
Retention of hunter-gatherer economies among maritime foragers from Caleta Vitor, northern Chile,...
13
Retention of hunteregatherer economies among maritime foragers from Caleta Vitor, northern Chile, during the late Holocene: evidence from stable carbon and nitrogen isotopic analysis of skeletal remains Amy Roberts a, * , F. Donald Pate a , Bianca Petruzzelli a , Chris Carter b , Michael C. Westaway c, h , Calogero M. Santoro d, e , Jaime Swift b , Todd Maddern f , Geraldine E. Jacobsen g , Fiona Bertuch g , Francisco Rothhammer d, e a Department of Archaeology, Flinders University, Sturt Rd., Bedford Park, Adelaide, SA 5001, Australia b School of Archaeology and Anthropology, Australian National University, Canberra, ACT 0200, Australia c Cultures and Histories, Queensland Museum, Brisbane, QLD 4101, Australia d Instituto de Alta Investigacion, Universidad de Tarapacá, Arica, Chile e Centro de Investigaciones del Hombre en el Desierto, Arica, Chile f Land and Water, Commonwealth Scientific and Industrial Research Organisation, Adelaide, SA 5064, Australia g Institute for Environmental Research, ANSTO, Locked Bag 2001, Kirrawee DC, NSW 2232, Australia h Griffith University, Nathan, Qld 4111, Australia article info Article history: Received 19 June 2012 Received in revised form 23 December 2012 Accepted 10 January 2013 Keywords: Isotope analysis Chile Human remains Caleta Vitor Atacama Marine economy Diet abstract On the basis of stable carbon and nitrogen isotope analysis of human remains, this paper provides evidence for the retention of hunteregatherer economies among coastal inhabitants in northern Chile during the late Holocene e at the same time that inland populations were adopting agricultural econ- omies. Coastal diets from the Caleta Vitor region of the Atacama Desert were dominated by marine-based foods, predominantly from upper trophic levels. The focus on reliable marine food resources is interpreted as a risk minimisation strategy in this marginal arid environment. Although these coastal hunteregatherers adopted other goods and traditions from agricultural populations, their participation in this larger interregional exchange network did not affect their basic subsistence economies. Associated new radiocarbon dates from the site are also discussed. Skeletal remains and artefacts associated with the Caleta Vitor shell middens generally originate from sites with dates ranging from c. 9000e476 BP. However, the human remains that are the subject of this research cover the time period c. 4000e476 BP. Ó 2013 Elsevier Ltd. All rights reserved. 1. Research aims The aims of this research were to employ stable isotopic data to test the hypothesis that marine economies were retained by coastal hunteregatherers in northern Chile during the late Holocene when other inland populations were adopting agricultural economies with diets dominated by cultivated plants. To test this hypothesis human remains were sampled from four archaeological sites in the Caleta Vitor area in the Atacama Region and the results were compared with previous data derived from coastal and inland Chilean sites. Associated dating was required to provide temporal control. Radiocarbon dating involved both direct dates on human remains as well as associated archaeological remains. 2. Regional background and relevant prior research 2.1. The Atacama Region The Atacama Region of northern Chile and southern Peru is one of the driest areas on earth e much of it receives less than 1 mm of rainfall per annum and is generally devoid of vegetation. Despite such harsh conditions humans settled the general area over 11,000 years ago (Núñez et al., 2002; Sandweiss et al., 1998; Santoro et al., 2005a). Indeed, there is growing evidence that “ancient people settled by the coast early and rapidly” (Arriaza et al., 2008). Early coastal sites in the region include: Quebradas Tacahuay, Jaguay, the Ring site, in southern Peru and Cascabeles on the coast of Taltal (Arriaza et al., 2008; Castelleti, 2007; Ugalde et al., 2012). Paleo-environmental reconstructions indicate that more water resources than today were available along the coast as * Corresponding author. Tel./fax: þ61 8 8201 2217. E-mail address: amy.roberts@flinders.edu.au (A. Roberts). Contents lists available at SciVerse ScienceDirect Journal of Archaeological Science journal homepage: http://www.elsevier.com/locate/jas 0305-4403/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.jas.2013.01.009 Journal of Archaeological Science 40 (2013) 2360e2372
Transcript of Retention of hunter-gatherer economies among maritime foragers from Caleta Vitor, northern Chile,...

at SciVerse ScienceDirect
Journal of Archaeological Science 40 (2013) 2360e2372
Contents lists available
Journal of Archaeological Science
journal homepage: http: / /www.elsevier .com/locate/ jas
Retention of hunteregatherer economies among maritime foragers from CaletaVitor, northern Chile, during the late Holocene: evidence from stable carbon andnitrogen isotopic analysis of skeletal remains
Amy Roberts a,*, F. Donald Pate a, Bianca Petruzzelli a, Chris Carter b, Michael C. Westaway c,h,Calogero M. Santoro d,e, Jaime Swift b, Todd Maddern f, Geraldine E. Jacobsen g, Fiona Bertuch g,Francisco Rothhammer d,e
aDepartment of Archaeology, Flinders University, Sturt Rd., Bedford Park, Adelaide, SA 5001, Australiab School of Archaeology and Anthropology, Australian National University, Canberra, ACT 0200, AustraliacCultures and Histories, Queensland Museum, Brisbane, QLD 4101, Australiad Instituto de Alta Investigacion, Universidad de Tarapacá, Arica, ChileeCentro de Investigaciones del Hombre en el Desierto, Arica, Chilef Land and Water, Commonwealth Scientific and Industrial Research Organisation, Adelaide, SA 5064, Australiag Institute for Environmental Research, ANSTO, Locked Bag 2001, Kirrawee DC, NSW 2232, AustraliahGriffith University, Nathan, Qld 4111, Australia
a r t i c l e i n f o
Article history:Received 19 June 2012Received in revised form23 December 2012Accepted 10 January 2013
Keywords:Isotope analysisChileHuman remainsCaleta VitorAtacamaMarine economyDiet
* Corresponding author. Tel./fax: þ61 8 8201 2217.E-mail address: [email protected] (A. R
0305-4403/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.jas.2013.01.009
a b s t r a c t
On the basis of stable carbon and nitrogen isotope analysis of human remains, this paper providesevidence for the retention of hunteregatherer economies among coastal inhabitants in northern Chileduring the late Holocene e at the same time that inland populations were adopting agricultural econ-omies. Coastal diets from the Caleta Vitor region of the Atacama Desert were dominated by marine-basedfoods, predominantly from upper trophic levels. The focus on reliable marine food resources isinterpreted as a risk minimisation strategy in this marginal arid environment. Although these coastalhunteregatherers adopted other goods and traditions from agricultural populations, their participationin this larger interregional exchange network did not affect their basic subsistence economies.
Associated new radiocarbon dates from the site are also discussed. Skeletal remains and artefactsassociated with the Caleta Vitor shell middens generally originate from sites with dates ranging from c.9000e476 BP. However, the human remains that are the subject of this research cover the time period c.4000e476 BP.
� 2013 Elsevier Ltd. All rights reserved.
1. Research aims
The aims of this research were to employ stable isotopic data totest the hypothesis that marine economies were retained by coastalhunteregatherers in northern Chile during the late Holocene whenother inland populations were adopting agricultural economieswith diets dominated by cultivated plants.
To test this hypothesis human remains were sampled from fourarchaeological sites in theCaletaVitorarea in theAtacamaRegionandthe results were compared with previous data derived from coastaland inland Chilean sites. Associated dating was required to providetemporal control. Radiocarbon dating involved both direct dateson human remains as well as associated archaeological remains.
oberts).
All rights reserved.
2. Regional background and relevant prior research
2.1. The Atacama Region
The Atacama Region of northern Chile and southern Peru is oneof the driest areas on earth emuch of it receives less than 1 mm ofrainfall per annum and is generally devoid of vegetation. Despitesuch harsh conditions humans settled the general area over 11,000years ago (Núñez et al., 2002; Sandweiss et al., 1998; Santoro et al.,2005a). Indeed, there is growing evidence that “ancient peoplesettled by the coast early and rapidly” (Arriaza et al., 2008). Earlycoastal sites in the region include: Quebradas Tacahuay, Jaguay, theRing site, in southern Peru and Cascabeles on the coast of Taltal(Arriaza et al., 2008; Castelleti, 2007; Ugalde et al., 2012).
Paleo-environmental reconstructions indicate that morewater resources than today were available along the coast as
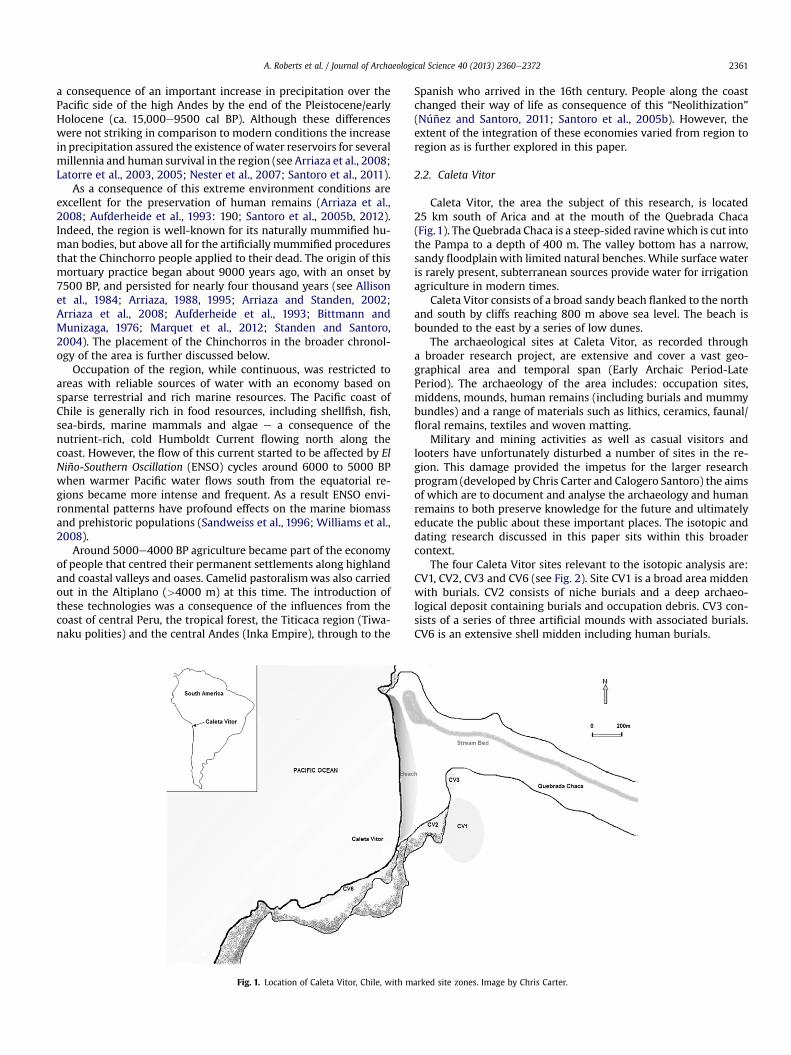
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e2372 2361
a consequence of an important increase in precipitation over thePacific side of the high Andes by the end of the Pleistocene/earlyHolocene (ca. 15,000e9500 cal BP). Although these differenceswere not striking in comparison to modern conditions the increasein precipitation assured the existence of water reservoirs for severalmillennia and human survival in the region (see Arriaza et al., 2008;Latorre et al., 2003, 2005; Nester et al., 2007; Santoro et al., 2011).
As a consequence of this extreme environment conditions areexcellent for the preservation of human remains (Arriaza et al.,2008; Aufderheide et al., 1993: 190; Santoro et al., 2005b, 2012).Indeed, the region is well-known for its naturally mummified hu-man bodies, but above all for the artificially mummified proceduresthat the Chinchorro people applied to their dead. The origin of thismortuary practice began about 9000 years ago, with an onset by7500 BP, and persisted for nearly four thousand years (see Allisonet al., 1984; Arriaza, 1988, 1995; Arriaza and Standen, 2002;Arriaza et al., 2008; Aufderheide et al., 1993; Bittmann andMunizaga, 1976; Marquet et al., 2012; Standen and Santoro,2004). The placement of the Chinchorros in the broader chronol-ogy of the area is further discussed below.
Occupation of the region, while continuous, was restricted toareas with reliable sources of water with an economy based onsparse terrestrial and rich marine resources. The Pacific coast ofChile is generally rich in food resources, including shellfish, fish,sea-birds, marine mammals and algae e a consequence of thenutrient-rich, cold Humboldt Current flowing north along thecoast. However, the flow of this current started to be affected by ElNiño-Southern Oscillation (ENSO) cycles around 6000 to 5000 BPwhen warmer Pacific water flows south from the equatorial re-gions became more intense and frequent. As a result ENSO envi-ronmental patterns have profound effects on the marine biomassand prehistoric populations (Sandweiss et al., 1996; Williams et al.,2008).
Around 5000e4000 BP agriculture became part of the economyof people that centred their permanent settlements along highlandand coastal valleys and oases. Camelid pastoralismwas also carriedout in the Altiplano (>4000 m) at this time. The introduction ofthese technologies was a consequence of the influences from thecoast of central Peru, the tropical forest, the Titicaca region (Tiwa-naku polities) and the central Andes (Inka Empire), through to the
Fig. 1. Location of Caleta Vitor, Chile, with m
Spanish who arrived in the 16th century. People along the coastchanged their way of life as consequence of this “Neolithization”(Núñez and Santoro, 2011; Santoro et al., 2005b). However, theextent of the integration of these economies varied from region toregion as is further explored in this paper.
