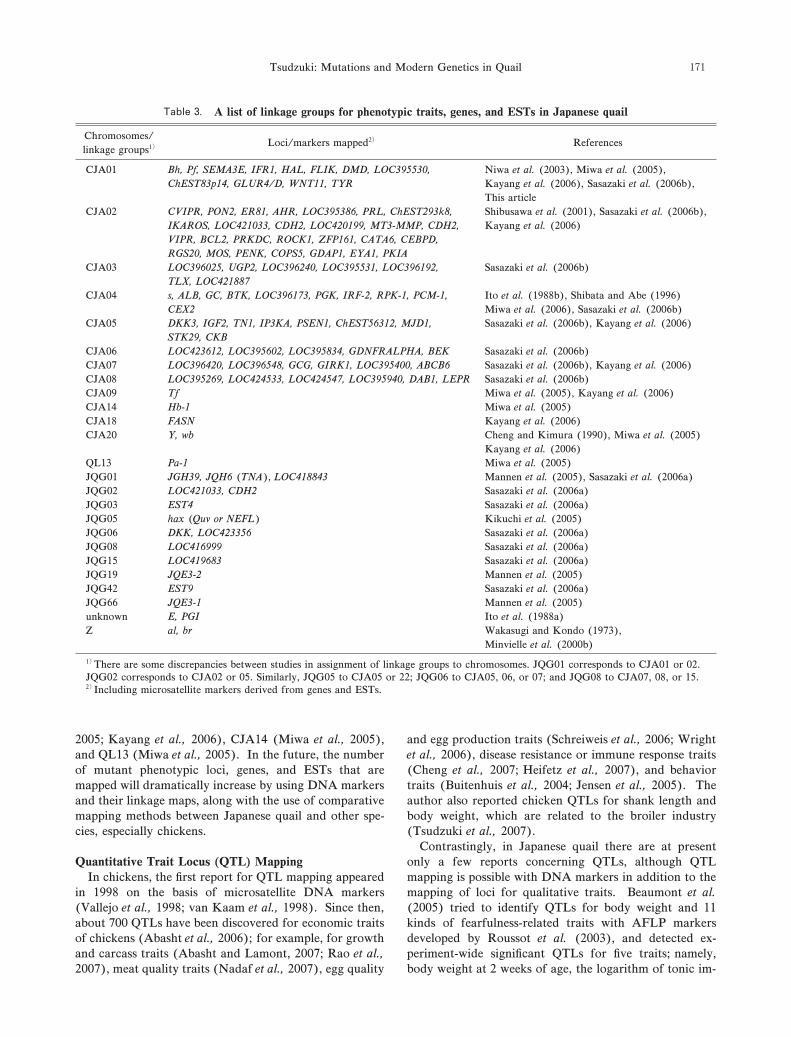
Table .
Chromosomes/Loci/markers mapped References
linkage groups
CJA Niwa ( ), Miwa ( ),
Kayang ( ), Sasazaki ( b),
This article
CJA Shibusawa ( ), Sasazaki ( b),
Kayang ( )
CJA Sasazaki ( b)
CJA Ito ( b), Shibata and Abe ( )
Miwa ( ), Sasazaki ( b)
CJA Sasazaki ( b), Kayang ( )
CJA Sasazaki ( b)
CJA Sasazaki ( b), Kayang ( )
CJA Sasazaki ( b)
CJA Miwa ( ), Kayang ( )
CJA Miwa ( )
CJA Kayang ( )
CJA Cheng and Kimura ( ), Miwa ( )
Kayang ( )
QL Miwa ( )
JQG ( ), Mannen ( ), Sasazaki ( a)
JQG Sasazaki ( a)
JQG Sasazaki ( a)
JQG ( ) Kikuchi ( )
JQG Sasazaki ( a)
JQG Sasazaki ( a)
JQG Sasazaki ( a)
JQG Mannen ( )
JQG Sasazaki ( a)
JQG Mannen ( )
unknown Ito ( a)
Z Wakasugi and Kondo ( ),
Minvielle ( b)
There are some discrepancies between studies in assignment of linkage groups to chromosomes. JQG corresponds to CJA or .
JQG corresponds to CJA or . Similarly, JQG to CJA or ; JQG to CJA , , or ; and JQG to CJA , , or .
Including microsatellite markers derived from genes and ESTs.
Tsudzuki: Mutations and Modern Genetics in Quail
; Kayang ), CJA (Miwa ), and egg production traits (Schreiweis ; Wright
and QL (Miwa ). In the future, the number ), disease resistance or immune response traits
of mutant phenotypic loci, genes, and ESTs that are (Cheng ; Heifetz ), and behavior
mapped will dramatically increase by using DNA markers traits (Buitenhuis ; Jensen ). The
and their linkage maps, along with the use of comparative author also reported chicken QTLs for shank length and
mapping methods between Japanese quail and other spe- body weight, which are related to the broiler industry
cies, especially chickens. (Tsudzuki ).
Contrastingly, in Japanese quail there are at present
only a few reports concerning QTLs, although QTL
In chickens, the first report for QTL mapping appeared mapping is possible with DNA markers in addition to the
in on the basis of microsatellite DNA markers mapping of loci for qualitative traits. Beaumont
(Vallejo ; van Kaam ). Since then, ( ) tried to identify QTLs for body weight and
about QTLs have been discovered for economic traits kinds of fearfulness-related traits with AFLP markers
of chickens (Abasht ); for example, for growth developed by Roussot ( ), and detected ex-
and carcass traits (Abasht and Lamont, ; Rao periment-wide significant QTLs for five traits; namely,
), meat quality traits (Nadaf ), egg quality body weight at weeks of age, the logarithm of tonic im-
Bh, Pf, SEMA E, IFR , HAL, FLIK, DMD, LOC , et al. et al.
ChEST p , GLUR /D, WNT , TYR et al. et al.
CVIPR, PON , ER , AHR, LOC , PRL, ChEST k , et al. et al.
IKAROS, LOC , CDH , LOC , MT -MMP, CDH , et al.
VIPR, BCL , PRKDC, ROCK , ZFP , CATA , CEBPD,
RGS , MOS, PENK, COPS , GDAP , EYA , PKIA
LOC , UGP , LOC , LOC , LOC , et al.
TLX, LOC
s, ALB, GC, BTK, LOC , PGK, IRF- , RPK- , PCM- , et al.
CEX et al. et al.
DKK , IGF , TN , IP KA, PSEN , ChEST , MJD , et al. et al.
STK , CKB
LOC , LOC , LOC , GDNFRALPHA, BEK et al.
LOC , LOC , GCG, GIRK , LOC , ABCB et al. et al.
LOC , LOC , LOC , LOC , DAB , LEPR et al.
Tf et al. et al.
Hb- et al.
FASN et al.
Y, wb et al.
et al.
Pa- et al.
JGH , JQH TNA LOC et al. et al.
LOC , CDH et al.
EST et al.
hax Quv or NEFL et al.
DKK, LOC et al.
LOC et al.
LOC et al.
JQE - et al.
EST et al.
JQE - et al.
E, PGI et al.
al, br
et al.
et al., et al., et al.,
et al., et al.,
et al., et al.,
et al., et al.,
et al.,
et al.
et al., et al.,
et al., et al.
et al.,
et al.,
A list of linkage groups for phenotypic traits, genes, and ESTs in Japanese quail
Quantitative Trait Locus (QTL) Mapping
171
-
,+
+
,
*+ ,**- ,**/,**0 ,**0
*, ,**+ ,**0,**0
*- ,**0
*. +322 +330,**0 ,**0
*/ ,**0 ,**0
*0 ,**0*1 ,**0 ,**0*2 ,**0*3 ,**/ ,**0+. ,**/+2 ,**0,* +33* ,**/
,**0+- ,**/
*+ ,**/ ,**0*, ,**0*- ,**0*/ ,**/*0 ,**0*2 ,**0+/ ,**0+3 ,**/., ,**000 ,**/
+322+31-
,***
*+ *+ *,*, *, */ */ */ ,, *0 */ *0 *1 *2 *1 *2 +/
,**/ ,**0 +. ,**/ ,**0+- ,**/ ,**0
,**1 ,**1,**. ,**/
,**1
+332+332 +332 ,**/ ++
1**,**0 ,**-
,**1,**1 ,**1 ,
- + -3//-*2- +. . ++
, 2+ -3/-20 ,3- 2.,+*-- , .,*+33 - ,
, + +0+ 0,* / + +-30*,/ , -30,.* -3//-+ -30+3,
.,+221-30+1- , + +
,- , + - + /0-+, +,3.,-0+, -3/0*, -3/2-.-30.,* -30/.2 + -3/.** 0-3/,03 .,./-- .,./.1 -3/3.* +
+
+-3 0 .+22.-.,+*-- ,.
