
Response of common purslane to dicamba treatment. - CORE
129
University of Massachusetts Amherst University of Massachusetts Amherst ScholarWorks@UMass Amherst ScholarWorks@UMass Amherst Doctoral Dissertations 1896 - February 2014 1-1-1972 Response of common purslane to dicamba treatment. Response of common purslane to dicamba treatment. Maria Stacewicz-Sapuncakis University of Massachusetts Amherst Follow this and additional works at: https://scholarworks.umass.edu/dissertations_1 Recommended Citation Recommended Citation Stacewicz-Sapuncakis, Maria, "Response of common purslane to dicamba treatment." (1972). Doctoral Dissertations 1896 - February 2014. 5903. https://scholarworks.umass.edu/dissertations_1/5903 This Open Access Dissertation is brought to you for free and open access by ScholarWorks@UMass Amherst. It has been accepted for inclusion in Doctoral Dissertations 1896 - February 2014 by an authorized administrator of ScholarWorks@UMass Amherst. For more information, please contact [email protected].
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Response of common purslane to dicamba treatment. - CORE

University of Massachusetts Amherst University of Massachusetts Amherst
ScholarWorks@UMass Amherst ScholarWorks@UMass Amherst
Doctoral Dissertations 1896 - February 2014
1-1-1972
Response of common purslane to dicamba treatment. Response of common purslane to dicamba treatment.
Maria Stacewicz-Sapuncakis University of Massachusetts Amherst
Follow this and additional works at: https://scholarworks.umass.edu/dissertations_1
Recommended Citation Recommended Citation Stacewicz-Sapuncakis, Maria, "Response of common purslane to dicamba treatment." (1972). Doctoral Dissertations 1896 - February 2014. 5903. https://scholarworks.umass.edu/dissertations_1/5903
This Open Access Dissertation is brought to you for free and open access by ScholarWorks@UMass Amherst. It has been accepted for inclusion in Doctoral Dissertations 1896 - February 2014 by an authorized administrator of ScholarWorks@UMass Amherst. For more information, please contact [email protected].


RESPONSE OF COMMON PURSLANE
TO DICAMSA TREATMENT
A Dissertation Presented
By
Maria Stacewicz-Sapuncakis
Submitted to the Graduate School of the University of Massachusetts in
partial, fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY
February, 1972
Plant and Soil Sciences

RESPONSE OF COMMON PURSLANE TO DICAMBA TREATMENT
A Dissertation Presented
By
Maria Stacewicz-Sapuncakis
Approved as to style and content by:
11 -Cii ' > /(^hairman of Committee)
C' ■/ : // ' ■
to -"Ml
r *
(Head of Department)
-7
(Member)
>ucy \ T ^ 7 ///cl/ -/'I
(Member)
(Member) U ^
(Month) (Year)

iii
ACKNOWL EDGEMENT S
Thanks are given to my advisor, Dr. J. Vengris, for helping to
choose the topic of this thesis.
I am grateful to Dr. H.V. Marsh and Dr. P.R. Jennings for their
valuable discussion and suggestions. My thanks are also due to them
for help in editing this thesis.
Helpful instruction of Dr. T. Robinson especially in the use of
radiochromatogram scanner is gratefully acknowledged.
Thanks are given to Dr. J.H. Baker for his supervision of the
experiments involving gas chromatography and atomic absorption
spectrophotometry.
I wish to express my sincere gratitude to Dr. F.W. Southwick,
Head of Plant and Soil Sciences Department, for his understanding
and encouragement.
Thanks are due to Velsicol Chemical Corporation for supplying
radioactive dicamba and other chemicals.

iv
TABLE OF CONTENTS
Page
INTRODUCTION. 1
LITERATURE REVIEW. 3
Characteristics of Common Purslane. 3
Dicamba as a Systemic Herbicide. 5
A. Characteristics of Dicamba. 5
B. Dissipation of Dicamba in Soil. 6
C. Anatomical and Morphological Responses of Plants to Dicamba. 6
D. Absorption of Dicamba in Plants. 8
E. Translocation of Dicamba in Plants'. 9
F. Metabolism of Dicamba in Plants. 13
Influence of Herbicides on Cell Membrane Permeability. 15
Influence of Herbicides on Protein Metabolism. 16
Role of Ethylene in Plant Growth. 20
A. Anatomical and Morphological Responses of Plants to Ethylene. 20
B. Physiological Actions of Ethylene in Plants. 22
C. Promotion of Ethylene Production in Plants. 27
D. Estimation of Internal Level of Ethylene in Plants... 33
MATERIALS AND METHODS. 35
Plant Material. 35
Cultural Procedure. 35
Effect of Dicamba on Growth and Development. 35
Effect of Ethylene on Common Purslane. 36

V
Page
Measurements of Cell Membrane Permeability. 36
A. Efflux of Electrolytes from Stem Segments. 36
B, Rubidium Release by Excised Leaves. 37
Protein Synthesis Studies. 38
A. Nitrate Reductase Activity. 39
B. Incorporation of Phenylalanine-C^'^. 40
Ethylene Evolution Measurements. 42
Studies of the Fate of Dicamba in Common Purslane. 43
A. Preparation of Plant Extracts. 44
B, Chromatography of Plant Extracts. 45
Statistical Evaluation. 46
RESULTS. 47
Morphological Response of Common Purslane Plants to Dicamba... 47
Influence of Dicamba on Growth and Fresh Weight of Purslane... 51
Morphological Response of Common Purslane Plants to Ethylene. 54
Studies of Cell Membrane Permeability in Common Purslane. 57
A. Efflux of Electrolytes from Purslane Stem Segments... 59
B. Efflux of Rubidium from Purslane Leaves. 62 i
Studies of Protein Metabolism in Purslane Leaves. 62
A. Nitrate Reductase Activity in Purslane Leaves. 64
B. Studies of Protein Synthesis in Purslane Leaves. 67
Effect of Dicamba on Ethylene Production in Purslane Plants... 72
Absorption and Translocation of Dicamba in Purslane Plants.... 75
Studies of Dicamba Metabolism in Purslane Plants. 83

vi
Page
DISCUSSION... 87
Susceptibility of Conmion Purslane to Dicamba. 87
Susceptibility of Common Purslane to Ethylene Fumigation. 88
Effect of Dicamba on Cell Membrane Permeability in
Purslane. 89
Effect of Dicamba on Protein Synthesis in Purslane. 91
Effect of Dicamba on Ethylene Production in Purslane. 92
Absorption and Translocation of Dicamba in Purslane Plants.... 95
Studies of Dicamba Detoxification in Common Purslane. 97
SUMMARY. 99
BIBLIOGRAPHY. 101
APPENDIX 114

Vll
LIST OF TABLES
Page
1. Morphological response of 4 week old purslane plants to various rates of foliarly applied dicamba. 48
2. Effect of dicamba on growth of 4 week old purslane plants as laeasured by the length of the main stem (cm). 52
3. Effect of dicamba on the fresh weight of purslane shoots harvested 25 days after application (in g per pot). 55
4. Morphological response of 4 week old purslane plants to
various levels of ethylene fumigation. 56
5. Efflux of electrolytes from purslane stem sections after dicamba treatment (in percent of total content of electrolytes). 60
6. Total content of electrolytes in purslane stem sections as
expressed in mhos/g fr. wt. 61
7. Nitrate reductase activity in leaves of purslane plants treated with dicamba. 65
8. Nitrate reductase activity in leaves of purslane plants treated with 1 ppm ethylene. 66
9. Phenylalanine-^^^C incorporation into protein in leaves of purslane plants treated with 10 yg dicamba. 68
10. Phenylalanine-^^C incorporation into protein in leaves of purslane plants treated with 1 ppm ethylene or 50 yg cycloheximide. 69
11. Phenylalanine-^^C uptake by leaves of purslane plants treated with 10 yg dicamba, 1 ppm ethylene or 50 yg
cycloheximide. 70
12. Protein content in leaves of purslane plants treated with
10 pg cycloheximide. 71
13. Absorption of dicamba-^^C in purslane plants expressed as
perccmt of the applied dose. 76
14. Translocation of dicamba-^^C in purslane plants after foliar application; in cpm per plant part. 78

viii
Page
15. Distribution of dicamba-^^C in purslane plants expressed in percent of the absorbed amount. 80
16. Concentration of radioactivity in \arious organs of purslane after foliar application of 0.1 yC of dicamba-^^C ; in cpm/g fr. wt. 81
17. Distribution of radioactivity along chromatograms of extracts from purslane plants 8 days after application of dicamba-^'^C to a single leaf. 85

ix
LIST OF FIGURES
Page
1. Formative effects of dicamba on common purslane plants. 50
2. Common purslane plants after various dicamba treatments. 53
3. Purslane plants 24 hrs after dicamba (10 yg/plant) and
ethylene (Ippm) treatment as compared to the untreated control. 58
4. Elfflux of rubidium from purslane leaves after dicamba and ethylene treatments. 63
5. Ethylene production in detached purslane leaves at various rates of dicamba treatment. 74
6. Translocation of dicamba-^'^C in purslane plants during the first day after treatment. 77
7. Concentration of radioactivity in purslane plant parts during the first day after application of 0.1 yC dicamba-C^^ to a single leaf. 82
8. Autoradiograph of thin layer chromatogram of extract of purslane plants treated with dicamba-^^C. 86
APPENDIX
Figure I. Standard curve for rubidium determination......... 114
Figure II. Standard curve for nitrite determination. 115
Figure III. Protein standard for biuret test. 116
Figure IV, Standard curves for ethylene determination by
gas chromatography. 117

1
INTRODUCTION
Dicamba (2~methoxy -3,6- dichlorobenzoic acid) is a systemic
herbicide which has been used effectively to control many broadleaved
weeds resistent to other commercial herbicides. Common purslane
(Portulaca oleracea L.) belongs to a group of plants which are sensitive
to dicamba treatments although not easily controlled by 2,A-D (2,4-
dichlorophenoxyacetic acid) or 2,4,5-T (2,4,5-trichlorophenoxyacetic
acid) .
The present investigation was planned to study the response of
common purslane to dicamba treatment with consideration of morphological
and physiological effects on the weed and to examine the absorption,
translocation and metabolism of the herbicide in the treated plants.
With this type of information it should be possible to determine the
mode of action of dicamba as it affects purslane.
Auxins, such as lAA (indole-3-acetic acid), NAA (1-naphtalene-
1-acetic acid) and systemic herbicides such as 2,4-D, 2,4,5-T and
picloram (4-amino-3,5,6 -trichloropicolinic acid) are known to
stimulate ethylene production in treated plants (1, 115, 126). Their
mode of action is partly dependent on the action of ethylene thus
produced. Preliminary observations indicated that some of the
responses of purslane to dicamba treatments were similar to well
known ethylene induced effects. Therefoie an attempt was made to
compare the physiological action of ethylene and dicamba in purslane
and to explore their interaction.
The literature on the subject suggests the possibility that many
herbicides and ethylene act upon protein synthesis and cell membrane

2
permeability (124, 144, 150). In the following work these two physio¬
logical aspects were investigated to compare the influence of dicamba
and ethylene.

3
LITERATURE REVIEW
Characteristics of Common Purslane
Common purslane (Portulaca oleracea L.)> 3. native plant of Eurasia,
is widespread in tropical and temperate lands (104, 135, 164). It is
a very troublesome, annual, broadleaved weed species commonly appearing
in vegetable gardens, fields, and turf areas. This weed is currently
spreading in the northeast region of the United States (163).
A prostrate growth pattern and the succulent character of this
plant, which has a thick stem and small leaves, make it very resistant
to eradication by common mechanical methods such as cultivation or
mowing (59, 133). Uprooted, mature purslane plants left in the field
can develop seeds due to internal water supplies and a very low
transpiration coefficient -292 g water per gram of fresh weight (28) .
Fragments of plants left on moist soil soon form new plants by the
formation of adventitious roots (51) . In the leaves of a related
species Portulaca grandiflora L. a root promoting substance was found
(122). It was named portulal and described as a bicyclic diterpene
containing a perhydroazulene nucleus. It appeared to influence
specifically the formation of adventitious root initials.
Another related species, Portulaca smalii P. Wilson, is very
adaptable to dry, unfavorable conditions (54). Members of the
Portulaca family vary greatly in chromosome sizes and ploidy, a
characteristic of an unstable, developing group of plants (159).
This condition might also explain the great variability in size
observable among the common purslane plants.

4
In the field, coiranon purslane seedlings do not appear until the
weather is warm (130). In laboratory studies common purslane seeds
failed to germinate below 10°C but at temperatures between 25®C and
40®C germination was over 80% (167). Hocombe (83) found that light
increased the germination of purslane. Growth and development of the
weed were studied under various outdoor light intensities (167).
Decreasing the light intensity by 25% did not affect purslane
emergence heading or maturity, but 50% and 75% reduction was pro¬
gressively detrimental to purslane growth and development. Laboratory
tests (60) showed that cool white light is the best for purslane
growth followed by green, blue, red and yellow light. However with
high temperatures during the day and low during the night best growth
was observed under green light. These conditions are similar to
those in late summer under the shade of crop foliage. Field competition
studies of common purslane with table beets (Beta vulgaris L.), snap
beans (Phaseolus vulgaris L.) and corn (Zea mays L.) showed that
purslane is an especially strong competitor in beets and cultivation
must be employed during the first two weeks after the crop emergence
to be effective (167, 168).
A well developed purslane plant may produce up to 193,000 seeds
(172) which have a great germination potential. Although the surface
germinating purslane seeds are susceptible to various pre-emergence
herbicidal creatments (151), the weed is not readily killed by
common broadleaf herbicides applied after emergence. The herbicidal
damage to the shoots often results in abundant development of lower
branches (155) .

5
There is some conflicting evidence on the effectiveness of
herbicidal control of common purslane. It has been reported (123)
that common purslane is very sensitive to post-emergence 2,4-D
applications, while according to another report (162) this herbicide
does not control purslane economically. Jagschitz (95, 96) reported
on excellent control of 6 week old purslane by 1-1.5 Ib/A of 2,4-D
or as little as 2 oz/A of dicamba.
Anatomical studies of common purslane* reveal developed parenchymatic
water storage tissue in the stem. The epidermis of leaves has stomata
evenly distributed on both the upper and lower surface with bulliform
cells located just beneath the lower epidermis. These large bulliform
cells are motor cells (64) and probably participate in leaf movement.
Dicamba as a Systemic Herbicide
A. Characteristics of Dicamba
2-methoxy -3,6 - dichlorobenzoic acid was first introduced in
field trials in 1958-59 by Velsicol Chemical Corporation under the
name "Velsicol 58-CS-ll." In 1963 this compound was recommended as
a herbicide by the Weed Society of America and given the common name
"dicamba."
Dicamba is readily soluble in ethanol but only slightly soluble
in water. However the dimethylamine derivative of dicamba is readily
soluble in water (88).
It is recommended especially for the control of phenoxy-tolerant
*Chang, T, C,, and G. B. Goddard, Univ. of Mass., personal communication.

6
annual and perennial broadleaved weeds and brush species in small grains,
corn, flax, pastures, certain vegetables and some orchards crops.
Both preemergence and postemergence applications are effective. As
little as 0.05 to 0.25 pounds per acre of dicamba has been found to
control broadleaved annuals economically.
B. Dissipation of Dicamba in Soil
An ideal herbicide effectively destroys weeds and then dissipates
to prevent an accumulation in the soil or harvested crop. Studies of
the detoxification of dicamba in the soil indicated that the dissipation
of the herbicide is greater in fine textured soils such as silty clay
loam soils, than in coarse sandy loam soils, and faster in the topsoil
than in the subsoil (76). The breakdown increased with increasing
moisture content and temperature (43) . The loss due to volatilization
was negligible, and photodecomposition was not a factor. Friesen (67)
reported that oxygen consumption in soil was greatly reduced by 10,000
ppm dicamba due to the elimination of microorganisms and he asserted
that dicamba could persist in the root zone for several years. However,
the adsorption of dicamba to soil particles or colloids was very
limited (67) with the exception of kaolinite clays (43).
The recent survey of Nebraska soils with field bioassays for
persistance of herbicides (44) indicated that dicamba was relatively
non-persistant. Beans could be grown successfully on most of the
locations two years after the application of 20 Ib/A of dicamba, and
after three years the pretreatment did not show any effect.
C. Anatomical and Morphological Responses of Plants to Dicamba
There are many reports about phytotoxicity of dicamba and the

7
nature of Injury it ellicits. Treated soybean (171) (Glycine max (L.)
Merr.) plants exhibited stunted growth, malformed leaves, which were
characteristically cupped and crinkled, as well as petiole and stem
curvature. Often there appeared branching from cotyledonary nodes.
Dicamba was especially injurious to soybeans at bloom stage. Flax
(Linum usitatissimum L.) (131) was susceptible when less than 2 inches
tall and during flowering. Killing of the terminal bud and subsequent
branching at the cotyledonary node or leaf axis were the typical
responses.
Hurtt e^ (90) found that the minimum dose of dicamba for
biological response of beans was less than .02 pg per plant. Dicamba
caused proliferate growth, stem swelling and a decrease in anthocyanin
content (146). In roots of pea (Pisum sativum L.) seedlings (154)
there was an inhibition of root elongation, radial enlargement and
lateral root proliferation.
Application of 4 oz/A before fourth leaf stage caused malformations
of wheat (Tricitum vulgare L.) and barley (Hordeum vulgare L.) st .ns
and leaves (68) and interferred with their further development. Root
tips grown in dicamba solutions (1-100 ppm) showed a reduction in
the number of dividing cells. Above the concentration of 10 ppm
there was some evidence of chromosome clumping and multinucleate
cells being formed.
In alligatorweed (Alternanthera philoxeroides (Mart.) Griseb.)
(132) destruction of phloem, cambium and associated parenchyma occurred
above and within the nodes of plants treated with 2 Ib/A of dicamba.
Four days after treatment, dissolution of the cell walls in the same

8
tissues occurred with the formation of apparent multinucleate
coagulated protoplast. Growth of root initials was induced by dicamba
in the lower portion of the nodes.
In Canada thistle (Cirsium arvense (L.) Scop.) (48) visible stem
bending appeared 10 hours after application of .01 to 1 mg of dicamba
per plant. Necrotic spots on the leaves developed in a few days.
Later on swelling of stems and deformative effects occurred or the
death of growing tips was observed.
Differences in susceptibility to dicamba treatments were in¬
vestigated by Chang (50). Tartary buckwheat (Fagopyrum tataricum (L.)
Gaertn.) was severely injured by 0.125 oz/A of dicamba, wild mustard
(Sinapis arvensis L.) by 1-4 oz/A. These rates did not harm wheat or
barley, which could survive even 64 oz/A.
The response of wild buckv;heat (Polygonum convolvulus L.) to
dicamba (23) depended on certain environmental factors. High temp¬
erature and relative humidity, especially after treatment, induced
maximum response to the foliar application of the herbicide.
The plant response to herbicide treatment is dependent on the
rate of absorption, translocation and detoxification in the treated
species.
D. Absorption of Dicamba in Plants
The absorption of a herbicide depends on the properties of the
epidermis uf the treated plant organs. various plant species exhibit
substantially different rates of herbicide penetration. For example,
the absorption of foliarly applied dicamba was much faster in Tartary
buckwheat and wild mustard than in wheat and barley (50). The studies

9
with Canada thistle (48) have shown that dicamba is readily absorbed
by both roots and shoots. In the case of quackgrass (Agropyron repens
(L.) Beauv.) and bromegrass (Bromus tectorum L.) the uptake by roots
was more effective than foliar uptake (165).
When oat (Avena sativa L.) or barley coleoptile sections were
treated with radioactive dicamba, the entry was much faster through
the cut surfaces (166). Terminal portions contained more herbicide
than the central ones. Temperature strongly affected the uptake of
dicamba, doubling it in the range of 10® to 25®C.
Quimby (138) followed the uptake of dicamba in leaf sections of
wheat and wild buckwheat. Pretreatment with dicamba facilitated the
later absorption of the labeled herbicide as if increasing the
number of binding sites. He concluded that the dicamba anions were
being adsorbed on basic proteins. The process was temperature
dependent and heat denaturation appeared to increase the number of
binding sites. Washing with unlabeled dicamba did not remove the
radioactivity from the leaf sections. Pretreatment with actinomycinD
and trypsin stimulated dicamba uptake apparently increasing the
number of binding sites. Dicamba uptake reached a steady state in
30 min after the application. Freshly cut wild buckv/heat leaf sections
absorbed significantly more dicamba than did wheat sections.
Also the pretreatment of detached bean leaves with 1000 ppm of
the dimethylamine derivative of dicamba facilitated the entry of
radioactive dicamba into the leaf (106).
E. Translocation of Dicamba in Plants
After entering the plants the transport of dicamba proceeds in the

10
symplast (protoplasm) and the apoplast (xylem, cell walls).
In the detached bean leaves basipetal symplastic transport was
prevalent, Dicamba accumulated in the lower part of the petiole and
leaked outside to the surrounding water being much more mobile than
phenoxyacids (106) .
In Quimby’s studies (138), radioautography showed that the
movement of label after foliar application was both symplastic and
apoplastic in wheat, only symplastic in wild buckwheat. The highest
concentration of label accumulated in the meristem of buckwheat and
the treated leaf tips of wheat.
Hull and Weisenberg (89) observed rapid symplastic transport of
dicamba when applied to the leaves of beans or Johnsongrass (Sorghum
halepense (L.) Pers.). However, the leakage from phloem caused
uniform distribution of label in all leaves above the site of
application. Some accumulation was noticed in the shoot meristerns
of beans. The basipetal movement of dicamba was markedly restricted.
When applied to the roots, dicamba moved slowly to the shoots with
little accumulation in the roots.
Dicamba is often excreted from the roots of treated plants and
may be reabsorbed by untreated ones growing nearby (48, 49). Studies
with aeration, nutrients and metabolic inhibitors (DMSO - dimethyl
sulfoxide) (65) suggested an active component in this root excretion
phenomena. According to Crafts and Yamaguchi (55) the ability of a
herbicide to leak from phloem to xylem is related to its exudation by
roots.
Comparing susceptible broadleaved weeds and resistant grasses

11
Chang (50) found that dicamba accumulated in meristematic tissues of
the first group and was evenly distributed in the second. Dicamba was
especially mobile in Tartary buckwheat. After both foliar and root
application it was translocated rapidly to meristematic tissues of the
shoot apex and in the leaf axils. Redistribution to young tissues
continued for at least 20 days with only a small amount of dicamba
immobilized in the dead marginal tissue. After root application the
pattern of distribution soon resembled that of foliar absorption but
with more herbicide remaining in the roots. Excretion from the roots
after foliar application reached 10% of the applied dose. He suggested
that dicamba was readily transported through both phloem and xylem.
In purple nutsedge (Cyperus rotundus L.) (Ill) dicamba was slowly
translocated by both s3nnplastic and apoplastic transport after leaf
application. The accumulation occurred mainly in the actively growing
plant parts. There was no accumulation in the underground parts,
although the translocation through rhizomes and tubers reached the
daughter plants. After root application the translocation was also
slow but more uniform and the accumulation was evident in the tips of
the leaves. The leaves of shade grown plants accumulated relatively
larger amounts of dicamba. Some herbicide was excreted from the
roots to the culture solution. The translocation occurred more
readily in vegetative than reproducing plants. Dicamba probably was
initially translocated in phloem following the foliar application,
but it migrated to the xylem at the stem and basal tuber connection.
Further distribution as well as that of root applied dicamba occurred
through the xylem.