2.2. Caleta Vitor
Caleta Vitor, the area the subject of this research, is located25 km south of Arica and at the mouth of the Quebrada Chaca(Fig.1). The Quebrada Chaca is a steep-sided ravinewhich is cut intothe Pampa to a depth of 400 m. The valley bottom has a narrow,sandy floodplainwith limited natural benches. While surface wateris rarely present, subterranean sources provide water for irrigationagriculture in modern times.
Caleta Vitor consists of a broad sandy beach flanked to the northand south by cliffs reaching 800 m above sea level. The beach isbounded to the east by a series of low dunes.
The archaeological sites at Caleta Vitor, as recorded througha broader research project, are extensive and cover a vast geo-graphical area and temporal span (Early Archaic Period-LatePeriod). The archaeology of the area includes: occupation sites,middens, mounds, human remains (including burials and mummybundles) and a range of materials such as lithics, ceramics, faunal/floral remains, textiles and woven matting.
Military and mining activities as well as casual visitors andlooters have unfortunately disturbed a number of sites in the re-gion. This damage provided the impetus for the larger researchprogram (developed by Chris Carter and Calogero Santoro) the aimsof which are to document and analyse the archaeology and humanremains to both preserve knowledge for the future and ultimatelyeducate the public about these important places. The isotopic anddating research discussed in this paper sits within this broadercontext.
The four Caleta Vitor sites relevant to the isotopic analysis are:CV1, CV2, CV3 and CV6 (see Fig. 2). Site CV1 is a broad area middenwith burials. CV2 consists of niche burials and a deep archaeo-logical deposit containing burials and occupation debris. CV3 con-sists of a series of three artificial mounds with associated burials.CV6 is an extensive shell midden including human burials.
arked site zones. Image by Chris Carter.

Fig. 2. View south across Caleta Vitor from the northern flank of the valley with marked site zones. Image by Chris Carter.
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e23722362
2.3. Chronological resolution of the archaeological landscape anddocumented dietary changes over time
The chronological resolution of the archaeological landscape atCaleta Vitor is also part of the broader research project occurring inthe area. In addition to the new and previously unreported datesdiscussed in this paper, further 14C and OSL dating is planned. Ingeneral terms, however, the chronology of cultural change innorthern Chile can be divided into several different periods (seeTable 1 for a summary).
Some documented dietary changes and relevant material cul-ture items are also noted in a general sense in this section to pro-vide context to later discussions. In particular, as also demonstratedin this isotope study, it should be noted that coastal sites such asthose at Caleta Vitor, in northern Chile’s Atacama region, provideevidence for the primary retention of hunteregatherer economiesand diets dominated by marine-based foods in comparison to theadoption of agricultural economies in other areas.
2.3.1. The Early Archaic Period (c. 10,000e8000 BP)The Early Archaic Period witnesses the arrival of the first
hunteregatherers in northern Chile as recognised by small campsof hunters and gatherers in the high Andes (Osorio et al., 2011),
Table 1Summary of chronological periods relevant to the Caleta Vitor area outlined in thispaper.
Time period (BP) Period Phases
c. 10,000e8000 Early Archaic Acha, Early Chinchorroc. 8000e6000 Middle Archaic Chinchorroc. 6000e4000 Late Archaic Late Chinchorroc. 4000e1500 Formative Final phase of Chinchorro
culture (4000e3600)Azapa (4000e2500)Alto Ramírez (2500e1500)
c. 1500e900 Middle Horizon (Tiwanakupolities expansion)
CabuzaMaitas Chiribaya
c. 1000e660 Late Intermediatec. 660e476 Late (Inka Empire)
while the earliest evidence for coastal hunteregathererefishers hasbeen dated to around 13,000 cal BP (DeFrance and Umire, 2004;Sandweiss et al., 1998). These people primarily survived on mol-luscs, sea-birds, sea mammals and fish (Santoro et al., 2005a). Sitesfrom this period have yielded obsidian artefacts, basalt choppers,bone tools for collecting shellfish, stemmed triangular projectilepoints and thick lanceolate points (Llagostera, 1979; Rivera, 1991;Santoro et al., 2005b; Standen et al., 2004). These coastal groupsmade long-distance movements through the valleys and up to theAltiplano looking for raw materials and other resources such asobsidian, game and plants, thus their movements were organisedon a seasonal basis (Núñez and Santoro, 2011; Rivera, 1991). Groupsfrom the highlands journeyed to the lowlands for the samepurposes.
2.3.2. The Middle Archaic Period (c. 8000e6000 BP)The Middle Archaic Period saw the inception of the Chinchorro
Culture (c. 8000e4000 BP). These groups were specialised hunteregatherers adapted to coastal living and reliant on marine resourcesas their primary means of subsistence (Alfonso et al., 2007;Rivadeneira et al., 2010). Arriaza et al. (2008:47) state that the“presence of large shell middens and cemeteries suggests thatharvesting was plentiful at many locations like Quiani, PlayaChinchorro-Las Machas, Caleta Vitor, Quebrada de Camarones, andPisagua”. Their primary food was mollusks such as Concholepasconcholepas and Fissurella spp. (limpets), fish and pelicans (Arriazaet al., 2008: 47). In addition, there is evidence that they huntedwildcamelids and marine mammals such as sea lions (Arriaza et al.,2008: 47).
Whilst Chinchorro populations may have been initially nomadic“there is growing bioarchaeological evidence that indicates theyhad a year-round occupation on the coast rather than seasonaloccupation” (Arriaza et al., 2008: 47 e see also Marquet et al., 2012;Núñez and Santoro, 2011; Santoro et al., 2012). Further, Arriaza et al.(2001) argue thatmobility can also be considered to be at oddswiththeir intensive mortuary practices. A distinct maritime technologyis also evident incorporating shell fishhooks, lines, weights andharpoons (Rivera, 1991; Santoro et al., 2005b).

A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e2372 2363
2.3.3. The Late Archaic Period (c. 6000e4000 BP)During the Late Archaic Period the Chinchorro began to occupy
dwelling siteswith simple architecture from approximately 5500 to3000 BP. Woven reed basketry and some cotton textiles are alsoaccredited to this period (Kelley et al., 1991; Santoro et al., 2005b;Standen et al., 2004). More Chinchorro sites are found along thecoast during this phase, which has been interpreted as indicative ofan increase in population and social complexity (Rivera, 1991;Marquet et al., 2012; Williams et al., 2008). It would appear thatsome inland groups during this period had adopted some domes-ticated plants, such as maize, sweet potatoes and manioc as well assome animals, including llamas and alpacas (Rivera, 1991; Ugaldeet al., 2012). Intensification in the extraction of marine resourcesis signalled by a variety of small hunting tools including triangular,lanceolate and rhomboidal points, as well as cactus thorn fishhooksand bone fishhooks (Grosjean et al., 1997; Llagostera, 1979; Rivera,1991; Rothhammer et al., 2009; Santoro et al., 2005b).
2.3.4. The Formative Period (c. 4000e1500 BP)During the Early Formative Period (c. 4000e2500 BP) the people
of northern Chile had become fully sedentary and during this finalphase of the Chinchorro Culture the incipient use of cultigens canbe observed. Achira, beans, gourds, squash, maize, sweetpotatoes, manioc, quinoa and possibly pecay were all beingcultivated in the Azapa Valley (Rivera, 1991; Ugalde et al., 2012).This period is also characterised by the introduction of ceramics,around 3000 BP, and woollen textiles (Alfonso et al., 2007; Rivera,1991).
The Late Formative Period (c. 2500e1500 BP) shows somemigration from the highlands to the coastal valleys and thebeginnings of an economy based on surplus goods (Kelley et al.,1991; Moraga et al., 2005; Rothhammer and Cocilovo, 2008). Inthe lowland valleys, these agriculturalists had irrigated land, theyields of which were complemented by highland products such asmeat, wool and jerky from camelids (Rivera, 1991). It is during thisperiod that the influences of pre-Tiwanaku ritual-political centresreached the western valleys such as Quebrada Chaca.
2.3.5. The Middle Horizon Period (c. 1500e1000 BP)In the Middle Horizon Period the Tiwanaku polities spread
throughout much of the central Andes and represent the first po-litical integration of the area. Sedentism developed towards ur-banisation, extensive irrigated agriculture, and population increaseallowed competition and cooperation between Tiwanaku politiesand other societal groups not subject to their system in the Alti-plano (Janusek, 2008; Rivera, 1991). In the valleys of northern Chilea number of populations were fully agricultural with cropsincluding maize, sweet potatoes, squash and beans grown by irri-gation along the valleys (Rivera, 1991). In general terms villagesduring the period produced fine textiles and pottery and there wasa differentiation of social and political roles (Rivera, 1991).
2.3.6. The Late Intermediate Period (c. 1000e660 BP)Throughout the Late Intermediate Period we see the appear-
ance of some defensive, walled sites, particularly in the highlands,which along with rock art and some funerary contexts from theperiod implies new forms of intergroup tension and interaction(Rivera, 1991). In general terms village populations were estimatedto be around 400e500 people and in many areas they continued tofarm from irrigated terraces and fields (Rivera, 1991). It is thoughtthat these people also had a sophisticated economic system andllama caravans crossed the desert for trade (Kelley et al., 1991).Larger settlements in the region show evidence of urban planningwith streets and sectors devoted to different activities (Rivera,1991).
2.3.7. The Late Period (c. 660e476 BP)The Late Period saw the consolidation of the Inka Empire.
Maintenance of roads and transportation are related to Inkaadministration and the region is managed by several mechanismsof interaction including verticality among other means of economicexchange between people from the coast, valleys, precordillera andAltiplano (Santoro et al., 2010). Some populations were controlledby force and mining became important to the empire with newtechniques introduced such as foundries and amalgamation pro-cedures (Rivera, 1991; Santoro et al., 2010). While the Inkan Em-pire’s administrative structure collapsed quite quickly as a result ofthe Spanish Invasion, it is likely that the economic structure atCaleta Vitor continued largely undisturbed for many decades(Santoro et al., 2010).
2.4. Prior stable carbon and nitrogen isotope research
Stable isotopic analyses of bones and teeth recovered fromarchaeological sites are now accepted methods used to independ-ently assess past diet and the related use of the landscape (see Gilet al., 2010; Katzenberg, 2000; Larsen, 1997; Pate, 1994, 1997, 2008;Turner et al., 2010). Indeed, stable carbon and nitrogen isotopeanalysis is increasingly being employed as an essential tool in thereconstruction of past human diets. Bone chemistry provides anindependent line of evidence to discussions based on archae-ofaunal and palaeobotanical data (see Ambrose, 1993; Van derMerwe, 1982). Unfortunately isotopic research in southern SouthAmerica is not well-represented in the international archaeologicalliterature (Barberena et al., 2009:128). Thus, this research aims tocontribute to the research in this area.
Variations in the stable carbon and nitrogen composition ofplants and animals allow inferences about the relative use of ma-rine versus terrestrial foods, plants employing C3 versus C4 pho-tosynthetic mechanisms and trophic level from analyses of faunaland human skeletal remains. Isotopic differences at the base of foodchains are passed up to higher level consumers.
Most elements exist as mixtures of two or more isotopes. Iso-topes are variants of the same element with differing numbers ofneutrons in their nuclei. They possess almost identical chemicalproperties, but have different atomic masses. These differences inatomic mass result in variable chemical reaction rates (Hoefs, 1987;Peterson and Fry, 1987; Urey, 1947). Kinetic variability allows plantsand animals to preferentially incorporate one isotope over othersduring biochemical reactions associated with photosynthesis andfood metabolism. Isotope ratios are expressed as delta (d) values inparts per thousand (&) relative to the isotopic composition ofstandards. The standards used are Pee Dee Belemnite (PDB) car-bonate for carbon isotopes and atmospheric nitrogen (AIR) for ni-trogen isotopes.
Stable carbon (d13C) and nitrogen (d15N) isotope values in bonecollagen are quantitatively related to the isotopic composition ofingested foods (DeNiro and Epstein, 1978, 1981; Katzenberg, 2000;Tieszen and Boutton, 1988). Due to slow turnover rates, the stableisotopic composition of normal bone proteins provide a record ofdietary intake over a period of decades and in many cases offera lifetime average (Libby et al., 1964; Sealy et al., 1995; Stenhouseand Baxter, 1979). Consequently, bone collagen stable isotopevalues will be related to long-term patterns of food consumptionand habitat use.
In regions of theworld where C3 plants (trees, shrubs, temperatecool season grasses/herbs) dominate the terrestrial landscape, bonecollagen d13C values can provide information regarding the relativeimportance of marine and terrestrial foods in the diets of prehis-toric populations who occupied coastal and adjacent inland habi-tats (Schoeninger et al., 1983; Schoeninger and DeNiro, 1984).

A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e23722364
However, because C4 plants (tropical and warm season grasses/herbs) have d13C values similar to marine foods, marine and ter-restrial dietary composition cannot be distinguished on the basis ofbone collagen stable carbon isotope values in habitats with largequantities of C4 plants. In contrast, in interior habitats with limitedaccess to marine foods, bone collagen d13C values can be employedto assess the relative proportions of C3 and C4 plants in prehistoricdiets.
Bone collagen stable nitrogen isotope values may also provideinformation regarding relative consumption of marine and terres-trial foods. Thus, in habitats with large quantities of C4 plants, bonecollagen d15N values can sometimes be used to address marineversus terrestrial dietary composition. However, because elevatedd15N values in arid-land fauna can result in an overlap betweenstable nitrogen isotope distributions for marine and terrestrialanimals (Ambrose, 1991; Ambrose and DeNiro, 1986; Heaton et al.,1986; Pate and Anson, 2008; Sealy et al., 1987), in arid-land eco-systems it is essential that isotopic analyses of local fauna beemployed to establish marine and terrestrial dietary distinctionsprior to applications involving prehistoric humans.