.,--/0.+0333.+302-- ,3- +

� �
mobility, the number of indictions, the number of jumps, The extended brown mutant shows blackish (or dark
and dejections. Furthermore, Minvielle ( b) brownish) plumage throughout the body (Somes, ;
carried out QTL analysis using microsatellite markers Truax and Johnson, ). Nadeau ( ) se-
developed by Kayang ( ) for traits responsible quenced an bp segment of the -bp single coding
for growth, feed consumption, egg production, tonic im- exon of melanocortin receptor ( ) gene of the
mobility, and body temperature. They found experiment- extended brown from a French flock, and observed perfect
wide significant QTLs for five traits, egg clutch length, association between a non-synonymous substitution (Glu
eggshell weight, egg number, feed intake, and residual Lys) and the mutant plumage. The “black” plumage
feed intake. color mutation (Chikamune and Kanai, ; Somes,
) also exhibits similar phenotype to the extended
brown. Nadeau ( ) also sequenced extended
With the exception of two examples, the causative genes brown from a Japanese flock, and found the same
for mutant phenotypes of Japanese quail were unclear at substitution as in the case of the extended brown from a
the molecular level in the th century, although detection French flock. The “extended brown” from a Japanese
of causative genes has continually been reported on and flock examined by Nadeau ( ) is thought to be
after . The first and second Japanese quail mutations, the “black” mutant discovered in Japan. Thus, it is very
on which molecular basis of the abnormality was dis- likely that the extended brown and black are the same
covered in the last decade of the th century, are “hy- mutants.
potrophic axonopathy ( )” (Mizutani ) and Pansy ( ), which exhibits tri-colored mutant plum-
“silver ( )” (Homma ). age, is a bottom recessive allele at the black ( ) locus
The mutant quail is characterized by quivering (Tsudzuki and Wakasugi, ; Tsudzuki ).
behavior and neurofilament deficiency in the axons of Additionally, it has been revealed that the pansy has a
the cervical spinal cord and the optic and sciatic nerves mutation in the locus (Wada and Tsudzuki, un-
(Yamasaki ). Ohara ( ) found a non- published data; accession no. AB in DDBJ). The
sense mutation in the neurofilament-L (NF-L) gene of the pansy mutant has insertion of two extra nucleotides (CG)
mutant quail. A point mutation from C to T resides at codon (nt insCG), which leads to a premature stop
at the nucleotide residue , resulting in a stop codon at codon and produces truncated transcript by a shift of the
the position corresponding to amino acid residue of reading frame. The redhead mutant ( ) (Truax
the NF-L protein. The expression level of NF-L mRNA ) is similar to the pansy in the phenotype, and it is
in the brain drastically decreases to of that in phenotypically unclear whether the two are identical
the wild-type quail. mutants. It is desired to investigate mutation in
The homozygote for the silver mutation ( / ) has pure the redhead, which will precisely determine the sameness
white plumage and eye defects (Homma ). In or di erence between the two mutations.
the / eyes, transdi erentiation from the retinal pigment Panda ( ) (Mizutani ) and do
epithelium into the neural retina occurs at the central
area (Fuji and Wakasugi, ). Mochii ( )
indicated that the microphthalmia-associated transcrip-
tion factor ( ) gene mutation is responsible for the
silver mutation. They revealed that there are two kinds of
nucleotide changes leading to amino acid changes in the
gene by complete sequencing of the cDNA from the
/ retinal pigment epithelial cells. One leads to a substi-
tution from histidine to arginine in the basic region of
MITF protein, and the other is a two-base deletion result-
ing in frame shift and a premature termination of the
polypeptide in the region following the zipper region. In
spite of having such mutations, the retinal pigment epithe-
lial cells of the / embryos express normal size
mRNA, but do not express normal MITF protein, which
causes a partial loss of function of the transactivation of
the promoter of the -kDa melanosomal matrix protein
( ) gene, a kind of pigment-cell-specific genes.
Other Japanese quail mutants, for which the causative
molecular mutations were revealed in this century, are
extended brown ( ), pansy ( ), panda ( ), imperfect
albinism ( ), sex-linked cinnamon ( ), roux ( ),
yellow ( ), and recessive black ( ).
ps
rh
ps
c
et al.
et al.
et al.
MC R
i.e.
et al.
MC R
et al.
hax et al., d
B et al., D
hax et al.,
MC R
et al., et al.
hax
e et al.,
hax
MC R
B B
et al.,
B B s et al.,
et al.
Mitf
Mitf
B B
B B Mitf
mmp
E d s
al al BR*R
Y Y*RB
Causative Genes for Mutant Phenotypes
,**/ +313+313 ,**0
,**. +- 2/ 3./+
3,+312
+32.,**0
,*,**0
,**0
,*+33,
+303+321 +33*
+33+ +33- ,*+0-0
,0 10-/,
++.+313
/
+303 ## +31.
+33- +332
++/
+
+
+
+
++/
�
J. Poult. Sci., ( )
tted white ( )
(Tsudzuki ) mutants show white plumage all
over the body with colored spots. The panda is dominant
over the dotted white (Tsudzuki ). Based on
the mapping result of the panda ( ) locus on Japanese
quail chromosome (CJA ) and the comparative mapp-
ing between Japanese quail and chickens, Miwa
( ) first suggested the endothelin receptor B (
) gene to be a candidate gene for the locus. According
to this finding, Miwa ( ) subsequently confirmed
that the causative gene for the locus is The
panda phenotype is associated with a missense mutation at
the nucleotide position (c. G A) in the
which leads to amino acid substitution (arginine to
histidine change) at the position (p. R H). On the
other hand, no amino acid substitution was found for the
dotted white phenotype. The expression level of the
mRNA was lower and drastically lower in the
panda and dotted-white mutants respectively, than in the
wild-type quail. In spite of the depression of
mRNA expression level and the amino acid change in
EDNRB protein, no significant di erence was observed
in the number or migration pattern of neural crest cells
between the wild-type and panda quail in . -day embryos.
dws
et al.,
et al.,
s
et al.
EDNRB
s
et al.
s EDNRB .
EDNRB
,
EDNRB
EDNRB
172 ./ -
+33,
+33-
. .
,**0
,**1
33/ 33/
--, --,
, #
, /
,
,
,
,
,

� �
�
�
� � �
�
�
Abasht B, Dekkers JCM and Lamont SJ. Review of quantitative
trait loci identified in the chicken. Poultry Science, :
. .
Abasht B and Lamont SJ. G
molecular level. In future, more numbers of DNA markers
Imperfect albinism ( ) and sex-linked cinnamon ( ) a cysteine residue, and removed the normal stop codon.
(Fig. P) are plumage color mutations and alleles at the On the other hand, the fawn- had two synonymous single
locus on Z chromosome. The is dominant over the nucleotide polymorphisms (c. T C and c. C T),
(Cheng and Kimura, ). Gunnarsson ( ) which raised the possibility that there might be another
found that the solute carrier family , member , protein regulatory mutation in the fawn- as seen in the yellow.
( ) gene, is the causative gene for the mutations The expression level of the recessive black is sig-
at the locus. In the imperfect albinism, a transversion nificantly lower than that of the wild type, whereas that of
from G to T resides at the splice acceptor site just preced- the fawn- is drastically higher than the wild-type and
ing exon . The splice site mutation leads to an in-frame yellow quail. The wild-type and yellow quail showed a
skipping of exon of and amino acids are similar level of expression of to each other as ob-
missing in the mature protein. Gunnarson ( ) served by Nadeau ( ).
also observed an association of a transition from C to A at As mentioned earlier, genetic studies of Japanese quail
nt (Ala Asp) in exon of with the sex- accelerated at the beginning of the st century at the
linked cinnamon allele.