12
In Canada thistle (48) the herbicide was translocated in both
phloem and xylem. After foliar application some dicamba was retained
in the treated leaf but the label moved ■’'eadily upward and downward
in the stem. The accumulation in the growing tip and young leaves
indicated a "source to sink" pattern of translocation in the assimilate
stream. But in one day some dicamba also entered mature leaves, which
may indicate some leakage from phloem to xylem and acropetal transport
in the transpiration stream. Canada thistle released about 0.5% of
applied dicamba through root exudation. Treatment of the roots with
dicamba resulted in rapid upward movement in the transpiration stream,
but the accumulation of the herbicide in young leaves indicated
redistribution via the phloem.
In studies with pea seedlings Scott (154) found that immediately
upon treatment the root tips accumulated dicamba above the external
concentration. However, the label moved out fast when the treatment
was removed.
Translocation of dicamba may be influenced by pretreatment with
other herbicides. Foliar application of 1000 ppm CEPA (2-chloro--
ethylphosphonic acid) a week before dicamba spot treatment on the
leaves of wild garlic (Allium vineale L.) (27) increased greatly basi-
petal translocation of dicamba. When the concentration of CEPA was
increased five times, the pretreatment caused a marked inhibition of
basipetal dicamba translocation, similar to that observed after pre¬
treatment with 100 ppm of 2,4-D. The action of ethylene on dicamba
transport may be suspected in these experiments, as CEPA is known to
release ethylene in alkaline solutions and in plants (174).

13
Besides the fast transport with assimilates in phloem and with
water in the xylem, there is a slow transport which occurs in all living
tissues. This different transport mechanism is conveniently invest¬
igated in segments of petioles or coleoptiles. There is evidence that
dicamba is transported in these tissues basipetally, similar to auxins
(47). The velocity of this transport in segments of Tartary buckwheat
and bean petioles and corn coleoptiles was calculated to be 0.8 mm/hr.
The transport was increased by the addition of sucrose and ATP (adenosine
triphosphate) and decreased by the addition of DNP (dinitrophenol) or
lowering the temperature. Active transport seemed to be responsible for
such movement. Acropetal movement was less affected by these factors,
suggesting a simple diffusion component. Light treatment promoted
the retention of dicamba by the tissue and reduced the transport.
The little investigated polar transport of dicamba in tissue segments
is of great importance as the first, slow and therefore rate limiting
step in the translocation of the herbicide in plants.
F. Metabolism of Dicamba in Plants
Detoxification mechanism in the treated plants is an Important
clue to their susceptibility to the applied herbicide. Dicamba is
relatively stable in the susceptible plants.
Scott (154) found only dicamba in the roots of treated pea seed¬
lings and the rate of decarboxylation was less than 0.3%. In beans
(89) only dicamba was recovered five days after treatment and there
was no indication that dicamba was metabolized in purple nutsedge
(111, 143).
In less susceptible barley and corn, the analysis of the excised

14
roots revealed 5-hydroxydicaiiiba as a major metabolite (143). 5-hydroxy-
dicamba is much less toxic than dicamba (47) and the process of
metabolism in this case must be considered as a detoxification
mechanism. Barley produced 3,6-dichlorogentisic acid and 3,6-dichloro-
salicylic acid as minor metabolites of exogenously added dicamba.
Hull reported (89) that two unidentified metabolites in Johnson-
grass accounted for about 50% degradation during five days. One of
them released dicamba upon acid hydrolysis. There was no noticeable
decarboxylation of the herbicide in this plant.
In Canada thistle (48) there appeared an additional unknown
product in chromatograms of the extract from treated leaves after
three days. Some decarboxylation, amounting to less than 10% of
the applied amount, was also recorded.
In studies comparing wheat and wild buckwheat, Quimby (138)
found that dicamba was conjugated or metabolized more rapidly and to
a greater extent in wheat main culms than in wild buckv7heat meristems.
The metabolites were suspected to be 3,6-dichlorosalicylic acid and
5-hydroxydicamba.
Tartary buckwheat (49) metabolized dicamba very slowly - with only
13.5Z being metabolized after forty days. Less than 1% underwent
decarboxylation. The major metabolite was 5-hydroxyd icamba present
as a glycoside. After acid hydrolysis of extracts, 3,6-dichlorosalicylic
acid was found but it was considered an artifact being a product of
dicamba hydrolysis. Wild mustard (50) also produced 5-hydroxyd icamba
and some dicamba conjugates at slightly higher rate of metabolism.
Resistant barley and wheat detoxified more than 85% of dicamba in the

15
course of twenty days forming the major metabolite 5-hydroxydicamba and
3,6-dichlorosalicylic acid as a minor one. The rates of dicamba detoxi¬
fication correlated with the resistance to dicamba for these plant
species.
The extent of metabolism was consistently greater in the treated
leaf than in the other plant parts. Studies with detached leaves of
Tartary buckwheat showed that the rate of metabolism was highest in
the treated leaf blade. However, the product, 5-hydroxydicamba, did
not move out of the treated leaf. The detoxification of dicamba is
closely related to immobilization, as the metabolites seem to be much
less mobile than dicamba itself.
Influence of Herbicides on Cell Membrane Permeability
According to Meinl and Gutenberg (120) plant growth hormones
increase water permeability of cell membranes. lAA in low concentrations
(10 ^ to 10 ^ M) reduced the time needed for deplasmolysis but high
_q _5 concentrations (10 ^ to 10 M) slowed down the process (79). Similarly
gibberellic acid at 10 yM caused maximum increase in water permeability
in onion (Allium cep a L.) epidermal cells, but 1 mM of GA decreased
the permeability (80).
Studies with betacyanin efflux (144) from red beet root tissues
"3 did not indicate any permeability change after treatments with 10 to
10“^ M picloram, 10"^ to 10"^ M dicamba, lO"*^ M 2,4-D and 10 ppm
ethylene. Significant pigment leakage occurred after the application
of 10“^ to 10”^ M phenylmercurie acetate, 10“^* M DNP, 10“^ M 2,4,5-T
and 10~^ M 2,4-D. Similar results were reported by Janes and Foy (97).

16
Within 24 hours of treatment with 10“^ to 10”^ M dicamba, paraquat
(l,l*-dijnethyl-4,4' bipyridinium ion) or atrazine (2-chloro-4-ethyl-
amino-6-isopropylamino-s-triazine) there was no increase in betacyanin
efflux. However after 48 hours, 10”^ M dicamba caused a significant
increase of pigment release (400%). The most pronounced effect on
efflux was that caused by DMSO (dimethylsulfoxide), especially when
added with 10”*^ M atrazine although atrazine itself did not facilitate
betacyanin release.
Magalhaes and Ashton (110) found a decrease in the permeability
of cell membranes of nutsedge as measured by electrolyte leakage five
days after treatment. This effect was proportional to the concentration
of dicamba within the range of 10“^ to 10“^ M.
Recently Bachelard and Ayling (17) found that 2,4-D application
(100 ppm) to pine (Pinus radiata Don.) needle segments caused severe
shrinkage of the protoplasts and loss of plasmalemma integrity as
measured by tissue resistance to electrical currents. Picloram (25 ppm)
and 2,4-D disrupted the chloroplast structure and the plasmalemma
integrity in stem tips of Eucalyptus viminalis Labill. The effects
were so rapid (4 to 8 hours) that the authors concluded the plasma-
lemma is a primary site of action of these herbicides.
Influence of Herbicides on Protein Metabolism
Herbicides may influence protein synthesis by decreasing the
amount of available substrates and energy for protein synthesis or
by interfering directly with the synthesis of nucleic acids or proteins.
Other possible effects include specific stimulation or inhibition of

17
certain enzymes.
There are some contradictions in the available reports about the
Influence of dicamba on the generation of metabolic energy. Oxygen
uptake by nutsedge leaves was inhibited by 40% five days after
spraying with 10“^ M dicamba solution (110). Isolated cucumber
(Cucmnis sativa L.) (66) mitochondria showed lower rates of succinate
utilization, consuming less oxygen, in the presence of increasing
dicamba concentrations. No such inhibition was seen when ^-keto-
glutarate was a substrate. It could indicate a suppression or
inactivation of succinate dehydrogenase by dicamba. However the ATP
content of dicamba treated soybean hypocotyls did not decrease
significantly (74).
Different herbicides have an ability to inhibit the increase in
proteolytic activity of the squash (Cucurbita maxima Duchesne)
cotyledons during germination (16) and to decrease the pool of
available amino acids for new protein synthesis. At 10”^ M, dicamba
inhibited proteolytic activity only slightly.
Low, auxinic levels of systemic herbicides usually enhance RNA
and protein synthesis, whereas at higher concentrations these processes
are-inhibited. The effects depend upon the sensitivity of the plants
and selectivity of a particular herbicide.
The application of 10“^ M 2,4-D to the elongating region of
soybean hypocotyl (85) caused marked swelling and inhibited growth
accompanied by a doubling in DNA, RNA and protein content. Chromatin
isolated from these auxin treated tissues showed 8 to 10 fold increase
in RNA synthetic capacity as measured in vitro (86). However, in apical

18
tissues, the nucleic acid accumulation and chromatin activity was de¬
creased by 2,4-D. The RNA synthesized by chromatin isolated from
treated tissue was different from that of control. The herbicide
seemed to facilitate RNA production on the template portions normally
left untranslated.
Benzoic acid derivatives and auxins were found to increase per¬
oxidase activity - the single isoperoxidase present in the exocellular
fraction in Xanthi tobacco (Nicotiana tabacum L.) (121).
Many herbicides were found to depress leucine-l-C^^ incorporation
(112) in barley and Sesbania exaltata (Raf.) Cory seedlings in con¬
centrations of 2 and 5 ppm. Amiben (3-amino-2.5-dichlorobenzoic acid)
was ineffective but it stopped the development of the seedlings. In
another experiment (124) dicamba at 6 x 10“^ M was found to inhibit
leucine incorporation by soybean hypocotyl sections but without
influence on ATP and orotic acid incorporation into RNA. Gibberellin
induction of °=-amylase in barley aleurone layers was very severely
reduced by dicamba. As the herbicide does not affect the activity of
oc-amylase i^ vitro, it was concluded that it interferred with the
formation of this enzyme.
It was reported that many broadleaved plants accumulated nitrates
after 2,4-D application, but in grasses the level of nitrates de¬
creased. Changes in nitrate reductase activity were found responsible
for this phenomenon. It decreased in cucumber four hours after 2,4-D
application (2.25 x 10”^ to 2.25 x 10”^ M) , but increased in corn
when treatexi with 4.5 x lO”^ to 4.5 x 10”^ M 2,4-D.
Another example of selectivity in the action of herbicides upon

19
protein metabolism in different plants was given recently by Arnold
and Nalewaja (15). They stated that dicamba increased RNA and protein
content in wild buckwheat and in wheat at the boot stage. However
examination of their data reveals that the increase in protein content
occurred only in the stems of young buckwheat plants, the protein in
the leaves and meristerns actually decreased sharply. In older buck¬
wheat plants, which were unaffected by the applied dicamba (140 g
dimethylamine salt/ha), the protein content increased after the treat¬
ment in leaves, stems and roots, but decreased in the meris terns.
Young wheat did not show any changes in protein content after dicamba
treatment. At the boot stage the content of protein increased in
wheat stems, but decreased in the treated leaves. The authors
associated these changes in RNA and protein content with the suscept¬
ibility of the plants to dicamba. Wild buckwheat is more susceptible
and the accumulation of dicamba in its meristematic tissues disrupted
normal growth and protein accumulation. Protein accumulation in stems
could be explained by the fact that stems were more meristematic after
dicamba treatment, producing callus and abnormal radial growth.
Working with calf th3mius DNA and histone, yeast RNA and serum albumin,
the same authors reported an increase in precipitation of nucleohistone
with increasing dicamba concentrations. More histone remained in the
supernatant, which would suggest that dicamba removed some histone
from DNA. This might explain the increased synthesis of RNA and
subsequently that of protein. Serum albumin was found capable of
binding dicamba and 2,4-D through arginine, suggesting that the
herbicides could directly influence protein conformation.

20
Role of Ethylene in Plant Growth
A. Anatomical and Morphological Responses of Plants to Ethylene
Ethylene, a simple olefin (C2H^), is known to influence a
variety of responses in plants. Among these are promotion of germ¬
ination, stem swelling, epinasty and abscission of leaves and fruits,
chlorosis, senescence, flower Induction, flower fading and fruit
ripening. It also affects tropistic responses and nastic movements,
inhibiting hypocotyl hook opening, geotropism, stem curvature and
night leaf closing (137).
The scope of this work limits this review to only a few of the
above mentioned responses.
Epinasty appears to be a specific effect of ethylene action (30)
as it cannot be prevented by auxins. High auxin concentrations can
induce epinasty by stimulation of ethylene formation. The epinastic
response is very sensitive to ethylene with the threshold being only
.025 to .05 ppm with maximum response at 1 ppm.
Abscission is induced by higher levels of ethylene than necessary
for epinastic response (2-10 ppm) and may be prevented by auxin appli¬
cation. Burg and Burg (38) are of the opinion that ethylene causes
abscission by influencing auxin metabolism and/or transport. According
to those authors abscission occurs when the level of auxin in a distal
part is much lower than in the proximal portion of a particular organ
and this gradient is caused by ethylene.
Another hypothesis (4), advanced by Abeles, maintains that
ethylene stimulates abscission through its influence upon RNA and

21
protein synthesis in the separation zone. This stimulation of RNA
and protein synthesis may occur only in the senescent tissues, which
are responsi''^e to ethylene. Increased protein synthesis is limited
to the abscission zone and involves stimulation of cell wall degrading
enzymes.
For some time Abeles tried to show that senescence and abscission
were two independent processes occurring at the same time, and that
ethylene hastened only abscission (9). Senescence, but not abscission,
may be delayed by lAA or cytokinin. Recently (5) the same author
described ethylene as a phytogerontological agent and found that it
does accelerate aging in concentrations of .3 to 10 ppm and plays a
dual role in abscission, accelerating senescence and specifically
stimulating cellulase activity.
Stem swelling and inhibition of elongation usually occur - in the
same elongating part of the stem under the influence of ethylene and
are due to the isodiametric expansion of cells in the cortical region.
Holm and Abeles (85) found that ethylene (1 to 10 ppm) reduced soybean
hypocotyl growth and induced its swelling together with a doubling of
DNA, RNA and protein content. However Burg and Burg (39) found that
the similar effects of ethylene on pea stem sections were independent
of protein synthesis. Addition of cycloheximide or actinomycin D did
not influence swelling or the inhibition of elongation. lAA was
effective in restoring elongation but it did not reduce the swelling
which is apparently another specific response to ethylene. Ethylene
inhibition of elongation may be also counteracted by GA (87).
In roots (139) ethylene also inhibits elongation, lateral root

22
formation and cambial activity. Apelbaum and Burg (14) found that
ethylene stops almost all cell division in etiolated pea hooks, retards
division in the root tips and buds (both apical and lateral on isolated
stem sections). DNA synthesis in apices was strongly inhibited. The
inhibition of bud growth on pea seedling nodes could be reversed by
kinetin (40).
Tropistic stem curvatures and nastic movements are often adversely
affected by ethylene. The gas is completely effective in preventing
spontaneous curvature development in straight growth test of pea stem
sections (36, 39), even in the presence of actinomycin D or cycloheximide.
At very low concentrations, which do not affect growth of stems and
roots, ethylene was capable of preventing the geotropic response of pea
root (46). The natural night leaf-closure of mesquite [Prosopis
juliflora (Swartz) DC. var. glandulosa (Torr.) Cockerell] and huisache
(Acacia farnesiana (L.) Willd.) is also inhibited by ethylene treatment
(20, 126).
B. Physiological Actions of Ethylene in Plants
The most intriguing characteristic of ethylene responses is the
observation that the concentration range is similar for them all (30).
The threshold of activity is usually as low as 0.01 ppm, and the
maximal physiological response often occurs at about 1 ppm. Only
defoliation and fruit ripening require higher levels (1 to 10 ppm).
The uniform concentration of ethylene required to start multiple
effects, and the fact that CO2 is a competitive inhibitor of ethylene
action in most of the described responses, indicate that a single
receptor may catalyze a single initial event. This primary target

could be:
23
an enhancement of cell membranes permeability,
changes in RNA and protein synthesis,
interactions with auxin (lAA),
effects on plant cell walls.
This last site of action is suggested by ethylene caused expansion
of cells in a radial direction instead of elongation and stimulation
of root hair development (45). Changes in microfibril deposition
could be responsible for these phenomena.
The influence of ethylene on cell membrane permeability is sug¬
gested by its effects on nastic leaf movements. Sacher (149, 150)
found that permeability of banana (Musa sapientum L.) fruit tissue
increased two days before the onset of the climacteric and continued
to increase during the respiratory rise. A significant increase in
ethylene production was noted just before the climacteric. It would
appear that enhanced ethylene production resulted from changes in
permeability. However, application of ethylene brought about increased
permeability of banana fruit peel, but not that of avocado (Persea
gratissima Gaertn. F.) fruit, bean endocarp or leaves of Rhoeo discolor
Hance. The rate of ion leakage of banana tissue during ripening was
examined also by Baur and Workman (21) and found to increase at the
onset of the clim.acteric. The major leaking cation was potassium.
Changes of water flux in cantaloupe (Cucumis m.elo L.) fruit
tissue (12) after ethylene application indicated a change in membrane
permeability. After an initial phase of water influx, an efflux
occurred.

24
A different approach was pursued by Burg e_t al^. (41) who found
sugar leakage proportional only to sugar content in potato (Solanum
tuberosum L.) tubers, apples (Pyrus malus L.)> bananas, pears (Pyrus
communis L.) and pea stems, and not influenced by auxin or ethylene.
Gibberellin induced “-amylase activity of barley aleurone layers
seemed to increase after ethylene treatment, Janes (98) reported
that ethylene did not affect the enzyme production, but its secretion
from aleurone cells. This might indicate an alteration of cell
membrane structure by ethylene.
Stimulation of swelling of mitochondria by ethylene is not an
ethylene specific response. Mitochondria were found to swell also
under the influence of similar rates of other aliphatic gases (101,
119). This does not exclude a possibility of cellular membranes as
a primary site of action of ethylene. Equilibrium dialysis experiments
to measure the binding of ethylene to various subcellular fractions
from tomato (Lycopersicum esculentum Mill.) fruits and bean petioles
(102) indicated that more ethylene was bound in responsive tissues,
and CO2 or deoxycholate depressed the binding.
The influence of ethylene on protein metabolism has been studied
with various and often contradictory results. The hormonal role of
ethylene in abscission seems to be expressed partially in a stimulation
of protein synthesis in the abscission zone (4, 7, 84). RNA synthesis
is also stimulated in this area. The increase in cell wall degrading
enzymes, such as cellulase, leads to the abscission. Ratner ^ al.
(142) found that the cellulase activity was markedly increased before
abscission in the abscission zone of orange (Citrus siniensis (L.)