Prior stable carbon and nitrogen isotope research that has takenplace in the region includes studies conducted by Aufderheide(1993), Aufderheide et al. (1993, 1994), Aufderheide and Santoro(1999), Knudson et al. (2007) and Slovak and Paytan (2009).Tykot et al. (2009) also provide a useful overview of stable isotoperesearch primarily in relation to central Chile and include isotopicvalues for a number of sites and temporal periods that suppliesuseful broader context.
Aufderheide et al. (1993) conducted stable carbon and nitrogenisotope and other analyses on hair, bone and soft tissues from sevenindividuals from the early to late Chinchorro Culture (dates rangedfrom 8970 � 255 BP to 4635 � 90 BP) from four sites in northernChile (Acha-2 [5 km east of Arica], Camarones-8 [80 km south ofArica], Camarones-17 [85 km south of Arica] and Chinchorro-1 [onthe northern edge of Arica]). Whilst only obtaining stable carbonand nitrogen results for a few individuals they did note that thed15N values were very high and indicative of a diet derived frommarine resources (Aufderheide et al., 1993: 198).
Aufderheide et al. (1994) conducted stable carbon and nitrogenisotope analysis on samples (hair and muscle) from Pisagua,a northern Chilean site approximately 130 km south of Arica. Thesamples were taken from spontaneously mummified Alto Ramirezbodies from a beach site (c. 2950 BP). They argue that although itwas previously believed that this cultural group “transferred theirhighland practices of agriculture and pastoralism” to lower valleysites that their analysis instead revealed that the group’s subsis-tence strategy was “indistinguishable from that of the coastalmaritime populations” (Aufderheide et al., 1994: 515). Further,Aufderheide et al. (1994: 521) found that:
.d15N values from hair specimens range from 21.9 to 27.0&.These very positive values are supported by d34S valuesfrom þ15.6 to 19.0&. These results clearly indicate thatthe.13C-enriched values were not the result of substantialingestion of a C4 cultigen such asmaize, but instead reflect a dietheavily dominated by foods of marine origin. At Pisagua thesewould include fish, shellfish, sea lions, seaweed, marine-feedingbirds and similar items.Aufderheide et al. (1994: 522) also notethat these results contrast with a later Alto Ramirez group fromc. 1600 BP that settled in the lower Azapa Valley (approximately15 km inland from Arica). This latter group appears to haveretained aspects of their agropastoral subsistence practices sothat marine foods constituted less than half of their dietarycomponents (see Aufderheide and Allison, 1992 e these resultswere based on trace analysis).
Aufderheide and Santoro (1999) employed stable carbon andnitrogen isotope analysis of bone collagen to investigate the humanpaleodiet of the Molle Pampa Este and Molle Pampa Medio popu-lation sub-groups. Their samples were collected from the LlutaValley, an area adjacent to the city of Arica (approximately 22.5 kminland from the valley mouth). The Molle Pampa Este populationwas mostly a Late Period group and the Molle Pampa Medio pop-ulation was exclusively from the Late Intermediate Period(Aufderheide and Santoro, 1999: 240). They concluded that “.theprincipal components in the diet of Molle Pampa populations wereterrestrial plants and marine meat resources with only a minorcontribution from terrestrial meat sources” (Aufderheide andSantoro, 1999: 237). Furthermore they noted that “the quiteaccessible marine resources were exploited by both studied groupsto a greater extent than the archaeological remains suggested”(Aufderheide and Santoro, 1999: 249). Indeed, they argue thatcarbon and nitrogen isotope ratio comparisons demonstrate thatapproximately 80% of the meat component of the diet was of ma-rine origin for the populations under study (Aufderheide andSantoro, 1999: 248). Their summary values for these populationsare plotted in comparison to the populations analysed in thisresearch later in this paper.
In contrast, Knudson et al.’s (2007) stable carbon and nitrogenisotope analyses on human hair from two Chiribaya-affiliated sites(southern Peru e approximately 150 km north of Arica) demon-strated that there was seasonal variability in relation to the con-sumption of marine products. They also argued that theconsumption of C4 plants was quite high for some individuals(Knudson et al., 2007).
Similarly, Slovak and Paytan’s (2009) study on individuals fromthe Andean Middle Horizon Period buried at the coastal site ofAncón, Peru (approximately 1000 km north of Arica), also indicateda mixed diet composed mainly of marine protein and C4 resourceswith reliance on maize increasing over time. Their results werebased on carbon and nitrogen isotope analyses on human bone andtooth enamel (Slovak and Paytan, 2009).
Tykot et al. (2009: 159) also provide useful comparative data forstable carbon and nitrogen isotope analyses (on human bones andteeth) (see also Falabella et al., 2007). They provide results forapproximately 100 human individuals from archaeological contextsspanning from c. 7000 to 400 BP in central Chile. Summary valuesfor some of these populations (i.e., where there are similar samplesizes from bone collagen analyses to compare) are plotted incomparison to the populations analysed in this research later in thispaper. In particular, we compare their averaged results from humancollagen for the following archaeological sites:
� Valle Verde e an inland site approximately 2000 km south ofArica (chronology ¼ c. 2150e1750 BP);
� Maria Pinto e an inland site approximately 2000 km south ofArica (chronology ¼ c. 950e550 BP);
� Laguna El Peral-Ce a coastal site approximately 2000 km southof Arica (chronology ¼ c. 1600e950 BP); and
� Las Brisas 10e14 a coastal site approximately 2000 km south ofArica (chronology ¼ c. 950e500 BP).
Their results suggest that the individuals from these sitesdemonstrate a low reliance on marine foods for most inhabitantsliving on the coast as well as “indirect evidence of mobility strat-egies that show differences between Early and Late Intermediatesocieties; increased dependence on maize with time, especiallyamong the Aconcagua people” (Falabella et al., 2007: 5). Theirresearch also revealed differences between the sexes in relation tomaize intake with men in Aconcagua groups showing higher levelsof consumption (Falabella et al., 2007: 5). Further, they note that

Table 2Summary of d13C and d15N values (bone collagen and plant tissues) of grave goodsfrom the Pisagua site, northern Chile (summarised and adapted from Aufderheideet al., 1994: 520).
Organism d13C (&) d15N (&)
Eel jay �14.4 16.1Sea lion bone �13.1 22.1Fish vertebrae �12.6 No dataFish vertebrae �11.3 20.3Fish vertebrae �12.9 20.3Seaweed �12.5 No dataMaize cob �12.6 No dataMaize cob �10.0 No dataMaize cob �11.4 No dataDistichlis �11.7 No dataSeed �14.6 No dataSeed �22.3 No dataPlant stem �10.3 No data
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e2372 2365
their studies revealed results that appear to reinforce some hy-potheses (e.g., the different dietary patterns between con-temporary Bato and Lolleo groups) but contradict other results (e.g.,the presumed reliance on a marine diet for coastal peoples)(Falabella et al., 2007: 5).
3. Materials and methods
3.1. Stable isotopic analysis
Bone specimens, ranging from approximately 1.0e3.0 g, weretaken for each individual depending on the amount of skeletalmaterial available for analysis. Following sample exclusions (seesections below) the sample size ultimately consisted of 10individuals.
Sample preparation involved cleaning of the bone pieces,demineralisation and sodium hydroxide treatment. Bone fragmentswere demineralised in dilute HCl according to the methods of Sealy(1986). Humic acids and other base-soluble contaminants wereremoved using a 0.125 M NaOH solution. Extracts were soaked andwashed thoroughly following acid and base treatments in order toremove dissolved contaminants. The remaining organic componentwas oven dried at approximately 40 �C.
Carbon and nitrogen concentrations were determined using anANCA SL elemental analyser coupled to a Geo 20-20 IRMS. Stablecarbon and nitrogen isotope values were determined by massspectrometry. Analytical precision was better than � 0.2& for d13Cand �0.3& for d15N. d values were placed on the VPDB scale usinga two-point calibration method, with the standards USGS40 andUSGS41 used as anchor points (Coplen et al., 2006; Qi et al., 2003). Anumber of samples were repeated during the mass spectrometryprocess to ensure accuracy. Where repeat testing was conductedthe values were averaged for use in the analysis and discussion forthis research.
Controls for post-mortem organic decomposition were imple-mented by excluding samples with: 1) Less than 5% collagen yieldfrom the bone specimens; or 2) Less than 5% carbon yield fromcollagen; or 3) Less than 0.5% nitrogen yield from collagen(Ambrose, 1990; Pate, 1997; Schoeninger et al., 1989). Atomic C:Nratios in modern collagen were employed to determine the pres-ence of acceptable collagen in archaeological extracts in relation tostable isotope analysis. These acceptable atomic C:N ratios weredeveloped by extracting collagen from large samples of modernmammals and measuring their C:N ratios. Results include atomicC:N ranges of 2.9e3.6 for 172 mammals (DeNiro, 1985) and 2.8e3.5for 79 mammals (Ambrose, 1990). C:N ratios for all samples utilisedin the analysis and discussion for this research were withinthe range of that reported for modern collagen e i.e., between 2.8and 3.6.
There are various methods employed to establish the relativecontributions of marine and terrestrial food staples in a diet viastable carbon and nitrogen isotope analysis of bone (Tykot et al.,2009). In this paper we have followed the isotopic ‘endpoints’used by Aufderheide and Santoro (1999: 246e247).
Plants using the C4 (HatcheSlack) pathway have markedlyhigher d13C values than plants employing the C3 (CalvineBenson)pathway. Mean d13C values of �13.1 � 1.2& for C4 plants and�27.1 � 2.0& for C3 plants are reported for a large number ofspecies grown under a variety of conditions (O’Leary, 1988). Carbonisotope tissue ratios vary from �7 to �16& in C4 plants and from�20 to �35& in C3 plants (Deines, 1980; O’Leary, 1981; Ehleringer,1989). Faunal controls for the region suggest that plant foods aredominated by species employing the C3 photosynthetic pathway. Incontrast, maize employs a C4 photosynthetic pathway (see Tables 2and 3).
Past research in the region in the vicinity of theMolle PampaEsteand Molle Pampa Medio sites (Tieszen and Chapman, 1995) foundthe d13C value of food resources to have a mean of �23.5& for C3terrestrial plants, �14.1& for C4 terrestrial plants and �15.2& formarine fish (all corrected for modern industrial fossil fuel burningeffects). Higher trophic level marine food sources (marine fish, sealions, sea birds) all contribute d13C similar to that of C4 plants, so thatby itself the 13C value from collagen cannot distinguish between C4plants and upper trophic marine foods.
Because stable carbon isotope values of ingested foods areenhanced by þ5& during the processes of digestion and proteinsynthesis (fractionation), human bone collagen values related tothe consumption of 100% C3 plant diets would be �18.5&, whilea value of �10.2& would represent a 100% C4 plant diet. A 100%marine fish diet would result in mean bone collagen carbon isotopevalues of approximately 9.1&.
Bone collagen d15N values may also be employed to estimate themarine protein fraction in human diets. Tieszen and Chapman(1995) found that terrestrial plants in this area rarely reveal d15Nvalues greater thanþ8& (with amean value ofþ3.9� 2.3&), whileupper trophic level marine foods averaged 24.9& (equivalent to27.9& in the consumer after applying a þ3& fractionation factorfor each trophic level).
3.2. Radiocarbon dating of bone samples
As noted above, some of the dates reported in this paper comefrom the direct dating of the skeletal remains from Caleta Vitor.While direct dating of ancient bones through the extraction andpurification of gelatin derived from the bone collagen is an estab-lished method (Bronk Ramsey et al., 2004; Longin, 1971; Staffordet al., 1987) the success of the method is largely dependent onunderstanding the degree of preservation of the bone and ensuringthe removal of contaminants (DeNiro and Weiner, 1988; Ambrose,1990).
Degraded collagen is susceptible to contamination from organicmaterials, particularly humics (Stafford et al., 1987; Van Klinkenand Hedges, 1995), thus it is essential that both prior to and dur-ing processing, the bone and extracted collagen are tested to assessthe degree of preservation and suitability for dating (Higham et al.,2006). Severalmethods for testing the degradation of the bone and/or extracted gelatin have been reported and are further discussedbelow (DeNiro and Weiner, 1988; Van Klinken, 1999). In addition,various methods have been reported for removing contaminationfrom the extracted gelatin (e.g. Stafford et al., 1987; Van Klinkenand Mook, 1990; Tripp et al., 2006). Separation of the high molec-ular weight gelatin components from low molecular weight
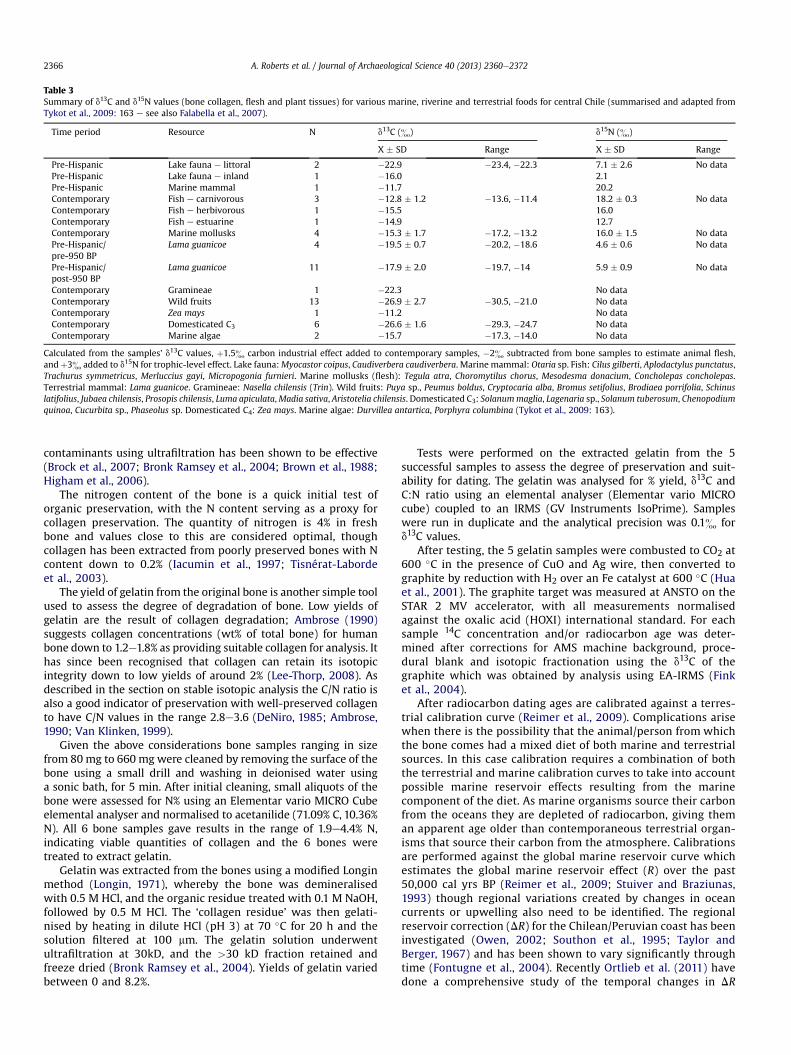
Table 3Summary of d13C and d15N values (bone collagen, flesh and plant tissues) for various marine, riverine and terrestrial foods for central Chile (summarised and adapted fromTykot et al., 2009: 163 e see also Falabella et al., 2007).