The roux ( ) is a recessive allele to the brown and genes both for qualitative and quantitative traits will
mutation at the brown ( ) locus on Z chromosome, and be mapped on the further integrated map, and further
exhibits slightly lighter brown color than the brown numbers of causative genes for mutant phenotypes will be
(Wakasugi and Kondo, ; Minvielle ). revealed. Such work will raise the value of Japanese quail
Nadeau ( ) revealed that the roux phenotype is to a higher level as a research model in the biomedical
expressed by the non-synonymous point mutation that fields, and as a bird of economic importance.
occurred at the nucleotide position (c. T C) of
the tyrosinase-related protein ( ) gene. The T
C results in a change of phenylalanine to serine at amino The author is deeply indebted to T. Goto, S. Onitsuka,
acid (p. Phe Ser). The expression level of and M. Yoshida for their great help in preparation of the
is the same between the dorsal skins of the wild-type and manuscript. The author is also grateful to F. Kanyinji for
roux quail, suggesting that the mutation is not due to a proof reading the manuscript. The author further wishes
regulatory mutation. to express gratitude to T. Maeda and Y. Yoshimura for
Yellow is a plumage color mutant that shows wheat- their encouragement. The author, M. Tsudzuki, was
straw color throughout the body with basically the same awarded the Scientist Prize of Japan Poultry Science
markings as the wild type (Homma, ). Nadeau Association for genetic studies on Japanese quail mutants.
( ) revealed that the yellow is a mutation of the reg-
ulatory region of agouti signaling protein ( ) gene,
and is not a mutation caused by coding sequence variation
at The yellow mutation lacks almost the entire
coding sequence of two upstream loci, hnRNP associated
with lethal yellow ( ) and eukaryotic translation
initiation factor B ( ), which results in the produc-
tion of a new transcript, because expression is con-
trolled by the promoter. These genotypic condi-
tions in the yellow mutant quail are quite similar to those
in the yellow mutant ( ) mouse (Michaud ).
Unlike the yellow mouse however, no significant di erence
was observed in the ASIP mRNA expression level in the
skins of the yellow and wild-type quail (Nadeau
).
Besides the study of Nadeau ( ) for the yellow
mutant quail, Hiragaki ( ) also investigated the
coding sequence of the gene in the yellow, fawn-
(Tsudzuki ), and recessive black (Fujiwara
) mutants and wild-type quail. The three mutants
are alleles at the locus, and the order of dominance is
fawn- yellow wild type recessive black (Tsudzuki
; Hiragaki ). The yellow mutant
showed no variation in the coding region, which is consis-
tent with the result of Nadeau ( ). The recessive
black showed -bp deletion (c. del), causing a
frame shift that changes the last six amino acids, including
c
c
y
al al
al
al al
et al.
SLC A ASIP
al
SLC A , ASIP
et al. et al.
SLC A
BR*R
br
et al.,
et al.
TYRP
TYRP
et al.
ASIP
ASIP.
RALY
EIF B
ASIP
RALY
A et al.,
et al.,
et al.
et al.
ASIP
et al., et
al.,
Y
et al., et al.,
et al.
Acknowledgments
References
2/,*13 ,*30 ,**0
+ ,-+ +**
+33* ,**1./ , ,
,.
. .1,**1 ,**2
,21 1, + ,+
+31- ,**-,**1
2./ 2./+ 2./
,2, ,2,
,**0+301
,**2
,
+33-#
,**2,**2
,**2,
+330,**/
,+330 ,**2
,**22 -1- -2*
./ ,
./ ,
./ ,
+
+
, �
�
�
�
Tsudzuki: Mutations and Modern Genetics in Quail
enome-wide association analysis
reveals cryptic alleles as an important factor in heterosis for
fatness in chicken F population. Animal Genetics, :
. .
Araki M, Yamao M and Tsudzuki M. Early embryonic interac-
tion of retinal pigment epithelium and mesenchymal tissue
induces conversion of pigment epithelium to neural retinal
fate in the silver mutation of the Japanese quail. Develop-
ment Growth and Di erentiation, : . .
Araki M, Takano T, Uemonsa T, Nakane Y, Tsudzuki M and
Kaneko T. Epithelia-mesenchyme interaction plays an essen-
tial role in transdi erentiation of retinal pigment epithelium
of silver mutant quail: localization of FGF and related
molecules and aberrant migration pattern of neural crest
cells during eye rudiment formation. Developmental Biolo-
gy, : . .
Beaumont C, Roussot O, Feve K, Vignoles F, Leroux S, Pitel F,
Faure JM, Mills AD, Guemene D, Sellier N, Mignon-
Grasteau S, Le Roy P and Vignal A. A genome scan with
AFLP markers to detect fearfulness-related QTLs in Japa-
nese quail. Animal Genetics, : . .
Braga IS rd, Tanaka S, Kimura T, Itakura C and Mizutani M.
Inherited muscular disorder in mutant Japanese quail
( ): Relationship between the de-Coturnix coturnix japonica
173
, -2 .3+.32 ,**1
# .* +01 +10 +332
#
,.. -/2 -1+ ,**,
-0 .*+ .*1 ,**/-

� �
��
��
��
� �
��
��
��
�
�
�
�
�
�
�
�
�
velopment of muscle lesions and age. Journal of Compara- barb: a feather structure mutation in Japanese quail. Poultry
tive Pathology, : . a. Science, : . c.
Braga IS rd, Oda K, Kikuchi T, Tanaka S, Shin Y, Sento M, Fulton JE, Jurilo DM, Cheng KM and Nichols CR. Defective
Itakura C and Mizutani M. A new inherited muscular feathers in Japanese quail: a two-locus model for a new trait.
disorder in Japanese quails ( ). The Journal of Heredity, : . .
Veterinary Pathology, : . b. Groenen MAM, Cheng HH, Bumstead N, Benkel BF, Briles
Buitenhuis AJ, Rodenburg TB, Siwek M, Cornelissen SJB, WE, Burke T, Burt DW, Crittenden LB, Dodgson J, Hillel
Nieuwland MGB, Crooijmans RPMA, Groenen MAM, J, Lamont S, de Leon AP, Soller M, Takahashi H and Vignal
Koene P, Bovenhuis H and van der Poel JJ. Identification of A. A consensus linkage map of the chicken genome.
QTLs involved in open-field behavior in young and adult Genome Research, : . .
laying hens. Behavior Genetics, : . . Gunnarsson U, Hellstrom AR, Tixier-Boichard M, Minvielle F,
Cheng HH, Zhang Y and Muir WM. Evidence for widespread Bed’hom B, Ito S, Jensen P, Rattink A, Vereijken A and
epistatic interactions influencing Marek’s disease virus Andersson L. Mutations in cause plumage color
viremia levels in chicken. Cytogenetic and Genome Re- variation in chicken and Japanese quail. Genetics, :
search, : . . . .
Cheng KM and Kimura M. Mutations and major variants in Hardiman JW, Collins WM and Urban WE Jr. Red egg-shell
Japanese quail. In: Poultry Breeding and Genetics color: a dominant mutation in Japanese quail. The Journal
(Crawford RD ed.) pp. . Elsevier. Amsterdam. of Heredity, : . .
. Hasegawa H, Murayama T, Takahashi A, Itakura C and
Chikamune T and Kanai Y. Studies on white-feathered and dark- Nomura Y. The monoamine content of the brain in the
feathered Japanese quail. . The relationship with wild neurofilament-deficient quail, (the quiver quail). Biological
plumage color (in Japanese with English summary). Japa- and Pharmaceutical Bulletin, : . .
nese Poultry Science, : . . Hasegawa H, Murayama T, Takahashi A, Itakura C and
Collins WM, Abplanalp H and Yoshida S. Early embryonic Nomura Y. Changes of GTP binding proteins, not neuro-
chondrodystrophy in Japanese quail. The Journal of Hered- filament-associated proteins, in the brain of the neuro-
ity, : . . filament-deficient quail, “Quiver”. Neurochemistry Interna-
Crawford RD. Origin and history of poultry species. In: Poultry tional, : . .