25
Osbeck) leaf explants. Ten ppm of ethylene increased cellulase
activity after a 6 hour lag period. Cycloheximide applied at different
times after ethylene fumigation indicated that synthesis of enzyme had
to be involved. There was no enhancement of protein synthesis in the
other parts of the leaves and petioles.
As ethylene production increased during the ripening of banana
fruit tissue (1A9), a decline in amino acid incorporation into protein
was noted. In another study (150) externally applied ethylene (7 and
45 ppm) had no effect on RNA and protein synthesis in avocado and
banana fruits, but it inhibited these processes in bean endocarp and
Rhoeo discolor leaves.
Marei and Romani (113) found that levels of ethylene which pro¬
moted growth and ripening of fig (Ficus carica L.) fruits, increased
RNA and protein synthesis in the fruit slices. The sedimentation
profiles of the induced RNA and ribosomes v/ere not different from the
untreated tissues.
In soybean seedlings treated with 1 ppm ethylene (86) there was a
small increase in RNA content in the elongating section of the hypocotyl
and the isolated chromatin showed some increase in capacity for RNA
synthesis. However, the accumulation of nucleic acids and chromatin
activity in the apical tissues was inhibited by ethylene. Chromatin
isolated from ethylene treated hypocotyl tissue produced RNA different
from that of auxin treated or control hypocotyls. Actually ethylene
inhibited auxin enhancement of nucleic acid synthesis, which itself
was much greater than the effect of ethylene in the hypocotyl region.
Wliile certain enz5mies are definitely stimulated by ethylene

26
application, the induction of invertase formation in sugar beet tissue
by GA was severely reduced by 10 ppm of ethylene (153). After exposure
to 8 ppm ethylene sweet potato (Ipomoea batatas Poir) tuber tissue
developed higher peroxidase and polyphenol oxidase activity (157).
Similar results were reported by Imaseki (92) who noticed that 1 ppm
of ethylene doubled the peroxidase activity and 10 ppm increased it
four times in sweet potato tubers. This increase could be prevented
by 5% CO2. In abscission zones of tobacco flower pedicels an increased
peroxidase and catalase activity was noted 5 hours after ethylene
exposure (81). In fruits of mango (Mangifera indica L.) (114) the
catalase and peroxidase activities were found to increase considerably
during the climacteric. This rise, however, was ascribed to dis¬
appearance of a heat-labile and nondialyzable inhibitor of these
enzymes. Ethylene fumigation (10 and 50 ppm) caused a remarkable
increase in catalase and peroxidase activity in slices of unripe
mango fruits, connected with the disappearance of the inhibitor.
Phenylalanine ammonia - lyase (PAL) activity often develops in
wounded tissues together with an increase in ethylene production.
Flavedo disks of grapefruit (Citrus paradisi Macf.) peel are an example
of this wounding response (145) . Exogenous ethylene stimulated PAL
activity in flavedo of intact fruits, and the enhancement could be
prevented by CO2 (20%) or cycloheximide application. Cycloheximide
also limited ethylene production by flavedo disks. Protein synthesis
appeared necessary both for ethylene evolution and ethylene promotion
of PAL activity. Maximum PAL stimulation was achieved at 100 ppm
ethylene, whereas half maximum was induced at 15 ppm. In segments

27
excised from the epicotyl of pea seedlings C91) marked increase in
PAL activity occurred in ethylene treated tissues after a 6 hour in¬
duction period. Maximal effect was caused by 10 ppm ethylene. Appli¬
cation of cycloheximide and actinomycin D at various stages of PAL
induction suggested de novo synthesis of protein. However, the rel¬
atively high levels of ethylene needed to stimulate PAL activity tend
to exclude it as a primary response to ethylene action.
C. The Promotion of Ethylene Production in Plants
A recognition of ethylene as a natural plant hormone controlling
plant growth and development came about as evidence accumulated of
ethylene production by various plant organs in various species. An
extensive survey of ethylene producing plants and their organs with
references was compiled by Burg (29). All plant parts produce
ethylene in various quantities: germinating seeds (10, 63, 100, 160),
young seedling hooks (40, 71), leaves and flowers (42, 78), roots (139)
and especially ripening fruits (32, 61, 108, 136).
Ethylene biosynthesis is not well understood, but the gas seems
to be produced by soluble cytoplasmic enz3mies rather than mitochondria
(103) . The only well established ethylene precursor is methionine,
more specifically - its third and fourth carbon atoms (18, 19, 107,
169) . The methionine conversion to ethylene seems to be catalyzed by
a copper requiring enzyme (107) and an involvement of a peroxidase
cannot be excluded (169).
Many physical stimuli and chemicals are effective in increasing
ethylene production. There are reports of ethylene increase after
irradiation with gamma rays (116, 117), applying chilling tem-peratures

28
(53), or various light treatments (8, 10, 71). In fact it could be
stated that most stress conditions appear to stimulate ethylene pro¬
duction.
Increased ethylene evolution is a typical wounding reaction.
Ethylene production by cantaloupe fruit slices was at least ten times
greater than that by intact fruit (109). Nonwounding mechanical stress
as that of pea epicotyls growing between glass beads, stimulated
ethylene production to a level high enough to reduce elongation and
increase the diameter of the stems (72).
Natural growth hormones as lAA, GA, kinetins and abscissic acid
were all found to influence ethylene production. Often their action
is indistinguishable from that mediated by ethylene. The interaction
with lAA is sometimes considered as a primary site of ethylene action.
Stanley and Ellen Burg carried on extensive investigations of this
relationship and found that ethylene production in pea stem sections
was initiated at lAA concentrations exceeding 10 ° M, which is the most
favorable for elongation (33). The elevation of ethylene evolution
depended upon the amount of auxin applied, with maximum stimulation
lasting 3 days (40). The ethylene produced caused reduction of growth,
swelling of stem and lateral buds inhibition with maximal response at
1 ppm concentration. Kinetin could reverse the inhibition of bud
growth although it too caused ethylene to increase. The authors felt
that apical dominance operated through lAA suppressing bud development
by stimulating ethylene formation in the nodes. High concentrations
of auxin induced ethylene production in excised root tips (46). The
ethylene produced halted their growth and inhibited root geotropism.

29
Abeles (1) found that the stimulation of ethylene evolution from
the seedlings of beans, sunflower (Helianthus annuus L.) and corn
occurred tvro hours after auxin application and could be impaired by
actinomycin D and puromycin. This work suggested an involvement of
the synthesis of specific enzymes required in ethylene biogenesis.
The interaction of ethylene with auxin is complicated by the fact
that ethylene seems to influence auxin transport. This might occur
by increased conjugation with aspartic acid and/or glucose (25, 99)
and result in immobilization (24, 25). In the presence of ethylene
the lateral transport of auxin in pea stem was reported to be almost
completely abolished (33, 35, 37, 38) which would explain the dis¬
appearance of tropistic responses. Burg and Burg (35) found that
polar transport of auxin was not influenced under these conditions,
except by long ethylene pretreatment (38). Other studies (24, 127)
indicated that in cotton (Gossypium hirsutum L.) and cowpea (Vigna
sinensis (L.) Endl.) polar auxin transport was inhibited by ethylene
action.
Different results were presented by Abeles (2) who did not find
any influence of ethylene on auxin transport in various plant tissues.
Fuchs and Lieberman (69) investigated effects of kinetin, lAA and
GA on ethylene production in pea seedlings. The kinetin stimulation
was limited to the third and fourth day of growth, but lAA stimulation
continued with age and could be increased by kinetin. The kinetin
induced system of ethylene production was much more sensitive to
cyclohexiraide than that of untreated plants. Kinetin (benzyladenine)
at 5 X 10“^ M promoted ethylene production in radish roots (139).

30
In agricultural practice (52) GA and abscissic acid are used
selectively to induce ethylene production in leaves, resulting in
abscission, without any effect on fruits. These substances stimulated
ethylene production and abscission in bean explants (6, 147, 148). The
effect of abscissic acid on beans and cotton was investigated in terms
of ethylene production and abscission break-force (56). At a concen¬
tration of 5 X 10*"^ M, ABA tripled ethylene production in cotton and
abscission was strongly accelerated.
Malformin B, v/hich consists of two closely related cyclic penta-
peptides produced by Aspergillus niger, causes responses in beans
similar to ethylene, such as epinasty, growth disturbances, stem
swelling and abscission in susceptible varieties. Curtis (57) found
that 2 X 10"*^ ymol per plant stimulated ethylene production within four
hours and the stimulation lasted up to eight days.
The effects of coumarin (129) on ethylene production are various
in different plants. It inhibited the opening of bean hypocotyl hook
by inducing ethylene production within 4 to 6 hours. But coumarin
also inhibited germination of lettuce (Lactuca sativa L.) seeds and
prevented the rise in ethylene production parallel to germination.
Among synthetic chemicals - the artificial auxins and some herb¬
icides proved to be ethylene stimulators.
NAA (1-naphtalene acetic acid) is widely used in agricultural
practice to induce defoliation and pineapple (Ananas sativus L.)
flowering (34) due to the increased ethylene production of treated
plants. The action of NAA is not however, limited to ethylene stim¬
ulation, as its auxinic properties can delay abscission (52) when

31
applied at certain concentrations. Beyer and Morgan (25) found that
NAA is metabolized less than lAA in plant tissues. In studies by
Abeles and Rubinstein (11, 148) NAA at 5 x 10”^ M stimulated ethylene
production from roots, stems and leaves of nine plant species (corn,
beans, tomato, Zebrina pendula Schnizl., Coleus blumei Benth., peas,
tobacco, Coffea arabica L.). Immature tomatoes also produced more
ethylene after NAA application, but in mature fruits the auxin inhibited
ethylene evolution.
2,4-D, a systemic herbicide, is even more effective in ethylene
induction than auxin. The increased ethylene production can be main¬
tained longer and at a higher level than with lAA (99), because of the
relative stability of the herbicide in plants. Abeles (1) found that
the action of 2,4-D on ethylene production is connected with protein
production, because it can be inhibited by actinomycin D and puromycin.
The ethylene rise was noticeable after two hours, and protein synthesis
after about three hours. In soybean hypocotyl (85) the action of 1 ppm
ethylene could be mimicked by 10”^ M 2,4-D, and 10 ppm ethylene - by 10“^
M 2,4-D. GA partially inhibited the ethylene rise caused by 2,4-D (87).
In another study (3) Abeles compared the action of 2,4-D on corn and
soybeans. Both plants exhibited ethylene evolution with 10”^ M 2,4-D,
many times greater than their normal production. Corn produced much
less ethylene than soybeans but seemed to be more sensitive in its
growth stunting response. Euonymus japonica L. leaves treated with
the butyl ester of 2,4-D underwent premature senescence and defoliation
(77), with their ethylene production increased 5 to 25 times.
Another phenoxyacetic acid, 2,4,5-T at a concentration of 20 ppm

32
(13-5) stimulated ethylene production in fruits and leaves of the fig
tree to levels responsible for fruit ripening, epinasty and senescence
of leaves. The stimulation was of long duration, with the peak on the
sixth day in the leaves and on the tenth day in the fruits.
Picloram (20, 126) applied to the roots of honey mesquite or
huisache inhibited leaf movement and induced epinasty. Subsequently
the leaves of huisache abscised. The responses were explained by
increased ethylene production in the treated plants and the internal
concentration of ethylene was correlated with the level of picloram in
mesquite stems.
The chemicals considered above probably stimulated ethylene pro¬
duction by influencing the ethylene synthesis system, be it de novo
synthesis of the enzymes or activation of existing enzymes. There are
no reports as to the specific action of these substances due partly to the
lack of information about the pathway of ethylene biogenesis.
Ethylene releasing compounds are available, whose mode of action
is well explained. CEPA (2-chloroethylphosphonic acid), its ester
derivatives and its anhydride cause many of the effects of ethylene
(170) i.e. ripening of bananas, tomatoes, flowering of pineapple;
epinasty and abscission. Loss of leaf movement and defoliation was
observed in mesquite and huisache (128), as well as growth of basal
buds and increase in the number of branches and leaves per node. In
cotton the youngest and the oldest leaves abscised after Ethrel (2-
chloroethylphosphonic acid and its ethyl ester) application leaving
only the fully expanded leaves (125).
In all these cases an increased ethylene evolution was observed due

33
to degradation of the applied chemical to ethylene. This occurred
nonenzymatically above pH 5.0 (170, 174), Also ethyl-propyl phos-
phonate was observed to generate ethylene in presence of reduced iron
and copper with an oxygen requirement. This chemical is a growth
retardant of pea seedlings (58).
D. Estimation of Internal Level of Ethylene in Plants
The problem of measuring the internal level of ethylene in plants
is a very important issue, because there, in plant tissues, ethylene
affects plant functions.
The rate of ethylene evolution is a helpful factor from which one
can deduct the internal ethylene level by comparing the responses
produced by the externally applied gas. Burg and Burg (33, 34), while
working with pea stems, compared auxin induced ethylene synthesis and
ethylene fumigation responses. They determined that the tissues should
contain 1 ppm ethylene while producing the gas at the rate of 5 yl per
kg fresh weight per hour. They also found that 1 ppm ethylene causes
maximal responses in preventing curvature and in growth inhibition.
The fruits are the best material for estimating the internal
ethylene content because of the presence of large air spaces enclosed
in the fruit tissues. Burg and Burg (31) found that when apples and
avocados produced 1 yl of ethylene per kg per hour the air in the
internal spaces contained 2 ppm of the gas.
Baur and Morgan (20) used this conversion factor in their estima¬
tion of the internal level of ethylene in mesquite and huisache after
picloram application. They divided the Burgs' factor of 2 ppm per yl
ethylene produced per kg per hour by surface to volume ratio for the

34
investigated plant parts, assuming that fruits have approximately
1 cm^/cm^. Multiplying then their reduced conversion factor by the
observed rate of ethylene production they estimated the internal level
of ethylene in plant tissues. This calculation did not account for any
difference in the epidermal resistance between fruits and other plant
organs. Similar calculations for pea epicotyl tissues resulted in a
factor of .13 ppm/yl per kg per hour (72), confirmed by the responses
to the externally applied ethylene.
Recently a method for direct determination of ethylene concentration
in the air spaces of plant tissues was developed by Beyer and Morgan
(26) , They extracted the gases from tissues by the use of vacuum and
then sampled the extracted air with a syringe whose content was injected
directly into a gas chromatograph. They applied their method to
various fruits, cotton, beans. Coleus blumei and more recently to
mesquite and huisache plants treated with picloram (126). The results
were similar to the previously estimated ones (20). The validity of
the new method was checked by ethylene fumigation experiments. The
authors predicted that the level of ethylene in fumigated plants
would be that of control plus external concentration. The predicted
internal level agreed with that determined by the new method within
the range 77 to 128%.

35
MATERIALS AND METHODS
Plant Material
All experiments were conducted with common purslane plants,
Portulaca pleracea L., using seeds from a uniform seed lot with
germination in the range of 80 to 90%.
Cultural Procedure
The purslane seeds were sown on sand in 4" plastic pots. The
pots were placed in trays filled with Hoagland's nutrient solution
(82) and allowed to absorb the medium through openings in the bottoms
of the pots protected only by cheesecloth. Hoagland’s solution was
added to the trays only after the previous addition had been completely
taken up by the plants. Purslane plants do not thrive under conditions
of excess moisture.
The plants were grown in a growth room under a 14 hour light
period and 10 hour dark period at a continuous temperature of 25 + 1°C.
Light was provided by Sylvania F48T12-WW-VHO fluorescent tubes at an
intensity of 1100 ft-C at the plant level. Relative humidity was 60%
+ 5% as measured continuously by a hygroscope. At the age of two
weeks the plants were thinned to three plants per pot.
Effect of Dicamba on Growth and Development
At the age of 4 and 10 weeks, purslane plants were treated with
various amounts of dicamba dissolved in water. Foliar application
was performed by placing 20 yl droplets on five fully developed leaves

36
of each plant. Root application was accomplished by watering each pot
with 30 ml of dicamba solution. The pots with treated plants were
placed in separate Petri dishes to avoid contamination with dicamba
from other treatments. Treatments were run in triplicate.
The plant height or main stem length was measured periodically
and after 25 days the fresh weight of the shoots was recorded.
The morphological effects of foliarly applied dicamba on four
week old purslane plants was followed visually during the first 24
hours after application and once each day thereafter for 14 days.
Again, three pots were used per treatment.
Effect of Ethylene on Common Purslane
Four week old potted plants (3 plants per pot) were placed in
10-1 desiccators in Petri dishes filled with nutrient solution. Each
desiccator contained a 50 ml beaker filled with saturated KOH solution
and supplied with a filter paper wick to trap carbon dioxide. The
desiccator outlets were sealed with rubber vaccine caps. Ethylene
was injected into the desiccator with a gas tight syringe to obtain
the desired concentrations. The effect of ethylene was followed
visually during the first 24 hours after application and once each
day thereafter for 7 days.
Measurements of Cell Membrane Permeability
A. Efflux of Electrolytes from Stem Segments
Because dicamba applications caused rapid bending of common
purslane plants, it was of interest to measure changes in cell membrane

37
permeability in the regions of stem bending.
Four week old purslane plants were treated by foliar application
of 0.05, 0.5 or 5 yg of dicamba per plant. After six hours when the
bending response occurred in plants treated with 5 yg of dicamba, one
cm stem sections were excised from the bending region. Three segments
taken from plants in one pot constituted one sample and ten replications
were used for each treatment. The samples were weighed to 0.1 mg and
placed into 25 ml of deionized water in plastic containers at 25° + 2°C
for 24 hours. The efflux of electrolytes from the stem sections was
measured according to the method of Wilner (173). After the incubation
period the conductance of the medium was recorded using a Serfass
Conductivity Bridge Model RCM15. The tissue was then boiled at 15 atm
for 30 min and the conductance again recorded.
The efflux of electrolytes was calculated as follows:
Percentage efflux of electrolytes = ^hos after incubation ^ _
mhos after boiling
efflux of electrolytes during incubation
total amount of electrolytes X 100.
B. Rubidium Release by Excised Leaves
Both dicamba and ethylene caused epinasty and loss of night
movement of common purslane leaves. The permeability of leaf tissues
after dicamba or ethylene treatment was examined by incubating excised
leaves with rubidium and then following thr efflux of Rb"^ ions into
the incubation medium.
Four week old purslane plants were treated with 10 yg dicamba per
plant applied to the leaves or 1 ppm ethylene in desiccators. After 24

38
hours the leaves were excised, avoiding the ones to which dicamba
was applied. Duplicate samples of 5 g of leaves were immersed for
two hours into 50 ml of a 10“^ M RbCl solution. Following the in¬
cubation period the leaves were washed three times with 50 ml demin¬
eralized water and then immersed in another 50 ml portion of this
deionized medium. At various intervals of the incubation period (0,
1.5 hr, 3 hr, 4.5 hr, 6 hr) samples were removed, blotted and dried
at yO^C overnight. Subsequently the rubidium content of the leaves
in the samples was determined according to Maynard and Baker (118).
The dry samples were crushed, weighed to .1 mg, ashed with 5 ml of
concentrated HNO^ and a few drops of H2O2 on a hot plate. Potassium
interference was eliminated by adding KCl to standards and samples to
the final concentration of 375 ppm. The samples were made up to 25 ml
with demineralized water.
Rubidium content was measured by atomic absorption with a Perkin
Elmer Spectrophotometer Model 214. The percent of absorption at
780 mu was recorded and converted to absorbance according to tables
(134). The concentration of rubidium was determined by reference to
a standard curve (Fig. I, Appendix) obtained with RbCl solutions.
Protein Synthesis Studies
Two systems were used to determine effects of dicamba on protein
synthesis in common purslane leaves. These were the incorporation of
labeled phenylalanine, and the activity of nitrate reductase, which
is known to turnover rapidly in plant material (152) .
Four week old purslane plants were treated with 10 yg of dicamba

39
per plant in foliar application or fumigated with 1 ppm ethylene.
A. Nitrate Reductase Activity
Triplicate four-gram leaf samples were excised 6, 12, 24 and 48 hr
after dicamba treatment from both the treated and control plants. Pre¬
liminary experiments had shown the activity of the enzyme was the
highest at noon time and the lowest at midnight. Nitrate reductase
activity was assayed according to the procedure of Hageman (75).
The samples were ground for 1 min in a Virtis 45 homogenizer with
16 ml of buffer containing 10“^ M TRIS (tris hydroxymethyl aminomethane),
10“^ M cysteine hydrochloride 10“^ M EDTA (ethylenedinitrilotetraacetic
acid), adjusted to pH 7.5 with 2N HCl. The homogenized material was
filtered through cheesecloth, the extract centrifuged at 20,000 x g
for 10 min and the supernatant used for the assay. All steps were
carried out at 1-4°C. The assay reaction mixture contained the
following:
O.IM potassium phosphate buffer (pH 7)
O.IM KNO3
1.3 x 10”^ NADH
H2O deionized
enzyme extract (supernatant)
Blank Assay
1.0 ml 1.0 ml
0.2 " 0.2 "
— 0.5 "
0.6 " 0.1 "
0.2 " 0.2 "
The ass'^y reaction was run for 15 mir at room temperature (25®C)
and stopped by adding 1 ml of 1% sulfanilamide in 1.5 N HCl. The
formed nitrite was estimated by adding 1 ml 0.02% N-l-naphtylethylene-
diaminedihydrochloride in 0.2 N HCl. The intensity of the resulting

40
pink color was measured after 30 min by recording optical density at
540 my with a Beckman DU-2 spectrophotometer against the blank. The
concentration of nitrite was calculated by reference to a standard
curve (Fig. II, Appendix). The nitrate reductase activity was ex¬
pressed in -.ymoles of nitrite formed per mg of protein per hour.
The protein content of the supernatant was determined by the
biuret method (105). One ml of the supernatant was mixed with 4 ml
of biuret reagent, and the optical density was measured after 30 min
at room temperature at 550 my with a Beckman DU-2 spectrophotometer
against a water blank. The protein concentration was obtained by
reference to a standard curve (Fig. Ill, Appendix) established x^^ith
serum albumin.
B. Incorporation of Phenylalanine-C^^
Following the dicamba application triplicate samples of purslane
leaves were excised every day for 6 days. For comparison purposes
1 ppm of ethylene and 50 yg of cycloheximide per plant (protein
synthesis inhibitor) were applied and the leaf samples excised after
1 day.
A one gram leaf sample was placed in 0.5 ml of the incubation
medium with only the petioles immersed. The incubation medium
contained the following chemicals (per 0.5 ml):
.01 M potassium phosphate buffer (pH 6)
1% sucrose
10 yg streptomycin
12.5 ymoles phenylalanine spiked with phenylalanine-C-14 (20,000 cpm per 0.5 ml) of specific activity 224 mC/mM.