Time period Resource N d13C (&) d15N (&)
X � SD Range X � SD Range
Pre-Hispanic Lake fauna e littoral 2 �22.9 �23.4, �22.3 7.1 � 2.6 No dataPre-Hispanic Lake fauna e inland 1 �16.0 2.1Pre-Hispanic Marine mammal 1 �11.7 20.2Contemporary Fish e carnivorous 3 �12.8 � 1.2 �13.6, �11.4 18.2 � 0.3 No dataContemporary Fish e herbivorous 1 �15.5 16.0Contemporary Fish e estuarine 1 �14.9 12.7Contemporary Marine mollusks 4 �15.3 � 1.7 �17.2, �13.2 16.0 � 1.5 No dataPre-Hispanic/pre-950 BP
Lama guanicoe 4 �19.5 � 0.7 �20.2, �18.6 4.6 � 0.6 No data
Pre-Hispanic/post-950 BP
Lama guanicoe 11 �17.9 � 2.0 �19.7, �14 5.9 � 0.9 No data
Contemporary Gramineae 1 �22.3 No dataContemporary Wild fruits 13 �26.9 � 2.7 �30.5, �21.0 No dataContemporary Zea mays 1 �11.2 No dataContemporary Domesticated C3 6 �26.6 � 1.6 �29.3, �24.7 No dataContemporary Marine algae 2 �15.7 �17.3, �14.0 No data
Calculated from the samples’ d13C values, þ1.5& carbon industrial effect added to contemporary samples, �2& subtracted from bone samples to estimate animal flesh,andþ3& added to d15N for trophic-level effect. Lake fauna:Myocastor coipus, Caudiverbera caudiverbera. Marine mammal: Otaria sp. Fish: Cilus gilberti, Aplodactylus punctatus,Trachurus symmetricus, Merluccius gayi, Micropogonia furnieri. Marine mollusks (flesh): Tegula atra, Choromytilus chorus, Mesodesma donacium, Concholepas concholepas.Terrestrial mammal: Lama guanicoe. Gramineae: Nasella chilensis (Trin). Wild fruits: Puya sp., Peumus boldus, Cryptocaria alba, Bromus setifolius, Brodiaea porrifolia, Schinuslatifolius, Jubaea chilensis, Prosopis chilensis, Luma apiculata,Madia sativa, Aristotelia chilensis. Domesticated C3: Solanummaglia, Lagenaria sp., Solanum tuberosum, Chenopodiumquinoa, Cucurbita sp., Phaseolus sp. Domesticated C4: Zea mays. Marine algae: Durvillea antartica, Porphyra columbina (Tykot et al., 2009: 163).
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e23722366
contaminants using ultrafiltration has been shown to be effective(Brock et al., 2007; Bronk Ramsey et al., 2004; Brown et al., 1988;Higham et al., 2006).
The nitrogen content of the bone is a quick initial test oforganic preservation, with the N content serving as a proxy forcollagen preservation. The quantity of nitrogen is 4% in freshbone and values close to this are considered optimal, thoughcollagen has been extracted from poorly preserved bones with Ncontent down to 0.2% (Iacumin et al., 1997; Tisnérat-Labordeet al., 2003).
The yield of gelatin from the original bone is another simple toolused to assess the degree of degradation of bone. Low yields ofgelatin are the result of collagen degradation; Ambrose (1990)suggests collagen concentrations (wt% of total bone) for humanbone down to 1.2e1.8% as providing suitable collagen for analysis. Ithas since been recognised that collagen can retain its isotopicintegrity down to low yields of around 2% (Lee-Thorp, 2008). Asdescribed in the section on stable isotopic analysis the C/N ratio isalso a good indicator of preservation with well-preserved collagento have C/N values in the range 2.8e3.6 (DeNiro, 1985; Ambrose,1990; Van Klinken, 1999).
Given the above considerations bone samples ranging in sizefrom 80 mg to 660 mg were cleaned by removing the surface of thebone using a small drill and washing in deionised water usinga sonic bath, for 5 min. After initial cleaning, small aliquots of thebone were assessed for N% using an Elementar vario MICRO Cubeelemental analyser and normalised to acetanilide (71.09% C, 10.36%N). All 6 bone samples gave results in the range of 1.9e4.4% N,indicating viable quantities of collagen and the 6 bones weretreated to extract gelatin.
Gelatin was extracted from the bones using a modified Longinmethod (Longin, 1971), whereby the bone was demineralisedwith 0.5 M HCl, and the organic residue treated with 0.1 M NaOH,followed by 0.5 M HCl. The ‘collagen residue’ was then gelati-nised by heating in dilute HCl (pH 3) at 70 �C for 20 h and thesolution filtered at 100 mm. The gelatin solution underwentultrafiltration at 30kD, and the >30 kD fraction retained andfreeze dried (Bronk Ramsey et al., 2004). Yields of gelatin variedbetween 0 and 8.2%.
Tests were performed on the extracted gelatin from the 5successful samples to assess the degree of preservation and suit-ability for dating. The gelatin was analysed for % yield, d13C andC:N ratio using an elemental analyser (Elementar vario MICROcube) coupled to an IRMS (GV Instruments IsoPrime). Sampleswere run in duplicate and the analytical precision was 0.1& ford13C values.
After testing, the 5 gelatin samples were combusted to CO2 at600 �C in the presence of CuO and Ag wire, then converted tographite by reduction with H2 over an Fe catalyst at 600 �C (Huaet al., 2001). The graphite target was measured at ANSTO on theSTAR 2 MV accelerator, with all measurements normalisedagainst the oxalic acid (HOXI) international standard. For eachsample 14C concentration and/or radiocarbon age was deter-mined after corrections for AMS machine background, proce-dural blank and isotopic fractionation using the d13C of thegraphite which was obtained by analysis using EA-IRMS (Finket al., 2004).
After radiocarbon dating ages are calibrated against a terres-trial calibration curve (Reimer et al., 2009). Complications arisewhen there is the possibility that the animal/person from whichthe bone comes had a mixed diet of both marine and terrestrialsources. In this case calibration requires a combination of boththe terrestrial and marine calibration curves to take into accountpossible marine reservoir effects resulting from the marinecomponent of the diet. As marine organisms source their carbonfrom the oceans they are depleted of radiocarbon, giving theman apparent age older than contemporaneous terrestrial organ-isms that source their carbon from the atmosphere. Calibrationsare performed against the global marine reservoir curve whichestimates the global marine reservoir effect (R) over the past50,000 cal yrs BP (Reimer et al., 2009; Stuiver and Braziunas,1993) though regional variations created by changes in oceancurrents or upwelling also need to be identified. The regionalreservoir correction (DR) for the Chilean/Peruvian coast has beeninvestigated (Owen, 2002; Southon et al., 1995; Taylor andBerger, 1967) and has been shown to vary significantly throughtime (Fontugne et al., 2004). Recently Ortlieb et al. (2011) havedone a comprehensive study of the temporal changes in DR
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e2372 2367
along the north Chilean e south Peruvian coast and have pro-posed mean DR values for different time periods over theHolocene.
The percentage of marine contribution to the bone collagencan be estimated by determining ‘end-point’ values for d13C at100% terrestrial and 100% marine (Arneborg et al., 1999). For thepurpose of calibrating ages from bone collagen, we need onlyconsider the protein sources in the diet as it is recognised that theprimary source of carbon in bone collagen is from dietary protein(Lee-Thorp, 2008). Consumption of C4 plants such as maize mayconfound the estimate of marine dietary contribution as the iso-topic signatures from the two sources can be difficult to separatebased on d13C of the collagen alone (Aufderheide and Santoro,1999), though it is likely that in a diet with a mix of low andhigh protein foods, the contribution of the low protein food to thebone collagen is underrepresented (Ambrose et al., 1997; Tykotet al., 2009).
Given the issues outlined above the Caleta Vitor ages werecalibrated by estimating the fraction of marine diet for each bonesample using mean d13C values of bone collagen taken from theChilean population groups listed in Table 5. Using the samemethod of Arneborg et al. (1999) two endpoints were determinedrepresenting 100% terrestrial diet and 100% marine diet. The d13Cmean value of �20.1& from the Valle Verde population group wastaken as representing 100% terrestrial diet endpoint (Table 5,Fig. 4). The diet of this population group does not include maize(Tykot et al., 2009), thus this is a reliable estimate for a 100%terrestrial diet.
The Molle Pampa Este population group provides the meand13C values closest to that expected for a 100% marine diet, witha mean d13C of �11.0&. However, using a range of isotopictechniques, Aufderheide and Santoro (1999) estimated that theMolle Pampa Este and Molle Pampa Medio diets consisted of 42%vegetal (including some maize), 47% marine meat and 11% ter-restrial meat with the protein diet comprising around 80% ma-rine protein. d15N values of the bone collagen also indicate thatdietary protein is primarily from marine sources (Table 5)(Aufderheide and Santoro, 1999). While maize is known to havebeen part of their diet for this calibration it is assumed that thecontribution of the maize C4 protein to the total protein diet issmall. If the protein component of the maize is assumed to bebetween 5 and 10% (Uarrota et al., 2011) then even if maize
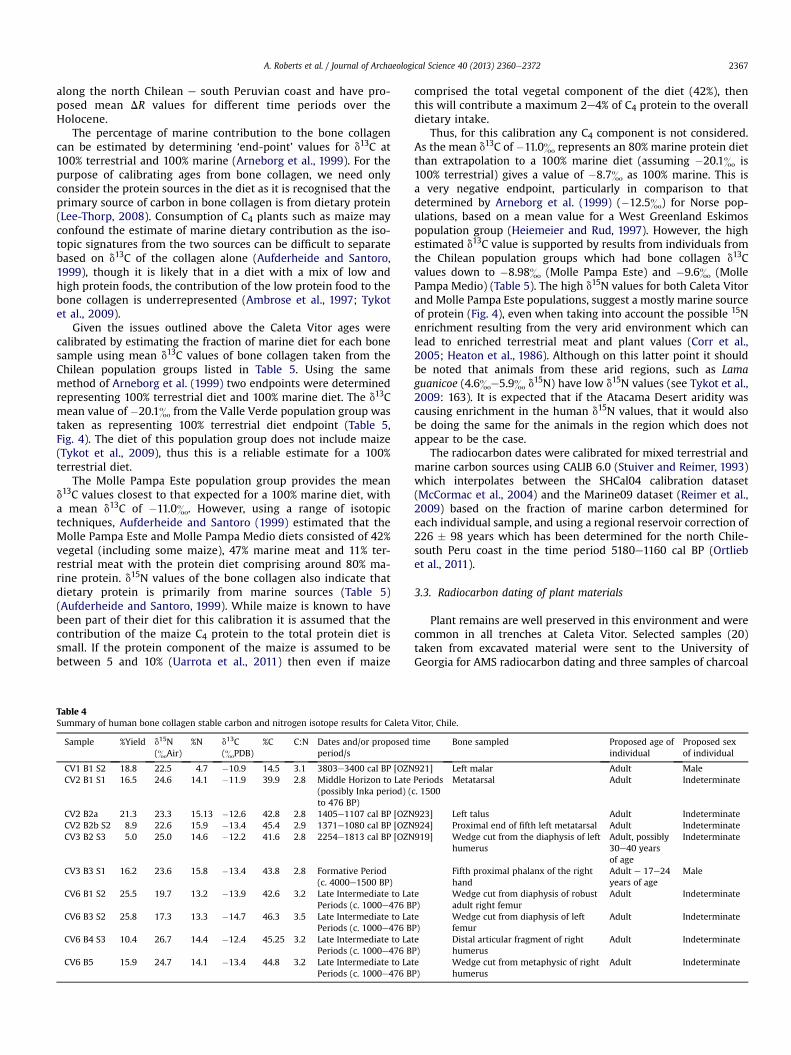
Table 4Summary of human bone collagen stable carbon and nitrogen isotope results for Caleta
Sample %Yield d15N(&Air)
%N d13C(&PDB)
%C C:N Dates and/or proposedperiod/s
CV1 B1 S2 18.8 22.5 4.7 �10.9 14.5 3.1 3803e3400 cal BP [OZNCV2 B1 S1 16.5 24.6 14.1 �11.9 39.9 2.8 Middle Horizon to Late
(possibly Inka period) (to 476 BP)
CV2 B2a 21.3 23.3 15.13 �12.6 42.8 2.8 1405e1107 cal BP [OZNCV2 B2b S2 8.9 22.6 15.9 �13.4 45.4 2.9 1371e1080 cal BP [OZNCV3 B2 S3 5.0 25.0 14.6 �12.2 41.6 2.8 2254e1813 cal BP [OZN
CV3 B3 S1 16.2 23.6 15.8 �13.4 43.8 2.8 Formative Period(c. 4000e1500 BP)
CV6 B1 S2 25.5 19.7 13.2 �13.9 42.6 3.2 Late Intermediate to LaPeriods (c. 1000e476 B
CV6 B3 S2 25.8 17.3 13.3 �14.7 46.3 3.5 Late Intermediate to LaPeriods (c. 1000e476 B
CV6 B4 S3 10.4 26.7 14.4 �12.4 45.25 3.2 Late Intermediate to LaPeriods (c. 1000e476 B
CV6 B5 15.9 24.7 14.1 �13.4 44.8 3.2 Late Intermediate to LaPeriods (c. 1000e476 B
comprised the total vegetal component of the diet (42%), thenthis will contribute a maximum 2e4% of C4 protein to the overalldietary intake.