Breeding and Genetics (Crawford RD ed.) pp. . Hays FA. Chondrodystrophy in Rhode Island reds. The Ameri-
Elsevier. Amsterdam. . can Naturalist, : . .
Crittenden LB, Bitgood JJ, Burt DW, Ponce de Leon FA and Heifetz EM, Fulton JE, O’Sullivan NP, Arthur JA, Wang J,
Tixier-Biochard M. Nomenclature for naming loci, alleles, Dekkers JCM and Soller M. Mapping quantitative trait loci
linkage groups and chromosomes to be used in poultry a ecting susceptibility to Marek’s disease virus in a back-
genome publications and databases. Genetics Selection Evo- cross
lution, : . .
Dkhissi O, Dalil-Thiney N and Minvielle F. Retinal distribution
of tyrosine hydroxylase immunoreactive cells in two strains
of quails Journal fur
Hirnforschung, : . .
Dkhissi O, Chanut E, Versaux-Botteri C, Minvielle F, Trouvin
JH and Nguyen-Legros J. Changes in retinal dopaminergic
cells and dopamine rhythmic metabolism during the devel-
opment of a glaucoma-like disorder in quails. Investigative
Ophthalmology and Visual Science, : . .
Dodson DL and Coleman TH. Crooked neck: an inherited con-
dition in Japanese quail. Poultry Science, : . .
Dodson DL and Coleman TH. Arostroperocephaly: a lethal con-
dition in Japanese quail. Poultry Science, : .
.
Fuji J and Wakasugi N. Transdi erentiation from retinal pig-
ment epithelium into neural retina due to plumage
mutant gene in Japanese quail. Development. Growth and
Di erentiation, : . .
Fujiwara A, Mizutani M, Ono T and Kagami H. “Recessive
black”: A plumage color mutant in Japanese quail. Journal
of Poultry Science, : . .
Fulton JE, Roberts CW and Cheng KM. Cinnamon: a mutant of
Japanese quail. Canadian Journal of Genetics and Cytology,
: . a.
Fulton JE, Roberts CW and Cheng KM. “Porcupine”: a feather
structure mutation in Japanese quail. Poultry Science, :
. b.
Fulton JE, Roberts CW, Nichols CR and Cheng KM. Short
Coturnix coturnix japonica
SLC A
Coturnix coturnix japonica.
Silver
++- +-+ +.- +33/ 0+ ,-+3 ,-,+ +32,- #
1. +2. +22 +32--, -/+ -0* +33/
+* +-1 +.1 ,***-. -,/ --- ,**.
+1/ 201++1 -+- -+2 ,**1 211 ,**1
--- -0, 00 +.+ +.- +31/+33*
++1 +*/- +*// +33.
+/ ,-0 ,.+ +312
/3 ,.2 ,/* +302,2 ,,+ ,,3 +330
+ .++33* 12 /. /2 +3..
#
,2 ,23 ,31 +330
-/ ,0- ,02 +33.
-1 ,--/ ,-.. +330
/+ 2, 2/ +31,
/, +0/. +0/1+31-
#
# -/ .21 .3. +33-
., 0. 03 ,**/
,. +0- +00 +32,
0+.,3 .-- +32,
./ ,
�
�
�
�
�
�
�
�
J. Poult. Sci., ( )
population of layer chickens. Genetics, :
. .
Hermes JC, Abbott UK, Johnston E and Owens M. A new
chondrodystrophy mutation in Japanese quail (
). The Journal of Heredity, : . .
Hill WG, Lloyd GL and Abplanalp H. Micromelia in Japanese
quail: an embryonic lethal mutant. The Journal of Heredity,
: . .
Hiragaki T, Inoue-Murayama M, Miwa M, Fujiwara A,
Mizutani M, Minvielle F and Ito S. Recessive black is allelic
to the yellow plumage locus in Japanese quail and associated
with a frameshift deletion in the gene. Genetics, :
. .
Hirai T, Mizutani M, Kimura T, Ochiai K, Uemura T and
Itakura C. Neurotoxic e ects of , -hexanedione on normal
and neurofilament-deficient quail. Toxicologic Pathology,
: . a.
Hirai T, Mizutani M, Ochiai K, Umemura T and Itakura C.
Distal axonopathy does not occur without neurofilament
accumulation in gamma-diketone neuropathy: comparative
studies of normal and neurofilament-deficient quail. Acta
Neuropathologica, : . b.
Homma K, Shumija S and Jinno M. Yellow feathered Japanese
quail ( ) (in Japanese with English
summary). The Japanese Journal of Zootechnical Science,
: . .
Homma K, Jinno M, Sato K and Ando A. Studies on perfect and
imperfect albinism in the Japanese quail (
) (in Japanese with English summary). The Japa-
nese Journal of Zootechnical Science, : . .
Coturnix
japonica
ASIP
Coturnix coturnix japonica
Coturnix coturnix
japonica
174 ./ -
+11 ,.+1,.-+ ,**1
2+ ,,, ,,. +33*
/. +22 +3* +30-
+1211+ 11/ ,**2
# , /
,1 -.2 -/- +333
31 //, //0 +333
-2 +0- +00 +301
-3 -.2 -/, +302

�
��
��
�
�
�
�
��
��
� �
� �
�
�
�
�
�
�
Homma K, Jinno M and Kito J. Studies on silver feathered ation in Japanese quail and helmeted guinea fowl. Animal
Japanese quail (in Japanese). The Japanese Journal of Science Journal, : . .
Zootechnical Science, : . . Kayang BB, Vignal A, Inoue-Murayama M, Miwa M,
Homma K. Japanese quail in biological research (in Japanese). Monvoisin JL, Ito S and Minvielle F. A first-generation
Japanese Poultry Science, : . . microsatellite linkage map of the Japanese quail. Animal
Homma K, Oki H and Watanabe G. A plumage color mutation Genetics, : . .
in Japanese quail associated with female specific sterility due Kayang BB, Fillon V, Inoue-Murayama M, Miwa M, Leroux S,
to oviduct dysfunction. Japanese Journal of Animal Repro- Feve K, Monvoisin JL, Pitel F, Vignoles M, Mouilhayrat C,
duction, : . . Beaumont C, Ito S, Minvielle F and Vignal A. Integrated
Inoue-Murayama M, Kayang BB, Kimura K, Ide H, Nomura A, maps in quail ( ) confirm the high degree of
Takahashi H, Nagamine Y, Takeda T, Hanada H, Tatsuda synteny conservation with chicken ( ) despite
K, Tsudzuki M, Matsuda Y, Mizutani M, Murayama Y and million years of divergence. BMC Genomics, : . .
Ito S. Chicken microsatellite primers are not e cient Kikuchi S, Fujima D, Sasazaki S, Tsuji S, Mizutani M, Fujiwara
markers for Japanese quail. Animal Genetics, : . A and Mannen H. Construction of a genetic linkage map of
. Japanese quail ( ) based on AFLP and
Ito S, Kimura M and Isogai I. A sex di erence in recombination microsatellite markers. Animal Genetics, : . .
values between extended brown and phosphoglucose Kikuchi T, Yang HW, Pennybacker M, Ichihara N, Mizutani M,
isomerase loci in Japanese quail. The Japanese Journal of Hove JLK and Chen YT. Clinical and metabolic correction
Zootechnical Science, : . a. of pompe disease by enzyme therapy in acid maltase-
Ito S, Kimura M and Isogai I. Linkage between panda plumage deficient quail. The Journal of Clinical Investigation, :
and albumin loci in Japanese quail. The Japanese Journal of . .
Zootechnical Science, : . b. Kimura M. Polymorphic proteins and enzymes in Japanese quail
Ito S, Tsudzuki M, Komori M and Mizutani M. Celadon: an a review (in Japanese). Japanese Poultry Science, :
eggshell color mutation in Japanese quail. The Journal of . .
Heredity, : . . Kubota Y, Nakamura A, Ito M and Shiojiri N. (black at
Ito S and Tsudzuki M. Light down-dominant plumage color hatch) gene appears to cause whole-body hemorrhage in
mutation with homozygous lethality in Japanese quail. The homozygous embryos of Japanese quail (
Journal of Heredity, : . . ). The Journal of Experimental Zoology, :
Ito S and Tsudzuki M. Orange: a plumage color mutation ac- . .
companied by semi-lethality in Japanese quail. The Journal Lauber JK. Sex-linked albinism in the Japanese quail. Science,
of Heredity, : . . : . .