41
The samples were placed in an incubator at a constant temperature
of 25°C and a light intensity of 500 ft-c for two hours. After this
period the leaves were washed three times T--ith 50 ml cold phenylalanine
solution of the same concentration. Subsequently the leaves were
ground in a prechilled mortar with a minimal amount of 0.01 M HgCl2
to arrest the incorporation. The homogenized material was made up to
9 ml with 0.01 M HgCl2. One ml aliquots of the homogenate were dried
on planchets over a hot plate and counted in a Nuclear Chicago gas
flow counter. The counts obtained were corrected for background and
used for the calculation of total phenylalanine uptake.
To determine phenylalanine incorporation, protein was extracted
from the crude homogenate as follows:
1) The protein was precipitated in the remaining 8 ml sample by
addition of 2 ml 50% TCA (trichloroacetic acid) to obtain a
final concentration of 10% TCA.
2) The precipitate was extracted three times with 5 ml of IN
NaOH at 100®C for 15 min, to dissolve the protein.
3) Combined protein extracts (15 ml) were precipitated again
with 50% TCA to a final concentration of 10% TCA and centri¬
fuged at 20,000 X g.
4) The precipitated protein was resuspended in 4 ml of 10% TCA
and then centrifuged in an International Clinical Centrifuge
for 10 min.
5) The protein was washed four times with 5% TCA (73). The
second washing included a 15 min incubation at 95®C to
remove nucleic acids.

42
6) The protein was washed with 10 ml of 95% ethanol.
7) The protein was washed three times with 8 ml of an absolute
ethanol: ether (3:1) mixture. All three washings included
3 min incubation at 60®C to remove fatty substances.
8) The purified protein was finally washed with 10 ml ether and
resuspended in a minimum amount of petroleum ether.
9) The suspension was dried on planchets, weighed to 0.1 mg and
counted as before.
The incorporation of labeled phenylalanine was expressed as
specific activity (cpm per mg protein).
Ethylene Evolution Measurements
To determine the effect of dicamba on ethylene evolution by purs¬
lane, four week old plants were enclosed in 10 1 desiccator jars with
CO2 traps as previously described. Various amounts of dicamba were
applied to the roots or foliage and the ethylene evolution was measured
by gas chromatography (108).
Five ml samples of air were periodically withdrawn with a gas
tight syringe and injected into a HI-FI Aerograph Model 600D gas
chromatograph equipped with a hydrogen flame detector. A stainless
steel column (6’ x 1/8") packed with aluminum oxide 60/80 mesh was
activated overnight at 110°C and sample chromatography run at ambient
oven temperature. Nitrogen was used as the carrier gas with a flow
rate of 25 ml/min. Hydrogen flow rate was also 25 ml/min, and that
of air was 250 ml/min.
Under these conditions ethylene had a retention time of 4 min.

43
The concentration of ethylene in air was measured by reference to the
standard curves (Fig. IV, Appendix) obtained with pure ethylene mixed
with air. The peak area proved to be a better measure of the ethylene
concentration than the peak height and the area was calculated using
the chart integrator recordings.
Because of the limitations of the sensitivity of the equipment
(5 X 10”^ yl C2H4) the ethylene production of whole plants was measur¬
able only beginning the second day after application of a relatively
large dose of dicamba (100 yg per pot). To measure the time course of
ethylene evolution after the application of sublethal doses of dicamba
an experiment with detached leaves was designed. Five grams of leaves
were placed in 50 ml Erlenmeyer flasks and 5 ml of water or dicamba
solution were added. The rates of dicamba were 1, 5, 10 and 60 yg per
flask. The flasks were sealed with vaccine rubber caps and samples
of air were periodically withdrawn for ethylene determination. The
measurements were made every four hours for at least three days.
After every sampling the flasks were opened, flushed with air, and
resealed. The evolution of ethylene by the vaccine rubber caps (93)
was found to be insignificant.
Studies of the Fate of Dicamba in Common Purslane
Four week old plants were transfered to 100 ml beakers containing
Hoagland’s solution and subjected to a foliar application of 0.1 yC of
dicamba in 10 yl of 50% ethanol solution to one of the upper
leaves (ca. 12 yg).
The radioactive dicamba was labeled in the carboxyl carbon and its

44
specific activity was 1.89 mc/mmole. It was 99.5% pure according to
the specification of the manufacturer, New England Nuclear Corporation
through the ^'ourtesy of Velsicol Chemical Corporation.
After treatment with radioactive dicamba, three plants were
harvested at each of the following time intervals: 1 hr, 4 hr, 1 day,
4 days, 8 days. At harvest the treated leaf was washed with 10 ml
of 50% ethanol and each plant was divided into the following parts:
treated leaf, youngest parts (including growing tips and youngest
leaves), leaves, stems and roots. When possible, the leaves were
divided into epinastic and normal. The excised parts were weighed
to 0.1 mg.
A. Preparation of Plant Extracts
The plant parts were ground in 95% ethanol with a Virtis 45
homogenizer, except for the treated leaf, which was ground with a
hand mortar and pestle.
The homogenized material was left overnight at room temperature,
then filtered in a Buchner funnel through Whatman No. 1 filter paper.
The residue was extracted again overnight in 95% ethanol. This pro¬
cedure was repeated until the residue radioactivity was insignificant.
The extracts were combined and concentrated under reduced pressure at
40®C, then brought to 10 ml volume with 95% ethanol. One ml aliquots
of these extracts were evaporated on planchets and counted. The
radioactivity, corrected for the background was used for the calcula¬
tions of dicamba distribution and concentration in the various plant
parts. The washings of the treated leaf and the nutrient solution were
also examined for the presence of radioactivity.

A5
B. Chromatography of Plant Extracts
The metabolism of dicamba in common purslane plants was studied
by the use of paper and thin layer chromatography of plant extracts
after radioactive dicamba application.
Various materials and solvent systems were tried in order to
accomplish the best separation of dicamba and its possible metabolites
- 5-hydroxydicamba, 3,6-dichlorosalicylic acid and 3,6-dichlorogentisic
acid, chromatographed as standards. The best separation was obtained
on Whatman No. 1 paper with isopropanol: ammonium hydroxide (28%) :
water (8:1:1) as a solvent by descending chromatography. Also glass
plates coated with silica gel of 0.25 mm thickness in combination with
the same solvent gave satisfactory results.
Detection of 3,6-dichlorosalicylic acid and 3,6-dichlorogentisic
acid was accomplished under UV light, but for dicamba and 5-hydroxy¬
dicamba another method had to be employed. Silver nitrate solution
(1.7 g AgNO^ in 10 ml water) was mixed with 5 ml NH^OH (28%) and
diluted to 200 ml with acetone (47). The chromatograms were sprayed
with this mixture, dried and exposed to strong sunlight.
Extracts of each excised plant part, washings of the treated
leaf and nutrient solution were concentrated and 10 to 50 pi chroma¬
tographed, depending on their radioactivity, along with radioactive
dicamba standard. The samples were spotted on Whatman No. 1 paper
strips ana separated by descending chromatography with isopropanol:
ammonia: water (8:1:1) solvents for a distance of 30 cm. After drying
the paper strips were scanned with a Packard Radiochromatogram Scanner.
More concentrated combined plant extracts, treated leaf washings

46
and radioactive dicamba standard were spotted on glass plates coated
with silica gel and developed by ascending chromatography in the
same solvent for a distance of 15 cm. Autoradiograms of the plates
were made with Kodak No-Screen Medical X-ray film. Exposure time
was four weeks.
Statistical Evaluation
The statistical methods used to analyse the data included:
a) calculation of standard deviation,
b) Duncan’s new multiple range test,
c) the least significant difference (Isd) according to Steel
and Torrie (158).

47
RESULTS
Morphological Response of Common Purslane Plants to Dicamba
The visual symptoms of dicamba injury to common purslane plants
depended on the method of application and stage of plant growth. The
amount of herbicide applied also determined the extent of damage.
The symptoms of dicamba injury invariably started with leaf epina-
sty. With foliar application this response could be seen after 4 hours
with 10 yg of dicamba per plant (_i.^. , 2 yg per leaf) . Younger leaves
were more strongly affected than the older ones. In the case of root
application the epinasty developed slower and mainly at the top of the
branches. Parallel to the epinastic response was the disappearance of
night leaf movement. Normally common purslane leaves change their posi¬
tion half an hour before the dark period, from more or less perpendicular
to the stem in the daytime into an upright, close to the stem position at
night. On the treated plants the leaves stayed perpendicular or strongly
bent downward in progressing epinasty.
Table 1 summarizes the observations of injury of four week old pur¬
slane plants treated with dicamba in the range of 0.01 to 10.0 yg per
plant.
At the lower rates of dicamba application (1 yg and below) the leaves
recovered from the epinasty with the exception of the treated ones in
foliar application, which remained twisced.
Another early symptom of dicamba action was bending of the main
steins. The bending zone included the first made region and caused the
plants to arch with their apices down, often below the pot level. This

48
Table 1. Morphological response of 4 week old purslane plants to
various rates of foliarly applied dicamba.
Rate Time of first
yg/plant response (hrs)
Description of
first response
Further development
.01 — none as control plants
.05 - none as control plants
.1 24 epinasty of treated leaves
slight bending of plants, later recovery,
but treated leaves remained twisted
.5 24 epinasty of treated leaves
slight bending of plants, later recovery, treated leaves re¬ mained twisted, new leaves small and abnormally shaped
1.0 6 bending of stems, epinasty of treated leaves
epinasty of all leaves, later partial recovery, abundant growth of lateral branches, new
leaves small, abnor¬ mally shaped
5.0 5 bending of stems, epinasty of treated leaves
epinasty of all leaves, chlorosis, necrotic spots, complete defoli¬ ation, swelling of stem, the plants dead in 2 week
10.0 4 bending of stems, epinasty of treated leaves
epinasty of all leaves, chlorosis, necrotic spots, complete defoli¬
ation, swelling of stem and callus ou^"- growths on the apex, the plants dead within 1 week

49
response also occurred very rapidly, within 4 to 6 hours after dicamba
treatment at 1 to 10 yg per plant. Older purslane plants normally have
a prostrate type of growth, with the basal portions of the stem lying on
the soil. Therefore no bending response could be observed in these
plants.
The bending of young purslane plants treated with 1 yg of dicamba
or less was reversible and the plants assumed a normal position within a
few days.
The growth of the treated plants was visibly impaired at dicamba
rates obove 0.5 yg per plant. New leaves were very small, twisted and
elongated. There was a proliferation of growth of small young branches
from the lateral buds in leaf axils which normally remained dormant.
The lateral branches were also extremely small with abnormal leaves.
Below 1 yg dicamba per plant this additional branching contributed to
the recovery, but at higher rates the whole plants eventually died. The
lethal responses started with the treated leaf which became yellowish,
developed black necrotic spots at the base and abscissed. Other epinastic
leaves soon followed this pattern. Often the defoliation was so complete
that only the stems remained. The stems were abnormally swollen in the
upper portions (Fig. 1) and the apex produced a cluster of yellow cal¬
lous intumescence.
The secondary symptoms of injury were identical for foliar and
root applications. The lethal dose of 1:camba was 5 yg for 4 week old,
20 yg for 10 week old purslane, as applied by either method. These re¬
spective doses killed the plants within two V7eeks.

50
Figure 1 Formative effects of dicamba on common purslane plants* a. deformed apex and twisted leaves (left) b* ’ swollen stem, epinastic lorves with miniature
lateral branches in their axils.

51
Influence of Dicamba on Growth and Fresh Weight of Purslane
The growth of the main stem of 4 week old purslane plants after
three rates of dicamba treatment appliea either to the foliage or roots
is shown in Table 2. Statistical analysis of the data revealed that the
first significant difference between the control and treated plants
occurred after 5 days. The stunting of growth progressed with time.
Within the treated plants the strongest effect was achieved by the high¬
est rates of dicamba with both foliar and root application. The differ¬
ence between the foliar and root application were not significant except
at the lowest rate of dicamba (0.5 yg), with root application being not¬
ab Iv less effective than foliar.
The appearance of the purslane plants in this experiment is re¬
corded in Figure 2. At the age of 6 weeks the plants were still growing
upright, but the dicamba treatments caused noticeable bending of the
stems and epinasty of the leaves. Foliar application of the herbicides
resulted in more visual damage. At the age of nearly 8 weeks the plants
assumed a prostrate growth pattern. Those treated with 5 yg of dicamba
per plant were most injured.
No significant changes were found in the growth of the plants which
were treated at 10 weeks of age with 0.5, 1, 5 and 10 yg of dicamba both
in foliar and root application. The length of the main stem was not sig¬
nificantly different from that of the control plants even after 25 days
of treatment. At this stage the elongation of the main stem was very
slow and the growth was expressed rather in branching of the prostrate
plants.

Table
2,
Eff
ect
of
dic
am
ba
on
gro
wth
of
4
week old p
urs
lan
e pla
nts
as
measu
red
by
the
len
gth
of
the
52
I
• CO
cO O CO
•H CO T3 > 4-1 X o P
U) CO c a (U
I—t
CO B 0)
(U p (U CO i-l
C •u •H
CO 6
4-‘ •H 0) 15 x:
H CO 4-i O • o u P
o (U a
U u 0)
o (X u
CO u p o C3
to iH Pu
CO fO •H r-i X O P
4-1 •H >
'O <u CO ■p p to o 0) (X M P ro
0) M <u (1) u
o (0 -H 4J tH
iH ^1
(X, -M
0) c •
c •H CO • CO >»
T—1 3 cO e CO 3 TO o u P
>-✓ 3 m (X (U
e 0) •3
P <u &
p t-H > 3 w O >
to 3 c p
•H <U C *3 to 0) 3 3 e 5 6 P
p 3 P (0 m 3 G) 3 O P 3
pz-i H a
4J a
■u to 0) M 4J U <u 4J 4-1
CO
CO
CO Q
to
’e CO u
4-1 O (U 4J •H CO
1
O T) *3
tH 3 >
>3 ' tH
3XXS O O 0*3. 00 <j\ o o to
• ••••••
vC CM o r** CX3 -d" 3 CM CM CM tH tH t-l »H
x o o
JC P
P 3
to 3 3 ^ O O U •^H
CO CO 00 fO o OO O i p o c es -O’ iH 00 vO vD to 3
CN CM iH iH tH tH iH
i
1
x> o o ■
U -H 4-» •H
§) -H
3a
7a
Ob
2b
6c 3C
3c
3
m P CM <T\ 'O' m . 3 CM iH iH tH tH :H fH j
i
I
u o o o
tp •44 •H *3
P O
3 X X> JO O JO ^ ] CM iH f" tH CM rO
C
o o tH
a
21
ab
18
b
15
ab
16
b
14
b 15
ab
15
•3
P 3 P P 3
• tH
O o^ mt 04 \o 3 m
00 vo CO ir> C3 lo 3 iH tH iH tH tH tH tH
3 1
m
3 .X3 • P P
n 3 3 3 3 3 3 3 to 3 r' vo tH 00 oo m c\ jx p
o -O' CO CO CM tH CO CM •*3 3
p 3 3
tH tH iH iH tH tH tH * 3 00
> s O 3
tH P tH O 3 tp tH
(X 3 H d P S tH
^ 2 »H a o u to
iH in o o «n o o a !••••••
^ S
bO
p 3 3
o iH m o iH »n
tH
•H 3 y
P c H 3 3.0
3 00 C C 3 -H 3 3
g o P P P P M p 3 P p O O O 3 3 3 3 p O O O H "H “H 3 P H
d P!5 PcJ 0e{ tH tH tH o o o o cj [S4 ^ Ci4
I

53
-
Fig
ure
2.
Com
mon purs
lane p
lan
ts afte
r v
ari
ou
s
dic
am
ba tr
eatm
en
ts;
a.1
and b
/ in ro
ot
appli
cati
on,
c/
an
d
d/
in foli
ar ap
pli
cati
on
. T
he
ph
oto
gra
ph
s
were
taken
12
day
s (a
/ and c/)
and
24
days
afte
r ap
pli
cati
on
(b/
and d/)
. E
very
ph
oto
gra
ph pre
sents
the pla
nts
treate
d
wit
h
foll
ow
ing
doses
of
dic
am
ba
(fro
m le
ft
to rig
ht)
: 5
yg
, ly
g,
0.5
yg/p
lant
and
the untr
eate
d co
ntr
ol
(C).

54
The fresh weight of purslane plants harvested at the end of these
two experiments is recorded in Table 3. In the case of the youngest
plants the'^e was a significant reduction in the fresh weight of the
shoots of all the treated plants when compared to the control. In all
cases the fresh weight decreased proportionally to the rate of dicamba
treatment with significant differences found between the lowest and the
highest dose of the herbicide. The differences between the foliar and
root application were pronounced. The older plants showed a signifi¬
cant decrease in the fresh weight of the shoots only when their roots
were treated with 5 and 10 yg of dicamba per plant. The foliar appli¬
cation was not effective. At this stage the differences in growth and
development were very difficult to follow because of the great varia¬
bility of mature plants. It was also noticed that common purslane
plants do' not increase in weight and size under growth room conditions
in proportion to their age.
Morphological Response of Common Purslane Plants to Ethylene
Many of the responses of purslane plants to dicamba were similar to
well known ethylene effects in other plants (20, 29, 137). No data are
available on the response of common purslane to this plant hormone.
Therefore a study was undertaken to establish the sensitivity of pur¬
slane to ethylene fumigation and to observe the visual responses.
Common purslane proved to be very sensitive when submitted to
ethylene fumigation in the range of 0.5 to 100 ppm (Table 4). Within
4 to 6 hours after application of 20 to 100 ppm rapid epinasty of
all the leaves occurred and abscission was initiated. After 24 hours

55
Table 3, Effect of dicamba on the fresh weight of purslane shoots
harvested 25 days after application (in g per pot).
Four and ten weeks old purslane plants were treated foliarly or to the roots with various rates of dicamba. Treatments were run
in triplicate (3 pots with 3 plants per pot). The shoots were harvested 25 days after application and the fresh weight recorded.
Site of
Treatment
Dicamba
yg/plant
Age at the time of treatment (weeks)
4 10
Control 69.ya^ 65.2a
Root 0.5 40.8b 43.3abc
Root 1,0 38.6bc 49.3abc
Root 5.0 17.4cd 38.3bc
Root 10.0 _b 29.4c
Foliar 0.5 36.3bc 57,6abc
Foliar 1.0 24.4bcd 41.5abc
Foliar 5.0 11.6d 43.2abc
Foliar 10.0 43,labc
^Means within columns followed by the same letter do not differ significantly at the 5% level according to Duncan’s multiple range test.
^Dashes indicate no treatment.