Thus, for this calibration any C4 component is not considered.As the mean d13C of �11.0& represents an 80% marine protein dietthan extrapolation to a 100% marine diet (assuming �20.1& is100% terrestrial) gives a value of �8.7& as 100% marine. This isa very negative endpoint, particularly in comparison to thatdetermined by Arneborg et al. (1999) (�12.5&) for Norse pop-ulations, based on a mean value for a West Greenland Eskimospopulation group (Heiemeier and Rud, 1997). However, the highestimated d13C value is supported by results from individuals fromthe Chilean population groups which had bone collagen d13Cvalues down to �8.98& (Molle Pampa Este) and �9.6& (MollePampa Medio) (Table 5). The high d15N values for both Caleta Vitorand Molle Pampa Este populations, suggest a mostly marine sourceof protein (Fig. 4), even when taking into account the possible 15Nenrichment resulting from the very arid environment which canlead to enriched terrestrial meat and plant values (Corr et al.,2005; Heaton et al., 1986). Although on this latter point it shouldbe noted that animals from these arid regions, such as Lamaguanicoe (4.6&e5.9& d15N) have low d15N values (see Tykot et al.,2009: 163). It is expected that if the Atacama Desert aridity wascausing enrichment in the human d15N values, that it would alsobe doing the same for the animals in the region which does notappear to be the case.
The radiocarbon dates were calibrated for mixed terrestrial andmarine carbon sources using CALIB 6.0 (Stuiver and Reimer, 1993)which interpolates between the SHCal04 calibration dataset(McCormac et al., 2004) and the Marine09 dataset (Reimer et al.,2009) based on the fraction of marine carbon determined foreach individual sample, and using a regional reservoir correction of226 � 98 years which has been determined for the north Chile-south Peru coast in the time period 5180e1160 cal BP (Ortliebet al., 2011).
3.3. Radiocarbon dating of plant materials
Plant remains are well preserved in this environment and werecommon in all trenches at Caleta Vitor. Selected samples (20)taken from excavated material were sent to the University ofGeorgia for AMS radiocarbon dating and three samples of charcoal
Vitor, Chile.
time Bone sampled Proposed age ofindividual
Proposed sexof individual
921] Left malar Adult MalePeriodsc. 1500
Metatarsal Adult Indeterminate
923] Left talus Adult Indeterminate924] Proximal end of fifth left metatarsal Adult Indeterminate919] Wedge cut from the diaphysis of left
humerusAdult, possibly30e40 yearsof age
Indeterminate
Fifth proximal phalanx of the righthand
Adult e 17e24years of age
Male
teP)
Wedge cut from diaphysis of robustadult right femur
Adult Indeterminate
teP)
Wedge cut from diaphysis of leftfemur
Adult Indeterminate
teP)
Distal articular fragment of righthumerus
Adult Indeterminate
teP)
Wedge cut from metaphysic of righthumerus
Adult Indeterminate
Marine Hunters
Maize Farmers
European Farmers
0
5
10
15
20
25
30
-25 -20 -15 -10 -5
δ15N
(‰)
13C (‰)δ
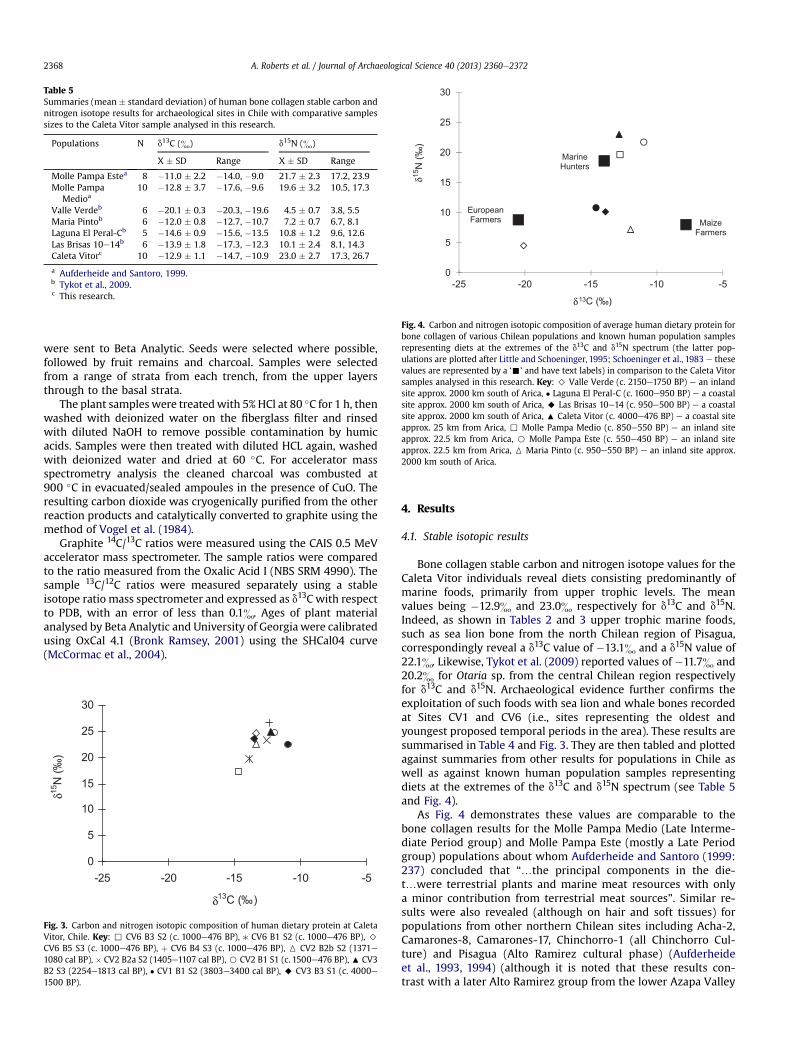
Fig. 4. Carbon and nitrogen isotopic composition of average human dietary protein forbone collagen of various Chilean populations and known human population samplesrepresenting diets at the extremes of the d13C and d15N spectrum (the latter pop-ulations are plotted after Little and Schoeninger, 1995; Schoeninger et al., 1983 e thesevalues are represented by a ‘-’ and have text labels) in comparison to the Caleta Vitorsamples analysed in this research. Key: > Valle Verde (c. 2150e1750 BP) e an inlandsite approx. 2000 km south of Arica, � Laguna El Peral-C (c. 1600e950 BP) e a coastalsite approx. 2000 km south of Arica, A Las Brisas 10e14 (c. 950e500 BP) e a coastalsite approx. 2000 km south of Arica, : Caleta Vitor (c. 4000e476 BP) e a coastal siteapprox. 25 km from Arica, , Molle Pampa Medio (c. 850e550 BP) e an inland siteapprox. 22.5 km from Arica, B Molle Pampa Este (c. 550e450 BP) e an inland siteapprox. 22.5 km from Arica, 6 Maria Pinto (c. 950e550 BP) e an inland site approx.2000 km south of Arica.
Table 5Summaries (mean � standard deviation) of human bone collagen stable carbon andnitrogen isotope results for archaeological sites in Chile with comparative samplessizes to the Caleta Vitor sample analysed in this research.
Populations N d13C (&) d15N (&)
X � SD Range X � SD Range
Molle Pampa Estea 8 �11.0 � 2.2 �14.0, �9.0 21.7 � 2.3 17.2, 23.9Molle Pampa
Medioa10 �12.8 � 3.7 �17.6, �9.6 19.6 � 3.2 10.5, 17.3
Valle Verdeb 6 �20.1 � 0.3 �20.3, �19.6 4.5 � 0.7 3.8, 5.5Maria Pintob 6 �12.0 � 0.8 �12.7, �10.7 7.2 � 0.7 6.7, 8.1Laguna El Peral-Cb 5 �14.6 � 0.9 �15.6, �13.5 10.8 � 1.2 9.6, 12.6Las Brisas 10e14b 6 �13.9 � 1.8 �17.3, �12.3 10.1 � 2.4 8.1, 14.3Caleta Vitorc 10 �12.9 � 1.1 �14.7, �10.9 23.0 � 2.7 17.3, 26.7
a Aufderheide and Santoro, 1999.b Tykot et al., 2009.c This research.
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e23722368
were sent to Beta Analytic. Seeds were selected where possible,followed by fruit remains and charcoal. Samples were selectedfrom a range of strata from each trench, from the upper layersthrough to the basal strata.
The plant samples were treatedwith 5% HCl at 80 �C for 1 h, thenwashed with deionized water on the fiberglass filter and rinsedwith diluted NaOH to remove possible contamination by humicacids. Samples were then treated with diluted HCL again, washedwith deionized water and dried at 60 �C. For accelerator massspectrometry analysis the cleaned charcoal was combusted at900 �C in evacuated/sealed ampoules in the presence of CuO. Theresulting carbon dioxide was cryogenically purified from the otherreaction products and catalytically converted to graphite using themethod of Vogel et al. (1984).
Graphite 14C/13C ratios were measured using the CAIS 0.5 MeVaccelerator mass spectrometer. The sample ratios were comparedto the ratio measured from the Oxalic Acid I (NBS SRM 4990). Thesample 13C/12C ratios were measured separately using a stableisotope ratio mass spectrometer and expressed as d13C with respectto PDB, with an error of less than 0.1&. Ages of plant materialanalysed by Beta Analytic and University of Georgia were calibratedusing OxCal 4.1 (Bronk Ramsey, 2001) using the SHCal04 curve(McCormac et al., 2004).
0
5
10
15
20
25
30
-25 -20 -15 -10 -5
δ
δ
13C (‰)
15N
(‰)
Fig. 3. Carbon and nitrogen isotopic composition of human dietary protein at CaletaVitor, Chile. Key: , CV6 B3 S2 (c. 1000e476 BP), CV6 B1 S2 (c. 1000e476 BP), >CV6 B5 S3 (c. 1000e476 BP), þ CV6 B4 S3 (c. 1000e476 BP), 6 CV2 B2b S2 (1371e1080 cal BP), � CV2 B2a S2 (1405e1107 cal BP), B CV2 B1 S1 (c. 1500e476 BP), : CV3B2 S3 (2254e1813 cal BP), � CV1 B1 S2 (3803e3400 cal BP), A CV3 B3 S1 (c. 4000e1500 BP).
4. Results
4.1. Stable isotopic results
Bone collagen stable carbon and nitrogen isotope values for theCaleta Vitor individuals reveal diets consisting predominantly ofmarine foods, primarily from upper trophic levels. The meanvalues being �12.9& and 23.0& respectively for d13C and d15N.Indeed, as shown in Tables 2 and 3 upper trophic marine foods,such as sea lion bone from the north Chilean region of Pisagua,correspondingly reveal a d13C value of �13.1& and a d15N value of22.1&. Likewise, Tykot et al. (2009) reported values of �11.7& and20.2& for Otaria sp. from the central Chilean region respectivelyfor d13C and d15N. Archaeological evidence further confirms theexploitation of such foods with sea lion and whale bones recordedat Sites CV1 and CV6 (i.e., sites representing the oldest andyoungest proposed temporal periods in the area). These results aresummarised in Table 4 and Fig. 3. They are then tabled and plottedagainst summaries from other results for populations in Chile aswell as against known human population samples representingdiets at the extremes of the d13C and d15N spectrum (see Table 5and Fig. 4).