Jensen P, Keeling L, Schutz K, Andersson L, Mormede P, Lin CY, Ho CH, Hsieh YH and Kikuchi T. Adeno-associated
Brandstrom H, Forkman B, Kerje S, Fredriksson R, virus-mediated transfer of human aci
Ohlsson C, Larsson S, Mallmin H and Kindmark A. Feather
pecking in chickens is genetically related to behavioural and
developmental traits. Physiology and Behavior, : .
.
Kawaguchi N, Ono T, Mochii M and Noda M. Spontaneous
mutation in gene causes osteopetrosis in silver homozy-
gote quail. Developmental Dynamics, : . .
Kawahara T. Body traits and behaviour in wild Japanese quail
(in Japanese with English sum-
mary). Tori (Bulletin of the Ornithological Society of
Japan), : . .
Kawahara T. A dark frayed feather nervous disorder mutant of
Japanese quail. Annual Report of the National Institute of
Genetics, Japan : . .
Kawahara T. Nervous disorder mutants in the poultry, especially
in Japanese quail (in Japanese). Experimental Animals, :
. .
Kayang BB, Inoue-Murayama M, Nomura A, Kimura K,
Takahashi H, Mizutani M and Ito S. Fifty microsatellite
markers for Japanese quail. The Journal of Heredity, :
. .
Kayang BB, Inoue-Murayama M, Hoshi T, Matsuo K,
Takahashi H, Minezawa M, Mizutani M and Ito S. Micro-
satellite loci in Japanese quail and cross-species amplification
in chicken and guinea fowl. Genetics Selection Evolution,
: . .
Kayang BB, Inoue-Murayama M, Takahashi H, Minezawa M,
Tsudzuki M, Mizutani M and Ito S. Twenty-eight new
microsatellite loci in chicken and their cross-species amplific-
Coturnix japonica
Gallus gallus
Coturnix japonica
Bh
Coturnix coturnix
japonica
Mitf
Coturnix coturnix japonica
1. ,// ,/3 ,**-.* +,3 +-* +303
1 +/1 +0/ +31*-/ +3/ ,** ,**.
-+ 2. 23 +32/
-/1 +*+ ,**0
$-, 1 ++
,**+# -0 ,,1 ,-+ ,**/
/3 2*+ 2*/ +322+*+
2,1 2-- +332/3 2,, 2,. +322
+3 ,++,,+ +32,
2. +./ +.1 +33-
2. ,,, ,,/ +33- ,1+ ..+./+ +33/
2/ /. /0 +33. +.0 3.2 3/* +30.
20 /, 0*,**/
,,* +-- +.* ,**+
,1 +*/ ++, +312
,3 +** +313
,33- 32 +32*
3+/*, /*/ ,***
-. ,-- ,/- ,**,
�
�
�
�
�
�
Tsudzuki: Mutations and Modern Genetics in Quail
d maltase gene results
in a transient reduction of glycogen accumulation in muscle
of Japanese quail with acid maltase deficiency. Gene Thera-
py, : . .
Mannen H, Murata K, Kikuchi S, Fujita D, Sasazaki S, Fujiwara
A and Tsuji S. Development and mapping of microsatellite
markers derived from cDNA in Japanese quail (
). The Journal of Poultry Science, : .
.
Matsui T, Kuroda S, Mizutani M, Kiuchi Y, Suzuki K and Ono
T. Generalized glycogen storage disease in Japanese quail
( ). Veterinary Pathology, :
. .
McVie-Wylie AJ, Ding EY, Lawson T, Serra D, Migone FK,
Pressley D, Mizutani M, Kikuchi T, Chen YT and
Amalfitano A. Multiple muscles in the AMD quail can be
“cross-corrected” of pathologic glycogen accumulation after
intravenous injection of an [E -, polymerase-] adenovirus
vector encoding human acid-alpha-glucosidase. The Journal
of Gene Medicine, : . .
Michaud EJ, Bultman SJ, Stubbs LJ and Woychik RP. The
embryonic lethality of homozygous lethal yellow mice ( /
) is associated with the disruption of a novel RNA-binding
protein. Genes and Development, : . .
Minezawa M and Wakasugi N. Studies on a plumage mutant
(black at hatch) in the Japanese quail. The Japanese Journal
of Genetics, : . .
Minvielle F, Hirigoyen E and Boulay M. Associated e ects of the
roux plumage color mutation on growth, carcass traits, egg
production, and reproduction of Japanese quail. Poultry
y
y
Coturnix
japonica
Coturnix coturnix japonica
A
A
175
3 //. /0- ,**,
., ,0- ,1+,**/
,*-+, -,+ +32-
+
/ -33 .*0 ,**-
1 +,*- +,+- +33-
/, +2- +3/ +311#

� �
� �
�
�
��
�
�
�
�
�
�
�
�
�
�
�
Science, : . . imental Animals, : . .
Minvielle F, Gandemer G, Maeda Y, Leborgne C, Hirigoyen E Mochii M, Ono T, Matsubara Y and Eguchi G. Spontaneous
and Boulay M. Carcase characteristics of a heavy Japanese transdi erentiation of quail pigmented epithelial cell is ac-
quail line under introgression with the roux gene. British companied by a mutation in the gene. Developmental
Poultry Science, : . a. Biology, : . .
Minvielle F, Ito S, Inoue-Murayama M, Mizutani M and Morse KA and Abplanalp H. A heritable head abnormality in
Wakasugi N. Genetic analyses of plumage color mutations chicken x coturnix hybrids. Poultry Science, : . .
on the Z chromosome of Japanese quail. The Journal of Nadaf J, Gilbert H, Pitel F, Berri CM, Feve K, Beaumont C,
Heredity, : . b. Duclos MJ, Vignal A, Porter TE, Simon J, Aggrey SE,
Minvielle F, Gourichon D and Monvoisin JL. Testing homology Cogburn LA and Le Bihan-Duval E. Identification of QTL
of loci for two plumage colors, “lavender” and “recessive controlling meat quality traits in an F cross between two
white”, with chicken and Japanese quail hybrids. The Jour- chicken lines selected for either low or high growth rate.
nal of Heredity, : . . BMC Genomics, : . .
Minvielle F, Gourichon D and Monvoisin JL. E ects of two- Nadeau NJ, Minvielle F and Mundy NI. Association of a Glu
locus combinations, using the roux, lavender, and beige Lys substitution in with extended brown in Japanese
mutations, on plumage color of Japanese quail. The Journal quail ( ). Animal Genetics, : .
of Heredity, : . . .
Minvielle F. The future of Japanese quail for research and Nadeau NJ, Mundy NI, Gourichon D and Minvielle F. Associa-
production. World’s Poultry Science Journal, : . tion of a single-nucleotide substitution in with roux
. in Japanese quail ( ). Animal Genetics, :
Minvielle F, Gourichon D and Moussu C. Two new plumage . .
mutations in the Japanese quail: “curly” feather and “rusty” Nadeau NJ, Minvielle F, Ito S, Inoue-Murayama M, Gourichon
plumage. BMC Genetics, : . a. D, Follett SA, Burke T and Mundy NI. Characterization of
Minvielle F, Kayang BB, Inoue-Murayama M, Miwa M, Vignal Japanese quail yellow as a genomic deletion upstream of the
A, Gourichon D, Neau A, Monvoisin JL and Ito S. avian homolog of the mammalian (agouti) gene. Ge-
Microsatellite mapping of QTL a ecting growth, feed con- netics, : . .
sumption, egg production, tonic immobility and body tem- Nakane Y and Tsudzuki M. Clench: a new autosomal recessive
perature of Japanese quail. BMC Genomics, : . b. mutation in Japanese quail. The Journal of Heredity, :
Minvielle F, Grossmann R and Gourichon D. Development and . a.
performances of a Japanese quail line homozygous for the Nakane Y and Tsudzuki M. A new morphogenetic lethal muta-
diabetes insipidus ( ) mutation. Poultry Science, : tion showing eye, beak, and leg abnormalities in Japanese
. a. quail. In: Proceedings of the th World Conference on
Minvielle F, Gourichon D, Ito S, Inoue-Murayama M and Animal Production. Vol.II. pp
Riviere S. E ects of the dominant lethal yellow mutation on
reproduction, growth, feed consumption, body temperature,
and body composition of the Japanese quail. Poultry Sci-
ence, : . b.