56
Table 4. Morphological response of 4 week old purslane plants to
various levels of ethylene fumigation.
Rate Time of first Description of Further development
ppm response (hrs) first response
.5 12 block of night leaf movement
epinasty of the leaves starting from the old¬ est, abscission within one week
1.0 12 block of night
leaf movement epinasty of the leaves starting from the old¬ est, 50% defoliation within 24 hrs, re¬ maining leaves epinastic and chlorotic
2 12 epinasty of leaves
75% defoliation within 24 hrs, remaining top leaves epinastic and chlorotic
5 8 epinasty and abscission of
leaves
nearly complete de-' foliation within 24
hrs, few top leaves remained epinastic and chlorotic
10 8 epinasty and abscission of
leaves
nearly complete de¬ foliation within 24 hrs, very few top leaves remained epin¬ astic and chlorotic
20 50 100
epinasty and abscission of leaves
complete defoliation within 24 hrs

57
defoliation was complete, with no leaves remaining on the stems. With
decreasing ethylene concentration the time of appearance of the first
response was delayed, and abscission occurred at a slower rate.
Still, 5 and 10 ppm of ethylene produced nearly complete defoliation
within 24 hours. The youngest leaves remained attached but they were
very chlorotic. It was also noticed that the few remaining leaves lost
their ability to close at night. This response became the first notice¬
able one in the plants treated with 0.5 and 1.0 ppm of ethylene. The
appearance of the purslane plants treated with 10 yg dicamba or 1 ppm
ethylene shown in Figure 3, as compared with untreated control. The
photograph was taken 24 hours after treatment. At this time about 50% of
the leaves of ethylene treated plants had already abscissed and the older
leaves exhibited epinasty. In contrast, dicamba treated plants bent
strongly (a response not seen with ethylene) with the youngest and middle
leaves exhibiting a strong epinastic response. Defoliation came much
later.
The lowest concentration of ethylene (0.5 ppm) caused epinasty of
the older leaves. However, the ethylene action was reversible. The re¬
moval of the plants from fumigation jars after 1-2 days of treatment re¬
sulted in remarkable recovery although there was some defoliation and
chlorosis during the first days. If left enclosed in the desiccator,
the plants lost all their leaves within a week.
Studies of Cell Membrane Permeability in Common Purslane
The nature of the first response to dicamba treatment (stem bend¬
ing and the epinasty of leaves) suggested that changes in cell

58
Figure 3. Purslane plants 24 hours after dicamba (10 yg/plant) and ethylene (1 ppin) treatment as compared to the
untreated control.
►

59
permeability might occur soon after treatment and the resulting water
and ion fluxes could account for the rapid and unusual movements of
the plant organs.
A. Efflux of Electrolytes from Purslane Stem Segments
Dicamba caused characteristic bending of the stem of the treated
purslane plants within 4 to 6 hours after the treatment with 1 to 10 yg
of the herbicide (Table 1). The bending always occurred in the basal
part of the main stem and was not gradual but occurred suddenly, in 15
to 30 min. The rapidity of this response probably cannot be accounted
for by assymetrical growth of the stem, but rather suggests sudden changes
in cell volumes due to rapid permeability changes.
A study of the efflux of ions from the excised bending regions of
the plants treated with 0.05 to 5.0 yg of dicamba as compared to the
untreated plants supports this hypothesis (Table 5). The release of the
electrolytes during the experiment (24 hrs) from the tissues treated
with 5 yg of dicamba was twice that of the control segments. The efflux
of the electrolytes increased with increased rates of dicamba. At the
rate of 0.5 yg per plant the bending took place after two days and was
not very strong. The 0.05 yg treatment never caused any noticeable
response, and the change of efflux of the electrolytes from the stems
was not significant statistically.
The data in Table 6 show that the differences in the efflux of
ions from the control and the sections excised from 0.5 and 5 yg di¬
camba treated plants were not due to any difference in the total con¬
tent of the electrolytes in these tissues.

60
Table 5, Efflux of electrolytes from purslane stem sections after
dicamba treatment (in percent of total content of electro¬
lytes) .
Four week old purslane plants were treated with foliar applied dicamba at three rates. Six hours after the treatment the basal portions of their stems were excised and immersed in 25 ml of demineralized water for a 24 hr incubation period. Three stems were used for one sample and every treatment and control had 10 replications. After the incubation the electrical conductance of the medium was measured. The samples were boiled at 15 atm pressure and the conductance measured again to find total content of the electrolytes. The data presented are expressed as efflux of the electrolytes after 24 hr incubation in percent of the total electrolyte content.
Dicamba Treatment
(yg/plant)
Efflux of electrolytes
(%)
Control 12.2a^
0.05 16.9ab
0.5 19.8bc
5.0 25.9c
^Means followed by the same letter do not differ significantly at the 5% level according to Duncan’s multiple range test.

61
Table 6. Total content of electrolytes in purslane stem sections as
expressed in mhos/g fr. wt.
The presented data were obtained in the same experiment as in Table 5, The conductance was measured in samples of 25 ml of
demineralized water in which three sections of purslane stems were immersed and boiled under a pressure of 15 atm. Every sample of stems was weighed before immersion. The values are the averages of 10 replications.
Dicamba Conductance Treatment
(yg/plant) (mhos/g fr. wt.)
Control 1084 + 116^
0.05 1002 + 109
0.5 971 + 104
5.0 1018 + 64
^The means are followed by their standard deviations.

62
B, Efflux of Rubidium from Purslane Leaves
Both dicamba and ethylene treatments caused epinasty of the common
purslane leases. Any rate of dicamba treatment above 0.1 yg per plant
gave this effect. The time of the response depended on the rate of the
treatment. Epinasty itself was more gradual than stem bending but
usually the change in the position of leaf was completed within three
hours. This fact and the inhibition of the night leaf movement by di¬
camba and ethylene suggested a possibility of changes in cell membrane
permeability, especially in potassium fluxes.
Rubidium was used to study further possible changes in the permea¬
bility of purslane leaves induced by dicamba or ethylene. Rubidium was
chosen as a convenient analog of potassium and as an element normally
not occurring in plants.
The results of studies of rubidium efflux from excised purslane
leaves pretreated with 10 yg of dicamba or 1 ppm ethylene during 24
hrs are shown in Figure 4. The control leaves did not lose a signifi¬
cant amount of rubidium during the six hr incubation period in deminer¬
alized water. Ethylene and dicamba treated leaves absorbed slightly
more rubidium during the two hr incubation period in RbCl solution and
subsequently lost half of their rubidium content during the first 3 hr
of the incubation in water. The efflux of rubidium from the dicamba
and ethylene treated purslane leaves was very similar both in the kine¬
tics and absolute values.
Studies of Protein Metabolism in Purslane Leaves
Numerous reports in the literature postulate that the action of

yeq
Rb
per
g fr
esh w
eig
ht
63
Figure 4. Efflux of rubidium from purslane leaves after dicamba and
ethylene treatment.
Four week old purslane plants were treated foliarly with 10 yg dicamba or fumigated with 1 ppm ethylene. After 24
hrs duplicate samples of 5 g leaves were excised and im¬ mersed for 2 hrs in 10~^M kbCl then transferred to demin¬ eralized water. At 1.5 hr intervals Rb content in samples was determined with atomic absorption spectrophotometry.
Each symbol represents the average of six replications, as
experiment was repeated 3X.

64
certain herbicides in plants is connected with their influence on plant
protein metabolism. The purpose of this study was to determine whether
dicamba and ethylene influence this process in common purslane leaves
and whether such influence could be considered as a primary site of
action,
A, Nitrate Reductase Activity in Purslane Leaves
Nitrate reductase was chosen to study as an inducible enzyme,
which induction might be blocked by systemic herbicides (22), The en¬
zyme is also readily detected in leaves of common purslane.
Table 7 shows the results of periodic assays of nitrate reductase
activity in the leaves of purslane plants treated foliarly with 10 yg of
dicamba as compared to a control. The absolute values of the activity
differed greatly depending on the time of the assay. The low value of
activity after 12 hours in both control and treated plants may be ex¬
plained by the fact that this assay was done in the evening, when ni¬
trate reductase activity temporarily dropped, to raise again next morn¬
ing, Despite strong epinastic response nitrate reductase activity in
the leaves taken from dicamba untreated plants did not change signifi¬
cantly until the second day when nearly 80% inhibition was noted.
The results of similar experiments performed with common purslane
plants treated with 1 ppm ethylene are presented in Table 8. Ethylene
fumigation of purslane caused a rapid decrease in nitrate reductase
activity in the leaves. After 6 hours the change was noticeable and
after 12 hours of fumigation 80% inhibition was noted, comparable to
the dicamba effect after two days.

65
Table 7. Nitrate reductase activity in leaves of purslane plants
treated with dicamba.
Four week old purslane plants were treated with 10 yg dicamba applied to the foliage. At indicated time intervals triplicate samples of 4 g of leaves were assayed for nitrate reductase ac- tivity and protein was determined in the enzyme reported values are averages of six samples each with triplicate samples).
extract. The (two experiments
Time after Nitrate reductase activity (ymoles NO2 /mg protein/hr)
application Control Treated Ratio
0 .154 + -013^ 100
6 hr .174 + .012 .154 + .011 91
12 hr .080 + .010 .076 + .001 95
1 day .225 + .011 .214 + .010 84
2 day .165 + .009 .038 + .007 23
^The last column represents the ratio of the activity of treated samples to that of control at the same time expressed in percent.
^The means are followed by their standard deviations.

66
Table 8. Nitrate reductase activity in leaves of purslane plants treated with 1 ppm ethylene.
Four week old purslane plants were enclosed in 10 1 desiccator jars
equipped with CO^ traps fumigated with 1 ppm ethylene or left un¬ treated. At indicated time intervals triplicate samples of 4 g of leaves were assayed for nitrate reductase activity and protein content was determined in the enzyme extract. The reported values
are averages of six samples each (two experiments with triplicate samples).
Nitrate reductase activity (ymoles NO2 /mg protein/hr)
Duration of Control Treated Ratio
fumigation (1 ppm
6 hr .270 + .020^ .200 + .015 74
12 hr .260 + .014 .050 + .013 19
The last column represents the ratio of the activity of treated samples to that of control at the same time expressed in percent.
^The means are followed by their standard deviations.

67
B. Studies of Protein Synthesis in Purslane Leaves
The activity changes of just one inducible enzyme cannot represent
the whole complex picture of protein systhesis and the studies of incor¬
poration of labeled amino acids provide more insight of total protein
synthesis.
The results of the studies of phenylalanine “ U-C^^ incorporation
into protein in the leaves of purslane treated with 10 yg dicamba are
presented in Table 9. During the first two days of treatment incorpora¬
tion did not change significantly. It decreased by half on the third
day, dropped by another 20% of control on the fourth day and then con¬
tinued to decrease very slowly. Thus the action of dicamba on total
protein synthesis was not an immediate response but occurred much later
than visible symptoms.
Another experiment included fumigation with 1 ppm ethylene and
treatment with cycloheximide, an inhibitor of protein synthesis. The
results of this study are shown in Table 10.
Cycloheximide, applied to the foliage at 50 yg per plant did not
cause any visible response in purslane plants during the time of the
experiment. However, within 24 hours cycloheximide reduced phenylalanine
incorporation by 70%, which was comparable with the dicamba effect after
4 days. At the same time ethylene decreased the incorporation by over
40%, similar to dicamba after 3 days.
The differences in phenylalanine incv. rporation did not result from
significant differences in the uptake of the radioactive amino acid, as
the counting of the homogenized leaves immediately after the incubation
had shown (Table 11). There were also no significant differences in the
total protein content as assayed in alkaline extracts (Table 12).

68
Table 9. Phenylalanine-^^C incorporation into protein in leaves of
purslane plants treated with 10 pg dicamba.
Four week old purslane plants were treated with 10 yg of foliar applied dicamba. At the same time each day triplicate samples of leaves (1 g) were excised and incubated with ^^C-labeled phenylalanine (spec, activity 224 mC/mM) in buffer solution for two hours. After the incubation period, protein was extracted from the samples, purified, weighed and its radioactivity assayed. The counts are corrected for background. The data are the means of six replications (two experiments with triplicate samples).
Time after Phenylalanine incorporation (cpm/mg protein)
treatment (days)
Control Treated (10 yg)
Ratio %
1 156 167 107
2 154 165 107
3 152 82 54
4 155 48 31
5 150 47 31
6 153 43 28
Isd^ 14 9
Isd = least significant difference at 5% probability level. The difference between two means is significant it it exceeds
Isd.

69
Table 10. Phenylalanine-^^C incorporation into protein in leaves
of purslane plants treated with 1 ppm ethylene or 50 yg
cycloheximide.
Four week old purslane plants were treated with 50 yg cyclo¬ heximide applied to the foliage or fumigated with 1 ppm ethylene. After a 24 hr treatment period triplicate samples of leaves (1 g) were excised and incubated with ^^C-labeled phenylalanine (spec, activity 224 mC/mM) in buffer solution for two hours. After the incubation period, protein was extracted from the samples, purified, weighed and its radioactivity assayed. The counts are corrected for background. The data are the means of six replications (two experiments with triplicate samples).
Treatment Phenylalanine incorporation (cpm/mg protein)
Percent of control
Control 118 100
Ethylene, 1 ppm 69 58
Cycloheximide, 50 yg 35 30
Isd^ 17 15
^Isd = the least significant difference at 5% probability level. The difference between two means is significant if it expeeds Isd.

70
Table 11. Phenylalanine-l^C uptake by leaves of purslane plants
treated with 10 yg dicamba, 1 ppm ethylene or 50 yg
cycloheximide.
Four week old purslane plants were treated with 10 yg dicamba or 50 yg cycloheximide applied to the foliage, or fumigated with 1 ppm ethylene. At indicated time intervals triplicate samples of leaves (1 g) were excised and incubated with I'^C-labeled phenylalanine (spec, activity 224 mc/mM) in buffer solution for two hours. After the incubation period
the leaves were ground in 9 ml of 0.01 M HgCl2 and aliquots of the homogenate assayed for radioactivity. The counts, corrected for the background represent total uptake of phenyl¬ alanine. The data are the means of six replications (two experiments with triplicate samples), followed by their standard deviations.
Phenylalanine uptake (cpm/g fr. wt.)
Treatment Control Treated
Dicamba, 10 P8» after 1 day 5253 + 314
II II ft It 2 days 5301 + 559
II II fl If 3 " 4839 + 120
11 /
II II If 4 " 5607 + 867
II II If It 5 ” 4821 + 315
II II If If 6 " 5238 + 602
Ethylene, 1 ppm. after 1 day 4257 + 157
Cycloheximide, 50 yg, after 1 day
4726 + 643
5694 + 900
5625 + 710
4639 + 589
4905 + 451
4496 + 504
4890 + 558
4693 + 603

71
Table 12► Protein content in leaves of purslane plants treated with
10 yg dicamba, 1 ppm ethylene or 50 yg cycloheximide.
Protein was extracted from crude homogenate of purslane leaves, obtained by grinding 1 g samples in 8 ml HgCl2 (0.01 M) . After precipitation with 10% TCA, the precipitate was extracted three times with 5 ml 1 N NaOH at 100®C for 15 min. Combined protein
extracts were used to assay protein content by recording optical density at 260 and 280 my (105). The data are the means of six replications (two experiments with triplicate samples), followed by their standard deviations.
Protein content (mg/g fr. wt.)
Treatment Control Treated
Dicamba, 10 yg, after 1 day
It
II
11
II
II
II II
11 II It
II It
II II
II II
ti
II ^ II
It
Ethylene, 1 ppm, after 1 day
day 40. .0 + 1. .6 39.5 + 5.2
days 41. ,5 + 3. .5 41.5 + 2.5
II 30. .0 Hr 3. ,0 34.9 + 5.4
11 37, .1 + 0. .5 33.5 + 0.5
II 33, .5 + 3, .5 36.0 + 6.0
11 39, .7 + 2, .3 36.2 + 2.9
. day 32, .6 + 4, .9 31.7 + 5.6
ter 1 day 34.0 + 6.3

72
Effect of Dicamba on Ethylene Production in Purslane Plants
The similarity of common purslane response to dicamba and ethylene
inevitably posed a question of ethylene pjoduction by the plant after
herbicide treatment. Normal ethylene production of the untreated plants
was so low that three well-developed plants placed in a 10 1 desiccator
produced barely detectable amounts of ethylene in a 48 hour period. Ne¬
vertheless these minute amounts of ethylene building up in the closed jar
did produce ethylene responses during one week enclosure and some defol¬
iation occurred. This indicates a great sensitivity of common purslane
to ethylene, as compared to some other plants. For example, bean plants
(Phaseolus vulgaris L. var. Red Kidney ) did not defoliate even at 8 ppm
ethylene in a similar experiment.
It is not surprising then, that application of 1, 5 and even 10 yg
of dicamba per plant failed to reveal any substantial increase in ethy¬
lene production by common purslane. The gas chromatograph available for
use was simply not sensitive enough to record eventual differences.
However, when a comparatively large dose (100 yg) of dicamba was
applied to the roots of purslane, the ethylene level in the desiccator
jar rose to 0.5 ppm during the first 24 hours and reached 1 ppm after
48 hours.
In order to study the effects of lower levels of dicamba, which were
used throughout this work, detached purslane leaves enclosed in 50 ml
Erlenmeyer flasks were used. The smaller volume of flask permitted a
more reliable determination of changes in ethylene evolution. The
course of ethylene evolution from excised leaves treated with various

73
doses of dicamba is recorded in Figure 5.
The control leaves were not stimulated by excision to produce a
large amount of ethylene. One yg of dicamba doubled the value of the
ethylene evolution of control leaves within 8 hours, which could be sig¬
nificant physiologically for very sensitive purslane. Five yg of di¬
camba tripled ethylene evolution within the first four hours and the
treated leaves maintained the rate of about 2 myl C2H^ per gram fresh
weight per hr during the next two days. After the application of 10 yg
dicamba the treated leaves achieved an ethylene evolution four times
greater than that of the control during the first four hours and the
production of ethylene continued to rise steadily during the next 16
hours. The ethylene evolution then reached a steady rate of about
5 myl per gram fresh weight per hour, which was maintained for a week
until death of the leaves.
The highest rate of dicamba (60 yg) resulted in the largest in¬
crease in ethylene production. During the first four hours it was seven
times greater than the control, and within 24 hours it achieved a rate of
10 myl/g per hour. The peak of ethylene evolution occurred after two
days, after which the production decreased slowly during the next 6 days
when it disappeared with the decomposition of the leaves.
These results indicate that dicamba causes an increase in ethylene
production in purslane tissues to levels many times higher than normal.
The increase of ethylene production depends on the applied rate of
dicamba, with higher doses causing greater response within the investi¬
gated range. The duration of the elevated ethylene production is also
a function of dicamba concentration with the longest period occurring at

74
4-J CO <U
4J CO ccj (U 0) > Pu cd 0) 0) M
CO (U cd
§ ® i-i 4J
CO C
U <U • D 6 CD C1.«H 0)
U tH 'd <u a Q) a, g ,c X cd O W CO cd U 0) <u • +J
Td <d cd rO o
C 6 ‘H •H cd rH
U P4 C -H d o'd Td
•H 4J M-l ^ O O U d
Td CO o <u M 4J CO a. cd q)
M d d CO +-> (u d
t-h o , ‘H 0)
rd M M 4-1 cd ^ W > 4-1
•H 15
B •H
0)
m (U V4 d bO
♦H PC4
aq X ‘ JiJ Trtui

75
10 pg of the herbicide. With lower doses ethylene production eventually
dropped to the control level, while higher dose (60 pg) hastened the de¬
composition 01 the tissues with complete loss of ethylene production.
The ethylene response is very rapid and differences may be noticed
during the first four hours after the dicamba application. Elevated
ethylene production thus precedes all the other observed visual and phy¬
siological responses of common purslane to the herbicide.
Absorption and Translocation of Dicamba in Purslane Plants
The absorption of radioactive dicamba applied in a droplet to a
single leaf of four week old purslane plants is shown in Table 13. The
values represent all the radioactivity recovered from the plants after
application of 0.1 pC dicamba-^^C (50,000 cpm) with subsequent washing
off of the residue with 50% ethanol. The radioactivity recovered
from the nutrient solution was also included.
The amount of dicamba applied, about 12 pg, was lethal for the
four week old purslane plants. The treated leaves absorbed only 10% of
the dose during 8 days with most of the herbicide absorbed during the
first day. The necrotic spots observed on the treated leaf could impair
both absorption and transport of the applied dicamba. Nevertheless, the
absorbed amount was high enough to cause the typical responses of stem
bending, leaf epinasty and subsequent death of the treated plants within
one to two weeks.
The translocation of the absorbed dicamba in purslane plants as
measured by the radioactivity of the extracts from various plant parts
is shown in Table 14 and Figure 6. From the treated leaf it moved

76
Table 13. Absorption of dicamba-^^C in purslane plants expressed as
percent of the applied dose.
Four v/eek old common purslane plants were treated with a 10 yl droplet containing 0.1 yC dicaraba-^^C^ (12 yg). Three plants were harvested at the indicated time intervals and the treated leaf washed v^ith 10 ml of 50% ethanol. The plants were dissected, ground and extracted with 95% ethanol. The extracts were counted
in a gas flow counter. The following values represent average total radioactivity recovered from the treated plants and their nutrient solution in percent of the applied dose (50,000 cpm).
Duration of absorption % of applied dose absorbed
1 hr .5
4 hr 1.0
1 day 8.6
4 days 9.5
8 days 10.4

77
2400
2000
1600
1200
800
400
Hours after Treatment
rigure 6. Translocation of dicamba in purslane plants during
the first day after treatment.
Dicamba-l^^C, (.1 yC) was applied as a 10 yl drop to a single leaf. The radioactivity was extracted from plant parts with 95% ethanol. Every symbol
represents an average of three replications.