As Fig. 4 demonstrates these values are comparable to thebone collagen results for the Molle Pampa Medio (Late Interme-diate Period group) and Molle Pampa Este (mostly a Late Periodgroup) populations about whom Aufderheide and Santoro (1999:237) concluded that “.the principal components in the die-t.were terrestrial plants and marine meat resources with onlya minor contribution from terrestrial meat sources”. Similar re-sults were also revealed (although on hair and soft tissues) forpopulations from other northern Chilean sites including Acha-2,Camarones-8, Camarones-17, Chinchorro-1 (all Chinchorro Cul-ture) and Pisagua (Alto Ramirez cultural phase) (Aufderheideet al., 1993, 1994) (although it is noted that these results con-trast with a later Alto Ramirez group from the lower Azapa Valley
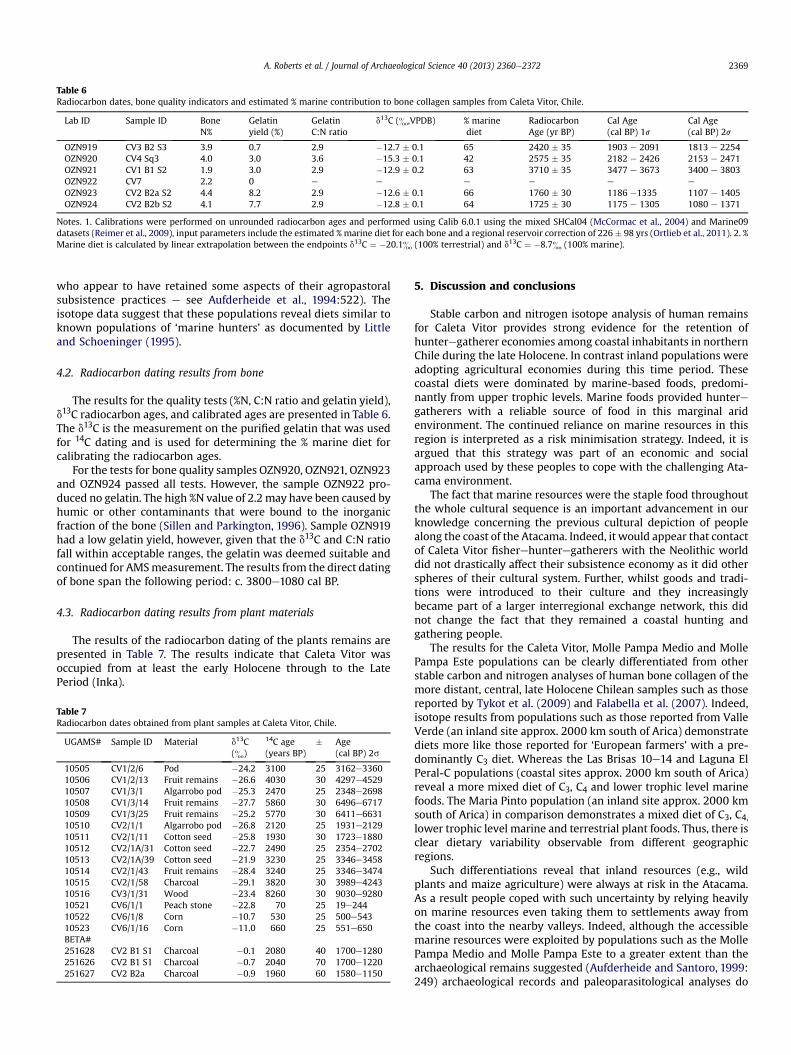
Table 6Radiocarbon dates, bone quality indicators and estimated % marine contribution to bone collagen samples from Caleta Vitor, Chile.
Lab ID Sample ID BoneN%
Gelatinyield (%)
GelatinC:N ratio
d13C (&,VPDB) % marinediet
RadiocarbonAge (yr BP)
Cal Age(cal BP) 1s
Cal Age(cal BP) 2s
OZN919 CV3 B2 S3 3.9 0.7 2.9 �12.7 � 0.1 65 2420 � 35 1903 e 2091 1813 e 2254OZN920 CV4 Sq3 4.0 3.0 3.6 �15.3 � 0.1 42 2575 � 35 2182 e 2426 2153 e 2471OZN921 CV1 B1 S2 1.9 3.0 2.9 �12.9 � 0.2 63 3710 � 35 3477 e 3673 3400 e 3803OZN922 CV7 2.2 0 e e e e e e
OZN923 CV2 B2a S2 4.4 8.2 2.9 �12.6 � 0.1 66 1760 � 30 1186 e1335 1107 e 1405OZN924 CV2 B2b S2 4.1 7.7 2.9 �12.8 � 0.1 64 1725 � 30 1175 e 1305 1080 e 1371
Notes. 1. Calibrations were performed on unrounded radiocarbon ages and performed using Calib 6.0.1 using the mixed SHCal04 (McCormac et al., 2004) and Marine09datasets (Reimer et al., 2009), input parameters include the estimated % marine diet for each bone and a regional reservoir correction of 226 � 98 yrs (Ortlieb et al., 2011). 2. %Marine diet is calculated by linear extrapolation between the endpoints d13C ¼ �20.1& (100% terrestrial) and d13C ¼ �8.7& (100% marine).
A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e2372 2369
who appear to have retained some aspects of their agropastoralsubsistence practices e see Aufderheide et al., 1994:522). Theisotope data suggest that these populations reveal diets similar toknown populations of ‘marine hunters’ as documented by Littleand Schoeninger (1995).
4.2. Radiocarbon dating results from bone
The results for the quality tests (%N, C:N ratio and gelatin yield),d13C radiocarbon ages, and calibrated ages are presented in Table 6.The d13C is the measurement on the purified gelatin that was usedfor 14C dating and is used for determining the % marine diet forcalibrating the radiocarbon ages.
For the tests for bone quality samples OZN920, OZN921, OZN923and OZN924 passed all tests. However, the sample OZN922 pro-duced no gelatin. The high %N value of 2.2 may have been caused byhumic or other contaminants that were bound to the inorganicfraction of the bone (Sillen and Parkington, 1996). Sample OZN919had a low gelatin yield, however, given that the d13C and C:N ratiofall within acceptable ranges, the gelatin was deemed suitable andcontinued for AMSmeasurement. The results from the direct datingof bone span the following period: c. 3800e1080 cal BP.
4.3. Radiocarbon dating results from plant materials
The results of the radiocarbon dating of the plants remains arepresented in Table 7. The results indicate that Caleta Vitor wasoccupied from at least the early Holocene through to the LatePeriod (Inka).
Table 7Radiocarbon dates obtained from plant samples at Caleta Vitor, Chile.
UGAMS# Sample ID Material d13C(&)
14C age(years BP)
� Age(cal BP) 2s
10505 CV1/2/6 Pod �24.2 3100 25 3162e336010506 CV1/2/13 Fruit remains �26.6 4030 30 4297e452910507 CV1/3/1 Algarrobo pod �25.3 2470 25 2348e269810508 CV1/3/14 Fruit remains �27.7 5860 30 6496e671710509 CV1/3/25 Fruit remains �25.2 5770 30 6411e663110510 CV2/1/1 Algarrobo pod �26.8 2120 25 1931e212910511 CV2/1/11 Cotton seed �25.8 1930 30 1723e188010512 CV2/1A/31 Cotton seed �22.7 2490 25 2354e270210513 CV2/1A/39 Cotton seed �21.9 3230 25 3346e345810514 CV2/1/43 Fruit remains �28.4 3240 25 3346e347410515 CV2/1/58 Charcoal �29.1 3820 30 3989e424310516 CV3/1/31 Wood �23.4 8260 30 9030e928010521 CV6/1/1 Peach stone �22.8 70 25 19e24410522 CV6/1/8 Corn �10.7 530 25 500e54310523 CV6/1/16 Corn �11.0 660 25 551e650BETA#251628 CV2 B1 S1 Charcoal �0.1 2080 40 1700e1280251626 CV2 B1 S1 Charcoal �0.7 2040 70 1700e1220251627 CV2 B2a Charcoal �0.9 1960 60 1580e1150
5. Discussion and conclusions
Stable carbon and nitrogen isotope analysis of human remainsfor Caleta Vitor provides strong evidence for the retention ofhunteregatherer economies among coastal inhabitants in northernChile during the late Holocene. In contrast inland populations wereadopting agricultural economies during this time period. Thesecoastal diets were dominated by marine-based foods, predomi-nantly from upper trophic levels. Marine foods provided hunteregatherers with a reliable source of food in this marginal aridenvironment. The continued reliance on marine resources in thisregion is interpreted as a risk minimisation strategy. Indeed, it isargued that this strategy was part of an economic and socialapproach used by these peoples to cope with the challenging Ata-cama environment.
The fact that marine resources were the staple food throughoutthe whole cultural sequence is an important advancement in ourknowledge concerning the previous cultural depiction of peoplealong the coast of the Atacama. Indeed, it would appear that contactof Caleta Vitor fisherehunteregatherers with the Neolithic worlddid not drastically affect their subsistence economy as it did otherspheres of their cultural system. Further, whilst goods and tradi-tions were introduced to their culture and they increasinglybecame part of a larger interregional exchange network, this didnot change the fact that they remained a coastal hunting andgathering people.
The results for the Caleta Vitor, Molle Pampa Medio and MollePampa Este populations can be clearly differentiated from otherstable carbon and nitrogen analyses of human bone collagen of themore distant, central, late Holocene Chilean samples such as thosereported by Tykot et al. (2009) and Falabella et al. (2007). Indeed,isotope results from populations such as those reported from ValleVerde (an inland site approx. 2000 km south of Arica) demonstratediets more like those reported for ‘European farmers’ with a pre-dominantly C3 diet. Whereas the Las Brisas 10e14 and Laguna ElPeral-C populations (coastal sites approx. 2000 km south of Arica)reveal a more mixed diet of C3, C4 and lower trophic level marinefoods. The Maria Pinto population (an inland site approx. 2000 kmsouth of Arica) in comparison demonstrates a mixed diet of C3, C4,lower trophic level marine and terrestrial plant foods. Thus, there isclear dietary variability observable from different geographicregions.
Such differentiations reveal that inland resources (e.g., wildplants and maize agriculture) were always at risk in the Atacama.As a result people coped with such uncertainty by relying heavilyon marine resources even taking them to settlements away fromthe coast into the nearby valleys. Indeed, although the accessiblemarine resources were exploited by populations such as the MollePampa Medio and Molle Pampa Este to a greater extent than thearchaeological remains suggested (Aufderheide and Santoro, 1999:249) archaeological records and paleoparasitological analyses do

A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e23722370
show that the pattern of moving marine foods to the interior isa practice that started early in the Archaic Period and continued tothe Late Period in the Atacama region (see Araujo de et al., 1985;Dorsey Vinton et al., 2009; Núñez and Hall, 1982; Santoro et al.,2003).
In contrast, the inland territories of central Chile, between thecoast and the Andes receive local rain and are traversed by per-manent rivers. Such environments offer a much better base tosubstantiate a terrestrial hunting and gathering economy withminor input from marine resources. Thus, for people in centralChile there was no need to transport marine foods to the interior asoccurred in the Atacama. In fact, even coastal groups in the lateHolocene such as the Las Brisas 10e14 and Laguna El Peral-Cpopulations (c. 950e500 and c. 1600e950 BP respectively) didnot have a full marine economy.
Thus, the difference in the importance of marine resources be-tween the Atacama and central Chilean populations is an importantgeographic distinction with broad implications. Simply becausepeople lived in a coastal environment at a particular time perioddoes not guarantee a marine diet. Rather it appears that peopleliving in such regions carefully considered the advantages and risksin relation to the integration of agriculture into their economies.
The stable isotope data relating to a marine based economy isfurther supported by the archaeological evidence at Caleta Vitor.The sites from which the human remains were obtained aredominated by middens including the remains of marine shell, sealions, marine fish bones, sea bird bones and whale bones. In addi-tion, one individual from CV1 exhibited bilateral auditory exostoseswhich is a pathological condition that reflects exposure to coldwater (including diving) over a long period of time. This providesadditional evidence of dependence on a marine economy.
Although the remains of corn cobs were present in thearchaeological deposits at some sites, e.g. CV6, the stable carbonisotope values demonstrate that corn was not a significant com-ponent in the diets at Caleta Vitor.
Associated new radiocarbon dates from the site have also beenpresented in this paper. On the basis of these results it can be statedthat skeletal remains and artefacts associated with the Caleta Vitorshell middens generally originate from sites with dates rangingfrom c. 9000e476 BP. However, the human remains that are thesubject of this research cover the time period c. 4000e476 BP.
With a sample size of 10 individuals we cannot address dietaryvariability through time. However, future research employinga larger sample would allow the investigation of any possible dif-ferences in diet over time and/or between classes of people fromthe same temporal period.
Acknowledgements
Funding for this isotope analysis was provided via the Faculty ofEducation, Humanities, Law and Theology, Flinders University.Thank you to Dr Rachel Popelka-Filcoff and other technical labo-ratory staff from the School of Chemical and Physical Sciences atFlinders University for facilitating access to laboratory space andequipment. Thanks are also due to technical laboratory staff at theCommonwealth Scientific and Industrial Research Organisation fortheir assistance in loading the samples for mass spectrometryanalysis. Calogero Santoro and Francisco Rothhammer are fundedby FONDECYT 1095006 and the Centro de Investigaciones delHombre en el Desierto (CIHDE), CONICYT-REGIONAL R07C1001.Calogero Santoro is also funded by a National Geographic ResearchGrant (#8690-09) lead by Dr Marcelo Rivadeneira. He also ac-knowledges the fellowship La Chaire des Amériques at L’universitéRennes 2 under which his final editions to this paper took place.Calogero and Chris Carter should also be acknowledged for
obtaining relevant permits and for dealing with the other ethicalconsiderations required. Thanks also to Linda Barry for doing thed13C, %N, C:N ratio analyses and Vladimir Levchenko for the AMSanalyses of the skeletal remains. Radiocarbon analyses of theskeletal remains were funded through an Australian Institute forNuclear Science and Engineering grant (ALNGRA11036). Thanksalso to Matt Cupper of the School of Earth Sciences, University ofMelbourne, for his assistance with calibrating the radiocarbondates for the plant samples.
References
Alfonso, M.P., Standen, V.G., Castro, M.V., 2007. The adoption of agriculture amongnorthern Chile populations in the Azapa Valley, 9000e1000 BP. In: Cohen, M.N.,Crane-Kramer, G.M.M. (Eds.), Ancient Health. University Press of Florida, Flor-ida, pp. 113e129.
Allison, M., Focacci, G., Arriaza, B., Standen, V., Rivera, M., Loweinstein, J., 1984.Chinchorro momias de preparación complicada. Chungara 13, 155e173.