Miwa M, Inoue-Murayama M, Kayang BB, Vignal A, Minvielle
F, Monvoisin JL, Takahashi H and Ito S. Mapping of
plumage colour and blood protein loci on the microsatellite
linkage map of the Japanese quail. Animal Genetics, :
. .
Miwa M, Inoue-Murayama M, Kobayashi N, Kayang BB,
Mizutani M, Takahashi H and Ito S. Mapping of panda
plumage color locus on the microsatellite linkage map of the
Japanese quail. BMC Genetics, : . .
Miwa M, Inoue-Murayama M, Aoki H, Kunisada T, Hiragaki T,
Mizutani M and Ito S. Endothelin receptor B ( )
is associated with the panda plumage colour mutation in
Japanese quail. Animal Genetics, : . .
Mizutani M, Chino K, Umezawa H and Kuramasu S. Genetic
analysis of a new plumage-panda in Japanese quail (in
Japanese). Experimental Animals, : . .
Mizutani M, Nunoya T, Yamasaki H and Itakura C. The
hypotrophic axonopathy mutant in Japanese quail. The
Journal of Heredity, : . .
Mizutani M. Inbred strains, closed clonies, and mutant strains
(animal models) in chicken and Japanese quail maintained
in NIBS (in Japanese). Journal of Animal Genetics, :
. .
Mizutani M. Establishment of inbred strains of chicken and
Japanese quail and their potential as animal models. Exper-
Mitf
MC R
Coturnix japonica
TYRP
Coturnix japonica
ASIP
di
EDNRB
,
12 +.13 +.2. +333 /+ .+1 .,3 ,**,
#
.+ .+ ./ ,*** +30 +./ +/3 +332
.3 +.+3 +31*
3+ .33 /*+ ,***
3- 1- 10 ,**, 2 +// ,**1# 3,
-1 ,21 ,233. /+1 /,, ,**- ,**0
0* /** /*1,**. -2
0*3 0+- ,**1
0 +. ,**/
# +12 111 120 ,**2
0 21 ,**/ 23-.1 -.2 +332
20 ,.3,/. ,**1 2
#
20 +0.0 +0/* ,**1
-0-30 .** ,**/
1 , ,**0
,
-2 +*- +*2 ,**1
,- /3 0+ +31.
2- ,-. ,-/ +33,
,2 .//2 ,***
+
+
,
�
�
�
�
�
�
�
for a new mutant of Japanese quail. In: Proceedings of the th
J. Poult. Sci., ( )
. . b.
Nakane Y and Tsudzuki M. Morphological and genetic studies
Asian Pacific Poultry Congress. pp. . c.
Nestor KE. Hereditary chondrodystrophy in the turkey. Poultry
Science, : . .
Nichols CR and Cheng KM. Fawn: a plumage color mutation in
Japanese quail. Poultry Science, : . .
Niwa T, Mochii M, Nakamura A and Shiojiri N. Plumage
pigmentation and expression of its regulatory genes during
quail development - histochemical analysis using (black
at hatch) mutants. Mechanisms of Development, :
. .
Niwa T, Shibusawa M, Matsuda Y, Terashima A, Nakamura A
and Shiojiri N. The (black at hatch) gene that causes
abnormal feather pigmentation maps to chromosome of
the Japanese quail. Pigment Cell Research, : .
.
Nunoya T, Tajima M and Mizutani M. A new mutant of
Japanese quail ( ) characterized
by generalized glycogenosis. Laboratry Animals, :
. .
Oda K. Fifteen Years’ Experiment for Japanese Quail Breeding
(in Japanese). Oda-Chorui-Kenkyusho. Tokyo. .
Ohara O, Gahara Y, Miyake T, Teraoka H and Kitamura T.
Neurofilament deficiency in quail caused by nonsense muta-
tion in neurofilament-L gene. The Journal of Cell Biology,
: . .
Ohmori S. Ten Years’ Japanese Quail Breeding. Tobundo-Shoten
(in Japanese). Tokyo. .
Bh
Bh
Coturnix coturnix japonica
176
0
./ -
+.2 +.3 +332
,., ,.- +332
/1 /11 /2* +312
01 +,1 +322
++2 +-3+.0 ,**,
++0 0/0 00+
,**-
+1 +-2+., +32-
+3+1
+,+ -21 -3/ +33-
+3+2

��
�
�
� �
� �
� �
��
��
��
�
�
�
�
�
Ono T and Wakasugi N. Abnormalities in liver morphogenesis and chromosomes . Cytogenetic and Genome Research,
attributed to the (Black at hatch) lethal gene in the : . .
Japanese quail. Japanese Poultry Science, : . Schmid M, Nanda I, Hoehn H, Schartl M, Haaf T, Buerstedde
a. JM, Arakawa H, Caldwell RB, Weigend S, Burt DW, Smith
Ono T and Wakasugi N. Observation on development of J, Gri n DK, Masabanda JS, Groenen MAM, Crooijmans
black at hatch ( ) lethal quail embryos with special refer- RPMA, Vignal A, Fillon V, Morisson M, Pitel F, Vignoles
ence to liver morphogenesis. Japanese Poultry Science, : M, Garrigues A, Gellin J, Rodionov AV, Galkina SA,
. b. Lukina NA, Ben-Ari G, Blum S, Hillel J, Twito T, Lavi U,
Padgett CA and Ivey WD. Coturnix quail as a laboratory re- David L, Feldman MW, Delany ME, Conley CA, Fowler
search animal. Science, : . . VM, Hedges SB, Godbout R, Katyal S, Smith C, Hudson Q,
Pang SW, Ritland C, Carlson JE and Cheng KM. Japanese quail Sinclair A and Mizuno S. Second report on chicken genes
microsatellite loci amplified with chicken-specific primers. and chromosomes . Cytogenetic and Genome Research,
Animal Genetics, : . . : . .
Petek M, Ozen Y and Karakas E. E ects of recessive white Schreiweis MA, Hester PY, Settar P and Moody DE. Identific-
plumage colour mutation on hatchability and growth of ation of quantitative trait loci associated with egg quality,
quail hatched from breeders of di erent ages. British Poul- egg production, and body weight in an F resource popula-
try Science, : . . tion of chickens. Animal Genetics, : . .
Poole HK. Egg shell pigmentation in Japanese quail: genetic Schrodl F, de Stefano ME, Minvielle F, Brehmer A and
control of the white egg trait. The Journal of Heredity, : Neuhuber L. Somatostatin immunoreactivity in quail
. . pterygopalatine ganglion. Journal of Anatomy, :
Rao Y, Shen X, Xia M, Luo C, Nie Q, Zhang D and Zhang X. . .
SNP mapping of QTL a ecting growth and fatness on Sheykholeslami K, Kaga K and Mizutani M. Auditory nerve
chicken GGA . Genetics Selection Evolution, : . fiber di erences in the normal and neurofilament deficient
. Japanese quail. Hearing Research, : . .
Roberts CW, Fulton JE and Barnes CR. Genetics of white- Shibata T and Abe T. Linkage between the loci for serum
breasted, white and brown colors and descriptions of feather albumin and vitamin D binding protein (GC) in the Japa-
patterns in Japanese quail. Canadian Journal of Genetics nese quail. Animal Genetics, : . .
and Cytology, : . . Shibusawa M, Minai S, Nishida-Umehara C, Suzuki T, Mano T,
Roberts CW and Fulton JE. Rough-textured: a feather structure Yamada K, Namikawa T and Matsuda Y. A comparative
mutant of Japanese quail. Canadian Journal of Genetics and cytogenetic study of chromosome homology between chick-
Cytology, : . . en and Japanese quail. Cytogenetics and Cell Genetics, :
Roussot O, Feve K, Plisson-Petit F, Pitel F, Faure JM, . .
Beaumont C and Vignal A. AFLP linkage map of the Shimakura K. Notes on the genetics of the Japanese quail. I. The
Japanese quail Genetics Selection Evolu- simpl
tion, : . .