Table
14.
Tra
nsl
ocati
on
of
dlc
amba-
-^‘C
in p
urs
lan
e pla
nts aft
er
foli
ar
ap
pli
cati
on
; in
cpra
78
CO XJ <ui o> o 4-1 CO O O *H >-i CO (U (U 4-1 tH cn •u X CO cn CO G)
•H V4 CO T3 CO CU •H
CX. x: (U 0) 4-1 u CO 4J Q) 4-1 •H
CO o > CO •H
cn CO 4-t 4-» 4-1 O c 4-» .-H CO CO ^4 3 O
t—t CO CO •H cx CX, (U TO
cO 0) x: i~i Q) E-* <U X d
> 4-* o •H
• <u 4-1 CO i-l d I—1 • cO tH
CO O cu 4-» CO CO
I—i O OJ O 3 4-1
CO M iH d H CO <u
'O > •H C c: V4
•H CO T3 4J (U d
n •> 4-» d cO CO M
E O <u 4-> (U a. X C 4-t a; H (U cn M E
•H .. 0) • CO X CO
O <U H 4-1 CX, > d ><: CO CO (U 0) . iH
tH rH CX <u O E * d d <0 cn CO d cn X
»-l 4.J X (i> CO <1> 4-1 x: fX
E 00 m o
E c o^ xi o d 4-1 V4 O X
U-l CO •H d
(U - 5 00 • 4-1 CO
4-1 cd CO 'V Xt V4 (U <u d CO TJ iH 4-» > Cu 0) O cO
4-> TO CO 4J c (I> >4 c GJ 4-i 4J CO 00 CO X E
I—1 <u (U (U cx cx u o
(X 4-1 TO N--' • u c •v 0) CO CD cO 00 d a. 4-» X d 4-1
q) 4-» TO •H d ’O c 4J d
O 3 d CO (U 4-» o d d x: c u o )-l
-H 00 o cx
+j
nj (X
C rH &.
'g (X o
4J
o CO o
•H
CO Pi
M *-»
e *-» H CO •»->
E rH d O mJ- CN
•H X TO 1 1 + 1 +1 + d 1 1 S o CO
CO CM CM
X CM CO X CM CM XO CO 4J o +1 + 1 + 1 +1 + o Pi vd o CM <!■
X m CO in X
X CM o CO CO CSJ CM '0- vO CM E d + 1 + i + 1 + 1 + 4-1 CO o c^ <1- 00
CM CM o 'O' X O X vO
X X
CO lO X CM
CO CM m CM 'O' X d > +1 + 1 + 1 +1 + CO d or c mT CO CO X rH CM CO CO m
CO MO o CM
X CO m o o CO
m X CM X vO 00 cn d 4J +1 +1 +1 +1 + d H O CO m o o CM pi cx CM X in o rH
X CM crt vO CM X X
CO o o X CO o
TO CO CM '0- 'O' CO d 4J 4-4 + 1 + 1 +1 +1 + CO CO d d <0^ 'O" o 00 d rH 'O- CM X m X
H nH CM vO m m
4-1 d d cn CO E >s 4J u M CO CO CO CO X X TO TO •d d Xi r—i MT X >0' 00
I
^T
he
mean
s are
foll
ow
ed
by th
eir
sta
nd
ard devia
tions.

79
rapidly into the stem and was translocated into young parts. Accumula¬
tion in the growing tips and buds was so rapid that it reached a maximum
after one day. Subsequently some of the '^icamba was apparently redis¬
tributed into the stems, leaves and roots. The roots exuded dicamba to
the nutrient solution, but altogether less than 0.5% of the applied dose.
The distribution of dicamba in different plant parts is more clearly
represented in Table 15, which shows the radioactivity of every part in
percent of the absorbed amount at each particular time. During the first
few hours most of the radioactivity x^as found in the treated leaf, but
after one day the share of the treated leaf dropped to 14%, and to 10%
after 8 days. The young parts accumulated dicamba very rapidly, account¬
ing for half of the total absorbed radioactivity after one day. The
leaves accumulated dicamba slowly, but at the end of the experiment they
contained 40% of total plant radioactivity. The translocating stems had
a fairly stable percentage of dicamba amounting to one fourth of the
total during the first four days. The roots apparently did not accumu¬
late dicamba, but exuded radioactivity into the nutrient solution, losing
4% of the absorbed amount during 8 days.
Some insight into the physiological significance of dicamba
translocation pattern may be gained from comparing the concentration of
the herbicide in various parts at different times (Table 16 and Fig. 7).
The highest concentration of label was found in the treated leaf, with
the peak occurring one day after the application. The concentration of
dicamba in the youngest parts of the shoots was also growing very rapidly
and the difference was very significant 4 hours after application when
compared with, stems, leaves and roots. One day after application the

80
Table 15. Distribution of dicamba-^^C in purslane plants expressed
in percent of the absorbed amount.
Time % radioactivity after " treatment Treated
leaf Young
parts Leaves Stems Roots Medium
1 hr 66.0 11.0 8.4 11.9 2.7 _a
4 hr 44.1 23.0 3.9 27.6 1.4 —
1 day 14.4 52.4 8.9 23.9 .2 .2
4 days 11.6 39.8 13.3 23.1 11.6 .6
8 days 10.0 31.0 39.5 12.5 2.6 4.3
^Dash stands for : no radioactivity was detected. The values within rows add up to 100%.

81
Table 16. Concentration of radioactivity in various organs of purslane
after foliar application of 0.1 pC of dicamba-^'^C; in cpm/g
fresh weight.
Time Radioactivity cpm/g fr. wt. after -
treatment Treated Young Leaves Stems Roots leaf parts
1 hr 1333 + 205^ 61 + 4 10 + 1 24 + 00 36 + 10
4 hr 2285 + 116 194 + 36 10 + 2 68 ± 9 34 + 6
1 day 9938 + 507 3390 + 315 163 + 44 720 + 58 129 + 31
4 days 5415 + 208 4422 + 310 839 + 61 1142 + 124 993 + 32
8 days 4440 + 205 1634 + 112 669 + 34 258 + 28 473 + 46
^The data are averages from three replications. Every mean is followed by its standard deviation.
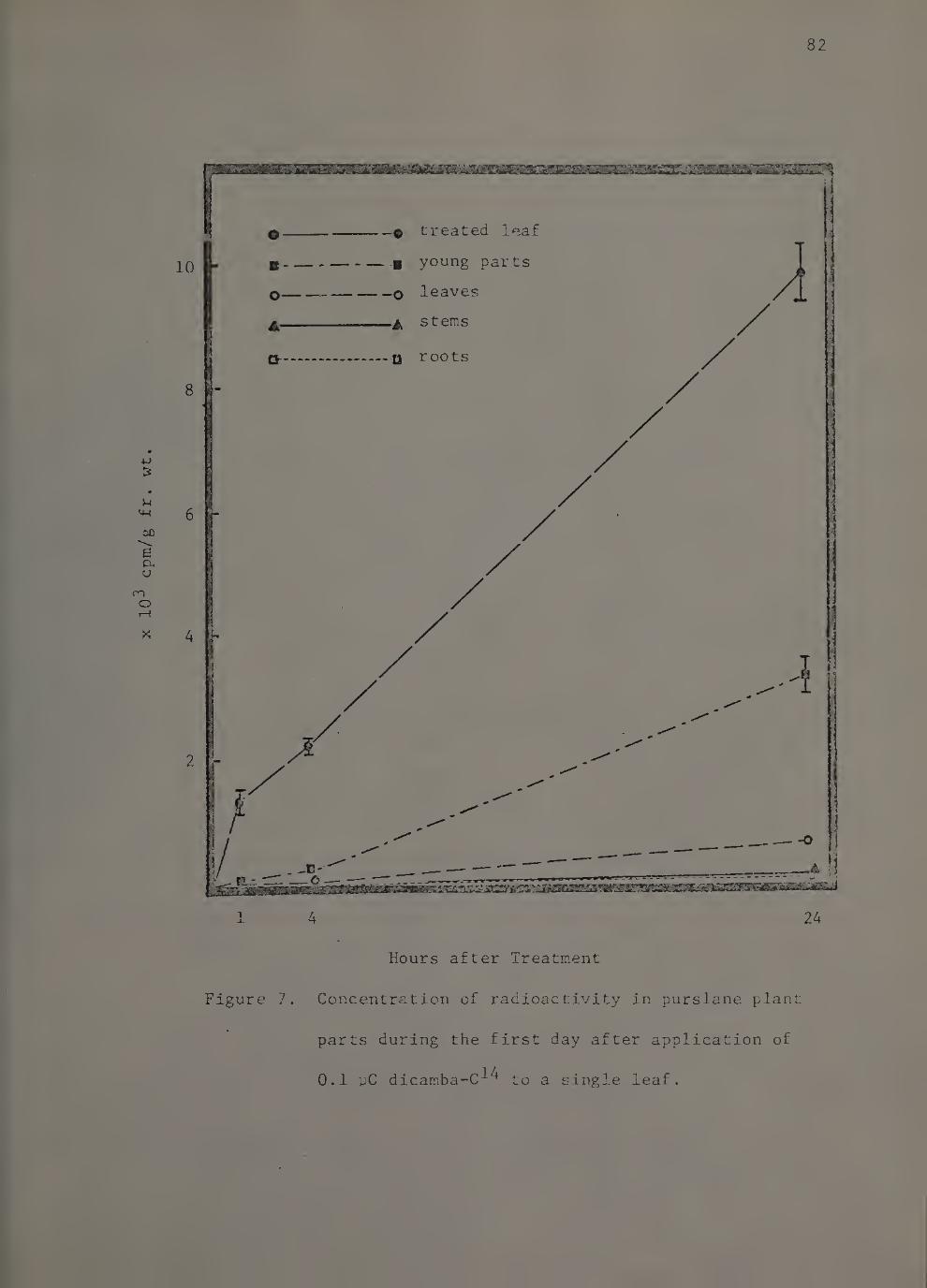
10
^
cp
m/g fr,
wt.
82
10
8
6
« 4
Hours after Treatment
Figure 7. Concentration of radioactivity in purslane plant
parts during the first day after application of
0.1 yC dicamba-C^^ to a single leaf.

83
content of label in the young parts was twenty-four times greater than
in leaves and roots, and the four-fold difference remained to the end
of the experiment. It is noteworthy that so much dicamba accumulates so
quickly in the young parts, considering that the young tissues are most
sensitive to herbicide action.
The concentration of radioactivity in the leaves, stems and roots
was uniformly low and similar in all these organs. However, the label
content of stems was notably higher during the first days of transloca¬
tion, presumably due to participation of stem vascular bundles in dicamba
movement out of the treated leaf.
After one day of treatment some of the mature leaves were affected
by dicamba and showed visual symptoms such as epinasty, while others still
appeared normal. Separate extraction of those two types of leaves re¬
vealed that the epinastic ones had 570 + 39 cpm/g fresh weight, while
the unaffected leaves showed only 79+11 cpm/g fresh weight. The ap¬
pearance of epinastic response was correlated with the concentration of
dicamba in leaf tissues. Later on all the leaves were epinastic and
their radioactivity content was in excess of that just mentioned.
Studies of Dicamba Metabolism in Purslane Plants
The radioactivity of the washing from the treated leaf surface
accounted for the bulk of the applied dicamba left unabsorbed (45,000
to 50,000 cpm). Therefore, the volatilization of dicamba from the leaf
surface or decarboxylation of the herbicide in the plant did not appear
significant.
The extract from plants, nutrient solution and leaf washings were

84
chromatographed along with dicamba-l'^C standard. Table 17 shows Rf values
of radioactive spots determined using a radiochromatogram scanner on the
chromatographs of the extracts from plants harvested 8 days after the
treatment with radioactive dicamba. Only one peak was found in every
case and it was identical with the dicamba standard. The radioactivity
of the nutrient solution was also determined to be pure dicamba.
The autoradiograph of dicamba standard, treated leaf washings and
combined plant extracts after an 8 day absorption period, made of thin
layer plate, confirmed the results of paper chromatography (Fig. 8).
The leaf washings and plant extract contained only dicamba as radioac¬
tive material. There was no decomposition on the leaf surface and no
metabolism of dicamba occurred during 8 days.

85
Table 17, Distribution of radioactivity along chromatograms of
extracts from purslane plants 8 days after application
of dicamba-^^C to a single leaf.
Chromatograms were run of dicamba-^^C standard, nutrient solution and ethanol extracts of the residue on the treated leaf, of the treated leaf, young parts, leaves, stems and roots of purslane plants with 0.1 yC dicamba-^'^C to a single leaf. Chromatrographi
solvent was isopropanol: ammonia: water (8:1:1) and paper strips of Whatman No. 1 type were used. The reported values are the average Rf of the peaks found with a Packard Radiochromatogram Scanner. The peaks usually covered the area of 0.1 Rf (+0.5 reported value).
Chromatrographed material Rf values
.78
Washings of treated leaf .77
Extract of treated leaf .75
Extract of young parts .75
Extract of leaves .74
Extract of stems .76
Extract of roots .76
Nutrient solution .77

: 7 -
Figure 8. Autoradiograph of silica gel thin layer plate chromatogram
of dicamba-C^^, washings from the treated leaves, and ex¬ tract of common purslane plant (in that order, from left to right), 8 days after treatment with 0.1 yc of dicamba-^^C to a single leaf. Developed in isoproponal: ammonia: water
(8:1:1).

87
DISCUSSION
Susceptibility of Common Purslane to Dicamba
The results presented indicate that common purslane is quite
sensitive to dicamba treatment. The lowest rate of dicamba application
which caused a visual response was 0.1 yg per plant. This is compar¬
able to the rates reported for other broadleaved plants, such as soy¬
beans (171), Canada thistle (48) and Tartary buckwheat (50). The
maximal response depended on the age of the purslane plants with the
observed killing rate being 5 yg for four week old and 20 yg for ten
week old plants.
Common purslane exhibited characteristic responses to dicamba.
Stem bending appeared promptly after application of the herbicide to
the young upright plants. Similar bending was reported for soybeans
(171) and Canada thistle (48) after comparable doses of dicamba.
Equally rapid was the development of epinasty in the treated leaf
(Table 1) and the leaf opposite to it. Other injury symptoms included
necrotic spots on the most affected leaves and finally defoliation.
The secondary response involved branching as in the case of soybeans
(171) and flax (131). Killing of the terminal buds was connected
with this process of removal of apical dominance. Inhibition of
elongation and stem swelling were observed as reported earlier for
pea seedlings (36). The reduction of growth was most pronounced in
younger plants (Table 2), gradually diminishing when the plants
assumed the natural prostrate growth pattern with the production of
many branches.

88
The influence of the herbicide was pronounced in the reduction of
the fresh weight of the shoots, due to both growth stunting and de¬
foliation. Formative effects of additicnil branching from the leaf
axils did not influence the fresh weight as those branches and leaves
were extremely thin and small.
The multitude of observed responses suggested a possibility of
intermediate steps in dicamba action. Many of the responses resembled
typical ethylene effects, such as stem swelling, epinasty and defol¬
iation with loss of apical dominance. On the other hand, the stem
curvature, which was the first response to dicamba action, is typical
for growth hormones, and could not be produced by ethylene. Therefore
any explanation of dicamba action must take into account this hormone
action as well as its eventual ability to stimulate ethylene production.
Susceptibility of Common Purslane to Ethylene Fumigation
The fumigation experiments should be considered with great caution,
because a uniform ethylene concentration was applied at the same time
to all plant tissues and organs. In natural conditions the youngest
plant parts produce more ethylene than other organs (40, 71). Con¬
siderable ethylene evolution has been noted from flowers and fruits
(29), but in the presence of natural air movement this ethylene does
not appear to influence the mature leaves and stems. Nevertheless,
fumigation with ethylene is helpful in the investigations of the gas
levels needed to cause typical responses.
Common purslane was very sensitive to ethylene and the plants lost
all their leaves at levels in excess of 5 ppm (Table 4) . The sequence

89
of abscission proceeded from the oldest to the youngest leaves.
The top leaves, although chlorotic, remained on the plants for the
longest time. This observation agrees wi'-h the theory that aging
proceeds abscission and makes the leaves more sensitive to ethylene
(9, 70).
Previous to the defoliation caused by ethylene it was observed
that the leaves lost their ability to close at night and assumed an
epinastic position. These two responses were more easily observed
at lower ethylene levels (0.5 and 1.0 ppm), where the defoliation was
much delayed.
The ethylene responses described resemble those caused by dicamba.
Swelling of the stem and deformation of the growing tip were not
observed because of difficulties in obtaining low levels of ethylene
fumigation which would not kill the plants. Even control plants
enclosed in the desiccators would not live long enough to produce
these responses due to their ethylene production and accumulation of
the evolved gas in the jar.
The only response produced by dicamba which could not be ascribed
to ethylene action was the early curvature of the stem, which indicates
auxin-like properties of dicamba.
Effect of Dicamba on Cell Membrane Permeability in Purslane
The bending response of common purslane stems to dicamba treat¬
ment resembled auxin response and was not produced by ethylene. It
occurred a few hours after the treatment and the actual time of change
was very short (30 min). Sudden changes of plant organ position are

90
best explained by changes in membrane permeability and efflux of ions
(94), specifically potassium (13). Therefore it was of interest to
investigate the permeability changes in the bending region of purslane
stems after dicamba application.
The investigation of electrolyte efflux from purslane stem frag¬
ments revealed that even small doses of dicamba increase the ion
release from the tissues (Table 5). At 5 yg of dicamba per plant the
efflux was twice that of the control, which could be of physiological
significance. Cell membrane permeability depended on the amount of
herbicide applied and translocated to the bending zone. The plants
treated with low doses of dicamba bent later if at all. However,
once the permeability change in a predisposed region of the stem
reached a certain threshold the efflux of ions would cause such
turgor changes that suddenly the whole stem changed its position
("an all or none response").
The epinasty of leaves and their loss of night movement belong
to a category of "fast responses", but they may also be caused by
ethylene. Nastic responses of Mimosa pudica leaves were connected with
reversible fluxes of potassium ions in the cells of the pulvinus (13).
It could be imagined that if the cell membrane permeability changed
irreversibly, the nastic movements would be arrested.
The results reported in Figure 4 indicate that rubidium was lost
from the lea\cs treated with dicamba and ethylene much faster than
from control leaves. In fact the control leaves did not appear to
lose any rubidium during the first six hours. This observation is in
agreement with many reports of rubidium and potassium efflux (140, 141)

91
from plant tissues. Rubidium is presumably absorbed by the mechanism
normally operating for potassium (62, 140, 156, 161).
It is reasonable to assume that if the bulliform cells, suspected
to participate in purslane leaf movement (64), irreversibly lost
potassium, they could not resume their rhythmical activity and the
leaves would remain open during the night.
An important feature of the increased ion efflux is the fact that
within 24 hours both 10 yg dicamba and 1 ppm ethylene produced nearly
identical results. It indicates that dicamba action might be connected
with that of superoptimal levels of ethylene in purslane.
Effect of Dicamba on Protein Synthesis in Purslane Leaves
The experiments with the influence of dicamba on nitrate reductase
indicated that the enzyme activity was not affected until two days
after the treatment (Table 7). It was not a "first response" as
opposed to many visible morphological changes. The drop in activity
occurred when the leaves were already in strong epinasty, the stems
were bent and the increase of the cell membrane permeability in the
stem bending regions and leaves were noticed.
The half life of nitrate reductase in corn is only four hours (152),
so the changes in the activity of this short-lived enzyme are a
sensitive tool of research of disturbances in protein metabolism. The
late, but strong inhibition of nitrate reductase activity (80%) in
purslane leaves might be caused not by dicamba itself but mediated
through changed cell membrane integrity, ion concentration in proto¬
plasm or other factors influenced by dicamba.