Ambrose, S.H., 1990. Preparation and characterization of bone and tooth collagenfor isotopic analysis. Journal of Archaeological Science 17, 431e451.
Ambrose, S.H., 1991. Effects of diet, climate and physiology on nitrogen isotopeabundances in terrestrial foodwebs. Journal of Archaeological Science 18, 293e318.
Ambrose, S.H., 1993. Isotopic analysis of paleodiets: methodological and inter-pretive considerations. In: Sandford, M.K. (Ed.), Investigations of Ancient Hu-man Tissue: Chemical Analysis in Anthropology. Gordon and Breach SciencePublishers, Pennsylvania, pp. 59e130.
Ambrose, S.H., Butler, B.M., Hanson, D.B., Hunter-Anderson, R.L., Krueger, H.W.,1997. Stable isotope analysis of human diet in the Marianas Archipelago,Western Pacific. American Journal of Physical Anthropology 104, 343e361.
Ambrose, S.H., DeNiro, M.J., 1986. The isotope ecology of East African mammals.Oecologia 69, 395e406.
Arneborg, J., Heinemeier, J., Lynnerup, N., Nielsen, H.L., Rud, N.,Sveinbjörnsdóttir, A.E., 1999. Change of diet of the Greenland Vikings deter-mined from stable isotope analysis and 14C dating of their bones. Radiocarbon41 (2), 157e168.
Arriaza, B.T., 1988. Modelo bioarqueológico para la búsqueda y acercamiento alindividual social. Chungara 21, 9e32.
Arriaza, B.T., 1995. Chinchorro bioarchaeology: chronology and mummy seriation.Latin American Antiquity 6, 35e55.
Arriaza, B.T., Standen, V.G., 2002. Death, Mummies, and Ancestral Rites: the Chin-chorro Culture. Universidad de Tarapaca, Arica.
Arriaza, B.T., Standen, V.G., Belmonte, E., Rosello, E., Nials, F., 2001. The peopling of theArica coast during the preceramic: a preliminary view. Chungara 33 (1), 16e31.
Arriaza, B.T., Standen, V.G., Cassman, V., Santoro, C.M., 2008. Chichorro culture:pioneers of the coast of the Atacama Desert. In: Silverman, H., Isbell, W.H. (Eds.),Handbook of South American Archaeology. Springer, New York, pp. 45e58.
Araujo de, A.J.G., Ferreira, L.F., Contalonieri, V.E.C., Riberiro Filho, B.M., Núñez, L.,1985. The finding of enteroblus vernicularis eggs in pre-columbian humancoprolites. Memorias do Instituto Oswaldo Cruz 80, 141e143.
Aufderheide, A.C., 1993. Reconstrucción química de la dieta del hombre de Acha. In:Muñoz, I., Arriaza, B., Aufderheide, A. (Eds.), Acha 2 y los Orígenes del Pobla-miento Humano en Arica, Ediciones. Universidad de Tarapacá, Arica, pp. 65e80.
Aufderheide, A.C., Allison, M.J., 1992. Chemical dietary reconstruction of north Chileprehistoric populations by trace mineral analysis. In: Manuscript Read at theFirst World Congress on Mummy Studies, Puerto de la Cruz, Canary Islands,Spain, 3e6 February.
Aufderheide, A.C., Kelly, M.A., Rivera, M.A., Gray, L., Tiezen, L.L., Iversen, E.,Krouse, H.R., Carevic, A., 1994. Contributions of chemical dietary re-constructions to the assessment of adaptations by highland immigrants tocoastal conditions at Pisagua, North Chile. Journal of Archaeological Science 21,515e524.
Aufderheide, A.C., Munoz, I., Arriaza, B., 1993. Seven Chinchorro mummies and theprehistory of northern Chile. American Journal of Physical Anthropology 91,189e201.
Aufderheide, A.C., Santoro, C.M., 1999. Chemical paleodietary reconstruction: hu-man populations at late prehistoric sites in the Lluta Valley of northern Chile.Revista Chilena de Historia Natural 72, 237e250.
Barberena, R., Gil, A.F., Neme, G.A., Tykot, R.H., 2009. Stable isotopes and archaeologyin Southern South America. Hunteregatherers, pastoralism and agriculture: anintroduction. International Journal of Osteoarchaeology 19, 127e129.
Bittmann, B., Munizaga, J., 1976. The earliest artificial mummification in the world?A study of the Chinchorro complex in northern Chile. Folk 18, 61e92.
Brock, F., Bronk Ramsey, C., Highham, T.F.G., 2007. Quality assurance of ultrafilteredbone dating. Radiocarbon 49 (2), 187e192.
Bronk Ramsey, C., 2001. Development of the radiocarbon calibration programOxCal. Radiocarbon 43 (2A), 355e363.
Bronk Ramsey, C., Higham, T., Bowles, A., Hedges, R., 2004. Improvements to thepretreatment of bone at Oxford. Radiocarbon 46 (1), 155e163.
Brown, T.A., Nelson, D.E., Vogel, J.S., Southon, J.R., 1988. Improved collagen extrac-tion by modified Longin method. Radiocarbon 30 (2), 171e177.

A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e2372 2371
Castelleti, J., 2007. Patrón de asentamiento y uso de recursos a través de la secuenciaocupacional prehispánica de la costa de Taltal. Tesis de Magíster en Antropo-logía con mención en Arqueología, Programa de Postgrado Universidad Católicadel Norte-Universidad de Tarapacá, Antofagasta-Arica.
Coplen, T.B., Brand, W.A., Gehre, M., Gröning, M., Meijer, H.A.J., Toman, B.,Verkouteren, R.M., 2006. New guidelines for d13C measurements. AnalyticalChemistry 78, 2439e2441.
Corr, L.T., Sealy, J.C., Horton, M.C., Evershed, R.P., 2005. A novel marine dietary in-dicator utilising compound-specific bone collagen amino acid d13C values ofancient humans. Journal of Archaeological Science 32 (3), 321e330.
DeFrance, S.D., Umire, A., 2004. Quebrada Tacahuay: un sitio marítimo del Pleis-toceno tardío en la costa sur del Perú. Chungara Revista de Antropología Chi-lena 36, 257e278.
Deines, P., 1980. The isotopic composition of reduced organic carbon. In: Fritz, P.,Fontes, J.Ch (Eds.), Handbook of Environmental Isotope Geochemistry, vol. 1.Elsevier, Amsterdam, pp. 329e406.
DeNiro, M.J., 1985. Postmortem preservation and alteration of in-vivo bone collagenisotope ratios in relation to paleodietary reconstructions. Nature 317, 806e809.
DeNiro, M.J., Epstein, S., 1978. Influence of diet on the distribution of carbon iso-topes in animals. Geochimica et Cosmochimica Acta 42, 495e506.
DeNiro, M.J., Epstein, S., 1981. Influence of diet on the distribution of nitrogenisotopes in animals. Geochimica et Cosmochimica Acta 45, 341e351.
DeNiro, M.J., Weiner, S., 1988. Chemical, enzymatic and spectroscopic characteri-sation of “collagen” and other organic fractions from archaeological bones.Geochimica et Cosmochimica Acta 52, 2425e2431.
Dorsey Vinton, S., Perry, L., Reinhard, K.J., Santoro, C.M., Teixeira-Santos, I., 2009.Impact of empire expansion on household diet: the Inka in northern Chile’sAtacama Desert. PLoS ONE 4 (11), e8069.
Ehleringer, J.R., 1989. Carbon isotope ratios and physiological processes in arid-landplants. In: Rundel, P.W., Ehleringer, J.R., Nagy, K.A. (Eds.), Stable Isotopes inEcological Research. SpringereVerlag, New York, pp. 41e54.
Falabella, F., Planella, M.T., Aspillaga, E., Sanhueza, L., Tykot, R.H., 2007. Dieta ensociedades alfareras de Chile central: aporte de analisis de isotopos estables.Chungara Revista de Antropologia 39, 5e27.
Fink, D., Hotchkis, M., Hua, Q., Jacobsen, G., Smith, A.M., Zoppi, U., Child, D.,Mifsud, C., van der Gaast, H., Williams, A., Williams, M., 2004. The ANTARESAMS facility at ANSTO. Nuclear Instruments and Methods B 223e224, 109e115.
Fontugne, M., Carré, M., Bentaleb, I., Julien, M., Lavallée, D., 2004. Radiocarbonreservoir age variations in the south Peruvian upwelling during the Holocene.Radiocarbon 46, 531e537.
Gil, A.F., Neme, G.A., Tykot, R.H., 2010. Isótopos Estables y Consumo de Maíz en elCentro Occidente Argentino: Tendencias Temporales y Espaciales. ChungaraRevista de Antropología Chilena 42, 497e513.
Grosjean, M., Núñez, L., Cartajena, I., Messerli, B., 1997. Mid-Holocene climate andculture change in the Atacama Desert, Northern Chile. Quaternary Research 48,239e246.
Heaton, T.H.E., Vogel, J.C., von la Chevallerie, G., Collett, G., 1986. Climatic influenceon the isotopic composition of bone nitrogen. Nature 322, 822e823.
Heiemeier, J., Rud, N., 1997. Danske arkæologiske AMS 14C dateringer, Århus 1996.Arkæologiske Udgravninger I Danmark 1996, 300e305.
Higham, T.F.G., Jacobi, R.M., Bronk Ramsey, C., 2006. AMS radiocarbon dating ofancient bone using ultrafiltration. Radiocarbon 48 (2), 179e195.
Hoefs, J., 1987. Stable Isotope Geochemistry. SpringereVerlag, New York.Hua, Q., Jacobsen, G.E., Zoppi, U., Lawson, E.M., Williams, A.A., Smith, A.M.,
McMann, M.J., 2001. Progress in radiocarbon target preparation at the ANTARESAMS Centre. Radiocarbon 43 (2a), 275e282.
Iacumin, P., Bocherens, H., Delgado Huertas, A., Mariotti, A., Longinelli, A., 1997.A stable isotope study of fossil mammal remains from the Paglicci cave,Southern Italy. N and C as palaeoenvironmental indicators. Earth and PlanetaryScience Letters 148, 349e357.
Janusek, J.W., 2008. Ancient Tiwanaku. Cambridge University Press, New York.Katzenberg, M.A., 2000. Stable isotope analysis: a tool for studying past diet,
demography and life history. In: Katzenberg, M.A., Saunders, S.R. (Eds.), Bio-logical Anthropology of the Human Skeleton. Wiley, New York, pp. 305e327.
Kelley, M.A., Levesque, D.R., Weidl, E., 1991. Contrasting patterns of dental disease infive early northern Chilean groups. In: Kelley, M.A., Larson, C.S. (Eds.), Advancesin Dental Anthropology. Wiley-Liss, New York, pp. 203e213.
Knudson, K.J., Aufderheide, A.E., Buikstra, J.E., 2007. Seasonality and paleodiet in theChiribaya polity of southern Peru. Journal of Archaeological Science 23 (3),451e462.
Latorre, C., Betancourt, J.L., Rylander, K.A., Quade, J., Matthei, O., 2003. A vegetationhistory from the arid Prepuna of Northern Chile (22e23�S) over the last 13,500years. Palaeogeography, Palaeoclimatology, Palaeoecology 194, 223e246.
Latorre, C., Betancourt, J.L., Rech, J.A., Quade, J., Holmgren, C., Placzek, C.,Maldonado, A., Vuille, M., Rylander, K.A., 2005. Late Quaternary history of theAtacama desert. In: Smith, M., Hesse, P. (Eds.), 23� South: The Archaeology andEnvironmental History of the Southern Desert. National Museum of Australia,Canberra, pp. 73e90.
Larsen, C.S., 1997. Bioarchaeology: Interpreting Behavior from the Human Skeleton.Cambridge University Press, New York.
Lee-Thorp, J.A., 2008. On isotopes and old bones. Archaeometry 50 (6), 925e950.Libby, W.F., Berger, R., Mead, J.F., Alexander, G.V., Ross, J.F., 1964. Replacement rates
for human tissue from atmospheric radiocarbon. Science 146, 1170e1172.Little, E.A., Schoeninger, M.J., 1995. The Late Woodland diet on Nantucket Island and
theproblemofmaize in coastalNewEngland.AmericanAntiquity60 (2), 351e368.
Llagostera, A., 1979. 9,700 years of maritime subsistence on the Pacific: an analysisby means of bioindicators in the north of Chile. Society for American Archae-ology 44, 309e324.
Longin, R., 1971. New method of collagen extraction for radiocarbon dating. Nature230, 241e242.
Marquet, P.A., Santoro, C.M., Latorre, C., Standen, V.G., Abades, S.R.,Rivadeneira, M.M., Arriaza, B., Hochberg, M.E., 2012. Emergence of socialcomplexity among coastal hunteregatherers in the Atacama Desert of northernChile. PNAS 109, 14754e14760.
McCormac, F.G., Hogg, A.G., Blackwell, P.G., Buck, C.E., Higham, T.F.G., Reimer, P.J.,2004. SHCal04 Southern Hemisphere calibration 0e11.0 cal kyr BP. Radiocarbon46, 1087e1092.
Moraga, M., Santoro, C.M., Standen, V., Carvallo, P., Rothhammer, F., 2005. Micro-evolution in prehistoric Andean populations: chronologic mtDNA variation inthe desert valleys of northern Chile. American Journal of Physical Anthropology127, 170e181.
Nester, P., Gayó, E., Latorre, C., Jordan, T.E., Blanco, N., 2007. Perennial stream dis-charge in the hyperarid Atacama Desert of northern Chile during the latestPleistocene. PNAS 104, 19724e19729.
Núñez, L., Grosjean, M., Cartajena, I., 2002. Human occupations and climate changein the Puna de Atacama, Chile. Science 298, 821e824.