Sasazaki S, Hinenoya T, Fujima D, Kikuchi S, Fujiwara A and
Mannen H. Mapping of expressed sequence tag markers
with a cDNA-amplified fragment length polymorphism
method in Japanese quail ( ). Animal Sci-
ence Journal, : . a.
Sasazaki S, Hinenoya T, Lin B, Fujiwara A and Mannen H. A
comparative map of macrochromosomes between chicken
and Japanese quail based on orthologous genes. Animal
Genetics, : . b.
Satoh H, Nakamura A and Shiojiri N. (black at hatch) gene
appears to be expressed in melanocytes of feather germs in
the Japanese quail ( ). Develop-
ment Growth and Di erentiation, : . .
Savage TF and Collins WM. Downless, a mutant in Japanese
quail. Poultry Science, : . a.
Savage TF and Collins WM. Wry neck, an abnormality in
Japanese quail. Poultry Science, : . b.
Savage TF and Collins WM. Inheritance of star-gazing in Japa-
nese quail. The Journal of Heredity, : . .
Schmid M, Nanda I, Guttenbach M, Steinlein C, Hoehn H,
Schartl M, Haaf T, Weigend S, Fries R, Buerstedde JM,
Wimmers K, Burt DW, Smith J, A’Hara S, Law A, Gri n
DK, Bumstead N, Kaufman J, Thomson PA, Burke T,
Groenen MAM, Crooijmans RPMA, Vignal A, Fillon V,
Morisson M, Pitel F, Tixier-Boichard M, Ladjali-
Mohammedi K, Hillel J, Maki-Tanila A, Cheng HH, Delany
ME, Burnside J and Mizuno S. First report on chicken genes
Bh
in vitro
Bh
Coturnix japonica.
Coturnix japonica
Bh
Coturnix coturnix japonica
,
,***3* +03 ,+2 ,***
,* +/2 +03 +32-
$
,*-1* -2* +32-
-* ,01 ,02 +3/3
,**/-* +3/ +33 +333 +*3 .+/ .13 ,**/
#
#./ 103 11. ,**. -1 +*0 ++, ,**0
//+-0 +-2 +30. ,*0 ,.3
,// ,**/#
+ -3 /03 /2, #,**1 +/3 ++1 +,. ,**+
,1 +3/ +31 +330,* + 2 +312
,+ ..- ..2 +313 3/+*- +*3 ,**+
-/ //3 /1, ,**-
11 ., .0 ,**0
-1 -+0 -,* ,**0
# -3 ../ ./* +331
/* +0,1 +31+
/* +0,1 +31+
0- 21 3* +31,
$
�
�
�
�
�
�
�
�
Tsudzuki: Mutations and Modern Genetics in Quail
e mendelian autosomal, recessive character, “brown-
splashed white”, of its plumage. The Japanese Journal of
Genetics, : . .
Shiojiri N, Niwa T, Wakamatsu K, Ito S and Nakamura A.
Chemical analysis of melanin pigments in feather germs of
Japanese quail (black at hatch) mutants. Pigment Cell
Research, : . .
Sittmann K and Abplanalp H. White-feathered Japanese quail.
The Journal of Heredity, : . .
Sittmann K, Richards WPC and Abplanalp H. Congenital loco in
a third species of domestic fowl. Canadian Journal of Genet-
ics and Cytology, : . .
Sittmann K, Wilson WO and McFarland LZ. Bu and albino
Japanese quail: description, inheritance, and fitness traits.
The Journal of Heredity, : . .
Sittmann K and Craig RM. Crooked neck dwarfs in Japanese
quail. The Journal of Heredity, : . .
Sittmann K, Abplanalp H and Abbott UK. Extended storage of
quail, chicken and turkey eggs. . Embryonic abnormalities
and the inheritance of twinning in quail. Poultry Science, :
. .
Somes RG Jr. Genetic bases for plumage color patterns in four
varieties of Japanese quail. The Journal of Heredity, :
. .
Somes RG Jr. International Registry of Poultry Genetic Stocks.
Storrs Agricultural Experiment Station. The University of
Connecticut. Srorrs. .
Somes RG Jr. International Registry of Poultry Genetic Stocks.
Storrs Agricultural Experiment Station. The University of
Bh
177
+0 +*0 +++ +3.*
+, ,/3 ,0/ +333
/0 ,,* ,,- +30/
1 0-0 0.* +30/#
/1 ++3 +,. +300
/2 .. .0 +301
,/*
1+. 1,, +31+
1* ,*/,+* +313
+32.

�
�
��
� �
��
�
�
��
��
��
� �
�
�
�
�
�
�
�
in Japanese quail. Experimental Animals, : . a.
Connecticut. Storrs. . associated with the throat tuft mutation ( ) in the Japa-
Soseidoh-Syujin. The Book of Japanese Quail (in Japanese). nese quail. Poultry Science, : . b.
Soseidoh. Edo. . Tsudzuki M and Wakasugi N. External and internal ab-
Takahashi A, Mizutani M and Itakura C. Acrylamide-induced normalities exhibited by the crossbreds between the ear-tuft
neurotoxicity in the central nervous system of Japanese ( ) and throat-tuft ( ) mutants of Japanese quail
quails: comparative studies of normal and neurofilament- ( ). Japanese Poultry Science, :
deficient quails. Journal of Neuropathology and Experimen- . .
tal Neurology, : . . Tsudzuki M, Wakasugi N, Morioka H and Esaki K. Stumpy
Takahashi A, Mizutani M and Itakura C. Acrylamide-induced limb - an embryonic lethal mutation in Japanese quail
peripheral neuropathy in normal and neurofilament-deficient ( ). Experimental Animals, :
Japanese quails. Acta Neuropathologica, : . . . .
Taka-Tsukasa N. The Birds of Nippon. Maruzen,Tokyo. . Tsudzuki M, Kusano S, Wakasugi N, Morioka H and Esaki K.
Tanaka S, Braga IS rd, Kimura T, Ochiai K, Itakura C and Dotted white - a plumage color mutant in Japanese quail
Mizutani M. Inherited muscular disorder in mutant Japa- ( ). Experimental Animals, :
nese quail ( ): an ultrastructural . .
study. Journal of Comparative Pathology, : . Tsudzuki M, Nakane Y, Wakasugi N and Mizutani M. Allelism
a. of panda and dotted white plumage genes in Japanese quail.
Tanaka S, Braga IS rd, Kimura T, Ochiai K, Itakura C and The Journal of Heredity, : . .
Mizutani M. Inherited muscular disorder in mutant Japa- Tsudzuki M. New findings in the stumpy-limb mutation of
nese quail ( ): an immunohis- Japanese quail. The Journal of Heredity, : . a.
tochemical study. Journal of Comparative Pathology, : Tsudzuki M. Light down lethal: a new autosomal recessive down
. b. color mutation in Japanese quail. The Journal of Heredity,
Toyoshima I, Kato K, Sugawara M, Wada C, Okawa S, : . b.
Kobayashi M, Masamune O and Watanabe S. Massive Tsudzuki M, Ito S, Sato K, Takahashi S and Uchida H. Fawn- :
accumulation of M and H subunits of neurofilament pro- a dominant plumage color mutation in Japanese quail. The
teins in spinal motor neurons of neurofilament deficient Journal of Heredity, : . .
Japanese quail, Quv. Neuroscience Letters, : . Tsudzuki M, Nakane Y and Wada A. Dilute down lethal: a new
. lethal mutation in Japanese quail. Experimental Animals,
Truax RE and Johnson WA. Genetics of plumage color mutants : . .
in Japanese quail. Poultry Science, : . . Tsudzuki M, Nakane Y and Wada A. Hereditary multiple mal-
Truax RE, Siegel PB and Johnson WA. Redhead, a plumage formation in Japanese quail: a possible powerful animal
color mutant in Japanese quail. The Journal of Heredity, : model for morphogenetic studies. The Journal of Heredity,
. . : . a.
Tsudzuki M and Wakasugi N. “Pansy”: a plumage color mutant Tsudzuki M, Nakane Y and Wada A. Short beak: a n
in Japanese quail. Japanese Poultry Science, : .
.