92
Ethylene fumigation at 1 ppm had a more rapid impact on nitrate
reductase activity in common purslane leaves (Table 8). Slight
changes were noticed 6 hours after the start of fumigation and at
12 hours the enzyme activity had decreased to 20% of control. The
effect of ethylene on nitrate reductase activity seems more immediate
than that of dicamba, and an increased level of ethylene in the tissues
due to dicamba could be a mediator of nitrate reductase suppression.
Total protein synthesis in purslane leaves as measured by ^^C-
phenylalanine incorporation showed inhibition by dicamba on the third
day after its application (Table 9), and steadily decreased afterwards.
Similar inhibition was achieved within 24 hours of ethylene fumigation
at 1 ppm. By comparison, 50 yg of cycloheximide inhibited protein
synthesis stronger than ethylene during the same time (Table 10).
The inhibition of total protein synthesis was so delayed that it
must be considered as a secondary effect of dicamba, possibly mediated
through earlier changes. Nevertheless, it is the one which can greatly
contribute to death of the leaves, as protein synthesis is reduced
80% six days after treatment.
Most of the available reports (74, 124) do not reveal strong
inhibition of protein synthesis by rather high rates of dicamba, but
the duration of treatment was possibly too short for slowly developing
responses.
Effect of Dicamba on Ethylene Production in Purslane
The similarity of responses of common purslane to dicamba and
ethylene suggested that dicamba might trigger an increased ethylene

93
production in this plant. Many of the observed effects would thus be
the results of ethylene action as an intermediate.
In author^s preliminary tests dicamba greatly increased ethylene
production in kidney bean plants. The increase of ethylene concentration
in desiccators containing three potted bean plants reached forty times
that of control just 24 hours after 100 pg of dicamba were applied to
the roots. Epinasty and stem swelling were observed later but no
defoliation occurred. Under these conditions purslane was able to
produce enough ethylene to achieve 0.5 ppm in 10 1 jars during the
first 24 hours after treatment and 1 ppm after 48 hours. Lower rates
of dicamba did not produce clearly observable ethylene increases.
However, measurements were limited due to the low sensitivity of the
gas chromatograph used. The levels detected in the desiccators after
the application of 100 yg of dicamba were above physiological levels
for ethylene response and caused much faster defoliation than in the
case of dicamba in the range of 0.5 to 10 yg per plant.
The investigations with detached leaves of purslane enclosed in
small flasks enabled the determination of ethylene production in
control untreated tissues and in those treated with low levels of
dicamba. The wounding response was not significant. These experiments
showed that the promotion of ethylene production by dicamba starts
within the first four hours after treatment. The maximum level of
ethylene evolution and duration of ethylene response depends on the
dicamba concentration. The maximum level of ethylene evolution was
proportional to the rate of dicamba treatment in the range of 1 to
6C yg (Fig. 5). The longest period of increased ethylene evolution was

94
achieved with 10 pg of dicamba which caused fairly stable ethylene
evolution at the level of 5 myl C2H^ per gram fresh weight per hour.
The ethylene evolution depends on the internal level of ethylene
in the tissues and so does the physiological response to ethylene.
Burg and Burg (33, 34) derived a factor for calculation of the internal
level of ethylene on the basis of ethylene evolution and confirmed it
by bioassays with pea and sunflower. Using their factor of 0.2 ppm
C2H4 as internal concentration when plants produce ethylene at the
rate of 1 yl per kg fresh weight per hour, the excised leaves of
purslane should contain 1 ppm ethylene 20 hours following the
application of 10 yg dicamba. This steady rate of ethylene evolution
(Fig. 5) permits the assumption of a stable internal level of the gas.
From the literature (29, 30, 34) it appears that the concentration of
1 ppm ethylene is saturating for many physiological responses in
various plants.
These considerations could explain similar changes in cell mem¬
brane permeability in the leaves after dicamba and ethylene treatment
(with 10 yg and 1 ppm, respectively). Another support comes from the
fact that the delayed decrease in protein synthesis was also com¬
parable in effects of 10 yg dicamba and 1 ppm ethylene. While it is
difficult to decide whether the permeability changes in the leaves
are mediated through ethylene because of their early appearance (one
day), the protein synthesis in the leaves does seem to be influenced
by the ethylene produced.
In conclusion, dicamba promotes a large increase in ethylene
production in common purslane plants immediately after treatment and

95
the evolved ethylene participates in causing morphological and physio¬
logical responses. The mode of action of dicamba upon this weed
species at least in part is connected with the production and effects
of ethylene.
Absorption and Translocation of Dicamba in Purslane Plants
In comparison with other plants, the data obtained with common
purslane absorption indicate an extremely low rate of dicamba pene¬
tration (Table 13). In a study by Chang (50), ten days after treat¬
ment with 0.1 yC of labeled dicamba, barley absorbed 40%, wheat 50%,
wild mustard 70%,and Tartary buckwheat nearly 90% of the applied dose.
Purslane absorbed 10.5% of the same dose during eight days. One
could speculate that the low rate of absorption was caused by special
cuticular properties of common purslane, a more draught resistant
species than Tartary buckwheat and wild mustard. Another explanation
could be offered by the fact of contact injury of the treated leaf in
the place of dicamba application (black necrotic spots), which possibly
impaired later penetration.
nevertheless, the 10% of the applied dose absorbed was enough to
cause extensive injury and to hasten the death of four week old plants.
Once in the treated plants, dicamba was translocated quickly. First
hints of this translocation were given by the observation of visual
responses - the epinasty of the treated leaf and the leaf opposite to
it, the bending of the main stem, later epinasty of the youngest
leaves on the treated branch, still later epinasty of the top leaves
on lateral branches and finally epinasty of all the leaves. It

96
indicated that the herbicide was translocated rapidly out of the
treated leaf into the stem, and from there with the movement of the
assimilates in the phloem into the youngest parts and downward to
the roots. Leakage from the phloem to xylem might have occurred as
indicated by the response of mature untreated leaves. The accumulation
of the herbicide in the top bud was indicated by growth stunting and
the deformation of the main stem tip (Fig. 1).
The rapid appearance of stem bending and leaf epinasty after root
application indicated that dicamba was absorbed and quickly translocated
in the xylem with the transpiration stream. However, the occurrence
of formative effects in the youngest organs showed that evidently
dicamba was leaking in this case into the phloem. The secondary redis¬
tribution resulted in foliar and root application finally bringing
about essentially the same effects.
The determination of the radioactivity in the plant extracts
substantiated these observations (Table 14). The treated leaf retained
only 10% of total radioactivity after eight days (Table 15). Most
herbicide accumulated in the young tissues at the top of branches and
the concentration of label in those parts was higher than in others
after four hours (Table 16).
There was some redistribution of label from the tops into the
leaves and roots after four days of treatment, and the leakage of
radioactivity from the roots into the nutrient medium was noticed.
Roots did not accumulate much of the herbicide. The loss of dicamba
through root exudation was detectable but not very significant (Table
15). It accounted for only 0.5% of the applied dose eight days after

97
treatment. A similar rate of exudation was observed for Canada thistle
(48), but Tartary buckwheat released 15% of the applied dose during
the same time (49).
When comparing the affected, epinastic leaves after one day of
dicamba treatment with other leaves on the same plant, the concentration
of dicamba in the responding ones was seven times greater than in the
others. The response seems to depend on the accumulation of the
herbicide in the leaves and so on the rate of its translocation.
The rapid translocation of dicamba in common purslane and its
accumulation in growing tips contribute to the explanation of the
sensitivity of this weed to dicamba. The herbicide was translocated
in common purslane in a way similar to other susceptible plants, such
as Canada thistle (48), Tartary buckwheat (49), wild mustard (50) and
beans (89). Resistant grasses (50) tend to retain more herbicide in
the treated leaf and accumulate it in the mature leaf tips.
Studies of Dicamba Detoxification in Purslane
The studies of translocation and the responses of common purslane
suggested that toxic properties of the herbicide were retained when
it reached growing tips and leaves. No significant loss through
volatilization from the treated leaf surface or decarboxylation was
indicated b]/ the high recovery of the applied dose. The radio¬
chroma togrdphy of plant extracts, the residue on the treated leaf and
the nutrient solution revealed only one radioactive compound moving
identically with dicamba-C-14 (Table 17).
Apparently common purslane does not metabolize dicamba at any

98
significant rate during 8 days of treatment. The lack of an effective
detoxifying mechanism contributes to the plant’s sensitivity, even
with low rates of dicamba absorption. Similar results were obtained
with sensitive peas (154), beans (89) and purple nutsedge (111, 113),
in which no indication of dicamba metabolism was found.
Dicamba, remaining unmetabolised for a long time, may play a role
in the stable rate of elevated ethylene production observed for many
days after the herbicide treatment, in the irreversibility of changes
in protein synthesis in the leaves and increased cell membrane
permeability. The formative effects in the youngest leaves and branches
appearing a considerable time after dicamba application contribute to
the mode of action of this herbicide and indicate that dicamba is still
present in plants in a toxic, mobile form, always translocated to new,
forming buds.

99
SUMMARY
Experiments were conducted under controlled conditions to deter¬
mine the response of common purslane to dicamba treatment, the fate of
dicamba in purslane and its mode of action. The results lead to the
following conclusions.
1. Common purslane is rather sensitive to dicamba treatment. The
lethal rate depends on the age of treated plants and increases with their
development. The signs of injury may be seen even at 0.1 yg of dicamba
on plants of any age. The visual response includes bending of the young
stems, epinasty of leaves and inhibition of nyctinasty, later necrosis
of leaves and defoliation, swelling of stems and death of the growing
tip with loss of apical dominance. Stem elongation is stunted, especi¬
ally at earlier growth stages, and the fresh weight of shoots strongly
reduced.
2. The visual responses are very similar to ethylene fumigation
effects and ethylene is a very powerful defoliant of common purslane.
At lower rates ( 1 ppm and below) it causes epinasty of leaves and the
inhibition of nyctinasty comparable to dicamba effects.
3. Dicamba causes a rapid increase of ethylene production in com¬
mon purslane plants to levels many times greater than those observed in
untreated tissue. The high level of ethylene production suggests that
the concein-ration of ethylene within the tissues may be responsible for
many observed dicamba effects.
4. Dicamba increases permeability of cell membranes in common
purslane tissues. An increased efflux of electrolytes was observed in

100
the bending region of dicamba treated plants. Epinastic leaves after
dicamba (10 yg) and ethylene (1 ppm) treatments showed an increased
efflux of rubidium. The permeability effects are seen within one day
after dicamba application.
5. Protein metabolism in the leaves was not influenced by di-
camba treatment until after two days as indicated by the decrease of
nitrate reductase activity. The inhibition of C^'^-phenylalanine in¬
corporation into protein was noticed after three days. Ethylene reduced
both phenylalanine incorporation and nitrate reductase activity within
one day.
6. It is assumed that ethylene produced by the influence of
dicamba in common purslane plants is a factor in the observed permea¬
bility changes and delayed protein synthesis inhibition.
7. Dicamba is absorbed slowly by common purslane leaves, but
the rate of translocation inside the plant is very rapid and may in¬
volve both phloem and xylem transport. Accumulation of dicamba in the
young tissues of growing tips and buds is extremely fast and extensive.
Some dicamba is exuded through the roots following foliar application.
8. There was no indication of dicamba metabolism in common pur¬
slane up to 8 days after the treatment. Only dicamba was recovered from
all parts of the plants and from the nutrient solution.

-101
BIBLIOGRAPHY
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
Abeles, F. Pla It
B. 1966. Auxin stimulation of ethylene evolution. Physiol. 41; 585-588.
Abeles, F. Plant
B. 1966. Effects of ethylene on auxin transport. Physiol. 41; 946-948.
Abeles, F. B. 1968. Herbicide induced ethylene production; role of the gas in sublethal doses of 2,4-D. Weed Sci. 16; 498-500.
Abeles, F. B. 1968. Role of RNA and protein synthesis in
abscission. Plant Physiol. 43; 1577-1586.
Abeles, F. B., L. E. Crocker, and G. R. Leather. 1970. Abscis¬
sion; the phytogerontological effects of ethylene. PI. Physiol. (Suppl.) 46; 32.
Abeles, F. B., and R. E. Holm. 1966. Evidence for an hormonal role for ethylene in abscission. Plant Physiol. (Suppl.) 41: nil.
Abeles, F. B., and R. E. Holm. 1966. Enhancement of RNA synthesis, protein synthesis, abscission by ethylene. Plant Physiol. 41; 1337-1342.
Abeles, F. B., R. E. Holm, and H. E. Gahagan. 1967. Photo¬ control of ethylene production. Plant Physiol. (Suppl.) 42; 5-9.
Abeles, F. B., R. E. Holm, and H, E. Gahagan. 1967. Abscission; the role of aging. Plant Physiol. 42; 1351-1356.
Abeles, F. B., and J. Lonski. 1969. Stimulation of lettuce seeds germination by ethylene. Plant Physiol. 44: 277-280.
Abeles, F. B., and B. Rubinstein. 1964. Regulation of ethylene evolution and leaf abscission by auxin. Plant Physiol. 39:
963-969.
Abrams, G. J. von, and K. K. Pratt. 1967. Effect of ethylene on .he permeability of excised ca.icaloupe fruit tissue.
Plant Physiol. 42; 299-301.
Allen, R. D. 1969. Mechanism of the seismonastic reaction in Mimosa pudica. Plant Physiol. 44; 1101-1107.
Apelbaum, A., and S. P. Burg. 1971. Ethylene effects on cellular division, expansion and differentiation. Plant
Physiol. (Suppl.) 47; 14.

102
15. Arnold, W. E., and J. D. Nalewaja. 1971. Effect of dicamba on RNA and protein. Weed Sci. 19: 301-305.
16. Ashton, F. M., D. Penner, and S. Hoffman. 1968. Effect of sev¬ eral herbicides on proteolytic ■'.ctivity of squash seedlings. Weed Sci. 16: 169-171.
17. Bachelard, E. P., and R. D. Ayling. 1971. The effects of picloram and 2,4-D on plant cell membranes. Weed Res. 11: 31-36.
18. Baur, A. H. and T. F. Yang. 1969. Precursors of ethylene. Plant Physiol. 44: 1347-1349.
19. Baur, A. H., T. F. Yang, H. K. Pratt, and J. B. Biale. 1971. Ethylene biosynthesis in fruit tissues. Plant Physiol. 47: 696-699.
20. Baur, J. R., and P. W. Morgan. 1969. Effects of picloram and ethylene on leaf movement in huisache and mesquite seed¬ lings. Plant Physiol. 44: 831-838.
21. Baur, J. R., and M. Workman. 1964. Relationship between cell permeability and respiration in ripening banana fruit tissue. Plant Physiol. 39: 540-543.
22. Beevers, L., D. M. Peterson, T. C. Shannon, and R. H. Hageman. 1963. Comparative effects of 2,4-D on nitrate metabolism in corn and cucumber. Plant Physiol. 38: 675-679.
23. Behrens, R. 1967. The influence of environment on the response of wild buckwheat to dicamba. Proc. NCWCC, p. 42.
24. Beyer, E. M., Jr., and P. W. Morgan. 1969. Ethylene modification of an auxin pulse in cotton stem sections. Plant Physiol. 44: 1690-1694.
25. Beyer, E. M., Jr., and P. W. Morgan. 1970. Effect of ethylene on uptake, distribution, metabolism of lAA-l-l^C and IAA-2-l^C and naphtaleneacetic acid-l-l^C. Plant Physiol. 46: 157-162.
26. Beyer, E. M., and P. W. Morgan. 1970. A method for determining ^he concentration of ethylene gas phase of vegetative plant tissues. Plant Physiol. 46: 352-354.
Binning, L. K., D. Penner, and W. F. Meggett. 1971. The effect of 2-chloroethylphosphonic acid on dicamba translocation
in wild garlic. Weed Sci. 19: 73-75.
27.

103
28. Briggs, L. J., and H. J. Shantz. 1914. Relative water require¬ ment of plants. J. Agr. Res. 3; 1-63.
29. Burg, S. P. 1962. The physiology of ethylene formation. Ann. Rev. Plant Physiol. 13: 265-302.
30. Burg, S. P. 1968. Ethylene plant senescence and abscission. Plant Physiol. 43; 1503-1511.
31. Burg, S. P., and Ellen A. Burg. 1962. Role of ethylene in fruit ripening. Plant Physiol. 37: 179-189.
32. Burg, S. P., and Ellen A. Burg. 1964. Evidence for a natural occurring inhibitor of fruit ripening. Plant. Physiol. (Suppl.) 39: X.
33. Burg, S. P., and Ellen A. Burg. 1966. The interaction between auxin and ethylene and its role in plant growth. Proc. Natl. Acad. Sci. U.S. 55: 262-269.
34. Burg, S. P., and Ellen A. Burg. 1966. Auxin induced ethylene formation: its relation to flowering in the pineapple. Science 152: 1269.
35. Burg, S. P., and Ellen A. Burg. 1966. Effect of ethylene on auxin metabolism and transport in etiolated pea stems. Plant Physiol. 41 (Suppl.): 1111.
36. Burg, S. P., and Ellen A. Burg. 1967. Molecular requirements for the biological activity of ethylene. Plant Physiol. 42: 144-152.
37. Burg, 3. P., and Ellen A. Burg. 1967. Lateral auxin transport in stems and roots. Plant Physiol. 42: 891-893.
38. Burg, S. P., and Ellen A. Burg. 1967. Inhibition of polar auxin transport by ethylene. Plant Physiol. 42: 1224-
1228.
39. Burg, S. P., and Ellen A. Burg. 1967. Ethylene action independ¬ ent of protein synthesis. Plant Physiol. 42 (Suppl.); 5-31.
40. Burg, S. P., and Ellen A. Burg. 1968. Ethylene formation in pea seedlings; its relation to the inhibition of bud growth caused by indole-3-acetic acid. Plant Physiol. 43: 1069-
1074.
Burg, S. P., Ellen A. Burg, and R. Marks. 1964. Relationship of solute leakage to solution tonicity in fruits and other plant tissues. Plant Physiol. 39: 185-195.
41.

104
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
Burg, S. P., and M. J. Dijkman. 1967. Ethylene and auxin
participation in pollen induced fading of Vanda orchid blossoms. Plant Physiol. 42: 1648-1650.
Burnside, 0. C., and T. L. Lavy. 1966. Dissipation of dicamba. Weeds 14: 211-214.
Burnside, 0. C., G. A. Wicks, and C. R. Fenster. 1971. Dissi¬ pation of dicamba, picloram and 2,3,6-TBA across Nebraska. Weed Sci. 19: 323-325.
Chadwick, A. W., and S. P. Burg. 1967. An explanation of the inhibition of root growth caused by indole-3-acetic acid. Plant Physiol. 42: 415-420.
Chadwick, A. W., and S. P. Burg. 1970. Regulation of root growth by auxin-ethylene interaction. Plant Physiol. 45: 192-200.
Chang, F. Y. 1969. Translocation and metabolism of 2-methoxy- 3,6-dichlorobenzoic acid in plants. Ph.D. Thesis. Univ. of Alberta, pp. 109.
Chang, F. Y., and W. H. Vanden Born, dicamba in Canada thistle. Weed
1968. Translocation of Sci. 16: 176-181.
Chang, F. Y., and W. H. Vanden Born, metabolism of dicamba in Tartary
107-112.
1971. Translocation and buckwheat. Weed Sci. 19:
Chang, F. Y., and W. H. Vanden Born. 1971. Dicamba uptake,
translocation, metabolism and selectivity. Weed Sci. 19: 113-117.
Connard, M. H., and P. W. Zimmerman. 1931. The origin of adventitious roots in cuttings of Portulaca oleracea L. Contrib. Boyce Thompson Institute 3: 337-346.
Cooper, W. C., G. K. Rasmussen, B, T. Rogers, P. C, Reece, W. H. Henry. 1968. Control of abscission in agricultural crops and its physiological basis. Plant Physiol. 43: 1560-1576.
Cooper, W. C., G. K. Rasmussen, E. S. Waldon. 1969. Ethylene evolution stimulated by chilling in citrus and Persea sp.
PI. Physiol. 44: 1194-1196.
54. Cotter, D. J., and R. B. Platt. 1959. Studies on the ecological life history of Portulaca smallii. Ecology 40: 651-658.