Núñez, L., Hall, H.J., 1982. Análisis de dieta ymovilidad en un campamento arcaico delnorte de Chile. Boletín del Instituto Francés de Estudios Andinos 11 (3e4), 91e113.
Núñez, L., Santoro, C.M., 2011. El tránsito Arcaico-Formativo en la circunpuna yvalles occidentales del Centro Sur Andino: hacia los cambios “neolíticos”.Chungara Revista de Antropología Chilena 43, 487e531.
O’Leary, M.H., 1981. Carbon isotope fractionation in plants. Phytochemistry 20,553e567.
O’Leary, M.H., 1988. Carbon isotopes in photosynthesis. BioScience 38, 328e336.Ortlieb, L., Vargas, G., Saliege, J.-L., 2011. Marine radiocarbon reservoir effect along
the northern Chile-southern Peru coast (14-24oS) throughout the Holocene.Quaternary Research 75, 91e103.
Osorio, D., Jackson, D., Ugalde, P.C., Latorre, C., de Pol-Holz, R., Santoro, C.M., 2011. Lithicanalyses from a late Pleistocene logistical camp, the Hakenasa cave and its rele-vance for the peoplingof the southernAndeanAltiplano. Antiquity 85,1194e1208.
Owen, B.D., 2002. Marine carbon reservoir age estimates for the far south coast ofPeru. Radiocarbon 44 (3), 701e708.
Pate, F.D., 1994. Bone chemistry and paleodiet. Journal of Archaeological Methodand Theory 1, 161e209.
Pate, F.D., 1997. Bone chemistry and paleodiet: reconstructing prehistoricsubsistence-settlement systems in Australia. Journal of AnthropologicalArchaeology 16, 103e120.
Pate, F.D., 2008. The use of human skeletal remains in landscape archaeology. In:David, B., Thomas, J. (Eds.), Handbook of Landscape Archaeology. Left CoastPress, Walnut Creek, California, pp. 502e520.
Pate, F.D., Anson, T.J., 2008. Stable nitrogen isotope values in arid-land kangarooscorrelated with mean annual rainfall: potential as a palaeoclimatic indicator.International Journal of Osteoarchaeology 18, 317e326.
Peterson, B.J., Fry, B., 1987. Stable isotopes in ecosystem studies. Annual Review ofEcology and Systematics 18, 293e320.
Qi, H., Coplen, T.B., Geilmann, H., Brand, W.A., Böhlke, J.K., 2003. Two new organicreferencematerials for d13C and d15Nmeasurements and a newvalue for the d13Cof NBS 22 oil. Rapid Communications in Mass Spectrometry 17, 2483e2487.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., BronkRamsey, C., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M.,Guilderson, T.P., Hajdas, I., Heaton, T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F.,Kromer, B., McCormac, F.G., Manning, S.W., Reimer, R.W., Richards, D.W.,Southon, J.R., Talamo, S., Turney, C.S.M., van der Plicht, J., Weyhenmeyer, C.E.,2009. IntCal09 and Marine09 radiocarbon age calibration curves, 0e50,000years cal BP. Radiocarbon 51, 1111e1150.
Rivadeneira, M., Santoro, C.M., Marquet, P.A., 2010. Reconstructing the history ofhuman impacts on coastal biodiversity in Chile: constraints and opportunities.Aquatic Conservation: Marine and Freshwater Ecosystems 20, 74e82.
Rivera, M.A., 1991. The prehistory of northern Chile: a Synthesis. Journal of WorldPrehistory 5, 1e47.
Rothhammer, F., Cocilovo, J.A., 2008. Microdiferenciación craneana en la costa deArica y valle de Azapa, norte de Chile. Resumen de resultados y conclusions.Chungara Revista de Antropología Chilena 40, 309e315.
Rothhammer, F., Santoro, C.M., Poulin, E., Moraga, M., Standen, V.G., 2009.Archeological and mtDNA evidence for tropical lowland migrations and culturalchange during the Late Archaic/Formative in northern Chile. Revista Chilena deHistoria Natural 82, 543e552.
Sandweiss, D.H., McInnis, H., Burger, R., Cano, A., Ojeda, B., Paredes, R., del CarmenSandweiss, M., Glascock, M., 1998. Quebrada Jaguay: early South Americanmaritime adaptations. Science 281, 1830e1832.
Sandweiss, D.H., Richardson, J.B., Reitz, E.J., Rollins, H.B., Maasch, K.A., 1996. Geo-archaeological evidence from Peru for a 5000 years B.P. onset of El Niño. Science273, 1531e1533.
Santoro, C., Arriaza, B., Standen, V., Marquet, P., 2005b. People of the coastal Ata-cama Desert: living between sand dunes and waves of the Pacific Ocean. In:Veth, P., Smith, M., Hiscock, P. (Eds.), Desert Peoples: Archaeological Perspec-tives. Blackwell Publishing, Melbourne, pp. 243e260.
Santoro, C.M., Dorsey Vinton, S., Reinhard, K.J., 2003. Inka expansion and parasitismin the Lluta valley: preliminary data. Memórias do Instituto Oswaldo Cruz 98(Supp. I), 161e163.

A. Roberts et al. / Journal of Archaeological Science 40 (2013) 2360e23722372
Santoro, C., Latorre, C., Salas, C., Osorio, D., Ugalde, P., Jackson, D., Gayó, E., 2011.Ocupación humana pleistocénica en el desierto de atacama. Primeros resulta-dos de la aplicación de un modelo predictivo interdisciplinario. ChungaraRevista de Antropologia Chilena 43, 353e366.
Santoro, C.M., Rivadeneira, M.M., Latorre, C., Rothhammer, F., Standen, V.G., 2012.Rise and decline of Chinchorro sacred landscapes along the hyperarid coast ofthe Atacama desert. Chungara Revista de Antropología Chilena 44, 637e653.
Santoro, C.M., Standen, V.G., Arriaza, B.T., Marquet, P.A., 2005a. Hunter gatherers onthe coast and hinterland of the Atacama Desert 17e278� south latitude. In:Smith, M., Hesse, P. (Eds.), 23� South: The Archaeology and EnvironmentalHistory of the Southern Desert. National Museum of Australia, Canberra,pp. 172e185.
Santoro, C.M., Williams, V., Valenzuela, D., Romero, Á., Standen, V.G., 2010. Anarchaeological perspective on the Inka provincial administration from theSouth Central Andes. In: Malpass, M.A., Alconini, S. (Eds.), Distant Provinces inthe Inka Empire: Toward a Deeper Understanding of Inka Imperialism. Uni-versity of Iowa Press, Iowa City, pp. 44e74.
Schoeninger, M.J., DeNiro, M.J., 1984. Nitrogen and carbon isotopic composition ofbone collagen from marine and terrestrial animals. Geochimica et Cosmochi-mica Acta 48, 625e639.
Schoeninger, M., DeNiro, M., Tauber, H., 1983. Stable nitrogen isotope ratios of bonecollagen reflect marine and terrestrial components of prehistoric human diet.Science 220, 1381e1383.
Schoeninger, M.J., Moore, K.M., Murray, M.L., Kingston, J.D., 1989. Detection of bonepreservation in archaeological and fossil samples. Applied Geochemistry 4,281e292.
Sealy, J.C., 1986. Stable Carbon Isotopes and Prehistoric Diets in the SouthwesternCape Province, South Africa. In: British Archaeological Reports, InternationalSeries 293. Oxford.
Sealy, J.C., Armstrong, R., Shrire, C., 1995. Beyond life-time averages: tracing lifehistories through isotopic analysis of different calcified tissues from archaeo-logical human skeletons. Antiquity 69, 290e300.
Sealy, J.C., van der Merwe, N.J., Lee-Thorp, J.A., Lanham, J.L., 1987. Nitrogen isotopicecology in southern Africa: implications for environmental and dietary tracing.Geochimica et Cosmochimica Acta 51, 2707e2717.
Sillen, A., Parkington, J., 1996. Digenesis of bones from Eland’s Bay Cave. Journal ofArchaeological Science 23, 535e542.
Slovak, N.M., Paytan, A., 2009. Fisherfolk and farmers: carbon and nitrogen isotopeevidence from Middle Horizon Ancón, Peru. International Journal of Osteo-archaeology 21 (3), 253e267.
Southon, J.R., Oakland Rodman, A., True, D., 1995. A comparison of marine andterrestrial radiocarbon ages from northern Chile. Radiocarbon 37, 389e393.
Stafford, T.W., Jull, A.J.T., Brendel, K., Duhamel, R.C., Donahue, D., 1987. Bone 14Cdating with Arizona TAMS. Radiocarbon 29 (1), 24e44.
Standen, V.G., Santoro, C.M., 2004. Patrón funerario arcaico temprano del sitioAcha-3 y su relación con Chinchorro (costa norte de Chile). Latin AmericanAntiquity 15, 89e109.
Standen, V.G., Santoro, C.M., Arriaza, B.T., 2004. Síntesis y propuestas para elperíodo Arcaico en el extremo norte de Chile. Chungara Revista de AntropologíaChilena Volumen Especial, 201e212.
Stenhouse, M.J., Baxter, M.S., 1979. The uptake of bomb 14C in humans. In: Berger, R.,Suess, H.E. (Eds.), Radiocarbon Dating. University of California Press, Berkeley,pp. 324e341.
Stuiver, M., Braziunas, T.F., 1993. Modelling atmospheric 14C influences and 14C agesof marine samples to 10,000 BC. Radiocarbon 35 (1), 137e189.
Stuiver, M., Reimer, P.J., 1993. Extended 14C data base and revised CALIB 3.0 14C agecalibration program. Radiocarbon 35, 215e230.
Taylor, R.E., Berger, R., 1967. Radiocarbon content of marine shells from the Pacificcoasts of Central and South America. Science 158 (3805), 1180e1182.
Tieszen, L.L., Boutton, T.W., 1988. Stable carbon isotopes in terrestrial ecosystemresearch. In: Rundel, P.W., Ehleringer, J.R., Nagy, K.A. (Eds.), Stable Isotopes inEcological Research. SpringereVerlag, New York, pp. 167e195.
Tieszen, L.L., Chapman, M., 1995. Carbon and nitrogen isotopic status of the majormarine and terrestrial resources in the Atacama desert of northern Chile. In:Proceedings of the First World Congress on Mummy Studies (February 3e6,1992).
Tisnérat-Laborde, N., Valladas, H., Kaltnecker, E., Arnold, M., 2003. AMS radiocarbondating of bones at LSCE. Radiocarbon 45 (3), 409e419.
Tripp, J.A., McCullagh, J.S.O., Hedges, R.E.M., 2006. Preparative separation ofunderivatized amino acids for compound-specific stable isotope analysis andradiocarbon dating of hydrolyzed bone collagen. Journal of Separation Science29, 41e48.
Turner, B.L., Kingston, J.D., Armelagos, G.J., 2010. Variation in dietary historiesamong the immigrants of Machu Picchu: carbon and nitrogen isotope evidence.Chungara Revista de Antropología Chilena 42, 515e534.
Tykot, R.H., Falabella, F., Planella, M.T., Aspillaga, E., Sanhueza, L., Becker, C., 2009.Stable isotopes and archaeology in central Chile: methodological insights andinterpretative problems for dietary reconstruction. International Journal ofOsteoarchaeology 19, 156e170.
Ugalde, P.C., Salas, C., Latorre, C., Osorio, D., Jackson, D., Santoro, C.M., 2012.Poblamiento temprano del norte de Chile (18e25�s): estudio interdisciplinarioarqueológico y paleoambiental. In: Castro, V., Cases, B. (Eds.), Actas XVIII Con-greso Nacional de Arqueología Chilena, pp. 197e206.
Uarrota, V.G., Schmidt, E.C., Bouzon, Z.L., Maaschin, M., 2011. Histochemical analysisand protein content of maize landraces (Zea mays L.). Journal of Agronomy 10(3), 93e98.
Urey, H.C., 1947. The thermodynamic properties of isotopic substances. Journal ofthe Chemical Society, 562.
Van der Merwe, N.J., 1982. Carbon isotopes, photosynthesis, and archaeology.American Scientist 70, 596e606.
Van Klinken, G.J., 1999. Bone quality indicators for palaeodietary and radiocarbonmeasurements. Journal of Archaeological Science 26, 687e695.
Van Klinken, G.J., Hedges, R.E.M., 1995. Experiments on collagen-humic in-teractions: speed of humic uptake, and effects of diverse chemical treatments.Journal of Archaeological Science 22, 263e270.
Van Klinken, G.J., Mook, W.G., 1990. Preparative high-performance liquid chroma-tographic separation of individual amino acids derived from fossil bone. Radi-ocarbon 32 (2), 155e164.
Vogel, J.S., Southon, J.R., Nelson, D.E., Brown, T.A., 1984. Performance of catalyticallycondensed carbon for use in accelerator mass spectrometry. Nuclear In-struments and Methods in Physics Research Section B: Beam Interactions withMaterials and Atoms 5 (2), 289e293.
Williams, A., Santoro, C., Smith, M., Latorre, C., 2008. The impact of ENSO in theAtacama Desert and the Australian Arid Zone: exploratory time-series anal-ysis of archaeological records. Chungara Revista de Antropología Chilena 40,245e259.
![Prospección arqueológica de la Caleta de San Marcos (Icod de los Vinos, Tenerife [Archaeological Survey of the Port of San Marcos (Icod de los Vinos, Tenerife, Canary Islands)]](https://static.fdokumen.com/doc/165x107/6323baa74d8439cb620d0c4d/prospeccion-arqueologica-de-la-caleta-de-san-marcos-icod-de-los-vinos-tenerife.jpg)



