Tsudzuki M and Wakasugi N. Back-drawer: a behavioral mutant
Tsudzuki M and Wakasugi N. A genetic defect in hyomandibular
furrow closure in the Japanese quail: the causes for ear-
opening abnormality and formation of an ear tuft. The
Journal of Heredity, : . b.
Tsudzuki M and Wakasugi N. Genetic head-skeleton ab-
normalities in the ET (ear tuft) strain of Japanese quail: a
defect associated with the incomplete closure of the hy-
omandibular furrow. Japanese Poultry Science, :
. c.
Tsudzuki M and Wakasugi N. Repair of the hyomandibular
furrow closure defect in the ET strain of Japanese quail.
Japanese Poultry Science, : . d.
Tsudzuki M and Wakasugi N. Throat tuft: a newly discovered
morphological mutation in Japanese quail. The Journal of
Heredity, : . .
Tsudzuki M, Fuji J and Wakasugi N. An allelism test for
plumage color mutant genes, (black) and (pansy), in
Japanese quail ( ). Japanese Poul-
try Science, : . .
Tsudzuki M and Wakasugi N. A genetic analysis on the throat-
tuft character of Japanese quail ( )
based on the head skeleton abnormality. Japanese Poultry
Science, : . a.
Tsudzuki M and Wakasugi N. Head skeletal abnormalities
Tt
Tt
hfd
hfd hfd
Coturnix coturnix japonica
Coturnix coturnix japonica
Coturnix coturnix japonica
Coturnix coturnix japonica
Coturnix coturnix japonica
D ps
Coturnix coturnix japonica
Coturnix coturnix japonica
-1 +-1 +.. +322
+32203 +,-0 +,.- +33*
+0.3
,2+-+ +.+ +33+
/- ,10 ,2- +33.
.*23 +1 ,, +33/ /,3 /-0 +33+
+301-
.+,/ -+ +33,
++. -,/ --1+330
- 2. ,,/ ,,3 +33-
20 00 02 +33/++/
+-3 +/* +33020 -*/ -*1 +33/
,
21 ,.2 ,/, +330-* +1/ +12
,***.0 3- 3. +331
/2 + 3 +313
1*.+- .+/ +313 23 ,. -+ +332
,. -,1 --/+321
13 +0* +0. +322
,/ ,*1,+1 +322
,/ ,12 ,21 +322
2* .-- ..+ +323
,1 ,00 ,03 +33*
,1 -3- -31 +33*
�
�
�
�
�
J. Poult. Sci., ( )
ew auto-
somal recessive semilethal mutation in Japanese quail. The
Journal of Heredity, : . b.
Tsudzuki M, Nakane Y, Wada A, and Ito S. A unique mode of
inheritance detected in a feather structure mutation of Jap-
anese quail. In: Proceedings of the th Asian Pacific Poultry
Congress. pp. . c.
Tsudzuki M, Onitsuka S, Akiyama R, Iwamizu M, Goto N,
Nishibori M, Takahashi H and Ishikawa A. Identification of
quantitative trait loci a ecting shank length, body weight
and carcass weight from the Japanese cockfighting chicken
breed, Oh-Shamo (Japanese Large Game). Cytogenetic and
Genome Research, : . .
Tsujino S, Kikuchi T, Tashiro T, Kinoshita N, Ikeda K, Ichihara
N, Kikuchi H, Hagiwara Y, Mizutani M and Sakuragawa
N. Adenovirus-mediated gene therapy for acid maltase de-
ficiency in Japanese quail. Jounal of Japanese Society for
Inherited Metabolic Diseases, : . .
Tsujino S, Kinoshita N, Tashiro T, Ikeda K, Ichihara T and
Sakuragawa N. Adenovirus-mediated transfer of human
acid maltase gene reduces glycogen accumulation in skeletal
muscle of Japanese quail with acid maltase deficiency.
Human Gene Therapy, : . .
Tsukamoto Y, Taira E, Nakane Y, Tsudzuki M, Kohama K,
Amin H, Miki N and Sasaki F. Expression of gicerin, a cell
adhesion molecule, in the abnormal retina in silver plumage
color mutation of Japanese quail ( ).
Neuroscience Letters, : . .
Ueda S, Ito H, Masai H and Kawahara T. Abnormal organiza-
Coturnix japonica
178 ./ -
23 +1/ +12 +332
0,.. ,./ +332
#
++1 ,22 ,3/ ,**1
+- .+3 +331
3 +0*3 +0+0 +332
,00 /- /0 +333

��
� �
�
�
�
�
� �
��
�
��
Tsudzuki: Mutations and Modern Genetics in Quail
tion of the cerebellar cortex in the mutant Japanese quail, Yamasaki H, Itakura C and Mizutani M. Hereditary hypotrophic
Brain Research, : . axonopathy with neurofilament deficiency in a mutant strain
. of the Japanese quail. Acta Neuropathologica, : .
Vallejo RL, Bacon LD, Liu HC, Witter RL, Groenen MAM, .
Hillel J and Cheng HH. Genetic mapping of quantitative Yamasaki H, Bennett GS, Itakura C and Mizutani M. Defective
trait loci a ecting susceptibility to Marek’s disease virus expression of neurofilament protein subunits in hereditary
induced tumors in F intercross chickens. Genetics, : hypotrophic axonopathy of quail. Laboratory Investigation,
. . : . .
van Kaam JBCHM, van Arendonk JAM, Groenen MAM, Yamashina Y. Quail breeding in Japan. The Journal of the
Bovenhuis H, Vereijken ALJ, Crooijmans RPMA, van der Bombay Natural History Society, : . .
Poel JJ and Veenendaal A. Whole genome scan for quantita- Yang HW, Kikuchi T, Hagiwara Y, Mizutani M, Chen YT and
tive trait loci a ecting body weight in chickens using a three Van Hove LK. Recombinant human acid alpha-glucosidase
generation design. Livestock Production Science, : corrects acid alpha-glucosidase-deficient human fibroblasts,
. . quail fibroblasts, and quail myoblasts. Pediatric Research,
Wakasugi N and Kondo K. Breeding methods for maintenance : . .
of mutant genes and establishment of strains in the Japanese Zhao JX, Ohnishi H and Murai Y. Smaller number of large
quail. Experimental Animals, (Suppl.): . . myelinated fibers and focal myelin thickening in mutant
Wakasugi N. Japanese quail. In: Evolution of Domesticated quails deficient in neurofilaments. Acta Neuropathologica,
Animals (Mason LL ed.). pp. . Longman. London. : . .
. Zhao JX, Ohnishi A, Itakura C, Mizutani M, Yamamoto T,
Wilson WO, Abbott UK and Abplanalp H. Evaluation of Hayashi H and Murai Y. Greater number of microtubules
coturnix (Japanese quail) as pilot animal for poultry. Poul- per axon of unmyelinated fibers of mutant quails deficient in
try Science, : . . neurofilaments: possible compensation for the absence of
Wright D, Kerje S, Lundstrom K, Babol J, Schutz K, Jensen P neurofilaments. Acta Neuropathologica, : . .
and Andersson L. Quantitative trait loci analysis of egg and Zhao JX, Ohnishi A, Itakura C, Mizutani M, Yamamoto T, Hojo
meat production traits in a red junglefowl x White Leghorn T and Murai Y. Smaller axon and unaltered numbers of
cross. Animal Genetics, : . . microtubules per axon in relation to number of myelin
Yakovlev AA, Kudryavtsev IV and Pigareva MD. A mutant lamellae of myelinated fibers in the mutant quail deficient in
Japanese quail ( ) with marble neurofilaments. Acta Neuropathologica, : . .
plumage. Soviet Genetics, : . .
Coturnix coturnix japonica.
Coturnix coturnix japonica
179
,
+11 +2- +22+313 2, .,1 .-.
+33+
#+.2
-.3 -0* +332 00 1-. 1.- +33,
/2 ,+0 ,,, +30+
#/. +--
+/* +332.- -1. -2* +332
,, +/+ +/3 +31-
-+3 -,+ 20 ,., ,.2 +33-+32.
.* 0/+ 0/1 +30+21 --, --0 +33.
-1 /,3 /-. ,**0
23 -*/ -+, +33/++ +,,/ +,-, +31/




