105
55. Crafts, A. S., and S. Yamaguchi. 1964. The Autoradiography of Plant Materials. Univ. of California Agr. Publ. Manual 35. Berkeley, Calif. 143 p.
56. Crocker, L. E., and F. B. Abeles. ‘>969. Abscission: role of abscissic acid. Plant Physiol. 44: 1144-1149.
57. Curtis, R. W. 1968. Mediation of a plant response to malformin by ethylene. Plant Physiol. 43: 76-80.
58. Dolliet, H. H. A., and J. Kumamoto. 3.970. Ethylene production
of ethyl propylphosphonate, Niagara 10637. Plant Physiol. 46: 786-789.
59. Dunham, R. S., R. G. Robinson, and R. N. Anderson. 1958. Crop rotation and associated tillage practices for controlling annual weeds in flax and reducing the weed seed population
in soil. Tech. Bull. Minn. Agric. Expt. Sta. 230. Weed
Abstr. 1960: 364.
60. Dunn, S. 1970. Light quality effects on the life of common purslane. Weed Sci. 18: 611-613.
61. Eaks, L. 1970. Respiratory response, ethylene production and response to ethylene of citrus fruits during antogeny. Plant Physiol. 45: 334-338.
62- El-Sheikh, A. M., T. C. Brayer, and A. Ulrich. 1971. Interaction of rubidium or sodium with potassium in absorption by intact sugar beet plants. Plant Physiol. 47: 709-712.
63. Esashi, Y., and A. C. Leopold. 1969. Dormancy regulation in subterranean clover seeds by ethylene. Plant Physiol. 44:
1470-1472.
64. Esau, C. 1965. Plant Anatomy. John Wiley and Sons, New York.
pp. 767.
65. Foy, C. L., and W. Hurtt. 1967. Further studies on root exudation of exogenous growth regulators in Phaseolus vulgaris. WSA
Meet. Abstr. 40.
66. Foy, C. L., and D. Penner. 1965. Effect of inhibitors and herbicides on tricarboxylic ac-Jd cycle substrate oxidation
by isolated cucumber mitochondria. Weeds 13: 226-231.
Friesen, H. A. 1965. The movement and persistence of dicamba
in soil. Weeds 13: 30-33. 67.

106
69. Fuchs, Y., and M. Lieberman. 1968. Effects of kinetin, lAA and gibberellin on ethylene production and their interaction in growth of seedlings. Plant Physiol. 43: 2029-2036.
70. dela Fuente, R. K., and A. C. Leopold. 1968. Senescence
processes in leaf abscission. Plant Physiol. 43: 1496- 1502.
71. Goeschl, J. D., H. K. Pratt, and B. A. Bonner. 1967. An effect of light on the production of ethylene and the growth of the plumular portion of etiolated pea seedlings. Plant Physiol. 42: 1077-1080.
72. Goeschl, J. D., L. Rappaport, and H. K. Pratt. 1966. Ethylene as a factor regulating the growth of pea epicotyls subjected to physical stress. Plant Physiol. 41: 877-884.
73. Greenberg, D. M. 1957. Preparation of samples of protein, amino acids, and other solids. In: Methods in Enzymology, V. N. Ac. Press Inc. New York, p: 708-711.
74. Gruenhagen, R. D., and D. E. Moreland. 1971. Effects of herbicides on ATP levels in excised soybean hypocotyls. Weed Sci. 19: 319-325.
75. Hageman, R. H., and D. Flescher. 1960. Nitrate reductase act¬ ivity in corn seedlings as affected by light and nitrate content of nutrient media. Plant Physiol. 35: 700-708.
76. Hahn, R. R., 0. C. Burnside, and T. L. Lavy. 1969. Dissipation and phytotoxicity of dicamba. Weed Sci. 17: 3-8.
77. Hallaway, M., and D. J. Osborne. 1969. Ethylene: a factor in defoliation induced by auxins. Science 163: 1067-1068.
78.. Heilman, M. D., and F. J. Meredith. 1971. Ethylene production in the cotton plant canopy and its effect on fruit abscission.
Crop Sci. 11: 25-27.
79. Heinrich, G. 1962. Fusarinsaure in ihrer Wirkung auf die Wasser- permeabilitat des Protoplasmas. Protoplasma 55: 320-356.
80. Heinrich, G. 1964. Huminsaiire und Permeabilitat. Protoplasma
58: 403-425.
Henry, E. W., J. G. Valdovinos, and T. E. Jensen. 1971. Ethylene enhanced peroxidases in abscission cells of tobacco flower pedicels. Plant Physiol. (Suppl.) 47: 14 (Abstr.).
81.

107
82. Hoagland, D, R., and D. I, Arnon. 1950. The water culture. Cali¬ fornia Agr. Exp. Sta, Circ. 347.
83. Hocombe, S. D. (n.d,). Simple experiments on the greenhouse germination of some East African weed species. Mis. Rep. Colon. Pest. Res. Unit Arusha, Tanganyika. 285, pp. 8. Weed Abstr. 1961: 505.
84. Holm, R. E., and F. B. Abeles. 1967. Abscission; the role of RNA synthesis. Plant Physiol. 42: 1094-1102.
85. Holm, R. E., and F. B. Abeles. 1967. 2,4-Dichlorophenoxyacetic acid-induced ethylene evolution: its role in soybean hypocotyl swelling. Plant Physiol. (Suppl.) 42: 30-1.
86. Holm, R. E., T. J. O’Brien, J. L. Key, and T. H. Cherry. 1970. The influence of auxin and ethylene on chromatin-directed ribonucleic acid synthesis in soybean hypocotyl. Plant Physiol. 45: 41-45.
87. Holm, R. E., and J. L. Key. 1969. Hormonal regulation of cell elongation in the hypocotyl of rootless soybean. An
evaluation of the role of DNA synthesis. Plant Physiol. 44: 1295-1302.
88. Hull, R. J. (Chief ed.) ^ al^* 1967. Herbicide Handbook of the Weed Society of America. 1st ed, W. F. Humpherey Press, Inc., Geneva, New York.
89. Hull, R. T., and M. R. Weisenberg. 1967. Translocation and metabolism of dicamba in Sorghum halepense and Phaseolus vulgaris. Plant Physiol. 42 (Suppl.): 49. (Abstr.).
90. Hurtt, W., and C. L. Foy. 1965. Some factors influencing the excretion of foliarly applied dicamba and picloram from roots of Black Valentine beans. Plant Physiol. (Suppl.)
40: xlviii. (Abstr.).
91. Hyado, H., and S. F. Yang. 1971. Ethylene-enhanced synthesis of phenylalanine ammonia-lyase in pea seedlings. Plant
Physiol. 47: 705-770.
92. Imaseki, H, 1970. Induction of peroxidase activity by ethylene
in sweet potato. Plant PhysioJ. 46: 172-174.
Jacobsen, J. V., and W. B. Glasson. 1970. Ethylene production by autoclaved rubber injection caps used in biological
systems. Plant Physiol. 45: 631.
93.

108
94. Jaffe, M. T., and A. W. Galston. 1967, Phytochrome control of rapid nyctinastic movements and membrane permeability in Albizzia julibrissin. Planta 77: 135-141,
95. Jagschitz, J. A. 1966. Chemical control of spurry, chickweed and purslane. Proc. NEWCC 20: 502-509.
96. Jagschitz, J. A., and A. R. Mazur. 1967. Chemical control of purslane and other broadleaf weeds in seedling turf. Proc. NEWCC 21: 520-523.
97. Janes, D. W., and C. L. Foy. 1971. Effect od dimethylsulfoxide, alone and in combination with three herbicides and surfactant in membrane permeability. WSSA Mtg. Abstr. p. 12.
98. Janes, R. L. 1968. Ethylene enhanced release of ^-amylase from barley aleurone cells. Plant Physiol. 43: 442-444.
99. Kang, B. G., and S. P. Burg. 1970. Ethylene synthesis and growth in relation to lAA metabolism. Plant Physiol. 46 (Suppl.): 20.
100. Ketring, D. L., and P. W. Morgan. 1969. Ethylene as a component of the emanations from germinating peanut seeds and its effect on dormant Virginia-type seeds. Plant Physiol. 44: 326-330.
101. Ku, H. S., and A. C. Leopold. 1970. Mitochondrial responses to ethylene and other hydrocarbons. Plant Physiol. 46: 842- 844.
102. Ku, H. S., and A. C. Leopold. 1970. Experiments on the mode of action of ethylene in plants. Plant Physiol. (Suppl.) 46:
20. 103. Ku, H. S., and H. K. Pratt. 1968. Active mitochondria do not
produce ethylene. Plant Physiol. 43: 999-1001.
104. Kutschera, L. 1960. Wurzelatlass. DLG - Verlags - GMBH, Frankfurt am Mein. 574 p.
105. Layne, E. 1957. Spectrophotometric and turbidimetric methods for measuring protein. In: Methods in Enzymology, v. III. Ac. Press Inc., New York, p: 447-454.
Leonard, 0. A., and R. K. Glenn. 1968. Translocation of herbicides in detached bean leaves. Weed Sci. 16: 352-356.
106.

109
107. Lieberman, M., and A. Kunishi. 1966. Stimulation of ethylene production in apple tissue slices by methionine. Plant Physiol. 41: 376-382.
108. Lyons, J. M., W. B. MacGlasson, H. K. Pratt. 1962. Ethylene production, respiration and internal gas concentrations in cantaloupe fruits at various stages of maturity. Plant Physiol. 37: 31-36.
109. McGlasson, W. B., and H. K. Pratt. 1964. Effect- of wounding on respiration and ethylene production by cantaloupe fruit tissue. Plant Physiol. 39: 128-132.
110. Magalhaes, A. C., and F. M. Ashton. 1969. Effect of dicamba on oxygen uptake and cell membrane permeability in leaf tissue
Cyperus rotundus L. Weed Res. 9: 48-52.
111. Magalhaes, A. C., F. M. Ashton, and C. L. Foy. 1968. Trans¬ location and fate of dicamba in purple mutsedge. Weed Sci. 16: 240-245.
112. Mann, J. D., L. S. Jordan, and B. E. Day. 1965. A survey of herbicides for their effect upon protein synthesis. Plant Physiol. 40: 840-843.
113. Marei, N., and R. Romani. 1971. Enhancement of ribosomal, RNA and protein synthesis in fig fruits by ethylene. Plant Physiol. (Suppl.) 47: 15.
114. Mattao, A. K., and V. V. Modi. 1969. Ethylene and ripening of mango. Plant Physiol. 44: 308-310.
115. Maxie, E. C., and J. C. Crane. 1967. 2,4,5-T. Effect on
ethylene production by fruits and leaves of fig tree. Science 155: 1548-1549.
116. Maxie, E. C., I. L. Eaks, N. F. Sommer, H. L. Roe, and S. El-Batal. 1965. Effect of gamma radiation on rate of ethylene and CO2 evolution by lemon fruit. Plant Physiol. 40: 407-409.
117. Maxie, E, C., N. F. Sommer, C. J. Muller, and H. L. Roe. 1966. Effect of gamma radiation on the ripening of Bartlett pears.
Plant Physiol. 41: 437-442.
118. Maynard, D. N., and J. H. Baker. 1965. The influence of rubidium and potassium levels on growth and ion accumulation in tomato. Plant and Soil 23: 137-140.
Mehard, C. W., and J. M. Lyons. 1970. A lack of specificity for ethylene induced mitochondrial changes. Plant Physiol. 45:
36-39.
119.

110
» «
120. Meinl, G., and H. von Guttenberg. 1952. Uber den Einfluss von
Wirkstoffen auf die PermeabilitSt des Protoplasmas. Planta 41: loy-ise.
121. Meudt, W. J., and K. J. Stecher. 1971. Peroxidase in Xaiithi tobacco plants treated with auxin and benzoic acid deriv¬ atives. Plant Physiol. (Suppl.) 47: 4.
122. Mitsuhashi, M., H. Shibaoka, and M. Shiraokoriyama. 1969. Portulal a rooting promoting substance in Portulaca leaves. Plant and Cell Physiol. 10: 715-723.
123. Monsanto Company. June 1953. Technical Bulletin No. 0-50.
124.. Moreland, D. E., S. S. Malhatra, R. D. Gruenhagen, and E. H. Shakraii. 1969. Effects of herbicides on RNA and protein synthesis. Weed Sci. 17: 556-562.
125. Morgan, P. W. 1969. Stimulation of ethylene evolution and abscission in cotton by 2-chloroethanephosphonic acid.
Plant Physiol. 44: 337-341.
126. Morgan, P. W., and T. R. Baur. 1970. Involvement of ethylene in picloram-induced leaf movement response. Plant Physiol. 46: 655-659.
127. Morgan, P. W., and H. W. Gausman. 1966. Effects of ethylene on auxin transport. Plant Physiol. 41: 45-52.
128. Morgan, P. W., R. E. Meyer, and M. G. Merkle. 1969. Chemical stimulation of ethylene evolution and bud growth. Weed Sci. 17: 353-355.
129. Morgan, P. W., and R. D. Powell. 1970. Involvement of ethylene in responses of etiolated bean hypocotyl hook to coumarin.
Plant Physiol. 45: 553-557.
130. Muensher, W. C. 1955. Weeds. 2nd edn. The Macmillan Co., New York, 560 p.
131. Nalev^aja, F. 1969. Reaction of flax to dicaraba applied at several stages. Weed Sci. 17: 385-387.
132. Pate, D. A., H. H. Funderburk, J. M. Lawrence, and D. E. Davis. J.965. The effect of dichlobenil and dicamba on nodal tissues of alligatorweed. Weeds. 13: 208-210.
Patek, V., and T. Elaha. 1959. The ability of vineyard weeds to regenerate. Polhospodarstwo 6 (3): 423. Weed Abstr.
1959: 1670.
133.

Ill
134. Perkin-Elmer Corp. 1971. Analytical methods for atomic absorption spectrophotometry. Instr. manual.
135. Poellnitz, Karl, von. 1934. Versuch zur einer Monographie der Gattung Portulaca L. Repertorium specierum novarum regni
vegetabilis. 37: 240-320. F. fedde (Publisher), Berlin- Dahlem.
136. Pratt, H. K., and J. D. Goeschl. 1964. Some relationships of ethylene to the respiratory patterns of honey dew melons. Plant Physiol. (Suppl.) 39: ix.
137. Pratt, H. K. and J. D. Goeschl. 1969. Physiological roles of ethylene in plants. Ann. Rev. Plant Physiol. 20: 541-584.
138. Quimby, P. C. 1967. Studies relating to the selectivity of dicamba for wild buckwheat vs Selkirk wheat and a possible mode of action. Ph.D. Thesis. N. Dakota Univ. pp. 87.
139. Radin, J. W., and R. S. Loomis. 1969. Ethylene and CO2 in the growth and development of cultured radish roots. Plant Physiol. 44: 1584-1589.
140. Rains, D. W. 1968. Kinetic and energetic of light-enhanced potassium absorption by corn leaf tissue. Plant Physiol. 43: 394-400.
141. Rains, D. W. 1969. Sodium and potassium absorption by bean stem tissue. Plant Physiol. 44: 547-554.
142. Ratner, A., R. Goren, and S. P. Monselise. 1969. Activity of pectin esterase and cellulase in the abscission zone of citrus leaf explants. Plant Physiol. 44: 1717-1723.
143. Ray, B. 1967. Chemical control of nutsedge (Cyperus rotundus L.) and the metabolism of 3,6-dichloro-a-anisic acid (dicamba).
Ph.D. Thesis. Univ. of Florida, pp. 87.
144. Reid, C. P., and W. Hurtt. 1970. Root permeability as affected by picloram and other chemicals. Physiol. Plantarum 23: 124-
130.
145. Riov, J., S. P. Monselise, and R. S. Kahan. 1969. Ethylene- controlled induction of phenylalanine ammonia-lyase in
citrus fruit peel. Plant Physiol 44: 631-635.
146. Rogerson, A. B., and C. L, Foy. 1968. Effect of dicamba and picloram on growth and anthocyanin content in Black Valentine beans under different environmental conditions.
Proc. SWC. 21: 347.

112
147. Rosen, L. A., and S. M. Siegel. 1963. Effect of oxygen tension on the course of ethylene and gibberellin-induced foliar abscission. Plant Physiol. 38: 189-191.
148. Rubinstein, B., and F. B. Abeles. 1965. Relationship between ethylene evolution and leaf abscission. Plant Physiol. CSuppl.) 40: xxvi.
149. Sachet, J. A. 1966. Perxaeability characteristics and amino acid incorporation during senescence (ripening) of banana tissue. Plant Physiol. 41: 701-708.
150. Sachet, J. A., and S. 0. Salninen. 1969. Comparative studies of eff-^cts of auxin and ethylene on permeability and syn¬ thesis of RKA and protein. Plant Physiol. 44: 1371-1377.
151. Santas, C. A., A. L. Das, and L. Leiderman. 1967. Preplanting and preenergence application of herbicides in potato fields. Biologico 33: 91-96. Weed Abstr. 1968: 95.
152. Schroder, L. E., G. L. PJ.ttenaur, G. L. Eilrich, and R. H. Hageman.
1968. Some characteristics of nitrate reductase from higher plants. Plant Physiol. 43: 930-940.
153. Scott, P. C., and A. C. Leopold. 1967. Opposing effects of gibberellin and ethylene. Plant Ph3/siol. 42: 1021-1022.
154. Scott, P. C., and R. 0. Morris. 1970. Quantitative distribution
and netabolism of auxin herbicides in roots. Plant Physiol. 46: 680-684.
155. Skoglund, S. A., and R. J. Coupland. 1960. Relative susceptibility of annual weeds to several herbicides. Proc. NCVCC 17: 78.
156. S’aith, R. C., and E. Epstein. 1964. Ion absorption by shoot tissue: kinetics of potassium and rubidium absorption by corn leaf tissue. Plant Physiol- 39: 992-996.
157. Stabeaxm, M. A., B. G. Clare, and W. Woodbury. 1966. Increased disease resistance and enzyme activity induced by ethylene aod ethylene production by black rot infected sweet potato
tissue. Plant Physiol. 41: 1505-1512.
158. Steel, R. G- D., and J. H. Torrie. 1960. Principles and Pro¬ cedures of Statistic. McGraw-Eill Book Co. New York,
Toronto, London, pp. 431.
Steiner, E. 1944. Cytogenetic studies on Talinum and Portulaca.
Bot. Gaz. 105: 374-379. 159.

113
160. Steward, E. R., and H. T. Freebairn. 1969. Ethylene, seed germination and epinasty. Plant Physiol. 44: 955-958.
161. Tanada, T, 1963. Kinetics of rubidium absorption by excised barley roots under changing rubidium concentrations. Effects of other cations on rubidium uptake. Plant Physiol. 38: 422-425.
162. tJ. S. Department of Agriculture. November 1948. Farmer’s Bulletin 2005.
163. U. S. Department of Agriculture, Agricultural Research Service. 1968. Extent and cost of weed control with herbicides and evaluation of important weeds. 1965. 85 p.
164. U. S. Department of Agriculture, Agricultural Research Service. 1970. Selected Weeds of the United States. Agricultural Handbook No. 366, 463 p.
165. Vanden Born, W. H. 1965. The effect of dicamba and picloram on quackgrass, bromegrass and Kentucky bluegrass. Weeds 13: 309-312.
166. Vanden Born, W. H. 1968. Uptake and release of dicamba by oat coleoptile sections. WSSA Abstr.: 14-15.
167. Vengris, J., and M. Stacewicz. 1969. Germination, growth, development and competitiveness of common purslane (Portulaca oleracea). Suppl. Proc. NEWCC 23: 39.
168. Vengris, J., and M. S. Sapuncakis. 1971. Common purslane competition in table beets and snap beans. Weed Sci. 19:
4-6.
169. Warner, H. L., and A. G. Leopold. 1968. Ethylene formation from methionine. Plant Physiol. (Suppl.) 43: 5-45.
170. Warner, H. L., and A. C. Leopold, 1969. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 44: 156-158.
171. Wax, L. M., L, A. Knuth, and F. W. Slife. 1969. Response of soybeans to 2,4-D, dicamba and picloram.. Weed Sci. 17:
388-393.
172. Wehsarg, 0. 1954. Ackerunkrauter. Akademie Verlag, Berlin.
173. Wiliier, T. (n.d.). Methods for evaluating winter hardiness of plants. Plant Res. Inst. Canada Dept, of Agriculture.
Yang, S. F. 1969. Ethylene evolution from 2-chloroethylphosphonic
acid. Plant Physiol. 44: 1203-1204. 174.

APPENDIX 114
081 josqV
J
KiK
'iro
1.
Rln
ncl
nn
l cu
rvr
l or
nib
itll
uin
iloto
rmln
nL
Io
n.

Ab
sorb
an
ce
540
my
115
.80
.70
.60
.50
.40
.30
,20
ao
.01 .02 .03 .04 .05 .06 .07
ymoles nitrite
Figure II. Standard curve for nitrite determination.

Abso
rbance
55
0
my
.40
.35
.30
.25
,20
.15
.10
.05
mg protein
Figure III. Protein standard for biuret test.

117
o o O O o o o O o o o MD CM 00 CM iH t—1
sijTun j[0 5t?.iSa:}ui





















